Introduction
Stroke, which is characterized by loss of
neurological function caused by ischemia of the brain,
intracerebral hemorrhage or subarachnoid hemorrhage (1), is associated with high morbidity and
mortality rates (2,3). It has been demonstrated that, by
inducing excitotoxicity, cerebral ischemia/reperfusion (I/R) injury
is a critical factor responsible for poor prognosis in patients
with ischemic stroke. Stroke disrupts calcium ion homeostasis,
causes overproduction of free radicals and inflammatory cytokines,
and promotes cell apoptosis (4).
Currently, although thrombolytic, endovascular and adjuvant novel
therapies have been developed for stroke (5,6),
they have been proven insufficient in achieving the desired
outcome. Therefore, a better understanding of the mechanisms
underlying the development of cerebral I/R injury is required.
Mild hypothermia (MH) exerts neuroprotective effects
against cerebral ischemia. It was previously reported that MH
reduces brain hemorrhage and blood-brain barrier disruption after
stroke (7), and that it may
alleviate cerebral ischemic injury in diabetic patients through
promoting autophagy and inhibiting pyroptosis (8). Researchers also demonstrated that
inhibition of Notch3 and Notch4 signaling is involved in the
protective effect of MH against cerebral ischemic injury (9). Moreover, MH promotes long-term white
matter integrity and inhibits neuroinflammation in mice with
ischemic brain injury (10). These
previous findings indicate that MH may be of therapeutic value in
cerebral I/R injury.
Edaravone (EDA; 3-methyl-1-phenyl-2-pyrazolin-5-one)
is a free radical scavenger. Evidence has indicated that EDA
protects the brain against cerebral ischemic injury, and it may
inhibit microglia-mediated neuroinflammation in rats with cerebral
ischemic injury (11). It has also
been demonstrated that EDA protects neuronal cells from ischemic
injury by inhibiting the translocation of 5-lipoxygenase to the
nuclear membrane, thereby blocking the 5-lipoxygenase signaling
pathway (12). Moreover, EDA
combined with MH significantly improves neuroprotection in rats
exposed to hypoxia (13). Thus, it
was inferred that EDA in combination with MH may exert a
synergistic effect against cerebral I/R injury.
The transcription factor nuclear factor erythroid
2-related factor 2 (Nrf2) is a central modulator in multiple
biochemical processes, such as redox, protein and metabolic
homeostasis. Nrf2-based therapeutics have been developed for
treating various cardiovascular, kidney and liver diseases
(14). It was previously
demonstrated that EDA protects the nervous system from toxicity
through activating the Nrf2 signaling pathway (15). However, whether the positive
effects of EDA on cerebral I/R injury are mediated through the
activation of the Nrf2 signaling pathway remains unclear.
Therefore, in the present study, a cerebral I/R
model in rats was constructed to explore the potential synergistic
effects and the mechanism underlying the combination of EDA with MH
in I/R injury. In addition, brusatol (Bru), an inhibitor of the
Nrf2 signaling pathway, was also used to investigate the effects of
Nrf2 signaling on I/R injury.
Materials and methods
MCAO model and drug treatment
A total of 60 healthy adult Sprague-Dawley male
rats, 9-10 weeks old and weighing 300-320 g, were purchased from
Vital River Laboratories Co., Ltd. All the animals were housed
under specific pathogen-free conditions with a 12-h dark/light
cycle at 25°C, and fed standard food and aseptic water. All the
experiments were approved by the Institutional Animal Ethics
Committee of Hainan Medical University (approval no.
C2017051922A).
Focal I/R in each rat was created by middle cerebral
artery occlusion (MCAO). Briefly, the rats were anesthetized by 3%
isoflurane at 50 mg/kg body weight (1235809; Sigma-Aldrich; Merck
KGaA) administered by intraperitoneal injection, while monitoring
the heart rate. Then, the end of the common cerebral artery (CCA)
nearer to the heart was closed by a nylon suture. A special nylon
suture with a spherical end (diameter 0.18 mm) was inserted into
the brain of each rat following the direction of the CCA and
stopped when the thread entered the middle cerebral artery (MCA).
After occlusion for 120 min, the thread was dismantled to allow
reperfusion for 24 h. The rats treated by MCAO were then randomly
divided into 5 groups as follows: MCAO, MCAO + MH, MCAO + EDA, MCAO
+ MH + EDA and MCAO + MH + EDA + Bru groups (n=10 rats per group).
Furthermore, 10 rats in the sham group were treated in a similar
manner but without arterial occlusion. Immediately after MCAO, ice
was placed under the skull of the rats treated by MH, with the
temperature of the temporalis muscle maintained at 32-34°C for 2 h.
EDA (M70800; Sigma-Aldrich; Merck KGaA) and Bru (SML1868,
Sigma-Aldrich; Merck KGaA) were diluted in physiological saline
(16,17), and 3.0 mg/kg EDA or 1.0 mg/kg Bru
were immediately intraperitoneally injected into the rats after
MCAO. No rats died during the experiments. The rats were euthanized
by 3% isoflurane (150 mg/kg body weight) through intraperitoneal
injection 24 h later and, after cardiac arrest was confirmed, their
brain tissues were collected for the different assays.
Neurological deficit score
The modified Longa's Scoring System (18) was used to evaluate the neurological
deficits of the rats at 24 h after reperfusion as follows: 0, no
deficit; 1, inability to stretch the contralateral forelimb fully;
2, circling to the contralateral side; 3, falling over to the
contralateral side; and 4, no spontaneous locomotor activity. A
higher score indicated a more severe neurological deficit.
Brain 2,3,5-triphenyltetrazolium chloride
(TTC) staining and infarct size calculation
The rats were euthanized and their brain tissues
were collected, cut into 3-mm sections, and placed in a water bath
with 2% TTC solution for 30 min at 37°C in the dark. The stained
sections were photographed and then analyzed by AlphaImager HP 1.0
software (Alpha Innotech Corp) to calculate the infarct area
rates.
Immunofluorescence staining
Immunofluorescent staining of NeuN was conducted to
determine the neuronal density in the ischemic hippocampi. Briefly,
the rats in each group were subjected to deep anesthesia 3 days
after MCAO. The brain tissues were isolated, exposed to 4%
paraformaldehyde (PFA) at 4°C overnight and then cryoprotected in
30% sucrose solution at 4°C overnight. When tissues sank to the
bottom, they were frozen in optimal cutting temperature compound,
and 25-µm brain sections were prepared using a Leica microtome
(CM3050 S; Leica Microsystems GmbH). Next, anti-rat NeuN antibody
(1:500; cat. no. PA5-78499, Thermo Fisher Scientific, Inc.) was
incubated with free-floating sections. Subsequently, secondary
antibody conjugated with Alexa Fluor® Plus 488 (cat. no.
