Introduction
It has been reported that over 80% of osteosarcoma
(OS) patients would die of metastases if they did not receive
chemotherapy (1). Although
multimodal treatment approaches have improved overall survival,
survival rates are still far from satisfactory. Currently, the
5-year overall survival rate of patients with OS is ~60%, even if
patients received intensive chemotherapy (2). Therefore, uncovering the underlying
mechanisms of tumorigenesis of OS may assist in the development of
improved treatments and targeting genes that promote progression of
OS may hold potential for improving the efficiency of treatments in
the future.
Epithelial-mesenchymal transition (EMT) is crucial
for progression of OS (3). Whilst
effectors of EMT, such as E-cadherin, N-cadherin, vimentin,
fibronectin, matrix metalloproteinase (MMP)-2 and MMP-9 have been
extensively studied (4), the
identities of the upstream modulators of these markers are less
clear. Zinc finger E-box-binding homeobox (ZEB) family members are
candidates for the regulation of EMT (3). In OS, increased expression of ZEB1
and ZEB2 has been associated with cancer development and poor
prognosis (5). Increased
expression levels of ZEB1 transcriptionally represses the
expression of E-cadherin (6),
which in turn, favours migration of cancer cells (3). Furthermore, by interacting with the
Wnt pathway or p73 proteins, ZEBs can also promote cell cycle
progression and cell survival in different types of cancer,
including colon, lung and pancreatic cancer (7). In this regard, investigating negative
regulators of ZEB family members may provide additional
opportunities for preventing or reversing EMT and serve as an
alternative anti-tumour therapy.
Emerging evidence has shown that non-coding RNAs
including long non-coding RNAs (lncRNAs; RNA transcripts >200 bp
in length) and microRNAs (miRNAs; RNA transcripts ~22 bp in length)
are intrinsically involved in regulation of protein expression at
all levels of the protein production process (8,9).
Transcriptome studies have demonstrated multiple series of
dysregulated miRNAs in various tumours, including OS (10,11).
miRNAs directly bind to protein-coding mRNAs and prevent them from
being translated into proteins, and during the pathogenesis of OS
this mechanism is used to suppress expression tumour suppressor
genes (12). In the field of
lncRNAs, the mechanisms of regulation of expression of genes is
more complicated and diverse. Through binding to chromatin, mRNAs
or proteins, lncRNAs can decrease or increase the expression of
protein-coding genes and have been reported to be involved in
almost all aspects of OS (13).
Previous studies on the crosstalk between lncRNAs and miRNAs have
revealed a new mechanism of protein-coding gene modulation
(14–16). LncRNAs can act as competing
endogenous RNAs (ceRNAs) and de-repress (increase) the expression
of protein-coding genes by sequestering miRNAs (17). CeRNA-dependent regulation,
therefore, represents a novel mechanism for the upregulation of
oncogenes.
Sprouty receptor tyrosine kinase signalling
antagonist 4-intronic transcript 1 (SPRY4-IT1) is a lncRNA that has
been associated with several types of cancer, including OS
(18). Overexpression of SPRY4-IT1
in OS cells promoted proliferation, survival and migration;
however, the downstream effectors of SPRY4-IT1 remain unknown
(19). As ZEBs promote tumour
development, it was hypothesized that SPRY4-IT1 may function by
regulating ZEB expression or function. miR-101, a tumour suppressor
gene, was reported to suppress EMT by targeting ZEB1 and ZEB2 in
ovarian carcinoma (20).
Furthermore, SPRY4-IT1 has been shown to promote the proliferation
and migration of bladder cancer cells by sponging miR-101 (21). However, the association between
SPRY4-IT1 and miR-101 and the functional interaction between
miR-101 and ZEB1/ZEB2 have not been explored in OS. The hypothesis
of the present study was that SPRY4-IT1 acted as a ceRNA to
sequester miR-101, resulting in an increase in the levels of ZEB1/2
proteins, which in turn promoted the proliferation, migration and
invasion of OS cells.
In the present study, the role of SPRY4-IT1 in the
tumorigenesis of OS was investigated. The aim of the present study
was to explore a miR-101 and ZEB1/ZEB2 axis as a potential
downstream effector of SPRY4-IT1 involved in proliferation,
survival, migration and invasion in OS. The present study may
provide insights into a novel strategy for treatment of OS.
Materials and methods
Cell culture
The human normal osteoblast cell line hFOB 1.19
(ATCC® CRL-11372™), the OS cell lines U2OS
(ATCC® HTB-96™), MG-63 (ATCC® CRL-1427™) and
Saos-2 cells (ATCC® HTB-85™), and normal human 293 cells
(ATCC® CRL-1573™) were obtained from ATCC. All cells
were maintained in DMEM (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% heat-inactivated FBS (Gibco; Thermo Fisher
Scientific, Inc.) and 1% penicillin-streptomycin solution
(Sigma-Aldrich; Merck KGaA) in a humidified incubator with 5%
CO2 at 37°C.
Transfection
miRNA mimics, inhibitor, scrambled control oligos,
pGPH1 plasmid containing short hairpin (sh)RNA targeting SPRY4-IT1
and pGPH1 plasmid containing scrambled control shRNA were purchased
from Shanghai GenePharma Co., Ltd. For transfection of miRNAs,
MG-63 or U2OS cells were seeded into 6-well plates and cultured
with supplemented media overnight. When the confluency of cells
reached ~80%, the medium was replaced with DMEM without serum. A
total of 25 pmol miR-101 mimics/inhibitor or its scrambled control
(NC) was mixed with 7.5 μl Lipofectamine® RNAiMAX
reagent (Thermo Fisher Scientific, Inc.) and added to the cells. A
total of 6 h later, the medium containing the oligos for
transfection and Lipofectamine® was removed and replaced
with supplemented DMEM without antibiotics. RNA or protein lysates
were collected 24 or 48 h after transfection, respectively. For RNA
interference studies, 1 mg pGPH1 plasmid and 3 ml
Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.)
were mixed and added to MG-63 or U2OS cells. Cells were cultured
with the transfection reagent and plasmid for 48 h before
subsequent assays were performed unless otherwise stated.
Cell proliferation assays
Cell proliferation was evaluated using MTT and
colony formation assays. For the MTT assay, MG-63 or U2OS cells
were seeded into 96-well plates at a density of 5×103
cells/well and transfected with the miR-101 mimics, inhibitor or
shSPRY4-IT1 for 24, 48, 72 or 96 h. After the indicated amount of
time, 10 μl MTT solution (Sigma-Aldrich; Merck KGaA) in PBS
(5 mg/ml) was added to each well and incubated in a cell culture
incubator for 3 h. The supernatant was removed carefully, and the
formazan crystals were dissolved in 100 μl DMSO
(Sigma-Aldrich; Merck KGaA). The cell viability in each well was
determined by measuring the optical density at 490 nm.
For the colony formation assay, MG-63 or U2OS cells
were seeded and transfected with miRNA mimics, inhibitors or
shSPRY4-IT1. Cells were then re-seeded in 6-well plates at a
density of 500 cells/well and allowed to grow for an additional 15
days. Cells were then fixed with 100% methanol for 10 min at −20°C
and stained with 0.5% crystal violet (in 25% methanol) for 10 min
at room temperature. The colonies were counted by two researchers
who were blinded to the experimental conditions.
Cell apoptosis analysis
MG-63 or U2OS cells were seeded in 12-well plates at
a density of 1×105 cells/well. After trans-fection, the
cells were trypsinized and harvested for staining using an Annexin
V-fluorescein isothiocyanate (FITC)/prop-idium iodide (PI)
Detection kit (Sigma-Aldrich; Merck KGaA) according to the
manufacturer's protocol. Cells were analysed by flow cytometry
(Becton, Dickinson and company). The
FITC+/PI− and FITC+/PI+
fractions were considered early and late apoptotic cells,
respectively. Analysis of flow data was performed using FlowJo
version X.10.0.7-1 (FlowJo LLC).
Assessment of cell cycle progression
Cell cycle progression was evaluated using flow
cytometry. For flow cytometry, after transfection as described
above, MG-63 or U2OS cells were trypsinized and harvested and fixed
with ice-cold 70% ethanol overnight at −20°C. Subsequently, the
cells were incubated with PBS solution containing PI (50
μg/ml) and RNase A (30 μg/ml) for 15 min at 37°C.
Cell cycle distribution was analysed by flow cytometry (Becton,
Dickinson and Company).
Cell migration and invasion assays
Cell migration and invasion were evaluated using a
wound healing assay and Transwell invasion assay, respectively. For
the wound healing assay, MG-63 or U2OS cells were seeded onto
12-well plates and transfected as described above. After 48 h of
transfection, a wound was created by scratching the cell monolayer
with a 1 ml pipette tip. Cells were then washed with PBS three
times and incubated for an additional 24 h in serum-free culture
medium. Distances were measured and analysed by ImageJ software
(NIH, US). Relative wound closure was measured using the following
formula: (W0h−W24 h)/W0 h ×100%;
where W is the width.
