Introduction
Triple-negative breast cancer (TNBC), which accounts
for 10-20% of all types of breast cancers (1,2), is
characterized by the lack of estrogen receptor (ER) and
progesterone receptor expression in addition to the absence of
human epidermal growth factor receptor 2 (HER2) amplification.
Although it exhibits positive responses to chemotherapy, TNBC
remain the most aggressive sub-type among younger patients since it
is associated with a worse outcome and a higher risk of relapse
within 5 years compared with other sub-types of breast cancer
(3-7). Treatment of patients with TNBC has
always been challenging due to the absence of effective therapeutic
targets. Recently, olaparib, an inhibitor of poly (adenosine
diphosphate-ribose) polymerase, has demonstrated promising
antitumor activity in HER2-negative patients with a germline
mutation in the BRCA gene (8).
Although BRCA germline mutations have a relatively higher
prevalence in TNBC compared with that in the non-TNBC sub-types,
its prevalence in TNBC is only 12% (9,10),
severely limiting the efficacy of this treatment strategy.
Consequently, there is an urgent need to identify promising
alternative treatment strategies for TNBC.
A hallmark of cancer is the dysregulation of cell
division, ultimately favoring aberrant proliferation that fuels
tumorigenesis and disease progression (11). The cyclin-dependent kinase
(CDK)-retinoblastoma (RB)-E2F axis controls transition through the
G1/S cell cycle checkpoint. CDK4/6 first forms a complex
with cyclin D1 to promote the phosphorylation of the RB protein
(12). Phosphorylated (p)-RB then
allows the dissociation of transcription factor E2F from the
pRb–E2F complex to promote G1/S transition (12). Inhibition of CDK4/6 results in RB
dephosphorylation, leading to cell cycle arrest (12). Since CDK4/6 activity is frequently
dysregulated and constitutively active in breast cancer cells,
inhibition of CDK4/6 may prove to be an effective therapeutic
intervention against breast cancer (2). Treatment using highly selective
CDK4/6 inhibitors, including palbociclib, ribociclib and
abemaciclib, have been previously reported to improve
progression-free survival (PFS) in the first and later lines of
ER+ metastatic breast cancer therapy (13-18).
Since TNBC is a group of highly prolifera-tive tumors enriched in
cell cycle genes, intrinsic resistance to CDK4/6 inhibitor
monotherapy has been observed owing to the loss or mutation of RB
(19-21). By contrast, it was also reported
that the loss of RB in TNBC can result in improved response to
chemotherapy (22-24). The Cancer Genome Atlas study
revealed that since only 20% of patients with TNBC carry mutations
in RB or are RB-negative, the use of CDK4/6 inhibitors may serve as
a promising therapeutic strategy for RB-positive TNBC (2).
Human sulfatase-1 (HSulf-1) is an endosulfatase that
selectively removes 6-O-sulfate from heparan sulfate proteoglycans
(HSPGs) in the extracellular matrix (25). The sulfation of HSPG residues is
required for their interaction with a number of heparin-binding
growth factors (HBGFs), including fibroblast growth factor 2
(FGF-2), hepatocyte growth factor (HGF) and heparin-binding
epidermal growth factor (HB-EGF), to promote their activation
(25,26). In normal human tissues, HSulf-1 is
stably expressed and serves important roles in cell proliferation
and differentiation (27).
However, HSulf-1 expression has been reported to be downregulated
in various tumors, including ovarian, breast and gastric carcinomas
(28,29). A number of studies previously
revealed that the down-regulation of HSulf-1 in cancers is mediated
by epigenetic modification, including CpG island methylation and
histone acetylation (28,29) Reduced expression of HSulf-1 is one
of the causes behind the increased sulfation of HSPGs, leading to
excessive activation of the signal transduction pathways downstream
of growth factor receptors. Ultimately, this results in cancer cell
proliferation, increased metastatic activity, inhibition of
apoptosis and reduced sensitivity to radio- and chemotherapy
(30-37). Previously, several studies revealed
that the overexpression of HSulf-1 inhibited tumor proliferation
and angiogenesis in xenografts derived from breast cancer cell
lines (36). Additionally, high
levels of HSulf-1 have also been found to correlate with increased
PFS and overall survival (OS) in patients with breast cancer
(38,39). Mechanistically, it was reported
that HSulf-1 inhibited autocrine cyclin D1 expression, leading to
decreased S phase fractions whilst increasing G2-M
fractions and increased cell death (39). However, the potential relationship
between HSulf-1 and CDK4/6 inhibition remains unexplored, which
serves as the subject of the present study.
In the present study, HSulf-1 expression level was
first measured in TNBC tissues and cell lines, following which the
function of HSulf-1 in TNBC was next demonstrated. Secondly, the
effects of HSulf-1 overexpression in combination with treatment
with the CDK4/6 inhibitor palbociclib on RB-positive TNBC was
investigated. Finally, further studies were performed to explore
the mechanism underlying this effect. Based on the present study,
it could be speculated that HSulf-1 may not only serve as a
potential therapeutic target but also a reliable efficacy predictor
for CDK4/6 inhibitors. Therefore, in clinical practice, patients
with TNBC and elevated HSulf-1 expression can potentially be
selected for palbociclib treatment, which may achieve superior
therapeutic efficacy.
Materials and methods
Human tissue specimens
In total, 86 tissue TNBC specimens and paired
adjacent normal breast tissues were collected from patients with
TNBC (sex, all female; age range, 25-73 years; median age, 47) at
the General Hospital of The Yangtze River Shipping (Wuhan, China),
Wuhan Polytechnic University (Wuhan, China) and the Henan
Provincial People's Hospital, Zhengzhou University (Zhengzhou,
China) between January 2000 and January 2019. All specimens were
collected by surgical resection and the distance between the tumour
and adjacent normal tissue was >1 cm. Non-TNBC patients,
patients receiving chemotherapy or radiotherapy prior to surgical
treatment and pregnancy concomitant with the diagnosis were
excluded from the present study. The follow-up time range was 6-210
months. Informed consent was obtained from all patients. This
experiment was approved by the Ethics Committee of the General
Hospital of The Yangtze River Shipping, Wuhan Polytechnic
University (approval no. 2017IEC0003). All specimens were
classified according to the 2003 World Health Organization
Consensus Classification and the Eighth Edition AJCC Cancer Staging
for breast cancer (40,41). Detailed clinicopathological
features are shown in Table I.
 | Table IAssociation between HSulf-1
expression and clinicopathologic characteristics of 86 TNBC
patients. |
Table I
Association between HSulf-1
expression and clinicopathologic characteristics of 86 TNBC
patients.
| Parameters | HSulf-1 expression
| P-valuea |
|---|
| Low (n=55) | % | High (n=31) | % |
|---|
| Age (years) | | | | | 0.4238 |
| ≤50 | 35 | 67 | 17 | 33 | |
| >50 | 20 | 59 | 14 | 41 | |
| Grade | | | | | 0.6237 |
| Low | 10 | 59 | 7 | 41 | |
| High | 45 | 65 | 24 | 35 | |
| Tumor size
(cm) | | | | | 0.5089 |
| ≤5 | 41 | 63 | 24 | 37 | |
| >5 | 14 | 67 | 7 | 33 | |
| Lymph node
metastasis | | | | | 0.7535 |
| Negative | 30 | 64 | 17 | 36 | |
| Positive | 25 | 64 | 14 | 36 | |
Immunohistochemistry
Tissue samples were first fixed in 4% formalin for
24 h at room temperature and then embedded in paraffin blocks.
Tissue sections (thickness, 4 µm) were subsequently deparaffinized
in a graded series of xylene followed by rehydration in a
descending series of ethanol before antigen retrieval in citrate
buffer (pH 9.0) at 100°C for 2 min. Then, the tissue sections were
blocked for 30 min with 3% BSA (Sigma-Aldrich; Merck KGaA) at room
temperature. After incubation with primary anti-HSulf-1 (1:250;
cat. no. ab32763; Abcam) at 4°C overnight and secondary antibodies
(biotin-conjugated goat anti-rabbit IgG, 1:400 dilution, cat. no.
SA00004-2; Proteintech Group, Inc.) at room temperature for 30 min,
the sections were then incubated with avidin-biotin
complex-horseradish peroxidase (HRP; Vectastain® ABC
kit, Maravai LifeSciences) at 37°C for 20 min followed by
incubation with 3'3-diaminobenzidine (Sigma-Aldrich; Merck KGaA)
and counterstained with hematoxylin (Sigma-Aldrich; Merck KGaA) for
5 min at room temperature. The percentage of positively stained
cells was scored on a scale of 0 to 4 as follows: i) 0, ≤5%; ii) 1,
6-25%); iii) 2, 26-50%); iv) 3, 50-75%; and v) 4, 76-100%. The
staining intensity was scored from 0 to 3 as follows: i) 0,
negative); ii) 1, weak; iii) 2, moderate; and iv) 3, strong). The
scores for the percentages of positive cells and staining
intensities were then multiplied to generate an immunoreactivity
score (IS) for each case. The IS ranged from 0-12 (0, 1, 2, 3, 4,
6, 8, 9 and 12). Cut-off values for this scoring system were
assigned as follows: i) High HSulf-1 expression, IS ≥4 (4, 6, 8, 9
and 12); ii) low HSulf-1 expression, IS <4 (0, 1, 2 and 3).
Tissue slides were visualized under a light microscope
(magnification, x400; Olympus FSX100; Olympus Corporation). Five
high-power fields were examined randomly for each section.
Quantifications of the proteins of interest were performed using
the Image Pro Plus 6.0 software (Media Cybernetics, Inc.).
Cell lines and culture
The breast cancer cell lines (Hs578T and MDA-MB-231)
human immortalized breast epithelial cell line (MCF10A) and 293T
cells were purchased from American Type Culture Collection (ATCC).
Hs578T MDA-MB-231 and 293T cells were cultured in DMEM (Gibco;
Thermo Fisher Scientific, Inc.) with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) in a humidified atmosphere at 37°C in 5%
CO2. MCF10A cells were cultured in DMEM/F12 (Procell
Life Science & Technology Co., Ltd.) containing 5% horse serum,
20 ng/ml EGF, 0.5 µg/ml hydrocortisone, 10 µg/ml insulin and 1%
NEAA (Procell Life Science & Technology Co., Ltd.) in a
humidified atmosphere at 37°C in 5% CO2. Palbociclib
(Sigma-Aldrich; Merck KGaA) was dissolved in DMSO, following which
the cells were treated with 500 nM palboci-clib for 72 h at 37°C.
Palbociclib (500 nM) was used for the majority of the experiments
as previously described (42,43).
An equivalent concentration of DMSO was used as negative control
treatment in parallel with palbociclib treatment.