A32731, Thermo Fisher Scientific, Inc.) was then incubated with the
sections at 25°C for 2 h. The images were captured using a confocal
laser scanning microscope (CLSM; LSM 510 META, Carl Zeiss AG), with
five fields examined for each sample (magnification, x400).
Neuronal changes were quantified against sham control or other
groups.
Barnes maze task
Barnes maze task was performed to examine the
spatial learning and memory capacity of the rats. Briefly, the
tests were conducted 11 days after MCAO. During testing, the time
that each rat spent on finding the hidden escape box was recorded
as escape latency. At 12 days after MCAO, the escape box was
removed, and the time that the rats spent on finding the target
quadrant within 90 sec was recorded as quadrant occupancy. The
behavioral traces of each rat were recorded by a camera and
analyzed by ANY-maze 4.8 video tracking software (Stoelting
Co.).
MitoTracker Red staining
Mitochondrial membrane potential (MMP) was detected
using MitoTracker Red staining, as previously described (12). Briefly, MitoTracker® Red
CMXRos (50 ng/ml in 100 µl saline; C1049, Beyotime Institute of
Biotechnology) was injected via the tail vein into the rats 5 min
before euthanasia. Then, the brain tissues were isolated and fixed
in 4% PFA at 4°C. Hippocampal tissue were cut into 25-µm sections
and stained by DAPI (D8417; Sigma-Aldrich; Merck KGaA) at 37°C for
10 min. Images were then captured using a Zeiss CLSM (Carl Zeiss
AG), and analyzed by LSM 510 META imaging software, version 2.01
(Carl Zeiss AG).
Oxidative stress parameters
For detecting oxidative stress parameters in rat
ischemic hemisphere tissues, lipid peroxidation assay kit (MAK085;
Sigma-Aldrich; Merck KGaA), glutathione (GSH) assay kit (CS0260;
Sigma-Aldrich; Merck KGaA), GSH peroxidase (PX) assay kit (S0058,
Beyotime Institute of Biotechnology), catalase (CAT; S0051,
Beyotime Institute of Biotechnology) and superoxide dismutase (SOD;
S0109, Beyotime Institute of Biotechnology) were purchased and
performed according to the manufacturer's instructions. Briefly,
the tissue (~10 mg) was homogenized on ice in 300 µl cell lysis
buffer (MCL1; Sigma-Aldrich; Merck KGaA). For the detection of
malondialdehyde (MDA) content, insoluble materials and proteins
were removed from homogenized tissues by centrifugation at 13,000 x
g at 4°C for 10 min; 150 µl water containing 3 µl butylated
hydroxytoluene (100X) and 2 N perchloric acid was then added to the
tissues and vortexed at 25°C. Next, 200 µl supernatant was mixed
with 600 µl thiobarbituric acid solution and incubated at 95°C for
60 min. Subsequently, 200 µl reaction mixture were added into a
96-well plate and the absorbance at 532 nm was determined by a
microplate reader (PLUS 384, Molecular Devices, LLC). For the
detection of GSH, 5 µl homogenized tissue was deproteinized using 5
µl 5% 5-sulfosalicylic acid solution. Subsequently, 10 µl mixture
was mixed with 150 µl working solution consisting of 1X assay
buffer, enzyme solution and DTNB, and incubated for 5 min at 25°C.
Next, 50 µl NADPH (0.16 mg/ml) was added into the mixture at 25°C,
and the absorbance at 412 nm was detected. For the detection of GSH
peroxidase (GSH-PX), 180 µl GSH-PX buffer, 5 µl of diluted sample,
11 µl GSH-PX working solution and 4 µl 15 mM superoxide (Cum-OOH)
were added into a 96-well plate and incubated for 20 min at 25°C,
and the absorbance at 340 nm was determined. For the detection of
CAT, 10 µl diluted sample were incubated with 10 µl 250 mM hydrogen
peroxide and 30 µl detection buffer at 25°C for 5 min, and then the
absorbance at 520 nm was read. For the detection of SOD, 20 µl
diluted sample were incubated with 160 µl NBT solution and 20 µl
reaction regent at 25°C for 30 min, and the absorbance at 560 nm
was read.
Mitochondrial dysfunction detection
Mitochondrial dysfunction in the ischemic hemisphere
tissues of the rats was evaluated by detecting the activity of
cytochrome c using a cytochrome c apoptosis assay kit (K257-100,
BioVision, Inc.). The cytochrome c released from mitochondria was
detected by western blotting using the cytochrome c antibody in the
kit. The relative content of cytochrome c in each group was
determined against that in the sham group. Caspase-3 enzyme
activity in rat ischemic hemisphere tissues was measured using a
commercial kit (CASP3C; Sigma-Aldrich; Merck KGaA).
Western blotting
After treatment for 24 h, the ischemic brain tissues
were separated and homogenized on ice. Nuclear and cytoplasmic
proteins were extracted using nuclear and cytoplasmic extraction
kits (SC-003, Invent Biotechnologies, Inc.). Equal amounts of
protein (50 µg) were separated on 12% SDS-PAGE and transferred onto
PVDF membranes (EMD Millipore). After blocking the membranes for 1
h in 5% non-fat milk, the bands were exposed to primary antibodies
against Nrf2 (1:2,000, cat. no. ab137550, Abcam), heme oxygenase-1
(HO-1; 1:2,000; cat. no. ab13243, Abcam), NAD(P)H quinone
oxidoreductase (NQO1; 1:2,000, cat. no. ab28947, Abcam), β-actin
(1:5,000, cat. no. ab8226, Abcam) and histone H3 (1:5,000, cat. no.
ab1791, Abcam) at 4°C overnight. Next, the bands were incubated
with anti-rabbit IgG (cat. no. SA00001-2, ProteinTech Group, Inc.)
or anti-mouse IgG (cat. no. SA00001-1, ProteinTech Group, Inc.) for
2 h. The bands were finally developed by ECL (Pierce; Thermo Fisher
Scientific, Inc.).
Primary culture of hippocampal
neurons
Briefly, the CA1 region of the rat hippocampus was
rapidly separated 3 days after MCAO and sectioned, and then exposed
to trypsin for 30 min at 37°C. After that, the digested cells were
cultured in DMEM (11965084; Thermo Fisher Scientific, Inc.) with
10% FBS (10100147; Gibco; Thermo Fisher Scientific, Inc.), then
transferred to an incubator with 5% CO2 at 37°C for 2 h.
The culture medium was replaced once every 2 days. After culture
for 24 h, hippocampal neurons were used for the subsequent
experiments. The purity of neurons was determined by
immunofluorescence staining of NeuN.
Cell viability
The viability of hippocampal neurons was measured by
MTT assay. Briefly, the neurons (2,000 cells/well) were cultured in
96-well culture plates for 24 h. Then, MTT solution (5 mg/ml) was
added into each well at 37°C for 4 h. Next, MTT was removed, and
150 µl DMSO were added into the wells to solubilize purple formazan
crystals. Absorbance at 570 nm was then read using a microplate
reader (PLUS 384, Molecular Devices, LLC).