Matrigel-coated Transwell inserts or uncoated
inserts with 5-μm pores were used (Corning, Corning, NY,
USA) in a 24 well plate for Transwell invasion and migration
assays, respectively. MG-63 or U2OS cells were transfected as
described above and subsequently 5×104 cells were added
to the top chamber of the inserts with 400 μl serum-free
culture medium. The bottom compartments were filled with DMEM
containing 20% FBS as a chemoattractant. After 24 h of culture, the
inserts were collected, the non-invading or non-migrating cells on
the upper surface were removed with a cotton swab, and the invaded
or migrated cells on the lower surface were fixed with 4%
paraformaldehyde for 10 min at room temperature and stained with
crystal violet for 10 min at room temperature according to
manufacturer's protocol (Sigma-Aldrich; Merck KGaA). The invaded or
migrated cells were counted under an inverted light microscope with
a ×100 magnification (Carl Zeiss AG) by researchers who were
blinded to the experimental conditions.
Dual-luciferase reporter assay
The predicted miR-101 binding sites on SPRY4-IT1
were predicted using RNAInter website (rna-society.org/raid/search.html). The predicted
miR-101 binding sites on ZEB1 or ZEB2 were predicted using starBase
(starbase.sysu.edu.cn/index.php). Dual-luciferase reporter assays
were performed to determine whether miR-101 was targeted by
SPRY4-IT1 and the 3′ untranslated region (UTR) ZEB1/2. To confirm
the interaction between miR-101 and SPRY4-IT1, SPRY4-IT1 gene
fragments containing a predicted miR-101 binding site or
corresponding mutant fragments were cloned into pGL3-basic vectors
(Promega Corporation). miR-101 mimics or inhibitor were used for
miR-101 overexpression or knockdown, respectively. A pRL Renilla
Luciferase vector (Promega Corporation) was used as the control
plasmid. During transfection, miR-101 mimics or inhibitor and
pGL3-basic vectors were co-transfected into 293T cells with
Lipofectamine® 2000 (Thermo Fisher Scientific, San Jose,
CA, USA). Scrambled oligos were used as the control. After
transfection for 48 h, firefly luciferase activity was determined
and adjusted based on the Renilla luminescence activity according
to the manufacturer's protocol (Promega Corporation).
For the 3′UTR assay, oligos containing putative
miR-101 binding sites were cloned from the ZEB1 or ZEB2 3′UTR and
inserted into pmir-GLO vectors (Promega Corporation). The ZEB1 and
ZEB2 3′UTR, with the predicted microRNA seed binding regions
mutated were generated through site-directed mutagenesis kit
(Promega Corporation). The primer sequence for mutation of ZEB1
3′UTR was 5′-TAAAAATGTTGCACAG-3′. The primer sequence for mutation
of ZEB2 3′UTR was 5′-CTTGAAATAAATAAAAT-3′. During transfection,
miR-101 mimics or inhibitor and pmir-GLO vectors were
co-transfected into 293T cells with Lipofectamine® 2000.
After transfection for 48 h, relative luciferase activity was
measured.
Establishment of stable cell lines
MG-63 or U2OS cells were transfected with shNC
(scramble control) or shSPRY4-IT1 plasmids. Transfected cells were
selected for using 400 mg/ml Geneticin (Thermo Fisher Scientific,
Inc.). Stably transfected cells were maintained in culture medium
containing 100 mg/ml Geneticin and then used in the nude mouse
xenograft study.
Xenograft tumour model
BALB/c nude mice were purchased from Shanghai SLAC
Laboratory Animal Center (Shanghai, China). The nude mouse study
was approved by the Animal Ethics Committee at the Third Affiliated
Hospital of Sun Yat-sen University (Guangzhou, China). A total of
24 nude mice were randomly assigned to 4 groups (n=6/group);
MG-63/shNC (control), MG-63/shSPRY4-IT1 (treatment), U2OS/shNC
(control) and U2OS/shSPRY4-IT1 (treatment). A total of
1×106 cells/100 ml MG-63 or U2OS cells, which were
stably transfected with shNC or shSPRY4-IT1, were inoculated
subcutaneously in the nude mice. The xenograft tumour size was
measured every 5 days using a Vernier scale. After 30 days, the
mice were sacrificed by cervical dislocation. The tumour tissues
were harvested for volume and weight measurement, RT-qPCR and
western blot analysis.
RNA isolation and reverse
transcription-quantitative (RT-q) PCR
Total RNA from xenograft tumours and cells was
extracted using TRIzol® reagent (Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol.
Briefly, tumours or cells were dissolved in 1 ml
TRIzol®, followed by total RNA extraction with 200
μl chloroform and RNA precipitation with 500 μl
isopropanol. cDNA was synthesized from 1 μg total RNA using
the PrimeScript RT reagent kit (Takara Bio, Inc.). Reverse
transcription as performed as follows: 25°C for 5 min, 37°C for 30
min and 85°C for 5 sec. cDNA was diluted 20-fold with
ddH2O and used for qPCR with SYBR Premix EX Taq kit
(Takara Bio, Inc.) in an ABI 7500HT real-time PCR system (Thermo
Fisher Scientific, Inc.). The thermocycling conditions were as
follows: Initial denaturation, 95°C for 5 sec; followed by 35
cycles of denaturation at 94°C for 15 sec, annealing at 55°C for 25
sec and extension at 70°C for 30 sec. The gene levels for all
samples were normalized to U6 small nuclear (sn)RNA (for miRNA) or
GAPDH levels using the 2−ΔΔCq method (22).
Western blotting
Total protein was extracted with cell lysis buffer
(50 mM Tris, 150 mM NaCl, 1% NP-40, 1 mM EDTA, pH 7.6) containing a
cocktail of protease inhibitors (Thermo Fisher Scientific, Inc.).
Protein concentration was determined using a bicinchoninic acid
assay kit (Pierce; thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Samples (30 μg protein/lane) were
loaded on a 10% SDS gel and resolved using SDS-PAGE and then
transferred onto PVDF membranes (0.22 μm pore, Roche). After
blocking with TBS-Tween buffer (20 mM Tris, 137 mM NaCl, 0.1%
Tween-20, pH 8.0) containing 5% non-fat milk for 1 h at room
temperature, the membranes were incubated with primary antibodies
against ZEB1 (cat. no. 3396), ZEB2 (cat. no. 97885), E-cadherin
(cat. no. 3195), Vimentin (cat. no. 5741), Fibronectin (cat. no.
ab2413), N-cadherin (cat. no. 13116), MMP-2 (cat. no. 40994), MMP-9
(cat. no. 13667) or GAPDH (cat. no. 5174) overnight at 4°C. All
primary antibodies were purchased from Cell Signaling Technology,
Inc. except for Fibronectin, which was purchased from Abcam, and
all antibodies were used at a dilution of 1:1,000. Subsequently,
membranes were incubated with a goat anti-rabbit (#7074)
horseradish peroxidase-conjugated secondary antibody (1:3,000; Cell
Signaling Technology, Inc.) for 1 h at room temperature. The
protein bands were visualized using Immobilon Western
Chemiluminescent horseradish peroxidase substrate (EMD Millipore).
The proteins were quantified using Quantity One version 4.2.1
(Bio-Rad Laboratories, Inc.).
Statistical analysis
All experiments were performed at least three times
in triplicate. Data are presented as the mean ± standard deviation.
All statistical analyses were performed in GraphPad Prism version 6
(GraphPad Software, Inc.). Statistical evaluation was performed
using a two-tailed Student's t-test between two groups or a one-way
ANOVA followed by a post hoc Tukey's test for multiple comparisons.
P<0.05 was considered to indicate a statistically significant
difference.
