Introduction
Pancreatic cancer is one of the leading causes of
cancer-related mortality, with a 5-year survival rate of 8%. This
disease is expected to surpass breast malignancy as the third
leading cause of cancer-related mortality in the USA (1,2).
Routine examinations of the pancreas are not readily performed and
consequently, the low survival rate noted in patients with
pancreatic cancer is largely due to the advanced stage of disease
at diagnosis (3). To date,
gemcitabine has been used as the standard treatment for pancreatic
cancer, extending patient survival by only a few months with
several adverse reactions, such as atrophy of hematopoietic organs
and immune suppression (4).
Therefore, the development of novel effective strategies to control
pancreatic cancer is urgently required.
Cancer cells always reprogram their metabolism and
enhance glycolysis to meet the constantly growing requirement for
intermediates, even in the presence of abundant oxygen, which is
known as the Warburg effect (5,6).
Recently, studies have indicated that the activation of the
PI3K/AKT signaling pathway can promote pancreatic cancer
proliferation and that the expression of several genes involved in
glycolysis can be upregulated by oncogenes, such as PI3K/Akt, NF-κB
and c-myc (7,8). Enzymes, such as glucose transporter-1
(GLUT-1), lactate dehydrogenase A (LDHA) and monocarboxylic acid
transporter-4, which are highly expressed in tumors and are
associated with a poor prognosis, are involved in the glycolytic
process (9). There is also
accumulating evidence to suggest that the inhibition of key
enzymes, such as GLUT-1 and LDHA, can effectively delay tumor
progression (10,11).
Based on known pharmacokinetic and safety profiles,
the existing clinical drugs can be further screened to identify
potential new indications that can significantly reduce the cost
and time of drug exploration. This method is considered one of the
most rapid and effective strategies in the field of drug discovery.
Certain agents can alter glucose metabolism or inhibit the
expression of glucose transporters to suppress tumor growth
(12). Canagliflozin (CANA) is an
important oral medication used for diabetes that acts by inhibiting
the activity of sodium-glucose cotransporter 2 (13). Recent studies have demonstrated
that CANA may suppress liver cancer progression, whereas the
effects of CANA against pancreatic cancer have not been fully
investigated (14,15).
In the present study, pancreatic cancer cells and a
PANC-1-derived xenograft tumor model were used to assess the
antitumor activity of CANA. The results suggested that CANA was
capable of suppressing pancreatic cancer growth through the
inhibition of glycolysis mediated primarily via the PI3K/AKT/mTOR
signaling pathway. These findings indicate its potential use in the
clinical treatment of pancreatic cancer.
Materials and methods
Cell culture and reagents
The two human pancreatic cancer cell lines, Capan-1
and PANC-1 (American Type Culture Collection), were cultured in
Dulbecco's modified Eagle's medium with high glucose (Gibco; Thermo
Fisher Scientific, Inc.) supplemented with 10% FBS, 100 U/ml
penicillin and 100 µg/ml streptomycin (Beyotime Institute of
Biotechnology) in a humidified incubator containing 5%
CO2 at 37°C. CANA (Meilun Biotechnology), gemcitabine
(Luoxin Biotechnology), LY294002 (MedChem Express) were dissolved
in DMSO or physiological saline for use in the in vitro
experiments.
Cell viability assay
The viability of the Capan-1 and PANC-1 cells was
assessed by MTT assay (Sigma-Aldrich; Merck KGaA). Briefly, the
cells were seeded in a 96-well plate at a density of 5,000
cells/well, incubated for 24 h and treated with 20, 40, 60 and 80
µM CANA, or 0.5 µM gemcitabine for 48 h. The number
of viable cells was quantified by measuring the OD at 490 nm with a
microplate reader (AC100; Meigu Molecular Instruments Co., Ltd.)
and the results were expressed as a percentage (%) of the control.
The IC50 values were calculated using GraphPad Prism 4.0 software
(GraphPad Software).
Colony formation assay
The Capan-1 and PANC-1 cells were seeded in 6-well
plates at a density of 500 cells/well in 2 ml culture medium.
Following 72 h of incubation (37°C, 5% CO2), the cells
were treated with 20, 40 and 60 µM CANA, or 0.03 µM
22 gemcitabine, or treated with the combination of 20 µM
CANA and 0.03 µM gemcitabine for 10 days (37°C, 5%
CO2). The cells were then rinsed with 1X PBS, fixed with
chilled at room temperature for 10 min and stained with 0.5%
crystal violet (Beijing Solarbio Science & Technology Co.,
Ltd.) at 25°C for 40 min. Cells morphology was observed and
photographed under a light microscope (Nicon Corporation). The
experiments were independently performed in triplicate.
Flow cytometric analysis
The ontrol and treated cells were stained with an
Annexin V/propidium iodide staining kit (Vazyme) to assess cell
death according to the manufacturer's instructions. The stained
cells were captured by flow cytometry (BD Biosciences) and 10,000
events were collected. All assays were repeated at least 3
times.
Glucose uptake and lactate release
assay
The DMEM culture medium contains high glucose, which
is taken up by cells to provide energy and produce lactate. The
supernatants from the control and treated cell cultures (40 and 60
µM CANA) were collected at 6, 12 and 24 h. The glucose
uptake rate was measured using a glucose assay kit (Jiancheng)
according to the manufacturer's instructions. Lactate release was
deter-mined using a lactate assay kit (Beijing Solarbio Science
& Technology Co., Ltd.).
In vivo antitumor activity assay
The PANC-1 cells (100 µl, 6×106
cells/ml) were suspended in saline and injected into the underarms
of 4-week-old male nude mice (weigh 18-20 g, n=25 in total)
(Balb/c, Yangzhou University, Jiangsu, China). The mice were kept
in an environment with a temperature of 22±1°C/40% relative
humidity and light exposure 12 h/day. They were provided with
sufficient food and water. All animal experiments were performed
according to the protocols approved by the Ethics Committee of
China Pharmaceutical University. After 1 week, the xenograft mice
were randomly divided into 5 groups according to the tumor volume.
Each group included 5 mice, with 25 mice in total. Mice in group 1
received the vehicle (physiological saline, 10 ml/kg), those in
group 2 were treated with an intraperitoneal injection of
gemcitabine at 10 mg/kg twice a week, and those in groups 3, 4 and
5 were orally administered CANA at doses of 25, 50 and 100 mg/kg
every day for 5 weeks. Gemcitabine was dissolved in normal saline
and CANA was resuspended in CMC-Na solution. All mice involved in
the experiment were euthanized by cervical dislocation following
the final administration of the drugs and the sizes of the
collected tumors were recorded. The tumor tissues were stored in a
refrigerator at -80°C. For hematoxylin and eosin (H&E)
staining, a standard staining procedure was performed. In brief,
tumor tissues were fixed with 10% formalin at room temperature for
24 h, 6-µm-thick slices were prepared. Subsequently,
hematoxylin staining was lasted for 3 min, as well as eosin
staining for 3 min (at room temperature). Finally, the specimens
were dried and observed under an inverted microscope (Nikon
Corporation).
