Introduction
Lung cancer is the most commonly diagnosed cancer
and the leading cause of cancer-related mortality worldwide, with
approximately 2.1 million new cases and 1.8 million deaths in 2018
(1). Non-small cell lung cancer
(NSCLC) is the major histological subtype of lung cancer, which
accounts for 80-85% of all cases (2). Despite recent advances in the
treatment of NSCLC, such as targeted therapy and immunotherapy, the
long-term prognosis for NSCLC patients remains poor, and the 5-year
survival rate is less than 15% (3). Therefore, it is urgently required to
elucidate the detailed molecular pathogenesis of NSCLC, so as to
identify more accurate prognostic markers and therapeutic
targets.
SET and MYND domain-containing protein 3 (SMYD3) is
a member of the SET and MYND domain family, a special class of
histone and non-histone lysine methyltransferase (4). SMYD3 functions by catalyzing lysine
methylation, mainly through the bi/trimethylation of histone H3 at
lysine 4 (H3K4), to render chromatin more accessible and activate
downstream gene transcription (5,6). In
addition, SMYD3 can transactivate downstream targets by binding to
the 5′-CCCTCC-3′ or 5′-GCAGGG-3′ motif in the promoter region of
target genes, and interacting with RNA polymerase II and RNA
helicase (5). In the past decade,
emerging evidence has suggested an oncogenic role of SMYD3 in human
cancer. The overexpression of SMYD3 has been observed in breast,
hepatocellular, colorectal, gastric, bladder, prostate, cervical,
glioma and esophageal squamous cell carcinomas, where it was
revealed to be involved in cell proliferation, migration, invasion,
epithelial-mesenchymal transition and autophagic activation
(7-14). Furthermore, a variable number of
tandem repeat (VNTR) polymorphism in the SMYD3 promoter
region, namely homozygosity for an allele with three tandem repeats
of a CCGCC (3/3), was found to be associated with an increased risk
of breast, hepatocellular and colorectal carcinomas (15). As for NSCLC, Barlesi et al
found that the SMYD3 VNTR genotype was not correlated with
either the tested clinicopathological characteristics or the risk
of NSCLC (16). However, the
expression of SMYD3 on protein levels in clinical NSCLC samples, as
well as the functional role of SMYD3 in NSCLC progression, remain
unknown.
In the present study, the expression of SMYD3 in
NSCLC and matched adjacent normal tissues was evaluated by
immunohistochemistry (IHC), and its clinical and prognostic
significance was analyzed. Moreover, the biological roles of SMYD3
in NSCLC cell proliferation, apoptosis, migration, invasion and
drug resistance, as well as the molecular basis for such effects,
were also characterized. The present findings demonstrated that
SMYD3 may serve as an oncogene in NSCLC development and
progression, recommending SMYD3 as a promising prognostic marker
and therapeutic target for NSCLC.
Materials and methods
Patients and tissue microarray (TMA)
Primary NSCLC and corresponding pericarcinous lung
tissues were consecutively collected from patients (89 men, 66
women; mean ± SD age 62.35 ± 10 years, range 38 to 81 years) who
underwent surgical resection at Fudan University Shanghai Cancer
Center (Shanghai, China) between 2010 and 2012. TMAs were
constructed as previously described (17). Two cores with a 1.6-mm diameter
were obtained from the original paraffin block of each sample.
Clinicopathological information was retrospectively reviewed, and
survival data was recorded on the basis of follow-up clinic visits
or telephone calls. The pathological stages were determined based
on the 7th edition of the lung cancer staging system proposed by
the International Association for the Study of Lung Cancer (IASLC)
(18). Patients who had received
neoadjuvant chemotherapy or radiotherapy, or had a history of other
malignancies were excluded. In total, 155 NSCLC cases were
enrolled. The present study was approved by the Ethics Committee of
Fudan University Shanghai Cancer Center. All experiments were in
accordance with the declaration of Helsinki and the approved
guidelines of our institution. Written informed consent was
obtained from all participants.
IHC
The TMAs were subjected to IHC using anti-SMYD3
antibody (1:800; cat. no. ab187149, Abcam) as previously described
(9). Concentration-matched
nonspecific rabbit IgG was used as an isotype control. The IHC
results were reviewed again by two trained pathologists. Staining
intensity was scored as follows: Negative (0), weak (1), moderate (2) and intense (3). The percentage of positive cells was
scored as follows: 0% (0), 1-25% (1), 26-50% (2), 51-75% (3) and 76-100% (4). The final SMYD3 IHC score was the
multiplication of these two scores, which were defined as low
(scores of 0-3) or high (scores of 4-12).
Cell culture and cell proliferation
assay
Human NSCLC H727, A549, NCI-H1299, H1650, NCI-H1975,
HCC4006, PC-9, HCC827, H2170 and H226 cell lines, and human
bronchial epithelial BEAS-2B cell line were obtained from the
American Type Culture Collection (ATCC), and cultured in RPMI-1640
or DMEM (Thermo Fisher Scientific, Inc.) supplemented with 10%
fetal bovine serum (FBS). All cells were authenticated by short
tandem repeat profiling (Thermo Fisher Scientific, Inc.) and
confirmed to be mycoplasma-free. The in vitro cell
proliferation was examined by sulforhodamine B (SRB) assay.
Briefly, the cells in 96-well plates were fixed by gently adding
cold trichloroacetic acid (100 µl; 10%) and incubated for 30
min at 4°C. The supernatant was discarded, and the plates were
washed three times with tap water and air dried. SRB solution (100
µl; 0.4% in 1% acetic acid) was added, and the plates were
incubated for 20 min at room temperature. The unconjugated SRB was
washed by 1% acetic acid, and the conjugated SRB was dissolved in
Tris solution (100 µl; 10 mM). The absorbance was measured
at 560 nm using a microplate reader.
siRNA Transfection
Cells (1×105 cells/well in 6-well plates)
were transfected with siRNA using Lipofectamine RNAiMAX (Thermo
Fisher Scientific, Inc.), following the manufacturer's
instructions. The target sequences of double-stranded nucleotides
used in siRNA transfection were
5′-GCAGAGTTGTCTTCAAACT-3′(SMYD3siRNA-1) and
5′-GGATGCTGATATGCTAACT-3′(SMYD3siRNA-2) for SMYD3 knockdown, and
5′-TTCTCCGAACGTGTCACGT-3′ as a negative control (NC) (Shanghai
GenePharma Co., Ltd.).
Lentiviral vectors and cell
infection
Lentiviral vectors were constructed with OBiO
Biotechnology, using pRLe nti-CMV-SMYD3-3FLAG-PGK-Puro plasmids,
which carry the full-length human SMYD3 (GenBank accession
NM_001167740.2) coding sequence. Cell infection was performed as in
our previous study (19).
Western blotting
Western blot analysis was performed as previously
described (9). The primary
antibodies, including anti-SMYD3 (cat. no. ab187149, Abcam),
anti-Bcl-2-like protein 11 (Bim; cat. no. 2933S), anti-Bcl-2
homologous antagonist/killer (Bak; cat. no. 6947S), anti-Bax (cat.
no. 14796S), anti-B-cell lymphoma 2 (Bcl-2; cat. no. 15071S),
anti-Bcl-xl (cat. no. 2764S), anti-matrix metalloproteinase-2
(MMP-2; cat. no. 13132S), anti-MMP-9 (cat. no. 13667S) and
anti-β-actin (cat. no. 3700S; all 1:1,000; all from Cell Signaling
Technology, Inc.) were used at the manufacturer's recommended
dilution. The secondary antibodies were peroxidase AffiniPure goat
anti-rabbit IgG (cat. no. 111-035-003) or anti-mouse IgG (cat. no.
115-035-003; both 1:3,000; both from Jackson ImmunoResearch, Inc.).
