Introduction
Lung cancer (LC) is the most fatal type of
malignancy world-wide and non-small cell LC (NSCLC) accounts for
85% of patients with LC (1).
NSCLC may be a predominant cause of the increasing number of deaths
resulting from tumors in all countries (2,3).
Older individuals (age, >65 years) constitute the most
susceptible group to NSCLC, with a rapidly increasing risk of
morbidity and death (4). The
known causes of NSCLC include excessive smoking, air contamination
and exposure to radon (5).
Supportive care, immunologic and biological therapies, chemotherapy
and radiotherapy are all promising choices for NSCLC treatment
(6). However, resistance derived
from a long-term use of chemotherapy markedly hinders its
efficiency (7). In addition, the
clinical repercussions and survival rate of individuals with NSCLC
have merely improved (8). To
date, biomarkers have been utilized in the diagnosis, prognosis and
attenuation of NSCLC (9). With
this background and the requirement to identify reliable biomarkers
of NSCLC and possible interventions to induce cell radiosensitivity
in NSCLC, the present study was performed to determine the
underlying mechanisms of NSCLC.
Long noncoding RNAs (lncRNAs) and microRNAs
(miRNAs/miRs) are being widely studied to identify medical
solutions targeting radioresistance involved in radiotherapies for
NSCLC (10). lncRNAs are the gold
standard for cancer diagnosis and prevention due to their wide
range of carcinogenic or antitumoral effects on the occurrence and
progression of tumors (11).
Initially, the lncRNA cytoskeleton regulator (CYTOR) was documented
to be sufficiently expressed in NSCLC and associated with cell
biological activities and frustrating survival rates (12). In addition, CYTOR overexpression
is associated with unsatisfactory clinical results and leads to
aggravated cancer cellular mobility, aggressiveness and the
epithelial-mesenchymal transition (EMT) of neoplasms, including
breast cancer, gastric cancer and colon cancer (13,14). Of note, CYTOR is also responsible
for the escalated relapse and resistance of various cancer types,
including oral squamous cell carcinoma and breast cancer (15,16). Since CYTOR affects most
malignancies by participating in competing endogenous RNA (ceRNA)
interactions (17-19), the present study sought to
investigate miRs that hold promise in alleviating NSCLC. Similarly,
alterations in certain miRNAs significantly influence cellular
growth, the EMT, metabolic pathways, apoptosis and radioresistance
in NSCLC (20). According to
certain previous studies, miR-206 is repeatedly acknowledged as a
major cytokine in the ceRNA network involved in cancer progression
(21,22). miR-206 is expressed at low levels
in NSCLC, resulting in the active aggressiveness, invasiveness and
dissemination of cells (23).
However, Samaeekia et al (24) revealed that miR-206 suppresses
breast tumor stemness and metastasis by inhibiting both
self-renewal and invasion. Furthermore, miR-206 was demonstrated to
reduce the resistance of LC toward effective drugs (25). When miRNAs are sponged in NSCLC
during the treatment process, certain downstream mRNAs may also be
regulated (26). Prothymosin α
(PTMA) is activated in several cancer types (such as esophageal
cancer and ovarian cancer), suggesting that it may represent a
target (27,28). Based on these findings, the
present study aimed to elucidate the effect of CYTOR on NSCLC
radiosensitivity by manipulating the miR-206/PTMA axis.
Materials and methods
Expression analysis of CYTOR
The online tool Starbase (http://starbase.sysu.edu.cn/) (29) was employed for expression analysis
of CYTOR on LUSC (n=501) and LUAD (n=526) data from TCGA.
Collection of clinical tissues
Between February 2013 and June 2015, 58 pairs of
NSCLC and paracancerous tissue specimens (≥5 cm away from tumor
tissue) were excised from patients with NSCLC during surgery at
Yantaishan Hospital (Yantai, China); clinically diagnosed and
histopathologically confirmed. None of these patients were treated
by radiotherapy or chemotherapy prior to surgery. After the
operation, each patient underwent radiotherapy and follow-up for at
least 60 months and their survival was recorded. If the tumor
recurred or the patient died, the follow-up ended; otherwise, the
data were recorded up to the last follow-up. The patients who
relapsed or died within 60 months were classified as the
radiotherapy-resistant group (n=28), and the remaining patients
were classified as the radiotherapy-sensitive group (n=30). The
present study was approved by the Ethics Committee of Yantaishan
Hospital (Yantai, China; accession no. for approval, YSLZ2021037).
All study procedures conformed to the Declaration of Helsinki.
Written informed consent was obtained from each patient. Supplementary Table SI provides the
general information of the clinically included patients.
Cell culture and transfection
A normal human lung epithelial cell line (BEAS-2B)
and NSCLC cell lines (H1650, H460 and A549; all from the China
Infrastructure of Cell Line Resource) were cultured in RPMI-1640
basic medium containing 10% fetal bovine serum, 100 mg/ml
streptomycin and 100 U/ml penicillin (all from Gibco; Thermo Fisher
Scientific, Inc.) in an incubator with 5% CO2 at
37°C.
Short hairpin RNA targeting CYTOR (sh-CYTOR),
miR-206 mimics, miR-206 inhibitor, mimics-negative control (NC),
inhibitor-NC, overexpression vector for PTMA (pcDNA3.1-PTMA) and NC
(empty pcDNA3.1 vector; Shanghai GenePharma Co., Ltd.) were
transfected with Lipofectamine® 3000 (Invitrogen; Thermo
Fisher Scientific, Inc.) following the manufacturer's protocol to
generate stably transfected cells (30). The plasmids and sequence
information are provided in Table
SII.
Establishment of radioresistant
cells
Radioresistant cells were established as previously
described (31). H460 and A549
cells were cultured to 90% confluence and then irradiated at a dose
of 0-8 Gy. After the X-ray irradiation, the medium was replaced
with fresh medium and the cells were allowed to grow in a 37°C
incubator. For the generation of radioresistant cells, the
radiation dose applied to the cells (90% confluence) was gradually
increased at a rate of 2.0 Gy/fraction until the final dose of 64
Gy was reached. The cells were subcultured 5 times at the same dose
and the radiation dose was then increased. The produced
radioresistant cell lines were named H460R and A549R and their
viability after radiation was tested by the Cell Counting Kit-8
(CCK-8) method.
Reverse transcription-quantitative PCR
(RT-qPCR)
According to a previous protocol (30), the total RNA was isolated from the
tissues or cells using TRIzol® reagent (Thermo Fisher
Scientific, Inc.). Following the instructions of the PrimeScript RT
kit (Takara Bio Inc.), the extracted RNA was reverse-transcribed
into cDNA, which was subsequently amplified by real-time qPCR
(Perfect Real Time; Takara Bio Inc.) to quantify the levels of
CYTOR, miR-206 and PTMA. The PCR amplification procedure was set as
two steps: The first step was pre-denaturation at 95°C for 10 min;
the second step was PCR for 40 cycles with denaturation at 95°C for
10 sec and annealing/extension at 60°C for 30 sec, and each sample
was set up in three wells. The expression of CYTOR and PTMA was
standardized to that of 18S RNA and GAPDH, respectively, and that
of miR-206 was standardized to that of U6. The relative expression
of each cytokine was attained using the 2−ΔΔCq method
(32). The primer sequences are
listed in Table I.
 | Table IPrimer sequences used for PCR. |
Table I
Primer sequences used for PCR.
| Gene | Primer
sequence |
|---|
| 18S RNA | F:
5′-CGTTCTTAGTTGGTGGAGCG-3′ |
| R:
5′-CCGGACATCTAAGGGCATCA-3′ |
| U6 | F:
5′-CGCTTCGGCAGCACATATAC-3′ |
| R:
5′-AATATGGAACGCTTCACGA-3′ |
| GAPDH | F:
5′-GGGAGCCAAAAGGGTCAT-3′ |
| R:
5′-GAGTCCTTCCACGATACCAA-3′ |
| miR-206 | F:
5′-GCTTCCCGAGGCCACATGCT-3′ |
| R:
5′-CACTTGCCGAAACCACACAC-3′ |
| CYTOR | F:
5′-GCGGTGCCTGAGCCCGTGCC-3′ |
| R:
5′-GGGCGGTTGGAACCAGGCC-3′ |
| PTMA | F:
5′-ATGTCAGACGCAGCCGTAG-3′ |
| R:
5′-CTAGTCATCCTCGTCGGTC-3′ |
CCK-8 assay
Based on the instructions of the CCK-8 kit (cat. no.
HY-K0301; MedChemExpress), differentially treated cells
(2×104 cells/well) were seeded into 96-well plates and
irradiated at 0, 2, 4, 6 or 8 Gy, followed by 24 h of culture at
37°C. Subsequently, the supernatant was discarded and 20 μl
CCK-8 solution in 180 μl fresh medium was added to each well,
followed by culture at 37°C for 1 h. The optical density of each
well at 450 nm was attained using a spectrophotometer.
Colony formation assay
The assay was performed as previously described
(31). The 5,000 cells in each
group were cultured for 14 days after X-ray irradiation at 4 Gy.
The colonies were subjected to 15 min of fixation with 4%
paraformaldehyde and 10 min of staining with 1% crystal violet at
room temperature. Colonies with >50 cells were scored and
counted under the microscope (Olympus Corporation), followed by
counting the number of colonies in five randomly selected areas.
Each procedure was performed 3 times per group.
Comet assay
A comet assay was applied to differently treated
cells according to a previous study (33). Specifically, the cell suspension
(1×106 cells/ml) was mixed with low melting point
agarose and dripped onto precoated slides (1% normal melting point
agarose) incubated with lysis solution (pH 10.0) and
electrophoresis buffer (pH >13.0) for 40 min and electrophoresed
for 40 min (25 V, 280 mA) (33).
Subsequently, the slides were placed in a neutralization solution
(pH 7.5) for 10 min, covered with sufficient staining solution (20
μg/ml ethidium bromide) for 10 min and immersed twice in PBS
(10 min each time) and distilled water. Subsequently, comets under
a 510-560 nm excitation filter and a 590 nm blocking filter were
observed and analyzed under a fluorescence microscope. All phases
of the comet assay were performed under at 4°C in the dark and all
solutions were freshly prepared and cooled for use.
γ-H2AX analysis
Cells with different treatments were cultured in
8-well slides for 24 h, fixed/permeabilized in cold (4°C) methanol
for 5 min and blocked with 1% BSA (Thermo Fisher Scientific, Inc.)
in PBS containing Tween-20 for 30 min at 37°C. Subsequently, the
cells were incubated with γ-H2AX antibody (1:250 dilution; cat. no.
ab81299; Abcam) overnight at 4°C, followed by immunoglobulin G
(1:250 dilution; cat. no. ab205781; Abcam) for 1 h. The cells were
stained with DAPI prior to mounting with coverslips. Finally,
images were captured and analyzed under a fluorescence
microscope.
Bioinformatics
The downstream target miRNAs of lncRNA CYTOR and the
downstream target genes of miR-206 were predicted through the
online websites Starbase (http://starbase.sysu.edu.cn/) (29), DIANA tools (http://carolina.imis.athena-innovation.gr/diana_tools/web/index.php?r=lncbasev2%2Findex-predicted)
(34), miRcode (http://www.mircode.org/?gene=HOTAIR&mirfam=&class=&cons=&trregion=)
(35), Targetscan(http://www.targetscan.org/vert_71/?tdsourcetag=s_pcqq_aiomsg)
(36) and miRTarBase (http://mirtarbase.cuhk.edu.cn/php/index.php) (37).
RNA fluorescence in situ hybridization
(FISH)
In order to detect the localization of CYTOR in
cells, RNA FISH was performed using a green fluorescent-labeled
CYTOR probe (Guangzhou RiboBio Co., Ltd.). The cells were fixed
with 4% paraformaldehyde for 15 min, permeated with 0.5% Triton
X-100 on ice for 10 min and then treated with prehybridization
buffer at 37°C for 30 min. Next, the cells were hybridized with the
fluorescent probe. After 12 h at 37°C, they were stained with DAPI.
