1. Introduction
The epithelium is a layer of cells that separates an
organism from the external environment; it is coated by mucous gel,
which is mainly composed of mucins (MUCs). MUCs are
high-molecular-weight O-linked glycoproteins synthesized by goblet
cells. The human MUC family contains 22 characterized members (MUC1
to MUC22), which are divided into secreted and transmembrane
glycoproteins (1,2). Secreted gel-forming MUCs, without a
transmembrane domain, include MUC2, MUC5AC, MUC5B, MUC6, MUC7, MUC8
and MUC9. Membrane-bound MUCs form rod-like structures that extend
from 200 to 500 nm beyond the glycocalyx, and members include
MUC3A, MUC3B, MUC4, MUC12, MUC13, MUC15, MUC16, MUC17, MUC20, MUC21
and MUC22 (3-6). To date, MUC1 is the
best-characterized MUC. Cancer-associated MUC1 notably differs from
that of normal cells with regards to its function, biochemical
features and cellular distribution (7,8).
It has been reported that MUC1, which is present in epithelial
cancer cells, is implicated in cell invasion, migration, adhesion,
proliferation, and resistance to apoptosis and chemoradiotherapy
(9). Therefore, MUC1 has been
attracting considerable attention as an oncogenic molecule due to
its crucial role in cancer progression (10). Apoptosis is a fundamental
biological process that allows organisms to remove unwanted cells.
One of the features of malignant transformation is the alteration
of the cell death pathways, which is associated with increased cell
survival. MUC1-dependent suppression of apoptosis causes an
imbalance between cell proliferation and cell death, which in turn
influences cancer progression (9). Therefore, the aim of the present
study was to summarize the divergent mechanisms of action of MUC1
with regard to the alteration of programmed cell death.
2. Apoptosis
Among all types of cell death, apoptosis (also known
as programmed cell death) is the most common; it is a genetically
controlled process, which eliminates unnecessary or damaged
individual cells (11). The
process is crucial in the regulation of cell death and survival
during the development of multicellular organisms and during normal
homeostasis. Apoptosis provides the conditions necessary for
appropriate regulation of numerous physiological processes, such as
embryonic development, immune system function and maintenance of
body homeostasis (12). In
cancerous diseases, an imbalance between cell division and death
occurs due to a lack of appropriate signaling. For example,
downregulation of the tumor suppressor gene p53 may result in
continuous cancer cell proliferation. Therefore, changes in the
ability of the cells to undergo apoptosis may cause malignant
transformation. In this context, the development of novel drugs
that target numerous steps of the apoptotic process is beneficial
in cancer treatment (11,13,14).
The induction of apoptosis is mediated via two main
pathways, namely the receptor- and mitochondria-mediated pathways
of apoptosis. Both pathways are regulated by the B-cell lymphoma
(Bcl)-2 family of proteins, which includes pro- and anti-apoptotic
members. Pro-apoptotic proteins include Bcl-2-associated X protein
(Bax), Bcl-2 antagonist/killer 1 (Bak), BH3 interacting domain
death agonist (Bid), Bad, Bax inhibitor motif (Bim), Bcl-2
interacting killer, Bcl-10, B-lymphocyte kinase, NADPH oxidase
activator and p53 upregulated modulator of apoptosis (PUMA), while
anti-apoptotic proteins mainly include Bcl-2, apoptosis regulator
Bcl-x, B-cell lymphoma-extra large (Bcl-xL), Bcl-extra small,
Bcl-w, induced myeloid leukemia cell differentiation protein
(Mcl-1) and Bcl-2-associated athanogene (12,15,16).
The mitochondrial pathway is also known as the
intrinsic pathway of apoptosis; it is initiated by various
non-receptor-mediated stimuli, such as increased reactive oxygen
species (ROS) or calcium ion levels, release of selected cytokines
[e.g., interferon (IFN)] and immune cells (e.g., T cells), hormone
deficiency, the presence of pathogens (e.g., viruses, bacteria and
their products) or induction of DNA damage (12). Pro-apoptotic signals induce the
translocation of Bax and Bak to the mitochondrial outer membrane
(MOM), where they form pores, thereby altering cell membrane
permeability. The assembly of Bax/Bak oligomers within MOM is
promoted by Bid and Bim (17).
Subsequently, mitochondrial cytochrome c is released into
the cytosol where it forms the apoptosome. The latter is a complex
including cytochrome c, apoptotic protease activating
factor-1 (18,19), pro-caspase-9 and deoxyadenosine
triphosphate (20). Other
molecules released from the mitochondria include second
mitochondria-derived activator of caspase/direct inhibitor of
apoptosis-binding protein with low pI (Smac/DIABLO) and the
mitochondrial serine protease high temperature requirement factor
A2, which contribute to activation of apoptosis inducing factor
(AIF), endonuclease G and caspases. The activation of inactive
caspase-9 pro-enzyme results in cleavage of effector
pro-caspases-3, -6 and -7 (21).
Caspases are cysteine proteases, which are divided
into inflammatory (caspase-1, -4 and -5) and apoptotic proteins.
The latter include initiator enzymes, such as caspase-2, -8 and -9,
and executioner enzymes, such as caspase-3, -6 and -7 (17,20,22). Caspases are synthesized as
pro-enzymes. Following apoptotic stimuli, they are converted into
mature enzymes. Caspase substrates include both enzymatic and
structural proteins. Specific examples include cytokeratins, the
plasma membrane cytoskeletal protein α-fodrin (spectrin-like
protein found in most cells) and microtubule-binding proteins,
which are involved in the formation of the spindle poles and the
segregation of chromosomes during mitosis (nuclear mitotic
apparatus protein). These ultimately lead to the morphological and
biochemical changes of the apoptotic cells. Proteolytic cleavage of
caspase substrates, such as poly (ADP-ribose) polymerase, renders
DNA repair impossible (22-24). Eventually, activation of caspases
results in cell death (14,21).
The initiation of the receptor-associated pathway of
apoptosis, which is also known as the extrinsic pathway of
apoptosis, involves receptor-ligand interactions. These ligands
belong to cytokines of the tumor necrosis factor (TNF) superfamily,
which are termed cognate ligands. Approximately 40 ligand-receptor
pairs have been characterized to date. The most widely known
ligands are fatty acid synthetase ligand (FasL), which binds to the
fatty acid synthetase receptor, TNF-α and TNF-related
apoptosis-inducing ligand or Apo2 ligand (TRAIL/Apo2L). TNF-α can
bind to TNF-receptor 1 (TNF-R1) and TNF-R2, and TRAIL/Apo2L
interacts with four cell surface receptors, namely TRAIL receptor-1
(TRAIL-R1), TRAIL-R2, TRAIL-R3 and TRAIL-R4 (25,26). These receptors are also termed
death receptor DR4, DR5/TRICK2/KILLER, decoy receptor
(DcR)1/TRID/LIT and DcR2/TRUNDD, respectively. The majority of
these receptors contain a C-terminal region, which includes an
80-amino acid (aa) death domain, an extracellular N-terminal region
and a transmembrane domain. The signal transduction machinery has
been described in detail using FasL/Fas and TNF-α/TNF-R1 models
(26). Receptor-ligand binding
results in receptor trimerization and the recruitment of adapter
proteins, which contain similar death domains. FasL activation
occurs following binding of this ligand with Fas, which leads to
the interaction of the receptor with the adapter protein
FAS-associated with death domain (FADD/MORT1). TNF-α binding to
TNF-R allows the recruitment of the adapter protein TNF receptor
type 1-associated death domain (TRADD), FADD and
receptor-interacting protein (20,26). TRADD participates in signal
transduction mediated by TNF-R, but not by TRAIL and Fas (25). Subsequently, FADD aggregates with
pro-caspase-8 via the death effector domain (DED) sequence, leading
to formation of the death-inducing signaling complex (DISC).
Following proteolytic cleavage of its precursor, active caspase-8
is released and triggers activation of executive caspases,
primarily caspase-3 (12). In
addition, caspase-8 cleaves and activates Bid to t-Bid, which is
its truncated form. The latter induces the release of mitochondrial
cytochrome c and pro-apoptotic factors, and links the
extrinsic with the intrinsic pathway of apoptosis (27-29). Cellular FADD-like IL-1β-converting
enzyme-inhibitory protein (FLIP) can bind to FADD and caspase-8,
causing their inactivation and, consequently, the inhibition of the
apoptotic response (12).
Anoikis is a type of programmed cell death that
prevents epithelial cells from seeding to abnormal sites and takes
place when cells lose adhesion to the extracellular matrix
(30,31). During anoikis, both apoptotic
pathways are activated (32,33). Detachment from the surrounding
matrix induces translocation of the pro-apoptotic protein Bim to
the mitochondria, where it binds to Bcl-xL, neutralizing its
anti-apoptotic functions. Moreover, decreased proteosomal
degradation of Bim promotes Bax/Bak oligomerization, whereas matrix
detachment results in upregulation of FasL and Fas expression, FLIP
downregulation and consequent activation of caspase-8 (34-36).
The apoptotic response of the cells may be
deregulated by various factors. One of the features of human cancer
is the high level of MUC1 expression. Accumulating evidence has
shown that MUC1 can modulate the apoptotic response in different
ways as a result of its interaction with multiple proteins that
participate in the regulation of this process (37,38).
3. MUC1 as a cancer-associated
membrane-bound molecule
MUC1 localization and structure
MUCs are highly glycosylated proteins, and are the
most abundant components of mucus on the epithelial surface. Among
transmembrane MUCs, MUC1 has been widely investigated due to its
role in oncogenesis (39-43). MUC1 is also known as episialin,
polymorphic epithelial MUC, H23 antigen, mucin-like
carcinoma-associated antigen, epithelial membrane antigen, cluster
of differentiation 227, Krebs von den Lungen-6, peanut-reactive
urinary mucin, carcinoma antigen CA15.3 and CA27.29 (the commonly
used serum markers for breast cancer), human milk fat globule
antigen, carcinoma-associated MUC and peanut-reactive urinary MUC
(44). MUC1 (for humans) and Muc1
(for other species) are the most commonly used names assigned at
the 1st International Workshop on Carcinoma-Associated Mucins (San
Francisco, USA; 1990) and are in accordance with the Human Genome
Project mapping conventions (45).
MUC1 is a heterodimeric transmembrane protein that
normally resides at the apical borders of epithelial cells of the
respiratory and gastrointestinal tracts, as well as in the ducts of
organs such as the liver, kidney, pancreas and mammary gland
(46,47). MUC1 is also expressed by
non-epithelial cells, such as hematopoietic cells, T cells and male
germ cells (48-51).