Densitometric analysis was performed using ImageJ software (v1.47,
Rawak Software, Inc.). The measured intensities for all samples
were first normalized to their respective internal controls and
then were compared with BEAS-2B cells, which was set to 100%.
Cell apoptosis analysis
NSCLC cells (1×105 cells per tube) were
harvested with EDTA-free trypsin 72 h after transfection, stained
using a FITC Annexin V Apoptosis Detection kit (BD Biosciences)
following the manufacturer's protocol, and then examined using a
flow cytometer (BD Biosciences). For the evaluation of cisplatin
resistance, SMYD3-silenced or -over-expressed NSCLC cells were
treated with cisplatin (Majorbio Bio-Pharm) at various doses (1, 2
and 4 µM for A549; 0.7, 1.5 and 3 µM for PC-9; 1, 2
and 5 µM for H1299) for 72 h, and then harvested for cell
apoptosis analysis.
Cell migration and invasion assays
Cell migration and invasion assays were conducted
using the Transwell method, as previously described (12). The number of migrated or invaded
cells were counted using a bright-field imaging system on a
microscope (Olympus BX51; Olympus Corporation) with five randomly
selected fields (magnification, ×200).
Statistical analysis
Statistical analysis was conducted with SPSS v19.0
(IBM Corp.). Comparisons between two groups were performed using
Student's t-test. Associations between two categorical variables
were examined using Chi-square test or Fisher's exact test. The
survival curves were plotted using the Kaplan Meier method, and
compared using the log-rank test. Univariate and multivariate Cox
proportional hazard models were employed to identify independent
predictors. All tests were two sided and P<0.05 was considered
to indicate a statistically significant difference.
Results
SMYD3 is upregulated in NSCLC and
associated with aggressive clinicopathological characteristics
The expression of SMYD3 was assessed in 155 pairs of
NSCLC and corresponding pericarcinous lung tissues by IHC.
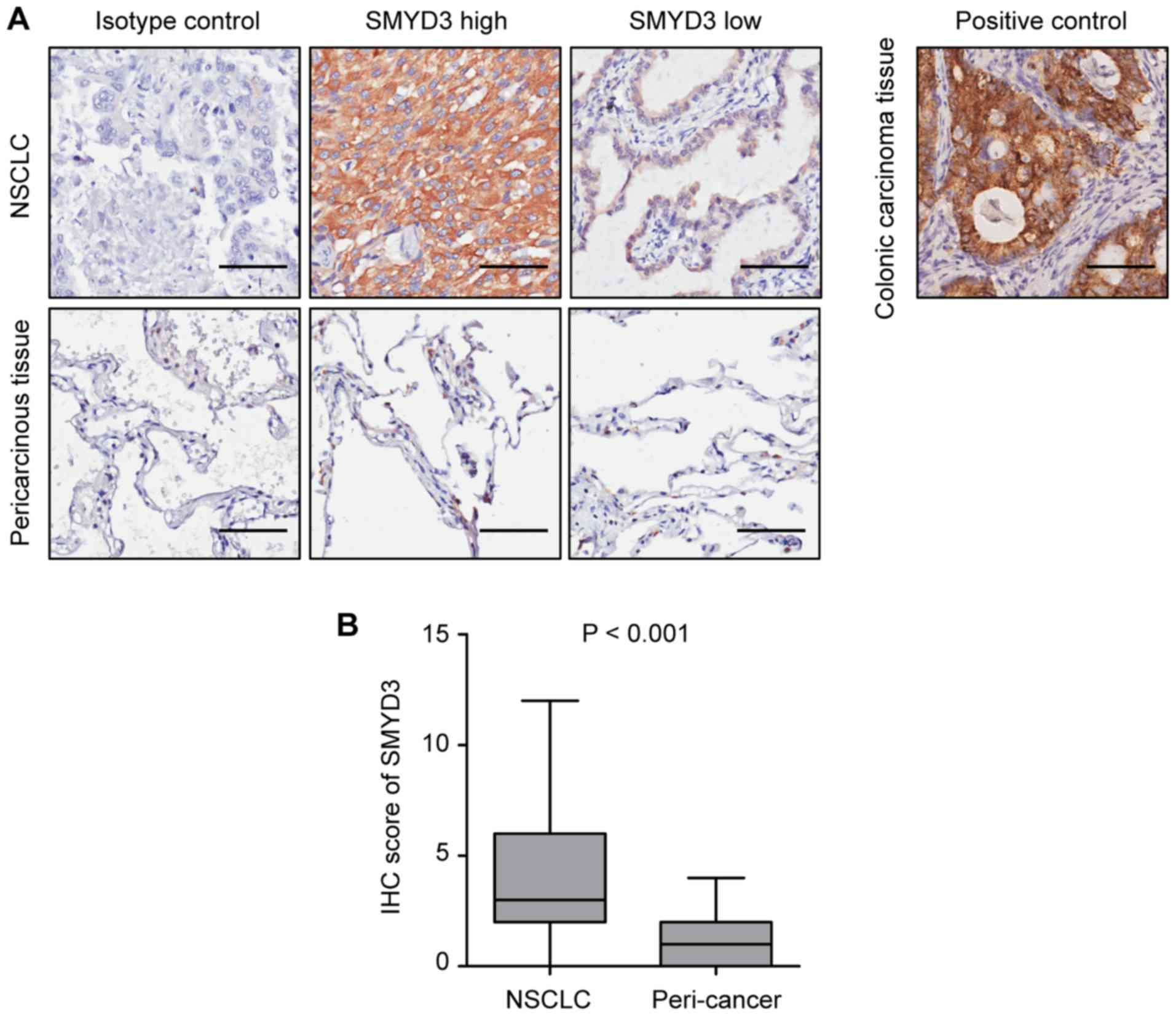
Representative images revealed that SMYD3 expression was mainly
localized in the cytoplasm of NSCLC cells (Fig. 1A), which was consistent with
previous studies (20,21). It was determined that SMYD3 was
significantly upregulated in NSCLC tissues compared with that in
pericarcinous lung tissues (Fig.
1B).
To analyze the association between SMYD3 and
clinicopathological characteristics, 155 NSCLC specimens were
divided into 'SMYD3 high' (scores of 4-12) and 'SMYD3 low' (scores
of 0-3) groups, according to the median expression value.
Consistent with the aforementioned findings, the proportion of
'SMYD3 high' samples was significantly higher in NSCLC than in
paired normal lung tissues (48.4 vs. 3.2%, P<0.001) (Table I). The present results revealed
that a high SMYD3 expression was significantly associated with
never-smoked history, more advanced pathological stage, larger
tumor size, and the presence of lymphovascular invasion (LVI),
pleural invasion (PI) and distant metastasis (Table II).
 | Table IProtein expression levels of SMYD3 in
NSCLC and corresponding pericarcinous lung tissues. |
Table I
Protein expression levels of SMYD3 in
NSCLC and corresponding pericarcinous lung tissues.
| Tissue sample | No. of
patients | SMYD3
| P-value |
|---|
| Low (%) | High (%) |
|---|
| NSCLC | 155 | 80 (51.6) | 75 (48.4) | <0.001a |
| Peri-cancer | 155 | 150 (96.8) | 5 (3.2) | |
| Table IIClinicopathological characteristics
according to SMYD3 protein expression in NSCLC. |
Table II
Clinicopathological characteristics
according to SMYD3 protein expression in NSCLC.
