Introduction
Prostate cancer is the most common malignant tumor
among men in developed countries (1). In recent years, prostate cancer has
contributed to a high morbidity and mortality in most native Asian
populations, with >297,000 new prostate cancer cases diagnosed
in Asia, accounting for 23.3% of the global incidence and ~33% of
deaths occurring in Asia in 2018 (2,3).
Currently, prostate gland removal and androgen blockade is an
effective treatment option for the majority of patients with
prostate cancer; however, a number of patients experience hormone
resistance, and prostatectomy is dangerous for older patients with
cardiovascular disease (4).
Hypoxia is a common biological feature of solid tumors, usually
associated with cancer progression, treatment resistance, and poor
prognosis (5). Hypoxia reportedly
triggers expression of adherens junction molecules to promote the
invasion and epithelial-mesenchymal transition (EMT) of prostate
cancer cells (6,7). Therefore, it is necessary to
elucidate the molecular mechanisms of prostate cancer progression
and explore novel targets for regulating its progression under
hypoxic conditions.
MicroRNAs (miRs) serve complicated roles in
pathophysiological tumor processes; for example, miR-150 regulates
TRPM4-mediated β-catenin pathway to impede the progression of
prostate cancer (8-10). Additionally, miRs serve as
prognostic markers of tumors or potential therapeutic targets
(11,12). miR-383-5p has been demonstrated to
be down-regulated in lung adenocarcinoma tissues and to be
associated with tumor size and differentiation; in addition,
miR-383-5p exerts an antiproliferative function, suggesting that it
may serve as a potential novel potential prognostic biomarker and
therapeutic target for lung adenocarcinoma (13). Aberrant expression of miR is
associated with dysregulation of cancer cell proliferation,
apoptosis, and migration. For example, miR-181a and miR-483-5p are
overexpressed in prostate cancer, leading to prostate cancer growth
(14,15). However, miR-129 is down-regulated
in prostate cancer tissues and cell lines, and restoring miR-129
expression reduces the viability, proliferation, and migration of
PC3 cells (16). A recent study
has demonstrated that miR-150 inhibits EMT in prostate cancer cells
(10). Patients with prostate
cancer recurrence following radical prostatectomy present with
reduced miR-137 in prostatectomy specimens (17). Li et al (18) have reported that miR-137 expression
is decreased in the mouse brain under hypoxic conditions. However,
the potential roles of miR-137 in hypoxia-mediated migration and
EMT of prostate cancer cells remain unclear.
The aim of the present study was to clarify the
function of miR-137 in prostate cancer cell migration, invasion and
EMT, as well as to elucidate the underlying mechanism under hypoxic
conditions in prostate cancer cells. This may help determine
whether miR-137 may serve as a potential therapeutic target for the
treatment of prostate cancer and provide a new mechanism of
hypoxia-induced migration, invasion and EMT.
Materials and methods
Cell culture
PC3 and DU145 cells were obtained from the American
Type Culture Collection (ATCC, Rockville, MD, USA). The cells were
cultured in Dulbecco's Modified Eagle's medium (DMEM; Gibco; Thermo
Fisher Scientific, Inc.) supplemented with 10% fetal bovine serum
(HyClone; Cytiva) in an incubator containing 95% air and 5%
CO2 at 37°C.
Cell transfection and hypoxia
treatment
The miR-137 mimic, miR-137 inhibitor, and miR-137
negative control (NC) were designed by and synthetized from
Shanghai GenePharma Co., Ltd. Leucine-rich repeat-containing G
protein-coupled receptor (4LGR4)-specific siRNA (siLGR4) and the
siRNA control (siCtrl) were obtained from Shanghai GenePharma Co.,
Ltd. PC3 and DU145 cells (2.5×105 cells/well) were
seeded in 6-well plates overnight and transfected with 50 pmol NC,
miR-137 mimic or miR-137 inhibitor using Lipofectamine®
2000 (Invitrogen; Thermo Fisher Scientific, Inc.), as well as 50
pmol siLGR4. The sequences used were as follows: miR-137 mimic,
5'-UUA UUG CUU AAG AAU ACG CGU AG-3'; miR-137 inhibitor, 5'-CUA CGC
GUA UUC UUA AGC AAU AA-3'; NC for miR-137, 5'-UCA CAA CCU CCU AGA
AAG AGU AGA-3'; siLGR4, 5'-GGU AAG AAA CUC CUA AUU AUU-3'; siCtrl,
5'-TTC TCC GAA CGT GTC ACG T-3'. Cells were cultured for 4-6 h at
37°C, and the medium was replaced with complete DMEM. At 48 h
post-transfection, cells were exposed to 24-h hypoxia (H; 1%
O2, 5% CO2 and 94% N2) in an
incubator (BioSpherix, Ltd.). For epidermal growth factor receptor
(EGFR) inhibition, 100 nM AG1478 (EMD Millipore) was added to the
cells 1 h prior to hypoxia.
Cell viability analysis
The viability of PC3 and DU145 cells was detected
using the Cell Counting Kit-8 (CCK-8) assay (Beyotime Institute of
Biotechnology). Cells (5×103 cells/well) were seeded in
a 96-well plate, transfected with the miR-137 mimic or inhibitor
and exposed to hypoxia. CCK-8 reagent was added to the cells (10
µl/well) and incubated for 2 h at 37°C. The absorbance of each well
was recorded at 450 nm by a microplate reader.
Apoptosis assessment
The Cell Death Detection ELISA PLUS kit (Roche
Diagnostics, GmbH) was used to evaluate the apoptosis of DU145 and
PC3 cells according to the manufacturer's instructions. In brief,
transfected cells (1×104 cells/well) were seeded in
96-well plates and subjected to hypoxic conditions. The cells were
lysed and centrifuged, and the supernatant was transferred into a
streptavidin-coated microplate and incubated with the immunoreagent
for 2 h at room temperature. Absorbance of each well was recorded
at 405 nm.
Transwell assay
A Transwell assay was used to measure migration and
invasion of PC3 and DU145 cells. For the migration assay, cells
(1.5×105 cells/well) were seeded in serum-free DMEM and
plated into the upper chamber of a 24-well Transwell plate with
8.0-µm pores (BD Biosciences). Subsequently, 600 µl DMEM
supplemented with 20% fetal bovine serum was added to the lower
chamber. After 24-h incubation at 37°C, the medium was discarded,
and the cells were washed with phosphate-buffered saline and fixed
with 4% paraformaldehyde at room temperature for 30 min. The cells
were stained with 0.1% crystal violet at room temperature for 20
min. Images of cells in five random fields (magnification, x200)
were captured and the cells were counted under an optical inverted
microscope. For the invasion assay, the cells (2.5×105
cells/well) were seeded into the upper chamber of a Transwell plate
pre-coated with Matrigel (BD Biosciences) at 37°C for 30 min, and
the protocol was the same as that of the migration assay.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA of PC3 and DU145 cells was extracted using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. A spectrophotometer
(Thermo Fisher Scientific) was used to quantify the concentration
of RNA. A PrimeScript II 1st strand cDNA Synthesis Kit (Takara Bio,
Inc.) was used to reverse-transcribe the RNA into cDNA at 42°C for
60 min and 70°C for 5 min. Quantitative PCR was performed with an
iQ SYBR® Green Super Mix (BioRad Laboratories, Inc.) on
an ABI PRISM 7000 Fluorescent Quantitative PCR System (Applied
Biosystems, Inc.). The thermocycling conditions were as follows:
95°C for 5 min; followed by 40 cycles of 95°C for 5 sec and 60°C
for 30 sec. U6 or GAPDH was used as the internal control, and the
expression levels of miR-137 or mRNAs were quantified using the
2-ΔΔCq method (19).
The primer sequences were as follows: miR-137 forward, 5'-TAT TGC
TTA AGA ATA CGC GTA G-3' and reverse, 5'-AAC TCC AGC AGG ACC ATG
TGA T-3'; U6 forward, 5'-CTC GCT TCG GCA GCA CA-3' and reverse,
5'-AAC GCT TCA CGA ATT TGC GT-3'; LGR4 forward, 5'-CTT TGT TTG CCA
TTT CCT A-3' and reverse, 5'-CTA GTG AGT TTA ATA GCA CTA A-3';
N-cadherin forward, 5'-CAT CCC TCC AAT CAA CTT GC-3' and reverse,
5'-ATG TGC CCT CAA ATG AAA CC-3'; Vimentin forward, 5'-TGT CCA AAT
CGA TGT GGA TGT TTC-3' and reverse, 5'-TTG TAC CAT TCT TCT GCC TCC
TG-3'; MMP2 forward, 5'-TCT TCA AGG ACC GGT TCA TTT G-3' and
reverse, 5'-GAT GCT TCC AAA CTT CAC GCT C-3'; GAPDH forward, 5'-GGA
GCG AGA TCC CTC CAA AAT-3' and reverse, 5'-GGC TGT TGT CAT ACT TCT
CAT GG-3'.