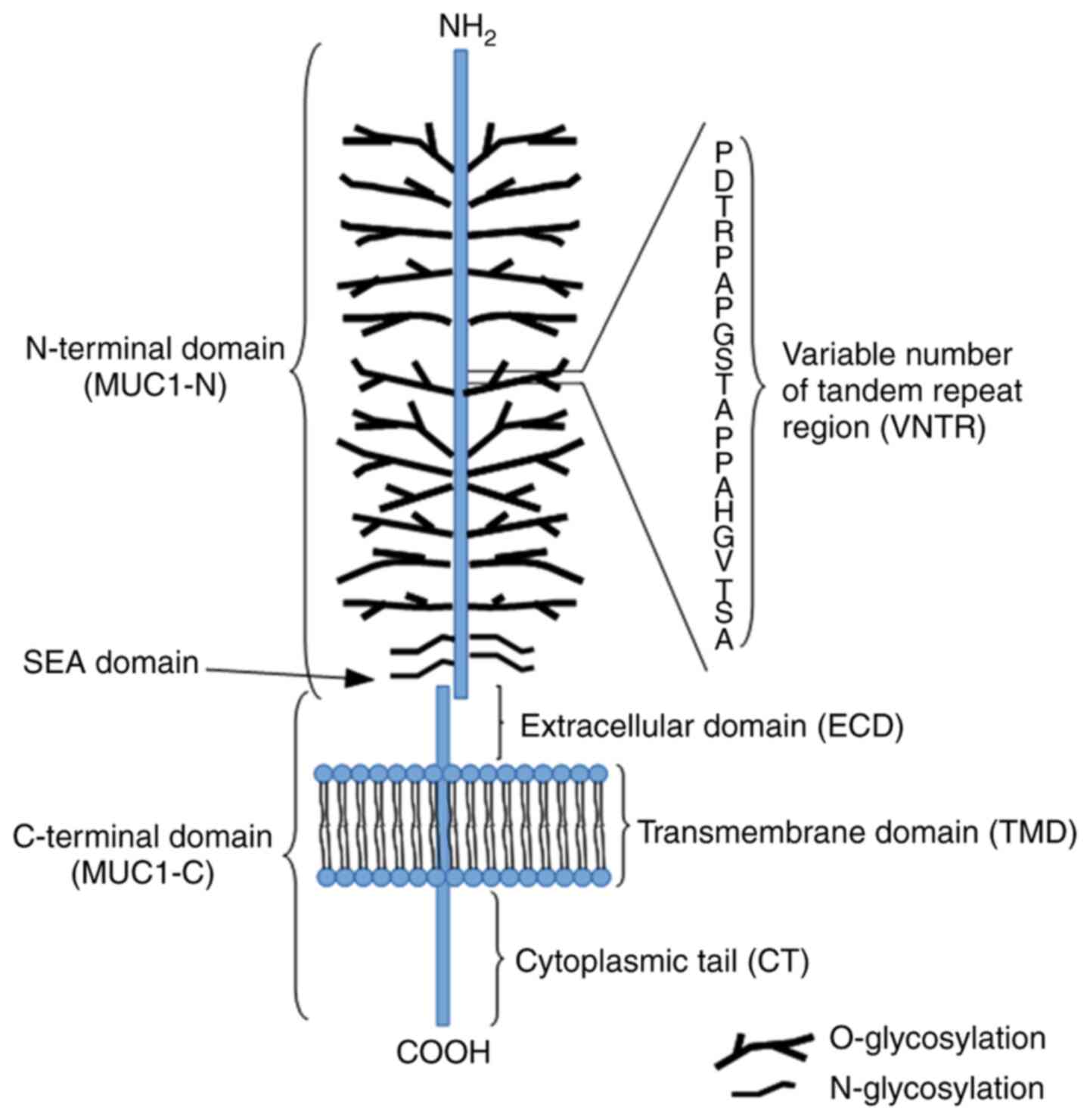
The MUC1 structure contains two distinctive
subunits, one large N-terminal subunit (MUC1-N; subunit α) and one
short C-terminal subunit (MUC1-C; subunit β) derived from
autoproteolytic cleavage of a single polypeptide chain at the
sea-urchin sperm protein enterokinase and agrin domain in the
endoplasmic reticulum. These subunits are bound by non-covalent
interactions (stable hydrogen bonds; Fig. 1) (5,52).
The MUC1-N domain contains an N-terminus (104 aa) and a variable
number of tandem repeat (VNTR) segment (20 aa) sequence
(PDTRPAPGSTAPPAHGVTSA), which is repeated 20-200 times. The domain
is also rich in serine and threonine residues that constitute
potential sites of O-glycosylation, and contains a large C-terminus
170 aa in length (5,52). The molecular mass range of MUC1 is
estimated to 1-40×106 Da. The majority of the protein
modifications include sugar moieties, which constitute 60-80% of
the total weight of the protein and are principally O-glycans. The
complex synthesis of these polysaccharides is based on enzymatic
attachment of monosaccharides to a polypeptide chain by ≥30 or more
glycosyltransferase enzymes (53,54). The detailed synthesis of the
O-glycan basic core structure is presented in Fig. 2. The VNTR region is followed by a
segment containing five N-linked glycosylation sites (5). The MUC1-C is composed of three
regions, including the extracellular domain (ECD), composed of 58
aa, which is responsible for anchoring MUC1 to the cell membrane,
the transmembrane domain, composed of 28 aa, and a cytoplasmic tail
(MUC1-CT), composed of 72 aa. Moreover, the MUC1-C ECD contains
asparagine residues that are used as N-glycosylation sites
(41,55). The MUC1-C
cysteine-glutamine-cysteine (CQC) motif is located below the
transmembrane region and is crucial for the formation of homodimers
(42).
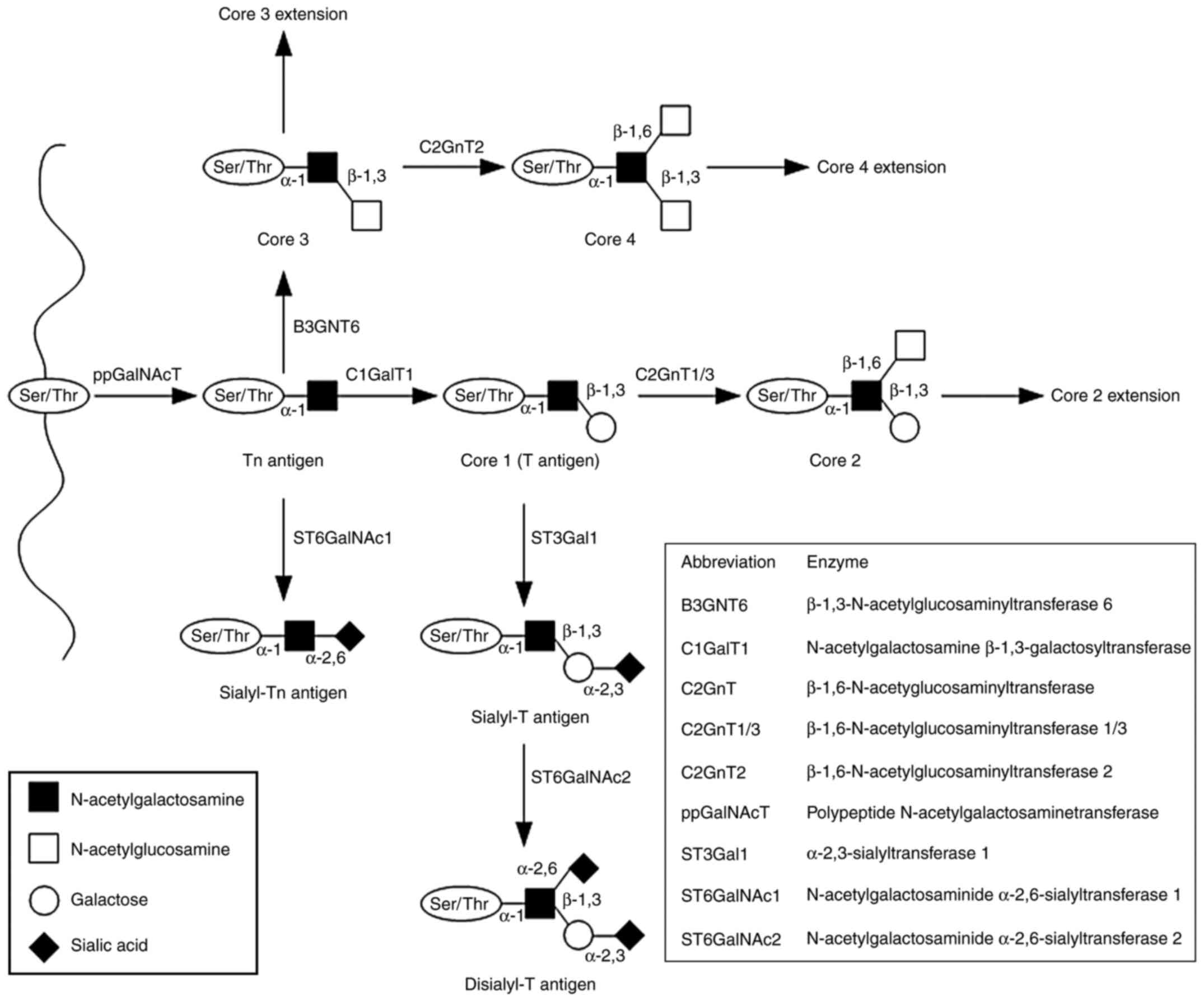 | Figure 2Mucin-type O-glycan synthesis.
O-glycosylation is initiated by attachment of GalNAc to the
hydroxyl groups of the Ser/Thr of the protein chain. The reaction
is catalyzed by ppGalNAcT enzymes and results in the formation of
the Tn antigen (GalNAcα1-O- Ser/Thr). The four basic core
structures are generated in the Golgi apparatus by specific
glycosyltransferases. The Tn antigen can be elongated through
galactose addition catalyzed by C1GalT1 or core 1 synthase, which
results in the synthesis of the T antigen or core 1. The antigen
can also be elongated by B3GNT6, which transfers GlcNAc to the
GalNAc-Ser/Thr structure to form core 3. Subsequent GlcNAc addition
to core 3 forms core 4. Core structures can be further elongated or
terminated by attachment of fucose or sialic acid. GalNAc,
N-acetylgalactosamine; Ser, serine; Thr, threonine; ppGalNAcT,
N-acetylgalactosaminyltransferase; C1GalT1, N-acetylgalactosamine
β-1,3-galactosyltransferase; Galβ1, galactosamine β1; T,
Galβ1-3GalNAcα1-O-Ser/Thr; Tn, GalNAcα1-O-Ser/Thr. |
Functions of MUC1 in physiology
In the past years, a considerable amount of data has
been gathered regarding MUC1 functions. It has been shown that this
MUC participates in complex interactions and is involved in the
regulation of a wide variety of cellular pathways, affecting both
physiological and pathological processes. Substantial glycosylation
of the MUC1 extracellular subunit in normal tissues aims to protect
them from the entry of harmful substances and provide lubrication
to the underlying epithelia (55). Therefore, the N-terminal subunit
can be shed from the cell surface and released into the
extracellular space as a result of proteolytic cleavage in order to
provide a barrier to invading pathogens. Such events may be induced
by a number of inflammatory stimuli, such as IFN-γ and TNF-α
(56,57). This process is also mediated by
specific enzymes, such as TNF-α-converting enzyme/a disintegrin and
metalloproteinase 17 (56,57).