| Variables | No. of
patients | SMYD3
| P-value |
|---|
| High (%) | Low (%) |
|---|
| Sex | | | | |
| Male | 89 | 39 (43.8) | 50 (56.2) | 0.186a |
| Female | 66 | 36 (54.5) | 30 (45.5) | |
| Age (years) | | | | |
| >60 | 94 | 41 (43.6) | 53 (56.4) | 0.14a |
| ≤60 | 61 | 34 (55.7) | 27 (44.3) | |
| Smoking
history | | | | |
| Ever | 68 | 26 (38.2) | 42 (61.8) | 0.025a |
| Never | 87 | 49 (56.3) | 38 (43.7) | |
| Stage | | | | |
| I | 66 | 25 (37.9) | 41 (62.1) | 0.024a |
| II-IV | 89 | 50 (56.2) | 39 (43.8) | |
| Tumor size
(cm) | | | | |
| > 3.0 | 76 | 43 (56.6) | 33 (43.4) | 0.045a |
| ≤ 3.0 | 79 | 32 (40.5) | 47 (59.5) | |
| LVI | | | | |
| Present | 41 | 34 (82.9) | 7 (17.1) | <0.001a |
| Absent | 114 | 41 (36.0) | 73 (64.0) | |
| PI | | | | |
| Present | 65 | 38 (58.5) | 27 (41.5) | 0.033a |
| Absent | 90 | 37 (41.1) | 53 (58.9) | |
| N status | | | | |
| N0 | 79 | 33 (41.8) | 46 (58.2) | 0.093a |
| N1/N2N3 | 76 | 42 (55.3) | 34 (44.7) | |
| M status | | | | |
| M0 | 150 | 70 (46.7) | 80 (53.3) | 0.025b |
| M1 | 5 | 5 (100) | 0 (0) | |
|
Differentiation | | | | |
| Well/moderate | 109 | 55 (50.5) | 54 (49.5) | 0.427a |
| Poor | 46 | 20 (43.5) | 26 (56.5) | |
High SMYD3 expression predicts poor
outcomes in NSCLC patients
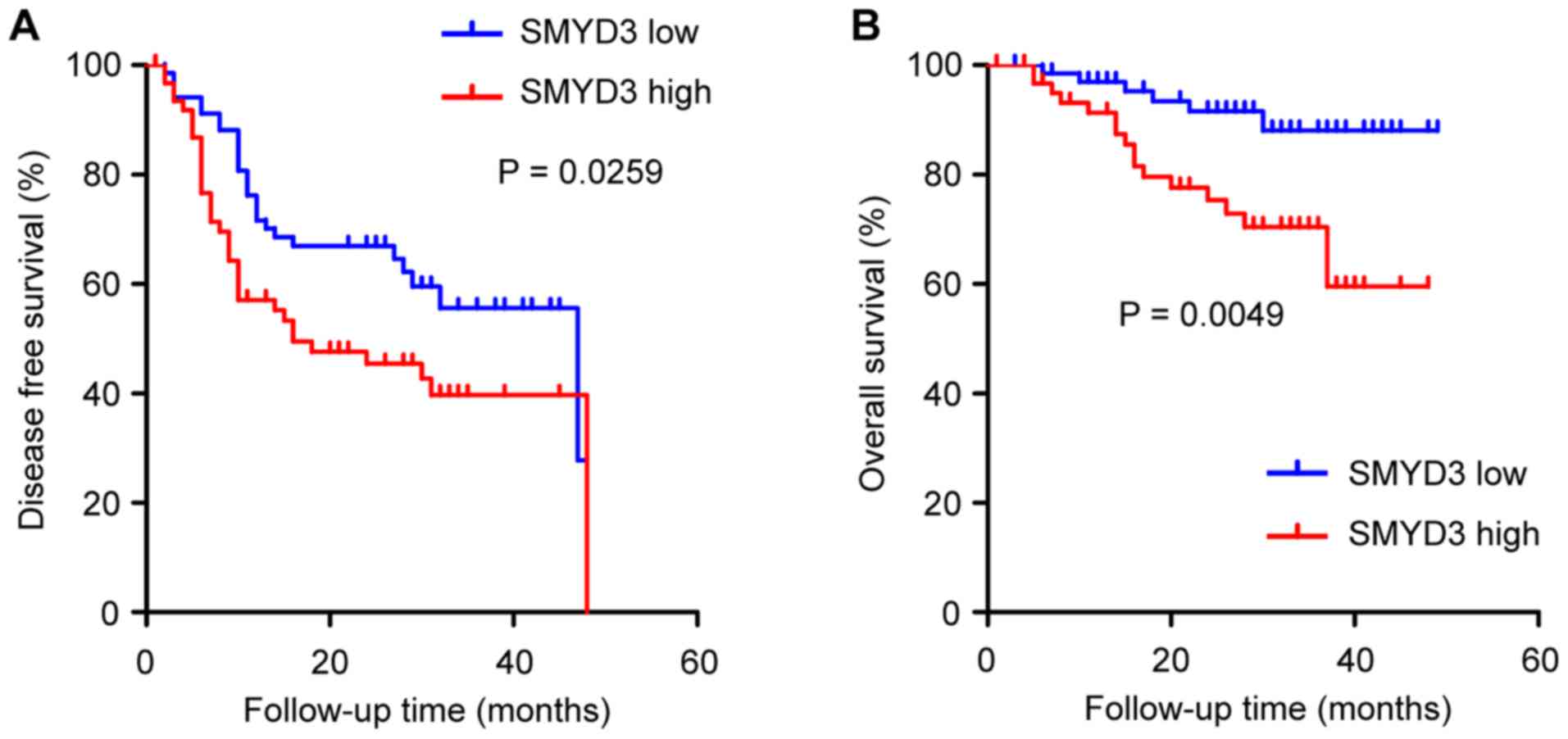
The prognostic significance of SMYD3 in NSCLC was
analyzed. One hundred and thirty-one NSCLC patients qualified for
survival analysis, due to sufficient follow-up. It was revealed
that disease-free survival (DFS) and overall survival (OS) were
significantly worse in patients with a high than in patients with a
low SMYD3 expression (Fig. 2A and
B).
It was also determined whether SMYD3 has predictive
value for clinical outcomes in NSCLC patients. Univariate Cox
regression analysis revealed that age at diagnosis (>60 years),
advanced pathological stage and PI were correlated with poor DFS in
NSCLC patients, and that age at diagnosis (>60 years), advanced
pathological stage and high SMYD3 expression were correlated with
poor OS (Tables III and IV). Furthermore, multivariate Cox
regression analysis demonstrated that advanced pathological stage
and PI were independent predictors for poor DFS, and that age at
diagnosis (>60 year), advanced pathological stage and high SMYD3
expression were independent predictors for poor OS (Table III and IV).
| Table IIIUnivariate and multivariate Cox
regression analysis for DFS in NSCLC. |
Table III
Univariate and multivariate Cox
regression analysis for DFS in NSCLC.
| Variables | Univariate analysis
| Multivariate
analysis
|
|---|
| HR | 95% CI | P-value | HR | 95% CI | P-value |
|---|
| Sex | | | | | | |
| Male | 1 | | | | | |
| Female | 1.586 | 0.665-3.78 | 0.298 | | | |
| Age (years) | | | | | | |
| ≤60 | 1 | | | 1 | | |
| >60 | 1.819 | 1.022-3.24 | 0.042 | 1.712 | 0.998-2.938 | 0.051 |
| Smoking
history | | | | | | |
| Never | | | | | | |
| Ever | 1.689 | 0.72-3.96 | 0.228 | | | |
| Stage | | | | | | |
| I | 1 | | | 1 | | |
| II-IV | 4.912 | 2.612-9.237 | <0.001 | 5.342 | 2.924-9.759 | <0.001 |
| Tumor size
(cm) | | | | | | |
| ≤3.0 | 1 | | | | | |
| >3.0 | 1.202 | 0.695-2.076 | 0.511 | | | |
| LVI | | | | | | |
| Absent | 1 | | | | | |
| Present | 1.221 | 0.639-2.331 | 0.545 | | | |
| PI | | | | | | |
| Absent | | | | 1 | | |
| Present | 1.812 | 1.054-3.114 | 0.031 | 1.927 | 1.142-3.251 | 0.014 |
|
Differentiation | | | | | | |
| Well/moderate | 1 | | | | | |
| Poor | 0.919 | 0.492-1.717 | 0.792 | | | |
| SMYD3 | | | | | | |
| Low | 1 | | | | | |
| High | 1.462 | 0.788-2.714 | 0.228 | | | |
| Table IVUnivariate and multivariate Cox
regression analysis for OS in NSCLC. |
Table IV
Univariate and multivariate Cox
regression analysis for OS in NSCLC.