Reverse transcription-quantitative PCR
(RT-qPCR) analysis
Total RNA was extracted from the control and treated
cells using TRIzol reagent. The RNA samples were transcribed into
cDNA with a Reverse Transcription kit (Vazyme). The qPCR assays
were performed with a ChamQ SYBR qPCR Master Mix kit (Vazyme) and
analyzed on the StepOne™ Real-Time PCR system (Thermo Fisher
Scientific, Inc.) All procedures were conducted according to the
manufacturers' protocols with 5 µl 2X ChamQ SYBR qPCR Master
Mix, 0.2 µl 50X ROX Reference Dye1, 2 µl template
cDNA (diluted with nuclease-free water), 0.2 µl primer 1 (10
µM), 0.2 µl primer 2 (10 µM) and 2.4 µl
ddH2O. The reaction was carried using specific thermal
cycling conditions, including the 3 following stages: Stage 1, 95°C
for 3 min; stage 2, 95°C for 10 sec, 56°C for 30 sec and 72°C for
30 sec; stage 3, 95°C for 15 sec, 60°C for 60 sec and 95°C for 15
sec. Each gene primer sequence was designed as follows: GAPDH
forward, 5′-AGG TCG GTG TGA ACG GAT TTG-3′ and reverse, 5′-TGT AGA
CCA TGT AGT TGA GGT CA-3′; aldolase A (ALDOA) forward, 5′-ATG CCC
TAC CAA TAT CCA GC-3′ and reverse, 5′-GAC AGC CCA TCC AAC CCT-3′;
glucose phosphate isomerase (GPI) forward, 5′-CAA GGA CCG CTT CAA
CCA CTT-3′ and reverse, 5′-CCA GGA TGG GTG TGT TTG ACC-3′; enolase
2 (ENO2) forward, 5′-CGT TAC TTA GGC AAA GGT GTC C-3′ and reverse,
5′-CTC CAG CAT CAG GTT GTC CAG T-3′; LDHA forward, 5′-ATG GCA ACT
CTA AAG GAT CAG C-3′ and reverse, 5′-CCA ACC CCA ACA ACT GTA ATC
T-3′; GLUT-1 forward, 5′-CGG GCC AAG AGT GTG CTA AA-3′ and reverse,
5′-TGA CGA TAC CGG AGC CAA TG-3′; and phosphofructokinase-1 (PFK1)
forward, 5′-GCC ATC AGC CTT TGA CAG A-3′ and reverse, 5′-CTC CAA
AAG TGC CAT CAC TG-3′. The relative mRNA expression levels were
normalized to those of the GAPDH and were determined using the
comparative Cq method (2−ΔΔCq) (16).
Western blot analysis
The cells were plated in 6-well plates and collected
for protein extraction following treatment with 60 µM CANA
and the PI3K/AKT/mTOR inhibitor, LY294002 (25 µM, MedChem
Express) for 24 h (17). The cells
were lysed in RIPA buffer in the presence of protease inhibitor
cocktail. The protein concentration was determined using a BCA
assay (Beyotime Institute of Biotechnology). The cell lysates were
subjected to SDS-PAGE gels (8% polyacrylamide sepa-ration gel and
5% concentrated gel) and transferred onto the PVDF membrane to
separate proteins (50 µg per lane). The following primary
antibodies were used: Mouse anti-β-actin (1:500; cat. no. sc-8432),
mouse anti-hypoxia-inducible factor (HIF)-1α (1:500; cat. no.
sc-13515), mouse anti-GLUT-1 (1:500; cat. no. sc-377228), mouse
anti-LDHA (1:500; cat. no. sc-137243), rabbit anti-phosphorylated
(p-)mammalian target of rapamycin (p-mTOR; 1:1,000; cat. no.
sc-293133), rabbit anti-mTOR (1:1,000; cat. no. sc-517464; all from
Santa Cruz Biotechnology, Inc.), rabbit anti-PI3K (1:1,000; cat.
no. 3011), rabbit anti-p-AKT (1:2,000; cat. no. 4060) and rabbit
anti-AKT (1:1,000; cat. no. 4691; all from Cell Signaling
Technology, Inc.). The blots were detected using a Bio-Imaging
System (Bio-Rad Laboratories, Inc.) and the ImageJ software was
used for expression analysis (http://rsbweb.nih.gov/ij/).
Statistical analysis
GraphPad Prism 4.0 software (GraphPad Software) was
used for all statistical analyses. The significant differences
between 2 groups were examined using the Student's t-test.
Statistical comparisons between ≥3 groups were tested using one-way
analysis of variance (ANOVA), followed by Tukey's multiple
comparison test. The data are presented as the means ± SD.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Pancreatic cancer is effectively
suppressed by CANA in vitro
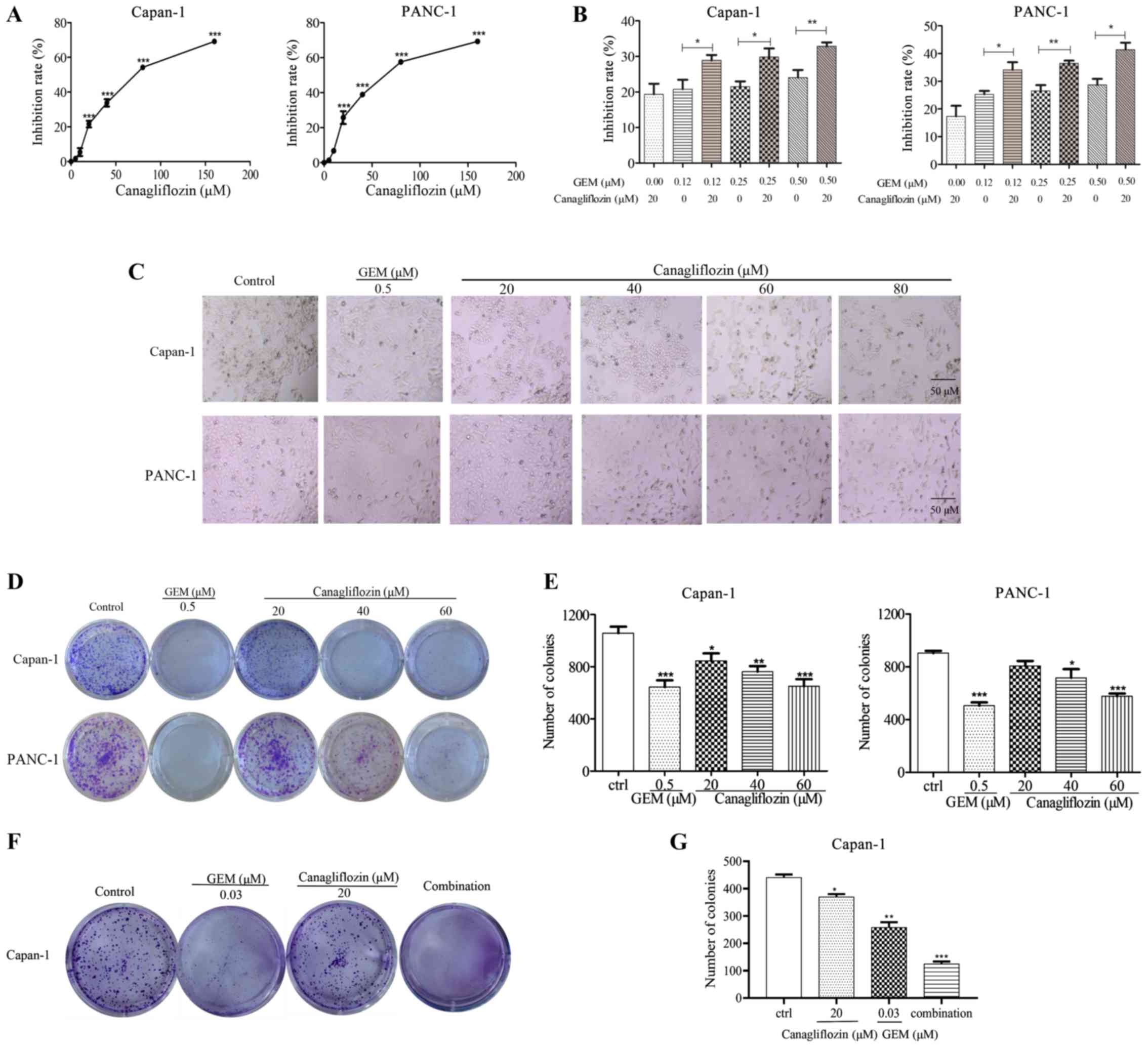
To examine the effects of CANA against pancreatic
cancer, the Capan-1 and PANC-1 cells were incubated with CANA for
48 h and the viability of the cells was measured by MTT assay. The
viability of the Capan-1 and PANC-1 cells was decreased by CANA in
a dose-dependent manner, with peak inhibition rates of 54.3 and
57.6% in the cultured Capan-1 and PANC-1 cells, respectively
treated with 80 µM CANA (Fig.