Western blotting
Total protein was collected using a RIPA lysis
buffer (Beyotime Institute of Biotechnology) according to
manufacturer's protocol. The concentration of each sample was
determined by BCA Protein Assay kit (Beyotime Institute of
Biotechnology). Equal amounts of protein from each sample (50
µg/lane) were separated by 6 or 10% SDS-PAGE and transferred to a
polyvinylidene fluoride membrane. The membrane was blocked with 5%
bovine serum albumin (Amresco, LLC) and then incubated with primary
antibodies against LGR4 (cat. no. ab137480; 1:1,000), N-cadherin
(cat. no. ab76057; 1:1,000), vimentin (cat. no. ab92547; 1:1,000;
all from Abcam), MMP2 (cat. no. 40994; 1:1,000), phosphorylated
(p)-EGFR (cat. no. 3777; 1:1,000), EGFR (cat. no. 4267; 1:1,000),
p-ERK (cat. no. 4370; 1:1,000), ERK (cat. no. 4695; 1:1,000; all
from Cell Signaling Technology, Inc.), β-actin (cat. no. sc-58673;
1:5,000) and GAPDH (cat. no. sc-47724; 1:5,000; both from Santa
Cruz Biotechnology, Inc.) overnight at 4°C. The membranes were
washed with 0.1% Tween in TBS (Boster Biological Technology) and
incubated with horseradish peroxidase-conjugated goat anti-rabbit
IgG (cat. no. ZDR-5306; 1:10,000) or goat anti-mouse IgG (cat. no.
ZDR-5307; 1:10,000; both from OriGene Technologies, Inc.) secondary
antibodies for 1 h at room temperature. Pierce Enhanced
chemiluminescence Western Blotting Substrate (Thermo Fisher
Scientific, Inc.) was used to visualize the protein bands, and the
optical density of the bands was analyzed using ImageJ version
1.8.0 software (National Institutes of Health). The relative
expression of the proteins was measured using β-actin or GAPDH as
an internal control.
Dual-luciferase reporter gene assay
Two bioinformatics software programs, miRDB
(http://mirdb.org/) and microRNA. org (http://www.microrna.org/microrna/home.do), were used
to determine whether LGR4 was a target gene of miR-137. The
wild-type (wt) 3'-untranslated region (UTR) of LGR4 was amplified
and cloned into a pMiR-reporter vector (Promega Corporation). A
mutation site of LGR4 in a seed sequence was generated and inserted
into the pMiR-reporter vector (mut-LGR4-3'-UTR). The LGR4 3'-UTR
was sequenced, and the nucleotide alignment was compared with that
in the NCBI database (https://blast.ncbi.nlm.nih.gov/Blast.cgi;
NM_018490.5). PC3 cells (4×104 cells/well) were seeded
into a 24-well plate and co-transfected with miR-137 mimic or NC
and wt or mut-LGR4-3'-UTR. After 48-h transfection, a
Dual-Luciferase Reporter assay (Promega Corporation) was conducted
to analyze the firefly and Renilla luciferase activities
according to the manufacturer's instructions. Firefly luciferase
activity was normalized to Renilla luciferase activity.
Statistical analysis
All experiments were repeated at least three times.
Data are presented as the mean ± SD and were analyzed using SPSS
11.0 software (SPSS, Inc.). One-way analysis of variance followed
by Bonferroni post hoc test was used to compare the differences
among multiple groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
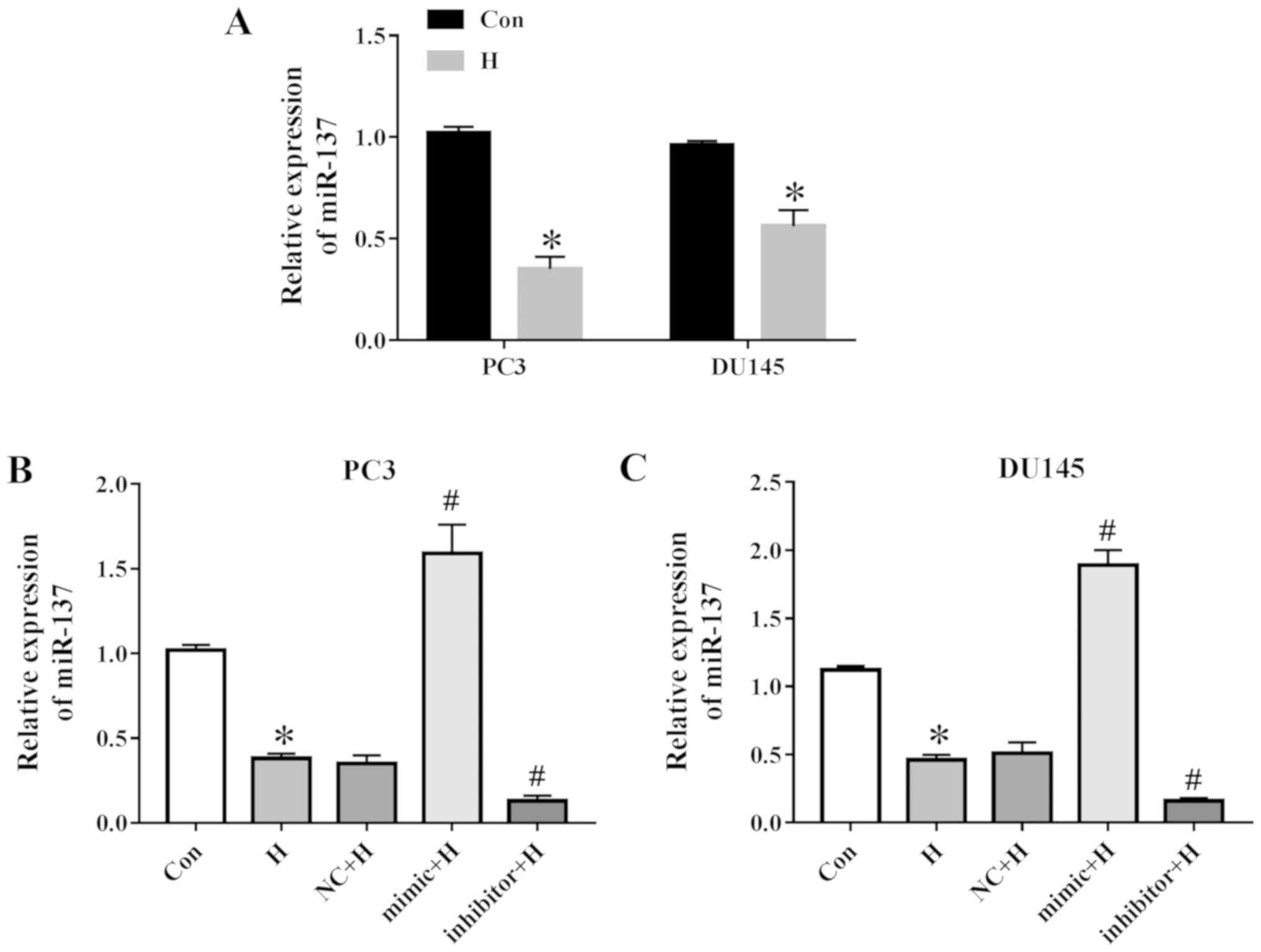
Hypoxia down-regulates miR-137 expression
in prostate cancer cells
The expression of miR-137 was determined in prostate
cancer cells under hypoxic conditions. PC3 and DU145 cells were
selected as they have been commonly used to study prostate cancer
progression and exhibit high levels of migratory and invasive
activity (20,21). As presented in Fig. 1A, PC3 and DU145 cells exposed to
hypoxia exhibited a decrease in miR-137. The results of RT-qPCR
also demonstrated high miR-137 levels following transfection with
the miR-137 mimic and low miR-137 levels following transfection
with the miR-137 inhibitor compared with those transfected with the
NC in PC3 and DU145 cells (Fig. 1B and
C).