Results
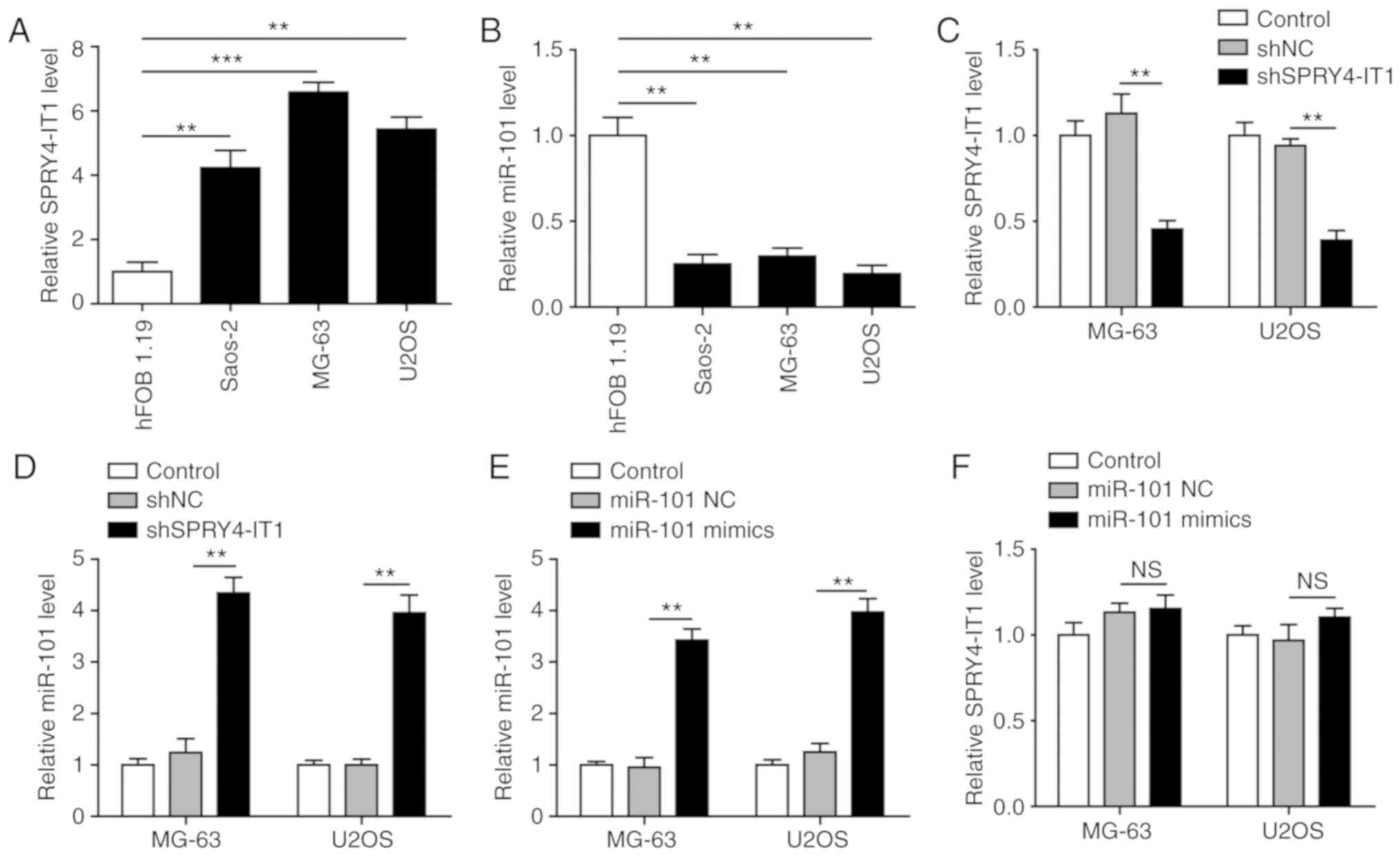
SPRY4-IT1 knockdown results in increased
miR-101 expression in OS cells
The expression of SPRY4-IT1 and miR-101 was detected
in three OS cell lines, Saos-2, MG-63 and U2OS, as well as the
normal osteoblast cell line hFOB 1.19. Compared with the hFOB 1.19
cells, SPRY4-IT1 was significantly upregulated in all three OS cell
lines (Fig. 1A). In contrast, a
reduction in miR-101 was observed in the three tumour cell lines
(Fig. 1B), highlighting a
potential functional interaction between SPRY4-IT1 and miR-101. To
examine this potential interaction, SPRY4-IT1 was knocked down in
MG-63 and U2OS cells. As shown in Fig.
1C, shSPRY4-IT1 transfection significantly decreased the
expression of SPRY4-IT1 compared with the control and shNC groups.
This was accompanied by a significant increase in miR-101
expression (Fig. 1D). However,
SPRY4-IT1 expression were not altered in cells transfected with
miR-101 (Fig. 1E and F). These
results suggest a unidirectional crosstalk between SPRY4-IT1 and
miR-101, such that SPRY4-IT1 may function as an upstream modulator
of miR-101 in OS cells.
SPRY4-IT1 silencing and miR-101
overexpression inhibits growth of OS cells
shSPRY4-IT1 or miR-101 mimic transfection was used
to study the molecular functions of SPRY4-IT1 and miR-101,
respectively. MTT and colony formation assays were first performed
to investigate the effects of SPRY4-IT1 and miR-101 on cell growth.
When compared with the scrambled control group, knockdown of
SPRY4-IT1 or overexpression of miR-101 significantly reduced the
growth of both MG-63 and U2OS cells in a time-dependent manner
(Fig. 2A). Consistent with these
results, colony formation assays revealed that SPRY4-IT1 knockdown
or miR-101 overexpression both significantly reduced the number of
colonies in MG-63 and U2OS cells (Fig.
2B and C). Furthermore, as shown in Fig. 2D and E, shSPRY4-IT1 transfection
resulted in a significant increase in the apoptotic ratio of MG-63
(shSPRY4-IT1, 30.51%; shNC, 8.56%) and U2OS (shSPRY4-IT1, 24.61%;
shNC, 7.39%) cells (both P<0.01). Similarly, overexpression of
miR-101 also greatly induced apoptosis in MG-63 (miR-101 mimics,
26.52%; miR-101 NC, 6.96%; P<0.05) and U2OS (miR-101 mimics,
31.43%; miR-NC 7.65%; P<0.01) cells. The effects of SPRY4-IT1
knockdown or miR-101 overexpression on cell cycle progression of OS
cells was assessed. shSPRY4-IT1 treatment of MG-63 and U2OS cells
resulted in a significant accumulation of cells in the G1 phase,
suggesting G1 phase arrest by SPRY4-IT1 knockdown (Fig. 2F and G). miR-101 mimic transfection
resulted in S phase cell cycle arrest (Fig. 2F and G). Together, these results
demonstrate that SPRY4-IT1 and miR-101 are involved in the growth
of OS cells at least partially by modulating proliferation,
apoptosis and cell cycle progression.
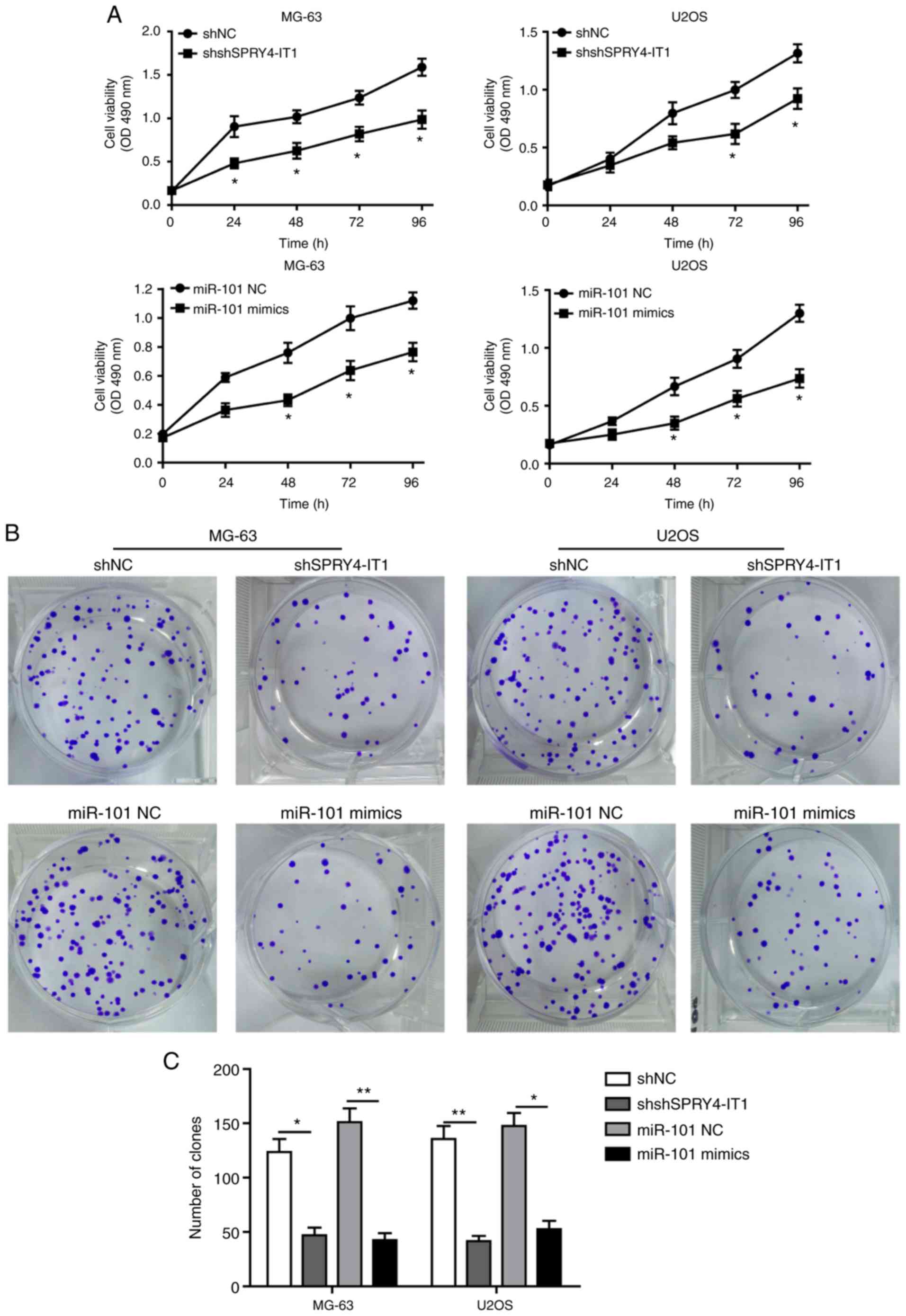 | Figure 2SPRY4-IT1 silencing or miR-101
overexpression reduces OS cell growth. (A) SPRY4-IT1 knockdown or
miR-101 overexpression significantly decreased proliferation of
MG-63 and U2OS cells as shown using an MTT assay. (B and C)
SPRY4-IT1 knockdown or miR-101 overexpression significantly reduced
colony formation in MG-63 and U2OS cells. Data are presented as the
mean ± standard deviation of three independent experiments.