1A). The IC50 values in the Capan-1 and PANC-1 cells
were 77 and 68 µM, respectively. The inhibitory rate
increased from 21.5 and 26.6% to 29.8 and 36.4% in the Capan-1 and
PANC-1 cells, respectively, following combined treatment with 20
µM CANA and gemcitabine compared to treatment with
gemcitabine (0.25 µM) alone (Fig. 1B). The data indicated that cell
proliferation was effectively inhibited by the combination of
gemcitabine and CANA compared with single treatment of the cells
with either compound. Moreover, the number of cells was decreased
and the morphology of the PANC-1 cells was altered from a round to
an elongated and shrunken shape following the treatment with 60
µM CANA (Fig. 1C). The
ability of 60 µM CANA treatment to inhibit the colony
formation of the cancer cells was comparable to that of the
positive control drug (Fig. 1D and
E). Combined treatment with 0.03 µM gemcitabine and 20
µM CANA significantly increased the ability of gemcitabine
to inhibit the colony formation of the Capan-1 cells compared to
treatment with gemcitabine alone (Fig.
1F and G). These data suggested that CANA inhibited pancreatic
cancer cell growth and colony formation. Moreover, CANA enhanced
the ability of gemcitabine to inhibit pancreatic cancer cell
proliferation and colony formation.
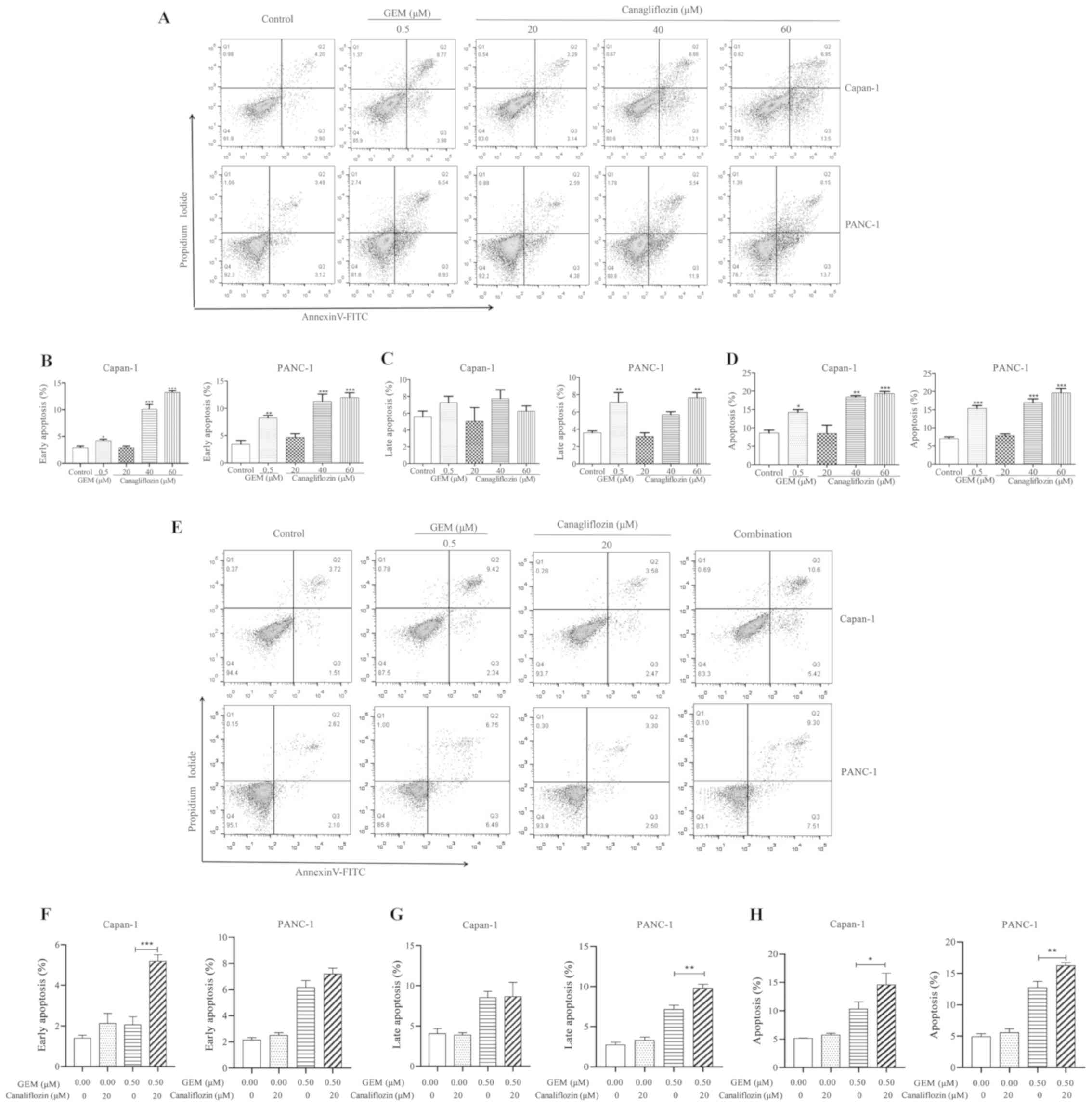
CANA induces pancreatic cancer cell
apoptosis
To further examine whether CANA suppresses cell
growth by inducing cell apoptosis, the tumor cells were stained
with Annexin V/propidium iodide following drug administration and
cell apoptosis was detected by flow cytometry. As shown in Fig. 2, the data in the flow cytometry
plots are representative images, while the data in the bar charts
represent the overall data. Thus, the data shown in the plots and
graphs do not necessarily represent each other. Following treatment
with 60 µM CANA, the apoptotic rate of the Capan-1 and
PANC-1 cells, notably of the early apoptotic cells, was
significantly increased, reaching 13.2 and 11.3%, respectively
(Fig. 2A and B). However, the
percentage of late apoptotic PANC-1 cells exhibited a more obvious
effect (compared to the control) following treatment with 60
µM CANA than that of the late apoptotic Capan-1 cells
(Fig. 2C), suggesting that the
PANC-1 cells may be more sensitive to CANA than the Capan-1 cells.
Moreover, the apoptotic rate of the Capan-1 and PANC-1 cells
increased in a dose-dependent manner, and the apoptotic rate of the
40 µM CANA-treated group reached 18.4% in the Capan-1 cells,
while that of the GEM-treated group was only 14.2% (Fig. 2D). The early apoptotic rate of the
Capan-1 cells was significantly increased when gemcitabine (0.5
µM) was used in combination with CANA (20 µM).
However, the late apoptotic rate of the PANC-1 cells was markedly
improved with this treatment combination (Fig. 2F and G). The total apoptotic rate
of the Capan-1 and PANC-1 cells was significantly increased from
11.8 and 12.5% to 14.9 and 16.3%, respectively when gemcitabine was
used in combination with CANA (Fig. 2E
and H). These data suggested that CANA inhibited tumor growth
by inducing the apoptosis of pancreatic cancer cells. Moreover,
CANA enhanced the effects of gemcitabine on the induction of the
apoptosis of pancreatic cancer cells.
