Introduction
Osteosarcoma is the most common primary osteogenic
malignant tumor in childhood and adolescence with a prevalence of 3
cases per million individuals annually worldwide (1,2). The
5-year survival rate of osteosarcoma patients has improved to
60-70% since 1970s due to the development of advanced radical
treatments and multi-agent chemotherapeutics; however, the survival
rate has remained unaltered over the past several decades (3). Additionally, the 5-year survival rate
of patients with recurrent or metastatic osteosarcoma is only ~20%
(4), and current therapies have
exhibited limited efficacy. Osteosarcoma remains the second highest
cause of cancer-related mortality among children and adolescents
(5,6), underscoring a critical need for the
development of novel therapeutic strategies.
A number of natural plant-derived ingredients have
been shown to exhibit promising antitumor therapeutic proper-ties
and are being evaluated in pre-clinical and clinical experiments
(7-9). Andrographolide has been widely used
for >60 years for its extensive therapeutic properties with
minimal adverse side-effects (10,11).
Andrographolide exhibits significant cytotoxic effects against
various cancer cell lines by inhibiting cell cycle progression,
reducing cell invasion and inducing apoptosis (12-14).
It has previously been reported that andrographolide treatment
results in the apoptosis of human leukemic cells through the
mitochondrial-mediated pathway (15). It has previously been demonstrated
that andrographolide inhibits the proliferation of prostate cancer
cells by inducing G2/M and G1 phase arrest, and initiating
caspase-8-mediated mitochondrial apoptosis (16). However, the effect of
andrographolide on osteosarcoma and the detailed underlying
molecular mechanisms have not yet been elucidated.
Apoptosis, also known as type I programmed cell
death, is a very orderly process during which the genome of the
cell is broken down, the cell is fragmented into smaller sections,
and the debris are consumed by nearby cells (phagocytes) that
'clean up' the cell fragments (17). Apoptosis serves a significant role
in the chemotherapy of various types of cancer (18). Reactive oxygen species (ROS),
active forms of oxygen, generated as by-products of cellular
metabolism, possess toxic effects on a variety of molecular targets
including lipids, DNA and proteins in most cells (19). Notably, cancer cells have been
found to be more sensitive to damage by ROS products since they
exhibit increased oxidative stress (20). Accumulating evidence indicates that
ROS are involved in various signaling pathways and serve a key role
in inducing cell apoptosis (21-24).
Excessive amounts of ROS can affect multiple signaling pathways,
such as MAPK family members, including JNK, p38 and ERK (25). JNK is a stress-activated protein
kinase of the MAPK family, and serves a critical role in a number
of cellular events, including apoptosis (26). Therefore, targeting ROS/JNK may be
an effective method for treating cancer.
The detailed mechanisms responsible for the
antitumor effects of andrographolide remain uncertain. It has been
shown that andrographolide can induce cytotoxicity and cell cycle
arrest in breast cancer, prostate cancer, laryngeal cancer and
gastric cancer by upregulating ROS production, leading to
programmed cell death (27-30).
Andrographolide has also been found to induce human hepatoma cell
death through JNK activation (31). Moreover, a recent study reported
that the ROS/JNK pathway was involved in the
andrographolide-induced cell death of HeLa cells (32). However, whether andrographolide can
induce apoptosis of osteosarcoma cells, and whether the antitumor
effect is mediated via the ROS/JNK pathway remains unknown, to the
best of our knowledge.
In the present study, the antitumor effects of
andrographolide against osteosarcoma cells in vitro and
in vivo were demonstrated. The possible molecular mechanisms
were further explored, and it was demonstrated that andrographolide
induced G2/M phase arrest and the apoptosis of osteosarcoma cells
via the ROS/JNK signaling pathway.
Materials and methods
Reagents and antibodies
Andrographolide (>98%) was purchased from Selleck
Chemicals and dissolved in DMSO (Sigma-Aldrich; Merck KGaA) at a
concentration of 100 mM. N-Acetyl-L-cysteine (NAC) and SP600125
(SP) were purchased from Sigma-Aldrich; Merck KGaA. The molecular
formula of andrographolide is C20H30O5 and its molecular weight is
350.45. FBS, DMEM, RPMI-1640 medium, penicillin, streptomycin, PBS
and 0.25% trypsin were obtained from Gibco; Thermo Fisher
Scientific, Inc. The following anti-bodies were used for western
blot analysis: Poly(ADP-ribose) polymerase (PARP, cat. no. 9542),
cleaved caspase-3 (cat. no. 9664), cleaved caspase-8 (cat. no.
9496), cleaved caspase-9 (cat. no. 9505), phospho-JNK (cat. no.
4668), JNK (cat. no. 9252) and GAPDH (cat. no. 5174). These were
obtained from Cell Signaling Technology, Inc.
Cells and cell culture
The osteosarcoma cell lines, HOS, U2OS, SAOS-2 and
MG-63, were purchased from the Shanghai Institute of Cell Biology,
Chinese Academy of Sciences (Shanghai, China). The HOS, SAOS-2 and
MG-63 cells were cultured in DMEM (Gibco; Thermo Fisher Scientific,
Inc.) and the U2OS cells were cultured in RPMI-1640 medium (Gibco;
Thermo Fisher Scientific, Inc.). A total of 6 patients with
osteosarcoma from the Musculoskeletal Tumor Center, Department of
Orthopedics, The Second Affiliated Hospital of Zhejiang University
School of Medicine were included in the present study. The cohort
included four males and two females, ranging with a median age of
20 (age range, 12-28). Tumor specimens from osteosarcoma patients
were mechanically disaggregated using razor blades at 37°C for 2 h
in DMEM with collagenase type IV (2 mg/ml), DNase (0.1 mg/ml),
hyaluronidase (0.1 mg/ml) and BSA (2 mg/ml) (all from
Sigma-Aldrich; Merck KGaA). Cell suspensions were passed through
70-µm filters to remove aggregates and then cultured in
DMEM. All media were supplemented with 10% FBS, 100 U/ml penicillin
and 100 µg/ml streptomycin. Cells were maintained at 37°C in
a 5% CO2 incubator.
Written informed consent was obtained from each
patient or their legal guardians where required and the study was
approved by the Human Research Ethics Committees of the Second
Affiliated Hospital, School of Medicine, Zhejiang University
(Hangzhou, China).
Cell viability assay
Briefly, HOS, U2OS, SAOS-2 and MG-63 cells were
seeded in 96-well tissue culture plates for one day at a density of
4×103 cells/well. Following treatment with various
concentrations of andrographolide (0, 20, 40 and 80 µM) for
12, 24, 48 and 96 h, cell viability was evaluated using an MTS kit
(cellTiter96AQ; Promega Corporation) according to the
manufacturer's protocol. The optical density value was measured on
an MR7000 microplate reader (Dynatech) at 490 nm.
Colony formation assay
Colony formation assays were performed to assess the
effect of andrographolide on osteosarcoma cell clonal ability.
Cells were seeded at 100/well in 6-well plates and treated with
various concentrations of andrographolide (0, 1.25, 2.5, 5 and 10
µM) for 2 weeks until visible colonies were observed. The
cells were then washed with PBS twice and fixed with 4%
paraformaldehyde at room temperature for 15 min. After washing
twice again, the cells were stained with 0.1% crystal violet at
room temperature for 15 min. Finally, the plates were imaged, and
the colonies were counted under a light microscope (magnification,
×4).
Morphological apoptosis
To determine the apoptotic morphological changes,
cells were treated with 40 µM andrographolide for 24 h and
then stained with Hoechst 33258 (Beyotime Institute of
Biotechnology) for 10 min at room temperature. After washing twice,
cell morphology was observed using a fluorescence microscope
(magnification, ×10; Olympus Corporation) to determine chromatin
condensation.
Cell cycle analysis
Cells were plated at 3×105/well in 6-well plates and
treated with andrographolide (0, 20, 40 and 80 µM) for 24 h.
The cells were then trypsinized and fixed with cold 75% ethanol at
−20°C overnight. The cells were stained with buffer containing
propidium iodide and RNase (BD Biosciences) at room temperature for
15 min and analyzed using a FACSCalibur (BD Biosciences) flow
cytometer and ModFit LT software (version 3.2; Verity
Software).
Apoptosis analysis
A total of 2.5×105 cells/well were plated
in 6-well plates. Following treatment with andrographolide (0-80
µM) for 24 h, the cells were incubated with Annexin V-PE and
7-AAD for 15 min at 37°C in the dark. Subsequently, cells were
washed and resuspended in 300 µl PB, assessed using a flow
cytometer (FACSCalibur, BD Biosciences) and the data were analyzed
using CellQuest™ Pro version 5.1 (Becton, Dickinson and
company).