Plasmids, lentivirus production and cell
transfection
The plasmids (pcDNA3.0-vector, pcDNA3.0-HSulf-1,
pcDNA3.0-cyclin D1, LV105-HSulf-1 and LV105-EGFP) and the vector
psi-H1 used for cloning the HSulf-1 short hairpin RNA (shRNA) were
purchased from iGene Biotechnology Co., Ltd. Hs578T and MDA-MB-231
cells (5x105 cells/well) were plated into six-well
plates 24 h prior to transfection. Plasmids (2.5 µg/well) were
transfected into cells using Lipofectamine® 3000
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol for in vitro assays. Cells were
incubated for 48 h at 37°C prior to further experimentation. The
lentivirus particles were produced by transfecting 293T cells
(ATCC) with pEZ-Lv105 lentiviral vectors encoding HSulf-1
(LV105-HSulf-1) and the control empty vector (LV105-EGFP) using the
LentiPac™ Expression packaging kit (GeneCopoeia, Inc.) according to
the manufacturer's protocols. The lentivirus-containing
supernatants were harvested 72 h following transfection and were
filtered through 0.45-µm PVDF filters (EMD Millipore). The
supernatant was then concentrated by ultracentrifugation at 100,000
x g for 2 h at room temperature. MDA-MB-231 cells (4x105
cells/well; multiplicity of infection, 10) were infected with the
lentiviral particles (2.03x108 TU/ml) where the stable
cell lines were established by treatment with puromycin (2.5 µg/ml)
for 2 weeks at 37°C for in vivo study. Transfection
efficiency was determined by reverse transcription-quantitative PCR
(RT-qPCR) and western blot analysis. The target sequences used for
shRNA were as follows: ShHSulf-1 1, 5'-CCC AAA TAT GAA CGG GTC
AAA-3' and shHSulf-1 2, 5'-CCA AGA CCT AAG AAT CTT GAT-3'. The
plasmid shHSulf-11 was chosen for further study based on its
superior silencing effect.
RT-qPCR
Total RNA was extracted from MDA-MB-231 cells
transfected with the HSulf-1 overexpression or vector plasmid using
RN07-EASYspin kit (Aidlab Biotechnologies Co., Ltd) according to
manufacturer's protocols. cDNA was then synthe-sized using the
PrimeScript™ RT Master Mix (Takara Bio, Inc.) from 1 µg RNA
according to manufacturer's protocols The following temperature
protocol was used for the reverse transcription reaction: 37°C for
15 min, followed by reverse transcriptase inactivation reaction:
85°C for 5 sec. qPCR reactions were performed using
SYBR® Premix Ex Taq™ (Takara Bio, Inc.) according to
manufacturer's protocols. The thermo-cycling conditions were as
follows: Initial denaturation at 95°C for 30 sec, followed by 40
cycles of 95°C for 5 sec and 60°C for 30 sec. Relative expression
was calculated using the 2-∆∆Cq method (44). GAPDH was used as an internal
control. The sequences of the primers were as follows: Cyclin D1
forward, 5'-CCC ACT CCT ACG ATA CGC-3' and reverse, 5'-AGC CTC CCA
AAC ACC C-3'; GAPDH forward, 5'-GGA GCG AGA TCC CTC CAA AAT-3' and
reverse, 5'-GGC TGT TGT CAT ACT TCT CAT GG-3'.
Western blotting
Protein extracts were prepared using RIPA buffer
(Thermo Fisher Scientific, Inc.) supplemented with protease and
phosphatase inhibitors. Protein concentrations were determined
using a bicinchoninic acid protein assay kit (Thermo Fisher
Scientific, Inc.). A total of 20 µg total protein was loaded per
lane and separated by SDS-PAGE (10 or 12% gels) before transferal
to polyvinylidene fluoride membranes (EMD Millipore). The membranes
were blocked in 5% skimmed milk diluted with Tris-buffered
saline/Tween-20 (0.1%) (TBS-T) at room temperature for 1 h and
subsequently incubated overnight at 4°C with the following primary
antibodies: Anti-RB (1:1,000, cat. no. 9309; Cell Signaling
Technology, Inc.), anti-p-RB (1:1,000, Ser780; cat. no. 9307; Cell
Signaling Technology, Inc.), anti-HSulf-1 (1:1,000, cat. no.
ab32763; Abcam), anti-E-cadherin (1:500, cat. no. ab15148; Abcam),
anti-vimentin (1:1,000, cat. no. ab92547; Abcam), anti-N-cadherin
(1:2,000, cat. no. ab76011; Abcam), anti-cyclin D1 (1:200, cat. no.
ab16663; Abcam), anti-STAT3 (1:1,000, cat. no. 30835; Cell
Signaling Technology, Inc.), anti-p-STAT3 (Y705; 1:2,000, cat. no.
9145; Cell Signaling Technology, Inc.), anti-JAK2 (1:1,000, cat.
no. 74987; Cell Signaling Technology, Inc.), anti-p-JAK2 (Tyr1007;
1:1,000, cat. no. 4406; Cell Signaling Technology, Inc.), anti-AKT
(1:1,000, cat. no. 9272; Cell Signaling Technology, Inc.),
anti-p-AKT (Ser473; 1:1,000, cat. no. 9271; Cell Signaling
Technology, Inc.), anti-ERK1/2 (1:1,000, cat. no. 4695; Cell
Signaling Technology, Inc.), anti-pERK1/2 (Thr202/Tyr204; 1:2,000,
cat. no. 4370; Cell Signaling Technology, Inc.), anti-GAPDH
(1:5,000, cat. no. 60004-1-Ig; Proteintech Group, Inc.), and
anti-β-actin (cat. no. 60008-1-Ig; Proteintech Group, Inc.). The
next day, the membranes were washed with TBS-T and then incubated
with secondary antibodies including horseradish
peroxidase-conjugated goat anti-mouse (1:10,000, cat. no.
SA00001-1; Proteintech Group, Inc.) and goat anti-rabbit (1:10,000,
cat. no. SA00001-2; Proteintech Group, Inc.) at 37°C for 1 h.
Immunoreactive bands were visualized using ECL reagent (Advansta,
Inc.) and detected using the ChemiDoc XRS+ system (Bio-Rad
Laboratories, Inc.). Gray values were calculated using ImageJ
software (version 1.52u; National Institutes of Health).
Cell counting kit-8 (CCK-8) assay
Hs578T and MDA-MB-231 cells (3,000 cells/well) were
first seeded into 96-well plates in triplicate, allowed to attach
for 24 h at 37°C and treated with the indicated concentrations of
palbociclib for 0, 24, 48 and 72 h at 37°C. Subsequently, 10 µl
CCK-8 solution (MedChemExpress) was added into each well before the
plates were subsequently incubated for 4 h at 37°C. The optical
density was measured at 450 nm using the SpectraMax M5 microplate
reader (Molecular Devices, LLC).
Colony formation assay
Cells were seeded in 6-well plates at a density of
1,000 cells per well. After 24 h of culture at 37°C, the Hs578T and
MDA-MB-231 cells were transfected with the HSulf-1 overexpression
or vector plasmid and then treated with 500 nM palbociclib for 14
days at 37°C. After 14 days, cell colonies were fixed with 4%
paraformaldehyde and stained with 0.1% crystal violet, each for 30
min at room temperature. Cell colonies containing >50 cells were
counted using the ImageJ software (version 1.52u; National
Institutes of Health) and images were taken with a camera (Canon,
Inc.).
Cell cycle assay
Hs578T and MDA-MB-231 cells were harvested and fixed
with 70% cold ethanol at 4°C overnight, following which the cells
(8x105/tube) were washed with PBS twice, mixed with 400
µl propidium iodide (PI) (Beyotime Institute of Biotechnology) and
incubated for 30 min in the dark at room temperature and then
analyzed by flow cytometry (Cytoflex, Beckman Coulter, Inc.) with a
CytExpert software (version 2.3; Beckman Coulter, Inc.). Each
sample was tested in triplicate.
Apoptosis analysis
Hs578T and MDA-MB-231 cell apoptosis was analyzed by
flow cytometry (CytoFLEX; Beckman Coulter, Inc.) as per the
FITC-conjugated Annexin V/PI method (Annexin V-FITC/PI Apoptosis
Kit; Beyotime Institute of Biotechnology). After 72 h of the
indicated treatment, adherent and suspended cells
(3x105/tube) were both harvested by centrifugation and
resuspended in 195 µl binding buffer, following which Annexin
V-FITC (5 µl) and PI (10 µl) were added to each sample and mixed.
Cells were incubated for 15 min in the dark at 4°C and then
analyzed by flow cytometry (Cytoflex; Beckman Coulter, Inc.) using
a CytExpert software (version 2.3; Beckman Coulter, Inc.). Each
sample was tested in triplicate.
Wound-healing assay
Hs578T and MDA-MB-231 cells were seeded into
six-well plates at a density of 5x105 cells per well and
allowed to attach for 24 h at 37°C. Plasmids (pcDNA3.0-vector,
pcDNA3.0-HSulf-1, shNC and shHSulf-1) were transfected into cells
and then treated with 500 nM palbociclib for 72 h 37°C. Cells were
harvested and reseeded into six-well plates (1x106
cells/well) and cultured to 90% confluence. A 200 µl sterile
pipette tip was then used to scratch a wound (time 0 h), following
which fresh low-serum (2% FBS) DMEM was immediately replaced after
three times of washing with PBS. Cell migration was imaged under a
light microscope (magnification, x100; Olympus Corporation) at 0
and 24 h after the injury. The wound width was evaluated by
measuring the distance between the two edges of the scratch at five
sites in each image. Cell migration was determined using the
following formula: Percentage of wound healing (%) = [(wound width
at the 0 h time point-wound width at the 24 h time point)/wound
width at the 0 h time point] x100.
Transwell assay
Transwell chambers (Corning, Inc.) were coated with
50 µl of DMEM-diluted Matrigel (1:8 dilution; BD Biosciences) which
was precooled at 4°C for 12 h and then incubated overnight at 37°C.
Cells were suspended in serum-free culture medium (5x104
cells/well). A total of 200 µl cell suspension was transferred into
the upper chambers whilst 600 µl DMEM supplemented with 10% FBS was
added to the lower chambers. After 24 h incubation, the cells that
migrated to the lower chambers were fixed with 4% paraformaldehyde
and stained with 0.1% crystal violet each for 30 min at room
temperature. The number of migratory cells were counted from three
randomly chosen fields per chamber under a light microscope
(magnification, x100; Olympus Corporation).
Tumor xenografts
Animal experiments were performed with the approval
of the Animal Care and Use Committee of the General Hospital of The
Yangtze River Shipping, Wuhan Polytechnic University (approval no.
2017IEC0003; Wuhan, China). A total of 12 four-week-old female
BALB/c nude mice were maintained under standard conditions (room
temperature, 22±2°C; relative humidity, 55±10%) on a 12-h
light/dark cycle (lights on at 6:00 a.m.), where they had ad
libitum access to food and water. A daily observation was
preformed to prevent the animals from anger, restlessness, fear,
anxiety, pain or damage to keep them at a normal status. These mice
(age, 4 weeks) were randomly divided into two groups (n=6
mice/group). MDA-MB-231 cells (2x107) stably transfected
with LV105-HSulf-1 plasmids or the control vector were suspended in
100 ml PBS and subcutaneously injected into the nude mice under the
axilla of the left forelimb. Once the tumors of the control vector
group reached ~300 mm3, mice with tumors of similar size
were distributed into the treatment cohorts (n=3/group). Mice were
orally treated with 50 mg/kg palbociclib dissolved in 0.5%
methylcellulose or vehicle (0.5% methylcellulose) daily for 21
days. Tumor sizes were monitored every 3 days for 3 weeks. At the
completion of 3 weeks of palbociclib treatment, all 12 mice were
euthanized by cervical dislocation. Death was confirmed by the
cessation of breathing, muscle relaxation and the absence of nerve
reflex. The experiment lasted a total of 6 weeks. No accidental
death was observed before these mice were euthanized. The tumors
were then excised and measured. The length (l) and width (w) of
each tumor were measured using digital calipers, where tumor
volumes were calculated according to the following formula: Tumor
volume (V)=lxw2x0.5.