Therefore, MUC1-N can act as a receptor when transmitting stress
signals to the interior of the cell (41). Furthermore, glycans can
participate in cell-cell and cell-matrix interactions and are
involved in the recognition of normal cells by the immune system
(58). Moreover, glycans
participate in the appropriate distribution of proteins, which are
newly synthesized in the endoplasmic reticulum. By affecting
proteolysis, they protect proteins from intra- or extracellular
degradation (58). In addition,
MUC1-CT may be phosphorylated by multiple kinases, including the
epidermal growth factor receptor (EGFR), glycogen synthase
kinase-3β, tyrosine-protein kinase MET, non-receptor tyrosine
kinase (Src), protein kinase C (PKC), glycogen synthase kinase 3
(GSK3) and tyrosine-protein kinase Abl. Previous studies have shown
that MUC1 interacts with diverse molecules and modulates their
activity (41,55).
MUC1 in cancer
Tumor progression is associated with a notable
increase in MUC1 expression, which has been reported in diverse
cancer types, such as colon, breast, lung, pancreatic and prostate
cancer, as well as in hematological malignancies (59-62).
The changes in the oligosaccharide structure of
glycoproteins have received considerable attention due to the key
role of glycans in processes such as cell proliferation,
differentiation, invasion, metastasis and immune surveillance
(63-66). It has been shown that, during the
process of neoplasia, the MUC1 ECD undergoes significant
modifications, including changes in the glycosylation profile via
the following main mechanisms: Incomplete synthesis and synthesis
of atypical forms of glycans (67). The attachment of the truncated
oligosaccharides to the VNTR region changes the MUC1 spatial
structure and increases the availability of the peptide backbone to
other proteins, which in turn affects potential protein-protein
interactions and intracellular signaling (52,68,69).
The majority of human carcinomas, including gastric
and colorectal cancer, contain truncated forms of glycans, such as
N-acetylgalactosamine (GalNAcα)1-O-serine (Ser)/threonine
(Thr) (Tn) and galactose (Gal) β1-3GalNAcα1-O-Ser/Thr (T) antigens
and their sialylated forms Neu5Acα2-6GalNAcα1-O-Ser/Thr (sTn) and
Neu5Acα2-3Galβ1-3GalNAcα1-O-Ser/Thr (sT). These forms are combined
with decreased levels of core 3 and core 4 structures (6,69,70) (Fig.
2); their presence is predominantly associated with changes in
the expression levels of different glycosyltransferases (71,72). For example, the activity of the
core 1 synthase, also known as N-acetylgalactosamine
β-1,3-galactosyltransferase (C1GalT1), depends on the co-expression
of the specific chaperone core 1 β3GalT-specific molecular
chaperone (COSMC). C1GalT1 is an enzyme that catalyzes galactose
addition to the Tn antigen. In the absence of COSMC, the enzyme
loses its function, resulting in higher levels of Tn antigen than
those noted in normal cells (52). Moreover, abnormal expression of
sialyltransferases, including N-acetylgalactosaminide
α-2,6-sialyltransferase 1 and β-galactoside α-2,3-sialyltransferase
1, may be associated with increased MUC1 sialylation and the
formation of sTn and sT antigens (52). The transfer of fucose residues to
oligosaccharides and proteins is catalyzed by fucosyltransferases.
Aberrant fucosylation is associated with neoplasia and may lead to
EGFR stimulation, which in turn affects the functions of integrins
and selectins, and the induction of apoptosis or oncogenesis. Large
amounts of sialylated and fucosylated core 1 and 2 structures have
been detected in gastric, ovarian, renal, colon and prostate cancer
(73-78). Sialylated and fucosylated glycans
(Lewis-type antigens) are composed of free monosaccharides, namely
N-acetylglucosamine, Gal and fucose, which differ in terms of their
corresponding glycosidic bonds. Core 1 structures contain the
Galβ1-3GlcNAc bond (Lewis a, Lewis b), whereas core 2 structures
contain the Galβ1-4GlcNAc bond (Lewis x, Lewis y). Further addition
of sialic acid leads to formation of their sialylated forms
(sLea and sLex); their expression may be
associated with intensification of the neoplastic transformation of
cells (69,74). The sLex and
sLea carbohydrate ligands have been shown to adhere to
E-selectin in vascular endothelial cells. The expression of this
protein may be induced by proinflammatory agents, such as cytokines
(69).
MUC1 function in cancer is associated with its
cellular localization. Loss of cell polarity during epithelial
transformation causes the translocation of the MUC1-N/MUC1-C
complex from the apical to the entire surface of the cell membrane
(5). Therefore, MUC1 may interact
with molecules normally expressed at the basolateral membrane, such
as receptor tyrosine kinases (RTKs), including EGFR. These RTKs, in
turn, activate several signaling pathways, including the
PI3K/protein kinase B/Akt, the p38 mitogen-activated protein (MAP)
kinase, the c-Jun N-terminal kinases (JNK), the Janus kinase/signal
transducers and activators of transcription (JAK/STAT) and the Src
pathways, which are involved in cell proliferation, survival and
differentiation under normal and pathological conditions (79). Formation of extracellular
connections between MUC1 and EGFR requires the MUC1-C/galectin-3
interaction (80). Galectin-3 is
member of a family of β-galactoside-binding lectins, which
influence biological processes, such as cell adhesion,
proliferation, differentiation, inflammation, angiogenesis and
oncogenesis (81,82). In addition, altered glycosylation
may also affect appropriate oligomerization of cell surface
receptors and, therefore, their sensitivity to stimulation
(55,83).
In vitro studies have shown that MUC1-C can
translocate from the cell membrane to the mitochondria, where it
most likely localizes to the MOM (84). The process may be stimulated by
phosphorylation of the MUC1-CT tyrosine induced by fibroblast
growth factor-1 (FGF-1), which results in MUC1 binding to the heat
shock protein HSP90 chaperone and its consequent translocation to
the mitochondria (85). It is
still not fully understood how these proteins are targeted and
anchored to the MOM. However, it has been shown that Bcl-xL and
Bcl-2 integrate with MOM by their C-termini (84). In cancer cells, MUC1-C is also
imported into the nucleus, where it directly interacts with
specific transcription factors, such as nuclear factor-κB (NF-κB),
and stimulates their transcription (86-89). In addition, MUC1 hypoglycosylation
affects its subcellular localization via increased intracellular
MUC endocytic trafficking by clathrin-coated pits (10).
Kufe (83) and Li
et al (90) demonstrated
that MUC1-C participated in signal transmission from the
Wnt/β-catenin pathway to the nucleus. This was facilitated
following its interactions with p53, STAT3 and estrogen receptor α
(ERα) (91). The aforementioned
pathways are associated with oncogenesis (41). In addition, MUC1 can contribute to
constitutive stimulation of various processes. For example, Ahmad
et al (92) demonstrated
that MUC1-C interacted with TNF-R1 in mammary epithelial MCF-10A
cells and participated in TAK1-mediated phosphorylation of IκB
kinase β (IKKβ), formation of the IKKβ-inhibitor of NF-κB kinase
subunit γ (IKKγ) complex and autophosphorylation of IKKβ.
Furthermore, MUC1 can be regulated by hormone receptors, such as
ERα, nuclear retinitis pigmentosa GTPase regulator and the androgen
receptor. Estrogen treatment combined with MUC1 knockdown results
in increased cell death in aromatase inhibitor-resistant cells
(93).
4. Role of MUC1 in apoptosis
Several articles have been published regarding the
effects of MUC1 on apoptosis. In the present study, the diverse
interactions of MUC1 with different factors that lead to the
inhibition of this process were assessed (Fig. 3).
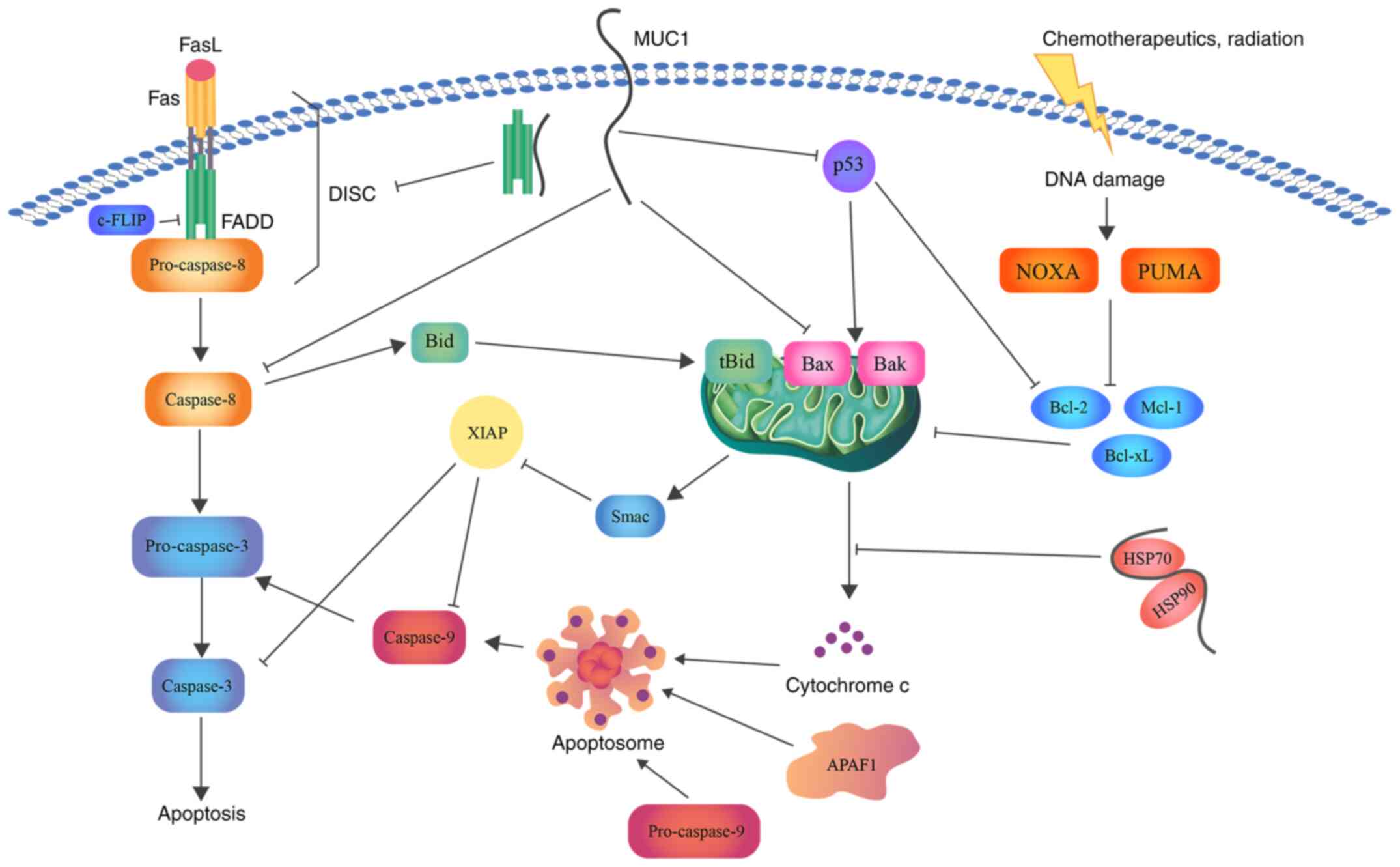 | Figure 3Schematic representation of the role
of MUC1 in apoptosis. MUC1 interacts with FADD DED, blocking the
formation of DISC and suppressing the induction of the extrinsic
apoptotic pathway. Direct association with caspase-8 inhibits its
activation. In addition, MUC1-C suppresses Bax translocation to the
MOM and cytochrome c release. Binding of MUC1 to the
HSP90/HSP70 complex weakens the activation of the mitochondrial
pathway. Moreover, direct binding of MUC1 to the p53 regulatory
domain is associated with stimulation of growth-arresting gene
transcription, thereby inhibiting apoptosis. Bcl, B-cell lymphoma;
Bax, Bcl-2-associated X protein; Bcl-xL, Bcl-extra large; MUC1,
mucin 1; FADD, FAS-associated with death domain; DED, death
effector domain; DISC, death inducing signaling complex; MUC1-C,
MUC1 C-terminal subunit; MOM, mitochondrial outer membrane; HSP,
heat shock protein; FasL, fatty acid synthetase ligand; FLIP, FLICE
(FADD-like IL-1β-converting enzyme)-inhibitory protein; Bid, BH3
interacting domain death agonist; Mcl-1, induced myeloid leukemia
cell differentiation protein; NOXA, NADPH oxidase activator; PUMA,
p53 upregulated modulator of apoptosis; Smac, second
mitochondria-derived activator of caspase; XIAP, X-linked inhibitor
of apoptosis protein. |
Role of MUC1 in the intrinsic apoptotic
pathway
The intrinsic (mitochondrial) pathway of apoptosis
is activated in response to various stimuli. It has been previously
mentioned that the treatment of cells with genotoxic anticancer
agents leads to the release of cytochrome c from the
mitochondria and the activation of the intrinsic pathway of
apoptosis (12,94). Ren et al (84) revealed that MUC1-C expression was
associated with decreased release of apoptogenic proteins,
including cytochrome c, Smac/DIABLO and AIF, as well as
attenuated activation of caspase-3 and PKC δ type (PKCδ) following
cisplatin treatment. It was shown that PKCδ may induce apoptosis in
lipopolysaccharide-activated macrophages via regulation of TNF
production (95). Similar results
were obtained when HCT116 colon carcinoma cancer cells were treated
with etoposide (84). Moreover,
the release of the aforementioned molecules was associated with
loss of the mitochondrial membrane potential. Therefore,
MUC1-C-dependent impairment of mitochondrial pro-apoptotic factor
release attenuates the apoptotic response, notably the intrinsic
pathway of apoptosis, in response to DNA damage. In contrast to
this finding, cisplatin treatment of A549 and ZR-75-1 cells with
transiently downregulated MUC1 expression increased the apoptotic
response (84).