| Variables | Univariate analysis
| Multivariate
analysis
|
|---|
| HR | 95% CI | P-value | HR | 95% CI | P-value |
|---|
| Sex | | | | | | |
| Male | 1 | | | | | |
| Female | 1.559 | 0.374-6.498 | 0.542 | | | |
| Age (years) | | | | | | |
| ≤60 | 1 | | | 1 | | |
| >60 | 5.321 | 1.651-17.142 | 0.005 | 4.474 | 1.485-13.479 | 0.008 |
| Smoking
history | | | | | | |
| Never | 1 | | | | | |
| Ever | 1.719 | 0.4-7.391 | 0.467 | | | |
| Stage | | | | | | |
| I | 1 | | | 1 | | |
| II-IV | 3.246 | 1.19-8.852 | 0.021 | 3.129 | 1.244-7.871 | 0.015 |
| Tumor size
(cm) | | | | | | |
| ≤3.0 | 1 | | | | | |
| >3.0 | 0.646 | 0.26-1.603 | 0.346 | | | |
| LVI | | | | | | |
| Absent | 1 | | | | | |
| Present | 0.864 | 0.302-2.47 | 0.785 | | | |
| PI | | | | | | |
| Absent | 1 | | | | | |
| Present | 1.369 | 0.548-3.421 | 0.501 | | | |
|
Differentiation | | | | | | |
| Well/moderate | 1 | | | | | |
| Poor | 1.581 | 0.561-4.457 | 0.387 | | | |
| SMYD3 | | | | | | |
| Low | 1 | | | 1 | | |
| High | 4.092 | 1.302-12.864 | 0.016 | 3.338 | 1.283-8.684 | 0.013 |
In combination, the present findings indicated that
SMYD3 was upregulated in NSCLC and associated with aggressive
clinicopathological characteristics. SMYD3 can serve as an adverse
prognostic marker for NSCLC, and may be involved in NSCLC
development and progression.
SMYD3 promotes the proliferation of NSCLC
cells in vitro
Next, the potential biological role of SMYD3 in
NSCLC was investigated. The expression levels of SMYD3 protein in
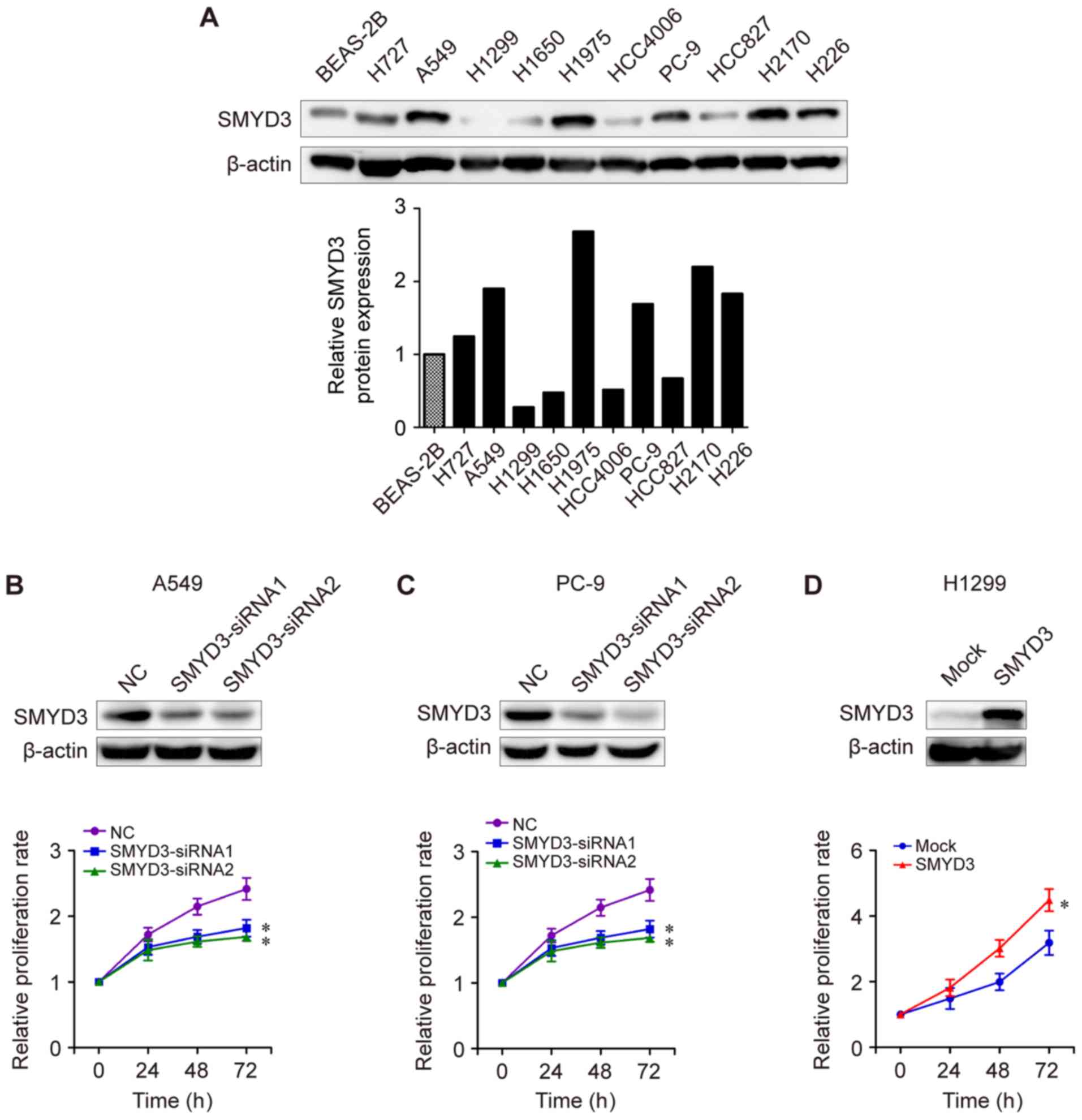
NSCLC cell lines were examined by western blot analysis. As
presented in Fig. 3A, SMYD3
protein expression was higher in most NSCLC cell lines (H727, A549,
H1975, PC-9, H2170 and H226) compared with that in human bronchial
epithelial BEAS-2B cells. H727, A549, H1975, PC-9, H2170 and H226
cells exhibited a higher SMYD3 expression than that of H1299,
H1650, HCC4006 or HCC827 cells (Fig.
3A). Next, SMYD3-siRNA transfections were performed in
endogenous high-SMYD3-expressing cells. It was determined that
SMYD3 knockdown led to a significant reduction in the proliferation
rate of A549 and PC-9 cells in vitro (Fig. 3B and C). By contrast, enforced
SMYD3 in endogenous low-SMYD3-expressing cells H1299 significantly
promoted cell proliferation compared to a mock group (Fig. 3D), indicating an important role of
SMYD3 in NSCLC cell proliferation.
SMYD3 knockdown triggers apoptosis by
affecting pro-apoptotic and anti-apoptotic proteins in NSCLC
cells
Apoptosis is considered an important approach in the
inhibition of cell proliferation. In order to elucidate the
mechanism underlying SMYD3 knockdown-induced proliferation
reduction in NSCLC cells, the apoptotic rate of
SMYD3-siRNA-transfected A549 and PC-9 cells was examined. Flow
cytometric data revealed that SMYD3 silencing significantly induced
apoptosis in both A549 and PC-9 cells (Fig. 4A-C).