Statistical analysis
Statistical data were analyzed using SPSS 19.0 (SPSS
Inc.) and are shown as mean ± standard deviation. Statistical
differences were analyzed by one-way analysis of variance followed
by Tukey's multiple comparisons test. P<0.05 was considered to
indicate statistically significant differences.
Results
Effects of MH, EDA and Bru on neuronal
survival in the CA1 region of the rat hippocampus after MCAO
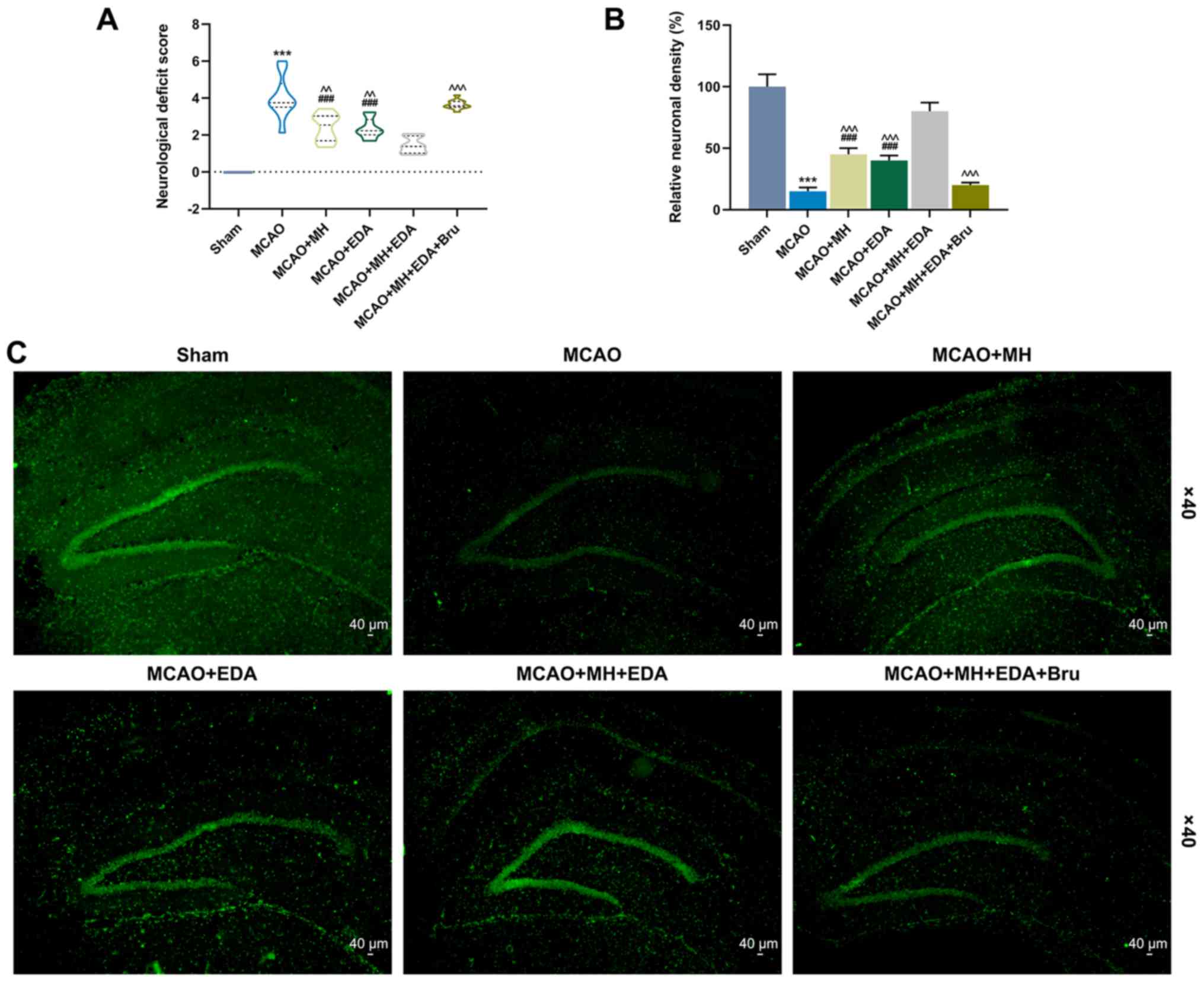
The neurological deficits were evaluated by the
five-point scale 24 h after I/R, and the results revealed that the
neurological deficit score of the rats subjected to MCAO was higher
compared with those in the sham group (P<0.001, Fig. 1A). The neurological deficit scores
of the rats treated by MH or EDA were reduced compared with those
in the MCAO group (P<0.01, Fig.
1A). The neurological deficit score was the lowest in rats
treated by MCAO, MH and EDA, whereas the protective effects of MH
and EDA on the nervous system of rats injected with the Nrf2
pathway inhibitor Bru were significantly reduced (P<0.001,
Fig. 1). Furthermore,
immunofluorescence staining and a neuronal cell marker, NeuN, were
used to analyze neuronal survival in the CA1 region of the rat
hippocampus after MCAO. As shown in Fig. 1B and C, the number of surviving
neurons was reduced in the MCAO group, but was increased in the
MCAO + MH and MCAO + EDA groups (Fig.
1B and C). In addition, the rats co-treated with MH and EDA
exhibited a more marked recovery of neuronal number after I/R,
while Bru reversed the protective effects of MH and EDA
(P<0.001, Fig. 1B).