Western blot analysis
HOS and U2OS cells were seeded in 60-mm dishes.
Following treatment with andrographolide for 24 h, the cells were
harvested, washed and resuspended in RIPA lysis buffer (Beyotime
Institute of Biotechnology) with phenylmethanesulfonyl fluoride for
30 min on ice. The lysates were centrifuged at 4°C and 13,000 × g
for 15 min. The protein concentrations were determined using a
bicinchoninic acid protein assay kit (Beyotime Institute
Biotechnology). Equal quantities of protein (40 µg) were
loaded on an SDS gel (10% or 12%), resolved using SDS-PAGE and
transferred to PVDF membranes (EMD Millipore). Membranes were
blocked with TBS-Tween (TBST) containing 5% skimmed milk, and then
incubated overnight at 4°C with the primary antibodies.
Subsequently, the membrane was washed with TBST, and incubated for
1 h at room temperature with a horseradish peroxidase-conjugated
goat secondary anti-rabbit IgG (Boster Biological Technology; cat.
no. BA1054). The membranes were incubated with an enhanced
chemiluminescence kit (EMD Millipore), and the protein bands were
then visualized using a ChemiDoc imaging system (Bio-Rad
Laboratories, Inc.). Densitometry analysis was performed using
ImageJ (Version 1.46; National Institutes of Health)
Measurement of mitochondrial membrane
potential (MMP)
JC-1 fluorescent probe (Beyotime Institute of
Biotechnology) was used to measure MMP. Briefly, cells were plated
at a density of 2.5×105/well in 6-well plates and
treated with andrographolide (0-80 µM) for 24 h. The cells
were then collected and incubated with JC-1 for 20 min at 37°C.
Subsequently, the stained cells were washed and analyzed using a
flow cytometry.
Measurement of ROS generation
ROS assays using DCFH-DA (Beyotime Institute of
Biotechnology) were used to measure intracellular ROS. Briefly, the
cells were treated with andrographolide (0-80 µM) for 12 h
prior to incubation with DCFH-DA (10 μM) for 30 min at 37°C.
Subsequently, the stained cells were detected using fluorescence
microscopy (magnification, ×10) and flow cytometry. ROS production
in vivo was determined using dihydroethidium (DHE) as
described previously (33).
Briefly, 24 h before sacrifice, each mouse received a 200 µl
intravenous injection of DHE at 25 mg/kg.
Animal experimental design
Female Balb/c-nu mice (5 weeks old, n=15) were
purchased from the Shanghai Laboratory Animal Center of Chinese
Academy of Sciences. HOS cells were transfected with luciferase
(HOS-Luc) using lentivirus for in vivo imaging. For
lentiviral infection, HOS cells were incu-bated with lentiviral
luciferase particles [pLV-Puro-CMV (Luc); Hanbio Biotechnology,
Co., Ltd.] in the presence of 5 µg/ml polybrene (Hanbio
Biotechnology, Co., Ltd.). After 12 h, the infection medium was
discarded, and the cells were cultured with fresh medium for 3 days
before being screened using puromycin (4 µg/ml;
Sigma-Aldrich; Merck KGaA) and then passaged for use in subsequent
experiments. Tumor xenografts were established by a subcutaneous
injection of 5×106 HOS-Luc cells suspended in 100 μl PBS
into the flanks of mice. Tumor size was monitored every two days
and calculated as follows: Length × width2/2. When the
tumors had grown for 10 days, reaching ~50 mm3, the mice
were randomly divided into three groups, each of which contained
five mice. The three groups of mice received intraperitoneal
injections with 100 µl 1% DMSO in PBS, or a low or high
concentration of andrographolide every two days, respectively.
Tumor sizes and body weights were measured every two days to
observe dynamic variations. Tumor inhibition rate was calculated as
follows: (1 - volume in experimental group/volume in control group)
× 100%. After two weeks of therapy, all mice were euthanized, and
tumors were excised and fixed in 10% neutral-buffered formalin for
24 h at 4°C for further analysis. The animal experiments were
approved by the Animal Care and Use Committee of Zhejiang
University, China.
Tumor histology
Tumor specimens were cut into serial sections
(3-µm-thick) after fixing with formalin and embedding in
paraffin. The slides were hydrated using a gradient of ethanol
solutions (100%, 100%, 95%, 85% and 75% ethanol, followed by PBS; 5
min each). Subsequently, the slides were stained with hematoxylin
at room temperature for 15 min, and then immersed in 1%
hydrochloric acid in 75% ethanol for 30 sec. The slides were then
washed and stained with eosin at room temperature for 5 min.
Following dehydration, the slides were immersed in xylene.
Immunohistochemical analysis
Formalin-fixed and paraffin-embedded tumor specimens
were cut into serial sections of 3 µm thickness. Slides were
treated with 3% H2O2 for 15 min to block
endogenous peroxidase activity. Antigen retrieval was performed by
boiling in sodium citrate buffer (pH 6.0) for 10 min. The slides
were then incubated with 10% goat serum (cat. no. SL038, Beijing
Solarbio Science & Technology Co., Ltd.) at room temperature
for 30 min. Slides were incubated with cleaved caspase-3 (1:50) and
phospho-JNK (1:50) antibodies at 4°C overnight and then washed with
PBS three times. Subsequently, the slides were treated with a
biotin-labeled secondary antibody (1:50) at room temperature for 30
min prior to incubation with DAB (Sigma-Aldrich; Merck KGaA) at
room temperature for ~30 sec. Then the slides were stained with
hematoxylin at room temperature for 15 min and the images were
obtained using a light microscope (magnification, ×20).
Statistical analysis
All experiments were performed in a minimum of
triplicates. Quantitative data are expressed as the mean ± standard
deviation, and the significance of the differences between
treatment groups were determined using one-way ANOVA with a post
hoc Dunnett's test or a Student's t-test. P<0.05 was considered
to indicate a statistically significant difference.
Results
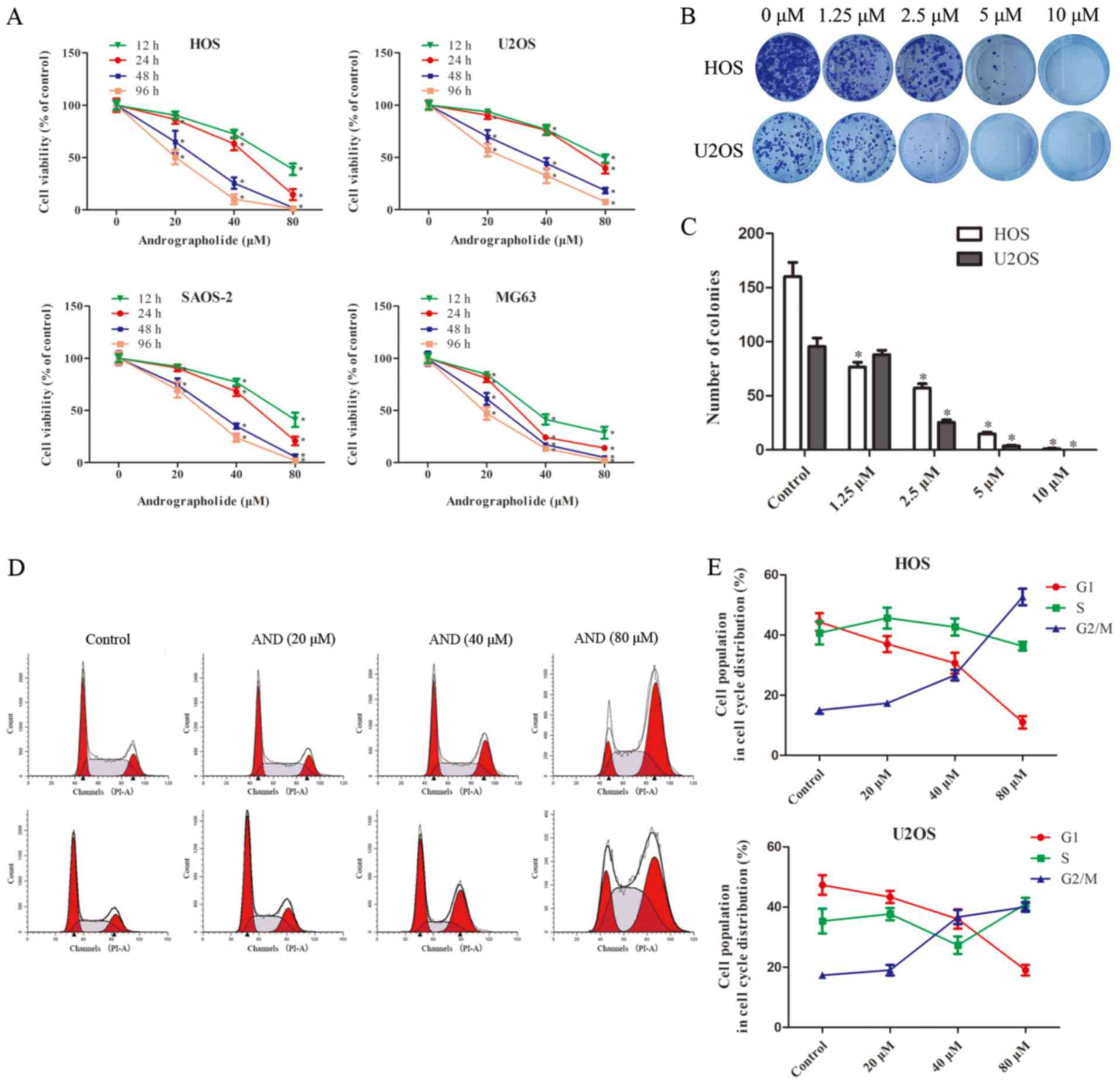
Andrographolide inhibit s the
proliferation of osteosarcoma cells in a time- and dose-dependent
manner
To evaluate theeffect of andrographolide on the
growth of osteosarcoma cells, HOS, U2OS, SAOS-2 and MG-63 cell
lines were treated with various concentrations of andrographolide
(0-80 µM) for 12, 24, 48 and 96 h (Fig. 1A). Based on the results of MTS
assay, the viability of the tumor cells was significantly reduced
by andrographolide in a time- and dose-dependent manner, and the
IC50 values for 24 h of treatment were 50.84 µM
for HOS cells, 68.42 µM for U2OS cells, 55.27 µM for
SAOS-2 cells and 30.87 µM for MG-63 cells. Although MG-63
seemed to be the most sensitive cell line, HOS and U2OS were
selected for further studies as HOS cells are more suitable for use
in animal studies (23,34) and U2OS cells are more stable for
use in the in vitro experiments used in the present study.