The mitochondrial localization of MUC1 impacts
signal transduction from the cell membrane to the mitochondria,
leading to decreased activation of the intrinsic apoptotic pathway
(96,97). Bax plays a crucial role among
other members of the Bcl-2 protein family in the activation of the
mitochondrial pathway of apoptosis (98). During the apoptotic process, Bax
localizes from the cytosol to MOM, where it undergoes
conformational changes followed by oligomerization to form a pore
necessary for cytochrome c release (99). Bax is composed of nine α-helices.
The α2 helix contains the Bcl-2 homology BH3 domain, which is
essential for Bax homo- or hetero-dimerization with other
cysteine-containing proteins, such as the Bcl-2, Bcl-xL and Mcl
proteins (99,100). Ahmad et al (101) showed that MUC1 interacted
directly with Bax in breast and colon cancer cells. Two cysteines
present in the MUC1-C CQC motif participated in the formation of
MUC1-C heterodimers. Studies have demonstrated that the MUC1-C CQC
motif binds to Cys-62 in the BH3 domain of Bax. This interaction
may block the ability of Bax to dimerize. It is not currently known
whether MUC1-C binds to a secondary Bax BH3 cysteine residue at
position 126. It has been shown that oxidative stress may promote
the association between MUC1-C and Bax (101-103). In conclusion, the current data
indicate that MUC1-C attenuates Bax dimerization, translocation to
MOM and, in turn, the release of cytochrome c, which
suppresses the activation of the intrinsic apoptotic pathway.
Mutations in the MUC1-C CQC motif impair MUC1-C transport to the
nucleus and MOM. This mutation abolishes the oncogenic function of
MUC1-C and can be used for the development of molecules that may
block homodimerization and, in turn, MUC1-C signal transduction
(55).
Nickel acetate (Ni2+) is an agent with
genotoxic abilities (104,105). The study by Castorina and Giunta
(106) revealed that cell death
resistance of human bronchial epithelial cells (Beas-2B) exposed to
Ni2+ was associated with direct stimulation of the EGFR
by MUC1. This glycoprotein had the ability to activate EGFR/ERK1/2
signaling. Decreased levels of cleaved caspase-3 were also noted
(106). These results are
consistent with those reported in the study by Schroeder et
al (107), which examined
the ability of MUC1 to enhance EGF binding to the EGFR in breast
tumors. In non-small lung cancer, MUC1 promoted activation of the
PI3K/Akt pathway (108), which
was necessary for NF-κB signaling activation (109). Previous studies have shown that
the interaction of MUC1-C with IKKβ/IKKγ complex activates NF-κB
signaling (92,110). Moreover, Akt has demonstrated an
anti-apoptotic effect by inhibiting the functions of specific Bcl-2
proteins, such as Bad, which interact with chaperone protein
14-3-3, resulting in the release of specific anti-apoptotic
proteins (111,112). Therefore, it has been
hypothesized that chronic exposure to nickel compounds in
combination with increased MUC1 expression enhances epithelial cell
resistance to apoptosis and promotes the development of
carcinogenesis (106).
An additional study has revealed that MUC1
overexpression may protect cells from oxidative stress-induced
apoptosis (85). The abundance of
ROS, which are derived from the mitochondria, leads to the
activation of cell death pathways (113,114). Yin et al (115) observed an increase in MUC1
transcription and translation in vitro following exposure to
H2O2. MUC1-positive cells further
demonstrated higher levels of antioxidant enzymes. Since
H2O2 easily diffuses across the cell
membrane, it was hypothesized that its transmembrane subunit may be
associated with a decrease in ROS levels in the cells. These
results indicated that MUC1 inhibited the apoptotic response to
oxidative stress by decreasing the concentration levels of
oxidative molecules (85). The
increase in ROS levels following MUC1-C inhibition was recently
confirmed in mouse embryonic stem cells (116).
Moreover, MUC1 can interact with forkhead box class
O (FOXO)3a, which is also known as forkhead in
rhabdomyo-sarcoma-like 1. FOXO3a is a member of the FOXO family of
transcription factors that mediate gene transcription following
dephosphorylation in the nucleus (115,117). MUC1 decreases its
phosphorylation by attenuating activation of the PI3K/Akt pathway.
Downregulation of MUC1 in breast cancer cells causes inactivation
of FOXO3a, which increases the necrotic cell response to oxidative
stress (115). Therefore, MUC1
can play a role in protecting the epithelium from apoptosis
following injury (84).
The tumor suppressor p53 protein is a product of the
TP53 gene. This gene is located in humans at the short arm of
chromosome 17 (17p13.1) (118).
p53 plays a role in induction of the cell apoptotic response by
stimulation of Bax/Bak oligomerization or inhibition of
anti-apoptotic Bcl-2 family member function. Loss of p53 activity
prevents cells from forming a normal response to DNA damage or
stress (119). Wei et al
(120) reported that MUC1
inhibited the cellular response to DNA damage mediated by p53. MUC1
direct binding to the regulatory domain of p53 was associated with
enhanced transcription of growth arrest genes and in turn depletion
of apoptosis.
Additional evidence has confirmed that MUC1
interacts with small non-coding RNA molecules and microRNAs
(miR/miRNAs). It is known that miRNAs, such as miR-136, are
implicated in specific biological processes, such as the cell
cycle, proliferation, migration and apoptosis (121,122). For example, miR-136 may function
as a suppressor in the development of multiple cancer types and its
decreased levels in human glioma cells can stimulate apoptosis via
inhibition of astrocyte elevated gene-1 and Bcl-2 proteins. It was
demonstrated that miR-136 upregulation contributed to the induction
of apoptosis in esophageal squamous cell carcinoma cells via MUC1
inhibition (121). An additional
study indicated that miR-145 decreased ovarian cancer cell
proliferation and invasion by suppressing MUC1 (123). Moreover, breast cancer cells
with silenced MUC1 expression and overexpression of miR-485-5p
demonstrated inhibition of cell proliferation, invasiveness and
migration (124).
Role of MUC1 in the extrinsic apoptotic
pathway
Cleavage of the pro-apoptotic protein Bid is
mediated by caspase-8 and results in the formation of tBid, which
may induce the mitochondrial release of cytochrome c. This
allows the interaction between the receptor-mediated and
mitochondrial pathways (27,28,125). Therefore, the modulation of the
extrinsic pathway response by MUC1 is important. Ren et al
(84) demonstrated that in HCT116
cells MUC1 attenuated TRAIL-induced apoptosis. This effect was
reversed by the addition of cycloheximide. It has also been shown
that MUC1-C may inhibit caspase-8 activation induced by TRAIL,
TNF-α and FasL, and consequently block death-receptor signaling
(91). Caspase-8 is composed of
an N-terminal region containing two DEDs (1-183 aa). The p18
(217-374 aa) and p10 (385-480 aa) fragments are derived following
cleavage of each region. The association of the adaptor protein
FADD with caspase-8 via DEDs leads to caspase-8 dimerization and
its cleavage to the p18/p10 fragments (91,126). It has been shown that MUC1-C can
bind directly to caspase-8 p18 via its specific regions, which
contain the aa residues 270-322 and 1-20, respectively. This
binding occurs by interactions other than disulfide bonds. MUC1
binding to the p18 region may interrupt interdimer processing and
block caspase activation (91).
MUC1 competition with caspase-8 for binding to the
FADD DED may interrupt formation of DISC in vitro. The same
region of MUC1-C can bind to other protein partners, such as
β-catenin, IKKγ or the HSP90/HSP70 complex (40,92,127). Based on this evidence, Agata
et al (91) suggested that
MUC1-C may have both transmembrane receptor and chaperone-like
functions. Moreover, MCF-10A cells with downregulated MUC1
expression exhibited an increase in caspase-8 activity following
TNF-α, FasL and TRAIL stimulation in comparison to non-transformed
MCF-10A cells. Therefore, the ability of MUC1-C to attenuate
caspase-8-mediated activation of apoptosis could be used by normal
epithelial or malignant cells to protect them from cell death under
inflammatory conditions or to enable their survival in an adverse
environment (91).
Certain enzymes, such as c-Jun N-terminal kinase 1
(JNK1), are involved in the regulation of the apoptotic process.
Chen et al (128)
reported that MUC1-C (1-45 aa) directly binds to JNK1. JNK1 belongs
to the superfamily of MAP kinases (129). The three following isoforms of
JNK have been identified: JNK1, JNK2 and JNK3. The first two are
expressed in a variety of tissues, whereas JNK3 is mainly limited
to the neurons and heart (130).
The JNK signaling pathway can be activated by different stimuli,
including genotoxic agents, TNF-α, MAP kinase 4 (MKK4) or MKK7.