Statistical analysis
Statistical analyses were performed using GraphPad
Prism 8.0 software (Graphpad Software, Inc.). Data are presented as
the means ± standard deviation. Data were analyzed using Student's
t-test or one-way ANOVA followed by either Dunnett's or Tukey's
post hoc test. Wilcoxon-signed rank sum test and the χ2
test was used to analyze the IHC data, whilst the log-rank test was
used to assess the statistical signifi-cance of the Kaplan-Meier
plots and Cox proportional-hazards model was used to determine the
hazard ratio and 95% CI. P<0.05 were considered to indicate a
statistically significant difference.
Results
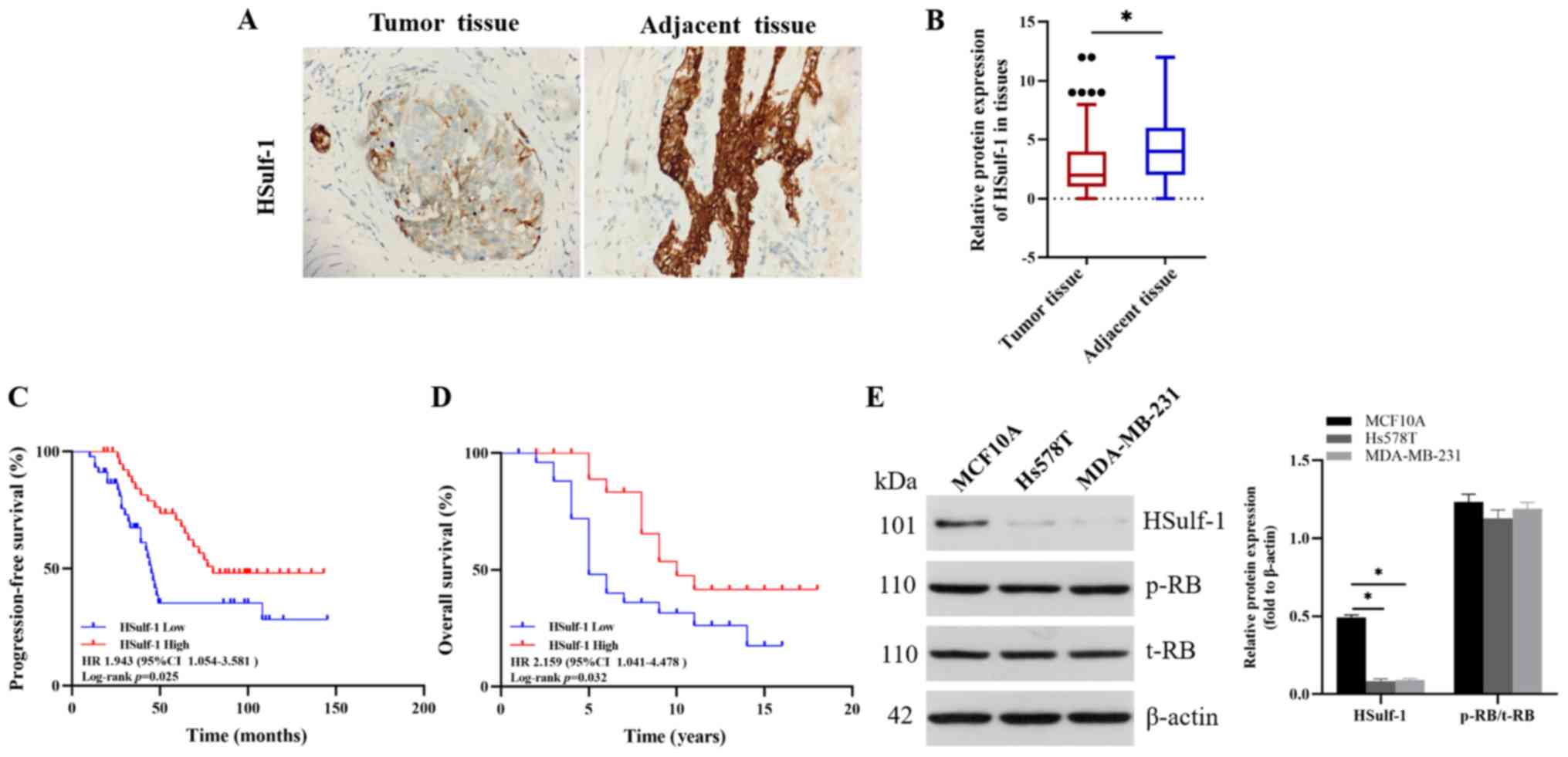
HSulf-1 is downregulated in TNBC tissues
and cell lines and high HSulf-1 levels associate with improved
prognosis in patients with TNBC
It was demonstrated previously that HSulf-1
expression was downregulated in ~60% of breast cancers, which was
in turn associated with poor patient prognosis (38,39).
Therefore, the expression levels of HSulf-1 in 86 pairs of TNBC and
adjacent normal mammary tissues were first measured, where it was
found that HSulf-1 expression was significantly downregulated in
TNBC tissues compared with that in the paired normal tissues
(Fig. 1A and B). In addition,
higher expression levels of HSulf-1 was found to significantly
associate with prolonged PFS (HR, 1.943; 95% CI, 1.054–3.581;
P=0.025) and OS (HR, 2.159; 95% CI, 1.041–4.478; P=0.032) in TNBC
(Fig. 1C and D). However, HSulf-1
levels showed no significant associations with the
clinicopathological indicators, including age, tumor grade, tumor
size and lymph node status (Table
I). A number of preclinical studies previously suggested that
CDK4/6 inhibitors exerted inhibitory effects in RB-positive TNBC
cell lines (43,45-47).
Therefore, two RB-positive cell lines Hs578T and MDA-MB-231 were
chosen to examine the expression levels of HSulf-1 and RB
phosphorylation in these two cell lines and one human immortalized
breast epithelial cell line MCF10A (43,47).
HSulf-1 expression also was also found to be significantly
downregulated in Hs578T and MDA-MB-231 compared with that in MCF10A
(Fig. 1E). Collectively, these
data suggested that HSulf-1 expression was downregulated in TNBC
tissues and cells, where reduced HSulf-1 expression was highly
predictive of poor patient prognosis.
Overexpression of HSulf-1 inhibits cell
proliferation and exhibits a synergistic antiproliferative effect
with palbociclib on RB-positive TNBC cells, both in vitro and in
vivo
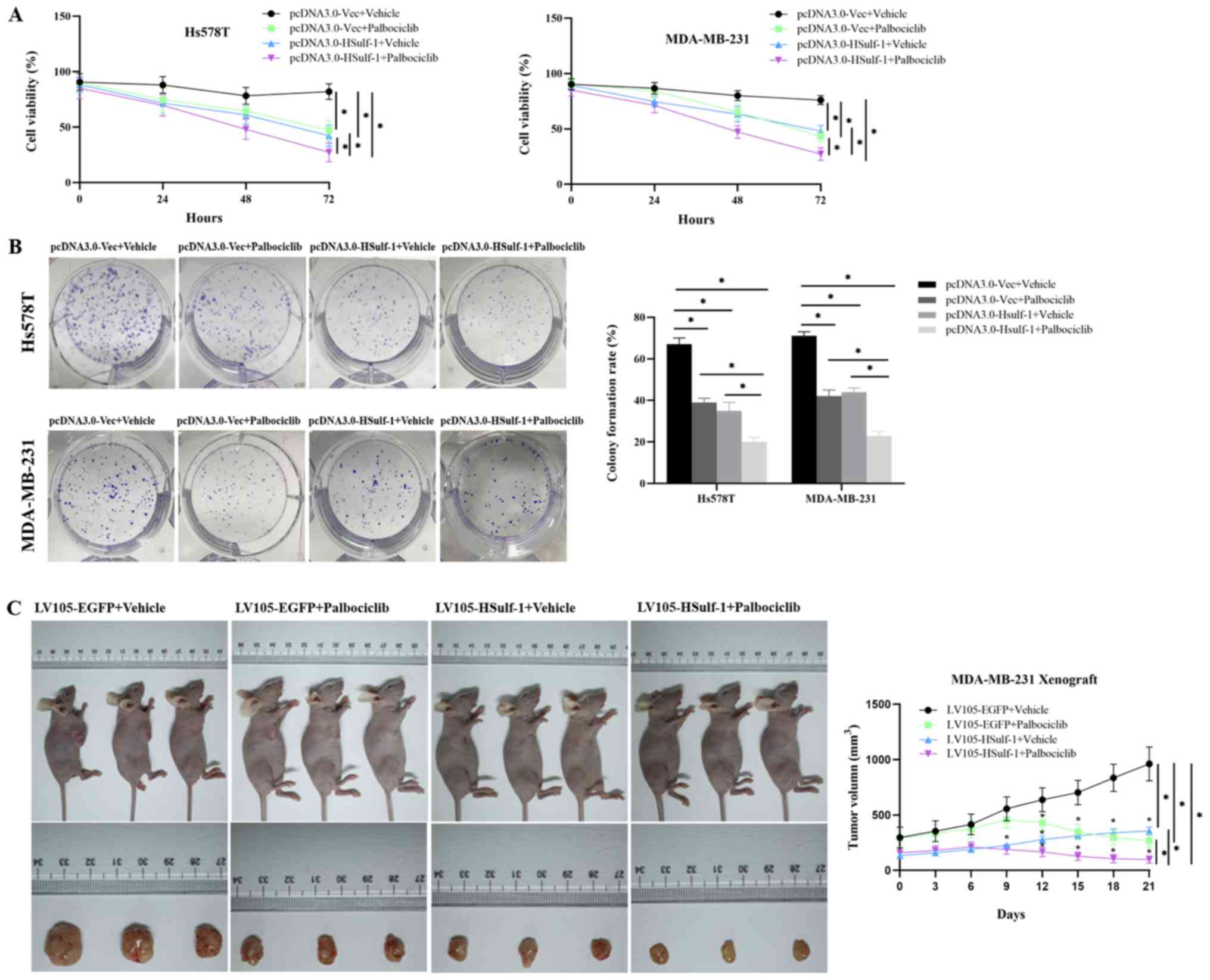
To explore the effects of HSulf-1 and/or palbociclib
treatment on TNBC cell proliferation, an HSulf-1 overexpression
plasmid was transfected into Hs578T and MDA-MB-231 cells. CCK-8
assay was performed to measure cell viability following treatment
with palbociclib for various periods of time (0, 24, 48 and 72 h).
Overexpression of HSulf-1 was found to significantly reduce the
viability of TNBC cells compared with those transfected with the
control vector (Fig. 2A). In
addition, combined HSulf-1 overexpression and palbociclib treatment
synergistically potentiated the reduction of cell viability
compared with that following either intervention alone (Fig. 2A). Colony formation assays yielded
similar trends of results (Fig.