Effects of MH, EDA and Bru on spatial
learning, memory and mitochondrial function of the rats after MCAO
treatment
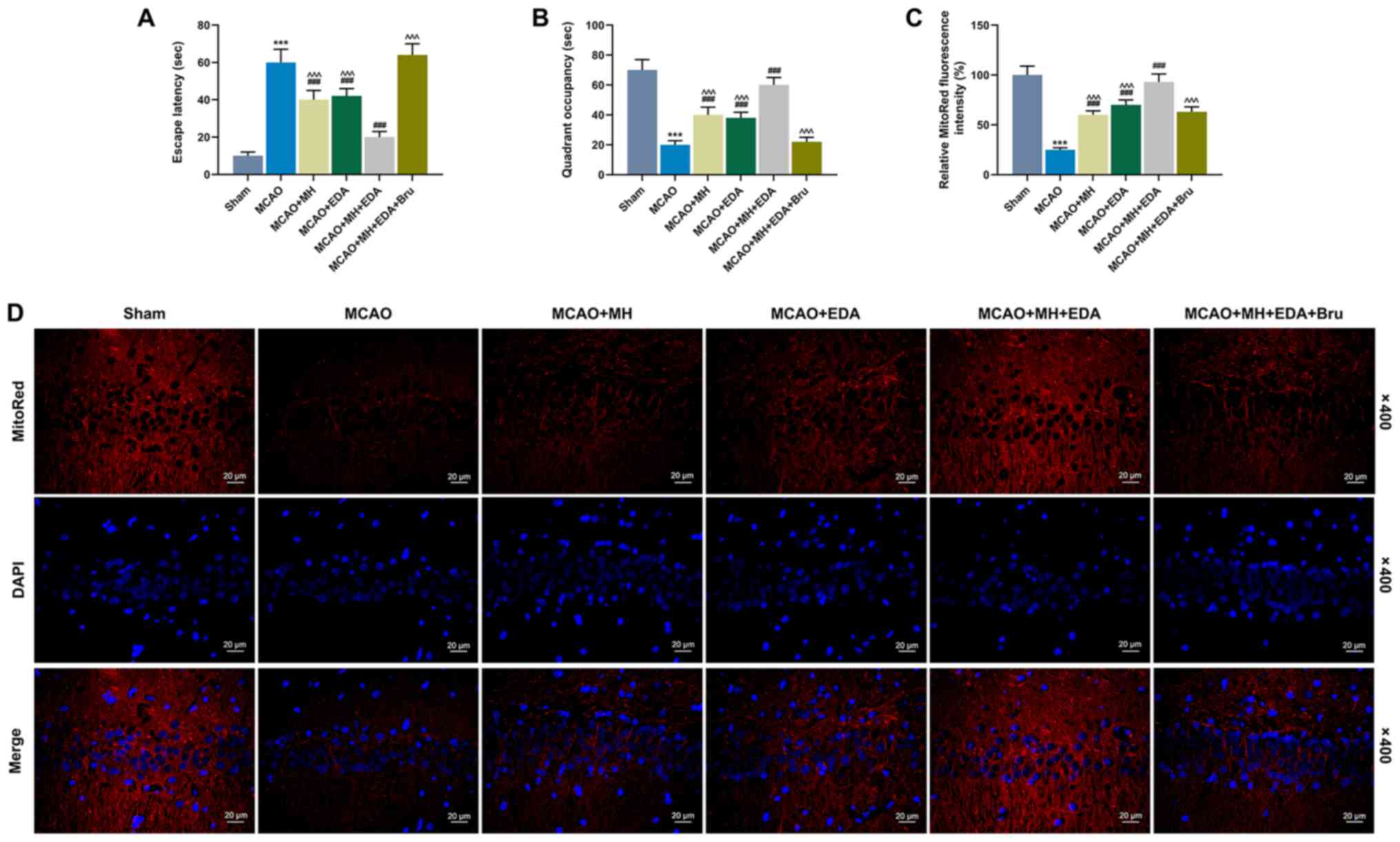
The escape latency and quadrant occupancy were
evaluated by performing the Barnes maze task to explore the effects
of MH, EDA and Bru on spatial learning and memory capacity of the
rats after MCAO treatment. It was observed that the rats in the
MCAO group spent a longer time on finding the hidden escape box
(escape latency) compared with those in the sham group (P<0.001,
Fig. 2A), whereas treatment with
MH or EDA alone, or combined treatment with MH and EDA, could
significantly shorten the escape latency compared with rats in the
MCAO group (P<0.001, Fig. 2A).
In addition, Bru injection increased the escape latency in rats
treated by MH and EDA (P<0.001, Fig. 2A). The rats in the MCAO group spent
a shorter time (quadrant occupancy) in the target quadrant compared
with those in the sham group (P<0.001, Fig. 2B). However, MH and EDA increased
the quadrant occupancy of the rats (P<0.001, Fig. 2B), and Bru injection blocked the
effect of co-treatment with MH and EDA (P<0.001, Fig. 2B). MitoRed fluorescence staining
was performed to detect the MMP, and the data indicated that MCAO
reduced the fluorescence intensity of MMP, which indicated
mitochondrial depolarization and collapse of the MMP, whereas MH
and EDA increased the fluorescence intensity. Additionally,
co-treatment with MH and EDA caused a more significant increase of
fluorescence intensity compared with either treatment alone
(P<0.001, Fig. 2C and D),
which, however, was reduced by Bru injection (P<0.001, Fig. 2C and D).
Effects of MH, EDA and Bru on the levels
of oxidative stress biomarkers, neuronal viability, cytotoxicity
and caspase-3 activity
Oxidative stress is involved in cerebral I/R. Thus,
oxidative stress was further assessed by detecting the levels of
MDA, GSH, GSH-PX, CAT and SOD in ischemic hemisphere tissues
(Fig. 3D and E). The results
demonstrated that the level of MDA was increased in the MCAO group
compared with that in the sham group, while MH or EDA treatment, or
MH and EDA co-treatment, could partly reverse the increase in MDA
content caused by MCAO (P<0.001, Fig. 3A). Additionally, Bru treatment
reversed the effect of co-treatment with MH and EDA on MDA levels
(P<0.001, Fig. 3A).
Furthermore, the levels of GSH, GSH-PX, CAT and SOD in rat ischemic
hemisphere tissues were reduced by MCAO, but increased by MH and
EDA treatment or co-treatment (P<0.001, Fig. 3B-E). Additionally, the levels of
GSH, GSH-PX, CAT and SOD in rat ischemic hemisphere tissues were
reduced in the MCAO + MH + EDA + Bru group compared with those in
the MCAO + MH + EDA group (P<0.001, Fig. 3B-E). The primary hippocampal
neurons were collected from the tissues of the rats in different
groups, and the cell viability was measured by the CCK-8 assay. The
result demonstrated that MCAO reduced the viability of primary
hippocampal neurons, which, however, was increased by MH and EDA
treatment or their co-treatment. Moreover, it was observed that Bru
injection suppressed the effects of MH and EDA (P<0.05, Fig. 3F). Furthermore, the activity of
cytochrome c oxidase and caspase-3 in the mitochondrial fraction
was detected, and the data demonstrated that MCAO increased the
levels of cytochrome c oxidase and caspase-3, while MH and EDA
weakened the effect of MCAO; moreover, Bru treatment blocked the
effects of MH and EDA on cytochrome c oxidase and caspase-3
activities (P<0.001, Fig. 3G and
H).