Finally, FISH results were obtained by confocal microscopy (Sp8;
Leica Microsystems).
Dual-luciferase reporter gene assay
CYTOR or PTMA fragments containing the miR-206
binding site were cloned into the pmirGLO dual oligosaccharase
vector (Promega Corporation) to construct the
pmirGLO-CYTOR-wild-type (WT), pmirGLO-PTMA-WT, pmirGLO-CYTOR-mutant
type (MUT) and pmirGLO-PTMA-MUT reporter vectors. After the cells
(6×104 cells/well) were seeded into 24-well plates,
pmirGLO-CYTOR-WT, pmirGLO-CYTOR-MUT, pmirGLO-PTMA-WT and
pmirGLO-PTMA-MUT were cotransfected with miR-206 mimics or
mimics-NC for 24 h. Subsequently, luciferase activity was assessed
using dual-luciferase assay kits (Promega Corporation). The
relative firefly luciferase activity was determined by
normalisation to Renilla luciferase activity.
RNA pull-down assay
To clarify the binding relationship between CYTOR
and miR-206 (38), biotinylated
CYTOR and CYTOR-NC probes (Thermo Fisher Scientific, Inc.) were
dissolved in washing/binding buffer according to the manufacturer's
protocol and then cultivated with streptavidin-conjugated magnetic
beads (Thermo Fisher Scientific, Inc.) for 2 h, followed by
cultivation for 2 h with cell lysates supplemented to the buffer to
remove RNA complexes conjugated to magnetic beads. Subsequently,
miR-206 expression was determined by RT-qPCR.
Xenograft tumors in nude mice
After a week of adaptive feeding (20-22°C, 50-60%
humidity, 12-h light/dark cycle; ad libitum access to food
and water), 3×106 H460R + sh-NC or H460R + sh-CYTOR
cells were subcutaneously inoculated into the right flank of BALB/c
nude mice (n=24; age, 5 weeks; body weight, 18-20 g; Beijing Vital
River Laboratory Animal Technology Co., Ltd.), with 12 mice per
group. The tumor dimensions were measured with calipers every 3
days and the volume was calculated as follows: Tumor volume =
(length × width2)/2. When the tumor volume reached
250-300 mm3, pentobarbital (50 mg/kg) was administered
via intraperitoneal injection and the tumors of the mice were
exposed to a single dose of 20 Gy ionizing radiation. For the tumor
irradiation, the anesthetized mice were fixed on a plate with the
right hind leg carrying the tumor exposed to the radiation field,
while the other parts were protected by a lead plate. After the
single irradiation, the tumor volume was examined every 3 days. The
health and behavior of the mice were monitored every 2 days. The
humane endpoints were as follows: Weight loss of >15% or the
nude mice suffering from the tumor load or a tumor length of
>1.50 cm and short diameter of >1.15 cm. Upon reaching the
humane endpoints, the mice were euthanized by an intraperitoneal
injection of an overdose of pentobarbital (>200 mg/kg). Death
was confirmed by observation of pupil dilation as well as ceasing
of breath (absence of chest fluctuation) and the heartbeat.
Subsequently, tumors were removed and weighed, and the tumors from
6 mice per group were washed, and paraffin-embedded sections were
prepared for the immunohistochemical analysis, while the remaining
tumors were ground into homogenate for the RT-qPCR analysis. The
animal experiments were performed in accordance with the
requirements of the guidelines for the use of experimental animals
(39), with the approval of the
Institutional Animal Care and Use Committee at Yantaishan Hospital
(Yantai, China). The accession number for this approval was
YSLZ2021026. Significant efforts were made to minimize both the
number of animals used and their respective suffering.
Immunohistochemistry
The tumor sections were deparaffinized, hydrated,
incubated with anti-ki67 (1:200 dilution; cat. no. ab16667; Abcam)
at 4°C and reacted with IgG (1:2,000 dilution; cat. no. ab205718;
Abcam) at 37°C for 2 h. After washing with PBS, the sections were
developed with diaminobenzidine, sealed and then observed and
analyzed under a microscope.
Statistical analysis
SPSS 21.0 software (IBM Corporation) was used for
data analysis. Values are expressed as the mean ± standard
deviation. The normality of distribution of all data was inspected
using the Kolmogorov-Smirnov test. The t-test was applied for
comparisons between two groups, while one-way or two-way ANOVA was
used to compare different groups, with Tukey's multiple-comparisons
test applied for pairwise comparisons after the ANOVA. The P-value
was attained using a two-tailed test and P<0.05 was considered
to indicate a statistically significant difference.
Results
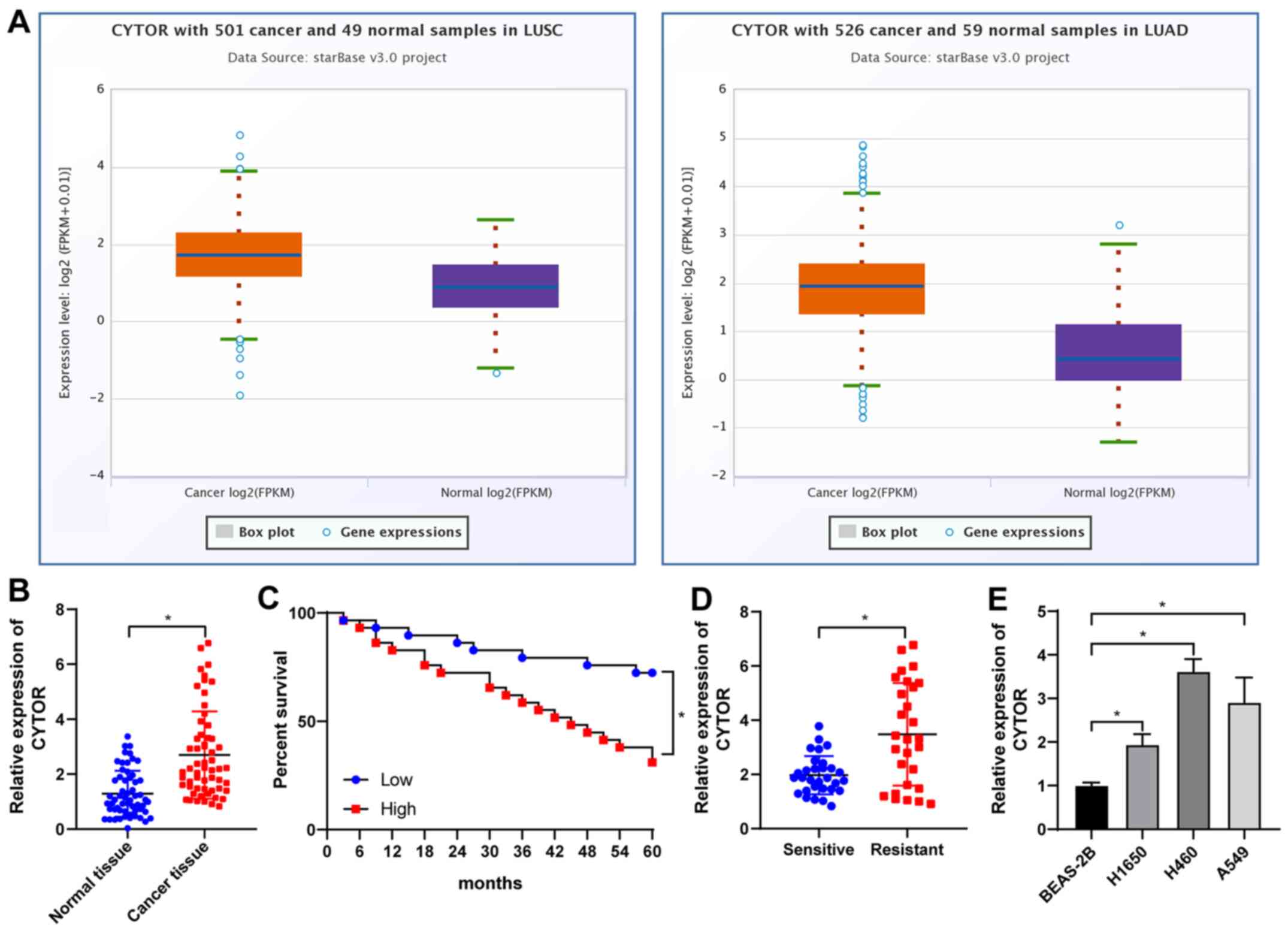
CYTOR is overexpressed in NSCLC and is
responsible for the poor prognosis of patients with NSCLC
Analysis with the starBase database (http://starbase.sysu.edu.cn/) revealed that CYTOR was
overexpressed in lung adenocarcinoma and lung squamous cell
carcinoma (Fig. 1A), suggesting
that CYTOR may be a possible target for NSCLC treatment. Analysis
of the 58 pairs of NSCLC and paracancerous tissues (average age,
51.83±7.22 years; males/females, 29/29). The basic information of
the cohort is provided in Table
SII. It was revealed that CYTOR was upregulated in NSCLC
tissues (P<0.05; Fig. 1B) and
after radiotherapy, the patients with high CYTOR expression had a
worse prognosis than those with low CYTOR expression (P<0.05;
Fig. 1C). The patients were then
divided into a sensitive group (n=30) and a resistant group (n=28)
according to their sensitivity to radiotherapy and the expression
of CYTOR in the two groups was analyzed. The results indicated that
the expression level of CYTOR in the resistant group was
significantly higher than that in the sensitive group (P<0.05;
Fig. 1D). Next, the expression of
CYTOR in different cell lines was detected and the results
indicated that CYTOR expression in the NSCLC cell lines was higher
than that in the non-cancerous cell line (P<0.05; Fig. 1E).