Rate of glycolysis in pancreatic cancer
cells is inhibited by treatment with CANA via the PI3K/AKT/mTOR
pathway
The majority of tumor cells exhibit the constitutive
upregulation of glycolysis, which is known as the Warburg effect
(18). Tumor cells meet their own
energy demands through glycolysis. Since CANA is a
metabolism-associated drug, it was hypothesized that it may affect
glycolysis in tumor cells. In the present study, the glucose uptake
and lactate release rates were examined in the Capan-1 and PANC-1
cells for a certain period of time following treatment with the
drugs. It is interesting to note that the glucose uptake rate was
reduced in the CANA-treated group (60 µM CANA) of Capan-1
and PANC-1 cells by 4.8 and 4.9%, respectively compared with that
of the control group (Fig. 3A).
The lactate release rate was reduced by 25.5 and 43.4% (Fig. 3B). In addition, the mRNA expression
levels of glycolysis-associated genes were detected by RT-qPCR at
24 h following treatment of the Capan-1 and PANC-1 cells with CANA.
Specifically, the mRNA expression levels of GLUT-1 and LDHA were
markedly decreased (Fig. 3C). CANA
was thus capable of inhibiting glycolysis by downregulating GLUT-1
and LDHA in pancreatic cancer cells.
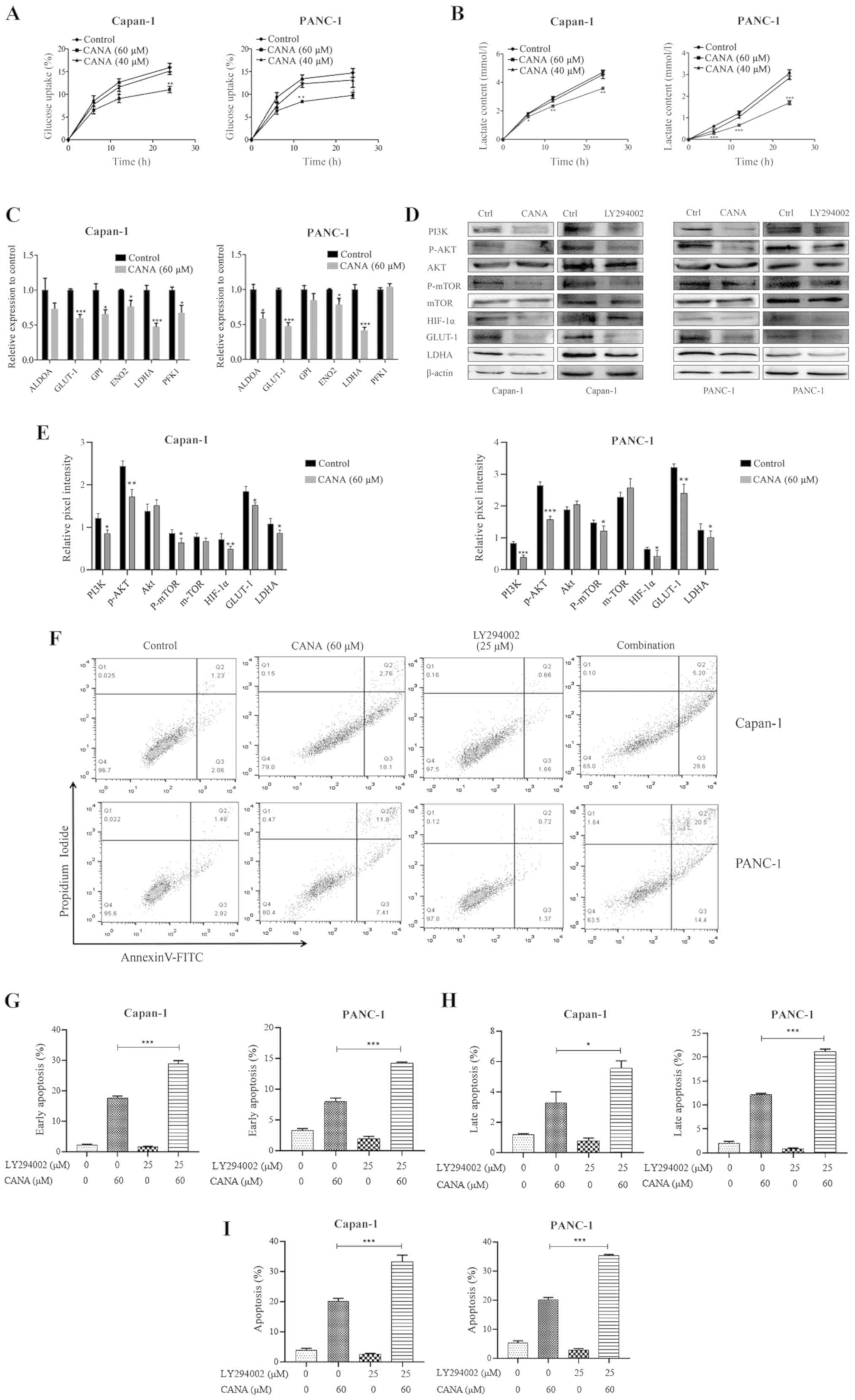 | Figure 3Glucose uptake, lactate release,
GLUT-1 and LDHA expression and PI3K/AKT/mTOR signaling were
inhibited by CANA in cultured Capan-1 and PANC-1 cells. The
concentrations of glucose and lactate in the supernatant were
assessed at 6, 12 and 24 h. (A) The glucose uptake rate was
measured in the control and treated groups of Capan-1 and PANC-1
cells. (B) Lactate release of control and treated cells. (C)
Capan-1 and PANC-1 cells were treated with 60 µM CANA. Total
RNA was extracted and the mRNA expression levels of ALDOA, GLUT-1,
GPI, ENO2, LDHA and PFK1 were analyzed by RT-qPCR. (D) The protein
levels of PI3K, p-AKT, p-mTOR, HIF-1α, GLUT-1 and LDHA were
determined by western blot analysis. (E) The relative expression
levels were normalized to the control. (F) Capan-1 and PANC-1 cells
were treated with CANA alone or in combination with LY294002. The
apoptotic rate of Capan-1 and PANC-1 cells was measured by an
Annexin V (AV)/propidium iodide (PI) apoptosis detection kit. (G-I)
The percentages of apoptotic cell are quantified. The data in the
flow cytometry plots are representative images, while the data in
the bar charts represent the overall data. Thus, the data in the
plots and graphs do not necessarily represent each other. The data
are presented as the means ± SD of 3 independent experiments.
*P<0.05, **P<0.01 and
***P<0.001 compared to the control. GLUT-1, glucose
transporter-1; LDHA, lactate dehydrogenase A; CANA, canagliflozin;
RT-qPCR, reverse transcription-quantitative PCR. |
The PI3K/AKT/mTOR pathway is involved in the
glycolytic process and HIF-1α is a major transcriptional regulator
of the response to hypoxia that can induce several genes encoding
glycolytic enzymes (19).