*P<0.05, **P<0.01. SPRY4-IT1 silencing
or miR-101 overexpression reduces OS cell growth. (D and E)
SPRY4-IT1 knockdown or miR-101 overexpression significantly
increased apoptosis in both OS cell lines. Data are presented as
the mean ± standard deviation of three independent experiments.
*P<0.05, **P<0.01. SPRY4-IT1 silencing
or miR-101 overexpression reduces OS cell growth. (F and G)
SPRY4-IT1 knockdown or miR-101 overexpression resulted in
significant arrest of the cell cycle in MG-63 and U2OS cells. Data
are presented as the mean ± standard deviation of three independent
experiments. *P<0.05, **P<0.01.
SPRY4-IT1, sprouty receptor tyrosine kinase signalling antagonist
4-intronic transcript 1; miR, microRNA; OS, osteosarcoma; sh, short
hairpin; NC, negative control; OD, optical density; PI, propidium
iodide. |
SPRY4-IT1 downregulation or miR-101
overexpression attenuates migration and invasion of OS cells
Increased cell migration and invasion have been
identified as features of cancer metastasis, which is one of the
hallmarks of cancer (23). The
oncogenic function of SPRY4-IT1 and tumour suppressor function of
miR-101 on cell migration and invasion were further explored. After
24 h, SPRY4-IT1 downregulation or miR-101 induction significantly
inhibited wound closure in both MG-63 and U2OS cells, suggesting
that the migratory ability was reduced (Fig. 3A and B). A Transwell assay was also
performed to evaluate the effect of SPRY4-IT1 and miR-101 on the
migratory and invasive capacities of OS cells. shSPRY4-IT1
transfection significantly reduced the number of migrated or
invaded MG-63 and U2OS cells (Fig.
3C-F). Similarly, overexpression of miR-101 also significantly
reduced the migration or invasion of MG-63 and U2OS cells (Fig. 3C-F). Taken together, our results
suggested that SPRY4-IT1 promoted cell migration and invasion,
whereas miR-101 functioned as a negative regulator of these
properties in OS cells.
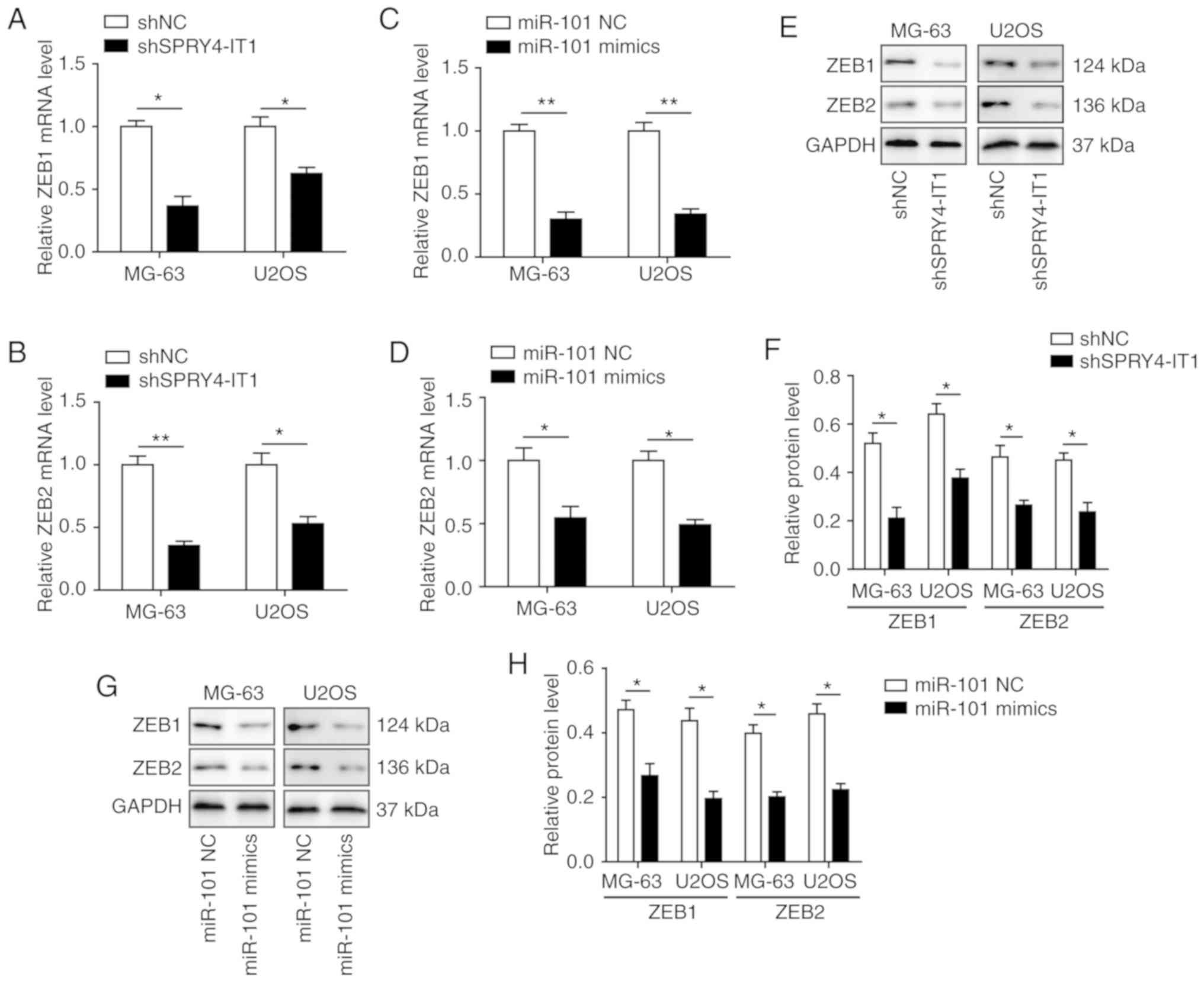
SPRY4-IT1 and miR-101 modulate the
expression of ZEB1 and ZEB2 in OS cells
Previously, it was reported that miR-101 may
regulate the expression of both ZEB1 and ZEB2, both of which have
been demonstrated to serve important roles in invasion and
metastasis of lung and ovarian carcinoma (20,24).
Therefore, the effects of SPRY4-IT1 and miR-101 on regulation of
ZEB1 and ZEB2 expression were determined. qPCR analysis showed that
transfection of shSPRY4-IT1 or miR-101 mimics was sufficient to
significantly reduce the mRNA expression levels of ZEB1 and ZEB2 in
both MG-63 and U2OS cells (Fig.
4A-D). Western blotting also showed that SPRY4-IT1 knockdown
and miR-101 overexpression significantly decreased the protein
expression levels of ZEB1 and ZEB2 (Fig. 4E-H). These findings suggest that
SPRY4-IT1 and miR-101 may modulate OS tumorigenesis by regulating
the expression of ZEB1 and ZEB2.
SPRY4-IT1 sponges miR-101 to regulate the
expression of ZEB1 and ZEB2
An increasing number of studies have revealed that
lncRNAs may sponge miRNAs and thus disinhibit the target genes of
these miRNAs (25). To investigate
the possibility of a SPRY4-IT1/miR-101/ZEB axis, the functional
interactions between SPRY4-IT1 and miR-101, as well as miR-101 and
ZEBs, were explored. RNAInter identified a putative miR-101 binding
sequence located in SPRY4-IT1 (Fig.
5A). Compared with the scrambled control, miR-101 mimics
significantly reduced SPRY4-IT1-WT-mediated relative luciferase
activity, whereas transfection with miR-101 inhibitor increased
luciferase activity (Fig. 5B).
Mutagenesis of the SPRY4-IT1 fragment (SPRY4-IT1-MUT) completely
abolished the effects of the miR-101 mimics and inhibitor. These
results suggest that SPRY4-IT1 may directly interact with miR-101
in OS cells.