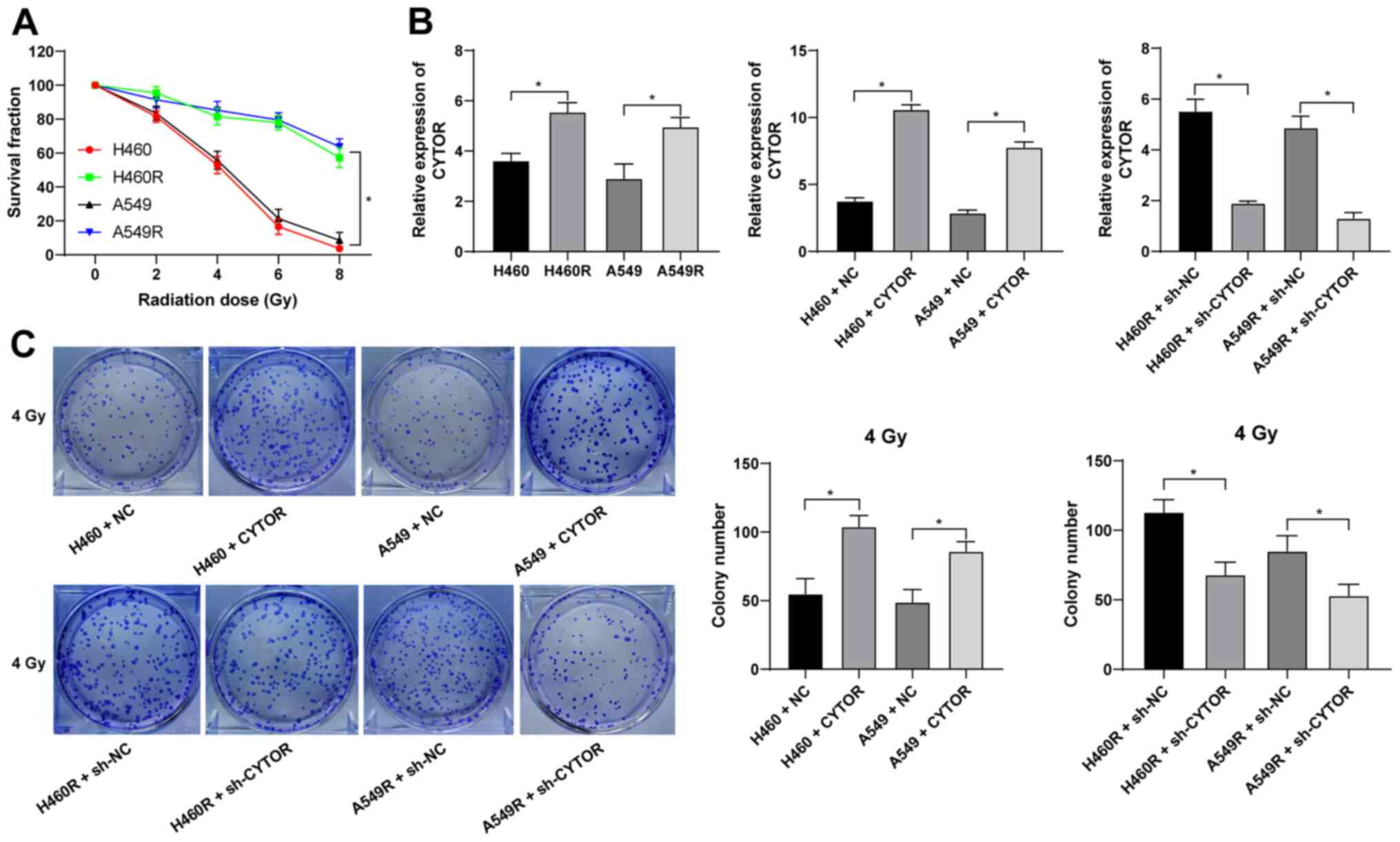
Silencing of CYTOR enhances
radiosensitivity of NSCLC cells
Radiotherapy is used as a prevalent therapy for
NSCLC. To investigate the role of CYTOR in the radiosensitivity of
NSCLC, radioresistant NSCLC cell lines (H460R and A549R) were
established. Compared with the parental cell lines, the
radioresistant cell lines had a better survival rate under
different doses of irradiation (P<0.05; Fig. 2A). The dose of 4 Gy, under which
the survival rate of the parental cells was ~50%, was selected for
the subsequent experiments. According to the RT-qPCR results, CYTOR
was highly expressed in the radioresistant cell lines (P<0.05;
Fig. 2B). CYTOR overexpression
vector was transfected into the parental cell lines (H460 and A549)
and sh-CYTOR was transfected into the radioresistant cell lines;
subsequently, the overexpression and knockdown efficiency was
verified (P<0.05; Fig. 2B).
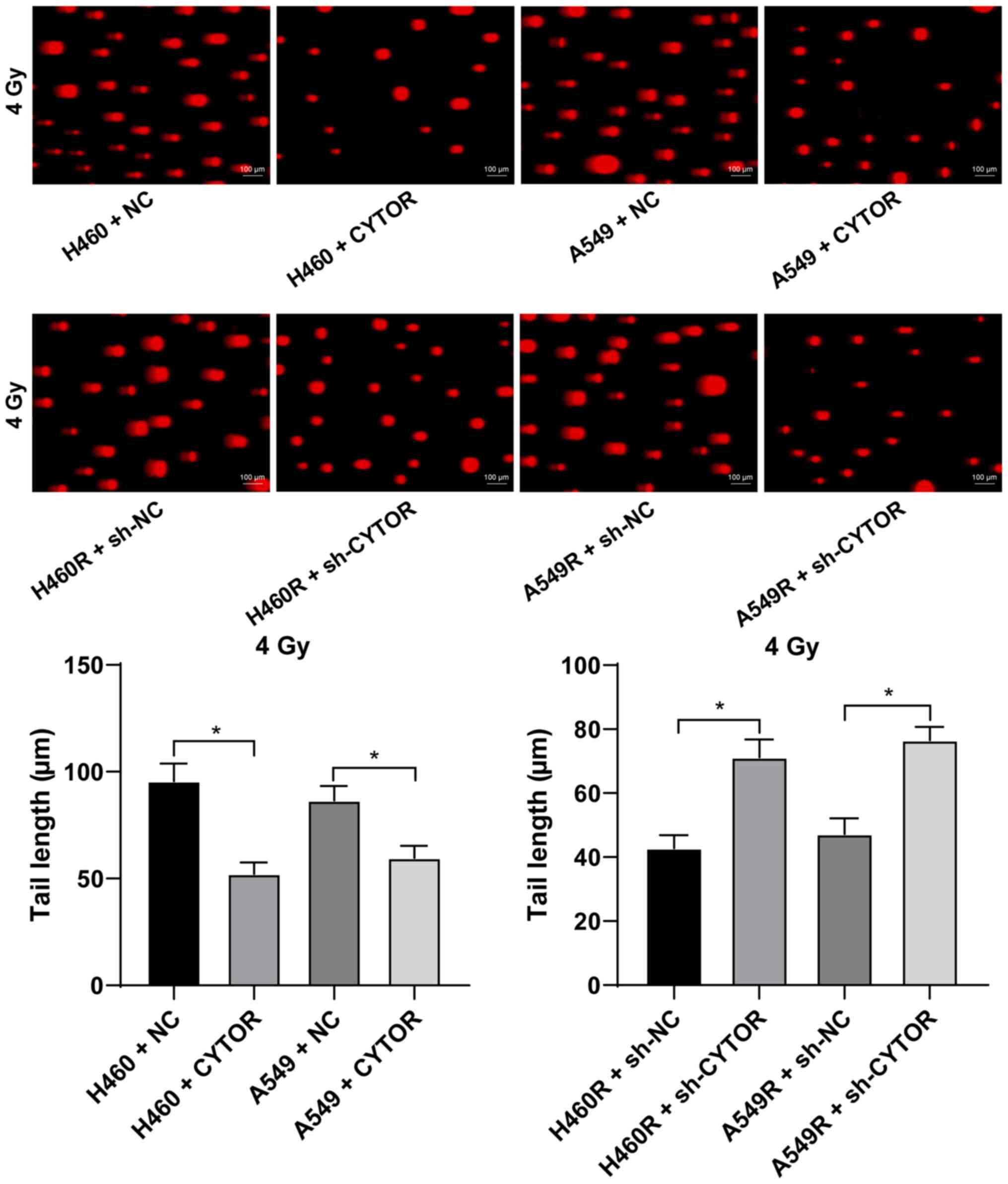
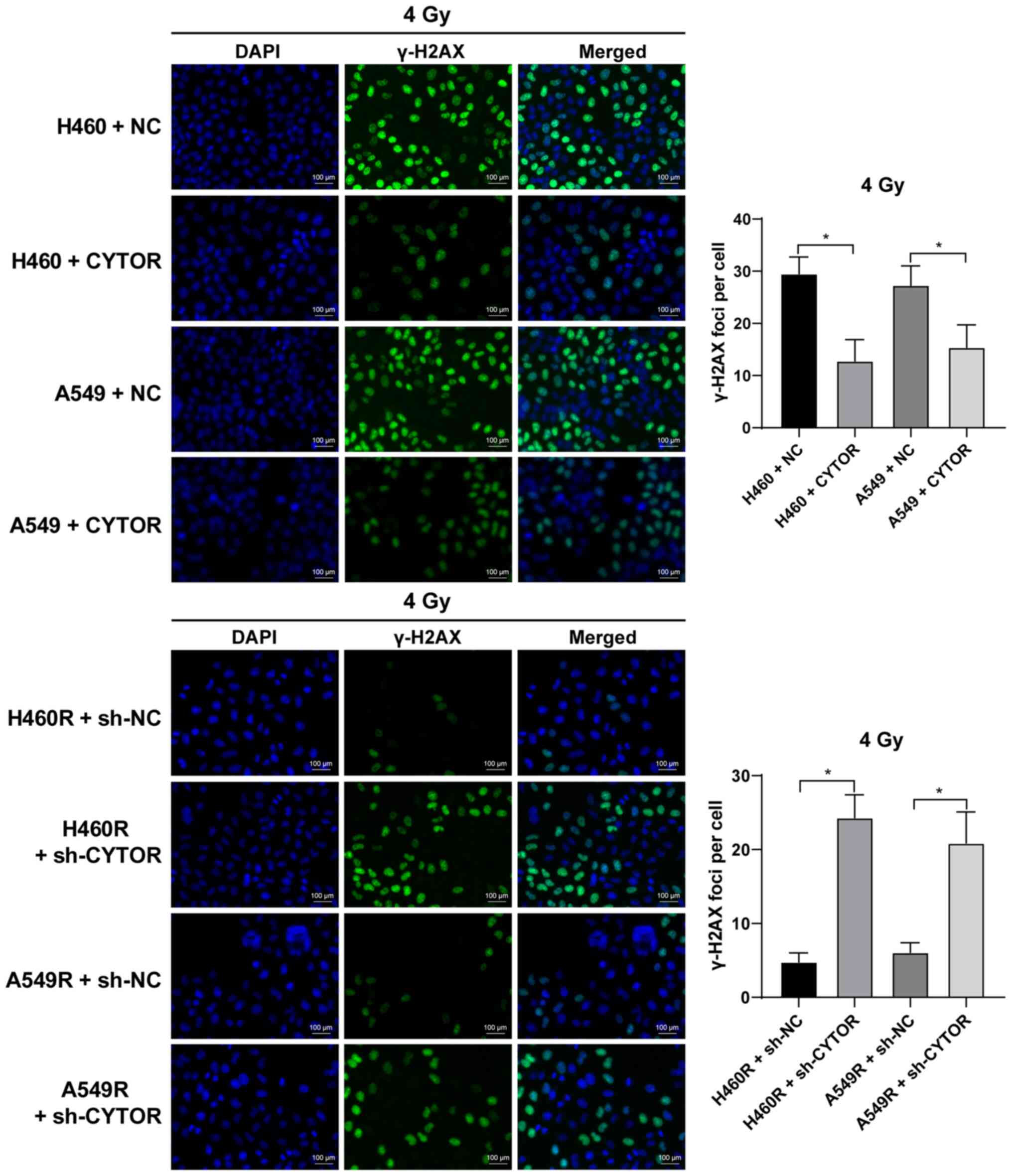
Furthermore, it was revealed that overexpression of CYTOR in the
parental cell lines impeded cell radiosensitivity (stronger colony
formation ability, shorter comet assay tailing and lower γ-H2AX
fluorescence intensity), while knockdown of CYTOR in the
radioresistant cell lines enhanced cell radiosensitivity (weakened
colony formation ability, longer comet assay tailing and increased
γ-H2AX fluorescence intensity) (P<0.05; Figs. 2C, 3 and 4). These results suggest that CYTOR
influences the radiosensitivity of NSCLC cells.