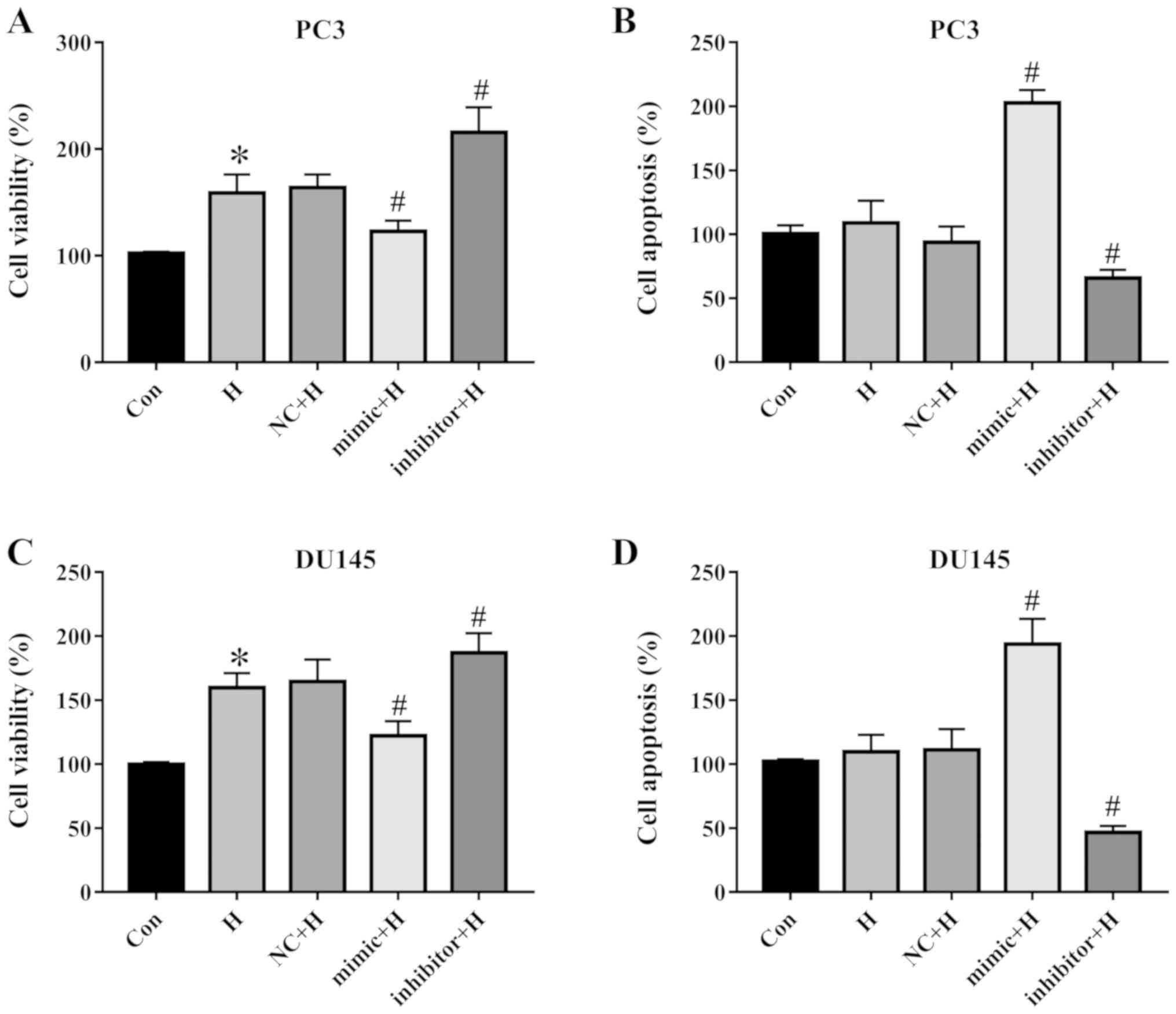
miR-137 affects prostate cancer cell
viability and apoptosis
Cell viability and apoptosis of PC3 and DU145 cells
were assayed. The results demonstrated that hypoxia induced cell
proliferation, transfection with the miR-137 mimic reduced cell
viability, and the miR-137 inhibitor promoted the survival of PC3
cells compared with the NC group (Fig.
2A). Hypoxia had no effect on apoptosis; however, miR-137
positively regulated apoptosis compared with the NC-transfected PC3
cells (Fig. 2B). Similar trends
were observed in DU145 cells, as hypoxia enhanced cell viability,
but did not affect apoptosis, whereas the miR-137 mimic reduced
cell viability and facilitated apoptosis, and inhibition of miR-137
promoted cell survival and decreased apoptosis (Fig. 2C and D). These results indicated
that miR-137 regulated the viability and apoptosis of PC3 and DU145
cells.
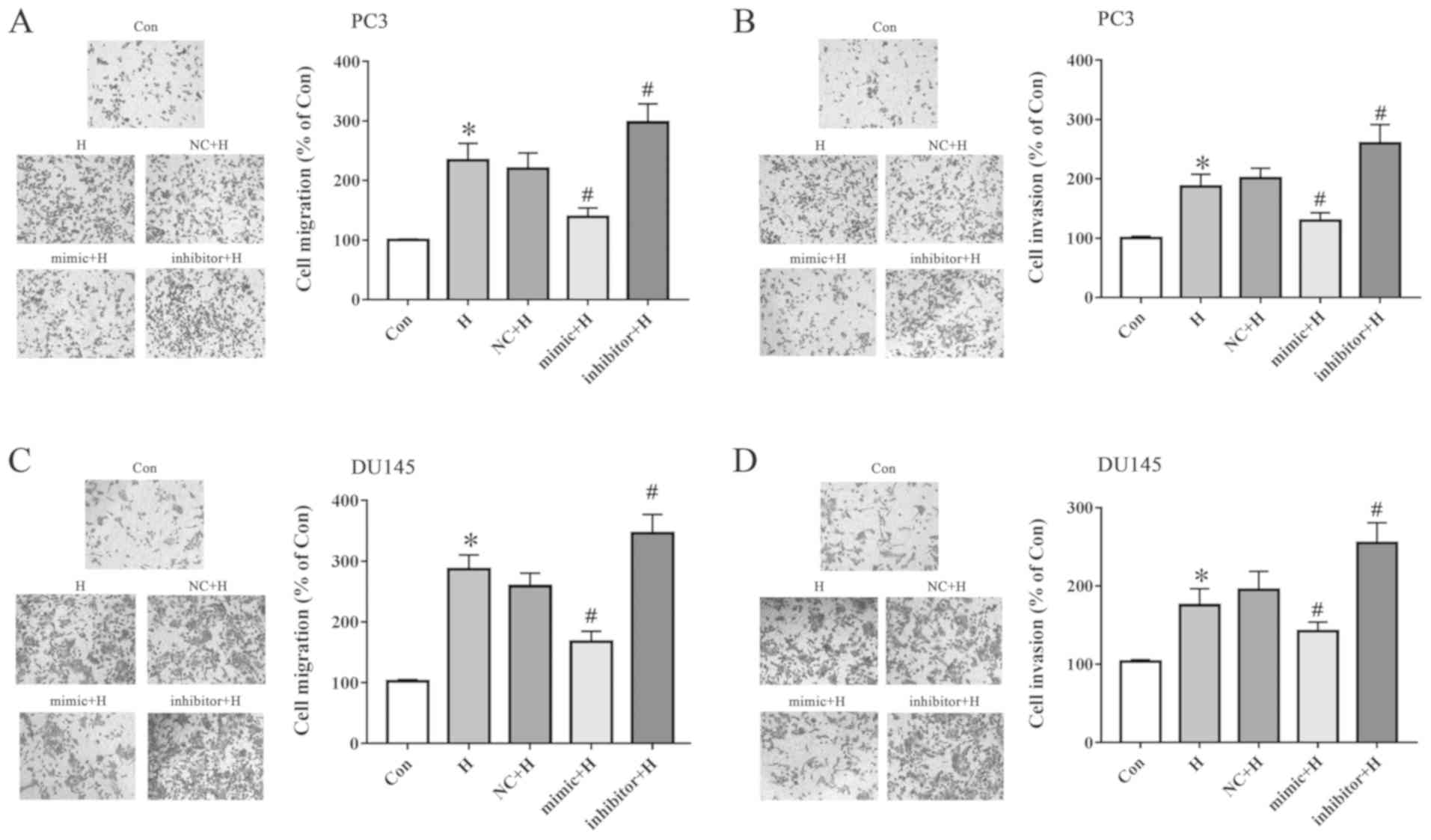
miR-137 inhibits cell migration and
invasion under hypoxia
PC3 and DU145 cells were exposed to hypoxia after
transfection with the miR-137 mimic or inhibitor. Cell migration
and invasion were assessed using Transwell assays. The miR-137
mimic inhibited hypoxia-induced cell migration (Fig. 3A) and invasion (Fig. 3B) compared with those in the NC + H
group. Inhibition of miR-137 displayed the opposite results in PC3
cells. In addition, the migration (Fig. 3C) and invasion (Fig. 3D) of DU145 cells were inhibited by
the miR-137 mimic and promoted by miR-137 inhibitor. These results
indicated that miR-137 blocked the hypoxia-induced migration and
invasion of prostate cancer cells.
miR-137 suppresses the hypoxia-induced
EMT
To investigate how miR-137 affects EMT in prostate
cancer cells, images of cell morphology were captured, and it was
observed that hypoxia induced a fusiform morphology; the miR-137
mimic reversed this effect, whereas the miR-137 inhibitor further
induced the EMT morphology of PC3 cells compared with the
NC-transfected cells (Fig. S1).