Following its activation, JNK1 localizes to the nucleus and
phosphorylates the effector protein c-Jun, which in turn influences
transcription of multiple target genes, including activator protein
1. MUC1 overexpression contributes to increased activation of JNK1
and its target c-Jun following treatment of the cells with
genotoxic anticancer agents, such as cisplatin or doxorubicin
(128). This process decreases
the cellular response to apoptotic stimuli (128).
Several other studies have also confirmed the
significant role of MUC1 in apoptosis. Zhang et al (131) demonstrated that cell
proliferation, invasion, migration, epithelial-mesenchymal
transition and apoptosis in oral squamous cell carcinoma (OSCC) may
be affected by the activity of specific transcription factors, such
as Snail and Slug. Slug is an invasion-promoting factor that plays
a major role in the inhibition of E-cadherin transcription and the
repression of the function of the pro-apoptotic protein PUMA, which
in turn leads to the induction of cell survival (132,133). It has been shown that MUC1
expression in OSCC is positively correlated with the expression of
Slug, whereas MUC1 gene silencing is correlated with a decrease in
Slug levels. Therefore, is has been suggested that MUC1 silencing
is associated with the induction of apoptosis and the inhibition of
cell proliferation, invasion and migration via downregulation of
Slug expression (131).
The JAK/STAT signaling pathway plays an important
role in transferring signals from cell membrane receptors to the
nucleus (134). Overexpression
of MUC1 was associated with decreased caspase-3 activation,
resulting in a decreased apoptotic response in the irradiated
hepatocellular carcinoma (HCC) SMMC-7721 cell line. An increase in
the expression levels of the anti-apoptotic proteins Mcl-1 and
Bcl-xL was also observed. Yi and Lu (135) reported that resistance to
irradiation-induced apoptosis was associated with JAK2/STAT3
signaling pathway activation by MUC1. Therefore, this protein
contributed to the radioresistance of the HCC cells. Escher et
al (93) demonstrated the
role of MUC1 in enhancing the activity of JAK/STAT, which
stimulated IFN-induced transmembrane protein 1 expression and an
aggressive phenotype in breast cancer cells resistant to aromatase
inhibitors.
Kato et al (136) demonstrated that cells treated
with polyinosinic:polycytidilic acid exhibited increased activation
of caspase-3 and -8, IFN regulatory factor 3, NF-κB and IFN-β
following MUC1 silencing in comparison to MUC1-expressing cells.
This study further showed that MUC1-CT attenuated Toll-like
receptor 3 (TRL3)-induced apoptosis in lung epithelial cells by
blocking the interaction between TRL3 and TIR-domain-containing
adapter-inducing IFN-β (136).
Moreover, inhibition of MUC1 significantly increased the
sensitivity of lung and pancreatic cancer cells to the induction of
apoptosis by anticancer drugs (137,138).
Role of MUC1 in anoikis
Loss of cell adhesion to the surrounding matrix or
its inappropriate adherence results in the activation of a specific
type of apoptosis, termed anoikis (139). Cell resistance to anoikis is a
biological process that precedes metastasis. Despite the current
understanding of the apoptotic process, the mechanism by which
metastatic cancer cells evade anoikis remains poorly defined. It
has been previously shown that MUC1 overexpression, which occurs
mainly in the ECD, blocks anoikis activation. This effect may be
associated with extensive glycosylation, which forms a specific
microenvironment on the cell surface and protects from activation
of anoikis-initiating factors and death receptors, such as
integrins and Fas, respectively (32,33). One of the key enzymes in the
O-glycosylation process is C1GalT1; its deficiency leads to the
formation of abnormal shortened forms of O-linked sugar chains and
is associated with increased availability of cell surface receptors
for integrin1β, E-cadherin or FasL. These molecular events result
in activation of the extracellular pathway of apoptosis (31,52). Piyush et al (140) demonstrated that suppression of
C1GalT1 expression resulted in increased expression of the Tn
antigen in MUC1-negative HCT116 cells (human colon cancer).
However, activation of anoikis with a concomitant increase in
caspase-8 activity due to binding of FasL to Fas was only noted in
MUC1-positive cells (SW620). Therefore, an evident association
between excessive O-glycosylation of MUC1 and anoikis resistance
was observed, primarily due to inhibition of anoikis-initiating
molecule activation.
5. Conclusions
MUC1 is a component of mucus that plays a protective
role in normal epithelial cells. However, during malignant
transformation, the changes in the expression and glycosylation
pattern of MUC1 modulate its interactions with other proteins,
which in turn regulate signal transmission. High levels of MUC1 are
correlated with a poor prognosis and shorter survival time in
patients with cancer. In addition, aberrant expression of this
protein may block drug diffusion through the cell membrane and
promote survival of cancerous cells, since it has been shown to
impact both extrinsic and intrinsic apoptotic pathways. Therefore,
the upregulation of MUC1-dependent attenuation of apoptotic
response indicates the potential role of this protein in cancer
therapy. Despite extensive evidence reported on the mechanism of
action of MUC1 with regard to cell death, a number of aspects
remain unresolved. Therefore, additional studies are necessary to
further elucidate such interactions.
Availability of data and materials
Not applicable.
Authors' contributions
KS developed the concept for the study and drafted
the manuscript. IR reviewed and edited the manuscript. All authors
read and approved the final manuscript. Data authentication is not
applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Abbreviations:
|
AIF
|
apoptosis-inducing factor
|
|
C1GalT1
|
N-acetylgalactosamine
β-1,3-galactosyltransferase (core 1 synthase)
|
|
CQC
|
cysteine-glutamine-cysteine motif
|
|
DED
|
death effector domain
|
|
DISC
|
death inducing signaling complex
|
|
ERα
|
estrogen receptor α
|
|
FADD/MORT1
|
FAS-associated with death domain
|
|
FasL
|
fatty acid synthase ligand
|
|
FLIP
|
FLICE (FADD-like IL-1β-converting
enzyme)-inhibitory protein
|
|
FOXO
|
forkhead box class O family of
transcription factors
|
|
FOXO3a
|
forkhead box class O3a
|
|
HCC
|
hepatocellular carcinoma
|
|
Gal
|
galactose
|
|
HSP90
|
heat shock protein 90
|
|
IKKβ
|
IκB kinase β
|
|
JAK/STAT
|
Janus kinase/signal transducers and
activators of transcription
|
|
MKK
|
mitogen-activated protein kinase
kinase
|
|
MOM
|
mitochondrial outer membrane
|
|
MUC1
|
mucin 1
|
|
OSCC
|
oral squamous cell carcinoma
|
|
PKC
|
protein kinase C
|
|
PKCδ
|
PKC δ type
|
|
ROS
|
reactive oxygen species
|
|
RTKs
|
receptor tyrosine kinases
|
|
Smac/DIABLO
|
second mitochondria-derived activator
of caspase/direct inhibitor of apoptosis-binding protein with low
pI
|
|
Src
|
non-receptor tyrosine kinase
|
|
TRADD
|
TNF receptor type 1-associated death
domain protein
|
|
TRAIL/Apo2L
|
tumor necrosis factor-related
apoptosis-inducing ligand
|
|
VNTR
|
variable number of tandem repeats
region
|
References
|
1
|
Behera SK, Praharaj AB, Dehury B and Negi
S: Exploring the role and diversity of mucins in health and disease
with special insight into non-communicable diseases. Glyconconj J.
32:575–613. 2015. View Article : Google Scholar
|
|
2
|
Dhanisha SS, Guruvayoorappan C, Drishya S
and Abeesh P: Mucins: Structural diversity, biosynthesis, its role
in pathogenesis and as possible therapeutic targets. Crit Rev Oncol
Hematol. 122:98–122. 2018. View Article : Google Scholar
|
|
3
|
Cornick S, Tawiah A and Chadee K: Roles
and regulation of the mucus barrier in the gut. Tissue Barriers.
3:e9824262015. View Article : Google Scholar
|
|
4
|
Moniaux N, Escande F, Porchet N, Aubert JP
and Batra SK: Structural organization and classification of the
human mucin genes. Front Biosci. 6:D1192–D1206. 2001. View Article : Google Scholar
|
|
5
|
Nath S and Mukherjee P: MUC1: A
multifaceted oncoprotein with a key role in cancer progression.
Trends Mol Med. 20:332–342. 2014. View Article : Google Scholar
|
|
6
|
Hanson RL and Hollingsworth MA: Functional
consequences of differential O-glycosylation of MUC1, MUC4, and
MUC16 (downstream effects on signaling). Biomolecules. 6:342016.
View Article : Google Scholar
|
|
7
|
Jonckheere N and Van Seuningen I: The
membrane-bound mucins: How large O-glycoproteins play key roles in
epithelial cancers and hold promise as biological tools for
gene-based and immunotherapies. Crit Rev Oncog. 14:177–196. 2008.
View Article : Google Scholar
|
|
8
|
Lau SK, Weiss LM and Chu PG: Differential
expression of MUC1, MUC2, and MUC5AC in carcinomas of various
sites: An immunohistochemical study. Am J Clin Pathol. 122:61–69.
2004. View Article : Google Scholar
|
|
9
|
Reynolds IS, Fichtner M, McNamara DA, Kay
EW, Prehn JHM and Burke JP: Mucin glycoproteins block apoptosis;
promote invasion, proliferation, and migration; and cause
chemoresistance through diverse pathways in epithelial cancers.
Cancer Metastasis Rev. 38:237–257. 2019. View Article : Google Scholar
|
|
10
|
Altschuler Y, Kinlough CL, Poland PA,
Bruns JB, Apodaca G, Weisz OA and Hughey RP: Clathrin-mediated
endocytosis of MUC1 is modulated by its glycosylation state. Mol
Biol Cell. 11:819–831. 2000. View Article : Google Scholar
|
|
11
|
Katoch B, Sebastian S, Sahdev S, Padh H,
Hasnain SE and Begum R: Programmed cell death and its clinical
implications. Indian J Exp Biol. 40:513–524. 2002.
|
|
12
|
Elmore S: Apoptosis: A review of
programmed cell death. Toxicol Pathol. 35:495–516. 2007. View Article : Google Scholar
|
|
13
|
D'Arcy MS: Cell death: A review of the
major forms of apoptosis, necrosis and autophagy. Cell Biol Int.
43:582–592. 2019. View Article : Google Scholar
|
|
14
|
Wong RS: Apoptosis in cancer: From
pathogenesis to treatment. J Exp Clin Cancer Res. 30:872011.
View Article : Google Scholar
|
|
15
|
Jan R and Chaudhry GE: Understanding
apoptosis and apoptotic pathways targeted cancer therapeutics. Adv
Pharm Bull. 9:205–218. 2019. View Article : Google Scholar
|
|
16
|
Papaliagkas V, Anogianaki A, Anogianakis G
and Ilonidis G: The proteins and the mechanisms of apoptosis: A
mini-review of the fundamentals. Hippokratia. 11:108–113. 2007.
|
|
17
|
Taylor R, Cullen S and Martin S:
Apoptosis: Controlled demolition at the cellular level. Nat Rev Mol
Cell Biol. 9:231–241. 2008. View Article : Google Scholar
|
|
18
|
Shakeri R, Kheirollahi A and Davoodi J:
Apaf-1: Regulation and function in cell death. Biochimie.