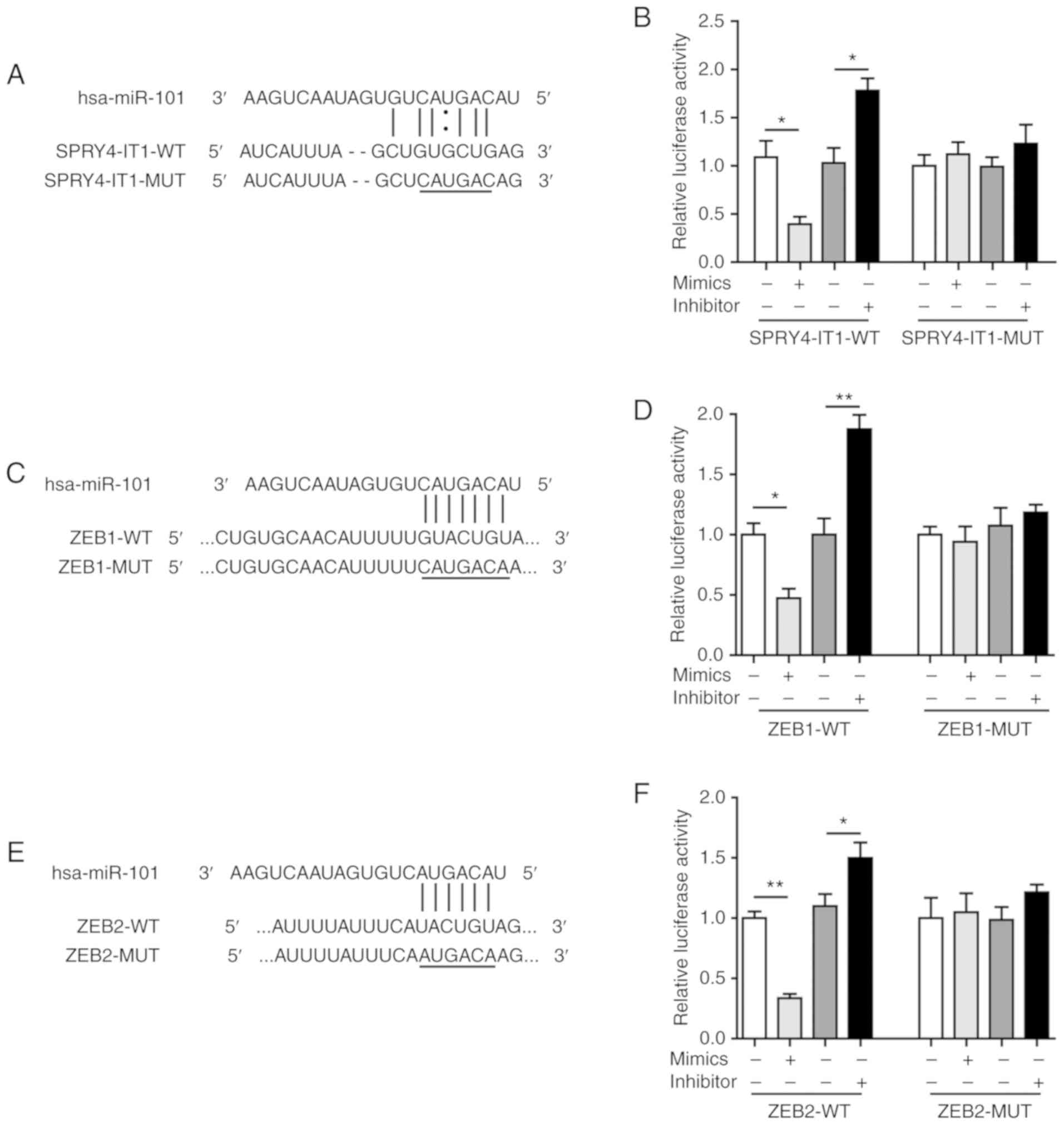 | Figure 5SPRY4-IT1 sponges miR-101 to regulate
the expression of ZEB1 and ZEB2. (A) Predicted binding site between
SPRY4-IT1 and miR-101. (B) miR-101 mimics reduced, whereas miR-101
inhibitor increased, luciferase activity in the SPRY4-IT1-WT group.
Luciferase activity was not altered by miR-101 mimics or inhibitors
in the SPRY4-IT1-MUT group. (C) Predicted binding site between
miR-101 and ZEB1. (D) miR-101 mimics reduced, whereas miR-101
inhibitor increased, luciferase activity in the ZEB1-WT group.
Luciferase activity was not altered by miR-101 mimics or inhibitors
in the ZEB1-MUT group. (E) Predicted binding site between miR-101
and ZEB2. (F) miR-101 mimics reduced, whereas miR-101 inhibitor
increased, luciferase activity in the ZEB2-WT group. Luciferase
activity was not altered by miR-101 mimics or inhibitors in the
ZEB2-MUT group. Data are presented as the mean ± standard deviation
of three independent experiments. *P<0.05,
**P<0.01. SPRY4-IT1, sprouty receptor tyrosine kinase
signalling antagonist 4-intronic transcript 1; miR, microRNA; ZEB,
zinc finger E-box-binding homeoboxes. |
Furthermore, bioinformatics analysis using starBase
identified a binding sequence for miR-101 in the ZEB1 3′UTR
(Fig. 5C). In addition, ZEB2 3′UTR
was also predicted to contain a 6 bp binding site for miR-101
(Fig. 5E). Dual-luciferase assays
showed that the relative luciferase activities were significantly
downregulated in the ZEB1-WT and ZEB2-WT groups by miR-101 mimic
transfection (Fig. 5D and F). In
contrast, the miR-101 inhibitor increased the relative luciferase
activity in the ZEB1-WT and ZEB2-WT groups (Fig. 5D and F). Mutagenesis of the seed
binding sequences completely eliminated the changes induced by
miR-101 mimics and inhibitor (Fig. 5D
and F). These findings confirmed that miR-101 could directly
target ZEB1 and ZEB2 in OS cells and together these data suggest
that SPRY4-IT1 may sponge miR-101 and thus disinhibit the
expression of ZEB1/2. The proposed SPRY4-IT1/miR-101/ZEB axis
represents a novel axis which may modulate tumorigenesis of OS.
SPRY4-IT1 knockdown inhibits cell growth
through upregulation of miR-101 in OS
As MG-63 cells exhibited the highest expression
levels of SPRY4-IT1 in the OS cells assessed, subsequent
experiments on the SPRY4-IT1/miR-101/ZEBs axis was performed in
MG-63 cells. In the present study, the effects of miR-101 inhibitor
on cell growth induced by SPRY4-IT1 knockdown wanted to be
determined. Therefore, shSPRY4-1T1 transfection alone group is an
essential control for this experiment, even though it had already
been performed in Fig. 4. Thus,
this transfection was repeated in Fig. shSPRY4-IT1 transfection
significantly reduced the expression of SPRY4-IT1 and increased
miR-101 levels (Fig. 4, and
6A and B). Although the miR-101
inhibitor had no effect on SPRY4-IT1 expression, transfection of
shSPRY4-IT1 with the inhibitors significantly reversed the miR-101
upregulation induced by SPRY4-IT1 knockdown (Fig. 6A and B). Consistent with Fig. 4E-H, shSPRY4-IT1 reduced the protein
levels of both ZEB1 and ZEB2 (Fig.
6C-D). However, these effects were partially reversed by
co-transfection with miR-101 inhibitors (Fig. 6C and D). Knockdown of SPRY4-IT1
significantly reduced cell growth and co-transfection with miR-101
inhibitor partially reversed the inhibitory effects of shSPRY4-IT1
(Fig. 6E). Similar effects were
also observed in the colony formation assay (Fig. 6F and G) where transfection of
miR-101 inhibitor reversed the inhibitory effects of shSPRY4-IT1 on
the colony formation of OS cells. Finally, an apoptosis assay also
revealed that the increase in apoptosis induced by SPRY4-IT1
knockdown was also reversed by co-transfection with the miR-101
inhibitor (Fig. 6H and I).