Similarly, in DU145 cells, the miR-137 mimic inhibited the
hypoxia-mediated EMT morphology, however, no differences were
observed in cell morphology between the NC + H and inhibitor + H
groups (Fig. S1). In order to
evaluate whether EMT occurred in PC3 and DU145 cells after
transfection with the miR-137 mimic or inhibitor and hypoxia, the
expression levels of EMT-related markers were detected. As
demonstrated in Fig. 4A, the
miR-137 mimic significantly attenuated the hypoxia-induced
increases in the mRNA levels of N-cadherin, vimentin and MMP2
compared with those in the NC + H group. Western blot analysis
revealed that compared with the NC + H group, the miR-137 mimic
down-regulated EMT-related protein expression levels, whereas the
miR-137 inhibitor further up-regulated EMT-related protein
expression in PC3 cells (Fig. 4B).
In DU145 cells, the miR-137 mimic blocked the hypoxia-induced
up-regulation of N-cadherin, vimentin and MMP2 mRNA levels compared
with the NC-transfected cells; however, ablation of miR-137
promoted hypoxia-induced increases in N-cadherin, vimentin, and
MMP2 mRNA levels (Fig. 4C). In
addition, the miR-137 mimic decreased the protein levels of
EMT-related proteins, whereas the miR-137 inhibitor increased the
protein levels of N-cadherin, vimentin, and MMP2 in DU145 cells
compared with those in the NC + H group (Fig. 4D). These results suggested that
miR-137 inhibited the EMT in prostate cancer cells exposed to
hypoxia.
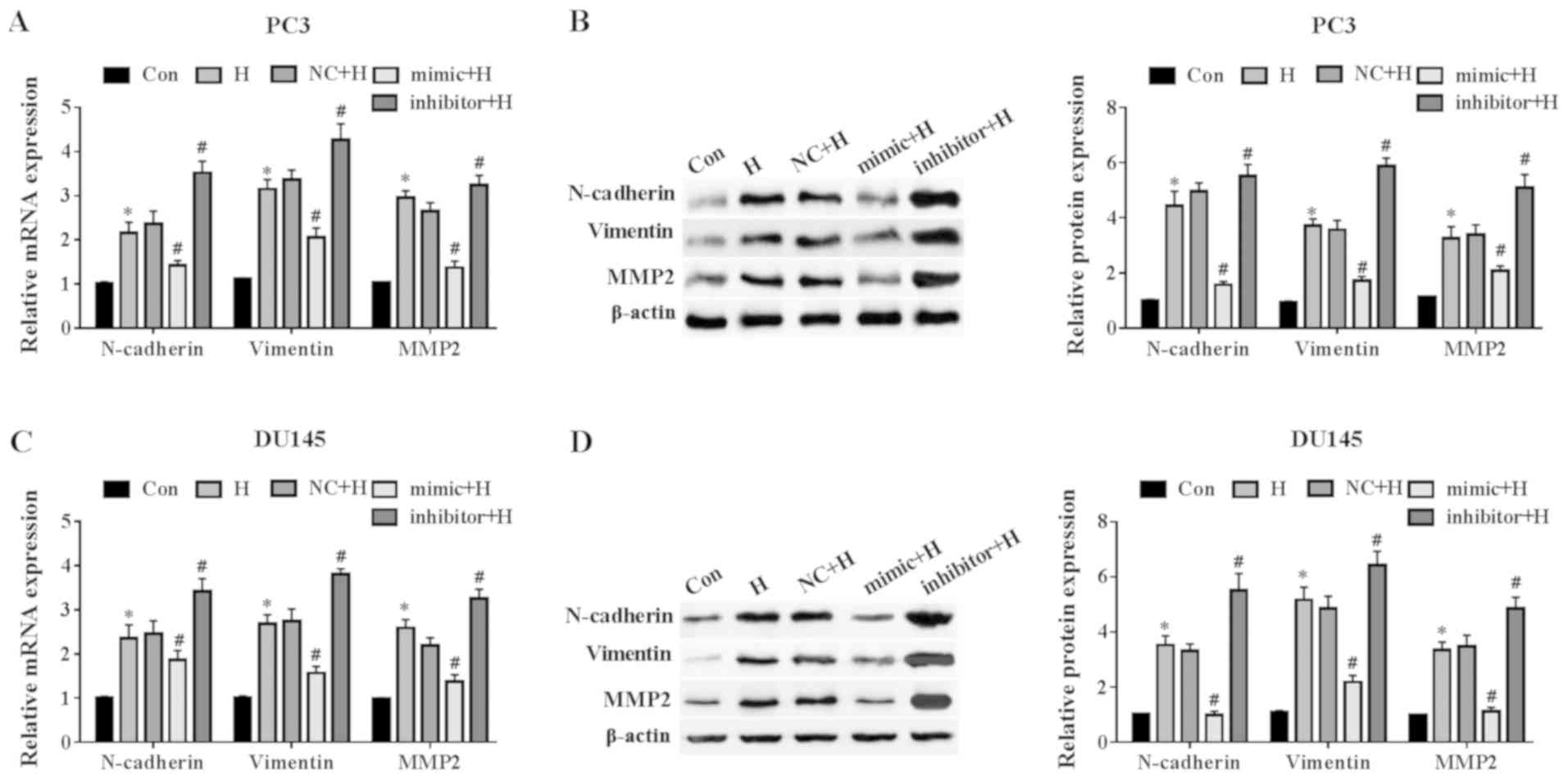 | Figure 4miR-137 inhibits hypoxia-induced
epithelial-mesenchymal transition in prostate cancer cells. PC3 and
DU145 cells were transfected with a miR-147 mimic or inhibitor for
48 h and exposed to hypoxia. (A and B) The N-cadherin, vimentin and
MMP2 (A) mRNA, as well as (B) protein levels in PC3 cells were
evaluated by RT-qPCR and western blotting, respectively. The (C)
mRNA and (D) protein levels of N-cadherin, vimentin and MMP2 in
DU145 cells were also tested by RT-qPCR and western blotting.
*P<0.05 vs. Con; #P<0.05 vs. NC + H.
miR, microRNA; Con, control; H, hypoxia; NC, miR-137 negative
control; RT-qPCR, reverse transcription-quantitative PCR; MMP2,
matrix metalloproteinase 2. |
miR-137 targets LGR4 in PC3 cells
Bioinformatics analysis identified LGR4 as a target
gene of miR-137. The binding site between miR-137 and LGR4-wt or
LGR4-mut is presented in Fig. 5A.
The LGR4-3'-UTR was sequenced, and then nucleotide alignment was
compared in NCBI. The similarity with NM_018490.5 was 99.09% (data
not shown). To confirm this result, a dual-luciferase reporter gene
assay was performed in PC3 cells after 48-h co-transfection with
the miR-137 mimic or NC and LGR4-wt or LGR4-mut. As presented in
Fig. 5B, luciferase activity
decreased following co-transfection with LGR4-wt and the miR-137
mimic compared with that in the NC group, whereas no significant
differences were observed in cells transfected with LGR4-mut.
Additionally, compared with the NC + H group, the miR-137 mimic
negatively regulated the expression of LGR4 at the mRNA (Fig. 5C) and protein (Fig. 5D) levels under hypoxic
conditions.