Therefore, the protein levels of PI3K, p-AKT, p-mTOR, HIF-1α,
GLUT-1 and LDHA were determined by western blot analysis in the
control and treatment groups. The data indicated that CANA not only
inhibited the PI3K, AKT and mTOR phosphorylation reactions, but it
also downregulated HIF-1α, GLUT-1 and LDHA expression. In addition,
the Capan-1 and PANC-1 cells were treated with the PI3K/AKT/mTOR
inhibitor, LY294002, for 24 h and the protein expression levels of
PI3K, P-AKT, P-mTOR, HIF-1α, GLUT-1 and LDHA were apparently
inhibited, as noted following treatment of the cells with CANA
(Fig. 3D and E). In addition,
effects of the combination of CANA with the PI3K/AKT pathway
inhibitor, LY294002, on the apoptosis of the PANC-1 and Capan-1
cells were also investigated. As shown in Fig. 3F-I, the data in the flow cytometry
plots are representative images, while the data in the bar charts
represent the overall data. Thus, the data in the plots and graphs
do not necessarily represent each other. The total apoptotic rate
of the Capan-1 and PANC-1 cells increased from 20.0 and 19.8% to
34.3 and 35.5%, respectively, when CANA (60 µM) was used in
combination with 25 µM LY294002 (Fig. 3F and I). The data mentioned above
suggest that LY294002 enhances the effect of CANA on the induction
of the apoptosis of pancreatic cancer cells.
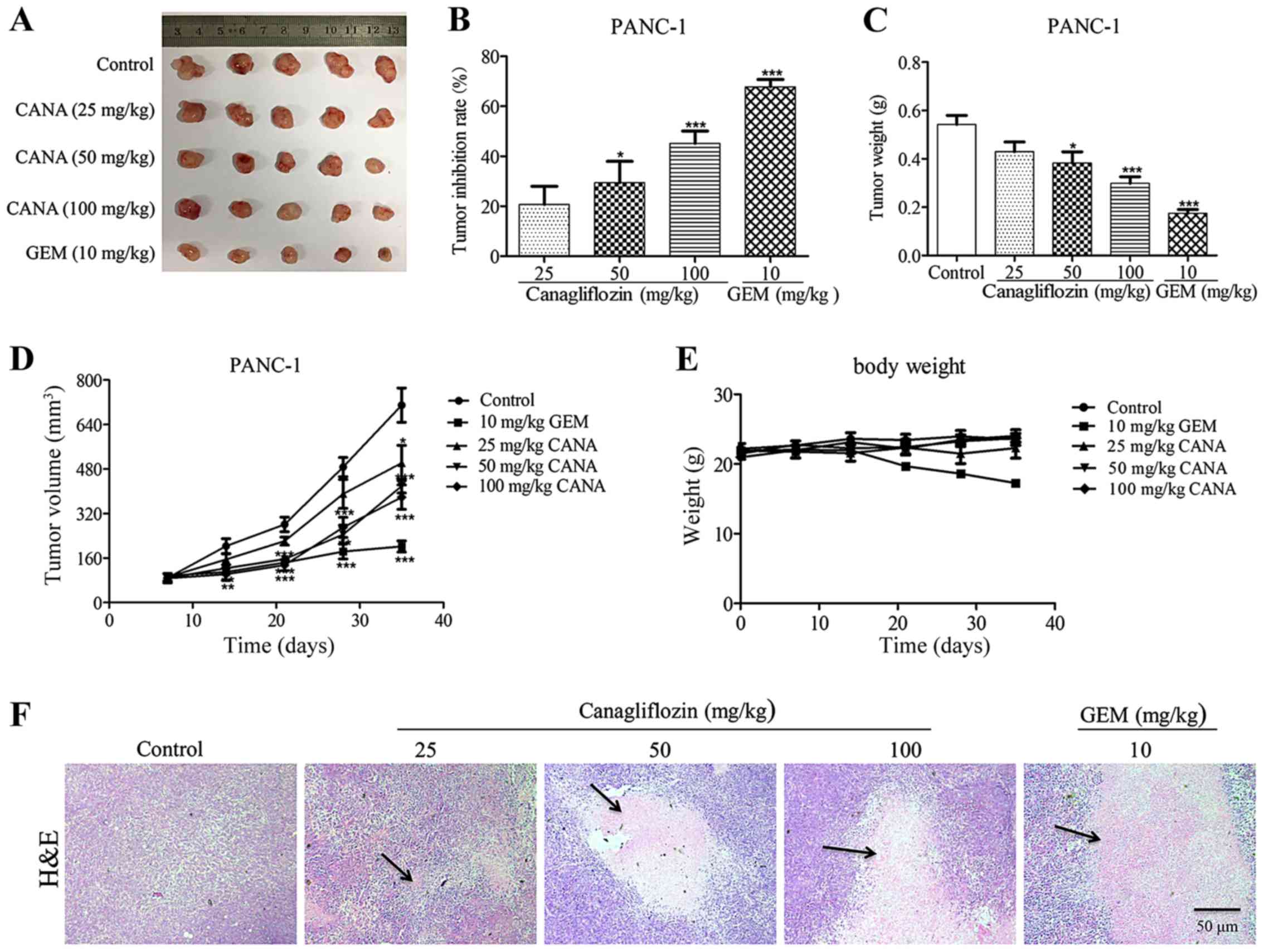
PANC-1 tumor growth is effectively
inhibited during the treatment of nude mice with CANA
As CANA suppressed the growth of pancreatic cancer
cells in vitro, the anti-tumor effects of this compound were
further investigated in nude mice. The mice were subcutaneously
implanted with PANC-1 cells (100 µl, 6×106
cells/ml) and treated with CANA at 3 different doses, with
gemcitabine alone or with the vehicle control. The group that
received CANA exhibited an effective inhibition of tumor growth
(Fig. 4A-D), with tumor inhibitory
rates of 20.7, 29.4 and 45.1% at the doses of 25, 50 and 100 mg/kg
in PANC-1 (Fig. 4B) nude mice,
respectively. Similarly, the weights and volumes of the tumors were
apparently decreased in a dose-dependent manner (Fig. 4C and D). Following 5 weeks of
administration, the mean body weight of the CANA-treated groups was
maintained at approximately 22 g, whereas that of the mice in the
GEM-treated group was significantly decreased to 17 g (Fig. 4E), suggesting that CANA was less
toxic than the positive control drug, gemcitabine. Moreover,
H&E staining indicated the necrosis of parenchymal tumor tissue
in the CANA-treated group compared with that of the control group
(Fig. 4F). Collectively, these
results indicated that CANA effectively inhibited the growth of
PANC-1 tumors with minimal toxicity to the mice.
Glycolysis-associated mRNA and protein
expression levels are suppressed in vivo
To investigate whether the antitumor effects of CANA
are associated with the inhibition of the glycolytic process, the
mRNA and protein levels of glucose metabolism-associated genes were
assayed by RT-qPCR and western blot analysis in the PANC-1-derived
tumor tissues. The data indicated that the mRNA expression levels
of GLUT-1 and LDHA were decreased by 71.5 and 56.6%, respectively,
at the dose of 25 mg/kg CANA (Fig. 5A
and B). Concomitantly, the protein levels of PI3K, p-AKT,
p-mTOR, HIF-1α, LDHA and GLUT-1 in the CANA-treated group were also
decreased in the PANC-1-derived tumors (Fig. 5C and D), suggesting that glycolysis
was inhibited by CANA via the PI3K/AKT/mTOR pathway in
vivo.