Colony formation assays showed that fewer colonies were formed
following andrographolide treatment (Fig. 1B and C).
Andrographolide treatment results in G2/M
phase cell cycle arrest
To determine whether andrographolide reduced cell
viability by inducing cell cycle arrest, cell cycle distribution of
the cells treated with andrographolide was assessed. Exposure to
andrographolide resulted in a marked increase in the proportion of
cells in the G2/M phase, and a corresponding decrease in the
proportion of cells in the G1 and S phases in both the HOS and U2OS
cells (Fig. 1D and E). The
percentage of cells in the G2 phase increased from 15.1 to 51.6% in
the HOS, and from 17.2 to 39.6% in the U2OS cells.
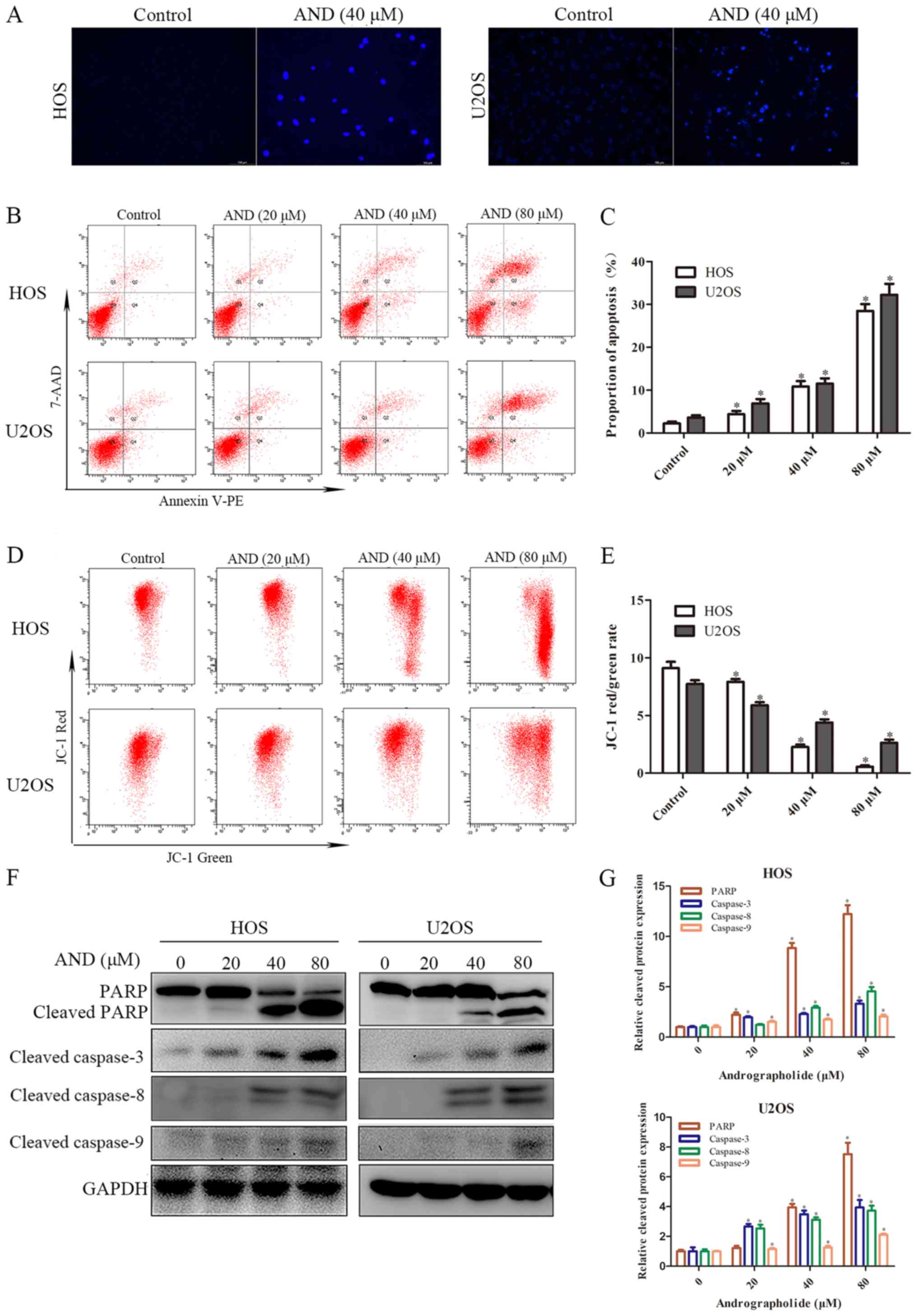
Andrographolide increases
mitochondrial-mediated apoptosis of osteosarcoma cells
To determine whether apoptosis was responsible for
the reduced cell growth induced by andrographolide, Hoechst
staining and flow cytometry assays were performed. The results
showed that apoptotic chromatin condensation was clearly observed
in the cells treated with andrographolide (Fig. 2A). To quantify apoptosis, tumor
cells treated with the indicated concentrations of andrographolide
were stained with Annexin V-PE/7-AAD. As shown in Fig. 2B and C, the proportion of apoptotic
cells was negligible for the control cells, whereas exposure of the
cells to andrographolide for 24 h resulted in a dose-dependent
increase in the proportion of both early and late apoptotic cells.
The effect of andrographolide on the mitochondria was also
determined. MMP was shown to decrease significantly in the presence
of andrographolide (Fig. 2D and
E). Subsequently, the expression of a downstream apoptotic
protein was measured. As shown in Fig.
2F and G, andrographolide significantly induced caspase-3, -8
and -9 activation, and resulted in PARP cleavage. Overall, these
results suggest that andrographolide induces mitochondrial-mediated
apoptosis.