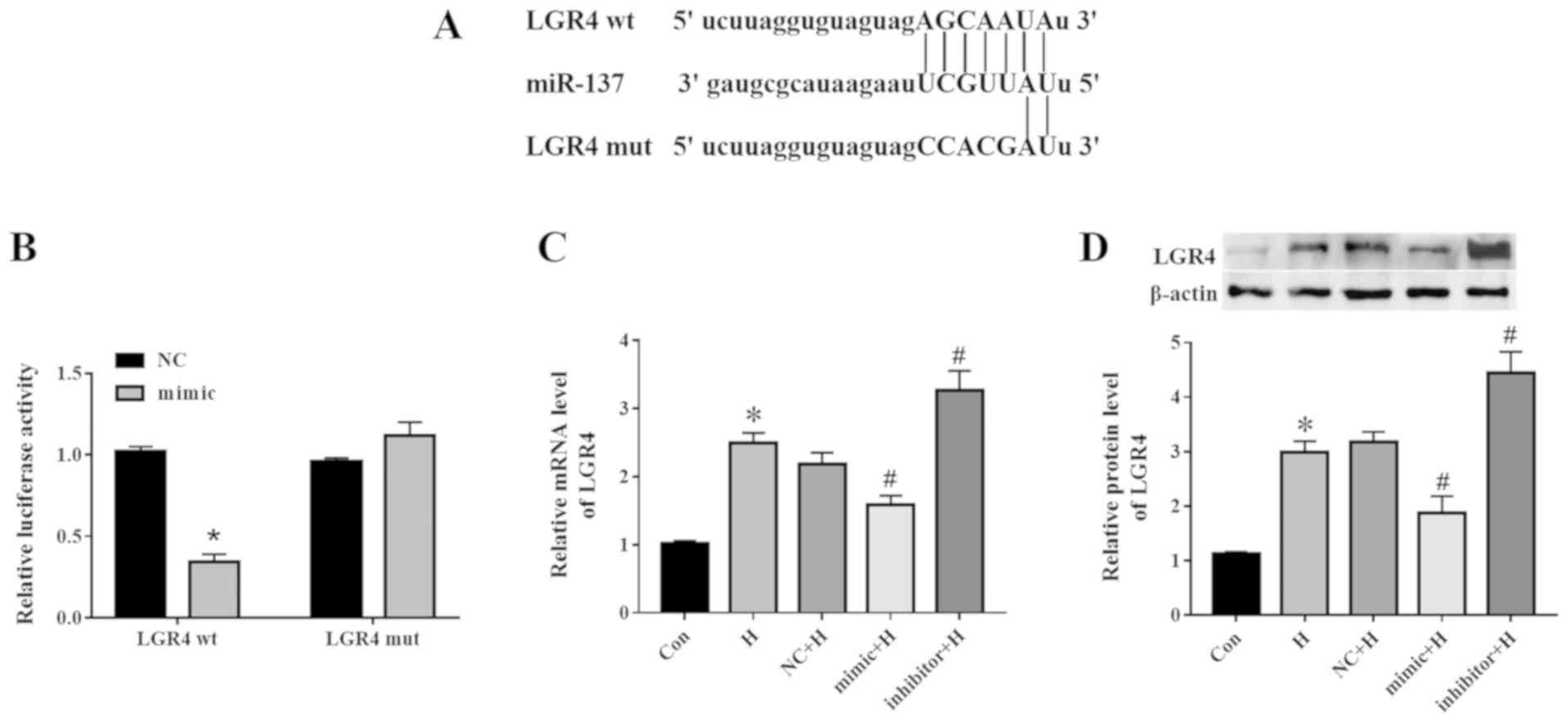 | Figure 5miR-137 directly targets LGR4 in PC3
cells. (A) The binding sites between miR-137 and LGR4-wt or
LGR4-mut. (B) PC3 cells were co-transfected with NC or miR-137
mimic and LGR4-wt or with LGR4-mut for 48 h, and relative
luciferase activity was determined. The (C) mRNA and (D) protein
levels of LGR4 were detected by RT-qPCR and western blotting,
respectively. *P<0.05 vs. Con or NC;
#P<0.05 vs. NC + H. miR, microRNA; Con, control; H,
hypoxia; NC, miR-137 negative control; LGR4, leucine-rich
repeat-containing G protein-coupled receptor 4; wt, wild-type; mut,
mutant; RT-qPCR, reverse transcription-quantitative PCR. |
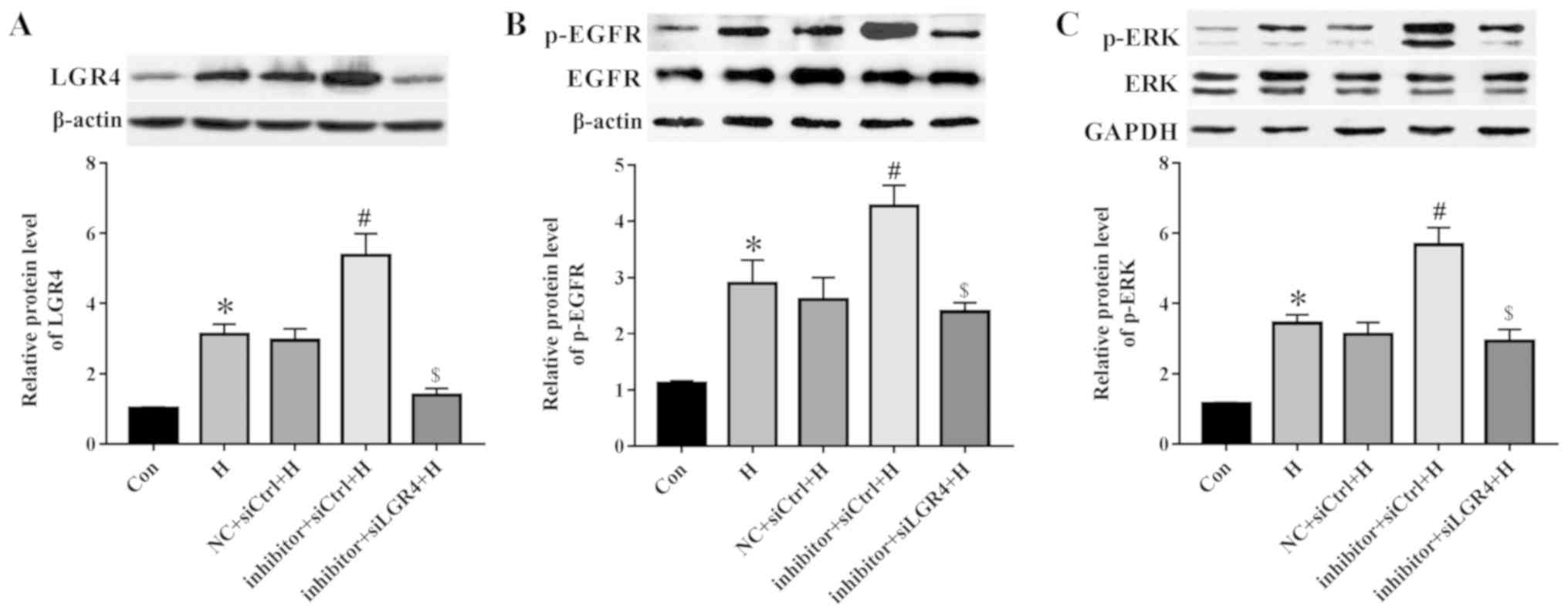
miR-137 regulates EGFR/ERK signaling by
targeting LGR4
To elucidate the mechanism of miR-137 in the
modulation of migration and EMT, PC3 cells were co-transfected with
the miR-137 inhibitor and siLGR4 and subjected to hypoxia. The
levels of LGR4 expression were decreased at the mRNA and protein
levels following transfection with siLGR4 compare with that in the
siCtrl group (Fig. S2). In
addition, the miR-137 inhibitor elevated the LGR4 protein levels
compared with those in the negative control group, whereas
knockdown of LGR4 reduced the expression of LGR4 compared with that
in the inhibitor + siCtrl group (Fig.
6A). Inhibition of miR-137 expression further up-regulated the
hypoxia-mediated augmentation of EGFR phosphorylation compared with
that in the NC + siCtrl + H group, whereas LGR4 silencing reversed
this effect (Fig. 6B). Consistent
with EGFR, the miR-137 inhibitor increased the phosphorylation of
ERK compared with that in the negative control group, whereas LGR4
knockdown decreased it (Fig. 6C).