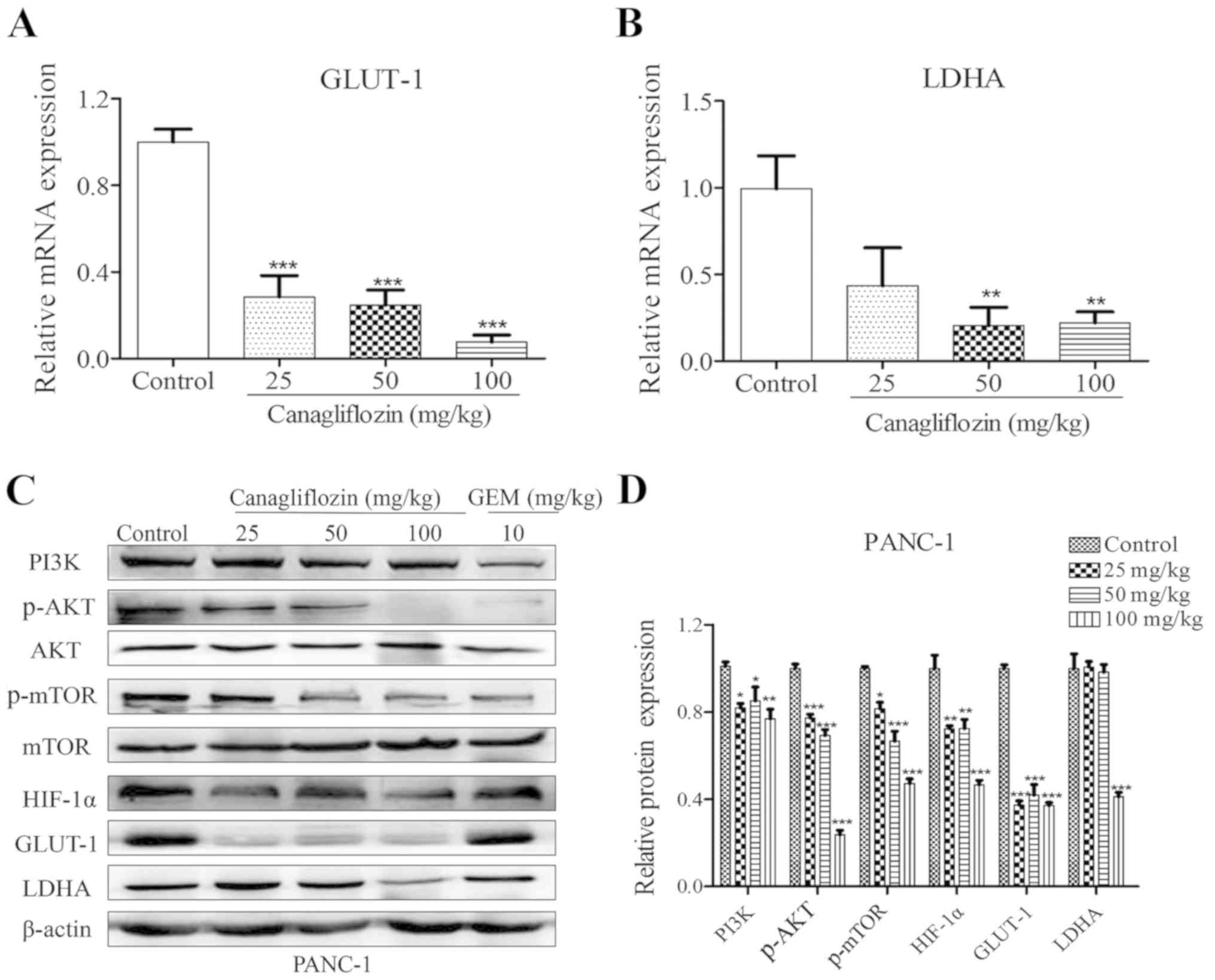 | Figure 5CANA dose-dependently suppresses the
expression of GLUT-1 and LDHA, and PI3K/AKT/mTOR signaling in
PANC-1 tumors. (A and B) Total RNA was extracted from PANC-1 tumors
and the mRNA expression levels of GLUT-1 and LDHA were analyzed by
RT-qPCR. (C) The protein levels of PI3K, p-AKT, p-mTOR, HIF-1α,
GLUT-1 and LDHA were determined by western blot analysis and
quantified by ImageJ software. (D) The relative expression levels
were normalized to the control. Data are presented as the means ±
SD of 3 independent experiments, *P<0.05,
**P<0.01 and ***P<0.001 compared to the
control. GLUT-1, glucose transporter-1; LDHA, lactate dehydrogenase
A; CANA, canagliflozin; RT-qPCR, reverse transcription-quantitative
PCR; p-, phosphorylated. |
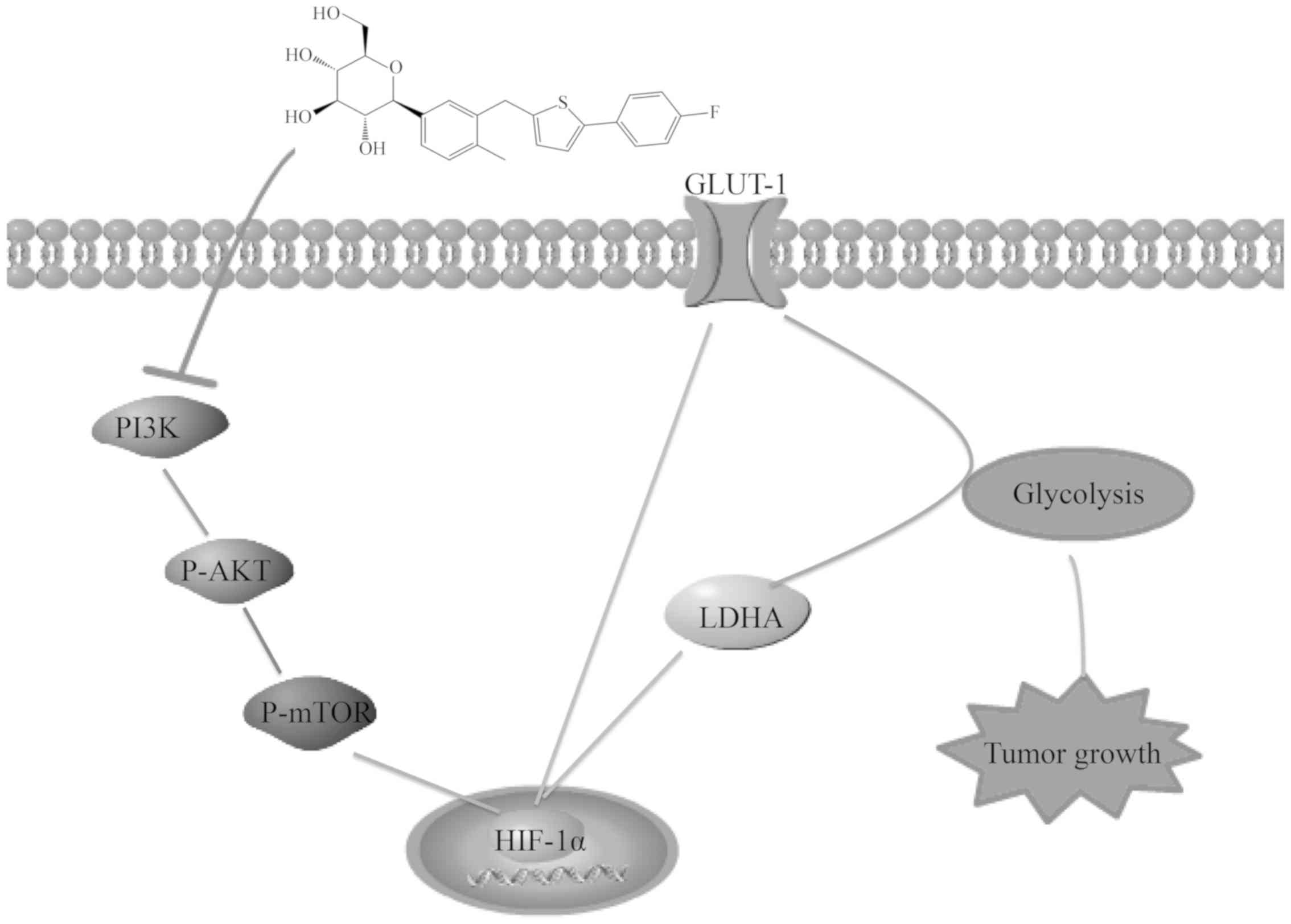
CANA is capable of suppressing pancreatic
cancer growth by inhibiting glycolysis
Cancer cells always reprogram their metabolism and
enhance glycolysis to meet the constantly growing requirement for
intermediate, the activation of the PI3K/AKT signaling pathway can
promote pancreatic cancer proliferation. HIF-1α is a major
transcriptional regulator of the response to hypoxia that can
induce several genes encoding glycolytic enzymes, such as glucose
transporter-1 (GLUT-1), lactate dehydrogenase A (LDHA), which are
highly expressed in tumors are involved in the glycolytic process
(9). The ability of CANA to
inhibit glycolysis is achieved by the downregulation of GLUT-1 and
LDHA at the mRNA and protein levels via the PI3K/AKT/mTOR/HIF-1α
signaling pathway which effectively inhibiting the growth of
pancreatic cancer (Fig. 6).