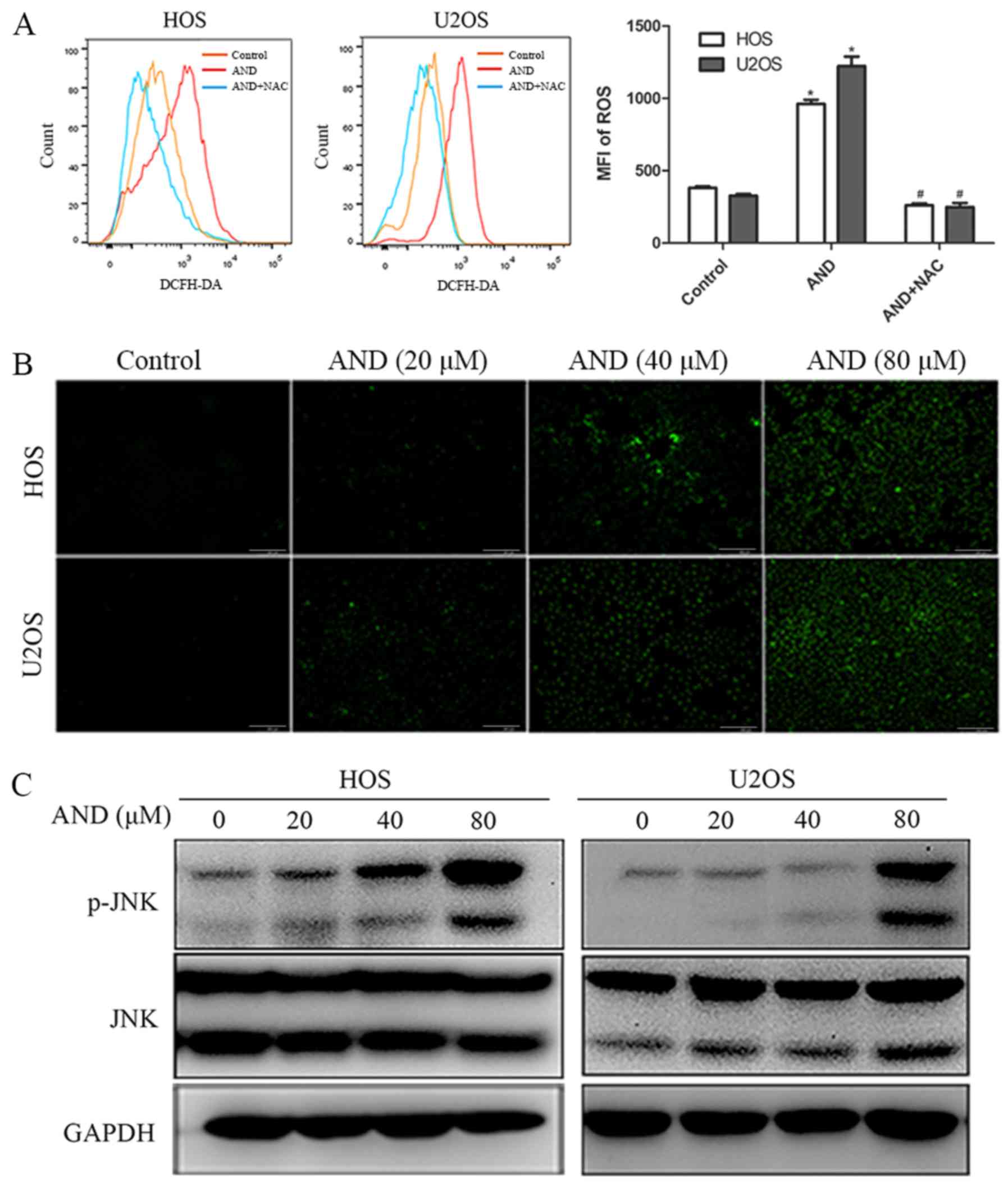
Andrographolide induces apoptosis by
promoting JNK activation
The role of JNK in andrographolide-induced apoptosis
was investigated. The results revealed that phosphorylated JNK
expression was increased by andrographolide in both the HOS and
U2OS cells (Fig. 3C). To determine
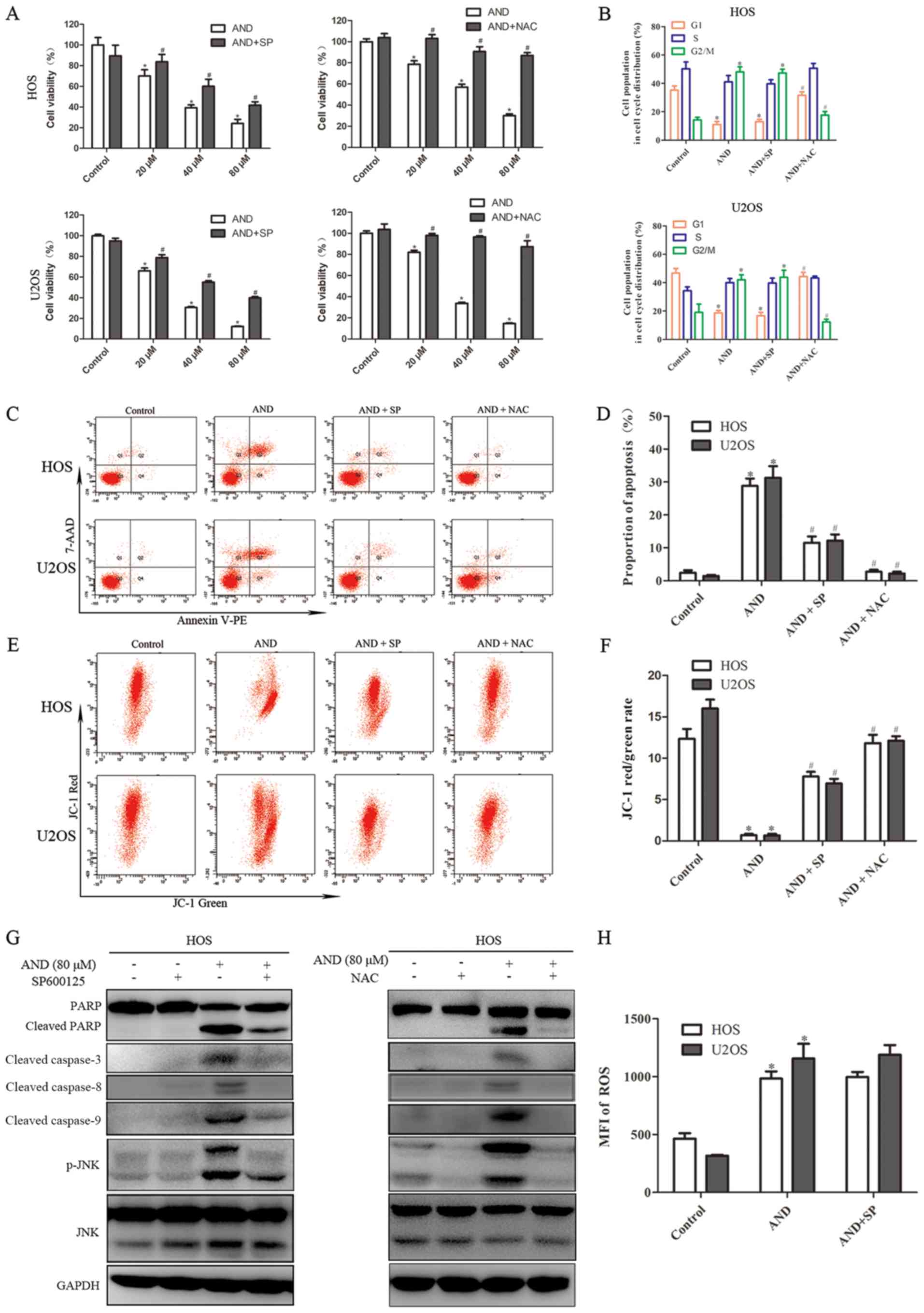
whether JNK activation contributes to andrographolide-induced
apoptosis and G2/M phase arrest, SP600125 (SP), a specific JNK
inhibitor, was used to block JNK phosphorylation. The MTS assays
showed that andrographolide-induced cytotoxicity against
osteosarcoma cells was reduced by SP (Fig. 4A). As shown by flow cytometry, SP
also inhibited andrographolide-induced apoptosis and reduced
mitochondrial depolarization (Fig.
4C-F). SP blocked JNK activation and inhibited the accumulation
of apoptosis-related proteins (Fig.
4G). However, SP failed to reverse G2/M phase cell cycle arrest
caused by andrographolide (Fig.
4B). These results suggest that JNK activation is required for
andrographolide-induced apoptosis, but was not involved in cell
cycle arrest.
Andrographolide promotes ROS generation,
which results in JNK activation and initiates
andrographolide-induced apoptosis and G2/M phase cell cycle
arrest
ROS have been reported to serve a significant role
in mediating apoptosis and cell cycle arrest (35,36),
and also to promote the sustained activation of JNK (37), thus, the ROS levels were detected.
ROS production was significantly increased by andrographolide
treatment and the increased ROS production was completely blocked
by NAC, an ROS scavenger (Fig.