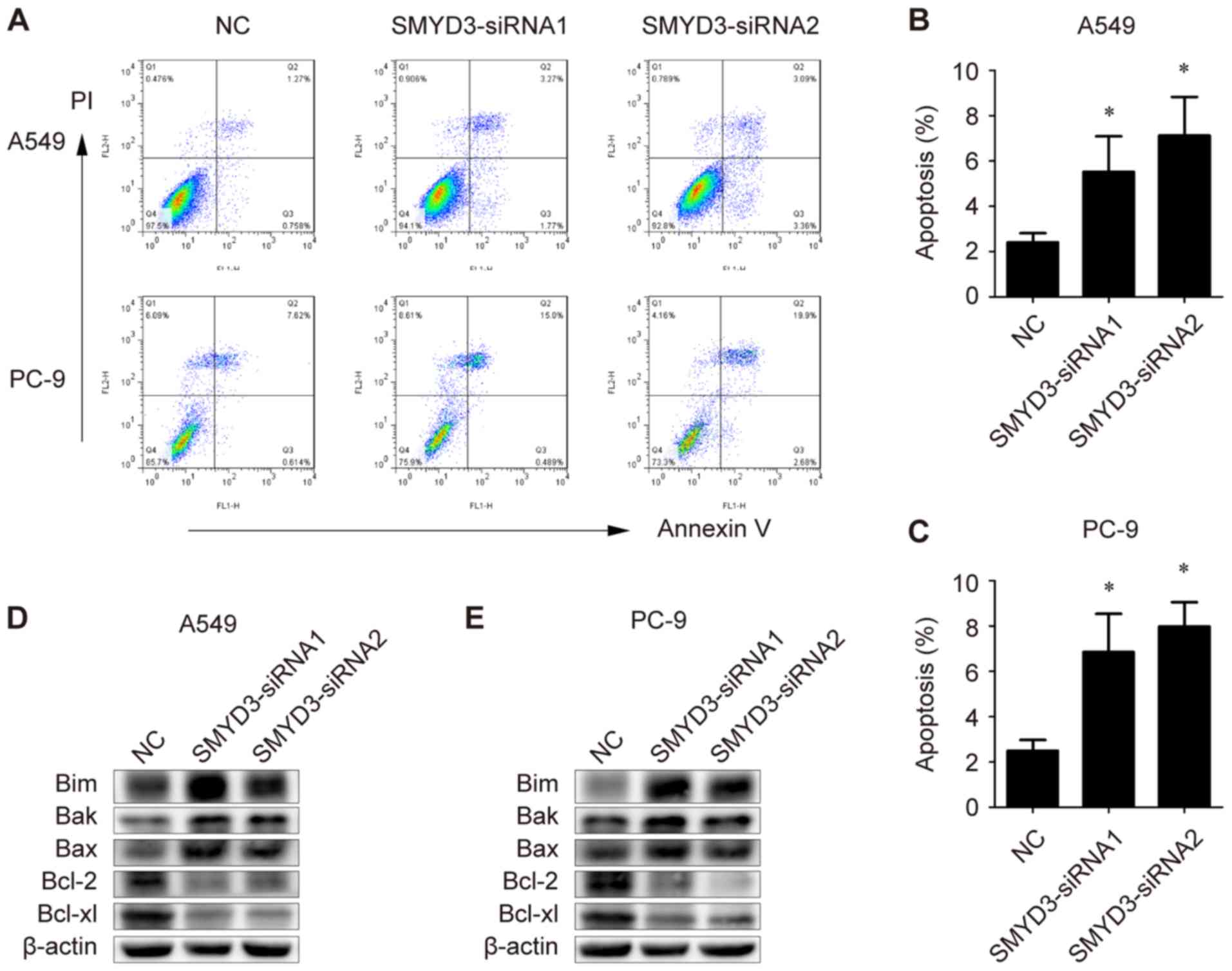 | Figure 4Effects of SMYD3 knockdown on
apoptosis and apoptosis-associated genes in NSCLC cells. A549 and
PC-9 cells were transfected with NC- or SMYD3-siRNAs. (A) The
apoptotic rate was measured using Annexin V-FITC/PI double staining
method in a flow cytometer 72 h after transfection. (B and C)
Statistical analysis of the apoptotic population in the transfected
(B) A549 and (C) PC-9 cells. (D and E) Expression of Bim, Bak, Bax,
Bcl-2 and Bcl-xl in the transfected (D) A549 and (E) PC-9 cells
were examined by western blotting. Data are presented as the mean ±
SD of at least 3 independent experiments. *P<0.05,
compared with the NC group. SMYD3, SET and MYND domain-containing
protein 3; NSCLC, non-small cell lung cancer; NC, negative control;
Bim, Bcl-2-like protein 11; Bak, Bcl-2 homologous
antagonist/killer; Bcl-2, B-cell lymphoma 2. |
To further explore the molecular basis for the
induced apoptosis, the expression of pro-apoptotic and
anti-apoptotic genes was evaluated. As revealed, the levels of Bim,
Bak and Bax were markedly increased in SMYD3-silenced A549 and PC-9
cells, while Bcl-2 and Bcl-xl proteins were decreased upon
SMYD3-siRNA transfection in A549 and PC-9 cells (Fig. 4D and E). Collectively, these
results indicated that SMYD3 knockdown triggered apoptosis in NSCLC
cells, probably by affecting key pro- and anti-apoptotic
proteins.
Effect of SMYD3 on cisplatin resistance
in NSCLC cells
Since the dysregulated apoptotic signals can result
in resistance to anticancer drugs in cancer cells (22), it was speculated that SMYD3 may be
involved in cisplatin resistance, a chemotherapy drug for advanced
NSCLC (23). As revealed,
cisplatin induced apoptosis in A549, PC-9 and H1299 cells in a
concentration-dependent manner (Fig.
5A-C). Specifically, SMYD3 silencing sensitized A549 and PC-9
cells to the apoptosis induced by cisplatin (Fig. 5A and B), whereas stable SMYD3
overexpression rendered H1299 cells more resistant to
cisplatin-induced apoptosis (Fig.
5C). These data indicated that SMYD3 was associated with
resistance to cisplatin in NSCLC cells.
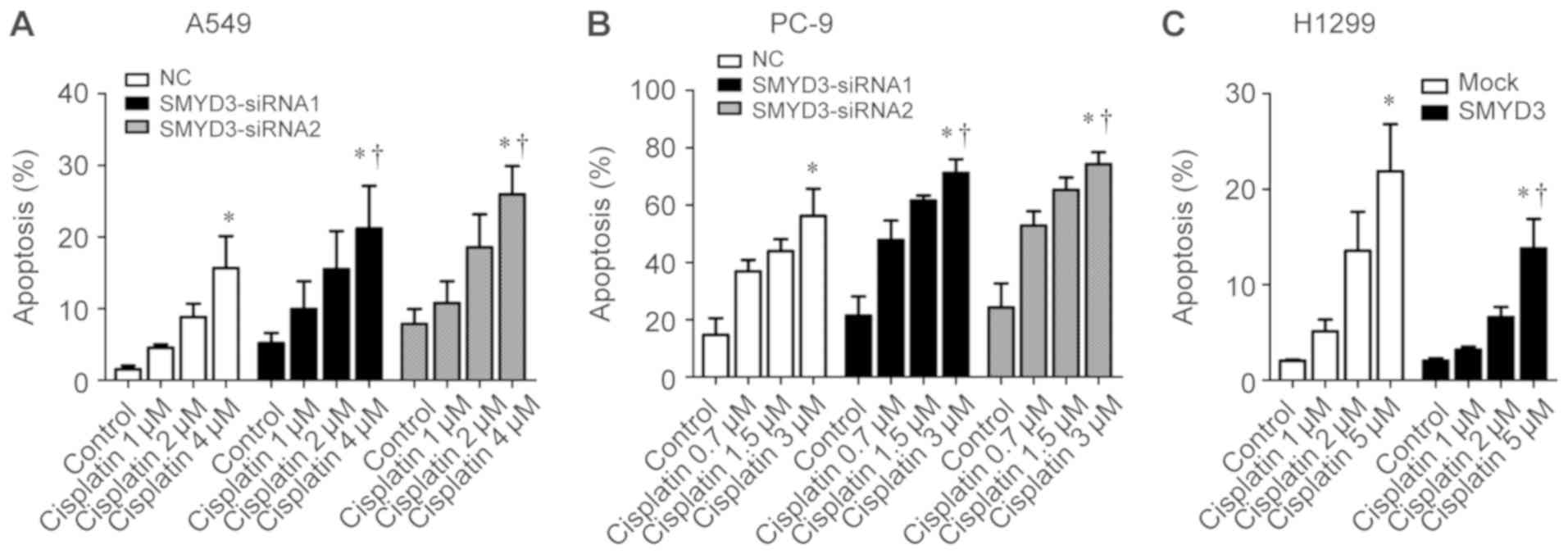 | Figure 5Effect of SMYD3 on cisplatin
resistance in NSCLC cells. (A) A549 and (B) PC-9 cells were
transfected with NC- or SMYD3-siRNAs for 24 h, and then treated
with increasing concentrations of cisplatin (1, 2, and 4 µM
for A549; 0.7, 1.5, and 3 µM for PC-9) for 72 h. (C) H1299
stable cell lines were treated with increasing concentrations of
cisplatin (1, 2, and 5 µM) for 72 h. The apoptotic rate was
measured using Annexin V-FITC/PI double staining method. Data are
presented as the mean ± SD of at least 3 independent experiments.