Discussion
As one of the most malignant and lethal solid
cancers, pancreatic cancer is associated with insidious clinical
symptoms, rapid progression, a poor prognosis and high mortality
rates (20-22). This is probably due to low
vascularization, prominent fibrotic stromal reactions, genomic
complexity and metabolic alterations (23,24).
Currently, the first-line drug for the clinical treatment of
pancreatic cancer causes significant side-effects, such as
nephrotoxicity, allergic reactions and edema (25). Therefore, it is imperative to
develop a safe and effective drug for treating pancreatic cancer.
In the present study, the data indicated that CANA effectively
inhibited the growth of pancreatic cancer cells (Fig. 1A and D) and suppressed the growth
of PANC-1 tumors. The highest tumor inhibition rate reached 45.1%
at doses of 100 mg/kg (Fig. 4A and
B), suggesting that pancreatic cancer could be effectively
inhibited during CANA treatment in vitro and/or in
vivo. Moreover, the combination of CANA with gemcitabine
improved the efficacy of gemcitabine in inhibiting pancreatic
cancer cell proliferation and colony formation and inducing
apoptosis in both cell lines (Figs.
1B, F and 2E). Furthermore, the apoptotic rate of the 40
µM CANA-treated group was higher than that of the
GEM-treated group, indicating that CANA could apparently induce
pancreatic cancer cell apoptosis (Fig.
2A and D). The mean body weight of the CANA-treated mouse
groups did not decrease, while that of the mice in the GEM-treated
group was significantly reduced (Fig.
4E), suggesting that the toxicity of CANA was much lower than
that of the positive drug gemcitabine. The data suggested that
pancreatic cancer could be effectively inhibited by the
low-toxicity drug CANA through the induction of apoptosis, which
indicated its potential efficacy in clinical applications. However,
further investigations are required to determine whether the cell
cycle and metastatic activity of pancreatic cancer cells are
affected by CANA. It was found by preliminary studies that the
combined anti-tumor efficacy of CANA with gemcitabine was better
than that of gemcitabine alone in Capan-1 and PANC-1 cells. In
addition, the efficacy of the combined group in the mouse
pancreatic cancer model and in the associated mechanisms can be
studied to provide theoretical support for future clinical
medication.
Otto Warburg was the first to demonstrate that tumor
cells depend more on aerobic glycolysis accompanied by accelerated
glucose uptake, which is currently known as the Warburg effect
(6). The Warburg effect is a
common trait of cancer, including pancreatic cancer (19,26).
Therefore, targeting metabolic reprogramming may be an extremely
promising strategy for the treatment of pancreatic cancer.
Everolimus is a clinically used drug for the treatment of advanced
cancer. It inhibits human pancreatic cancer cell proliferation and
induces apoptosis by interfering with glycolysis. The sodium
glucose cotransporter 2 inhibitor, CANA, is currently used
clinically in the treatment of diabetes. In the present study, the
glucose uptake and lactate release rates of PANC-1 cells were
reduced in the treated group (60 µM CANA) by 4.9 and 43.3%,
respectively compared with those of the control group (Fig. 3A and B). This suggested that CANA
inhibited pancreatic cancer progression by suppressing glucose
metabolism, which is consistent with previous studies. It could be
concluded that suppressing metabolic reprogramming is a potential
option for pancreatic cancer treatment. However, only preliminary
studies have been conducted on the suppression of glucose
metabolism during treatment with CANA. To fully examine the
mechanism by which CANA inhibits pancreatic cancer, it is necessary
to further determine whether this compound affects the metabolic
function of mitochondria.
Due to its polar hydrophilicity, glucose cannot
freely pass through the cell membrane lipid bilayer structure and
requires transportation via GLUT. Recent studies have reported that
extracellular glucose can be transported into cancer cells via
GLUT-1 in order to participate in the glycolytic process (27,28).
GLUT-1 and LDHA are overexpressed in pancreatic cancer (29,30).
The identification of a drug targeting GLUT-1 and LDHA may have
significant applications in the treatment of pancreatic cancer. It
has also been reported that HIF-1α is associated with the
PI3K/AKT/mTOR pathway (31) and
that it induces the expression of genes involved in glycolysis
(32,33). Moreover, HIF-1α stimulates the
production of glycolytic energy by transactivating the gene GLUT-1
and by inducing LDHA to catalyze lactic acid conversion (34,35).
The results indicated that the mRNA expression levels of the
glycolytic genes, including GLUT-1 and LDHA, were downregulated
(Figs. 3C, 5A and B), whereas the protein levels of
PI3K, p-AKT, p-mTOR, HIF-1α, GLUT-1 and LDHA were suppressed by
CANA treatment to pancreatic cells. In order to determine whether
CANA functions through the PI3K/AKT pathway, the combination of
CANA with the PI3K/AKT pathway inhibitor, LY294002, on the
apoptosis of PANC-1 and Capan-1 cells was investigated. Apparently,
the total apoptotic rate of the Capan-1 and PANC-1 cells increased
from 20.0 and 19.8% to 34.3 and 35.5%, respectively (Fig. 3F and I), suggesting that LY294002
can enhance the effect of CANA on inducing apoptosis of pancreatic
cancer cells. However, there has no enough evidence to clarify if
it only functioned through the PI3K/AKT pathway or other pathways,
which warrants further investigations. In contrast to these
findings, gemcitabine did not induce these effects (Figs. 3D and 5C), suggesting that glycolysis in
pancreatic cancer was reduced by the downregulation of GLUT-1 and
LDHA expression caused through the PI3K/AKT/mTOR signaling pathway
(Fig. 6). These effects were
associated with the tumorigenic processes, including cell
proliferation (Fig. 6). Currently,
the additional molecules or pathways regulated by CANA have not
been identified and warrant further investigation in future
studies.
The exploration of new applications for existing
drugs can reduce the cost and time required during drug
development. The current findings provide novel evidence and
demonstrate that CANA effectively inhibits the growth of pancreatic
cancer in addition to its anti-diabetic role. CANA is capable of
suppressing pancreatic cancer growth by inhibiting glycolysis,
which is achieved by the downregulation of GLUT-1 and LDHA at the
mRNA and protein levels via the PI3K/AKT/mTOR/HIF-1α signaling
pathway (Fig. 6). It is also
interesting to note that the combination of CANA and gemcitabine
exhibited an optimal therapeutic effect on pancreatic cancer cells,
providing a theoretical basis and guidance for clinical drug use.
These data provide important information for future targeted
research on the use of CANA as a chemo-therapeutic drug for other
tumor types. It is expected that the addition of CANA as a novel
chemotherapeutic drug may benefit pancreatic cancer patients
treated with the first-line drug gemcitabine.