3A). As shown in Fig. 3B, the
results further revealed that ROS production was initiated by 20
µM andrographolide and was significantly increased when
cells were treated with 80 µM. MTS assays showed that NAC
reduced the cell death caused by andrographolide (Fig. 4A). In addition, in contrast to the
JNK inhibitor, NAC was found to have a notably more potent effect
on reducing apoptosis and attenuating the decrease in MMP levels
induced by andrographolide (Fig.
4C-F). Furthermore, NAC was demonstrated to completely inhibit
andrographolide-induced increase in expression of
apoptosis-associated proteins (Fig.
4G), and restore the andrographolide-induced increase in the
proportion of cells in the G2/M phase (Fig. 4B). NAC significantly blocked JNK
activation, and the JNK inhibitor did not exert a notable effect on
ROS suppression (Fig. 4G and H).
Together, these results suggest that ROS generation results in JNK
phosphorylation and initiates andrographolide-induced apoptosis and
cell cycle arrest.
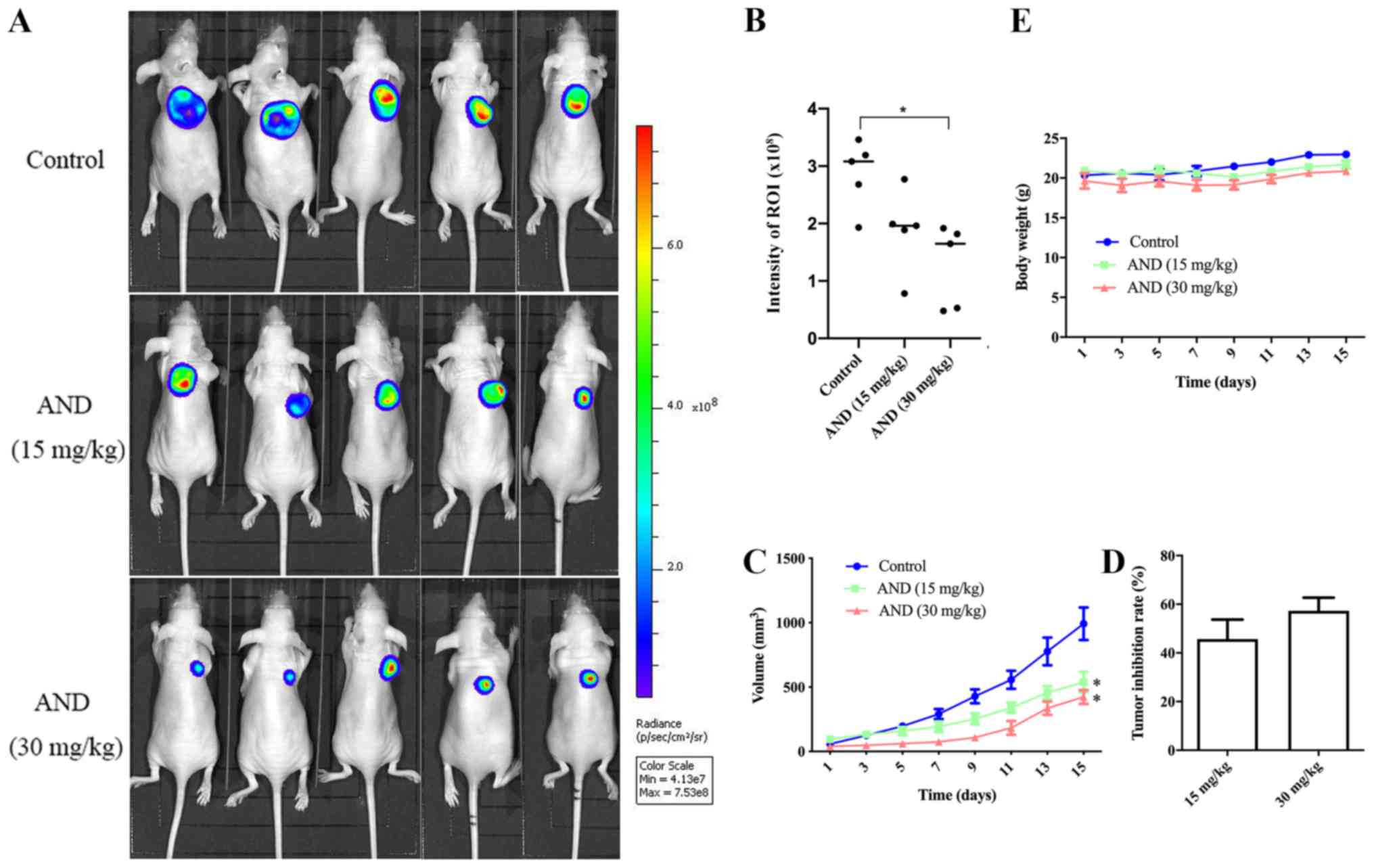
Andrographolide inhibits the growth of
osteosarcoma in vivo
To examine the in vivo effects of
andrographolide on osteosarcoma growth, equal numbers of HOS-Luc
cells were injected subcutaneously into nude mice. Once the tumors
reached ~50 mm3, the mice were treated with an
intra-peritoneal injection of control or andrographolide treatment
every other day. Compared with the control, tumor growth in the
groups receiving 15 and 30 mg/kg was decreased by 45.6 and 57.4%,
respectively, after 14 days of treatment (Fig. 5A-D). There was no difference in
body weight according to the data (Fig. 5E). Tumor tissue was harvested from
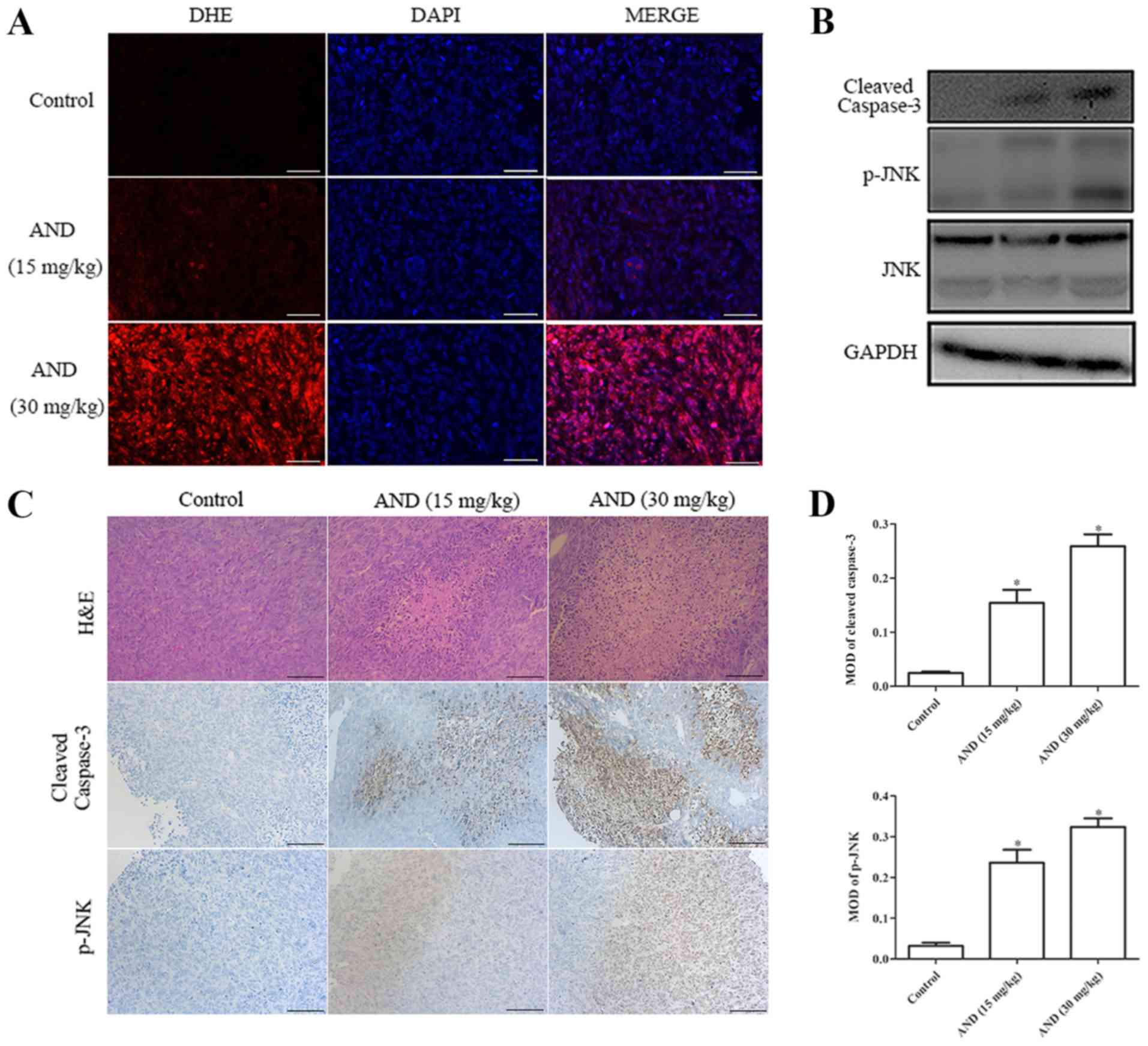
the control- and andrographolide-treated mice for further analysis.