*P<0.05, compared with the control group.
†P<0.05, compared with the NC or mock group. SMYD3,
SET and MYND domain-containing protein 3; NSCLC, non-small cell
lung cancer; NC, negative control. |
SMYD3 knockdown blocks NSCLC cell
migration and invasion in vitro
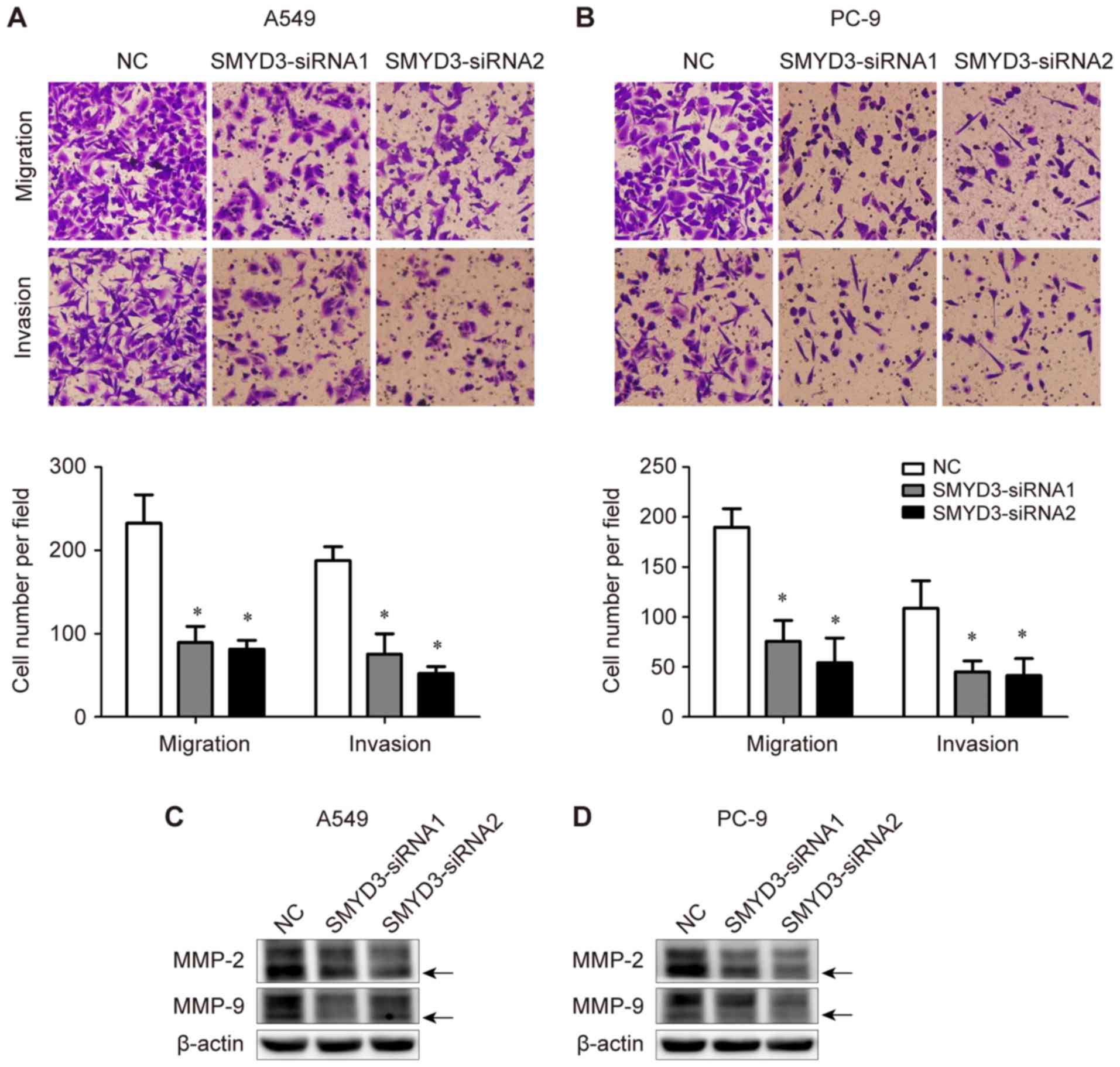
The effects of SMYD3 on NSCLC cell migration and
invasion were determined using Transwell assays. It was determined
that SMYD3 knockdown significantly suppressed the migratory and
invasive abilities of both A549 and PC-9 cells (Fig. 6A and B). These data indicated the
critical role of SMYD3 in the migration and invasion of NSCLC
cells.
In addition, the expression of invasion-associated
MMP-2 and MMP-9 was assessed in the same transfection experiments.
It was observed that the expression levels of both pro- and
active-form MMP-2 and MMP-9 were down-regulated upon SMYD3
silencing in A549 and PC-9 cells (Fig.
6C and D).
Discussion
SMYD3 is a lysine methyltransferase that directs the
fine regulation of chromatin structure and gene expression
(4,24). Recently, it has been revealed that
SMYD3 plays significant roles in human carcinogenesis and tumor
progression, including in breast, hepatocellular, colorectal and
gastric carcinomas (7,8,10).
In the present study, it was demonstrated that SMYD3 was
upregulated in NSCLC tissues, and associated with aggressive
clinicopathological characteristics and poor survival. Multivariate
analysis revealed that SMYD3 served as an independent predictor of
poor OS in NSCLC patients. It was also revealed that SMYD3 played a
critical role in NSCLC cell proliferation, migration, invasion and
cisplatin resistance, as well as the regulation of associated
genes. The present study is the first, to the best of our
knowledge, to comprehensively analyze the expression of SMYD3 in
clinical NSCLC samples, and offer insights into the oncogenic
potential of SMYD3 in NSCLC carcinogenesis.
Barlesi et al have reported that the
SMYD3 VNTR genotype was not correlated with either
clinicopathological features or the risk of NSCLC in a Caucasian
population (16). However, in the
same cohort of NSCLC, there was no correlation between the proposed
VNTR genotype and the level of H3K4 dimethylation either, implying
that the SMYD3 VNTR genotype may not be equal to the
expression or activity of SMYD3 protein in NSCLC. Herein, it was
determined that SMYD3 protein was significantly upregulated in
NSCLC tissues when compared with paired adjacent normal tissues.