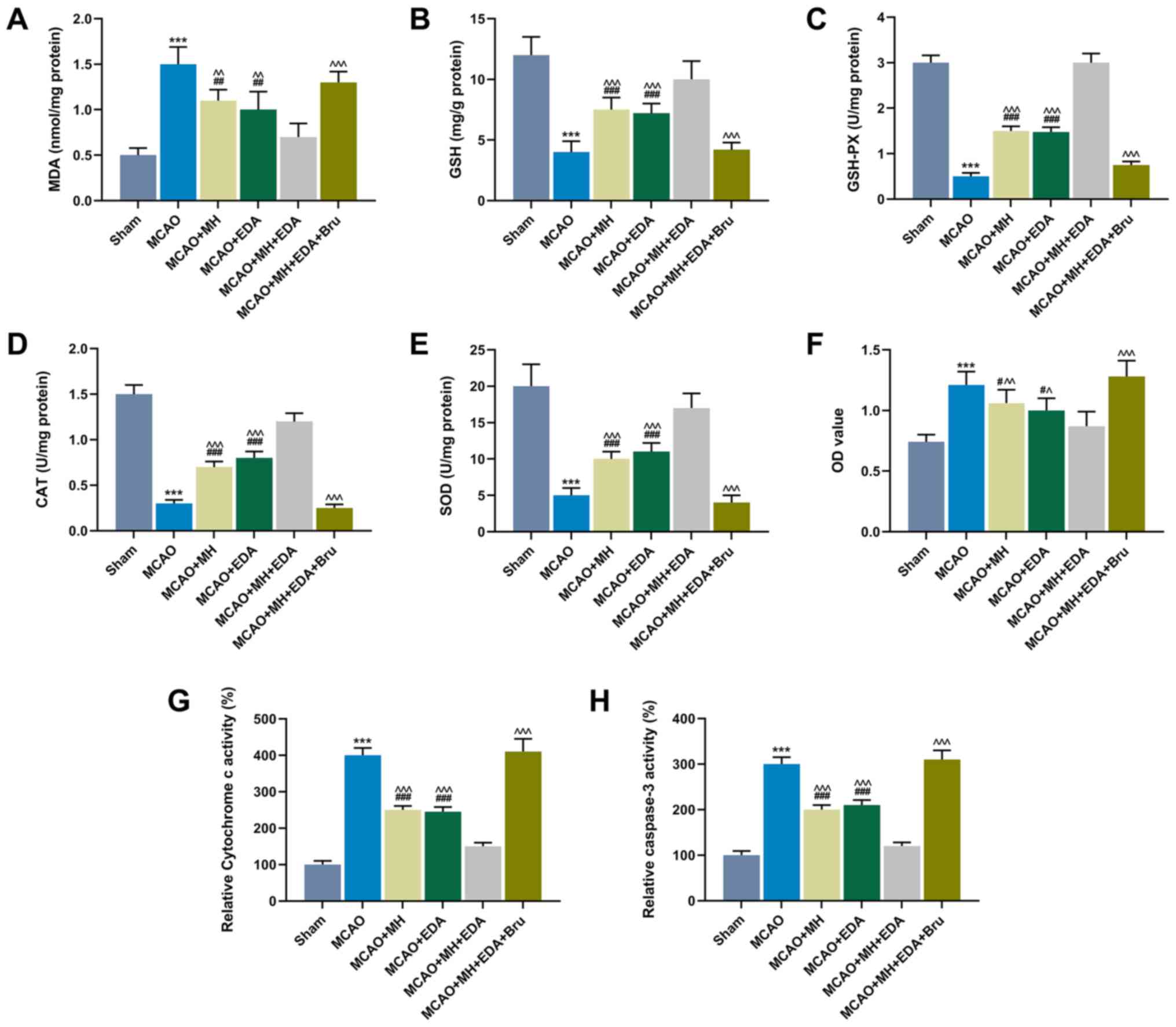 | Figure 3Effects of MH, EDA and Bru on the
levels of oxidative stress indicators, neuronal viability,
cytotoxicity and caspase-3 activity. (A-E) The levels of MDA, GSH,
GSH-PX, CAT and SOD in rat ischemic hemispheres tissues were
determined by colorimetric methods; n=10 mice per group. (F) The
viability of neurons from the CA1 region of hippocampi was
determined by the MTT assay. (G and H) Cytochrome c and caspase-3
in the mitochondrial fractions from each group was detected by
ELISA. ***P<0.001 vs. sham. #P<0.1,
##P<0.01 and ###P<0.001 vs. MCAO.
^P<0.1, ^^P<0.01 and
^^^P<0.001 vs. MCAO + MH + EDA. MCAO, middle cerebral
artery occlusion; MH, mild hypothermia; EDA, edaravone; Bru,
brusatol; MDA, malondialdehyde; GSH, glutathione; GSH-PX,
glutathione peroxidase; CAT, catalase; SOD, superoxide
dismutase. |
Effects of MH, EDA and Bru on the
Nrf2/HO-1 pathway
The Nrf2 pathway is known to act protectively
against oxidative stress. The expressions of cytoplasmic Nrf2
(cNrf2), nuclear Nrf2 (nNrf2), and those of its downstream genes
HO-1 and NQO1 in rat ischemic hemisphere tissues were measured by
western blotting. The data demonstrated that MCAO significantly
increased the expression of cNrf2 (P<0.001) and slightly
increased the expressions of Nrf2, HO-1 and NQO1 (P<0.01 or
P<0.001, Fig. 4A-E). In
addition, MH and EDA markedly increased the levels of HO-1, NQO1
and nNrf2, and simultaneously reduced the level of cNrf2, as
compared with the MCAO group (P<0.001, Fig. 4A-E). The combined treatment with MH
and EDA enhanced the effects caused by treatment with either MH or
EDA alone on the Nrf2/HO-1 pathway, whereas these effects were
reversed by Bru treatment (P<0.05 or P<0.001, Fig. 4A-E).