Funding
The present study was supported by the MOST of China
(2018YFA0902000, 2019ZX09721-001-04-01), the '111 Project' from the
Ministry of Education of China (grant no. 111-2-07) and 'Double
First-Class' University Project (grant no. CPU2018GF/GY16).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
DX, YZ and CZ initiated the study, and conceived and
performed the experiments. XX, LH and JD contributed to the data
analysis. SP designed the combined administration protocol and
provided the data. BS and CZ contributed to the conception of the
study and assisted with data interpretation. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were performed according to
the protocols approved by the Ethics Committee of China
Pharmaceutical University (approval no. SYXK 2016-0011).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Garrido-Laguna I and Hidalgo M: Pancreatic
cancer: From state-of-the-art treatments to promising novel
therapies. Nat Rev Clin Oncol. 12:319–334. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Skandalakis LJ, Rowe JS Jr, Gray SW and
Skandalakis JE: Surgical embryology and anatomy of the pancreas.
Surg Clin North Am. 73:661–697. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Burris HA III, Moore MJ, Andersen J, Green
MR, Rothenberg ML, Modiano MR, Cripps MC, Portenoy RK, Storniolo
AM, Tarassoff P, et al: Improvements in survival and clinical
benefit with gemcitabine as first-line therapy for patients with
advanced pancreas cancer: A randomized trial. J Clin Oncol.
15:2403–2413. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Levine AJ and Puzio-Kuter AM: The control
of the metabolic switch in cancers by oncogenes and tumor
suppressor genes. Science. 330:1340–1344. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Warburg O: On the origin of cancer cells.
Science. 123:309–314. 1956. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Vidal C, Rauly I, Zeggari M, Delesque N,
Esteve JP, Saint-Laurent N, Vaysse N and Susini C: Up-regulation of
somatostatin receptors by epidermal growth factor and gastrin in
pancreatic cancer cells. Mol Pharmacol. 46:97–104. 1994.PubMed/NCBI
|
|
8
|
Miller DM, Thomas SD, Islam A, Muench D
and Sedoris K: c-Myc and cancer metabolism. Clin Cancer Res.
18:5546–5553. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Blum R and Kloog Y: Metabolism addiction
in pancreatic cancer. Cell Death Dis. 5:e10652014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Goldberg MS and Sharp PA: Pyruvate kinase
M2-specific siRNA induces apoptosis and tumor regression. J Exp
Med. 209:217–224. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fantin VR, St-Pierre J and Leder P:
Attenuation of LDH-A expression uncovers a link between glycolysis,
mitochondrial physiology, and tumor maintenance. Cancer Cell.
9:425–434. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim YS and Milner JA: Bioactive food
components and cancer-specific metabonomic profiles. J Biomed
Biotechnol. 2011:7212132011. View Article : Google Scholar
|
|
13
|
Chao EC and Henry RR: SGLT2 inhibition-a
novel strategy for diabetes treatment. Nat Rev Drug Discov.
9:551–559. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kaji K, Nishimura N, Seki K, Sato S,
Saikawa S, Nakanishi K, Furukawa M, Kawaratani H, Kitade M, Moriya
K, et al: Sodium glucose cotransporter 2 inhibitor canagliflozin
attenuates liver cancer cell growth and angiogenic activity by
inhibiting glucose uptake. Int J Cancer. 142:1712–1722. 2018.
View Article : Google Scholar
|
|
15
|
Shiba K, Tsuchiya K, Komiya C, Miyachi Y,
Mori K, Shimazu N, Yamaguchi S, Ogasawara N, Katoh M, Itoh M, et
al: Canagliflozin, an SGLT2 inhibitor, attenuates the development
of hepatocellular carcinoma in a mouse model of human NASH. Sci
Rep. 8:23622018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
17
|
Jiang H, Fan D, Zhou G, Li X and Deng H:
Phosphatidylinositol 3-kinase inhibitor(LY294002) induces apoptosis
of human nasopharyngeal carcinoma in vitro and in vivo. J Exp Clin
Cancer Res. 29:342010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Koppenol WH, Bounds PL and Dang CV: Otto
Warburg's contributions to current concepts of cancer metabolism.
Nat Rev Cancer. 11:325–337. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gatenby RA and Gillies RJ: Why do cancers
have high aerobic glycolysis? Nat Rev Cancer. 4:891–899. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kamisawa T, Wood LD, Itoi T and Takaori K:
Pancreatic cancer. Lancet. 388:73–85. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chu LC, Goggins MG and Fishman EK:
Diagnosis and detection of pancreatic Cancer. Cancer J. 23:333–342.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dreyer SB, Chang DK, Bailey P and Biankin
AV: Pancreatic cancer genomes: Implications for clinical management
and therapeutic development. Clin Cancer Res. 23:1638–1646. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Le A, Rajeshkumar NV, Maitra A and Dang
CV: Conceptual framework for cutting the pancreatic cancer fuel
supply. Clin Cancer Res. 18:4285–4290. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sousa CM and Kimmelman AC: The complex
landscape of pancreatic cancer metabolism. Carcinogenesis.
35:1441–1450. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Heinrich S and Lang H: Neoadjuvant therapy
of pancreatic Cancer: Definitions and benefits. Int J Mol Sci.
18:16222017. View Article : Google Scholar :
|
|
26
|
Garber K: Energy deregulation: Licensing
tumors to grow. Science. 312:1158–1159. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yoon SO, Jeon TJ, Park JS, Ryu YH, Lee JH,
Yoo JS, Kim JK, Yoon DS and Oh EJ: Analysis of the roles of glucose
transporter 1 and hexokinase 2 in the metabolism of glucose by
extrahepatic bile duct cancer cells. Clin Nucl Med. 40:e178–e182.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang TB, Zhao Y, Tong ZX and Guan YF:
Inhibition of glucose-transporter 1 (GLUT-1) expression reversed
Warburg effect in gastric cancer cell MKN45. Int J Clin Exp Med.
8:2423–2428. 2015.PubMed/NCBI
|
|
29
|
Sun HC, Qiu ZJ, Liu J, Sun J, Jiang T,
Huang KJ, Yao M and Huang C: Expression of hypoxia-inducible
factor-1 alpha and associated proteins in pancreatic ductal
adenocarcinoma and their impact on prognosis. Int J Oncol.
30:1359–1367. 2007.PubMed/NCBI
|
|
30
|
Rong Y, Wu W, Ni X, Kuang T, Jin D, Wang D
and Lou W: Lactate dehydrogenase A is overexpressed in pancreatic
cancer and promotes the growth of pancreatic cancer cells. Tumor
Biol. 34:1523–1530. 2013. View Article : Google Scholar
|
|
31
|
Li X, Wenes M, Romero P, Huang SC, Fendt
SM and Ho PC: Navigating metabolic pathways to enhance antitumour
immunity and immunotherapy. Nat Rev Clin Oncol. 16:425–441. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang V, Davis DA, Haque M, Huang LE and
Yarchoan R: Differential gene up-regulation by hypoxia-inducible
factor-1alpha and hypoxia-inducible factor-2alpha in HEK293T cells.
Cancer Res. 65:3299–3306. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gordan JD, Bertout JA, Hu CJ, Diehl JA and
Simon MC: HIF-2alpha promotes hypoxic cell proliferation by
enhancing c-myc transcriptional activity. Cancer Cell. 11:335–347.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen C, Pore N, Behrooz A, Ismail-Beigi F
and Maity A: Regulation of glut1 mRNA by hypoxia-inducible
factor-1. Interaction between H-ras and hypoxia. J Biol Chem.
276:9519–9525. 2001. View Article : Google Scholar
|
|
35
|
Firth JD, Ebert BL and Ratcliffe PJ:
Hypoxic regulation of lactate dehydrogenase A. Interaction between
hypoxia-inducible factor 1 and cAMP response elements. J Biol Chem.
270:21021–21027. 1995. View Article : Google Scholar : PubMed/NCBI
|