CYTOR targets miR-206 expression in NSCLC
cells
To clarify the specific mechanism of CYTOR to
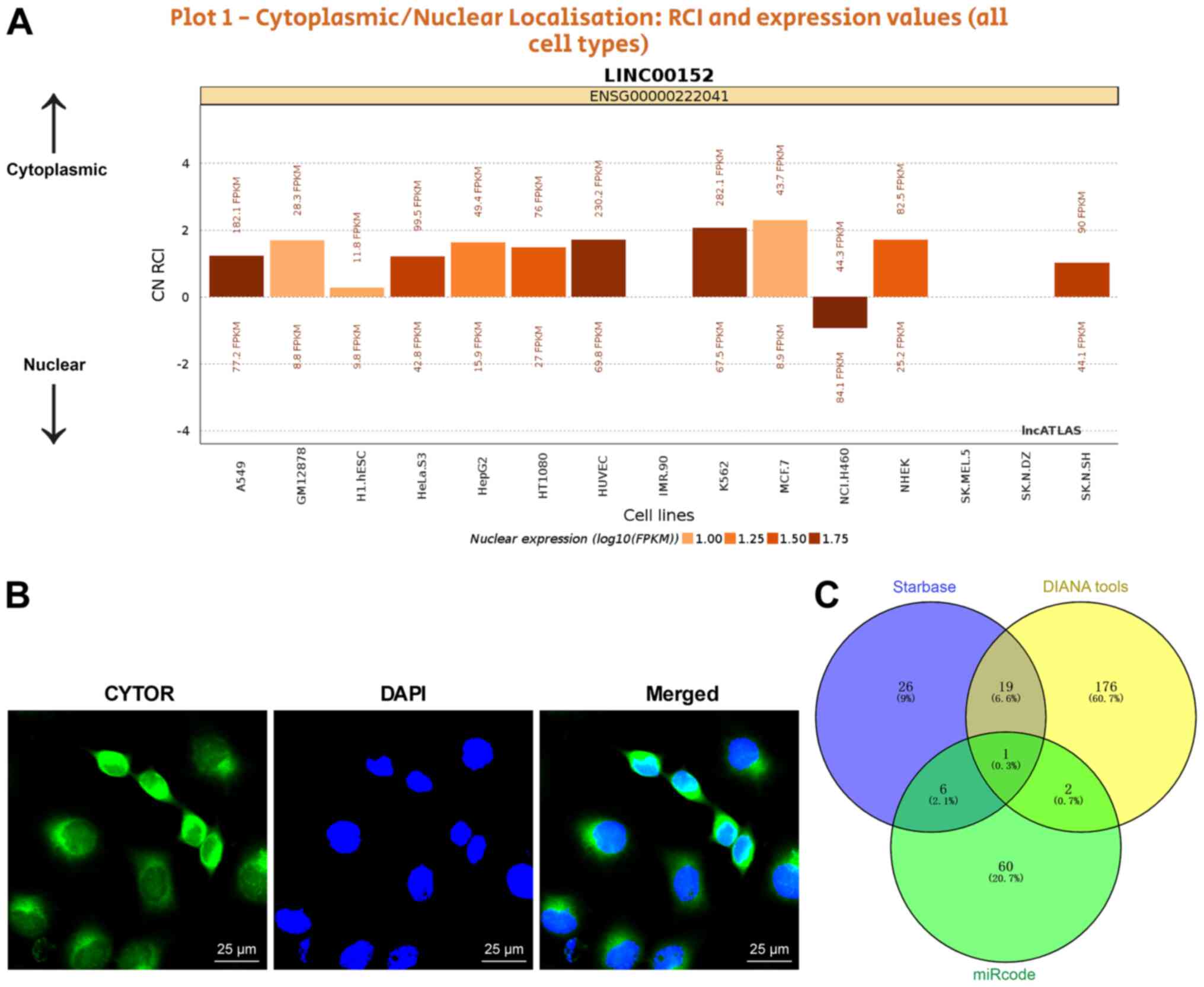
regulate the radiosensitivity of NSCLC cells, the subcellular
localization of CYTOR in the cytoplasm was initially predicted by a
database (http://lncatlas.crg.eu/?tdsourcetag=s_pcqq_aiomsg) and
verified by FISH (Fig. 5A and B),
indicating that CYTOR may affect the radiosensitivity of NSCLC
cells via the ceRNA mechanism. Thus, multiple databases were
employed to search for the downstream targets of CYTOR, and
miR-206, the intersecting target, was identified (Fig. 5C). It has been reported that
miR-206 improves the radiosensitivity of nasopharyngeal carcinoma
(40). Therefore, it may be
speculated that CYTOR affects NSCLC cell radiosensitivity by
targeting miR-206. The StarBase database (http://starbase.sysu.edu.cn/) predicted the binding
site between CYTOR and miR-206 (Fig.
6A), and subsequently, the binding association between CYTOR
and miR-206 was confirmed by a dual-luciferase reporter gene assay
and an RNA pull-down assay (Fig. 6B
and C). Next, compared with the normal adjacent tissues and
normal cell line, respectively, miR-206 expression was determined
to be decreased in both NSCLC tissues and cells, and it was even
further decreased in the radioresistant cells (P<0.05; Fig. 6D and E). Furthermore, miR-206
expression was reduced when CYTOR was overexpressed, while it was
promoted when CYTOR was inhibited (P<0.05; Fig. 6E). In summary, the results
suggested that CYTOR directly targets miR-206 in NSCLC cells.
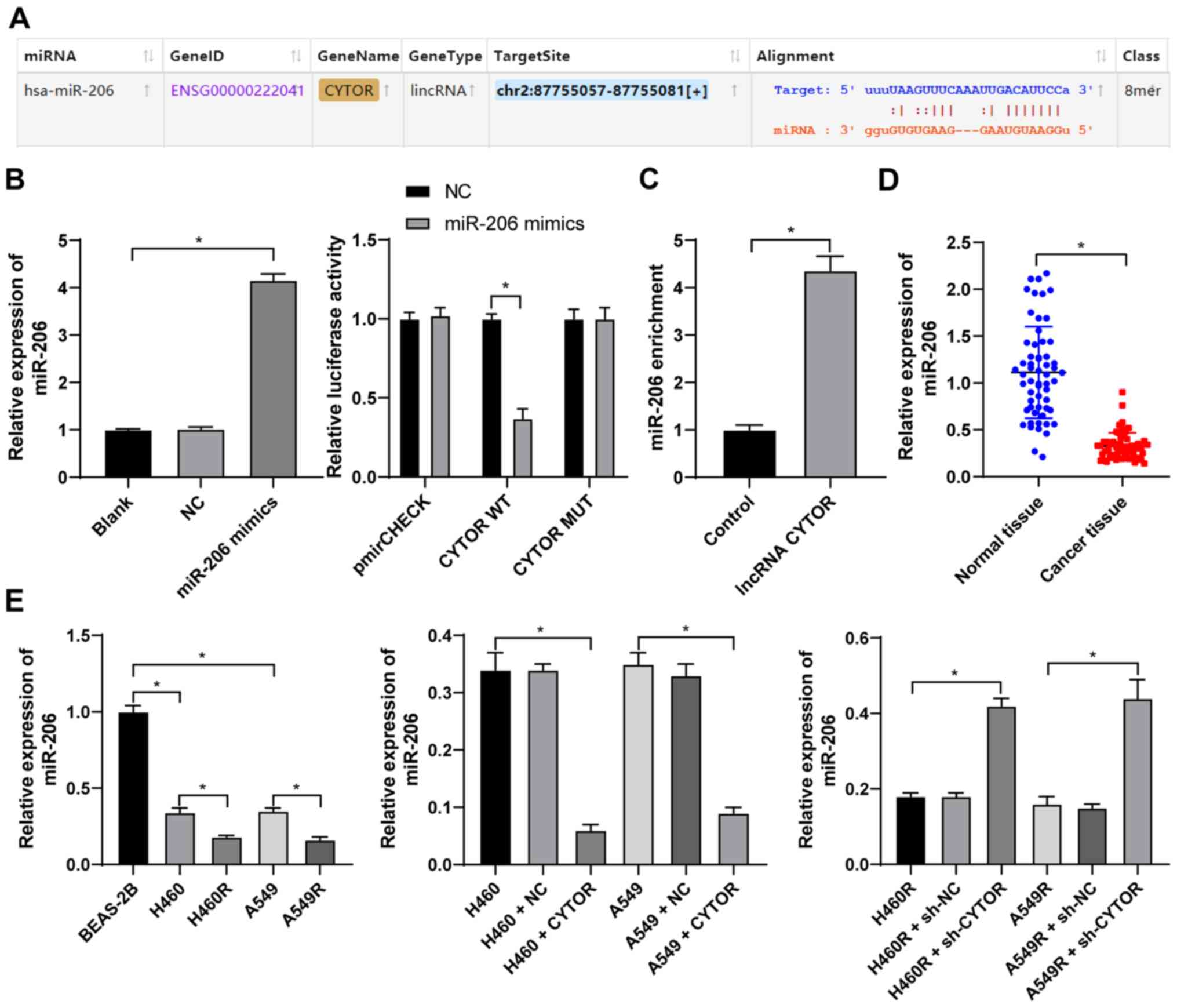 | Figure 6CYTOR targets miR-206. (A) The
binding site between CYTOR and miR-206 predicted via starBase
(http://starbase.sysu.edu.cn/). (B) The
transfection of miR-206 mimics was confirmed by RT-qPCR and the
binding association between CYTOR and miR-206 was verified by a
dual-luciferase reporter assay. (C) The binding relationship
between CYTOR and miR-206 in H460 cells was confirmed by an RNA
pull-down assay. (D and E) miR-206 expression in (D) patients with
NSCLC and (E) NSCLC cell lines as detected by RT-qPCR. Values are
expressed as the mean ± standard deviation. Cell experiments were
performed as three repeats. *P<0.05. RT-qPCR, reverse
transcription-quantitative PCR; NSCLC, non-small cell lung cancer;
CYTOR, long noncoding RNA cytoskeleton regulator; NC, negative
control; CYTOR, long noncoding RNA cytoskeleton regulator;
sh-CYTOR, short hairpin RNA targeting CYTOR; A549R, A549 with
radioresistance; miR/miRNA, microRNA; lincRNA, long intergenic
non-coding RNA; WT, wild-type; MUT, mutant; hsa, Homo
sapiens. |
miR-206 knockdown neutralizes the