Notably, a high SMYD3 expression was associated with aggressive
clinicopathological features, such as more advanced pathological
stage, larger tumor size and distant metastasis, as well as poor
DFS and OS in NSCLC patients. Multivariate Cox regression analysis
revealed that SMYD3 overexpression was an independent predictor for
poor OS, indicating that SMYD3 may be involved in NSCLC development
and progression. In line with our findings, Mazur et al have
revealed that abrogating SMYD3 catalytic activity inhibited the
formation of Ras-driven lung adenocarcinomas in mouse models
(25).
It was previously reported that SMYD3 is essential
for the proliferation of cervical, breast, esophageal,
hepatocellular and colorectal cancer cells (7,8,14,20).
Consistently, the present data revealed that SMYD3 knockdown by
siRNA led to a decreased proliferation rate in NSCLC cells in
vitro, whereas stable SMYD3 overexpression significantly
promoted NSCLC cell proliferation. These data confirmed our
hypothesis that SMYD3 behaves as an oncogene in NSCLC
carcinogenesis. Given the fact that apoptosis induction is a
pivotal mechanism underlying the inhibition of cancer cell
proliferation, the apoptosis of SMYD3-silenced NSCLC cells was
examined, and it was revealed that SMYD3 silencing significantly
induced apoptosis in NSCLC cells. The Bcl-2 protein family members,
which can be divided into two classes (pro-apoptotic and
anti-apoptotic), have been acknowledged as key regulators of
caspase activation and apoptosis (26). Further experiments were then
performed to explore the molecular basis for the induced apoptosis
in SMYD3-silenced NSCLC cells. It was determined that the levels of
pro-apoptotic Bim, Bak and Bax were markedly increased in
SMYD3-silenced NSCLC cells, while anti-apoptotic Bcl-2 and Bcl-xl
were decreased by SMYD3-siRNA. Our study extends the knowledge
derived from Ren et al, who reported that the downregulation
of SMYD3 decreased the ratio of Bcl-2/Bax in MDA-MB-231 cells
(27), indicating that SMYD3 has a
broad effect on apoptosis-associated genes.
Defective or inefficient apoptosis has been
considered as a critical mechanism of resistance to anticancer
drugs in cancer cells (22). It
was then determined whether SMYD3 is involved in resistance to
cisplatin, which remains a standard chemotherapy drug for advanced
NSCLC (23). The present data
revealed that SMYD3 silencing sensitized NSCLC cells to the
apoptosis induced by cisplatin, whereas stable SMYD3 overexpression
rendered NSCLC cells more resistant to cisplatin-induced apoptosis.
This is probably attributed to the change of apoptotic signals in
SMYD3-silenced or -over-expressed NSCLC cells. Consistently,
previous studies have indicated that cisplatin resistance can be
mediated by an inhibition of the apoptotic machinery derived from
intracellular events (HER-2/neu amplification, or p53
mutation) in human malignancies (28,29).
In the present study, it was observed that SMYD3
knockdown significantly suppressed NSCLC cell migration and
invasion in vitro, highlighting the oncogenic role of SMYD3
in NSCLC progression. Similarly, recent studies revealed that SMYD3
promoted the migration and invasion of multiple cancer types,
including hepatocellular, breast and gastric carcinomas (30-32).
MMPs are zinc-dependent endopeptidases that participate in tumor
progression and metastasis by degrading the extracellular matrix,
as well as regulating adherin, cytoskeletal proteins, growth
factors, chemokines and cytokines (33). To date, 24 MMPs have been
recognized in human beings. In the present study, it was determined
that both pro- and active-form MMP-2 and MMP-9 proteins were
decreased upon SMYD3 silencing in NSCLC cells, which probably
contributed to the impaired invasive activities in those cells. The
present results are supported by a previous study, which
demonstrated that SMYD3 promoted the trimethylation of H3K4 in the
MMP-9 promoter in a reversible model of cancer, thereby inducing
MMP-9 expression and increasing cell migration and invasion
(6). Based on the present data, it
was reasoned that other MMPs, except for MMP-2 and MMP-9, may also
be regulated by SMYD3 and involved in SMYD3-mediated cancer
migration and invasion, although further studies are required to
establish this hypothesis.
The present study has some limitations. First, we
were unable to reveal the detailed mechanisms of SMYD3-modulated
apoptosis- or invasion-associated genes in NSCLC cells. In
addition, another limitation is that although SMYD3 has been
recognized to regulate multiple downstream target genes, which are
implicated in cell adhesion, proliferation, invasion, apoptosis,
cell cycle regulation and signal transduction (5,34,35),
only the effects of SMYD3 on NSCLC cell proliferation, apoptosis,
migration and invasion were evaluated in the present study. More
studies are required to elucidate the effects of other functions of
SMYD3 in NSCLC development and progression.
In conclusion, it was demonstrated that SMYD3 was
upregulated in NSCLC tissues, and associated with aggressive
clinicopathological characteristics and poor survival. SMYD3
overexpression independently predicted poor OS in NSCLC patients.
Functionally, SMYD3 played a critical role in NSCLC cell
proliferation, migration, invasion and cisplatin resistance, as
well as the regulation of apoptosis- and invasion-associated genes.
The present study suggested SMYD3 as a novel oncogene in NSCLC
carcinogenesis, which is anticipated to be a potential prognostic
marker and therapeutic target for NSCLC patients.
Abbreviations:
|
NSCLC
|
non-small cell lung cancer
|
|
SMYD3
|
SET and MYND domain-containing protein
3
|
|
H3K4
|
histone H3 at lysine 4
|
|
VNTR
|
variable number of tandem repeats
|
|
IHC
|
immunohistochemistry
|
|
TMA
|
tissue microarray
|
|
IASLC
|
International Association for the
Study of Lung Cancer
|
|
ATCC
|
American Type Culture Collection
|
|
FBS
|
fetal bovine serum
|
|
SRB
|
sulforhodamine B
|
|
NC
|
negative control
|
|
MMP
|
matrix metalloproteinase
|
|
LVI
|
lymphovascular invasion
|
|
PI
|
pleural invasion
|
|
DFS
|
disease-free survival
|
|
OS
|
overall survival
|
Acknowledgments
Not applicable.
Funding
The present study was supported in part by grant
nos. 81902325 and 81903633 from the National Natural Science
Foundation of China, grant no. 2017WS026 from the Health and Family
Planning Commission of Shandong Province (China), and grant nos.
2019GSF107042 and 2019GSF107051 from the Key Research and
Development Plan of Shandong Province (P.R. China).
Availability of data and materials
All data generated or analyzed in this study are
available from the corresponding author on reasonable request.
Authors' contributions
WY devised the conceptual idea. JL, YP and XM
acquired and analyzed the clinical data. JL, LZ and LL performed
the in vitro experiments. JL, LZ and WW analyzed the data
and prepared the figures. JL, LZ and WY wrote, reviewed, and
revised the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Fudan University Shanghai Cancer Center. All
experiments complied with the Helsinki declaration and the approved
guidelines of our institution. Written informed consent was
obtained from all participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zheng M: Classification and Pathology of
Lung Cancer. Surg Oncol Clin N Am. 25:447–468. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rosell R and Karachaliou N: Lung cancer:
Maintenance therapy and precision medicine in NSCLC. Nat Rev Clin
Oncol. 10:549–550. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Spellmon N, Holcomb J, Trescott L,
Sirinupong N and Yang Z: Structure and function of SET and MYND
domain-containing proteins. Int J Mol Sci. 16:1406–1428. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hamamoto R, Furukawa Y, Morita M, Iimura
Y, Silva FP, Li M, Yagyu R and Nakamura Y: SMYD3 encodes a histone
methyl-transferase involved in the proliferation of cancer cells.