These results suggested that miR-137 may regulate the EGFR/ERK
signaling pathway by targeting LGR4.
miR-137 regulates PC3 cell migration and
EMT by targeting LGR4 via the EGFR/ERK signaling pathway
To examine the potential mechanisms of miR-137 in
the regulation of the migration and EMT of PC3 cells, the cells
were pretreated with AG1478, which is a selective inhibitor of
EGFR. As presented in Fig. 7A and
B, the miR-137 inhibitor increased cell migration and invasion
under hypoxic conditions compared with the NC + H group, whereas
down-regulation of LGR4 or disruption of the EGFR/ERK signaling
pathway reversed these effects. Furthermore, the mRNA levels of
N-cadherin, vimentin and MMP2 increased after transfection with the
miR-137 inhibitor compared with those in the NC-transfected cells,
but decreased following LGR4 knockdown or inhibition of the
EGFR/ERK signaling pathway (Fig.
7C). Similarly, the effects of the miR-137 inhibitor on the
regulation of N-cadherin, vimentin and MMP2 protein levels were
abolished by LGR4 silencing or EGFR/ERK signaling pathway
inhibition in presence of hypoxia (Fig. 7D). These results suggested that
miR-137 regulated the migration and EMT of PC3 cells by targeting
LGR4 via the EGFR/ERK signaling pathway.
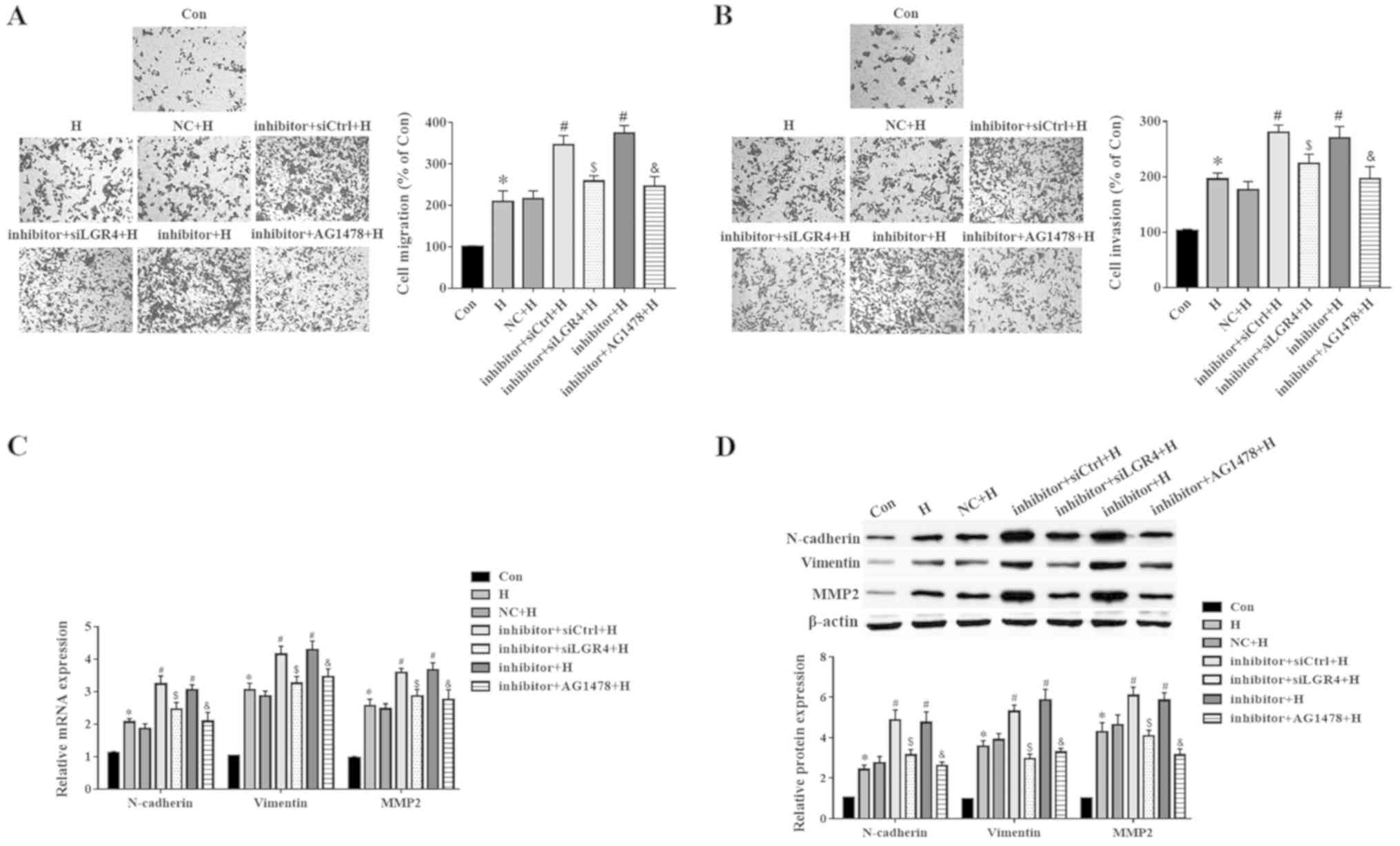 | Figure 7LGR4 silencing or EGFR/ERK blockade
reverses the biological roles of miR-137 in PC3 cells. Cells were
transfected with miR-137 inhibitor, and then treated with 100 nM of
AG1478 for 1 h, and exposed to hypoxia. (A and B) A Transwell assay
was used to evaluate the (A) migration and (B) invasion of PC3
cells. (C) The mRNA levels of N-cadherin, vimentin and MMP2 were
analyzed by RT-qPCR. (D) The protein levels of N-cadherin, vimentin
and MMP2 were detected using western blotting.
*P<0.05 vs. Con or NC; #P<0.05 vs. NC +
H; $P<0.05 vs. inhibitor + siCtrl + H;
&P<0.05 vs. inhibitor + H. miR, microRNA; Con,
control; H, hypoxia; NC, miR-137 negative control; si, small
interfering RNA; LGR4, leucine-rich repeat-containing G
protein-coupled receptor 4; siCtrl, negative control for siLGR4;
EGFR, epidermal growth factor receptor; MMP2, matrix
metalloproteinase 2; AG1478, a selective inhibitor of EGFR;
RT-qPCR, reverse transcription-quantitative PCR. |
Discussion
Hypoxia is a common feature of solid tumors and a
negative prognostic and predictive factor that contributes to tumor
progression and chemoresistance (22). MicroRNAs are abnormally expressed
when exposed to hypoxia, lipopolysaccharides or tumor necrosis
factor-α (23,24). miR-137 is downregulated in the
mouse brain during hypoxia (18).
Consistent with these findings, the results of the present study
demonstrated decreased levels of miR-137 in PC3 and DU145 cells
after exposure to hypoxia, suggesting that miR-137 was involved in
the hypoxia-mediated prostate cancer progression.
The responses of tumor cells to hypoxia include cell
proliferation, differentiation and resistance to apoptosis
(25). The sensitivity to
apoptosis is likely to be one of the critical factors in
determining whether cancer cells survive (26). In the present study, hypoxia
promoted cell survival, but had no effect on apoptosis in PC3 and
DU145 cells. Coffey et al (27) previously demonstrated that PC3 and
DU145 cells were resistant to apoptosis under hypoxic conditions,
but LNCaP and PWR-1E cells were susceptible, which may be due to
androgen-independent PC3 and DU145 expressing high levels of NLR
family apoptosis inhibitory protein, cellular inhibitor of
apoptosis 1 (cIAP-1) and cIAP-2, among which cIAP-2 expression has
been identified to be associated with apoptosis resistance. The
results of the present study demonstrated that miR-137 prevented
the proliferation and triggered the apop-tosis of PC3 and DU145
cells. Consistent with these results, Huang et al (28) demonstrated that miR-137
overexpression reduced cell proliferation and increased apoptosis
in acute lymphoblastic leukemia cells, which has also been observed
in pancreatic cancer cells (29).
miR-137 is a suppressor of a number of tumors,
including gastric cancer (30),
hepatocellular (31) and renal
cell (32) carcinoma. miR-137
attenuates the proliferation, migration, invasion and EMT of
multiple types of cancer cells, including endometrial (33), colon (34) and triple-negative breast (35) cancer cells. Consistent with these
findings, the results of the present study demonstrated that
miR-137 reduced the hypoxia-induced cell viability, migration and
invasion, as well as EMT-related protein expression of prostate
cancer cells.