135:111–125. 2017. View Article : Google Scholar
|
|
19
|
Shimizu S, Narita M and Tsujimoto Y: Bcl-2
family proteins regulate the release of apoptogenic cytochrome c by
the mitochondrial channel VDAC. Nature. 399:483–487. 1999.
View Article : Google Scholar
|
|
20
|
Savitskaya MA and Onishchenko GE:
Mechanisms of apoptosis. Biochemistry (Mosc). 80:1393–1405. 2015.
View Article : Google Scholar
|
|
21
|
O'Brien MA and Kirby R: Apoptosis: A
review of pro-apoptotic and anti-apoptotic pathways and
dysregulation in disease. J Vet Emerg Crit Care (San Antonio).
18:572–585. 2008. View Article : Google Scholar
|
|
22
|
Julien O and Wells JA: Caspases and their
substrates. Cell Death Differ. 24:1380–1389. 2017. View Article : Google Scholar
|
|
23
|
Porter AG and Jänicke RU: Emerging roles
of caspase-3 in apoptosis. Cell Death Differ. 6:99–104. 1999.
View Article : Google Scholar
|
|
24
|
Van Ba H and Hwang I: Role of caspase-9 in
the effector caspases and genome expressions, and growth of bovine
skeletal myoblasts. Dev Growth Differ. 56:131–142. 2014. View Article : Google Scholar
|
|
25
|
Dempsey PW, Doyle SE, He JQ and Cheng G:
The signaling adaptors and pathways activated by TNF superfamily.
Cytokine Growth Factor Rev. 14:193–209. 2003. View Article : Google Scholar
|
|
26
|
Srivastava RK: TRAIL/Apo-2L: Mechanisms
and clinical applications in cancer. Neoplasia. 3:535–546. 2001.
View Article : Google Scholar
|
|
27
|
Li H, Zhu H, Xu CJ and Yuan J: Cleavage of
BID by caspase 8 mediates the mitochondrial damage in the Fas
pathway of apoptosis. Cell. 94:491–501. 1998. View Article : Google Scholar
|
|
28
|
Luo X, Budihardjo I, Zou H, Slaughter C
and Wang X: Bid, a Bcl2 interacting protein, mediates cytochrome c
release from mitochondria in response to activation of cell surface
death receptors. Cell. 94:481–490. 1998. View Article : Google Scholar
|
|
29
|
Valentijn AJ and Gilmore AP: Translocation
of full-length Bid to mitochondria during anoikis. J Biol Chem.
279:32848–32857. 2004. View Article : Google Scholar
|
|
30
|
Woods NT, Yamaguchi H, Lee FY, Bhalla KN
and Wang HG: Anoikis, initiated by Mcl-1 degradation and Bim
induction, is deregulated during oncogenesis. Cancer Res.
67:10744–10752. 2007. View Article : Google Scholar
|
|
31
|
Zhao Q, Piyush T, Chen C, Hollingsworth
MA, Hilkens J, Rhodes JM and Yu LG: MUC1 extracellular domain
confers resistance of epithelial cancer cells to anoikis. Cell
Death Dis. 5:e14382014. View Article : Google Scholar
|
|
32
|
Kim YN, Koo KH, Sung JY, Yun UJ and Kim H:
Anoikis resistance: An essential prerequisite for tumor metastasis.
Int J Cell Biol. 2012:3068792012. View Article : Google Scholar
|
|
33
|
Paoli P, Giannoni E and Chiarugi P:
Anoikis molecular pathways and its role in cancer progression.
Biochim Biophys Acta. 1833:3481–3498. 2013. View Article : Google Scholar
|
|
34
|
Yang MC, Lin RW, Huang SB, Huang SY, Chen
WJ, Wang S, Hong YR and Wang C: Bim directly antagonizes Bcl-xl in
doxorubicin-induced prostate cancer cell apoptosis independently of
p53. Cell Cycle. 15:394–402. 2016. View Article : Google Scholar
|
|
35
|
Aoudjit F and Vuori K: Matrix attachment
regulates Fas-induced apoptosis in endothelial cells: A role for
c-flip and implications for anoikis. J Cell Biol. 152:633–643.
2001. View Article : Google Scholar
|
|
36
|
Marconi A, Atzei P, Panza C, Fila C,
Tiberio R, Truzzi F, Wachter T, Leverkus M and Pincelli C:
FLICE/caspase-8 activation triggers anoikis induced by
beta1-integrin blockade in human keratinocytes. J Cell Sci.
117:5815–5823. 2004. View Article : Google Scholar
|
|
37
|
Hattrup CL and Gendler SJ: Structure and
function of the cell surface (tethered) mucins. Annu Rev Physiol.
70:431–457. 2008. View Article : Google Scholar
|
|
38
|
Singh PK and Hollingsworth MA: Cell
surface-associated mucins in signal transduction. Trends Cell Biol.
16:467–476. 2006. View Article : Google Scholar
|
|
39
|
Hagiwara M, Yasumizu Y, Yamashita N,
Rajabi H, Fushimi A, Long MD, Li W, Bhattacharya A, Ahmad R, Oya M,
et al: MUC1-C Activates the BAF (mSWI/SNF) complex in prostate
cancer stem cells. Cancer Res. 81:1111–1122. 2021. View Article : Google Scholar
|
|
40
|
Hanson JM, BroweIl DA, Cunliffe WJ, Varma
J, Allen A, Hemming D, Shenton BK, Young JR, Higgs MJ, Brotherick I
and Pearson JP: MUC1 expression in primary breast cancer: The
effect of tamoxifen treatment. Breast Cancer Res Treat. 67:215–222.
2001. View Article : Google Scholar
|
|
41
|
Kufe DW: Mucins in cancer: Function,
prognosis and therapy. Nat Rev Cancer. 9:874–885. 2009. View Article : Google Scholar
|
|
42
|
Raina D, Ahmad R, Rajabi H, Panchamoorthy
G, Kharbanda S and Kufe D: Targeting cysteine-mediated dimerization
of the MUC1-C oncoprotein in human cancer cells. Int J Oncol.
40:1643–1649. 2012.
|
|
43
|
Yang J: Identification of novel
biomarkers, MUC5AC, MUC1, KRT7, GAPDH, CD44 for gastric cancer. Med
Oncol. 37:342020. View Article : Google Scholar
|
|
44
|
Apostolopoulos V, Stojanovska L and
Gargosky SE: MUC1 (CD227): A multi-tasked molecule. Cell Mol Life
Sci. 72:4475–4500. 2015. View Article : Google Scholar
|
|
45
|
Taylor-Papadimitriou J: Report on the
first international workshop on carcinoma-associated mucins. Int J
Cancer. 49:1–5. 1991. View Article : Google Scholar
|
|
46
|
Gendler SJ and Spicer AP: Epithelial mucin
genes. Annu Rev Physiol. 57:607–634. 1995. View Article : Google Scholar
|
|
47
|
Hanisch FG and Müller S: MUC1: The
polymorphic appearance of a human mucin. Glycobiology. 10:439–449.
2000. View Article : Google Scholar
|
|
48
|
Agrawal B, Krantz MJ, Parker J and
Longenecker BM: Expression of MUC1 mucin on activated human T
cells: Implications for a role of MUC1 in normal immune regulation.
Cancer Res. 58:4079–4081. 1998.
|
|
49
|
Dent GA, Civalier CJ, Brecher ME and
Bentley SA: MUC1 expression in hematopoietic tissues. Am J Clin
Pathol. 111:741–747. 1999. View Article : Google Scholar
|
|
50
|
Franke FE, Kraus S, Eiermann C, Pauls K,
Lalani EN and Bergmann M: MUC1 in normal and impaired
spermatogenesis. Mol Hum Reprod. 7:505–512. 2001. View Article : Google Scholar
|
|
51
|
Seo JT, Lee JS, Jun JH and Yang MH:
Expression of mucin genes in the human testis and its relationship
to spermatogenesis. Yonsei Med J. 46:667–672. 2005. View Article : Google Scholar
|
|
52
|
Cascio S and Finn OJ: Intra- and
extra-cellular events related to altered glycosylation of MUC1
promote chronic inflammation, tumor progression, invasion, and
metastasis. Biomolecules. 6:392016. View Article : Google Scholar
|
|
53
|
Bennett EP, Mandel U, Clausen H, Gerken
TA, Fritz TA and Tabak LA: Control of mucin-type O-glycosylation: A
classification of the polypeptide GalNAc-transferase gene family.
Glycobiology. 22:736–756. 2012. View Article : Google Scholar
|
|
54
|
Taherali F, Varum F and Basit AW: A
slippery slope: On the origin, role and physiology of mucus. Adv
Drug Deliv Rev. 124:16–33. 2018. View Article : Google Scholar
|
|
55
|
Raina D, Agarwal P, Lee J, Bharti A,
McKnight CJ, Sharma P, Kharbanda S and Kufe D: Characterization of
the MUC1-C cytoplasmic domain as a cancer target. PLoS One.
10:e01351562015. View Article : Google Scholar
|
|
56
|
McAuley JL, Corcilius L, Tan HX, Payne RJ,
McGuckin MA and Brown LE: The cell surface mucin MUC1 limits the
severity of influenza A virus infection. Mucosal Immunol.
10:1581–1593. 2017. View Article : Google Scholar
|
|
57
|
Thathiah A, Blobel CP and Carson DD: Tumor
necrosis factor-alpha converting enzyme/ADAM 17 mediates MUC1
shedding. J Biol Chem. 278:3386–3394. 2003. View Article : Google Scholar
|
|
58
|
Tarp MA and Clausen H: Mucin-type
O-glycosylation and its potential use in drug and vaccine
development. Biochim Biophys Acta. 1780:546–563. 2008. View Article : Google Scholar
|
|
59
|
Awaya H, Takeshima Y, Yamasaki M and Inai
K: Expression of MUC1, MUC2, MUC5AC, and MUC6 in atypical
adenomatous hyperplasia, bronchioloalveolar carcinoma,
adenocarcinoma with mixed subtypes, and mucinous bronchioloalveolar
carcinoma of the lung. Am J Clin Pathol. 121:644–653. 2004.
View Article : Google Scholar
|
|
60
|
Horm TM and Schroeder JA: MUC1 and
metastatic cancer: Expression, function and therapeutic targeting.
Cell Adh Migr. 7:187–198. 2013. View Article : Google Scholar
|
|
61
|
Krishn SR, Kaur S, Smith LM, Johansson SL,
Jain M, Patel A, Gautam SK, Hollingsworth MA, Mandel U, Clausen H,
et al: Mucins and associated glycan signatures in colon
adenoma-carcinoma sequence: Prospective pathological implication(s)
for early diagnosis of colon cancer. Cancer Lett. 374:304–314.