Upregulated levels of ROS were detected in tumor tissue from mice
treated with andrographolide (Fig.
6A). Hematoxylin and eosin staining showed that the proportion
of dead cells in the andrographolide-treated tumor tissue was
higher (Fig. 6C). Both
immunohistochemistry and western blot analysis showed that the
expression levels of cleaved caspase-3 and phosphorylated JNK were
increased by andrographolide treatment (Fig. 6B-D). All these data suggest that
andrographolide inhibits the growth of osteosarcoma in
vivo.
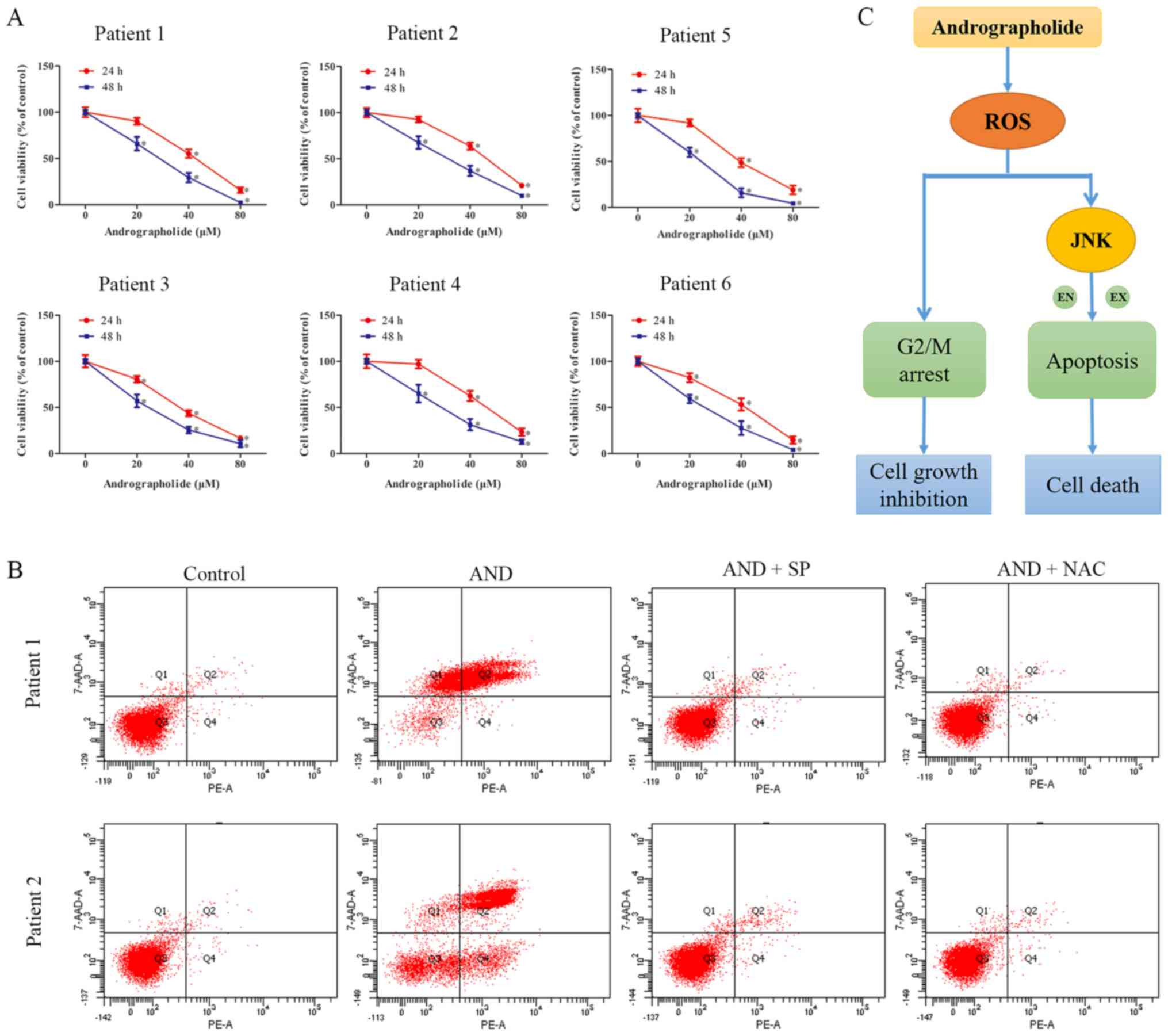
Andrographolide exerts similar antitumor
effects on primary osteosarcoma cells as it does on cell lines
Primary osteo-sarcoma cells were obtained from six
patients who suffered from osteosarcoma and were used to assess the
effects of andrographolide on clinical specimen. The
characteristics of the patients from which specimen were obtained
are presented in Table SI.
Andrographolide significantly inhibited the proliferation of all
primary tumor cells (Fig. 7A).
Flow cytometry analysis showed there was an increase in the number
of apoptotic cells following andrographolide treatment, similar to
the results obtained with the cell lines (Fig. 7B). Thus, these results suggest that
andrographolide exerts similar effects on primary cells as it does
on immortalized cell lines.
Discussion
The 5-year survival rate for osteosarcoma patients
has increased to >60% since 1970s (38). Despite the fact that the prognosis
of localized osteosarcoma has significantly improved due to
advances in surgery and multiagent chemotherapy, the long-term
survival of patients has remained unaltered for several decades
(38). Thus, novel treatments are
required to improve the outcomes of patients with osteosarcoma.
Andrographolide has been reported to be a promising therapeutic for
treatment of multiple types of cancer, with potent antitumor
effects and minimal toxicity (11,29,39-42).
However, little is known about the effects and biochemical
mechanisms of action of andrographolide in osteosarcoma. In the
present study, the results demonstrated that andrographolide
effectively inhibited the growth of osteosarcoma cells in
vitro and in vivo through cell cycle arrest at the G2/M
phase and increasing apoptosis, and that these effects were
mediated via the ROS/JNK signaling pathway.
Apoptosis, an energy-dependent genetically
programmed cell death mechanism, is a major route for the
elimination of cancer. Apoptosis can be induced through either the
extrinsic or the intrinsic pathway. Caspase-dependent apoptosis is
the major form of controlled cell death in cancer cells. The
results of the present study indicated that andrographolide induced
apoptosis by activating both extrinsic and intrinsic pathways. In
the present study, andrographolide treatment resulted in the
cleavage of caspase-3, -8 and -9, confirming the involvement of
caspase-dependent apoptosis in andrographolide-induced cytotoxicity
against osteosarcoma cells.
ROS are unstable molecules which contain oxygen, and
are predominantly generated by enzymatic activity and organelles,
such as the mitochondria, endoplasmic reticulum and peroxisomes
(43). ROS can function as key
indicators that serve a prominent role in the mediation of both
cell survival and death following exposure to various stimuli,
including chemotherapeutic agents, radiation, senescence and host
defense (43,44). The interaction of ROS with proteins
that are involved in cancer generation, survival and metastasis has
been previously demonstrated (45). The presence of ROS has been found
to increase cell stress and damage, which result in cell death, via
a mechanism known as oxidative stress (46). Moreover, ROS can also induce
temporary or permanent cell cycle arrest, and act as signaling
molecules that lead to enhanced cytotoxicity and apoptosis by
increasing oxidative stress (47,48).
Thus, intracellular ROS levels have become a key indicator for a
number of chemotherapeutic agents in the induction of cancer cell
apoptosis (49-51). In the present study,
andrographolide significantly increased the intracellular levels of
ROS. Furthermore, the andrographolide-induced cell death, increase
in apoptosis, loss of MMP and the increase in cleaved PARP and
caspase levels were significantly reversed by an ROS scavenger.
Additionally, increased levels of ROS production were also detected
following andrographolide treatment in vivo. All these data
highlight the critical role of ROS in andrographolide-induced
cytotoxicity against osteosarcoma cells.
The activation of JNKs is mediated by several
stimuli such as heat shock, Fas ligation, oxidative stress and
chemotherapeutic agents (52).