2B). By contrast, downregulation of HSulf-1 expression using
shRNAs was found to significantly increase proliferation compared
with cells transfected with the control vector (Fig. S1A-C).
The effects of HSulf-1 overexpression and/or
palbociclib treatment on tumor proliferation was next investigated
in vivo. MDA-MB-231 cells stably expressing either the
control vector or HSulf-1 overexpression plasmid were injected into
female nude mice to form xenografts. The xenograft models were
treated with 50 mg/kg palbociclib dissolved in 0.5% methylcellulose
or vehicle daily for 21 consecutive days. Treatment with
palbociclib or overexpression of HSulf-1 alone was found to
significantly hinder tumor growth compared with tumors in the
Lv105-EGFP + vehicle group. (Fig.
2C). Combined treatment with palbociclib and HSulf-1
overexpres-sion potentiated the reduction in tumor volume compared
with that following either interventions alone (Fig. 2C). Collectively, these results
suggested that the combination of HSulf-1 over-expression and
palbociclib treatment co-operatively inhibited the proliferation of
RB-positive TNBC cells both in vitro and in vivo.
HSulf-1 and palbociclib exhibit a
cooperative antitumor effect by inducing G1/S cell cycle
arrest, inhibiting migration, invasion and
epithelial-to-mesenchymal transition (EMT) in RB-positive TNBC in
vitro
To investigate the biological effects of HSulf-1 and
palbociclib in RB-positive TNBC cells further, the following
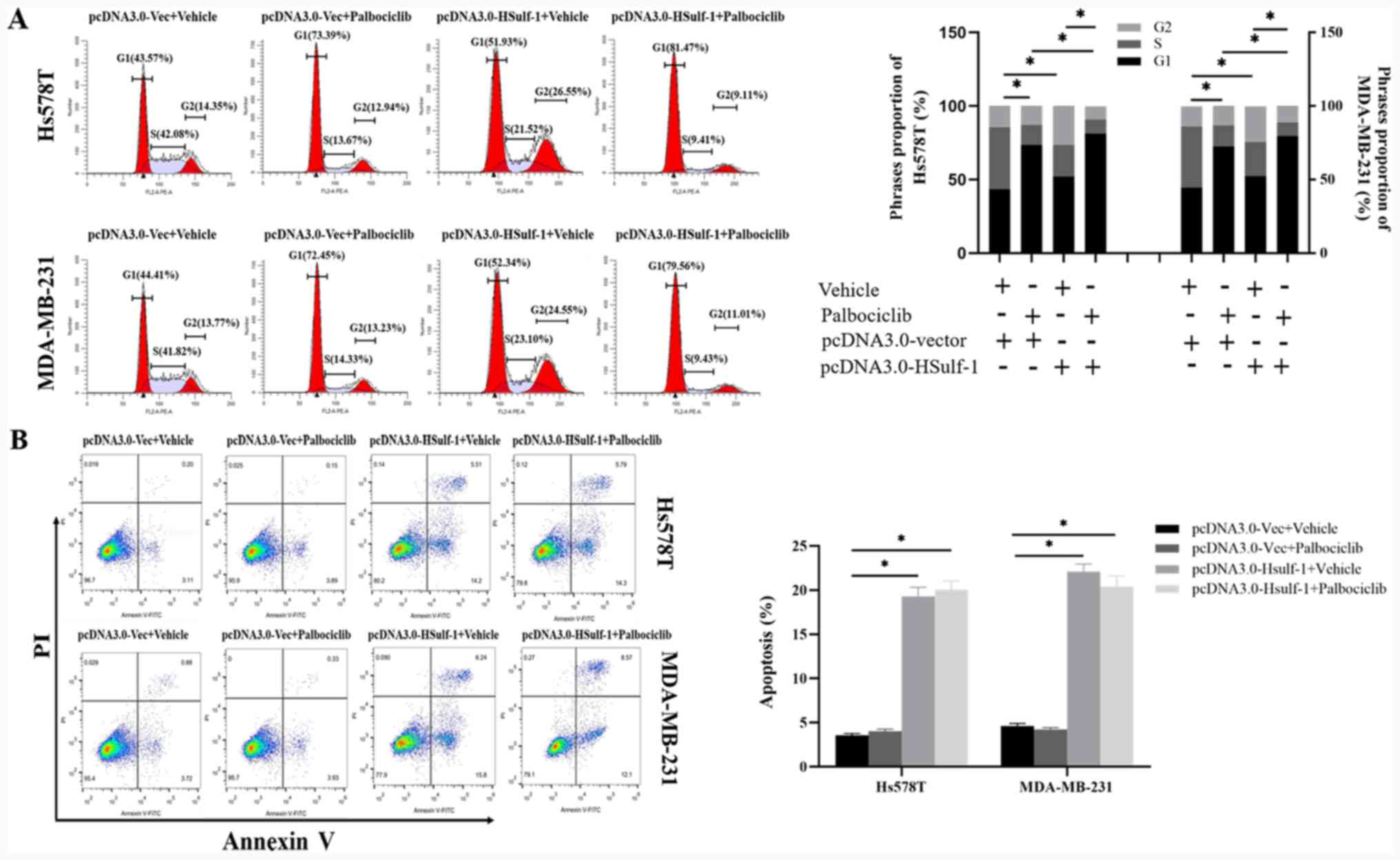
experiments were performed. Cell cycle analysis indicated that
palbociclib treatment significantly inhibited S phase entry, whilst
HSulf-1 overexpression increased the % of cells in the
G1 and G 2phases of the cell cycle (Fig. 3A). The combination of HSulf-1
overexpression and palbociclib treatment enhanced G1
arrest in RB-positive TNBC cells further compared with either
treatments alone (Fig. 3A).
Apoptosis analysis revealed that palbociclib treatment had no
impact on the apoptosis of TNBC cells (Fig. 3B). By contrast, significantly
increased apoptosis was observed in the HSulf-1-overexpressing TNBC
cells compared with those transfected with the control vector
(Fig. 3B). These results suggested
that the cooperative antiproliferative effect of the combinatorial
treatment was dependent on the induction of G1/S cell
cycle arrest instead of the apoptosis of RB-positive TNBC
cells.
The anti-metastatic effect of palbociclib on TNBC
remains controversial (48-51).
As a result of these contradictory reports, the potential
anti-migratory and anti-invasive properties of palbociclib in the
two RB-positive TNBC cell lines were investigated. Overexpression
of HSulf-1 or palbociclib treatment alone significantly impaired
the migration and invasion of Hs578T and MDA-MB-231 cells compared
with those treated with vehicle and transfected with the control
vector (Fig. 3C and D). By
contrast, when HSulf-1 expression was downregulated using shRNAs,
it was shown that the migratory and invasive abilities of Hs578T
and MDA-MB-231 cells were significantly increased compared with
those transfected with the control vector (Fig. S1D and E). Additionally,
palbociclib treatment combined with HSulf-1 overexpression
significantly enhanced the anti-migratory and invasive effects
further compared with those following either treatment alone
(Fig. 3C and D). Since EMT is
typically the first crucial process in cancer metastasis (52-55),
potential changes in the expression of EMT markers was examined.
Hs578T and MDA-MB-231 cells treated with palbociclib or transfected
with the HSulf-1 overexpression plasmid showed significantly
increased expression levels of E-cadherin, along with signifi-cant
reductions in those of vimentin and N-cadherin compared with cells
in the control vector+vehicle group (Fig. 3E). When the two interventions were
combined, a significant synergistic effect was observed (Fig. 3E). Collectively, these data
demonstrated the cooperative antitumor effects of combining
palbociclib with HSulf-1 overexpression was mediated via the
induction of G1/S cell cycle arrest and the inhibition
of migration, invasion and EMT in vitro.
HSulf-1 inhibits cell cycle progression
by suppressing the expression of cyclin D1 via non-canonical
AKT/STAT3 and ERK1/2/STAT3 signaling
Previous studies reported that HSulf-1 downregulated
cyclin D1 expression by inhibiting the STAT3 signaling pathway in
hepatocellular carcinoma cells (56). In addition, it was reported that
constitutive STAT3 activation occurs in >40% of breast cancers,
especially in the triple-negative sub-type (31,57).
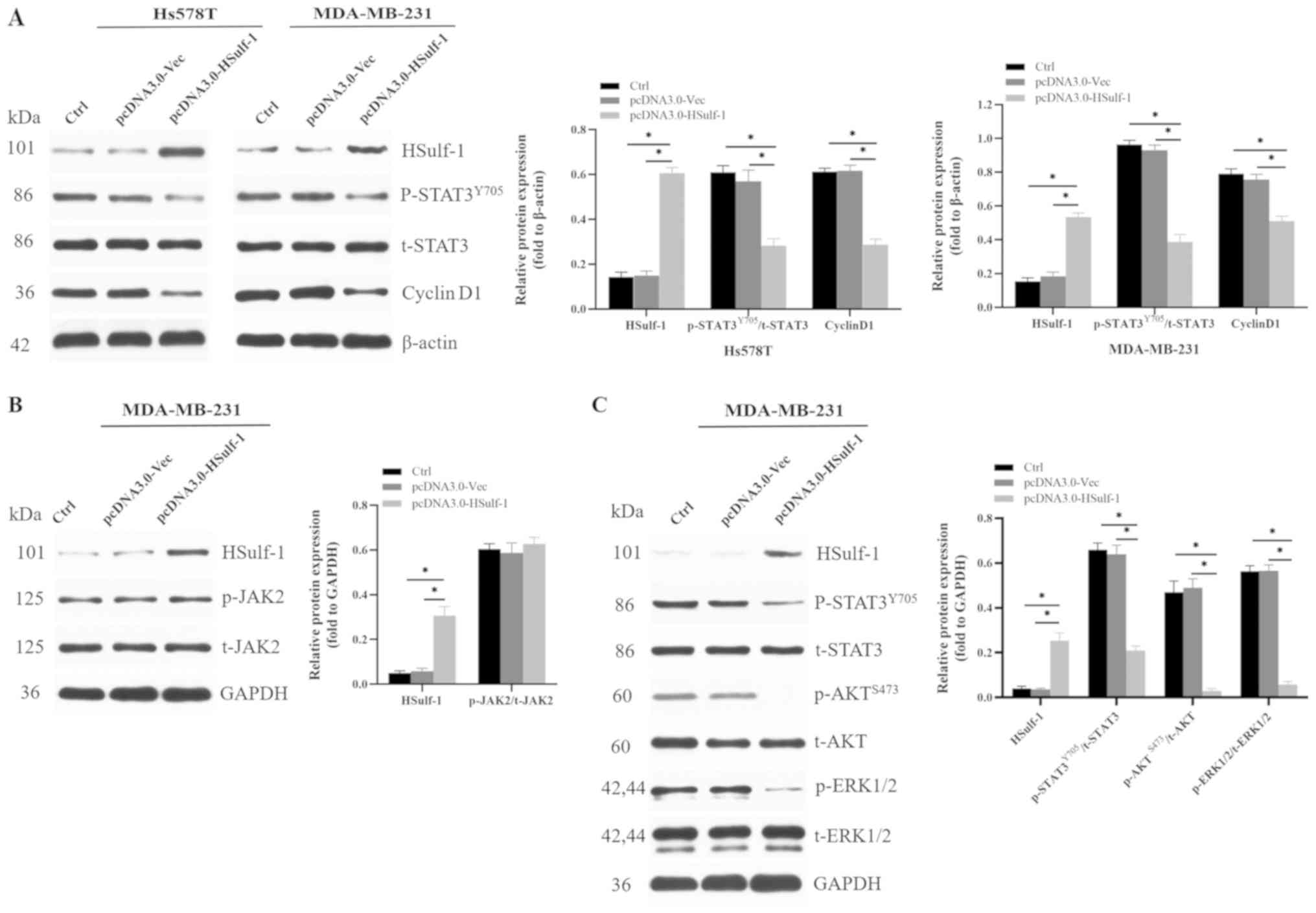
Therefore, to ascertain if HSulf-1 regulated cyclin D1 expression
in TNBC cells via the STAT3 pathway, HSulf-1 was overexpressed in
TNBC cells, following which the expression levels of the associated
critical signaling components were evaluated in vitro.