Overall, SPRY4-IT1 knockdown inhibited cell growth and promoted
cell apoptosis through upregulation of miR-101. Inhibition of
miR-101 in the shSPRY4-IT1 transfected cells abolished the
anticancer effects of SPRY4-IT1 knockdown. These results suggested
that miR-101 may be a direct downstream effector of SPRY4-IT1.
 | Figure 6SPRY4-IT1 knockdown reduces cell
growth through upregulation of miR-101 in MG-63 cells. (A)
Transfection of shSPRY4-IT1 significantly decreased SPRY4-IT1 mRNA
expression levels, whereas transfection of miR-101 inhibitor did
not affect SPRY4-IT1 mRNA expression levels. (B) SPRY4-IT1
knockdown significantly increased, whereas miR-101 inhibitor
significantly reduced, miR-101 levels. The increase in miR-101
induced by shSPRY4-IT1 was significantly reversed by simultaneous
transfection of miR-101 inhibitors. (C and D) SPRY4-IT1 knockdown
significantly decreased, whereas miR-101 inhibitor increased ZEB1
and ZEB2 protein expression levels. The reduction in ZEB1 and ZEB2
by shSPRY4-IT1 was partially abolished by transfection of miR-101
inhibitor. (E) SPRY4-IT1 knockdown decreased OS cell growth in the
MTT assay, and simultaneous transfection with miR-101 inhibitor
reversed the effects of shSPRY4-IT1 on cell growth. (F and G)
SPRY4-IT1 knockdown significantly decreased, whereas miR-101
inhibitor increased colony formation. Reduced colony formation in
cells transfected with shSPRY4-IT1 was partially abolished by
simultaneous transfection with miR-101 inhibitor. (H and I)
SPRY4-IT1 knockdown significantly increased, whereas miR-101
inhibitor decreased apoptosis. shSPRY4-IT1-induced apoptosis was
partially reversed by simultaneous transfection with miR-101
inhibitor. Data are presented as the mean ± standard deviation of
three independent experiments. *P<0.05,
**P<0.01. SPRY4-IT1, sprouty receptor tyrosine kinase
signalling antagonist 4-intronic transcript 1; miR, microRNA; OS,
osteosarcoma; sh, short hairpin; NC, negative control; OD, optical
density; PI, propidium iodide; ZEB, zinc finger E-box-binding
homeoboxes. |
miR-101 inhibition reverses the
shSPRY4-IT1-mediated suppression of cell migration and
invasion
As miR-101 inhibition in the shSPRY4-IT1 transfected
cells reversed the effects SPRY4-IT1 knockdown on ZEB expression,
colony formation and apoptosis, the effects on migration and
invasion were determined using the co-transfected cells.
Transfection of shSPRY4-IT1 treatment reduced wound closure,
whereas inhibition of miR-101 alone promoted wound closure.
Co-transfection of the miR-101 inhibitor reversed the inhibitory
effects of shSPRY4-IT1 on wound closure (Fig. 7A and B). In the Transwell invasion
assay, SPRY4-IT1 knockdown significantly reduced the number of
invaded cells and inhibition of miR-101 in the SPRY4-IT1 knockdown
cells partially restored the invasive capacity of cells (Fig. 7C and 7D). These data suggest that SPRY4-IT1 may
alter the migratory and invasive capacities of OS cells by
regulating miR-101 expression.
 | Figure 7miR-101 inhibitor reverses the
shSPRY4-IT1-mediated suppression of cell migration and invasion in
MG-63 cells. (A and B) SPRY4-IT1 knockdown significantly delayed
wound closure, whereas miR-101 inhibitor increased wound closure.
The shSPRY4-IT1-induced decrease in migratory rate was partially
reversed by miR-101 inhibitor. (C and D) SPRY4-IT1 knockdown
significantly reduced cell invasion, whereas miR-101 inhibitor
increased cell invasion. miR-101 expression in SPRY4-IT1 knockdown
cells reversed the effects of SPRY4-IT1 knockdown on cell invasion.
(E and F) shSPRY4-IT1 increased E-cadherin protein expression
levels, whereas vimentin, fibronectin, N-cadherin, MMP-2 and MMP-9
levels were decreased by SPRY4-IT1 knockdown, whereas cells
transfected with miR-101 inhibitors exhibited the opposite changes
in protein expression levels of epithelial-mesenchymal
transition-associated proteins. Transfection of miR-101 inhibitor
reversed the effects of shSPRY4-IT1 transfection. Data are
presented as the mean ± standard deviation of three independent
experiments. *P<0.05, **P<0.01.
SPRY4-IT1, sprouty receptor tyrosine kinase signalling antagonist
4-intronic transcript 1; miR, microRNA; sh, short hairpin; NC,
negative control; MMP, matrix metalloproteinase. |
EMT serves a critical role in cell migration and
invasion (26,27), and E-cadherin has been reported to
reduce cell migration in a number of different types of cancer
(26). ZEB1/2 increase migration
and invasion by transcriptionally repressing E-cadherin expression
(28). As shown in Fig. 7E and F, SPRY4-IT1 knockdown
significantly increased the protein expression levels of
E-cadherin. In contrast, transfection of miR-101 inhibitor alone
was sufficient to decrease E-cadherin protein expression levels.
Upregulation of E-cadherin by shSPRY4-IT1 was partially reversed by
co-transfection with the miR-101 inhibitor. The expression levels
of vimentin, fibronectin, N-cadherin, MMP-2 and MMP-9, proteins
which may promote cell migration and invasion, were also detected.
SPRY4-IT1 knockdown reduced the expression of all these proteins,
whereas miR-101 inhibition increased their expression. Inhibition
of miR-101 in the shSPRY4-IT1 cells was sufficient to reverse the
changes in the expression of all these proteins. Based on these
results, it was hypothesized that SPRY4-IT1 promoted cell migration
and invasion by sponging miR-101, which subsequently disinhibited
ZEB1/2, leading to the upregulation of the EMT-associated
proteins.
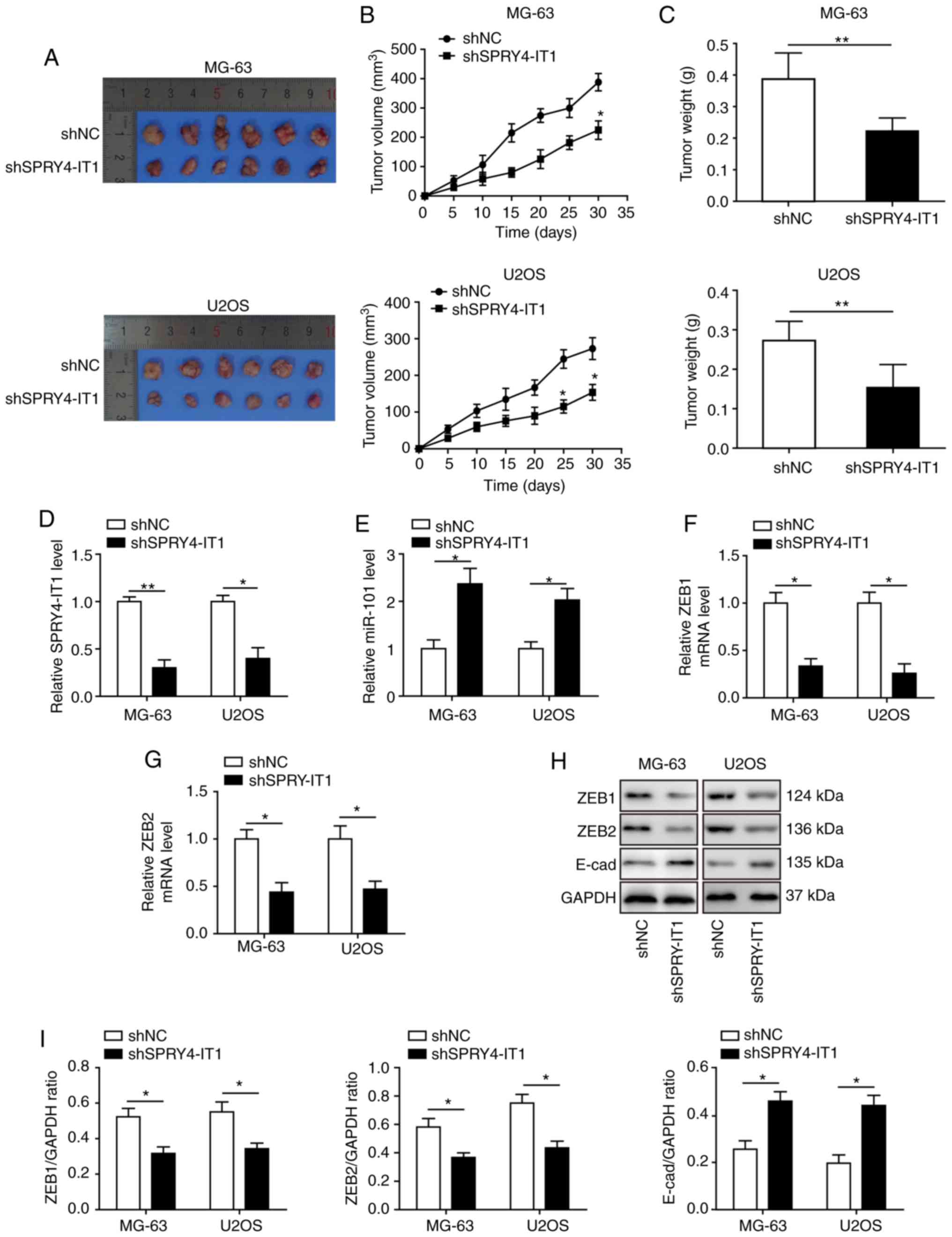
SPRY4-IT1 knockdown decreases tumour
growth in vivo
The anti-tumour function of SPRY4-IT1 knockdown was
evaluated in a xenograft tumour model. MG-63 and U2OS cells stably
transfected with shSPRY4-IT1 or shNC were subcutaneously injected
into nude mice. A total of 30 days after model establishment, the
mice were sacrificed for tumour collection (Fig. 8A). Knockdown of SPRY4-IT1 was
sufficient to reduce tumour growth of both MG-63 and U2OS OS cells
in vivo, as indicated by the significant reduction in tumour
volume and weight (Fig. 8B and C).