inhibitory effect of CYTOR on radiosensitivity of NSCLC cells
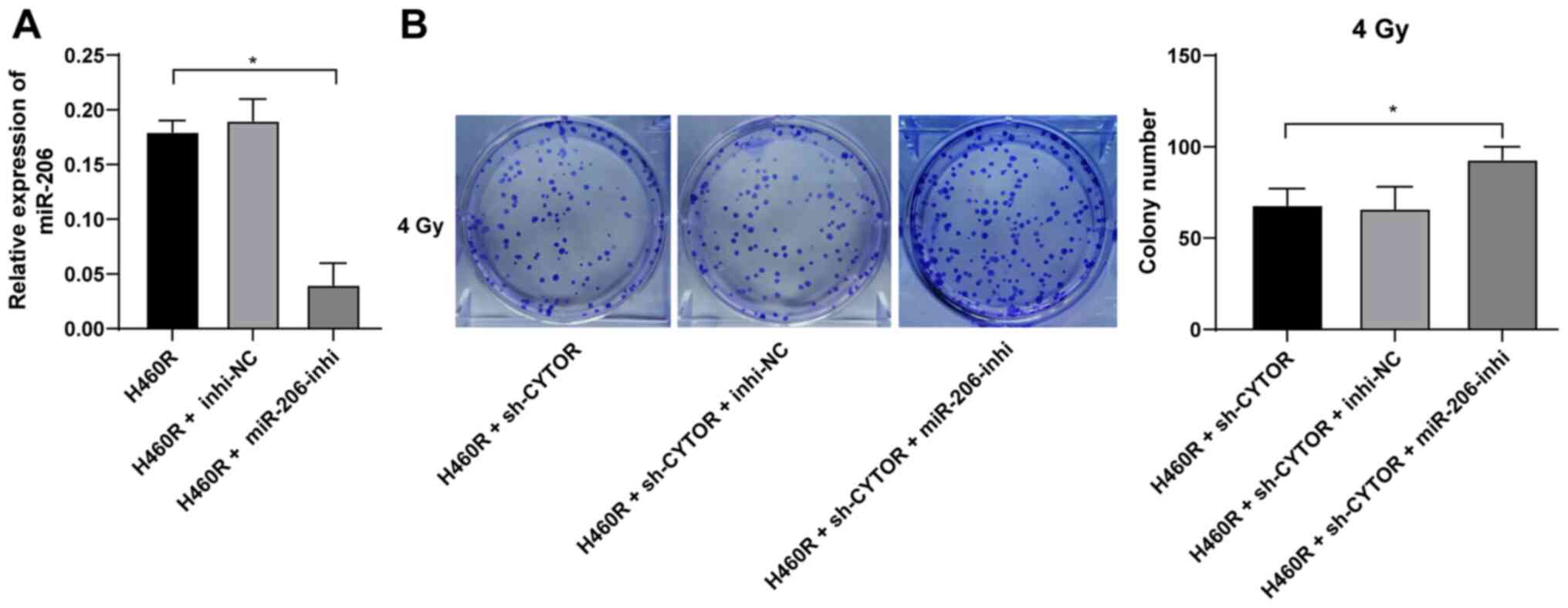
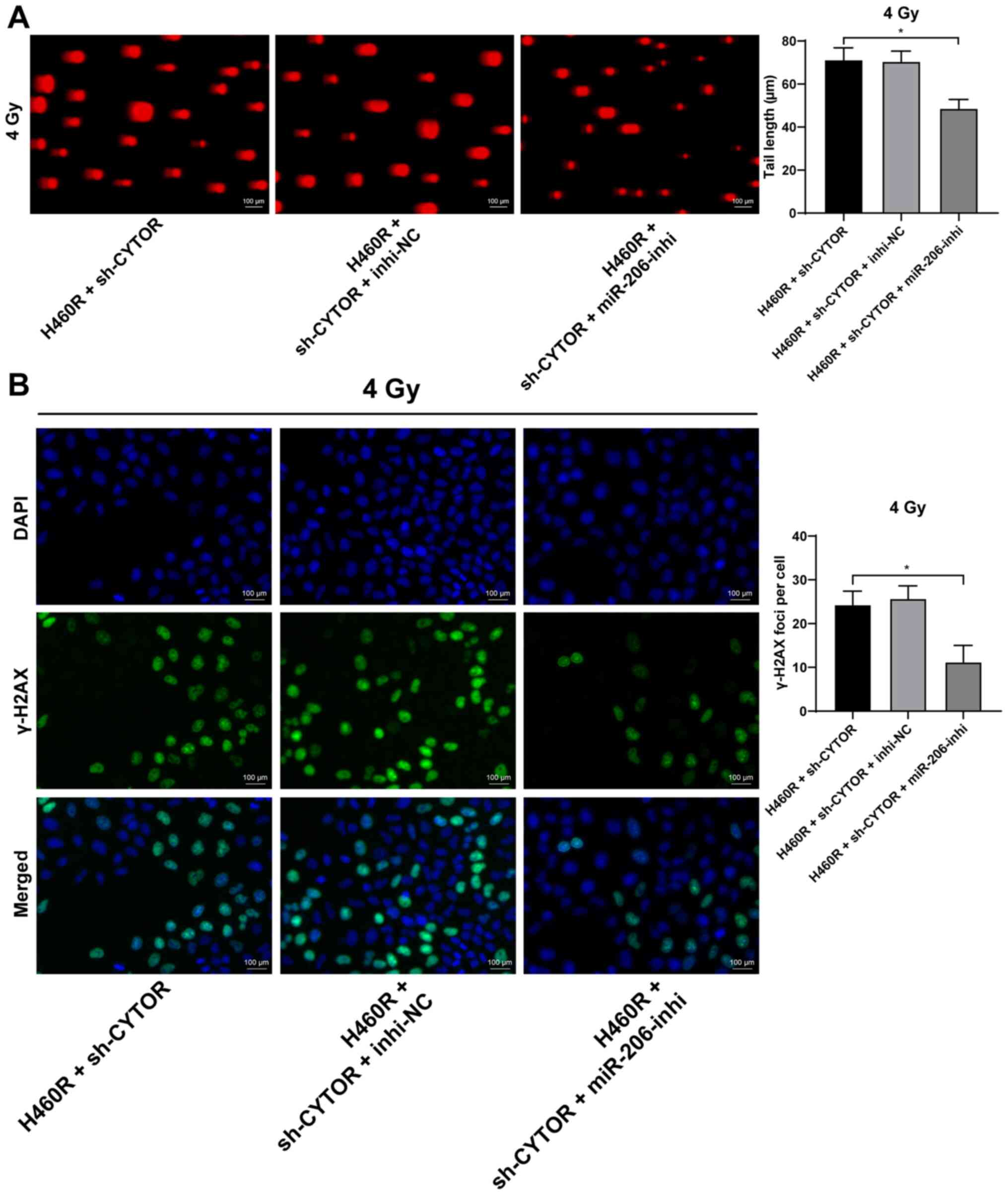
To further validate the role of miR-206 in the
CYTOR-regulated radiosensitivity of NSCLC cells, a miR-206
inhibitor was utilized to degrade miR-206 expression in H460R cells
(P<0.01; Fig. 7A);
subsequently, miR-206 inhibitor was combined with sh-CYTOR for
joint application. The results suggested that the radiosensitivity
of the H460R cells induced by sh-CYTOR was moderately inhibited
(P<0.01; Figs. 7B and 8A and B), indicating that CYTOR
suppressed the radiosensitivity of NSCLC cells by targeting
miR-206.
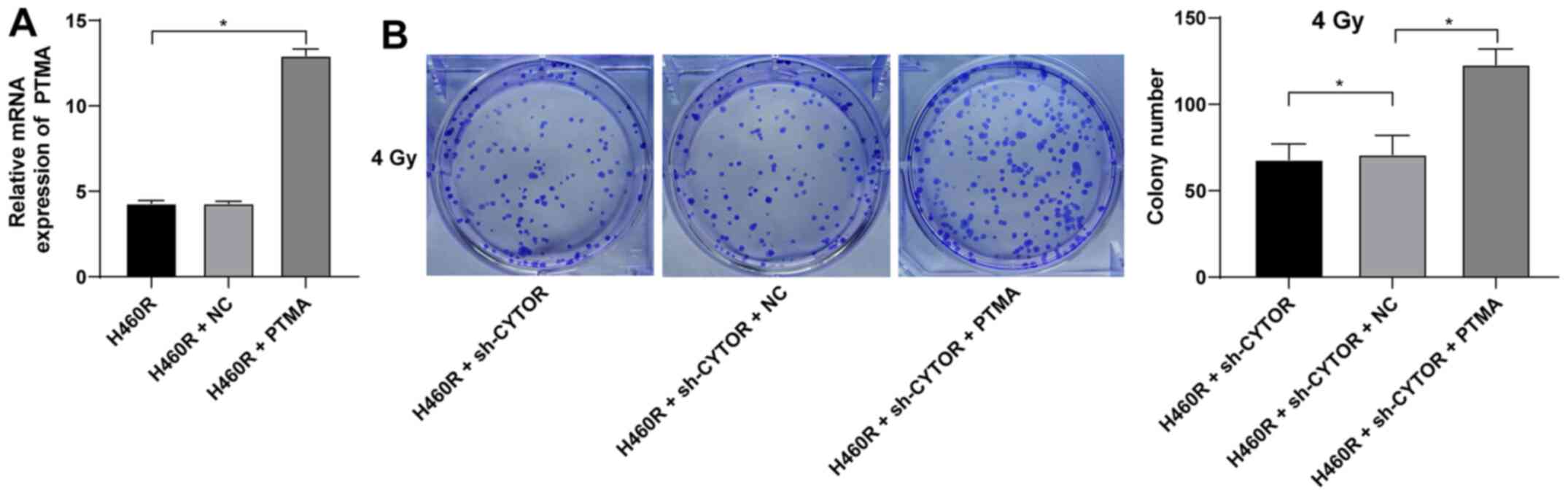
miR-206 targets PTMA expression
Subsequently, the intersection among the target
genes downstream of miR-206 predicted from multiple databases was
attained (Fig. 9A) and PTMA was
identified. PTMA was reported to be highly expressed in a
radiation-resistant colorectal cancer cell line (41). PTMA expression in the NSCLC
tissues and cell lines was measured and it was indicated that the
level of PTMA mRNA was elevated in both the NSCLC tissues and
radioresistant cell lines (P<0.05; Fig. 9B). The target binding relationship
between miR-206 and PTMA was verified by a dual-luciferase reporter
gene assay (P<0.05; Fig. 9C).
In addition, increased intracellular CYTOR expression led to
upregulation of PTMA mRNA and knockdown of CYTOR expression led to
the opposite result; of note, simultaneous CYTOR knockdown and
inhibition of miR-206 promoted PTMA mRNA expression (P<0.05;
Fig. 9D), indicating that miR-206
targeted PTMA in NSCLC cells and that CYTOR competitively bound to
miR-206 to upregulate PTMA expression.
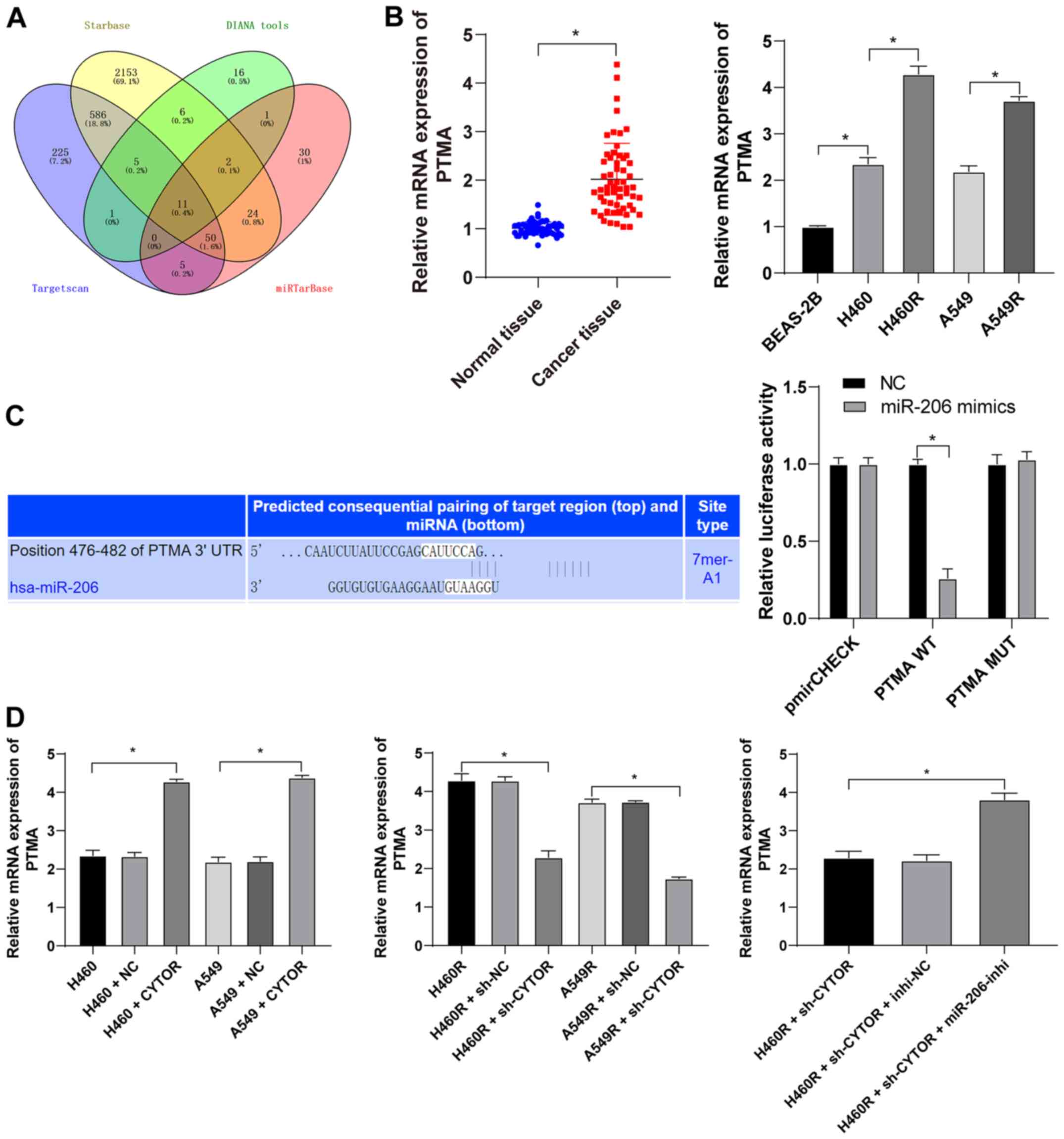 | Figure 9miR-206 targets PTMA expression. (A)
Target genes downstream of miR-206 were analyzed using various
databases and their overlapping results were presented in a Venn
diagram, with yellow representing starBase (http://starbase.sysu.edu.cn/), green representing
DIANA tools (http://carolina.imis.athena-innovation.gr/diana_tools/web/index.php?r=lncbasev2%2Findex-predicted),
blue representing TargetScan (http://www.targetscan.org/vert_71/?tdsourcetag=s_pcqq_aiomsg)
and red representing miRTarBase (http://mirtarbase.cuhk.edu.cn/php/index.php). (B)
Level of PTMA mRNA in 58 pairs of non-small cell lung cancer
tissues and cell lines as detected by RT-qPCR. (C) The binding site
between miR-206 and PTMA was predicted via starBase (http://starbase.sysu.edu.cn/) and their binding
interaction was verified by a dual-luciferase reporter assay. (D)
The level of PTMA mRNA in differently treated cells as verified by
RT-qPCR. Values are expressed as the mean ± standard deviation.