LGRs belong to the seven-transmembrane protein
family with receptors for glycoprotein, thyrotropin, and
luteinizing and follicle-stimulating hormones (36). LGR4, also termed G protein-coupled
receptor 48, is closely associated with LGR5 and LGR6 (37). LGR4 serves a central role in the
development of the male reproductive tract (38). LGR5 mostly occurs on the surface of
gastrointestinal proliferative stem cells, where it serves as a
specific molecular marker of stem cells (39). LGR6 appears on the surface of skin
pluripotent stem cells (40).
LGR4, LGR5 and LGR6 were initially considered to be orphan
receptors until it was discovered that R-spondin ligands directly
linked these receptors to signal transduction in stem cells
(41). Recently, LGR4 has been
implicated in tumor progression, including papillary thyroid
carcinoma (42) and prostate
cancer (43).
The short-term survival rate in patients with high
expression of LGR4 is lower compared with that in patients with low
LGR4 expression, and LGR4 silencing inhibits the migration,
invasion and scarring of the prostate cancer cell line DU145
(43). In addition, overexpression
of LGR4 exacerbates tumorigenesis of prostate cancer cells
(44,45). PC3 and DU145 cells are commonly
used to study prostate cancer progression due to high levels of
migratory and invasive activity (20,21).
However, in all experiments in the present study, PC3 cells
exhibited higher reproducibility of the results and more
significant differences among groups compared with DU145 cells.
Thus, PC3 cells were selected to study the association between
miR-137 and LGR4, and the roles of LGR4 knockdown in hypoxia and
miR-137 inhibitor-mediated cell migration, invasion and EMT-related
protein expression. The results demonstrated that miR-137 directly
targeted the 3'-UTR of LGR4 in PC3 cells. Of note, miR-137
negatively regulated the expression of LGR4, and LGR4 knockdown
reversed the biological effects of the miR-137 inhibitor on PC3
cell migration and EMT, suggesting that miR-137 protected against
hypoxia-mediated migration and EMT by inhibiting LGR4.
EGFR belongs to the receptor tyrosine kinase family
and initiates tyrosine kinase activity after activation of EGF and
other ligands, and further activates the downstream signal
transduction pathways (46).
Activation of EGFR/ERK signaling contributes to EMT and migration
in prostate cancer, and LGR4 deficiency reduces phosphorylation of
EGFR and ERK. EGFR/ERK signaling is associated with tumorigenesis
and is activated in lung cancer (47). EGFR/ERK signaling also inhibits
cell proliferation and facilitates apoptosis in renal cell
carcinoma (48). Blocking this
pathway promotes cell death in prostate cancer (49). A previous report has demonstrated
that LGR4 deficiency downregulates the phosphorylation of EGFR and
ERK (50). The results of the
present study demonstrated that miR-137 negatively regulated the
EGFR/ERK signaling pathway by targeting LGR4 and that AG1478
abrogated the biological activities of miR-137. These results
suggested that miR-137 regulated the EGFR/ERK signaling pathway by
targeting LGR4 to impede the migration and EMT of prostate cancer
cells.
In conclusion, the results of the present study
revealed that hypoxia led to decreases in miR-137 expression. The
miR-137 mimic inhibited, whereas the miR-137 inhibitor aggravated
the hypoxia-induced cell migration, invasion and EMT in prostate
cancer cells. Additionally, a luciferase reporter assay verified
that miR-137 directly targeted LGR4. miR-137 negatively regulated
EGFR/ERK by targeting LGR4. Knockdown of LGR4 abolished the effects
of the miR-137 inhibitor on cell migration and EMT. AG1478
treatment also abrogated the effects of the miR-137 inhibitor on
PC3 cell migration, invasion and EMT. Taken together, these results
demonstrated that miR-137 may regulate the hypoxia-mediated
migration and EMT in prostate cancer by targeting LGR4 via the
EGFR/ERK signaling pathway. These results suggest a possible
therapeutic strategy for prostate cancer treatment.
Supplementary Data
Funding
This study was supported by The Medical Science and
Technology Research Projects of Henan Province (grant no.
201602335).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HZ and FL conceived and designed the study. HZ, FL
and JY designed the experiments. HZ, FL, JY, PL, JW, ZW, HL, DC and
JD performed experiments and analyzed the data. HZ wrote the draft
of manuscript. KZ and PD analyzed the data and revised the paper.
All authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
Not applicable.
References
|
1
|
Bashir MN: Epidemiology of prostate
cancer. Asian Pac J Cancer Prev. 16:5137–5141. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kazuto I: Prostate cancer in Asian men.
Nat Rev Urol. 11:197–212. 2014. View Article : Google Scholar
|
|
3
|
Zhu Y, Freedland SJ and Ye D: Prostate
cancer and prostatic diseases best of Asia, 2019: Challenges and
opportunities. Prostate Cancer Prostatic Dis. Dec 6–2019, Epub
ahead of print. PubMed/NCBI
|
|
4
|
Marusic G, Vojinov S and Levakov I:
Treatment of locally advanced prostatic cancer. Med Pregl.
63:689–695. 2010.In Serbian. View Article : Google Scholar
|
|
5
|
Noman MZ, Hasmim M, Messai Y, Terry S,
Kieda C, Janji B and Chouaib S: Hypoxia: A key player in antitumor
immune response. A review in the theme: Cellular responses to
hypoxia. Am J Physiol Cell Physiol. 309:C569–C579. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ramteke A, Ting H, Agarwal C, Mateen S,
Somasagara R, Hussain A, Graner M, Frederick B, Agarwal R and Deep
G: Exosomes secreted under hypoxia enhance invasiveness and
stemness of prostate cancer cells by targeting adherens junction
molecules. Mol Carcinog. 54:554–565. 2015. View Article : Google Scholar
|
|
7
|
Deep G and Panigrahi GK: Hypoxia-induced
signaling promotes prostate cancer progression: Exosomes role as
messenger of hypoxic response in tumor microenvironment. Crit Rev
Oncog. 20:419–434. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Roldo C, Missiaglia E, Hagan JP, Falconi
M, Capelli P, Bersani S, Calin GA, Volinia S, Liu CG, Scarpa A and
Croce CM: MicroRNA expression abnormalities in pancreatic endocrine
and acinar tumors are associated with distinctive pathologic
features and clinical behavior. J Clin Oncol. 24:4677–4684. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Farazi TA, Hoell JI, Morozov P and Tuschl
T: MicroRNAs in human cancer. Adv Exp Med Biol. 774:1–20. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hong X and Yu JJ: MicroRNA-150 suppresses
epithelial mesenchymal transition, invasion and metastasis in
prostate cancer through the TRPM4-mediated β-catenin signaling
pathway. Am J Physiol Cell Physiol. 316:C463–C480. 2019. View Article : Google Scholar
|
|
11
|
Blenkiron C, Goldstein LD, Thorne NP,
Spiteri I, Chin SF, Dunning MJ, Barbosa-Morais NL, Teschendorff AE,
Green AR, Ellis IO, et al: MicroRNA expression profiling of human
breast cancer identifies new markers of tumor subtype. Genome Biol.
8:R2142007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kota J, Chivukula RR, O'Donnell KA,
Wentzel EA, Montgomery CL, Hwang HW, Chang TC, Vivekanandan P,
Torbenson M, Clark KR, et al: Therapeutic microRNA delivery
suppresses tumorigenesis in a murine liver cancer model. Cell.
137:1005–1017. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao S, Gao X, Zang S, Li Y, Feng X and
Yuan X: MicroRNA-383-5p acts as a prognostic marker and inhibitor
of cell proliferation in lung adenocarcinoma by cancerous inhibitor
of protein phosphatase 2A. Oncol Lett. 14:3573–3579. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhiping C, Shijun T, Linhui W, Yapei W,
Lianxi Q and Qiang D: miR-181a promotes epithelial to mesenchymal
transition of prostate cancer cells by targeting TGIF2. Eur Rev Med
Pharmacol Sci. 21:4835–4843. 2017.PubMed/NCBI
|
|
15
|
Yang ZG, Ma XD, He ZH and Guo YX:
miR-483-5p promotes prostate cancer cell proliferation and invasion
by targeting RBM5. Int Braz J Urol. 43:1060–1067. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xu S, Ge J, Zhang Z and Zhou W: miR-129
inhibits cell proliferation and metastasis by targeting ETS1 via
PI3K/AKT/mTOR pathway in prostate cancer. Biomed Pharmacother.
96:634–641. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pashaei E, Ahmady M, Ozen M and Aydin N:
Meta-analysis of miRNA expression profiles for prostate cancer
recurrence following radical prostatectomy. PLoS One.