Oxidative stress strongly associated with elevated levels of ROS.
JNK activation can be induced by both exogenous and endogenous ROS
(53). In addition, JNK activation
serves a pivotal role in the induction of apoptotic cells (54). In the present study, it was
demonstrated that treatment with andrographolide significantly
increased the expression levels of phosphorylated JNK in
osteosarcoma cells. Furthermore, the inhibition of JNK activation
attenuated andrographolide-induced cleavage of PARP, caspase-3, -8
and -9, and cell apoptosis. However, the JNK inhibitor failed to
block G2/M phase arrest, suggesting that andrographolide-induced
cell cycle arrest was modulated by ROS, but not by JNK.
Specifically, JNK phosphorylation was significantly inhibited by
ROS scavenger, whereas ROS generation was minimally attenuated by
the JNK inhibitor, suggesting that ROS activity is a proximal event
of JNK. Based on these results, it can be concluded that
andrographolide induced the apoptosis of osteosarcoma cells through
the ROS/JNK signaling pathway, and that ROS serves a significant
role in the modulation of andrographolide-induced G2/M phase cell
cycle arrest.
In conclusion, the present study showed that
andrographolide inhibited the growth of osteosarcoma cells by
causing G2/M phase cell cycle arrest and inducing apoptosis via the
ROS/JNK signaling pathway (Fig.
7C). In the animal experiments, andrographolide was shown to
exhibit significant antitumor activity with minimal toxicity. To
the best of our knowledge, the present study is the first study to
demonstrate the antitumor effects and mechanisms of andrographolide
against osteosarcoma. However, there are some limitations. Despite
significant tumor growth inhibition in the andrographolide-treated
group in the animal study, andrographolide failed to completely
prevent tumor growth. Further studies are required to explore the
combined therapy of andrographolide with other established
treatments to enhance the antitumor effects. Additionally, only six
clinical specimens from patients with osteosarcoma were used to
preliminarily show the potential antitumor effects of
andrographolide in primary cells. Larger cohorts are required in
future studies to confirm these results. The results of the present
study demonstrate that andrographolide may be a promising
therapeutic agent for treatment of osteosarcoma and the findings of
the present study may contribute to an improved understanding of
the benefits and clinical applications of andrographolide
therapy.
Supplementary Data
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China (grant no. 81872173), the Medical and
Health Science and Technology Plan of the Department of Health of
Zhejiang Province (grant nos. WKJ-ZJ-1821 and 2018KY405), Zhejiang
Provincial Natural Science Foundation of China (grant nos.
LY20H160018, LY19H160045 and LQ20H060006).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
PL and SW designed the study. SW, HL, SC and TC
performed the experiments. YY assisted with the animal experiments.
PL, ZW and ZY analyzed the data. PL and SW wrote this manuscript.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Specimen collection was approved by the Human
Research Ethics Committees of the Second Affiliated Hospital,
School of Medicine, Zhejiang University (Hangzhou, China). Written
informed consent was obtained from each patient. The animal
experiments were approved by the Animal Care and Use Committee of
Zhejiang University (Hangzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Agarwal M, Anchan C, Shah M, Puri A and
Pai S: Limb salvage surgery for osteosarcoma: Effective low-cost
treatment. Clin Orthop Relat Res. 459:82–91. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Biermann JS, Adkins DR, Benjamin RS,
Brigman B, Chow W, Conrad EU III, Frassica DA, Frassica FJ, George
S, Hande KR, et al: National Comprehensive Cancer Network Bone
Cancer Panel: Bone cancer. J Natl Compr Canc Netw. 8:688–712. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ando K, Heymann MF, Stresing V, Mori K,
Rédini F and Heymann D: Current therapeutic strategies and novel
approaches in osteosarcoma. Cancers (Basel). 5:591–616. 2013.
View Article : Google Scholar
|
|
4
|
Harrison DJ, Geller DS, Gill JD, Lewis VO
and Gorlick R: Current and future therapeutic approaches for
osteosarcoma. Expert Rev Anticancer Ther. 18:39–50. 2018.
View Article : Google Scholar
|
|
5
|
Bielack S, Carrle D and Casali PG; ESMO
Guidelines Working Group: Osteosarcoma: ESMO clinical
recommendations for diagnosis, treatment and follow-up. Ann Oncol.
20(Suppl 4): 137–139. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Durfee RA, Mohammed M and Luu HH: Review
of Osteosarcoma and Current Management. Rheumatol Ther. 3:221–243.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cassileth BR and Deng G: Complementary and
alternative therapies for cancer. Oncologist. 9:80–89. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kaur R, Kapoor K and Kaur H: Plants as a
source of anticancer agents. J Nat Prod Plant Resour. 1:119–124.
2011.
|
|
9
|
Mayzlish-Gati E, Fridlender M, Nallathambi
R, Selvaraj G, Nadarajan S and Koltai H: Review on Anti-Cancer
Activity in Wild Plants of the Middle East. Curr Med Chem.
25:4656–4670. 2018. View Article : Google Scholar
|
|
10
|
Lim JCW, Chan TK, Ng DS, Sagineedu SR,
Stanslas J and Wong WS: Andrographolide and its analogues:
Versatile bioactive molecules for combating inflammation and
cancer. Clin Exp Pharmacol Physiol. 39:300–310. 2012. View Article : Google Scholar
|
|
11
|
Islam MT, Ali ES, Uddin SJ, Islam MA, Shaw
S, Khan IN, Saravi SSS, Ahmad S, Rehman S, Gupta VK, et al:
Andrographolide, a diterpene lactone from Andrographis paniculata
and its therapeutic promises in cancer. Cancer Lett. 420:129–145.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pratheeshkumar P, Sheeja K and Kuttan G:
Andrographolide induces apoptosis in B16F-10 melanoma cells by
inhibiting NF-κ B-mediated bcl-2 activation and modulating
p53-induced caspase-3 gene expression. Immunopharmacol
Immunotoxicol. 34:143–151. 2012. View Article : Google Scholar
|
|
13
|
Huang K, Chen Y, Zhang R, Wu Y, Ma Y, Fang
X and Shen S: Honokiol induces apoptosis and autophagy via the
ROS/ERK1/2 signaling pathway in human osteosarcoma cells in vitro
and in vivo. Cell Death Dis. 9:1572018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhou J, Zhang S, Ong CN and Shen HM:
Critical role of pro-apoptotic Bcl-2 family members in
andrographolide-induced apoptosis in human cancer cells. Biochem
Pharmacol. 72:132–144. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cheung HY, Cheung SH, Li J, Cheung CS, Lai
WP, Fong WF and Leung FM: Andrographolide isolated from
Andrographis paniculata induces cell cycle arrest and
mitochondrial-mediated apoptosis in human leukemic HL-60 cells.
Planta Med. 71:1106–1111. 2005. View Article : Google Scholar
|
|
16
|
Wong HC, Wong CC, Sagineedu SR, Loke SC,
Lajis NH and Stanslas J: SRJ23, a new semisynthetic andrographolide
derivative: In vitro growth inhibition and mechanisms of cell cycle
arrest and apoptosis in prostate cancer cells. Cell Biol Toxicol.
30:269–288. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Burgess DJ: Apoptosis: Refined and lethal.
Nat Rev Cancer. 13:792013. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zimmermann KC, Bonzon C and Green DR: The
machinery of programmed cell death. Pharmacol Ther. 92:57–70. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fruehauf JP and Meyskens FL Jr: Reactive
oxygen species: A breath of life or death? Clin Cancer Res.
13:789–794. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pelicano H, Carney D and Huang P: ROS
stress in cancer cells and therapeutic implications. Drug Resist
Updat. 7:97–110. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang G, Zhang T, Sun W, Wang H, Yin F,
Wang Z, Zuo D, Sun M, Zhou Z, Lin B, et al: Arsenic sulfide induces
apoptosis and autophagy through the activation of ROS/JNK and
suppression of Akt/mTOR signaling pathways in osteosarcoma. Free
Radic Biol Med. 106:24–37. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Smolensky D, Rhodes D, McVey DS, Fawver Z,
Perumal R, Herald T and Noronha L: High-Polyphenol Sorghum Bran
Extract Inhibits Cancer Cell Growth Through ROS Induction, Cell
Cycle Arrest, and Apoptosis. J Med Food. 21:990–998. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li H-Y, Zhang J, Sun L-L, Li BH, Gao HL,
Xie T, Zhang N and Ye ZM: Celastrol induces apoptosis and autophagy
via the ROS/JNK signaling pathway in human osteosarcoma cells: An
in vitro and in vivo study. Cell Death Dis. 6:e16042015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jian SL, Chen WW, Su YC, Su YW, Chuang TH,
Hsu SC and Huang LR: Glycolysis regulates the expansion of
myeloid-derived suppressor cells in tumor-bearing hosts through
prevention of ROS-mediated apoptosis. Cell Death Dis. 8:e27792017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zeng KW, Song FJ, Wang YH, Li N, Yu Q,
Liao LX, Jiang Y and Tu PF: Induction of hepatoma carcinoma cell
apoptosis through activation of the JNK-nicotinamide adenine
dinucleotide phosphate (NADPH) oxidase-ROS self-driven death signal
circuit. Cancer Lett. 353:220–231. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kyriakis JM, Banerjee P, Nikolakaki E, Dai
T, Rubie EA, Ahmad MF, Avruch J and Woodgett JR: The
stress-activated protein kinase subfamily of c-Jun kinases. Nature.