Overexpression of HSulf-1 significantly reduced cyclin D1
expression and STAT3 phosphorylation without affecting total STAT3
expression in TNBC cells compared with those transfected with the
control vector (Fig. 4A).
Additionally, it was hypothesized that HSulf-1 could inhibit the
expression of cyclin D1 through the canonical JAK2/STAT3 pathway.
Subsequent measurement of JAK2 phosphorylation and total JAK2
expression in parental and HSulf-1-overexpressing cells showed that
neither were significantly unaffected by HSulf-1 overexpression
(Fig. 4B).
Previous studies have also reported that HSulf-1
inhibited the AKT and ERK signaling pathways, leading to cell cycle
arrest, apoptosis, suppression of EMT and suppression of metastasis
(34,39,58-61).
Additionally, it has been demonstrated that ERK1/2 can directly
phosphorylate Ser727 at the C-terminus of STAT3 (62). Malanga et al previously
showed that suppression of AKT reduces the phosphorylation and
transcriptional activity of STAT3, whereas the opposite effects
were observed following AKT activation in non-small cell lung
cancer cells (63). Based on these
previous observations, it was hypothesized that HSulf-1 may
suppress STAT3 activity via the ERK1/2 and AKT signaling pathways.
Overexpression of HSulf-1 was found to significantly attenuate
ERK1/2, AKT and STAT3 phosphorylation in MDB-MB-231 cells compared
with those transfected with the control vector (Fig. 4C). In conclusion, these
observations suggest that HSulf-1 suppressed cyclin D1 expression
via non-canonical AKT/STAT3 and ERK1/2/STAT3 signaling but not the
canonical JAK2/STAT3 pathway, leading to cell cycle arrest and the
inhibition of proliferation of TNBC cells both in vitro and
in vivo.
HSulf-1 cooperates with palbociclib to
induce antiproliferative effects by reducing palbociclib-induced
cyclin D1 accumulation
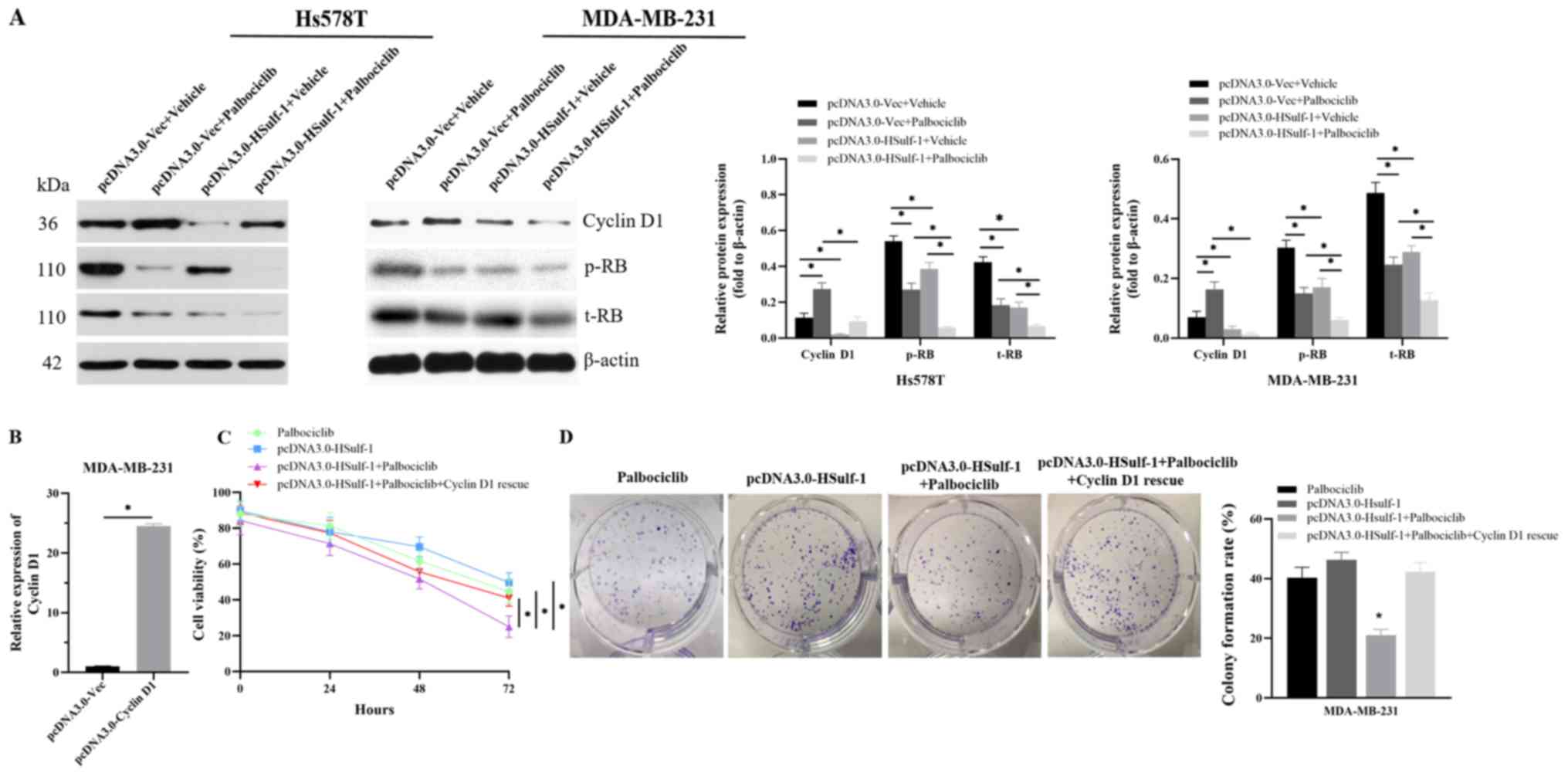
A recently published preclinical study reported that
a novel CDK4/6 inhibitor, SHR6390, increased the expression of
cyclin D1 in RB-positive MCF7 cells without affecting the
RB-negative TNBC cell line MDA-MB-468 (64). To determine if palbociclib can
exert a similar effect on RB-positive TNBC cells, cyclin D1
expression was next measured following palbociclib treatment. It
was found that following palbociclib treatment, the levels of RB
expression and phosphorylation were significantly reduced, whilst
that of cyclin D1 expression was significantly increased compared
with those treated with vehicle (Fig.
5A). In addition, palbociclib-induced cyclin D1 accumulation
was found to be significantly reduced following the overexpression
of HSulf-1, leading to significantly lower levels of total RB
expression and RB phosphorylation compared with cells transfected
with the control vector (Fig. 5A).
Finally, the possibility that the aforementioned synergistic
effects may be due to a reduction in cyclin D1 expression caused by
HSulf-1 overexpression was next considered. Therefore, a rescue
experiment was performed on MDA-MB-231 cells, where the
overexpression of cyclin D1 abolished the synergistic effects of
HSulf-1 and palbociclib treatment on cell proliferation (Fig. 5C and D). Consequently, it was
concluded that the mechanism by which HSulf-1 cooperated with
palbociclib to exert antiproliferative effects was due to the
reversal of palbociclib-induced cyclin D1 accumulation.
Discussion
TNBC is the most heterogeneous sub-type of breast
cancer, where gene expression profiling identified six distinct
molecular subgroups: Luminal androgen receptor (LAR), mesenchymal
stem-like, mesenchymal, two basal-likes and immunomodula-tory
(21,65). Due to this heterogeneity, patients
with TNBC frequently undergo a variety of clinical courses, where a
and diverse set of therapeutic responses can occur (21). Therefore, specific therapeutic
strategies should be identified based on these molecular sub-types
and clinicopathological features. A number of preclinical studies
have reported that the RB-positive LAR subgroup of TNBC is
sensitive to CDK4/6 inhibitors (43,47,66).
However, the anti-metastatic effect of palbociclib on TNBC remains
controversial. Lamb et al (48) revealed that treatment with
palbociclib increased migration and mammosphere formation and had
the potential to increase the migratory capacity of TNBC cells,
which could in turn increase metastasis and cancer recurrence.
Conversely, other studies have previously demonstrated that
palbociclib reduced the migration and invasion of TNBC cells in
addition to suppressing metastasis in a xenograft metastasis model
derived from TNBC cells (49-51).
In the present study, RB-positive TNBC cell lines were found to be
sensitive to palbociclib, which reduced tumor proliferation both
in vitro and in vivo. Furthermore, palbociclib was
found to inhibit TNBC cell migration, invasion and EMT by
upregulating E-cadherin expression whilst downregulating that of
vimentin and N-cadherin, thus further validating the antitumor
effect of palbociclib on RB-positive TNBC.
In recent years, the roles of HSulf-1 in human
cancers have attracted attention. HSulf-1 expression was reported
to be downregulated in breast cancers, which is considered an early
event in the tumorigenesis of breast cancer (37). Additionally, previous studies have
demonstrated that HSulf-1 functions as a tumor suppressor in breast
cancer; where it can be applied in predicting clinical outcomes
(36,38,39).
However, these previous studies aforementioned focused primarily on
breast cancer without considering the subtype. It is well
documented that breast cancer is a highly heterogeneous disease
with a complex mechanism of pathogenesis that exhibits a wide range
of clinical behaviours and treatment responses. Therefore, it
becomes necessary to validate the role of HSulf-1 in TNBC. The
present study was specifically performed on the RB-positive TNBC
subtype both in vitro and in vivo. Accordingly, it
was found that HSulf-1 expression was downregulated in TNBC tissues
and cells compared with their non-cancerous counterparts. Notably,
reduced HSulf-1 expression was highly predictive of poor PFS and OS
in patients with TNBC. However, no association was observed between
HSulf-1 expression and the other clinicopathological indicators
tested, including age, tumor grade, tumor size and lymph nodal
status in TNBC tissues. Collectively, these results indicate that
HSulf-1 is likely to serve as a reliable prognostic biomarker of
patient outcomes for TNBC.
Previous studies have demonstrated that HSulf-1
induces cell cycle arrest and inhibits tumorigenesis and
angiogenesis in TNBC cells in vivo (36,39).
However, the potential effects of HSulf-1 on other physiological
processes in TNBC, including apoptosis, migration and invasion,
remained unclear. In the present study, it was found that HSulf-1
overexpression inhibited tumor proliferation by inducing
G1/S and G2/M cell cycle arrest and
apoptosis, in accordance with previous reports (19,45,47).