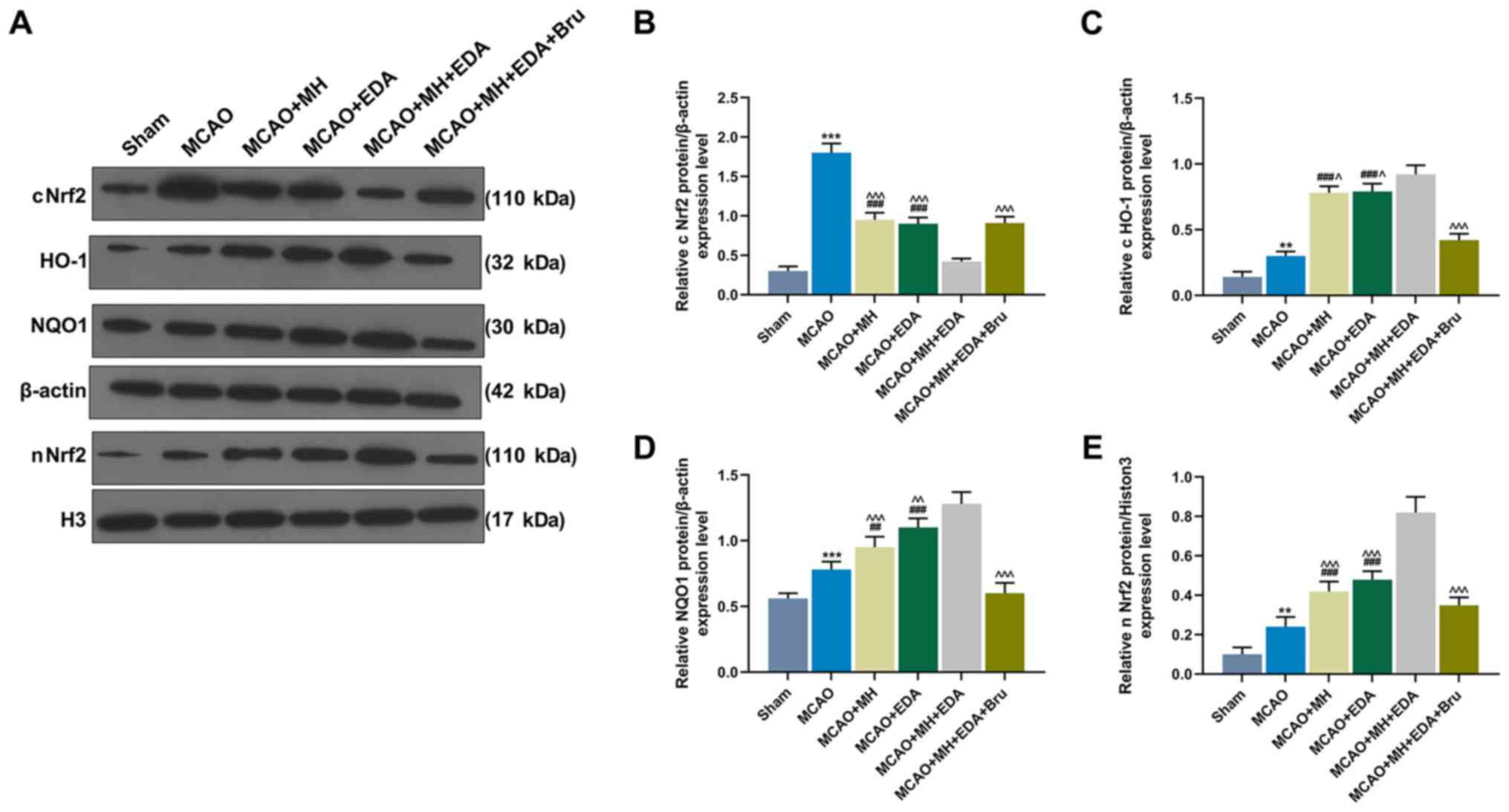 | Figure 4Effects of MH, EDA and Bru on the
activation of the Nrf2/HO-1 pathway. (A-E) Western blot analyses of
expressions of cNrf2, HO-1, NQO1, β-actin, nNrf2 and histone H3 in
rat ischemic hemisphere tissues; n=10 mice per group.
**P<0.01 and ***P<0.001 vs. sham.
##P<0.001 and ###P<0.001 vs. MCAO.
^P<0.1, ^^P<0.01 and
^^^P<0.001 vs. MCAO + MH + EDA. MCAO, middle cerebral
artery occlusion; MH, mild hypothermia; EDA, edaravone; Bru,
brusatol; Nrf2, nuclear factor erythroid 2-related factor 2; cNrf2,
cytoplasmic Nrf2; nNrf2, nuclear Nrf2; HO-1, heme oxygenase-1;
NQO1, NAD(P)H quinone oxidoreductase. |
Discussion
It has been demonstrated that MH used together with
other drugs or treatments may enhance the protective effects of MH
against cerebral I/R injury. For example, a recent research
revealed that MH with bone mesenchymal stem cell transplantation
improved the prognosis of rats with cerebral ischemia (19). Combined therapy with phenothiazines
and MH also enhanced neuroprotection in ischemic rats through
regulating the PI3K/Akt pathway (20). It has also been reported that the
Notch inhibitor DAPT combined with MH exerted a synergistic effect
on post-stroke seizures (21). It
was reported that MH in combination with phenothiazine drugs
achieved a better long-term motor performance after acute stroke
(22). Moreover, MH used with
hydrogen sulfide treatment suppressed apoptosis and pathological
injury of hippocampi of rats with cerebral I/R injury (23). These findings indicated that MH
combined with other treatments may be a promising therapy in the
management of cerebral I/R injury. The present study demonstrated
that the neurological deficit score was significantly reduced and
neuronal density was increased in the CA1 region of the rat
hippocampi, suggesting that EDA combined with MH exerted a
synergistic effect against I/R injury.
Although neurological deficit score evaluation and
Barnes maze task test were performed, it may be a limitation that
the TTC dying method was not used to assess the brain infarct area.
The hippocampus is responsible for spatial learning and memory
(24), which may be disturbed when
the hippocampus sustains cerebral I/R injury (25). Therefore, the Barnes Maze task that
helps evaluate spatial learning and memory ability was performed in
the present study, and it was observed that EDA combined with MH
could improve the spatial learning and memory ability of rats with
cerebral I/R injury. MitoRed fluorescence staining was used to
evaluate MMP. Under normal physiological conditions, mitochondria
with high MMP can be marked by Mito-Tracker Red CMXRos, while MMP
is reduced in mitochondrial dysfunction and therefore cannot be
marked by Mito-Tracker Red CMXRos under pathological conditions or
when cell apoptosis occurs. The present study demonstrated that
mitochondrial dysfunction in rat hippocampi was induced by MCAO,
but was significantly restored by the combined therapy of EDA with
MH.
The CAT, SOD, GSH and GSH-PX enzymes are involved in
the antioxidant defense system against cell oxidative stress
(26), and MDA is indicative of
IR-induced mitochondrial oxidative damage (27). In the present study, it was
observed that MDA was increased, but CAT, SOD, GSH and GSH-PX were
reduced in rats subjected to MCAO, suggesting that mitochondrial
oxidative damage was induced by cerebral I/R injury. Importantly,
restoration of dysregulated MDA, CAT, SOD, GSH and GSH-PX levels
was detected in rats treated by MCAO and in those treated by
combined EDA and MH. Thus, it may be inferred that EDA combined
with MH alleviated the mitochondrial dysfunction induced by
cerebral I/R.
Neurogenesis may be enhanced under pathological
conditions, including cerebral ischemic injury, and it promotes
repair of brain injury after stroke. It was demonstrated that
neurogenesis can be suppressed by EDA in rats with cerebral I/R
injury (28). However, it has also
been shown that EDA protects hippocampal neurons pre- and
post-treatment and enhances neurogenesis after global cerebral
ischemia (29). In the present
research, the viability of neurons separated from the CA1 region of
the hippocampus was increased by MCAO, but was suppressed by MH and
EDA. In addition, the increased activities of cytochrome c and
caspase-3 were partially reversed by the combination of MH with EDA
in rats with cerebral I/R. I/R induces dysfunction of mitochondria
and reduces cytochrome c oxidase activity in rat hippocampal
neurons (30,31). Increased activity of cytochrome c
increases oxidative stress and cell apoptosis. Caspase-3 is
considered to be a cell apoptosis marker; thus, it was inferred
that MCAO may promote neurogenesis and increase the levels of
apoptosis-associated factors, whereas MH and EDA may inhibit this
process. It should be noted that the effect of the combined therapy
with MH and EDA on cell apoptosis require further
investigation.