2016. View Article : Google Scholar
|
|
62
|
Singh AP, Chauhan SC, Bafna S, Johansson
SL, Smith LM, Moniaux N, Lin MF and Batra SK: Aberrant expression
of transmembrane mucins, MUC1 and MUC4, in human prostate
carcinomas. Prostate. 66:421–429. 2006. View Article : Google Scholar
|
|
63
|
Gao Y, Liu Z, Feng J, Sun Q, Zhang B,
Zheng W and Ma W: Expression pattern of polypeptide
N-acetylgalactosaminyltransferase-10 in gastric carcinoma. Oncol
Lett. 5:113–116. 2013. View Article : Google Scholar
|
|
64
|
Guda K, Moinova H, He J, Jamison O, Ravi
L, Natale L, Lutterbaugh J, Lawrence E, Lewis S, Willson JK, et al:
Inactivating germ-line and somatic mutations in polypeptide
N-acetylgalactosaminyltransferase 12 in human colon cancers. Proc
Natl Acad Sci USA. 106:12921–12925. 2009. View Article : Google Scholar
|
|
65
|
Stowell SR, Ju T and Cummings RD: Protein
glycosylation in cancer. Annu Rev Pathol. 10:473–510. 2015.
View Article : Google Scholar
|
|
66
|
Liesche F, Kölbl AC, Ilmer M, Hutter S,
Jeschke U and Andergassen U: Role of
N-acetylgalactosaminyltransferase 6 in early tumorigenesis and
formation of metastasis. Mol Med Rep. 13:4309–4314. 2016.
View Article : Google Scholar
|
|
67
|
Hakomori S: Aberrant glycosylation in
cancer cell membranes as focused on glycolipids: Overview and
perspectives. Cancer Res. 45:2405–2414. 1985.
|
|
68
|
Radziejewska I, Supruniuk K, Nazaruk J,
Karna E, Popławska B, Bielawska A and Galicka A: Rosmarinic acid
influences collagen, MMPs, TIMPs, glycosylation and MUC1 in
CRL-1739 gastric cancer cell line. Biomed Pharmacother.
107:397–407. 2018. View Article : Google Scholar
|
|
69
|
Syrkina MS, Maslakova AA, Potashnikova DM,
Veiko VP, Vassetzky YS and Rubtsov MA: Dual role of the
extracellular domain of human mucin MUC1 in metastasis. J Cell
Biochem. 118:4002–4011. 2017. View Article : Google Scholar
|
|
70
|
Ho WL, Hsu WM, Huang MC, Kadomatsu K and
Nakagawara A: Protein glycosylation in cancers and its potential
therapeutic applications in neuroblastoma. J Hematol Oncol.
9:1002016. View Article : Google Scholar
|
|
71
|
Liu B, Pan S, Xiao Y, Liu Q, Xu J and Jia
L: LINC01296/miR-26a/GALNT3 axis contributes to colorectal cancer
progression by regulating O-glycosylated MUC1 via PI3K/AKT pathway.
J Exp Clin Cancer Res. 37:3162018. View Article : Google Scholar
|
|
72
|
Mao Y, Zhang Y, Fan S, Chen L, Tang L,
Chen X and Lyu J: GALNT6 promotes tumorigenicity and metastasis of
breast cancer cell via β-catenin/MUC1-C signaling pathway. Int J
Biol Sci. 15:169–182. 2019. View Article : Google Scholar
|
|
73
|
Bäckström M, Thomsson KA, Karlsson H and
Hansson GC: Sensitive liquid chromatography-electrospray mass
spectrometry allows for the analysis of the O-glycosylation of
immunoprecipitated proteins from cells or tissues: Application to
MUC1 glycosylation in cancer. J Proteome Res. 8:538–545. 2009.
View Article : Google Scholar
|
|
74
|
Blanas A, Sahasrabudhe NM, Rodríguez E,
van Kooyk Y and van Vliet SJ: Fucosylated antigens in cancer: An
alliance toward tumor progression, metastasis, and resistance to
chemotherapy. Front Oncol. 8:392018. View Article : Google Scholar
|
|
75
|
Jia L, Zhang J, Ma T, Guo Y, Yu Y and Cui
J: The Function of Fucosylation in Progression of Lung Cancer.
Front Oncol. 8:5652018. View Article : Google Scholar
|
|
76
|
Chen Z, Gulzar ZG, St Hill CA, Walcheck B
and Brooks JD: Increased expression of GCNT1 is associated with
altered O-glycosylation of PSA, PAP, and MUC1 in human prostate
cancers. Prostate. 74:1059–1067. 2014. View Article : Google Scholar
|
|
77
|
Nakamori S, Kameyama M, Imaoka S, Furukawa
H, Ishikawa O, Sasaki Y, Kabuto T, Iwanaga T, Matsushita Y and
Irimura T: Increased expression of sialyl Lewisx antigen correlates
with poor survival in patients with colorectal carcinoma:
Clinicopathological and immunohistochemical study. Cancer Res.
53:3632–3637. 1993.
|
|
78
|
Ricardo S, Marcos-Silva L, Valente C,
Coelho R, Gomes R and David L: Mucins MUC16 and MUC1 are major
carriers of SLe(a) and SLe(x) in borderline and malignant serous
ovarian tumors. Virchows Arch. 468:715–722. 2016. View Article : Google Scholar
|
|
79
|
Mori Y, Akita K, Yashiro M, Sawada T,
Hirakawa K, Murata T and Nakada H: Binding of galectin-3, a
β-galactoside-binding lectin, to MUC1 protein enhances
phosphorylation of extracellular signal-regulated kinase 1/2
(ERK1/2) and Akt, promoting tumor cell malignancy. J Biol Chem.
290:26125–26140. 2015. View Article : Google Scholar
|
|
80
|
Ramasamy S, Duraisamy S, Barbashov S,
Kawano T, Kharbanda S and Kufe D: The MUC1 and galectin-3
oncoproteins function in a microRNA-dependent regulatory loop. Mol
Cell. 27:992–1004. 2007. View Article : Google Scholar
|
|
81
|
Sciacchitano S, Lavra L, Morgante A,
Ulivieri A, Magi F, De Francesco GP, Bellotti C, Salehi LB and
Ricci A: Galectin-3: One molecule for an alphabet of diseases, from
A to Z. Int J Mol Sci. 19:3792018. View Article : Google Scholar
|
|
82
|
Zhao Q, Guo X, Nash GB, Stone PC, Hilkens
J, Rhodes JM and Yu LG: Circulating galectin-3 promotes metastasis
by modifying MUC1 localization on cancer cell surface. Cancer Res.
69:6799–6806. 2009. View Article : Google Scholar
|
|
83
|
Kufe DW: MUC1-C oncoprotein as a target in
breast cancer: Activation of signaling pathways and therapeutic
approaches. Oncogene. 32:1073–1081. 2013. View Article : Google Scholar
|
|
84
|
Ren J, Agata N, Chen D, Li Y, Yu WH, Huang
L, Raina D, Chen W, Kharbanda S and Kufe D: Human MUC1
carcinoma-associated protein confers resistance to genotoxic
anticancer agents. Cancer Cell. 5:163–175. 2004. View Article : Google Scholar
|
|
85
|
Yin L and Kufe D: Human MUC1 carcinoma
antigen regulates intracellular oxidant levels and the apoptotic
response to oxidative stress. J Biol Chem. 278:35458–35464. 2003.
View Article : Google Scholar
|
|
86
|
Baldwin AS: Control of oncogenesis and
cancer therapy resistance by the transcription factor NF-κB. J Clin
Invest. 107:241–246. 2001. View Article : Google Scholar
|
|
87
|
Dyomin VG, Palanisamy N, Lloyd KO, Dyomina
K, Jhanwar SC, Houldsworth J and Chaganti RS: MUC1 is activated in
a B-cell lymphoma by the t(1;14)(q21;q32) translocation and is
rearranged and amplified in B-cell lymphoma subsets. Blood.
95:2666–2671. 2000. View Article : Google Scholar
|
|
88
|
Nakshatri H, Bhat-Nakshatri P, Martin DA,
Goulet RJ Jr and Sledge GW Jr: Constitutive activation of NF-κB
during progression of breast cancer to hormone-independent growth.
Mol Cell Biol. 17:3629–3639. 1997. View Article : Google Scholar
|
|
89
|
Stroopinsky D, Rosenblatt J, Ito K, Mills
H, Yin L, Rajabi H, Vasir B, Kufe T, Luptakova K, Arnason J, et al:
MUC1 is a potential target for the treatment of acute myeloid
leukemia stem cells. Cancer Res. 73:5569–5579. 2013. View Article : Google Scholar
|
|
90
|
Li Y, Yu WH, Ren J, Huang L, Kharbanda S,
Loda M and Kufe D: Heregulin targets γ-catenin to the nucleolus by
a mechanism dependent on the DF3/MUC1 protein. Mol Cancer Res.
1:765–775. 2003.
|
|
91
|
Agata N, Ahmad R, Kawano T, Raina D,
Kharbanda S and Kufe D: MUC1 oncoprotein blocks death
receptor-mediated apoptosis by inhibiting recruitment of caspase-8.
Cancer Res. 68:6136–6144. 2008. View Article : Google Scholar
|
|
92
|
Ahmad R, Raina D, Trivedi V, Ren J, Rajabi
H, Kharbanda S and Kufe D: MUC1 oncoprotein activates the IkappaB
kinase beta complex and constitutive NF-kappaBsignalling. Nat Cell
Biol. 9:1419–1427. 2007. View Article : Google Scholar
|
|
93
|
Escher TE, Lui AJ, Geanes ES, Walter KR,
Tawfik O, Hagan CR and Lewis-Wambi J: Interaction between MUC1 and
STAT1 drives IFITM1 overexpression in aromatase inhibitor-resistant
breast cancer cells and mediates estrogen-induced apoptosis. Mol
Cancer Res. 17:1180–1194. 2019. View Article : Google Scholar
|
|
94
|
Pistritto G, Trisciuoglio D, Ceci C,
Garufi A and D'Orazi G: Apoptosis as anticancer mechanism: Function
and dysfunction of its modulators and targeted therapeutic
strategies. Aging (Albany NY). 8:603–619. 2016. View Article : Google Scholar
|
|
95
|
Comalada M, Xaus J, Valledor AF,
López-López C, Pennington DJ and Celada A: PKC epsilon is involved
in JNK activation that mediates LPS-induced TNF-alpha, which
induces apoptosis in macrophages. Am J Physiol Cell Physiol.
285:C1235–1245. 2003. View Article : Google Scholar
|
|
96
|
Rajabi H and Kufe D: MUC1-C oncoprotein
integrates a program of EMT, epigenetic reprogramming and immune
evasion in human carcinomas. Biochim Biophys Acta Rev Cancer.
1868:117–122. 2017. View Article : Google Scholar
|
|
97
|
Rajabi H, Hiraki M and Kufe D: MUC1-C
activates polycomb complexes and downregulates tumor suppressor
genes in human cancer cells. Oncogene. 37:2079–2088. 2018.
View Article : Google Scholar
|
|
98
|
Wei MC, Zong WX, Cheng EH, Lindsten T,
Panoutsakopoulou V, Ross AJ, Roth KA, MacGregor GR, Thompson CB and
Korsmeyer SJ: Proapoptotic BAX and BAK: A requisite gateway to
mitochondrial dysfunction and death. Science. 292:727–730. 2001.
View Article : Google Scholar
|
|
99
|
Westphal D, Dewson G, Czabotar PE and
Kluck RM: Molecular biology of Bax and Bak activation and action.