In addition, HSulf-1 was also found to suppress migration, invasion
and EMT. Collectively, these results demonstrated that HSulf-1
serves as a tumor suppressor in TNBC. Following the discovery that
HSulf-1 reduced cyclin D1 expression to induce G1/S
arrest, the hypothesis that HSulf-1 overexpression combined with
palbociclib treatment may exhibit synergistic antitumor effects on
TNBC cells was investigated. HSulf-1 and palbociclib in combination
exerted additive antitumor effects on the induction of
G1/S cell cycle arrest, inhibition of migration,
invasion and EMT in vitro. Accordingly, this combination may
serve as a good alternative therapeutic option for RB-positive
TNBC. However, in clinical practice, a suitable vector, including
the likes of polyamidoamine dendrimers, transferrin-polyethylene
glycol-polyethylenimine and adeno-associated viral vectors, may be
required for the effective delivery of the HSulf-1 gene into tumors
for patients with TNBC in the future (67-71),
which require further clinical validation.
Several studies have previously demonstrated that
HBGFs, such as HGF binding to its corresponding receptors c-Met and
HSPGs, form a ternary complex to activate downstream tyrosine
kinases, resulting in the phosphorylation of substrate proteins,
including PI3K-AKT and Ras-MAPK. Desulfation of HSPGs by HSulf-1
prevents the formation of these ternary complexes, followed by the
inactivation of downstream signaling pathways (33,34,38,72,73).
Therefore, in the context of HBGF signaling, HSPG serves as a
core-ceptor to facilitate the interaction between the HBGFs and
their cognate transducing receptors, activating the phosphorylation
cascade. HSulf-1 inhibits the sulfation of cell-surface HSPGs and
abrogates growth factor signaling. Mechanisms underlying the
regulation of cyclin D1 by HSulf-1 have been proposed. Liu et
al (56) previously documented
that HSulf-1 downregulates cyclin D1 expression by suppressing
STAT3 signaling in hepatocellular carcinoma. Notably, the
CCND1 gene was demonstrated to be a target of STAT3, as
STAT3-binding sites were identified in the CCND1 promoter
(74). In the present study,
HSulf-1 was identified to regulate cyclin D1 expression via
non-canonical AKT/STAT3 and ERK1/2/STAT3 signaling instead of the
canonical JAK2/STAT3 pathway. In addition, overexpres-sion of
HSulf-1 prevented the apparent increase in cyclin D1 levels caused
by palbociclib treatment. Therefore, data from the present study
hinted at a potential crosstalk between the
HSulf-1/ERK1/2/AKT/STAT3 and CDK-RB-E2F pathways to control cell
cycle progression (Fig. 6).
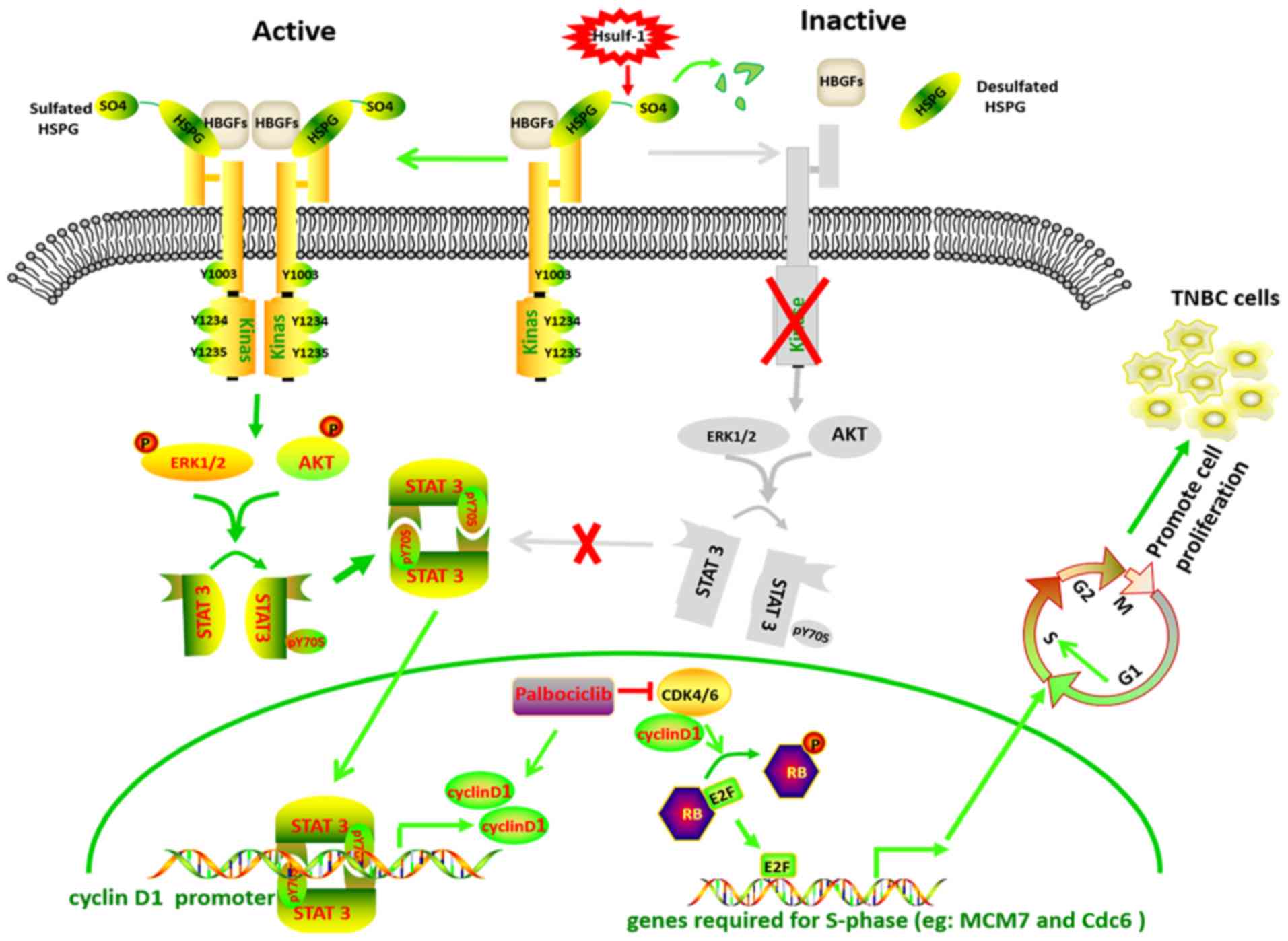 | Figure 6Schematic diagram depicting the
possible mechanism of HSulf-1-mediated inactivation of STAT3 via
the PI3K/AKT and Ras/ERK1 pathways and crosstalk with CDK-RB-E2F
signaling in TNBC cells. The initiation of HBGF signaling requires
the binding of extracellular HBGF (FGF, EGF, HGF) and its cognate
tyrosine kinase receptor to HSPGs to form a ternary
ligand-HSPG-receptor complex at the cell surface. Ligand binding
induces receptor dimerization and subsequent phosphorylation of the
receptor tyrosine kinase, leading to a downstream phosphorylation
cascade including the PI3K/AKT/STAT3 and Ras/ERK1/2/STAT3 pathways.
Phosphorylated STAT3 forms a dimer and translocates into the
nucleus, promoting cyclin D1 expression. Desulfation of HSPGs by
HSulf-1 prevents formation of the ternary complex, leading to a
reduction in HBGF signaling and cyclin D1 expression. Hsulf-1,
human sulfatase 1; CDK, cyclin-dependent kinase; RB,
retinoblastoma; YNBC, triple-negative breast cancer; HBGF,
heparin-binding growth factors; HSPG, heparan sulfate
proteoglycans; MCM7, minichromosome maintenance complex component
7; cdc6, cell division cycle 6; EGF, epidermal growth factor; FGF,
fibroblast growth factor; HGF, hepatocyte growth factor. |
A number of preclinical studies have reported that
ER-positive breast cancer cells adapt rapidly to CDK4/6 inhibition
to evade cytostasis, in part via non-canonical cyclin
D1-CDK2-mediated S phase entry (75). This adaptation was found to be
prevented by combining CDK4/6 inhibitors with PI3K/mTOR inhibitors,
which reduced the levels of cyclin D1 and other G1-S
phase cyclins (75,76). In the present study, it was also
found that the levels of cyclin D1 were increased following
palbociclib treatment, consistent with previously reported results
(64). This may be due to an
adaptive response to damaging stimuli or CDK4/6 inhibitors have a
similar negative feedback effect of preventing CDK4/6 to form a
complex with cyclin D1, which may increase cyclin D1 expression and
further activate the crosstalk between the cyclin D1-CDK2 signaling
pathway, which require further study (77). It was previously revealed that
STAT3 phos-phorylation was significantly increased in
palbociclib-resistant ER-positive breast cancer cells, where
ER-positive metastatic breast cancer patients acquired resistance
after treatment with Palbociclib (78). Furthermore, palbociclib-resistant
cells appear to be more sensitive to the STAT3 inhibitor TTI-010,
which directly targets the tyrosine-phosphorylated peptide-binding
pocket within the STAT3 SH2 domain (78). Based on these observations, HSulf-1
may have the potential to prevent the early adaptation and prolong
the durability of palbociclib efficacy in TNBC patients.
Although the present study demonstrated that HSulf-1
over-expression in conjunction with palbociclib has a synergistic
antitumor effect in RB-positive TNBC cells. Only the synergistic
anti-proliferative mechanism by which HSulf-1 reduced
palbociclib-induced cyclin D1 accumulation was clarified.
Additionally, although it was demonstrated that HSulf-1 inhibited
cyclin D1 expression through the ERK1/2/STAT3 and AKT/STAT3
pathways, it remains unclear whether HSulf-1 could suppresses
cyclin D1 expression through other pathways. For the wound healing
experiments, it was found that the majority of cells died after 24
h incubation in medium without FBS following scratching a wound
introduction. Therefore, medium containing 2% FBS was used, where
little or no cell proliferation or cell death were observed.
Accordingly, further studies are required to explore these
possibilities.
In conclusion, the present study demonstrated that
HSulf-1 expression is downregulated in TNBC tissues and cells,
which serves an anti-oncogenic role by inducing cell cycle arrest
and apoptosis in addition to preventing proliferation, EMT,
migration and invasion in TNBC. In addition, it was found that
downregulation of HSulf-1 was associated with poor clinical
outcomes in patients with TNBC. HSulf-1 inhibited cyclin D1
expression via the non-canonical AKT/STAT3 and ERK1/2/STAT3
signaling pathways in TNBC. In particular, the combination of
HSulf-1 overexpression and treatment with the CDK4/6 inhibitor
palbociclib exerted additive, synergistic antitumor effects on
RB-positive TNBC, which may serve as an effective alternative
therapeutic option for patients with RB-positive TNBC.
Supplementary Data
Funding
This work was supported by the National Natural
Science Foundation of China (grant no. 81904231 to FFP) and the
Health and Family Planning Commission of Wuhan Municipality (grant
no. WX17Q38 to FXC).
Availability of data and materials
The datasets generated during the current study are
available from the corresponding author on reasonable request.