Tumours were lysed for qPCR and western blot analysis. SPRY4-IT1
expression levels were significantly lower in the tumours of mice
injected with the shSPRY4-IT1 cells (Fig. 8D), and miR-101 expression was
increased in the tumours (Fig.
8E). ZEB1 and ZEB2 mRNA (Fig. 8F
and G) and protein expression levels (Fig. 8H and I) were significantly reduced
in the tumours of mice injected with the shSPRY4-IT1 cells, and the
expression of E-cadherin, a target gene of ZEB1 and ZEB2, was
significantly increased (Fig. 8H and
I). These results demonstrate that a dysregulated
SPRY4-IT1/miR-101/ZEBs axis may promote tumour growth in
vivo and targeting this axis highlights a potentially novel
approach for treating patients with OS.
Discussion
OS, the most frequent type of malignant bone tumour,
often afflicts children and accounts for 2.4% of all malignancies
and ~20% of all types of primary bone cancer in paediatric patients
(1,2). Despite considerable advances in
treatment and diagnosis, the survival rates of patients with OS
remains poor, and the rate of recurrence is as high as 30–50% for
patients with initial localized disease (29). In the present study, SPRY4-IT1 was
determined to function as a ceRNA which regulated the expression of
the oncogenes ZEB1 and ZEB2 by competing for miR-101 binding in OS
cells. Inhibition of SPRY4-IT1 restored the expression of miR-101,
which then inhibited cell growth, migration and invasion by
regulating the levels of ZEB1 and ZEB2 in OS cells.
Previous studies have highlighted the diagnostic
and/or prognostic values of lncRNAs in various types of cancer
(30,31). Non-coding RNAs, including lncRNAs
and miRNAs, have been frequently reported to be differentially
expressed in the cancer tissues of patients (30,31).
Recently, it was reported that the serum levels of miR-101 were
significantly lower in patients with OS compared with the healthy
controls (32). Therefore, miR-101
may represent a potentially useful biomarker for predicting the
survival of patients with OS. Consistent with the previous study,
miR-101 expression was demonstrated to be significantly
downregulated in three OS cell lines compared with the normal
osteoblast cell line in the present study. Expression of SPRY4-IT1,
which was recently demonstrated to modulate miR-101 in
cholangiocarcinoma (33), was
upregulated in OS cells compared with the normal cells. These
results suggest that a functional interaction between SPRY4-IT1 and
miR-101 may be observed during the pathogenesis of OS. Indeed,
knockdown of SPRY4-IT1 was sufficient to restore miR-101 levels in
OS cells. However, overexpression of miR-101 did not affect
SPRY4-IT1 levels, implying that SPRY4-IT1 was the upstream
modulator of miR-101. Furthermore, in silico analysis
indicated that SPRY4-IT1 may bind to a complementary sequence in
miR-101. This binding was confirmed using a dual-luciferase assay,
which showed that the relative luciferase activity of the
SPRY4-IT1-WT group was reduced in the presence of miR-101 mimics.
Therefore, it was plausible that SPRY4-IT1 sponged miR-101,
resulting in the disruption of miR-101-mediated tumour suppression
in OS.
To test the hypothesis that SPRY4-IT1 acted as a
sponge of miR-101, the effect of SPRY4-IT1 and miR-101 interactions
on cancer cells was further investigated. shSPRY4-IT1 or miR-101
mimic transfection was used to study the functional effects of
SPRY4-IT1 and miR-101, respectively. Knockdown of SPRY4-IT1 alone
was sufficient to decrease cell growth, cause cell cycle arrest and
induce apoptosis in OS cells. Wound healing and Transwell assays
also showed that shSPRY4-IT1 attenuated cell migration and
invasion. This was further confirmed by the upregulation of the
epithelial marker E-cadherin and downregulation of the mesenchymal
markers vimentin, fibronectin, N-cadherin, MMP-9 and MMP-2 when
SPRY4-IT1 was knocked down. Notably, similar anticancer effects
were also observed in cells treated with miR-101 mimics, whereas
transfection of the miR-101 inhibitor resulted in the opposite
outcomes. Through MTT, colony formation, flow cytometry, wound
healing and Transwell invasion assays, the effects on cell growth,
migration, invasion and cell cycle progression induced by SPRY4-IT1
knockdown were partially abolished when miR-101 was simultaneously
inhibited in vitro. These results demonstrate that SPRY4-IT1
sequestered miR-101 in OS and that inhibition of SPRY4-IT1 was
sufficient to impede growth of OS through restoration of miR-101
function. The results of the present study are in agreement with
previous studies which showed that increased SPRY4-IT1 levels could
sponge miR-101 and thus confer oncogenic properties, such as
proliferation and invasion, on colorectal cancer and
cholangiocarcinoma cells (33,34).
Accompanying these findings, both ZEB1 and ZEB2 mRNA
and protein expression levels were reduced by SPRY4-IT1 knockdown
or miR-101 overexpression. The effects of shSPRY4-IT1 on ZEB
expression were also partially attenuated by co-transfection with
the miR-101 inhibitor. Together with the dual-luciferase assays
showing that ZEB1 and ZEB2 were target genes of miR-101, it was
concluded that knockdown of SPRY4-IT1 de-repressed miR-101, leading
to the degradation of ZEB1/2. Previously, the expression levels of
ZEB1 and ZEB2 were shown to be positively associated with
metastatic status in patients with OS (5,35),
whereas knockdown of ZEB1 was sufficient to reduce the invasive
capacity of OS cells (35). Other
published studies also suggested that miR-101 inhibited
proliferation, migration and invasion in osteosarcoma cells by
targeting ROCK1 or ZEB2 (36,37).
However, the present study primarily focused on the
SPRY4-IT1/miR-101/ZEB1 or ZEB2 axis in osteosarcoma progression,
which have not been reported on by others before, to the best of
our knowledge. From a mechanistic perspective, the EMT-modulating
functions of ZEBs may contribute to the anti-OS actions of ZEBs
(38). For example, it has been
demonstrated that ZEB1 can directly bind to the promoter of CDH1,
resulting in the suppression of E-cadherin (product of CDH1) and
thus, subsequent induction of EMT (6). In this regard, miR-101 overexpression
anticancer actions in vitro may be attributed to reversal of
E-cadherin suppression as ZEB1/2 was downregulated.
OS cells which stably expressed SPRY4-IT1 shRNA
exhibited significantly lower growth rates in the OS xenograft
models. Expression levels of miR-101 were increased in the
shSPRY4-IT1 xenograft tumour tissues. Accordingly, the mRNA and
protein expression levels of ZEB1 and ZEB2, the target genes of
miR-101, were reduced in the SPRY4-IT1 knockdown tumour tissues,
and this was accompanied by upregulation of E-cadherin expression
in vivo. Therefore, it was concluded that elevated SPRY4-IT1
contributed to the decrease in miR-101 levels in OS cells. The
interaction between SPRY4-IT1 and miR-101 then disrupted the
inhibition of EMT by upregulating ZEB1 and ZEB2, leading to
dysregulated cell growth, migration and invasion.
Although the data in the present study suggested
that targeting the SPRY4-IT1/miR-101/ZEBs axis could be a promising
approach for the treatment of OS, there were certain limitations.
Firstly, despite the data providing evidence that SPRY4-IT1 could
interact with miR-101, an RNA pull-down assay, which was not
included in the current study, would further strengthen this
conclusion. Secondly, the associations between gene expression
levels (such as SPRY4-IT1 and miR-101) and the clinical
characteristics of patients (such as tumour stage and metastatic
status) have not yet been examined, thus, the clinical significance
of the dysregulation of this signalling axis is unknown. Future
studies on the associations are required and would potentially
provide insight into stage-specific therapy. Finally, the mechanism
by which SPRY4-IT1 was dysregulated was not investigated in the
present study. The upstream regulators of SPRY4-IT1, such as
transcription factors, are more likely to be targeted by small
molecule chemicals, which are clinically more feasible than shRNA
oligos. Future studies on addressing these limitations may
demonstrate the clinical significance of SPRY4-IT1/miR-101.