Nat Cell Biol. 6:731–740. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cock-Rada AM, Medjkane S, Janski N, Yousfi
N, Perichon M, Chaussepied M, Chluba J, Langsley G and Weitzman JB:
SMYD3 promotes cancer invasion by epigenetic upregulation of the
metalloproteinase MMP-9. Cancer Res. 72:810–820. 2012. View Article : Google Scholar :
|
|
7
|
Hamamoto R, Silva FP, Tsuge M, Nishidate
T, Katagiri T, Nakamura Y and Furukawa Y: Enhanced SMYD3 expression
is essential for the growth of breast cancer cells. Cancer Sci.
97:113–118. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sarris ME, Moulos P, Haroniti A,
Giakountis A and Talianidis I: Smyd3 Is a transcriptional
potentiator of multiple cancer-promoting genes and required for
liver and colon cancer development. Cancer Cell. 29:354–366. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhu Y, Zhu MX, Zhang XD, Xu XE, Wu ZY,
Liao LD, Li LY, Xie YM, Wu JY, Zou HY, et al: SMYD3 stimulates EZR
and LOXL2 transcription to enhance proliferation, migration, and
invasion in esophageal squamous cell carcinoma. Hum Pathol.
52:153–163. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu Y, Deng J, Luo X, Pan Y, Zhang L,
Zhang R and Liang H: Overexpression of SMYD3 was associated with
increased STAT3 activation in gastric cancer. Med Oncol.
32:4042015. View Article : Google Scholar
|
|
11
|
Shen B, Tan M, Mu X, Qin Y, Zhang F, Liu Y
and Fan Y: Upregulated SMYD3 promotes bladder cancer progression by
targeting BCLAF1 and activating autophagy. Tumour Biol.
37:7371–7381. 2016. View Article : Google Scholar
|
|
12
|
Liu C, Wang C, Wang K, Liu L, Shen Q, Yan
K, Sun X, Chen J, Liu J, Ren H, et al: SMYD3 as an oncogenic driver
in prostate cancer by stimulation of androgen receptor
transcription. J Natl Cancer Inst. 105:1719–1728. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dai B, Wan W, Zhang P, Zhang Y, Pan C,
Meng G, Xiao X, Wu Z, Jia W, Zhang J, et al: SET and MYND
domain-containing protein 3 is overexpressed in human glioma and
contributes to tumorigenicity. Oncol Rep. 34:2722–2730. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang SZ, Luo XG, Shen J, Zou JN, Lu YH and
Xi T: Knockdown of SMYD3 by RNA interference inhibits cervical
carcinoma cell growth and invasion in vitro. BMB Rep. 41:294–299.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tsuge M, Hamamoto R, Silva FP, Ohnishi Y,
Chayama K, Kamatani N, Furukawa Y and Nakamura Y: A variable number
of tandem repeats polymorphism in an E2F-1 binding element in the
5′ flanking region of SMYD3 is a risk factor for human cancers. Nat
Genet. 37:1104–1107. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Barlési F, Giaccone G, Gallegos-Ruiz MI,
Span SW, Lefesvre P, Kruyt FA and Rodriguez JA: Genotype analysis
of the VNTR polymorphism in the SMYD3 histone methyltransferase
gene: Lack of correlation with the level of histone H3 methylation
in NSCLC tissues or with the risk of NSCLC. Int J Cancer.
122:1441–1442. 2008. View Article : Google Scholar
|
|
17
|
Fedor HL and De Marzo AM: Practical
methods for tissue microarray construction. Methods Mol Med.
103:89–101. 2005.
|
|
18
|
Detterbeck FC, Boffa DJ and Tanoue LT: The
new lung cancer staging system. Chest. 136:260–271. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
You W, Chen B, Liu X, Xue S, Qin H and
Jiang H: Farnesoid X receptor, a novel proto-oncogene in non-small
cell lung cancer, promotes tumor growth via directly
transactivating CCND1. Sci Rep. 7:5912017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dong SW, Zhang H, Wang BL, Sun P, Wang YG
and Zhang P: Effect of the downregulation of SMYD3 expression by
RNAi on RIZ1 expression and proliferation of esophageal squamous
cell carcinoma. Oncol Rep. 32:1064–1070. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kunizaki M, Hamamoto R, Silva FP,
Yamaguchi K, Nagayasu T, Shibuya M, Nakamura Y and Furukawa Y: The
lysine 831 of vascular endothelial growth factor receptor 1 is a
novel target of methylation by SMYD3. Cancer Res. 67:10759–10765.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Indran IR, Tufo G, Pervaiz S and Brenner
C: Recent advances in apoptosis, mitochondria and drug resistance
in cancer cells. Biochim Biophys Acta. 1807:735–745. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rossi A and Di Maio M: Platinum-based
chemotherapy in advanced non-small-cell lung cancer: Optimal number
of treatment cycles. Expert Rev Anticancer Ther. 16:653–660. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Giakountis A, Moulos P, Sarris ME, Hatzis
P and Talianidis I: Smyd3-associated regulatory pathways in cancer.
Semin Cancer Biol. 42:70–80. 2017. View Article : Google Scholar
|
|
25
|
Mazur PK, Reynoird N, Khatri P, Jansen PW,
Wilkinson AW, Liu S, Barbash O, Van Aller GS, Huddleston M, Dhanak
D, et al: SMYD3 links lysine methylation of MAP3K2 to Ras-driven
cancer. Nature. 510:283–287. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cory S and Adams JM: The Bcl2 family:
Regulators of the cellular life-or-death switch. Nat Rev Cancer.
2:647–656. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ren TN, Wang JS, He YM, Xu CL, Wang SZ and
Xi T: Effects of SMYD3 over-expression on cell cycle acceleration
and cell proliferation in MDA-MB-231 human breast cancer cells. Med
Oncol. 28(Suppl 1): S91–S98. 2011. View Article : Google Scholar
|
|
28
|
Zhou BP, Liao Y, Xia W, Zou Y, Spohn B and
Hung MC: HER-2/neu induces p53 ubiquitination via Akt-mediated MDM2
phosphorylation. Nat Cell Biol. 3:973–982. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Perego P, Giarola M, Righetti SC, Supino
R, Caserini C, Delia D, Pierotti MA, Miyashita T, Reed JC and
Zunino F: Association between cisplatin resistance and mutation of
p53 gene and reduced bax expression in ovarian carcinoma cell
systems. Cancer Res. 56:556–562. 1996.PubMed/NCBI
|
|
30
|
Zhou Z, Jiang H, Tu K, Yu W, Zhang J, Hu
Z, Zhang H, Hao D, Huang P, Wang J, et al: ANKHD1 is required for
SMYD3 to promote tumor metastasis in hepatocellular carcinoma. J
Exp Clin Cancer Res. 38:182019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Luo XG, Zhang CL, Zhao WW, Liu ZP, Liu L,
Mu A, Guo S, Wang N, Zhou H and Zhang TC: Histone methyltransferase
SMYD3 promotes MRTF-A-mediated transactivation of MYL9 and
migration of MCF-7 breast cancer cells. Cancer Lett. 344:129–137.
2014. View Article : Google Scholar
|
|
32
|
Zou JN, Wang SZ, Yang JS, Luo XG, Xie JH
and Xi T: Knockdown of SMYD3 by RNA interference down-regulates
c-Met expression and inhibits cells migration and invasion induced
by HGF. Cancer Lett. 280:78–85. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gonzalez-Avila G, Sommer B, Mendoza-Posada
DA, Ramos C, Garcia-Hernandez AA and Falfan-Valencia R: Matrix
metal-loproteinases participation in the metastatic process and
their diagnostic and therapeutic applications in cancer. Crit Rev
Oncol Hematol. 137:57–83. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sims RJ III and Reinberg D: From chromatin
to cancer: A new histone lysine methyltransferase enters the mix.
Nat Cell Biol. 6:685–687. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Huang L and Xu AM: SET and MYND domain
containing protein 3 in cancer. Am J Transl Res. 9:1–14.
2017.PubMed/NCBI
|