Authors' contributions
FP and FC conceived the study and participated in
its design and coordination. YY and ZZ performed the experiments
and contributed to data collection. FP and FC analyzed the data and
drafted the manuscript. ZZ and QL assisted in designing experiments
and provided technical expertise in conducting experiments and
reviewed the manuscript. All authors read and approved the final
version of the manuscript.
Ethics approval and consent to
participate
The use of patient tissue samples was approved by
the Ethics Committee of the General Hospital of The Yangtze River
Shipping, Wuhan Polytechnic University (approval no. 2017IEC0003;
Wuhan, China). Informed consent was obtained from all patients.
Animal experiments were performed with the approval of the Animal
Care and Use Committee of the General Hospital of The Yangtze River
Shipping, Wuhan Polytechnic University (approval no. 2017IEC0003;
Wuhan, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Perou CM, Sørlie T, Eisen MB, Van De Rijn
M, Jeffrey SS, Rees CA, Pollack JR, Ross DT, Johnsen H, Akslen LA,
et al: Molecular portraits of human breast tumours. Nature.
406:747–752. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cancer Genome Atlas Network: Comprehensive
molecular portraits of human breast tumours. Nature. 490:61–70.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rakha EA, El-Rehim DA, Paish C, Green AR,
Lee AH, Robertson JF, Blamey RW, Macmillan D and Ellis IO: Basal
phenotype identifies a poor prognostic subgroup of breast cancer of
clinical importance. Eur J Cancer. 42:3149–3156. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Haffty BG, Yang Q, Reiss M, Kearney T,
Higgins SA, Weidhaas J, Harris L, Hait W and Toppmeyer D:
Locoregional relapse and distant metastasis in conservatively
managed triple negative early-stage breast cancer. J Clin Oncol.
24:5652–5657. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dent R, Trudeau M, Pritchard KI, Hanna WM,
Kahn HK, Sawka CA, Lickley LA, Rawlinson E, Sun P and Narod SA:
Triple-negative breast cancer: Clinical features and patterns of
recurrence. Clin Cancer Res. 13:4429–4434. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liedtke C, Mazouni C, Hess KR, André F,
Tordai A, Mejia JA, Symmans WF, Gonzalez-Angulo AM, Hennessy B,
Green M, et al: Response to neoadjuvant therapy and long-term
survival in patients with triple-negative breast cancer. J Clin
Oncol. 26:1275–1281. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Carey LA, Dees EC, Sawyer L, Gatti L,
Moore DT, Collichio F, Ollila DW, Sartor CI, Graham ML and Perou
CM: The triple negative paradox: Primary tumor chemosensitivity of
breast cancer subtypes. Clin Cancer Res. 13:2329–2334. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Robson M, Im SA, Senkus E, Xu B, Domchek
SM, Masuda N, Delaloge S, Li W, Tung N, Armstrong A, et al:
Olaparib for metastatic breast cancer in patients with a germline
BRCA mutation. N Engl J Med. 377:523–533. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Winter C, Nilsson MP, Olsson E, George AM,
Chen Y, Kvist A, Törngren T, Vallon-Christersson J, Hegardt C,
Häkkinen J, et al: Targeted sequencing of BRCA1 and BRCA2 across a
large unselected breast cancer cohort suggests that one-third of
mutations are somatic. Ann Oncol. 27:1532–1538. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Atchley DP, Albarracin CT, Lopez A, Valero
V, Amos CI, Gonzalez-Angulo AM, Hortobagyi GN and Arun BK: Clinical
and pathologic characteristics of patients with BRCA-positive and
BRCA-negative breast cancer. J Clin Oncol. 26:4282–4288. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
O'Leary B, Finn RS and Turner NC: Treating
cancer with selective CDK4/6 inhibitors. Nat Rev Clin Oncol.
13:417–430. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Finn RS, Crown JP, Lang I, Boer K,
Bondarenko IM, Kulyk SO, Ettl J, Patel R, Pinter T, Schmidt M, et
al: The cyclin-dependent kinase 4/6 inhibitor palbociclib in
combination with letrozole versus letrozole alone as first-line
treatment of oestrogen receptor-positive, HER2-negative, advanced
breast cancer (PALOMA-1/TRIO-18): A randomised phase 2. study
Lancet Oncol. 16:25–35. 2015. View Article : Google Scholar
|
|
14
|
Turner NC, Ro J, André F, Loi S, Verma S,
Iwata H, Harbeck N, Loibl S, Huang Bartlett C, Zhang K, et al:
Palbociclib in hormone-receptor-positive advanced breast cancer. N
Engl J Med. 373:209–219. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Finn RS, Martin M, Rugo HS, Jones S, Im
SA, Gelmon K, Harbeck N, Lipatov ON, Walshe JM, Moulder S, et al:
Palbociclib and letrozole in advanced breast cancer. N Engl J Med.
375:1925–1936. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hortobagyi GN, Stemmer SM, Burris HA, Yap
YS, Sonke GS, Paluch-Shimon S, Campone M, Blackwell KL, André F,
Winer EP, et al: Ribociclib as first-line therapy for HR-positive,
advanced breast cancer. N Engl J Med. 375:1738–1748. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tripathy D, Im SA, Colleoni M, Franke F,
Bardia A, Harbeck N, Hurvitz SA, Chow L, Sohn J, Lee KS, et al:
Ribociclib plus endocrine therapy for premenopausal women with
hormone-receptor-positive, advanced breast cancer (MONALEESA-7): A
randomised phase 3 trial. Lancet Oncol. 19:904–915. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sledge GW Jr, Toi M, Neven P, Sohn J,
Inoue K, Pivot X, Burdaeva O, Okera M, Masuda N, Kaufman PA, et al:
MONARCH 2: Abemaciclib in combination with fulvestrant in women
with HR+/HER2-advanced breast cancer who had progressed while
receiving endocrine therapy. J Clin Oncol. 35:2875–2884. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Finn RS, Dering J, Conklin D, Kalous O,
Cohen DJ, Desai AJ, Ginther C, Atefi M, Chen I, Fowst C, et al: PD
0332991, a selective cyclin D kinase 4/6 inhibitor, preferentially
inhibits proliferation of luminal estrogen receptor-positive human
breast cancer cell lines in vitro. Breast Cancer Res. 11:R772009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
DeMichele A, Clark AS, Tan KS, Heitjan DF,
Gramlich K, Gallagher M, Lal P, Feldman M, Zhang P, Colameco C, et
al: CDK 4/6 inhibitor palbociclib (PD0332991) in Rb+ advanced
breast cancer: Phase II activity, safety, and predictive biomarker
assessment. Clin Cancer Res. 21:995–1001. 2015. View Article : Google Scholar
|
|
21
|
Lehmann BD, Bauer JA, Chen X, Sanders ME,
Chakravarthy AB, Shyr Y and Pietenpol JA: Identification of human
triple-negative breast cancer subtypes and preclinical models for
selection of targeted therapies. J Clin Invest. 121:2750–2767.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Witkiewicz AK and Knudsen ES:
Retinoblastoma tumor suppressor pathway in breast cancer:
Prognosis, precision medicine, and therapeutic interventions.
Breast Cancer Res. 16:2072014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Treré D, Brighenti E, Donati G, Ceccarelli
C, Santini D, Taffurelli M, Montanaro L and Derenzini M: High
prevalence of retinoblastoma protein loss in triple-negative breast
cancers and its association with a good prognosis in patients
treated with adjuvant chemotherapy. Ann Oncol. 20:1818–1823. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Witkiewicz AK, Ertel A, McFalls J,
Valsecchi ME, Schwartz G and Knudsen ES: RB-pathway disruption is
associated with improved response to neoadjuvant chemotherapy in
breast cancer. Clin Cancer Res. 18:5110–5122. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lamanna WC, Frese MA, Balleininger M and
Dierks T: Sulf loss influences N-, 2-O-, and 6-O-sulfation of
multiple heparan sulfate proteoglycans and modulates fibroblast
growth factor signaling. J Biol Chem. 283:27724–27735. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dhoot GK, Gustafsson MK, Ai X, Sun W,
Standiford DM and Emerson CP Jr: Regulation of Wnt signaling and
embryo patterning by an extracellular sulfatase. Science.
293:1663–1666. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Takashima Y, Keino-Masu K, Yashiro H, Hara
S, Suzuki T, van Kuppevelt TH, Masu M and Nagata M: Heparan sulfate
6-O-endosulfatases, Sulf1 and Sulf2, regulate glomerular integrity
by modulating growth factor signaling. Am J Physiol Renal Physiol.
310:F395–F408. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen Z, Fan JQ, Li J, Li QS, Yan Z, Jia
XK, Liu WD, Wei LJ, Zhang FZ, Gao H, et al: Promoter
hypermethylation correlates with the Hsulf-1 silencing in human
breast and gastric cancer. Int J Cancer. 124:739–744. 2009.
View Article : Google Scholar
|
|
29
|
Staub J, Chien J, Pan Y, Qian X, Narita K,
Aletti G, Scheerer M, Roberts LR, Molina J and Shridhar V:
Epigenetic silencing of HSulf-1 in ovarian cancer: Implications in
chemoresistance. Oncogene. 26:4969–4978. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lai JP, Sandhu DS, Shire AM and Roberts
LR: The tumor suppressor function of human sulfatase 1 (SULF1) in
carcinogenesis. J Gastrointest Cancer. 39:149–158. 2008. View Article : Google Scholar
|
|
31
|
Marotta LL, Almendro V, Marusyk A,
Shipitsin M, Schemme J, Walker SR, Bloushtain-Qimron N, Kim JJ,
Choudhury SA, Maruyama R, et al: The JAK2/STAT3 signaling pathway
is required for growth of CD44(+)CD24(-) stem cell-like breast
cancer cells in human tumors. J Clin Invest. 121:2723–2735. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dai Y, Yang Y, MacLeod V, Yue X, Rapraeger
AC, Shriver Z, Venkataraman G, Sasisekharan R and Sanderson RD:
HSulf-1 and HSulf-2 are potent inhibitors of myeloma tumor growth
in vivo. J Biol Chem. 280:40066–40073. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lai JP, Chien JR, Moser DR, Staub JK,
Aderca I, Montoya DP, Matthews TA, Nagorney DM, Cunningham JM,
Smith DI, et al: hSulf1 Sulfatase promotes apoptosis of
hepatocellular cancer cells by decreasing heparin-binding growth
factor signaling. Gastroenterology. 126:231–248. 2004. View Article : Google Scholar
|
|
34
|
Lai JP, Chien J, Strome SE, Staub J,
Montoya DP, Greene EL, Smith DI, Roberts LR and Shridhar V: HSulf-1
modulates HGF-mediated tumor cell invasion and signaling in head
and neck squamous carcinoma. Oncogene. 23:1439–1447. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Roy D, Mondal S, Wang C, He X, Khurana A,
Giri S, Hoffmann R, Jung DB, Kim SH, Chini EN, et al: Loss of
HSulf-1 promotes altered lipid metabolism in ovarian cancer. Cancer
Metab. 2:132014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Narita K, Staub J, Chien J, Meyer K, Bauer
M, Friedl A, Ramakrishnan S and Shridhar V: HSulf-1 inhibits
angiogenesis and tumorigenesis in vivo. Cancer Res. 66:6025–6032.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lai J, Chien J, Staub J, Avula R, Greene
EL, Matthews TA, Smith DI, Kaufmann SH, Roberts LR and Shridhar V:
Loss of HSulf-1 up-regulates heparin-binding growth factor
signaling in cancer. J Biol Chem. 278:23107–23117. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Khurana A, Liu P, Mellone P, Lorenzon L,
Vincenzi B, Datta K, Yang B, Linhardt RJ, Lingle W, Chien J, et al:
HSulf-1 modulates FGF2- and hypoxia-mediated migration and invasion
of breast cancer cells. Cancer Res. 71:2152–2161. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Narita K, Chien J, Mullany SA, Staub J,
Qian X, Lingle WL and Shridhar V: Loss of HSulf-1 expression
enhances autocrine signaling mediated by amphiregulin in breast
cancer. J Biol Chem. 282:14413–14420. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Böcker W: WHO Classification of Breast
Tumors and Tumors of the Female Genital Organs: Pathology and
Genetics. Verh Dtsch Ges Pathol. 86:116–119. 2002.