In conclusion, the present study identified a novel
SPRY4-IT1/miR-101/ZEBs axis underlying the tumorigenesis of OS, to
the best of our knowledge. SPRY4-IT1 may act as a ceRNA to
sequester miR-101 in OS as inhibition of SPRY4-IT1 increased
function of miR-101, which in turn caused degradation of ZEBs,
leading to decreased cell growth, migration and invasion of OS
cells. Additionally, the association between SPRY4-IT1 and miR-101
have not been examined in OS previously and the present study is
the first to have studied two direct targets (ZEB1 and ZEB2) of
miR-101 in OS.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analysed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YH and HG conceived the study. YH, HG, WQY and XWB
collected the data. YH and HG analysed the data. YH, HG, WQY and
XWB performed the experiments. ZHQ and XYC provided the resources
and supervised the study. YH, HG, WQY and XWB wrote the original
draft. ZHQ and XYC reviewed and edited the manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
The animal experiments were approved by the Animal
Ethics Committee at the Third Affiliated Hospital of Sun Yat-sen
University (Guangzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Anninga JK, Gelderblom H, Fiocco M, Kroep
JR, Taminiau AHM, Hogendoorn PC and Egeler RM: Chemotherapeutic
adjuvant treatment for osteosarcoma: Where do we stand? Eur J
Cancer. 47:2431–2445. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang Y, He Z, Duan Y, Wang C, Kamar S,
Shi X, Yang J, Yang J, Zhao N, Han L, et al: Does intensified
chemotherapy increase survival outcomes of osteosarcoma patients? A
meta-analysis. J bone Oncol. 12:54–60. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yang G, Yuan J and Li K: EMT transcription
factors: Implication in osteosarcoma. Med Oncol. 30:6972013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ye X and Weinberg RA:
Epithelial-mesenchymal plasticity: A central regulator of cancer
progression. Trends Cell Biol. 25:675–686. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang X and Lei X: Expression of Zeb1 and
Zeb2 indicates metastasis and unfavorable prognosis in
osteosarcoma. Int J Clin Exp Pathol. 10:611–617. 2017.
|
|
6
|
Eger A, Aigner K, Sonderegger S, Dampier
B, Oehler S, Schreiber M, Berx G, Cano A, Beug H and Foisner R:
DeltaEF1 is a transcriptional repressor of E-cadherin and regulates
epithelial plasticity in breast cancer cells. Oncogene.
24:2375–2385. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Browne G, Emre Sayan A and Tulchinsky E:
ZEB proteins link cell motility with cell cycle control and cell
survival in cancer. Cell Cycle. 9:886–891. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schmitt AM and Chang HY: Long noncoding
RNAs in cancer pathways. Cancer Cell. 29:452–463. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Suzuki HI, Young RA and Sharp PA:
Super-enhancer-mediated RNA processing revealed by integrative
microRNA network analysis. Cell. 168:1000–1014.e15. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gulino R, Forte S, Parenti R, Memeo L and
Gulisano M: MicroRNA and pediatric tumors: Future perspectives.
Acta Histochem. 117:339–354. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fan H, Lu S, Wang S and Zhang S:
Identification of critical genes associated with human osteosarcoma
metastasis based on integrated gene expression profiling. Mol Med
Rep. 20:915–930. 2019.PubMed/NCBI
|
|
12
|
Sampson VB, Yoo S, Kumar A, Vetter NS and
Kolb EA: MicroRNAs and potential targets in osteosarcoma: Review.
Front Pediatr. 3:692015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang Z, Li X, Yang Y, He Z, Qu X and Zhang
Y: Long noncoding RNAs in the progression, metastasis and prognosis
of osteosarcoma. Cell Death Dis. 7:e23892016. View Article : Google Scholar
|
|
14
|
Ergun S and Oztuzcu S: Oncocers:
ceRNA-mediated cross-talk by sponging miRNAs in oncogenic pathways.
Tumor Biol. 36:3129–3136. 2015. View Article : Google Scholar
|
|
15
|
López-Urrutia E, Bustamante Montes LP,
Ladrón de Guevara Cervantes D, Pérez-Plasencia C and Campos-Parra
AD: Crosstalk between long non-coding RNAs, micro-RNAs and mRNAs:
Deciphering molecular mechanisms of master regulators in cancer.
Front Oncol. 9:6692019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yamamura S, Imai-Sumida M, Tanaka Y and
Dahiya R: Interaction and cross-talk between non-coding RNAs. Cell
Mol Life Sci. 75:467–484. 2018. View Article : Google Scholar :
|
|
17
|
Karger S, Liu T, He S, Li Z and Dou P:
Application of long noncoding RNAs in osteosarcoma: Biomarkers and
therapeutic targets. Cell Physiol Biochem. 42:1407–1419. 2017.
View Article : Google Scholar
|
|
18
|
Li Z, Shen J, Chan MTV and Wu WKK: The
long non-coding RNA SPRY4IT1: An emerging player in tumorigenesis
and osteosarcoma. Cell Prolif. 51:e124462018. View Article : Google Scholar
|
|
19
|
Xu J, Ding R and Xu Y: Effects of long
non-coding RNA SPRY4-IT1 on osteosarcoma cell biological behavior.
Am J Transl Res. 8:5330–5337. 2016.
|
|
20
|
Guo F, Cogdell D, Hu L, Yang D, Sood AK,
Xue F and Zhang W: miR-101 suppresses the epithelial-to-mesenchymal
transition by targeting ZEB1 and ZEB2 in ovarian carcinoma. Oncol
Rep. 31:2021–2028. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu D, Li Y, Luo G, Xiao X, Tao D, Wu X,
Wang M, Huang C, Wang L, Zeng F and Jiang G: LncRNA SPRY4-IT1
sponges miR-101-3p to promote proliferation and metastasis of
bladder cancer cells through up-regulating EZH2. Cancer Lett.
388:281–291. 2017. View Article : Google Scholar
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
23
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Han L, Chen W, Xia Y, Song Y, Zhao Z,
Cheng H and Jiang T: miR-101 inhibits the proliferation and
metastasis of lung cancer by targeting zinc finger E-box binding
homeobox 1. Am J Transl Res. 10:1172–1183. 2018.PubMed/NCBI
|
|
25
|
Thomson DW and Dinger ME: Endogenous
microRNA sponges: Evidence and controversy. Nat Rev Genet.
17:272–283. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Son H and Moon A: Epithelial-mesenchymal
transition and cell invasion. Toxicol Res. 26:245–252. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wong TS, Gao W and Chan JY: Transcription
regulation of E-cadherin by zinc finger E-box binding homeobox
proteins in solid tumors. Biomed Res Int. 2014:9215642014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Daw NC, Chou AJ, Jaffe N, Rao BN, Billups
CA, Rodriguez-Galindo C, Meyers PA and Huh WW: Recurrent
osteosarcoma with a single pulmonary metastasis: A
multi-institutional review. Br J Cancer. 112:278–282. 2015.
View Article : Google Scholar :
|
|
30
|
Sánchez Y and Huarte M: Long non-coding
RNAs: Challenges for diagnosis and therapies. Nucleic Acid Ther.
23:15–20. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Faruq O and Vecchione A: microRNA:
Diagnostic Perspective. Front Med (Lausanne). 2:512015.
|
|
32
|
Yao ZS, Li C, Liang D, Jiang XB, Tang JJ,
Ye LQ, Yuan K, Ren H, Yang ZD, Jin DX, et al: Diagnostic and
prognostic implications of serum miR-101 in osteosarcoma. Cancer
Biomark. 22:127–133. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xu Y, Yao Y, Jiang X, Zhong X, Wang Z, Li
C, Kang P, Leng K, Ji D, Li Z, et al: SP1-induced upregulation of
lncRNA SPRY4-IT1 exerts oncogenic properties by scaffolding
EZH2/LSD1/DNMT1 and sponging miR-101-3p in cholangiocarcinoma. J
Exp Clin Cancer Res. 37:812018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jin J, Chu Z, Ma P, Meng Y and Yang Y:
Long non-coding RNA SPRY4-IT1 promotes proliferation and invasion
by acting as a ceRNA of miR-101-3p in colorectal cancer cells.
Tumor Biol. 39:10104283177162502017. View Article : Google Scholar
|
|
35
|
Shen A, Zhang Y, Yang H, Xu R and Huang G:
Overexpression of ZEB1 relates to metastasis and invasion in
osteosarcoma. J Surg Oncol. 105:830–834. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jiang R, Zhang C, Liu G, Gu R and Wu H:
MicroRNA-101 inhibits proliferation, migration and invasion in
osteosarcoma cells by targeting ROCK1. Am J Cancer Res. 7:88–97.
2017.PubMed/NCBI
|
|
37
|
Lin H, Zheng X, Lu T, Gu Y, Zheng C and
Yan H: The proliferation and invasion of osteosarcoma are inhibited
by miR-101 via targetting ZEB2. Biosci Rep. 39:BSR201812832019.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang P, Sun Y and Ma L: ZEB1: At the
crossroads of epithelial-mesenchymal transition, metastasis and
therapy resistance. Cell Cycle. 14:481–487. 2015. View Article : Google Scholar : PubMed/NCBI
|