Cell experiments were performed as three repeats.
*P<0.05. RT-qPCR, reverse transcription-quantitative
PCR; miR, microRNA; PTMA, prothymosin α; MUT, mutant; WT,
wild-type; hsa, Homo sapiens; CYTOR, long noncoding RNA
cytoskeleton regulator; sh-CYTOR, short hairpin RNA targeting
CYTOR; NC, negative control; inhi, inhibitor; miR, microRNA; H460R,
H460 cells with radioresistance. |
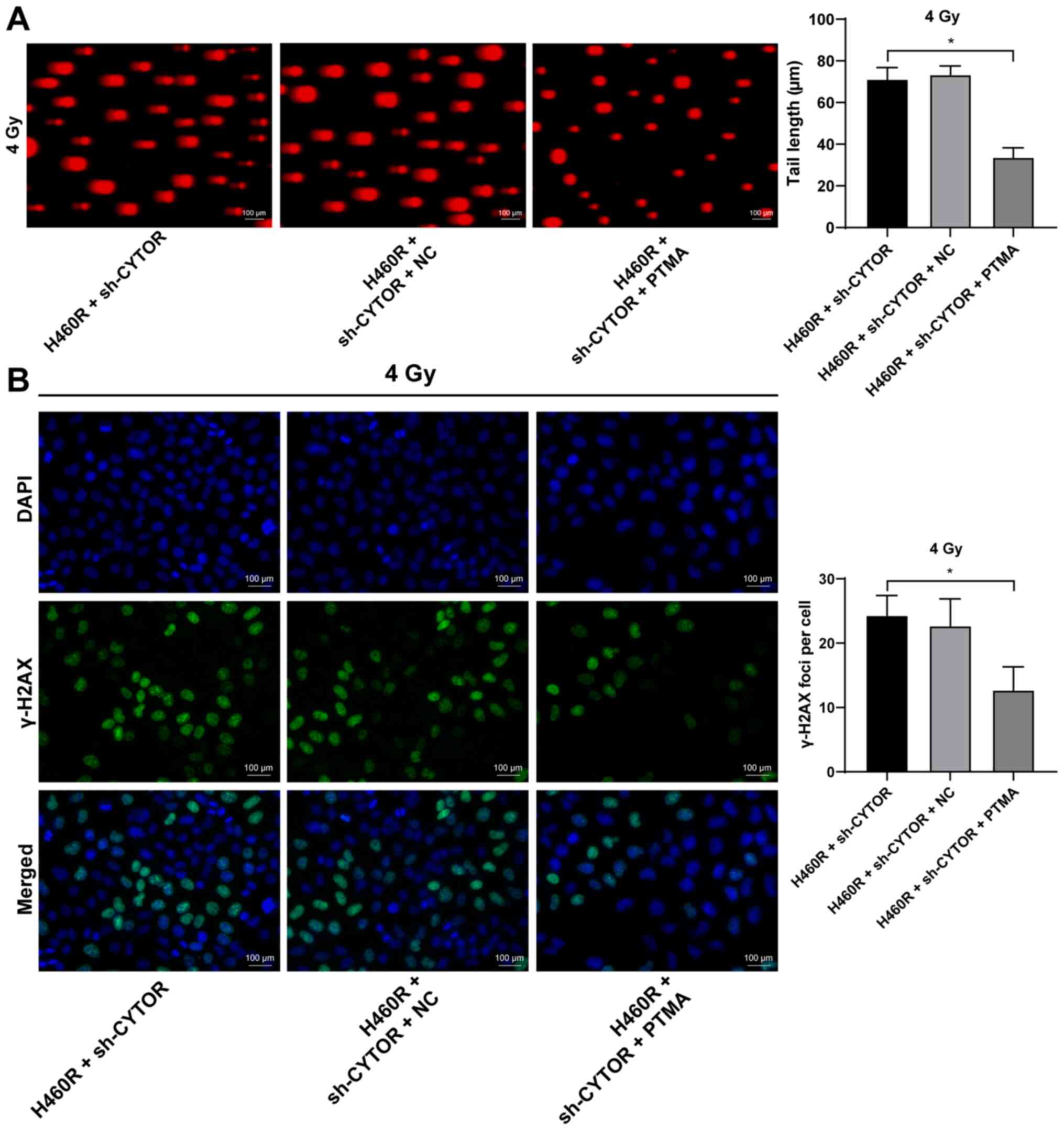
PTMA overexpression debilitates the
inhibitory effect of CYTOR depletion on NSCLC cell
radiosensitivity
To investigate the role of PTMA in the
CYTOR-regulated radiosensitivity of NSCLC cells, pcDNA3.1-PTMA was
constructed and transfected into H460R cells to upregulate PTMA
expression (P<0.05; Fig.
10A). It was indicated that overexpression of PTMA partially
reversed the reduced radioresistance of the sh-CYTOR-treated H460R
cells as demonstrated by the enhanced clonogenic ability of the
cells (Fig. 10B), shorter comet
assay tailing (Fig. 11A) and
diminished γ-H2AX immunofluorescence (Fig. 11B). These results suggested that
overexpression of PTMA retarded the inhibitory role of sh-CYTOR on
NSCLC cell radiosensitivity.
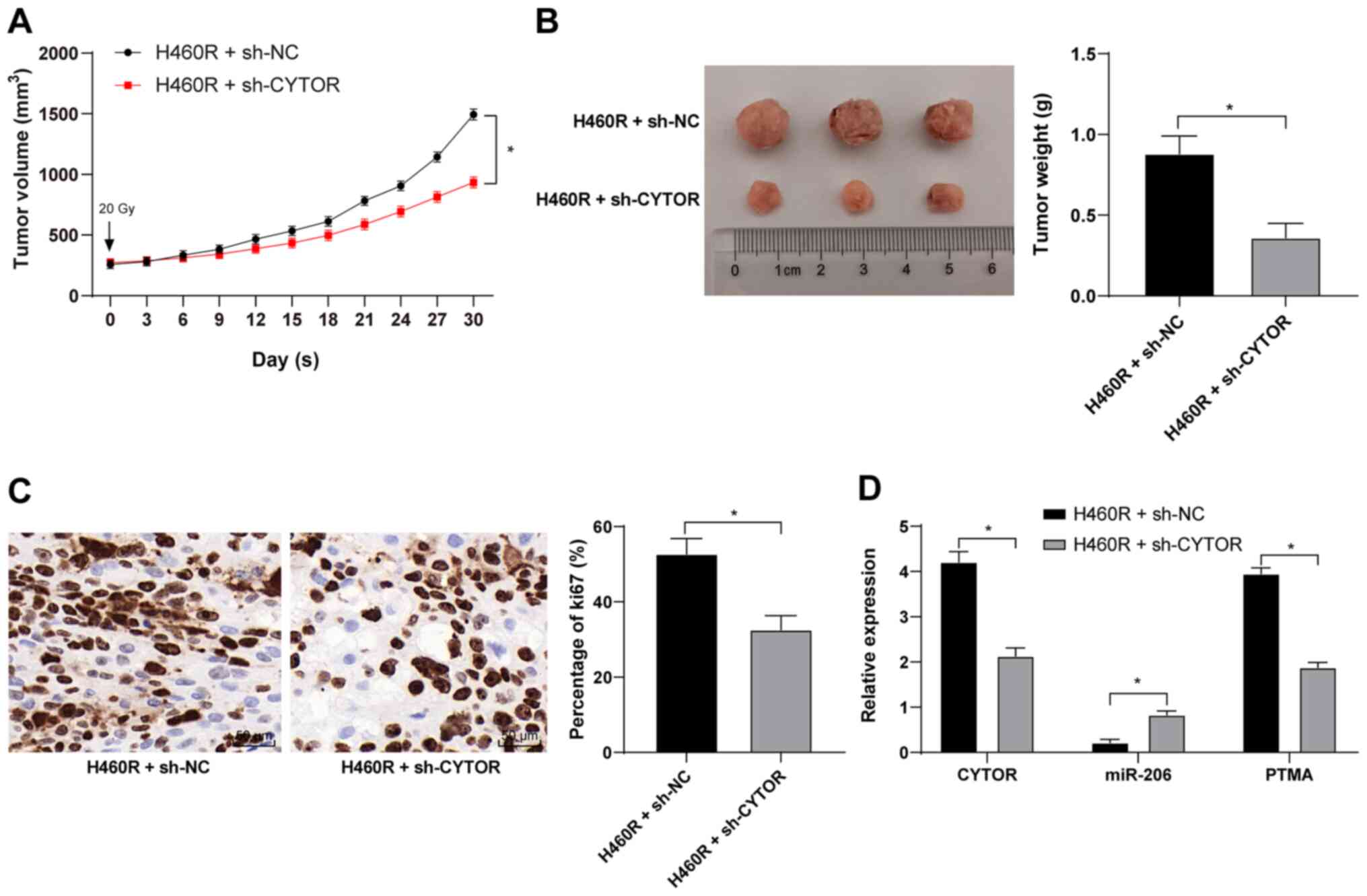
CYTOR knockdown in vivo enhances the
radiosensitivity of xenograft tumors in mice
Xenograft tumors were established in mice by
subcutaneous injection of differently treated H460R cells. After
the tumors were treated by irradiation, compared with the H460R +
sh-NC group, the H460R + sh-CYTOR group exhibited a reduced tumor
weight and size and ki67-positive rate (P<0.05; Fig. 12A-C), decreased expression of
CYTOR and PTMA in the tumors and enhanced miR-206 expression
(P<0.05; Fig. 12D). The above
results indicated that CYTOR enhances NSCLC radioresistance by
competitively binding miR-206 to upregulate PTMA at the
posttranscriptional level in vivo.
Discussion
Although significant advances have been achieved in
the field of medicine targeting LC, the prognostic repercussions
and survival rate of NSCLC remain disappointing and mortality is
high, partially due to escalated chemoresistance (42). lncRNAs may serve as tumor
promotors or inhibitors by modulating the growth and development of
a variety of cancer types, such as NSCLC (43). Although the specific regulatory
mechanisms of CYTOR in NSCLC have yet to be clarified, studies have
indicated that CYTOR is activated in multiple malignancies,
including gastric cancer, renal cell carcinoma and hepatocellular
carcinoma, and predicts metastatic tumors and unfavorable prognosis
(44). Therefore, CYTOR is an
oncogenic factor and marker of neoplasms. Hence, the present study
attempted to uncover the role of CYTOR in NSCLC, including the
molecular signaling involved in the relevant axis.