369:156–160. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Banerjee M, Chattopadhyay S, Choudhuri T,
Bera R, Kumar S, Chakraborty B and Mukherjee SK: Cytotoxicity and
cell cycle arrest induced by andrographolide lead to programmed
cell death of MDA-MB-231 breast cancer cell line. J Biomed Sci.
23:402016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wei RJ, Zhang XS and He DL:
Andrographolide sensitizes prostate cancer cells to TRAIL-induced
apoptosis. Asian J Androl. 20:200–204. 2018. View Article : Google Scholar :
|
|
29
|
Lim SC, Jeon HJ, Kee KH, Lee MJ, Hong R
and Han SI: Andrographolide induces apoptotic and non-apoptotic
death and enhances tumor necrosis factor-related apoptosis-inducing
ligand-mediated apoptosis in gastric cancer cells. Oncol Lett.
13:3837–3844. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mao W, He P, Wang W, Wu X and Wei C:
Andrographolide sensitizes Hep-2 human laryngeal cancer cells to
carboplatin-induced apoptosis by increasing reactive oxygen species
levels. Anticancer Drugs. 30:e07742019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhou J, Lu GD, Ong CS, Ong CN and Shen HM:
Andrographolide sensitizes cancer cells to TRAIL-induced apoptosis
via p53-mediated death receptor 4 up-regulation. Mol Cancer Ther.
7:2170–2180. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Alzaharna M, Alqouqa I and Cheung HY:
Taxifolin synergizes Andrographolide-induced cell death by
attenuation of autophagy and augmentation of caspase dependent and
independent cell death in HeLa cells. PLoS One. 12:e01713252017.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Rodríguez-Muela N, Germain F, Mariño G,
Fitze PS and Boya P: Autophagy promotes survival of retinal
ganglion cells after optic nerve axotomy in mice. Cell Death
Differ. 19:162–169. 2012. View Article : Google Scholar :
|
|
34
|
Li B, Zhu X, Sun L, Yuan L, Zhang J, Li H
and Ye Z: Induction of a specific CD8+ T-cell response to
cancer/testis antigens by demethylating pre-treatment against
osteosarcoma. Oncotarget. 5:10791–10802. 2014.PubMed/NCBI
|
|
35
|
Scherz-Shouval R and Elazar Z: Regulation
of autophagy by ROS: Physiology and pathology. Trends Biochem Sci.
36:30–38. 2011. View Article : Google Scholar
|
|
36
|
Neumann J, Yang Y, Köhler R, Giaisi M,
Witzens-Harig M, Liu D, Krammer PH, Lin W and Li-Weber M: Mangrove
dolabrane-type of diterpenes tagalsins suppresses tumor growth via
ROS-mediated apoptosis and ATM/ATR-Chk1/Chk2-regulated cell cycle
arrest. Int J Cancer. 137:2739–2748. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kamata H, Honda S, Maeda S, Chang L,
Hirata H and Karin M: Reactive oxygen species promote
TNFalpha-induced death and sustained JNK activation by inhibiting
MAP kinase phosphatases. Cell. 120:649–661. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Isakoff MS, Bielack SS, Meltzer P and
Gorlick R: Osteosarcoma: Current Treatment and a Collaborative
Pathway to Success. J Clin Oncol. 33:3029–3035. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Deng Y, Bi R, Guo H, Yang J, Du Y, Wang C
and Wei W: Andrographolide Enhances TRAIL-Induced Apoptosis via
p53-Mediated Death Receptors Up-Regulation and Suppression of the
NF-кB Pathway in Bladder Cancer Cells. Int J Biol Sci. 15:688–700.
2019. View Article : Google Scholar :
|
|
40
|
Peng T, Hu M, Wu TT, Zhang C, Chen Z,
Huang S and Zhou XH: Andrographolide suppresses proliferation of
nasopharyngeal carcinoma cells via attenuating NF-κB pathway.
BioMed Res Int. 2015:7350562015. View Article : Google Scholar
|
|
41
|
Zhang QQ, Ding Y, Lei Y, Qi CL, He XD, Lan
T, Li JC, Gong P, Yang X, Geng JG, et al: Andrographolide suppress
tumor growth by inhibiting TLR4/NF-κB signaling activation in
insulinoma. Int J Biol Sci. 10:404–414. 2014. View Article : Google Scholar :
|
|
42
|
Peng Y, Wang Y, Tang N, Sun D, Lan Y, Yu
Z, Zhao X, Feng L, Zhang B, Jin L, et al: Andrographolide inhibits
breast cancer through suppressing COX-2 expression and angiogenesis
via inactivation of p300 signaling and VEGF pathway. J Exp Clin
Cancer Res. 37:2482018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cui Q, Wang JQ, Assaraf YG, Ren L, Gupta
P, Wei L, Ashby CR Jr, Yang DH and Chen ZS: Modulating ROS to
overcome multidrug resistance in cancer. Drug Resist Updat.
41:1–25. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Gupta SC, Hevia D, Patchva S, Park B, Koh
W and Aggarwal BB: Upsides and downsides of reactive oxygen species
for cancer: The roles of reactive oxygen species in tumorigenesis,
prevention, and therapy. Antioxid Redox Signal. 16:1295–1322. 2012.
View Article : Google Scholar :
|
|
45
|
Kumari S, Badana AK, G MM, G S and Malla
RR: G MM, G S and Malla R: Reactive Oxygen Species: A Key
Constituent in Cancer Survival. Biomark Insights. 13:919146892018.
View Article : Google Scholar
|
|
46
|
Nita M and Grzybowski A: The Role of the
Reactive Oxygen Species and Oxidative Stress in the Pathomechanism
of the Age-Related Ocular Diseases and Other Pathologies of the
Anterior and Posterior Eye Segments in Adults. Oxid Med Cell
Longev. 2016:31647342016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Boonstra J and Post JA: Molecular events
associated with reactive oxygen species and cell cycle progression
in mammalian cells. Gene. 337:1–13. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Bridge G, Rashid S and Martin SA: DNA
mismatch repair and oxidative DNA damage: Implications for cancer
biology and treatment. Cancers (Basel). 6:1597–1614. 2014.
View Article : Google Scholar
|
|
49
|
Zhou G-Z, Li A-F, Sun Y-H and Sun G-C: A
novel synthetic curcumin derivative MHMM-41 induces ROS-mediated
apoptosis and migration blocking of human lung cancer cells A549.
Biomed Pharmacother. 103:391–398. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Pardhasaradhi BVV, Ali AM, Kumari AL,
Reddanna P and Khar A: Phycocyanin-mediated apoptosis in AK-5 tumor
cells involves down-regulation of Bcl-2 and generation of ROS. Mol
Cancer Ther. 2:1165–1170. 2003.PubMed/NCBI
|
|
51
|
Deeb D, Gao X, Jiang H, Janic B, Arbab AS,
Rojanasakul Y, Dulchavsky SA and Gautam SC: Oleanane triterpenoid
CDDO-Me inhibits growth and induces apoptosis in prostate cancer
cells through a ROS-dependent mechanism. Biochem Pharmacol.
79:350–360. 2010. View Article : Google Scholar
|
|
52
|
Shen HM and Liu ZG: JNK signaling pathway
is a key modulator in cell death mediated by reactive oxygen and
nitrogen species. Free Radic Biol Med. 40:928–939. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Dhanasekaran DN and Reddy EP: JNK
signaling in apoptosis. Oncogene. 27:6245–6251. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Krajarng A, Imoto M, Tashiro E, Fujimaki
T, Shinjo S and Watanapokasin R: Apoptosis induction associated
with the ER stress response through up-regulation of JNK in HeLa
cells by gambogic acid. BMC Complement Altern Med. 15:262015.
View Article : Google Scholar : PubMed/NCBI
|