Nrf2 is a regulator of endogenous antioxidant
defense and may be a potential target for oxidative stress in
stroke (32). Nrf2 induces
HO-1expression and release of GSH to resist oxidative stress
(33). It was reported that BRCA1
protects neurons from cerebral I/R injury through activating the
Nrf2-associated antioxidant pathways (34). It has been reported that EDA
enhances the oxidant defense capacity in brain tissue through
promoting Nrf2/HO-1 signaling (35). MH was also reported to protect
cerebral cortex and hippocampus neurons from cerebral I/R injury
through the Nrf2/ARE pathway (36). In the present study, by injecting
the Nrf2 pathway inhibitor Bru into rats treated by MH combined
with EDA, it was observed that the protective effect of MH combined
with EDA is linked to the Nrf2 pathway. A previous study
demonstrated that nuclear-translocated Nrf2 binds to ARE and
p300/CBP to promote the transcription of downstream target genes,
such as HO-1 (37). The present
study revealed that nuclear location of Nrf2 was increased and the
expression of the Nrf2 downstream genes NQO1 and HO-1 was
upregulated in the treated by MH combined with EDA.
In conclusion, a rat cerebral ischemia model was
constructed in the present study, and demonstrated that combined
treatment with MH and EDA improved rat neurological function and
increased neuronal survival. In addition, the Nrf2 pathway
inhibitor, Bru, could block the protective effect of MH and EDA.
These findings indicated that MH combined with EDA exert a
synergistic effect against cerebral I/R injury through the
Nrf2/HO-1 pathway. However, a limitation of the present study was
that there was no detection of more indicators of oxidative stress,
and further research on these findings is required.
Funding
The present study was supported by the Hainan
Provincial Health and Family Planning Research Project (grant no.
18A200002), the Hainan Provincial Higher Education Research Project
(grant no. Hnky2018-50) and the National Natural Science Foundation
of China (grant no. 81660270).
Availability of data and materials
The data sets generated and/or analyzed during the
present study are available from the corresponding author on
reasonable request.
Authors' contributions
Substantial contributions to conception and design:
HY and ZW; data acquisition, data analysis and interpretation: XW,
CG, RL, FK and MD; drafting the article and critically revising it
for important intellectual content: HY and ZW. All the authors have
read and approved the final version to be published and agree to be
accountable for all aspects of the work in ensuring that questions
related to the accuracy or integrity of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
All the animal experiments were approved by the
Institutional Animal Ethics Committee of Hainan Medical University
(no. C2017051922A).
Patient consent for publication
Not applicable.
Competing interests
All the authors declare that they have no competing
interests.
Acknowledgements
Not applicable.
References
|
1
|
Alkhachroum Am, Miller B, Chami T,
Tatsuoka C and Sila C: A troponin study on patients with ischemic
stroke, intracerebral hemorrhage and subarachnoid hemorrhage: Type
II myocardial infarction is significantly associated with stroke
severity, discharge disposition and mortality. J Clin Neurosci.
64:83–88. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Feigin VL, Mensah GA, Norrving B and
Murray CJ: Atlas of the Global Burden of Stroke (1990–2013): The
GBD 2013 Study. Neuroepidemiology. 45:230–236. 2015. View Article : Google Scholar :
|
|
3
|
Vos T, Abajobir AA, Abate KH, Abbafati C,
Abbas KM, Abd-Allah F, Abdulkader RS, Abdulle AM, Abebo TA, Abera
SF, et al: GBD 2016 Disease and Injury Incidence and Prevalence
Collaborators: Global, regional, and national incidence,
prevalence, and years lived with disability for 328 diseases and
injuries for 195 countries, 1990–2016: A systematic analysis for
the Global Burden of Disease Study 2016. Lancet. 390:1211–1259.
2017. View Article : Google Scholar
|
|
4
|
Cheung RT: The utility of melatonin in
reducing cerebral damage resulting from ischemia and reperfusion. J
Pineal Res. 34:153–160. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zerna C, Thomalla G, Campbell BCV, Rha JH
and Hill MD: Current practice and future directions in the
diagnosis and acute treatment of ischaemic stroke. Lancet.
392:1247–1256. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Leng T, Shi Y, Xiong ZG and Sun D:
Proton-sensitive cation channels and ion exchangers in ischemic
brain injury: New therapeutic targets for stroke? Prog Neurobiol.
115:189–209. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tang XN, Liu L, Koike MA and Yenari MA:
Mild hypothermia reduces tissue plasminogen activator-related
hemorrhage and blood brain barrier disruption after experimental
stroke. Ther Hypothermia Temp Manag. 3:74–83. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tu Y, Guo C, Song F, Huo Y, Geng Y, Guo M,
Bao H, Wu X and Fan W: Mild hypothermia alleviates diabetes
aggravated cerebral ischemic injury via activating autophagy and
inhibiting pyroptosis. Brain Res Bull. 150:1–12. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang GS, Zhou XY, An XF, Liu XJ, Zhang YJ
and Yu D: Mild hypothermia inhibits the Notch 3 and Notch 4
activation and seizure after stroke in the rat model. Pathol Res
Pract. 214:1008–1016. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu LQ, Liu XR, Zhao JY, Yan F, Wang RL,
Wen SH, Wang L, Luo YM and Ji XM: Brain-selective mild hypothermia
promotes long-term white matter integrity after ischemic stroke in
mice. CNS Neurosci Ther. 24:1275–1285. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yuan Y, Zha H, Rangarajan P, Ling EA and
Wu C: Anti-inflammatory effects of Edaravone and Scutellarin in
activated microglia in experimentally induced ischemia injury in
rats and in BV-2 microglia. BMC Neurosci. 15:1252014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li L, Yang R, Li P, Lu H, Hao J, Li L,
Tucker D and Zhang Q: Combination Treatment with Methylene Blue and
Hypothermia in Global Cerebral Ischemia. Mol Neurobiol.
55:2042–2055. 2018. View Article : Google Scholar
|
|
13
|
Shibuta S, Varathan S, Kamibayashi T and
Mashimo T: Small temperature variations alter edaravone-induced
neuroprotection of cortical cultures exposed to prolonged hypoxic
episodes. Br J Anaesth. 104:52–58. 2010. View Article : Google Scholar
|
|
14
|
Dodson M, de la Vega MR, Cholanians AB,
Schmidlin CJ, Chapman E and Zhang DD: Modulating NRF2 in Disease:
Timing Is Everything. Annu Rev Pharmacol Toxicol. 59:555–575. 2019.
View Article : Google Scholar :
|
|
15
|
Shou L, Bei Y, Song Y, Wang L, Ai L, Yan Q
and He W: Nrf2 mediates the protective effect of edaravone after
chlorpyrifos-induced nervous system toxicity. Environ Toxicol.