Biochim Biophys Acta. 1813:521–531. 2011. View Article : Google Scholar
|
|
100
|
Peña-Blanco A and García-Sáez AJ: Bax, Bak
and beyond - mitochondrial performance in apoptosis. FEBS J.
285:416–431. 2018. View Article : Google Scholar
|
|
101
|
Ahmad R, Alam M, Rajabi H and Kufe D: The
MUC1-C oncoprotein binds to the BH3 domain of the pro-apoptotic BAX
protein and blocks BAX function. J Biol Chem. 287:20866–20875.
2012. View Article : Google Scholar
|
|
102
|
D'Alessio M, De Nicola M, Coppola S,
Gualandi G, Pugliese L, Cerella C, Cristofanon S, Civitareale P,
Ciriolo MR, Bergamaschi A, et al: Oxidative Bax dimerization
promotes its translocation to mitochondria independently of
apoptosis. FASEB J. 19:1504–1506. 2005. View Article : Google Scholar
|
|
103
|
Leng Y, Cao C, Ren J, Huang L, Chen D, Ito
M and Kufe D: Nuclear import of the MUC1-C oncoprotein is mediated
by nucleoporin Nup62. J Biol Chem. 282:19321–19330. 2007.
View Article : Google Scholar
|
|
104
|
Cangul H, Broday L, Salnikow K, Sutherland
J, Peng W, Zhang Q, Poltaratsky V, Yee H, Zoroddu MA and Costa M:
Molecular mechanisms of nickel carcinogenesis. Toxicol Lett.
127:69–75. 2002. View Article : Google Scholar
|
|
105
|
Lu H, Shi X, Costa M and Huang C:
Carcinogenic effect of nickel compounds. Mol Cell Biochem.
279:45–67. 2005. View Article : Google Scholar
|
|
106
|
Castorina A and Giunta S: Mucin 1 (MUC1)
signalling contributes to increase the resistance to cell death in
human bronchial epithelial cells exposed to nickel acetate.
Biometals. 27:1149–1158. 2014. View Article : Google Scholar
|
|
107
|
Schroeder JA, Masri AA, Adriance MC,
Tessier JC, Kotlarczyk KL, Thompson MC and Gendler SJ: MUC1
overexpression results in mammary gland tumorigenesis and prolonged
alveolar differentiation. Oncogene. 23:5739–5747. 2004. View Article : Google Scholar
|
|
108
|
Raina D, Kosugi M, Ahmad R, Panchamoorthy
G, Rajabi H, Alam M, Shimamura T, Shapiro GI, Supko J, Kharbanda S
and Kufe D: Dependence on the MUC1-C oncoprotein in non-small cell
lung cancer cells. Mol Cancer Ther. 10:806–816. 2011. View Article : Google Scholar
|
|
109
|
Andjelic S, Hsia C, Suzuki H, Kadowaki T,
Koyasu S and Liou HC: Phosphatidylinositol 3-kinase and NF-kappa
B/Rel are at the divergence of CD40-mediated proliferation and
survival pathways. J Immunol. 165:3860–3867. 2000. View Article : Google Scholar
|
|
110
|
Ahmad R, Raina D, Joshi MD, Kawano T, Ren
J, Kharbanda S and Kufe D: MUC1-C oncoprotein functions as a direct
activator of the nuclear factor-kappaB p65 transcription factor.
Cancer Res. 69:7013–7021. 2009. View Article : Google Scholar
|
|
111
|
Datta SR, Dudek H, Tao X, Masters S, Fu H,
Gotoh Y and Greenberg ME: Akt phosphorylation of BAD couples
survival signals to the cell-intrinsic death machinery. Cell.
91:231–241. 1997. View Article : Google Scholar
|
|
112
|
Datta SR, Katsov A, Hu L, Petros A, Fesik
SW, Yaffe MB and Greenberg ME: 143-3 proteins and survival kinases
cooperate to inactivate BAD by BH3 domain phosphorylation. Mol
Cell. 6:41–51. 2000. View Article : Google Scholar
|
|
113
|
Fleury C, Mignotte B and Vayssière JL:
Mitochondrial reactive oxygen species in cell death signaling.
Biochimie. 84:131–141. 2002. View Article : Google Scholar
|
|
114
|
Redza-Dutordoir M and Averill-Bates DA:
Activation of apoptosis signalling pathways by reactive oxygen
species. Biochim Biophys Acta. 1863:2977–2992. 2016. View Article : Google Scholar
|
|
115
|
Yin L, Huang L and Kufe D: MUC1
oncoprotein activates the FOXO3a transcription factor in a survival
response to oxidative stress. J Biol Chem. 279:45721–45727. 2004.
View Article : Google Scholar
|
|
116
|
Park JA, Park S, Choi JK, Han MK and Lee
Y: Inhibition of MUC1-C Increases ROS and Cell Death in Mouse
Embryonic Stem Cells. Int J Stem Cells. 14:180–190. 2021.
|
|
117
|
Liu Y, Ao X, Ding W, Ponnusamy M, Wu W,
Hao X, Yu W, Wang Y, Li P and Wang J: Critical role of FOXO3a in
carcinogenesis. Mol Cancer. 17:1042018. View Article : Google Scholar
|
|
118
|
Levine AJ, Momand J and Finlay CA: The p53
tumour suppressor gene. Nature. 351:453–456. 1991. View Article : Google Scholar
|
|
119
|
Amaral JD, Xavier JM, Steer CJ and
Rodrigues CM: The role of p53 in apoptosis. Discov Med. 9:145–152.
2010.
|
|
120
|
Wei X, Xu H and Kufe D: Human MUC1
oncoprotein regulates p53 responsive gene transcription in the
genotoxic stress response. Cancer Cell. 7:167–178. 2005. View Article : Google Scholar
|
|
121
|
Huang HZ, Yin YF, Wan WJ, Xia D, Wang R
and Shen XM: Up-regulation of microRNA-136 induces apoptosis and
radiosensitivity of esophageal squamous cell carcinoma cells by
inhibiting the expression of MUC1. Exp Mol Pathol. 110:1042782019.
View Article : Google Scholar
|
|
122
|
Wang JJ, Li ZF, Li XJ, Han Z, Zhang L and
Liu ZJ: Effects of microRNA-136 on melanoma cell proliferation,
apoptosis, and epithelial-mesenchymal transition by targetting PMEL
through the Wnt signaling pathway. Biosci Rep. 37:BSR201707432017.
View Article : Google Scholar
|
|
123
|
Wang L, Wu X, Wang B, Wang Q and Han L:
Mechanisms of miR-145 regulating invasion and metastasis of ovarian
carcinoma. Am J Transl Res. 9:3443–3451. 2017.
|
|
124
|
Wang X, Zhou X, Zeng F, Wu X and Li H:
miR-485-5p inhibits the progression of breast cancer cells by
negatively regulating MUC1. Breast Cancer. 27:765–775. 2020.
View Article : Google Scholar
|
|
125
|
Du C, Fang M, Li Y, Li L and Wang X: Smac,
a mitochondrial protein that promotes cytochrome c-dependent
caspase activation by eliminating IAP inhibition. Cell. 102:33–42.
2000. View Article : Google Scholar
|
|
126
|
Tummers B and Green DR: Caspase-8:
Regulating life and death. Immunol Rev. 277:76–89. 2017. View Article : Google Scholar
|
|
127
|
Ren J, Raina D, Chen W, Li G, Huang L and
Kufe D: MUC1 oncoprotein functions in activation of fibroblast
growth factor receptor signaling. Mol Cancer Res. 4:873–883. 2006.
View Article : Google Scholar
|
|
128
|
Chen Q, Li D, Ren J, Li C and Xiao ZX:
MUC1 activates JNK1 and inhibits apoptosis under genotoxic stress.
Biochem Biophys Res Commun. 440:179–183. 2013. View Article : Google Scholar
|
|
129
|
Dunn C, Wiltshire C, MacLaren A and
Gillespie DA: Molecular mechanism and biological functions of c-Jun
N-terminal kinase signalling via the c-Jun transcription factor.
Cell Signal. 14:585–593. 2002. View Article : Google Scholar
|
|
130
|
Yarza R, Vela S, Solas M and Ramirez MJ:
c-Jun N-terminal Kinase (JNK) signaling as a therapeutic target for
Alzheimer's disease. Front Pharmacol. 6:3212016. View Article : Google Scholar
|
|
131
|
Zhang AM, Chi XH, Bo ZQ, Huang XF and
Zhang J: MUC1 gene silencing inhibits proliferation, invasion, and
migration while promoting apoptosis of oral squamous cell carcinoma
cells. Biosci Rep. 39:BSR201821932019. View Article : Google Scholar
|
|
132
|
Tanaka Y, Terai Y, Kawaguchi H, Fujiwara
S, Yoo S, Tsunetoh S, Takai M, Kanemura M, Tanabe A and Ohmichi M:
Prognostic impact of EMT
(epithelial-mesenchymal-transition)-related protein expression in
endometrial cancer. Cancer Biol Ther. 14:13–19. 2013. View Article : Google Scholar
|
|
133
|
Vega S, Morales AV, Ocaña OH, Valdés F,
Fabregat I and Nieto MA: Snail blocks the cell cycle and confers
resistance to cell death. Genes Dev. 18:1131–1143. 2004. View Article : Google Scholar
|
|
134
|
Renauld JC: Class II cytokine receptors
and their ligands: Key antiviral and inflammatory modulators. Nat
Rev Immunol. 3:667–676. 2003. View Article : Google Scholar
|
|
135
|
Yi FT and Lu QP: Mucin 1 promotes
radioresistance in hepatocellular carcinoma cells through
activation of JAK2/STAT3 signaling. Oncol Lett. 14:7571–7576.
2017.
|
|
136
|
Kato K, Lillehoj EP and Kim KC: MUC1
regulates epithelial inflammation and apoptosis by PolyI:C through
inhibition of Toll/IL-1 receptor-domain-containing adapter-inducing
IFN-β (TRIF) recruitment to Toll-like receptor 3. Am J Respir Cell
Mol Biol. 51:446–454. 2014. View Article : Google Scholar
|
|
137
|
Tréhoux S, Duchêne B, Jonckheere N and Van
Seuningen I: The MUC1 oncomucin regulates pancreatic cancer cell
biological properties and chemoresistance. Implication of p42-44
MAPK, Akt, Bcl-2 and MMP13 pathways. Biochem Biophys Res Commun.
456:757–762. 2015. View Article : Google Scholar
|
|
138
|
Xu X, Wells A, Padilla MT, Kato K, Kim KC
and Lin Y: A signaling pathway consisting of miR-551b, catalase and
MUC1 contributes to acquired apoptosis resistance and
chemoresistance. Carcinogenesis. 35:2457–2466. 2014. View Article : Google Scholar
|
|
139
|
Frisch SM and Ruoslahti E: Integrins and
anoikis. Curr Opin Cell Biol. 9:701–706. 1997. View Article : Google Scholar
|
|
140
|
Piyush T, Rhodes JM and Yu LG: MUC1
O-glycosylation contributes to anoikis resistance in epithelial
cancer cells. Cell Death Discov. 3:170442017. View Article : Google Scholar
|