The most important finding of the present study was
that CYTOR was overexpressed in NSCLC and high expression of CYTOR
was associated with poor prognosis of patients with NSCLC. Early
studies discovered that CYTOR functions as a promoter of various
neoplasms, including LC, kidney cancer, gastric cancer, gallbladder
cancer and colon cancer, as it expedites cellular metastasis,
invasiveness and motility (45).
CYTOR acts as a driving factor in a considerable number of
malignancies by regulating cell biological behaviors, facilitating
lymph node metastasis and incurring relapse, consequently being
associated with poor prognosis (46,47). Of note, proactively expressed
CYTOR was commonly observed in tumors that received no treatment,
with large tumor volume and advanced stage NSCLC (48). However, CYTOR is a
well-acknowledged therapeutic target and biomarker in NSCLC and
CYTOR knockdown causes cell inactivation and even death (49). Furthermore, silencing of CYTOR
enhanced the radiosensitivity of NSCLC cells (50). In a previous study, high CYTOR
expression was inextricably linked to limited apoptosis and
enhanced drug resistance (13).
In addition, CYTOR knockout was able to restore chemosensitivity
and enhance the apoptotic rate in epithelial ovarian cancer
(51). In summary, silencing
CYTOR may represent a strategy for treating NSCLC from the
perspective of strengthening cell sensitivity to drugs or
radiotherapy.
Of note, CYTOR targeted miR-206 expression in NSCLC
cells. CYTOR is involved in ceRNA mechanisms and sponges miR-497 in
thyroid tumors, which increases tumor progression and metastasis,
while CYTOR depletion, in turn, restrains cancer cell growth,
viability and aggressiveness (52). CYTOR functions as an upstream gene
to sponge miR-193a/b-3p, augmenting hepatocellular carcinoma
(53). In a relevant study, CYTOR
was determined to be potent in sponging miR-195 in NSCLC, therefore
enhancing malignancy and impaired radiosensitivity of NSCLC
(50), suggesting that CYTOR
mediates NSCLC development by acting as a sponge in ceRNA
interactions. Similarly, miR-206, a crucial cytokine connecting
homeobox transcript antisense intergenic RNA and cyclin D1 in the
ceRNA network, was reduced in ovarian cancer and catalyzed cell
survival, motility and proliferation (54). In addition, when miR-206 was
competitively bound by metastasis-associated lung adenocarcinoma
transcription 1, cyclin-dependent kinase 9, a direct target of
miR-206, was activated to stimulate osteosarcoma augmentation
(55), strengthening the
feasibility of proceeding with research concerning miR-206 in
NSCLC. Subsequently, it was uncovered that miR-206 knockdown
neutralized the inhibitory effect of CYTOR on NSCLC cell
radiosensitivity. miR-206 contributes to quenching various types of
carcinoma, such as thyroid cancer, prostate cancer and ovarian
cancer, by restricting cellular pathogenesis, viability and
motility (56-58). miR-206 was also beneficial in
preventing drug resistance and inducing necrocytosis in papillary
thyroid carcinoma (59). In
addition, miR-206 inhibited aerobic glycolysis to ameliorate NSCLC
(60). Subsequently, the present
study indicated that, as a target of miR-206, PTMA overexpression
debilitated the inhibitory role of CYTOR depletion in NSCLC cell
radiosensitivity, as evidenced by the improved Ki67-positive rate.
As a downstream factor in the ceRNA network, the upregulation of
PTMA caused the outgrowth of exacerbated colorectal cancer
(61). Furthermore, PTMA
exhibited a negative association with the radiosensitivity of
colorectal cancer (41). In
addition, Ki67 is a dependable indicator of cancer cellular
duplication and migration (62).
Studies have suggested that in diverse cancers, Ki67 activation
usually concurs with upregulated CYTOR, low expression of miR-206
or promoted PTMA (63-65), which is consistent with the
results of the present study. Based on the above, the
CYTOR/miR-206/PTMA axis may be valuable in NSCLC research.
In conclusion, the present study illustrated that
silencing CYTOR potentiated NSCLC cell radiosensitivity by
upregulating miR-206 and suppressing PTMA. This suggests
therapeutic implications for NSCLC alleviation. Experimental
revelations and realistic applications in medical practice require
extensive validation. It is esteemed that the present results
contribute to NSCLC research.
There are certain limitations to the present study.
First, since the role of CYTOR in the radiosensitivity of NSCLC
cells was discussed, patients who did not receive any radiotherapy
were not included. This is also a limitation of the present study.
This point will also be considered for investigation in the future.
Furthermore, due to limitations regarding the sample volume, other
possible miRs or mRNAs downstream of CYTOR that may affect NSCLC
radiosensitivity were not fully investigated. Future research by
our group will aim to further identify the possible downstream
genes or axes of PTMA and in-depth experiments targeting other miRs
related to CYTOR will be designed. Finally, the present study
mainly focused on the effect of CYTOR on the radiosensitivity of
NSCLC and its downstream mechanism. Therefore, the present results
were all obtained with the background of radiation and the role of
CYPOR in common NSCLC cell lines was not explored. Certain studies
have reported the effect of CYPOR on NSCLC. One previous study
investigated the mechanism of CYTOR in the migration and invasion
of NSCLC cells and revealed that CYTOR promoted cell proliferation,
migration and invasion ability, and induced radioresistance in
NSCLC cells (50). Another study
assessed the role of CYTOR in NSCLC, including the cell cycle and
apoptosis and found that CYTOR knockdown also promoted cell
apoptosis and induced cell cycle arrest in G1 phase (12). Our group aims to further explore
the role of the CYPOR/miR-206/PTMA axis in the malignant behavior
of NSCLC cells and cancer metastasis in the future.
Supplementary Data
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
GJ and FZ conceived and designed the present study.
GJ, HY, ZL and FZ performed the experiments. GJ, HY, ZL and FZ
analyzed and interpreted the data. GJ, HY and FZ wrote, reviewed
and/or revised the manuscript. GJ and FZ confirm the authenticity
of all the raw data. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
This study was approved by the Ethics Committee of
Yantaishan Hospital (Yantai, China; accession no. YSLZ2021037).
Informed consent was obtained from all subjects. The animal
experiments were carried out in accordance with the requirements of
the guidelines for the use of experimental animals (38), with the approval of the
Institutional Animal Care and Use Committee at Yantaishan Hospital
(Yantai, China; accession no. YSLZ2021026). Significant efforts
were made to minimize both the number of animals used and their
respective suffering.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Chen Z, Fillmore CM, Hammerman PS, Kim CF
and Wong KK: Non-small-cell lung cancers: A heterogeneous set of
diseases. Nat Rev Cancer. 14:535–546. 2014. View Article : Google Scholar
|
|
2
|
Tandberg DJ, Tong BC, Ackerson BG and
Kelsey CR: Surgery versus stereotactic body radiation therapy for
stage I non-small cell lung cancer: A comprehensive review. Cancer.
124:667–678. 2018. View Article : Google Scholar
|
|
3
|
Rafei H, El-Bahesh E, Finianos A,
Nassereddine S and Tabbara I: Immune-based therapies for non-small
cell lung cancer. Anticancer Res. 37:377–387. 2017. View Article : Google Scholar
|
|
4
|
Elias R, Morales J and Presley C:
Checkpoint inhibitors for non-small cell lung cancer among older
adults. Curr Oncol Rep. 19:622017. View Article : Google Scholar
|
|
5
|
Gridelli C, Rossi A, Carbone DP, Guarize
J, Karachaliou N, Mok T, Petrella F, Spaggiari L and Rosell R:
Non-small-cell lung cancer. Nat Rev Dis Primers. May 21–2015.Epub
ahead of print. View Article : Google Scholar
|
|
6
|
Skřičková J, Kadlec B, Venclíček O and
Merta Z: Lung cancer. Cas Lek Cesk. 157:226–236. 2018.
|
|
7
|
Liang J, Lu T, Chen Z, Zhan C and Wang Q:
Mechanisms of resistance to pemetrexed in non-small cell lung
cancer. Transl Lung Cancer Res. 8:1107–1118. 2019. View Article : Google Scholar
|
|
8
|
Tabchi S, Kassouf E, Rassy EE, Kourie HR,
Martin J, Campeau MP, Tehfe M and Blais N: Management of stage III
non-small cell lung cancer. Semin Oncol. 44:163–177. 2017.
View Article : Google Scholar
|
|
9
|
Pennell NA, Arcila ME, Gandara DR and West
H: Biomarker testing for patients with advanced non-small cell lung
cancer: Real-world issues and tough choices. Am Soc Clin Oncol Educ
Book. 39:531–542. 2019. View Article : Google Scholar
|
|
10
|
Huang Q: Predictive relevance of ncRNAs in
non-small-cell lung cancer patients with radiotherapy: A review of
the published data. Biomarkers Med. 12:1149–1159. 2018. View Article : Google Scholar
|
|
11
|
Bhan A, Soleimani M and Mandal SS: Long
noncoding RNA and cancer: A New Paradigm. Cancer Res. 77:3965–3981.
2017. View Article : Google Scholar
|
|
12
|
Yu H and Li SB: Role of LINC00152 in
non-small cell lung cancer. J Zhejiang Univ Sci B. 21:179–191.
2020. View Article : Google Scholar
|
|
13
|
Yue B, Liu C, Sun H, Liu M, Song C, Cui R,
Qiu S and Zhong M: A positive feed-forward loop between
LncRNA-CYTOR and Wnt/β-catenin signaling promotes metastasis of
colon cancer. Mol Ther. 26:1287–1298. 2018. View Article : Google Scholar :
|
|
14
|
Moradi MT, Hatami R and Rahimi Z:
Circulating CYTOR as a potential biomarker in breast cancer. Int J
Mol Cell Med. 9:83–90. 2020.
|
|
15
|
Liu Y, Li M, Yu H and Piao H: lncRNA CYTOR
promotes tamoxifen resistance in breast cancer cells via sponging
miR 125a 5p. Int J Mol Med. 45:497–509. 2020.
|
|
16
|
Chen S, Yang M, Wang C, Ouyang Y, Chen X,
Bai J, Hu Y, Song M, Zhang S and Zhang Q: Forkhead box D1 promotes
EMT and chemoresistance by upregulating lncRNA CYTOR in oral
squamous cell carcinoma. Cancer Lett. 503:43–53. 2021. View Article : Google Scholar
|
|
17
|
Zhu H, Shan Y, Ge K, Lu J, Kong W and Jia
C: LncRNA CYTOR promotes pancreatic cancer cell proliferation and
migration by sponging miR-205-5p. Pancreatology. 20:1139–1148.
2020. View Article : Google Scholar
|
|
18
|
Li M, Wang Q, Xue F and Wu Y: lncRNA-CYTOR
works as an Oncogene through the CYTOR/miR-3679-5p/MACC1 axis in
colorectal cancer. DNA Cell Biol. 38:572–582. 2019. View Article : Google Scholar
|
|
19
|
Hu B, Yang XB, Yang X and Sang XT: LncRNA
CYTOR affects the proliferation, cell cycle and apoptosis of
hepatocellular carcinoma cells by regulating the
miR-125b-5p/KIAA1522 axis. Aging (Albany NY). 13:2626–2639. 2020.
View Article : Google Scholar
|
|
20
|
Petrek H and Yu AM: MicroRNAs in non-small
cell lung cancer: Gene regulation, impact on cancer cellular
processes, and therapeutic potential. Pharmacol Res Perspect.
7:e005282019. View
Article : Google Scholar
|
|
21
|
Shengnan J, Dafei X, Hua J, Sunfu F,
Xiaowei W and Liang X: Long non-coding RNA HOTAIR as a competitive
endogenous RNA to sponge miR-206 to promote colorectal cancer
progression by activating CCL2. J Cancer. 11:4431–4441. 2020.
View Article : Google Scholar
|
|
22
|
Cao HL, Liu ZJ, Huang PL, Yue YL and Xi
JN: lncRNA-RMRP promotes proliferation, migration and invasion of
bladder cancer via miR-206. Eur Rev Med Pharmacol Sci.