12:e01795432017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li W, Zhang X, Zhuang H, Chen HG, Chen Y,
Tian W, Wu W, Li Y, Wang S, Zhang L, et al: MicroRNA-137 is a novel
hypoxia-responsive microRNA that inhibits mitophagy via regulation
of two mitophagy receptors FUNDC1 and NIX. J Biol Chem.
289:10691–10701. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
20
|
Pulukuri SM, Gondi CS, Lakka SS, Jutla A,
Estes N, Gujrati M and Rao JS: RNA interference-directed knockdown
of urokinase plasminogen activator and urokinase plasminogen
activator receptor inhibits prostate cancer cell invasion,
survival, and tumorigenicity in vivo. J Biol Chem. 280:36529–36540.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Viticchie G, Lena AM, Latina A, Formosa A,
Gregersen LH, Lund AH, Bernardini S, Mauriello A, Miano R, Spagnoli
LG, et al: miR-203 controls proliferation, migration and invasive
potential of prostate cancer cell lines. Cell Cycle. 10:1121–1131.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Vaupel P and Mayer A: Hypoxia in cancer:
Significance and impact on clinical outcome. Cancer Metastasis Rev.
26:225–239. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shin S, Moon KC, Park KU and Ha E:
MicroRNA-513a-5p mediates TNF-α and LPS induced apoptosis via
downregulation of X-linked inhibitor of apoptotic protein in
endothelial cells. Biochimie. 94:1431–1436. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chan SY, Zhang YY, Hemann C, Mahoney CE,
Zweier JL and Loscalzo J: MicroRNA-210 controls mitochondrial
metabolism during hypoxia by repressing the iron-sulfur cluster
assembly proteins ISCU1/2. Cell Metab. 10:273–284. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tang J, Chen Y, Cui R, Li D, Xiao L, Lin
P, Du Y, Sun H, Yu X and Zheng X: Upregulation of fractalkine
contributes to the proliferative response of prostate cancer cells
to hypoxia via promoting the G1/S phase transition. Mol Med Rep.
12:7907–7914. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shannon AM, Bouchier-Hayes DJ, Condron CM
and Toomey D: Tumour hypoxia, chemotherapeutic resistance and
hypoxia-related therapies. Cancer Treat Rev. 29:297–307. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Coffey RN, Morrissey C, Taylor CT,
Fitzpatrick JM and Watson RW: Resistance to caspase-dependent,
hypoxia-induced apoptosis is not hypoxia-inducible factor-1 alpha
mediated in prostate carcinoma cells. Cancer. 103:1363–1374. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Huang Y, Zou Y, Zheng R and Ma X: miR-137
inhibits cell proliferation in acute lymphoblastic leukemia by
targeting JARID1B. Eur J Haematol. 103:215–224. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ding F, Zhang S, Gao S, Shang J, Li Y, Cui
N and Zhao Q: miR-137 functions as a tumor suppressor in pancreatic
cancer by targeting MRGBP. J Cell Biochem. 119:4799–4807. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Du Y, Chen Y, Wang F and Gu L: miR-137
plays tumor suppressor roles in gastric cancer cell lines by
targeting KLF12 and MYO1C. Tumour Biol. 37:13557–13569. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Huang B, Huang M and Li Q: miR-137
suppresses migration and invasion by targeting EZH2-STAT3 signaling
in human hepatocellular carcinoma. Pathol Res Pract. 214:1980–1986.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang M, Gao H, Qu H, Li J, Liu K and Han
Z: miR-137 suppresses tumor growth and metastasis in clear cell
renal cell carcinoma. Pharmacol Rep. 70:963–971. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang W, Chen JH, Shan T,
Aguilera-Barrantes I, Wang LS, Huang TH, Rader JS, Sheng X and
Huang YW: miR-137 is a tumor suppressor in endometrial cancer and
is repressed by DNA hypermethylation. Lab Invest. 98:1397–1407.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bi WP, Xia M and Wang XJ: miR-137
suppresses proliferation, migration and invasion of colon cancer
cell lines by targeting TCF4. Oncol Lett. 15:8744–8748.
2018.PubMed/NCBI
|
|
35
|
Lee SJ, Jeong JH, Kang SH, Kang J, Kim EA,
Lee J, Jung JH, Park HY and Chae YS: MicroRNA-137 inhibits cancer
progression by targeting Del-1 in Triple-negative breast cancer
cells. Int J Mol Sci. 20:pii: E6162. 2019. View Article : Google Scholar
|
|
36
|
Schoore GV, Mendive F, Pochet R and
Vassart G: Expression pattern of the orphan receptor LGR4/GPR48
gene in the mouse. Histochem Cell Biol. 124:35–50. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Carmon KS, Gong X, Lin Q, Thomas A and Liu
Q: R-spondins function as ligands of the orphan receptors LGR4 and
LGR5 to regulate Wnt/beta-catenin signaling. Proc Natl Acad Sci
USA. 108:11452–11457. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hoshii T, Takeo T, Nakagata N, Takeya M,
Araki K and Yamamura K: LGR4 regulates the postnatal development
and integrity of male reproductive tracts in mice. Biol Reprod.
76:303–313. 2007. View Article : Google Scholar
|
|
39
|
Koo BK and Clevers H: Stem cells marked by
the R-spondin receptor LGR5. Gastroenterology. 147:289–302. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Leushacke M and Barker N: Lgr5 and Lgr6 as
markers to study adult stem cell roles in self-renewal and cancer.
Oncogene. 31:3009–3022. 2012. View Article : Google Scholar
|
|
41
|
Dongli W, Binlu H, Senyan Z, Xiaojuan Y,
Wei W and Xinquan W: Structural basis for R-spondin recognition by
LGR4/5/6 receptors. Genes Dev. 27:1339–1344. 2013. View Article : Google Scholar
|
|
42
|
Kang YE, Kim JM, Kim KS, Chang JY, Jung M,
Lee J, Yi S, Kim HW, Kim JT, Lee K, et al: Upregulation of
RSPO2-GPR48/LGR4 signaling in papillary thyroid carcinoma
contributes to tumor progression. Oncotarget. 8:114980–114994.
2017. View Article : Google Scholar
|
|
43
|
Luo W, Tan P, Rodriguez M, He L, Tan K,
Zeng L, Siwko S and Liu M: Leucine-rich repeat-containing G protein
coupled receptor 4 (Lgr4) is necessary for prostate cancer
metastasis via epithelial-mesenchymal transition. J Biol Chem.
292:15525–15537. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhang J, Qi L, Zhang S, Xu Q and Wang T:
Lgr4 promotes prostate tumorigenesis through the Jmjd2a/AR
signaling pathway. Exp Cell Res. 349:77–84. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Fang L, Junmin Y, Junyong W, Lijuan Z, Rui
F, Hao Z and Qingsong Z: GPCR48/LGR4 promotes tumorigenesis of
prostate cancer via PI3K/Akt signaling pathway. Med Oncol.
32:492015. View Article : Google Scholar
|
|
46
|
Orton RJ, Adriaens ME, Gormand A, Sturm
OE, Kolch W and Gilbert DR: Computational modelling of cancerous
mutations in the EGFR/ERK signalling pathway. BMC Syst Biol.
3:1002009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yang Y, Zhao W, Xu QW, Wang XS, Zhang Y
and Zhang J: IQGAP3 promotes EGFR-ERK signaling and the growth and
metastasis of lung cancer cells. PLoS One. 9:e975782014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Li L, Gao Y, Zhang L, Zeng J, He D and Sun
Y: Silibinin inhibits cell growth and induces apoptosis by caspase
activation, down-regulating survivin and blocking EGFR-ERK
activation in renal cell carcinoma. Cancer Lett. 272:61–69. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Oh HY, Lee EJ, Yoon S, Chung BH, Cho KS
and Hong SJ: Cholesterol level of lipid raft microdomains regulates
apoptotic cell death in prostate cancer cells through EGFR-mediated
Akt and ERK signal transduction. Prostate. 67:1061–1069. 2010.
View Article : Google Scholar
|
|
50
|
Pan H, Cui H, Liu S, Qian Y, Wu H, Li L,
Guan Y, Guan X, Zhang L, Fan HY, et al: Lgr4 gene regulates corpus
luteum maturation through modulation of the WNT-mediated EGFR-ERK
signaling pathway. Endocrinology. 155:3624–3637. 2014. View Article : Google Scholar : PubMed/NCBI
|