|
|
41
|
Amin MB, Edge S, Greene F, Byrd DR,
Brookland RK, Washington MK, Gershenwald JE, Compton CC, Hess KR,
Sullivan DC, et al: AJCC cancer staging manual. 8th ed. New York:
Springer; 2017, View Article : Google Scholar
|
|
42
|
Dean JL, McClendon AK and Knudsen ES:
Modification of the DNA damage response by therapeutic CDK4/6
inhibition. J Biol Chem. 287:29075–29087. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Asghar US, Barr AR, Cutts R, Beaney M,
Babina I, Sampath D, Giltnane J, Lacap JA, Crocker L, Young A, et
al: Single-cell dynamics determines response to CDK4/6 inhibition
in triple-negative breast cancer. Clin Cancer Res. 23:5561–5572.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
45
|
Yamamoto T, Kanaya N, Somlo G and Chen S:
Synergistic anti-cancer activity of CDK4/6 inhibitor palbociclib
and dual mTOR kinase inhibitor MLN0128 in pRb-expressing
ER-negative breast cancer. Breast Cancer Res Treat. 174:615–625.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Foidart P, Yip C, Radermacher J, Blacher
S, Lienard M, Montero-Ruiz L, Maquoi E, Montaudon E,
Château-Joubert S, Collignon J, et al: Expression of MT4-MMP, EGFR,
and RB in triple-negative breast cancer strongly sensitizes tumors
to erlotinib and palbociclib combination therapy. Clin Cancer Res.
25:1838–1850. 2019. View Article : Google Scholar
|
|
47
|
Liu CY, Lau KY, Hsu CC, Chen JL, Lee CH,
Huang TT, Chen YT, Huang CT, Lin PH and Tseng LM: Combination of
palbociclib with enzalutamide shows in vitro activity in RB
proficient and androgen receptor positive triple negative breast
cancer cells. PLoS One. 12:e01890072017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lamb R, Lehn S, Rogerson L, Clarke RB and
Landberg G: Cell cycle regulators cyclin D1 and CDK4/6 have
estrogen receptor-dependent divergent functions in breast cancer
migration and stem cell-like activity. Cell cycle. 12:2384–2394.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhong Z, Yeow WS, Zou C, Wassell R, Wang
C, Pestell RG, Quong JN and Quong AA: Cyclin D1/cyclin-dependent
kinase 4 interacts with filamin A and affects the migration and
invasion potential of breast cancer cells. Cancer Res.
70:2105–2114. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Liu T, Yu J, Deng M, Yin Y, Zhang H, Luo
K, Qin B, Li Y, Wu C, Ren T, et al: CDK4/6-dependent activation of
DUB3 regulates cancer metastasis through SNAIL1. Nat Commun.
8:139232017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Qin G, Xu F, Qin T, Zheng Q, Shi D, Xia W,
Tian Y, Tang Y, Wang J, Xiao X, et al: Palbociclib inhibits
epithelial-mesenchymal transition and metastasis in breast cancer
via c-Jun/COX-2 signaling pathway. Oncotarget. 6:41794–41808. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Tsai JH and Yang J: Epithelial-mesenchymal
plasticity in carcinoma metastasis. Genes Dev. 27:2192–2206. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Kang Y and Massague J:
Epithelial-mesenchymal transitions: Twist in development and
metastasis. Cell. 118:277–279. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Ye X and Weinberg RA:
Epithelial-mesenchymal plasticity: A central regulator of cancer
progression. Trends Cell Biol. 25:675–686. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Liu L, Ding F, Chen J, Wang B and Liu Z:
hSulf-1 inhibits cell proliferation and migration and promotes
apoptosis by suppressing stat3 signaling in hepatocellular
carcinoma. Oncol Lett. 7:963–969. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Banerjee K and Resat H: Constitutive
activation of STAT3 in breast cancer cells: A review. Int J Cancer.
138:2570–2578. 2016. View Article : Google Scholar :
|
|
58
|
Bao L, Yan Y, Xu C, Ji W, Shen S, Xu G,
Zeng Y, Sun B, Qian H, Chen L, et al: MicroRNA-21 suppresses PTEN
and hSulf-1 expression and promotes hepatocellular carcinoma
progression through AKT/ERK pathways. Cancer Lett. 337:226–236.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Liu H, Fu X, Ji W, Liu K, Bao L, Yan Y, Wu
M, Yang J and Su C: Human sulfatase-1 inhibits the migration and
proliferation of SMMC-7721 hepatocellular carcinoma cells by
downregulating the growth factor signaling. Hepatol Res.
43:516–525. 2013. View Article : Google Scholar
|
|
60
|
Xu G, Ji W, Su Y, Xu Y, Yan Y, Shen S, Li
X, Sun B, Qian H, Chen L, et al: Sulfatase 1 (hSulf-1) reverses
basic fibroblast growth factor-stimulated signaling and inhibits
growth of hepatocellular carcinoma in animal model. Oncotarget.
5:5029–5039. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Zhang H, Newman DR and Sannes PL: HSULF-1
inhibits ERK and AKT signaling and decreases cell viability in
vitro in human lung epithelial cells. Respir Res. 13:692012.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Corsetti G, Yuan Z, Romano C,
Chen-Scarabelli C, Fanzani A, Pasini E, Dioguardi FS, Onorati F,
Linardi D, Knight R, et al: Urocortin induces phosphorylation of
distinct residues of signal transducer and activator of
transcription 3 (STAT3) via different signaling pathways. Med Sci
Monit Basic Res. 25:139–152. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Malanga D, De Marco C, Guerriero I,
Colelli F, Rinaldo N, Scrima M, Mirante T, De Vitis C, Zoppoli P,
Ceccarelli M, et al: The Akt1/IL-6/STAT3 pathway regulates growth
of lung tumor initiating cells. Oncotarget. 6:42667–42686. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Long F, He Y, Fu H, Li Y, Bao X, Wang Q,
Wang Y, Xie C and Lou L: Preclinical characterization of SHR6390, a
novel CDK 4/6 inhibitor, in vitro and in human tumor xenograft
models. Cancer Sci. 110:1420–1430. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Burstein MD, Tsimelzon A, Poage GM,
Covington KR, Contreras A, Fuqua SA, Savage MI, Osborne CK,
Hilsenbeck SG, Chang JC, et al: Comprehensive genomic analysis
identifies novel subtypes and targets of triple-negative breast
cancer. Clin Cancer Res. 21:1688–1698. 2015. View Article : Google Scholar :
|
|
66
|
Asghar U, Herrera-Abreu MT, Cutts R,
Babina I, Pearson A and Turner NC: Identification of subtypes of
triple negative breast cancer (TNBC) that are sensitive to CDK4/6
inhibition. J Clin Oncol. 33:110982015. View Article : Google Scholar
|
|
67
|
Rein DT, Breidenbach M and Curiel DT:
Current developments in adenovirus-based cancer gene therapy.
Future Oncol. 2:137–143. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Maruyama TH, Harada Y, Matsumura T, Satoh
E, Cui F, Iwai M, Kita M, Hibi S, Imanishi J, Sawada T and Mazda O:
Effective suicide gene therapy in vivo by EBV-based plasmid vector
coupled with polyamidoamine dendrimer. Gene Ther. 7:53–60. 2000.
View Article : Google Scholar
|
|
69
|
Kursa M, Walker GF, Roessler V, Ogris M,
Roedl W, Kircheis R and Wagner E: Novel shielded
transferrin-polyethylene glycol-polyethylenimine/DNA complexes for
systemic tumor-targeted gene transfer. Bioconjug Chem. 14:222–231.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Griffin JM, Fackelmeier B, Clemett CA,
Fong DM, Mouravlev A, Young D and O'Carroll SJ: Astrocyte-selective
AAV-ADAMTS4 gene therapy combined with hindlimb rehabilitation
promotes functional recovery after spinal cord injury. Exp Neurol.
327:1132322020. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Salameh JW, Zhou L, Ward SM, Chalaarca CF,
Emrick T and Figueiredo ML: Polymer-mediated gene therapy: Recent
advances and merging of delivery techniques. Wiley Interdiscip Rev
Nanomed Nanobiotechnol. 1:e15982020.
|
|
72
|
Selva EM and Perrimon N: Role of heparan
sulfate proteoglycans in cell signaling and cancer. Adv Cancer Res.
83:67–80. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Rubin JS, Day RM, Breckenridge D, Atabey
N, Taylor WG, Stahl SJ, Wingfield PT, Kaufman JD, Schwall R and
Bottaro DP: Dissociation of heparan sulfate and receptor binding
domains of hepatocyte growth factor reveals that heparan
sulfate-c-met interaction facilitates signaling. J Biol Chem.
276:32977–32983. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Bromberg JF, Wrzeszczynska MH, Devgan G,
Zhao Y, Pestell RG, Albanese C and Darnell JE Jr: Stat3 as an
oncogene. Cell. 98:295–303. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Herrera-Abreu MT, Palafox M, Asghar U,
Rivas MA, Cutts RJ, Garcia-Murillas I, Pearson A, Guzman M,
Rodriguez O, Grueso J, et al: Early adaptation and acquired
resistance to CDK4/6 inhibition in estrogen receptor-positive
breast cancer. Cancer Res. 76:2301–2313. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Michaloglou C, Crafter C, Siersbaek R,
Delpuech O, Curwen JO, Carnevalli LS, Staniszewska AD, Polanska UM,
Cheraghchi-Bashi A, Lawson M, et al: Combined inhibition of mTOR
and CDK4/6 is required for optimal blockade of E2F function and
long-term growth inhibition in estrogen receptor-positive breast
cancer. Mol Cancer Ther. 17:908–920. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Javle MM, Shroff RT, Xiong H, Varadhachary
GA, Fogelman D, Reddy SA, Davis D, Zhang Y, Wolff RA and Abbruzzese
JL: Inhibition of the mammalian target of rapamycin (mTOR) in
advanced pancreatic cancer: Results of two phase II studies. BMC
Cancer. 10:3682010. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Kettner NM, Vijayaraghavan S, Durak MG,
Bui T, Kohansal M, Ha MJ, Liu B, Rao X, Wang J, Yi M, et al:
Combined inhibition of STAT3 and DNA repair in
palbociclib-resistant ER-positive breast cancer. Clin Cancer Res.
25:3996–4013. 2019. View Article : Google Scholar : PubMed/NCBI
|