23:1012–1021. 2019.
|
|
23
|
Liao M and Peng L: MiR-206 may suppress
non-small lung cancer metastasis by targeting CORO1C. Cell Mol Biol
Lett. 25:222020. View Article : Google Scholar
|
|
24
|
Samaeekia R, Adorno-Cruz V, Bockhorn J,
Chang YF, Huang S, Prat A, Ha N, Kibria G, Huo D, Zheng H, et al:
miR-206 inhibits stemness and metastasis of breast cancer by
targeting MKL1/IL11 pathway. Clin Cancer Res. 23:1091–1103. 2017.
View Article : Google Scholar
|
|
25
|
Jiao D, Chen J, Li Y, Tang X, Wang J, Xu
W, Song J, Li Y, Tao H and Chen Q: miR-1-3p and miR-206 sensitizes
HGF-induced gefitinib-resistant human lung cancer cells through
inhibition of c-Met signalling and EMT. J Cell Mol Med.
22:3526–3536. 2018. View Article : Google Scholar
|
|
26
|
Weidle UH, Birzele F and Nopora A:
MicroRNAs as potential targets for therapeutic intervention with
metastasis of non-small cell lung cancer. Cancer Genomics
Proteomics. 16:99–119. 2019. View Article : Google Scholar
|
|
27
|
Zhu Y, Qi X, Yu C, Yu S, Zhang C, Zhang Y,
Liu X, Xu Y, Yang C, Jiang W, et al: Identification of prothymosin
alpha (PTMA) as a biomarker for esophageal squamous cell carcinoma
(ESCC) by label-free quantitative proteomics and Quantitative Dot
Blot (QDB). Clin Proteomics. 16:122019. View Article : Google Scholar
|
|
28
|
Sandow JJ, Rainczuk A, Infusini G, Makanji
M, Bilandzic M, Wilson AL, Fairweather N, Stanton PG, Garama D,
Gough D, et al: Discovery and validation of novel protein
biomarkers in ovarian cancer patient urine. Proteomics Clin Appl.
12:e17001352018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42:D92–D97. 2014. View Article : Google Scholar
|
|
30
|
Yan F, Zhao W, Xu X, Li C, Li X, Liu S,
Shi L and Wu Y: LncRNA DHRS4-AS1 inhibits the stemness of NSCLC
cells by sponging miR-224-3p and upregulating TP53 and TET1. Front
Cell Dev Biol 2020. 8:5852512020. View Article : Google Scholar
|
|
31
|
Li Z, Qu Z, Wang Y, Qin M and Zhang H:
miR-101-3p sensitizes non-small cell lung cancer cells to
irradiation. Open Med (Wars). 15:413–423. 2020. View Article : Google Scholar
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
33
|
Zamani M, Etebari M and Moradi SH: The
increment of genoprotective effect of melatonin due to 'Autooptic'
effect versus the genotoxicity of mitoxantrone. J Biomed Phys Eng.
10:771–782. 2020.
|
|
34
|
Paraskevopoulou MD, Vlachos IS, Karagkouni
D, Georgakilas G, Kanellos I, Vergoulis T, Zagganas K, Tsanakas P,
Floros E, Dalamagas T, et al: DIANA-LncBase v2: Indexing microRNA
targets on non-coding transcripts. Nucleic Acids Res. 44:D231–D238.
2016. View Article : Google Scholar
|
|
35
|
Jeggari A, Marks DS and Larsson E:
miRcode: A map of putative microRNA target sites in the long
non-coding transcriptome. Bioinformatics. 28:2062–2063. 2012.
View Article : Google Scholar
|
|
36
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
eLife. 4:20152015. View Article : Google Scholar
|
|
37
|
Huang HY, Lin YC, Li J, Huang KY, Shrestha
S, Hong HC, Tang Y, Chen YG, Jin CN, Yu Y, et al: miRTarBase 2020:
Updates to the experimentally validated microRNA-target interaction
database. Nucleic Acids Res. 48:D148–D154. 2020.
|
|
38
|
Wang L, Tong X, Zhou Z, Wang S, Lei Z,
Zhang T, Liu Z, Zeng Y, Li C, Zhao J, et al: Circular RNA
hsa_circ_0008305 (circPTK2) inhibits TGF-β-induced
epithelial-mesenchymal transition and metastasis by controlling
TIF1γ in non-small cell lung cancer. Mol Cancer. 17:1402018.
View Article : Google Scholar
|
|
39
|
Jones-Bolin S: Guidelines for the care and
use of laboratory animals in biomedical research. Curr Protoc
Pharmacol 2012 Appendix. 4:4B2012.
|
|
40
|
Wang T, Dong XM, Zhang FL and Zhang JR:
miR-206 enhances nasopharyngeal carcinoma radiosensitivity by
targeting IGF1. Kaohsiung J Med Sci. 33:427–432. 2017. View Article : Google Scholar
|
|
41
|
Ojima E, Inoue Y, Miki C, Mori M and
Kusunoki M: Effectiveness of gene expression profiling for response
prediction of rectal cancer to preoperative radiotherapy. J
Gastroenterol. 42:730–736. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chiu YH, Hsu SH, Hsu HW, Huang KC, Liu W,
Wu CY, Huang WP, Chen JY, Chen BH and Chiu CC: Human non small cell
lung cancer cells can be sensitized to camptothecin by modulating
autophagy. Int J Oncol. 53:1967–1979. 2018.
|
|
43
|
Osielska MA and Jagodziński PP: Long
non-coding RNA as potential biomarkers in non-small-cell lung
cancer: What do we know so far? Biomed Pharmacother. 101:322–333.
2018. View Article : Google Scholar
|
|
44
|
Liang J, Wei X, Liu Z, Cao D, Tang Y, Zou
Z, Zhou C and Lu Y: Long noncoding RNA CYTOR in cancer: A TCGA data
review. Clin Chim Acta. 483:227–233. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yu Y, Yang J, Li Q, Xu B, Lian Y and Miao
L: LINC00152: A pivotal oncogenic long non-coding RNA in human
cancers. Cell Prolif. 50:20172017.
|
|
46
|
Quan FY, Jiang J, Zhai YF, Li B, Wu XH and
Nie W: The prognostic effect of LINC00152 for cancer: A
meta-analysis. Oncotarget. 8:75427–75433. 2017. View Article : Google Scholar
|
|
47
|
Mao Y, Tie Y, Du J and He J:
LINC00152-promotes the proliferation of gastric cancer cells by
regulating B-cell lymphoma-2. J Cell Biochem. 120:3747–3756. 2019.
View Article : Google Scholar
|
|
48
|
Li N, Feng XB, Tan Q, Luo P, Jing W, Zhu
M, Liang C, Tu J and Ning Y: Identification of circulating long
noncoding RNA Linc00152 as a novel biomarker for diagnosis and
monitoring of non-small-cell lung cancer. Dis Markers.
2017:74396982017. View Article : Google Scholar
|
|
49
|
Feng S, Zhang J, Su W, Bai S, Xiao L, Chen
X, Lin J, Reddy RM, Chang AC, Beer DG and Chen G: Overexpression of
LINC00152 correlates with poor patient survival and knockdown
impairs cell proliferation in lung cancer. Sci Rep. 7:29822017.
View Article : Google Scholar
|
|
50
|
Zhang J and Li W: Long noncoding RNA CYTOR
sponges miR-195 to modulate proliferation, migration, invasion and
radiosensitivity in nonsmall cell lung cancer cells. Biosci Rep.
38:20182018. View Article : Google Scholar
|
|
51
|
Zou H and Li H: Knockdown of long
non-coding RNA LINC00152 increases cisplatin sensitivity in ovarian
cancer cells. Exp Ther Med. 18:4510–4516. 2019.
|
|
52
|
Sun Z, Guo X, Zang M, Wang P, Xue S and
Chen G: Long non-coding RNA LINC00152 promotes cell growth and
invasion of papillary thyroid carcinoma by regulating the
miR-497/BDNF axis. J Cell Physiol. 234:1336–1345. 2019. View Article : Google Scholar
|
|
53
|
Ma P, Wang H, Sun J, Liu H, Zheng C, Zhou
X and Lu Z: LINC00152 promotes cell cycle progression in
hepatocellular carcinoma via miR-193a/b-3p/CCND1 axis. Cell Cycle.
17:974–984. 2018. View Article : Google Scholar
|
|
54
|
Chang L, Guo R, Yuan Z, Shi H and Zhang D:
LncRNA HOTAIR regulates CCND1 and CCND2 expression by sponging
miR-206 in ovarian cancer. Cell Physiol Biochem. 49:1289–1303.
2018. View Article : Google Scholar
|
|
55
|
Ren D, Zheng H, Fei S and Zhao JL: MALAT1
induces osteosarcoma progression by targeting miR-206/CDK9 axis. J
Cell Physiol. 234:950–957. 2018. View Article : Google Scholar
|
|
56
|
Wang Y, Xu H, Si L, Li Q, Zhu X, Yu T and
Gang X: MiR-206 inhibits proliferation and migration of prostate
cancer cells by targeting CXCL11. Prostate. 78:479–490. 2018.
View Article : Google Scholar
|
|
57
|
Wang P, Gu J, Wang K, Shang J and Wang W:
miR-206 inhibits thyroid cancer proliferation and invasion by
targeting RAP1B. J Cell Biochem. 120:18927–18936. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Dai C, Xie Y, Zhuang X and Yuan Z: MiR-206
inhibits epithelial ovarian cancer cells growth and invasion via
blocking c-Met/AKT/mTOR signaling pathway. Biomed Pharmacother.
104:763–770. 2018. View Article : Google Scholar
|
|
59
|
Liu F, Yin R, Chen X, Chen W, Qian Y, Zhao
Y, Jiang Y, Ma D, Hu T, Yu T, et al: Over-expression of miR-206
decreases the Euthyrox-resistance by targeting MAP4K3 in papillary
thyroid carcinoma. Biomed Pharmacother. 114:1086052019. View Article : Google Scholar
|
|
60
|
Jia KG, Feng G, Tong YS, Tao GZ and Xu L:
miR-206 regulates non-small-cell lung cancer cell aerobic
glycolysis by targeting hexokinase 2. J Biochem. 167:365–370. 2020.
View Article : Google Scholar
|
|
61
|
Yang L, Sun H, Liu X, Chen J, Tian Z, Xu
J, Xiang B and Qin B: Circular RNA hsa_circ_0004277 contributes to
malignant phenotype of colorectal cancer by sponging miR-512-5p to
upregulate the expression of PTMA. J Cell Physiol. Jan 21–2020.Epub
ahead of print. View Article : Google Scholar
|
|
62
|
Yang C, Zhang J, Ding M, Xu K, Li L, Mao L
and Zheng J: Ki67 targeted strategies for cancer therapy. Clin
Transl Oncol. 20:570–575. 2018. View Article : Google Scholar
|
|
63
|
Wierzbicka-Tutka I, Sokołowski G,
Bałdys-Waligórska A, Adamek D, Radwańska E and Gołkowski F:
Prothymosin-alpha and Ki-67 expression in pituitary adenomas.
Postepy Hig Med Dosw. 70:1117–1123. 2016. View Article : Google Scholar
|
|
64
|
Wang Y, Li M, Dong C, Ma Y, Xiao L, Zuo S,
Gong Y, Ren T and Sun B: Linc00152 knockdown inactivates the
Akt/mTOR and Notch1 pathways to exert its anti-hemangioma effect.
Life Sci. 223:22–28. 2019. View Article : Google Scholar
|
|
65
|
Du L, Huang GH, Mou KJ, Xiang Y, Tang JH,
Xu W, Xia SL, Zhao JN and Lv SQ: MiR-206 is down-regulated and
suppresses cell proliferation by targeting FOXP1 in brain gliomas.
Int J Clin Exp Pathol. 11:3405–3415. 2018.
|