34:626–633. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li C, Mo Z, Lei J, Li H, Fu R, Huang Y,
Luo S and Zhang L: Edaravone attenuates neuronal apoptosis in
hypoxic-ischemic brain damage rat model via suppression of TRAIL
signaling pathway. Int J Biochem Cell Biol. 99:169–177. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cen J, Zhao N, Huang WW, Liu L, Xie YY,
Gan Y, Wang CJ and Ji BS: Polyamine analogue QMA attenuated
ischemic injury in MCAO rats via ERK and Akt activated Nrf2/HO-1
signaling pathway. Eur J Pharmacol. 844:165–174. 2019. View Article : Google Scholar
|
|
18
|
Longa EZ, Weinstein PR, Carlson S and
Cummins R: Reversible middle cerebral artery occlusion without
craniectomy in rats. Stroke. 20:84–91. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bi M, Wang J, Zhang Y, Li L, Wang L, Yao
R, Duan S, Tong S and Li J: Bone mesenchymal stem cells
transplantation combined with mild hypothermia improves the
prognosis of cerebral ischemia in rats. PLoS One. 13:pp.
e01974052018, View Article : Google Scholar : PubMed/NCBI
|
|
20
|
An H, Duan Y, Wu D, Yip J, Elmadhoun O,
Wright JC, Shi W, Liu K, He X, Shi J, et al: Phenothiazines Enhance
Mild Hypothermia-induced Neuroprotection via PI3K/Akt Regulation in
Experimental Stroke. Sci Rep. 7:74692017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang GS, Zhou XY, An XF, Liu XJ, Zhang YJ
and Yu D: Synergistic effect of mild hypothermia and the Notch
inhibitor DAPT against post stroke seizures. Biomed Pharmacother.
96:675–684. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu S, Geng X, Forreider B, Xiao Y, Kong
Q, Ding Y and Ji X: Enhanced beneficial effects of mild hypothermia
by phenothiazine drugs in stroke therapy. Neurol Res. 37:pp.
454–460. 2015, View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dai HB, Xu MM, Lv J, Ji XJ, Zhu SH, Ma RM,
Miao XL and Duan ML: Mild hypothermia combined with hydrogen
sulfide treatment during resuscitation reduces hippocampal neuron
apoptosis via NR2A, NR2B, and PI3K-Akt signaling in a rat model of
cerebral ischemia-reperfusion injury. Mol Neurobiol. 53:4865–4873.
2016. View Article : Google Scholar
|
|
24
|
Jeffery KJ: The Hippocampus: From Memory,
to Map, to Memory Map. Trends Neurosci. 41:64–66. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xu F, Zhang G, Yin J, Zhang Q, Ge MY, Peng
L, Wang S and Li Y: Fluoxetine mitigating late-stage cognition and
neurobehavior impairment induced by cerebral ischemia reperfusion
injury through inhibiting ERS-mediated neurons apoptosis in the
hippocampus. Behav Brain Res. 370:1119522019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Das KK, Gupta AD, Dhundasi SA, Patil AM,
Das SN and Ambekar JG: Protective role of L-ascorbic acid on
antioxidant defense system in erythrocytes of albino rats exposed
to nickel sulfate. Biometals. 20:177–184. 2007. View Article : Google Scholar
|
|
27
|
Yang Y, Duan W, Jin Z, Yi W, Yan J, Zhang
S, Wang N, Liang Z, Li Y, Chen W, et al: JAK2/STAT3 activation by
melatonin attenuates the mitochondrial oxidative damage induced by
myocardial ischemia/reperfusion injury. J Pineal Res. 55:275–286.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang P, Li W, Li L, Wang N, Li X, Gao M,
Zheng J, Lei S, Chen X, Lu H, et al: Treatment with edaravone
attenuates ischemic brain injury and inhibits neurogenesis in the
subventricular zone of adult rats after focal cerebral ischemia and
reperfusion injury. Neuroscience. 201:297–306. 2012. View Article : Google Scholar
|
|
29
|
Lei S, Zhang P, Li W, Gao M, He X, Zheng
J, Li X, Wang X, Wang N, Zhang J, et al: Pre- and posttreatment
with edaravone protects CA1 hippocampus and enhances neurogenesis
in the subgranular zone of dentate gyrus after transient global
cerebral ischemia in rats. ASN Neuro. 6:17590914145584172014.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Racay P, Tatarková Z, Drgová A, Kaplan P
and Dobrota D: Ischemia-reperfusion induces inhibition of
mitochondrial protein synthesis and cytochrome c oxidase activity
in rat hippocampus. Physiol Res. 58:127–138. 2009.
|
|
31
|
Zhong S, Li Z, Huan L and Chen BY:
Neurochemical mechanism of electroacupuncture: Anti-injury effect
on cerebral function after focal cerebral ischemia in rats. Evid
Based Complement Alternat Med. 6:51–56. 2009. View Article : Google Scholar :
|
|
32
|
Zhang R, Xu M, Wang Y, Xie F, Zhang G and
Qin X: Nrf2-a promising therapeutic target for defensing against
oxidative stress in stroke. Mol Neurobiol. 54:6006–6017. 2017.
View Article : Google Scholar
|
|
33
|
Wu L, Li HH, Wu Q, Miao S and Liu ZJ:
Lipoxin A4 activates Nrf2 pathway and ameliorates cell damage in
cultured cortical astrocytes exposed to oxygen-glucose
deprivation/reperfusion insults. J Mol Neurosci. 56:848–857. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xu P, Liu Q, Xie Y, Shi X, Li Y, Peng M,
Guo H, Sun R, Li J, Hong Y, et al: Breast cancer susceptibility
protein 1 (BRCA1) rescues neurons from cerebral
ischemia/reperfusion injury through NRF2-mediated antioxidant
pathway. Redox Biol. 18:158–172. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li Z, Yulei J, Yaqing J, Jinmin Z, Xinyong
L, Jing G and Min L: Protective effects of tetramethylpyrazine
analogue Z-11 on cerebral ischemia reperfusion injury. Eur J
Pharmacol. 844:156–164. 2019. View Article : Google Scholar
|
|
36
|
Xia D and Zhang H: Effects of mild
hypothermia on expression of NF-E2-related factor 2 and
heme-oxygenase-1 in cerebral cortex and hippocampus after
cardiopulmonary resuscitation in rats. Iran J Basic Med Sci.
20:1002–1008. 2017.PubMed/NCBI
|
|
37
|
Kim SW, Lee HK, Shin JH and Lee JK:
Up-down regulation of HO-1 and iNOS gene expressions by ethyl
pyruvate via recruiting p300 to Nrf2 and depriving It from p65.
Free Radic Biol Med. 65:468–476. 2013. View Article : Google Scholar : PubMed/NCBI
|