Introduction
Protein disulfide isomerases (PDIs) are involved in
a wide range of biological pathways as mediators of oxidative
folding in the endoplasmic reticulum (1,2),
in the formation, breakage, and rearrangement of disulfide bonds
(1,3), and in the regulation of apoptosis,
exerting both pro-apoptotic and pro-survival effects (4,5).
The PDI family member prolyl 4-hydroxylase β (P4HB, also known as
PDIA1) functions as a chaperone that assists protein folding by
inhibiting the aggregation of partially folded or damaged
polypeptides (6). Recent findings
have implicated PDIA1 in the processes of antigen processing and
presentation, immunomodulation and tumor immunorecognition
(4,7-9).
PDIA1 has been shown to play an important role in
various stages of carcinogenesis in a wide variety of cancers,
being involved in the early stages of carcinogenesis possibly by
contributing to the management of misfolded and non-functional
proteins (5) as well as in the
evasion of the immunosurveillance (4). Evidence has been presented that PDI
family members are involved in the proliferation and metastasis of
brain, kidney, and lung cancers (10,11). Overexpression of PDI family
members has been positively correlated with metastasis and invasion
of breast cancer (12). For
instance, significantly higher PDIA1 levels have been observed in
axillary lymph node metastatic breast tumor compared to primary
breast tumors (13). In addition,
PDIA1 mRNA levels have been found to be positively correlated with
malignant glioma metastasis and invasion (14) and the epithelial-mesenchymal
transition (EMT) of liver cancer cells (15-17). The potential molecular mechanisms
by which PDIA1 contributes to metastasis include the regulation of
the hypoxia-inducible factor-1α (HIF-1α) pathway (17,18), and induction of matrix
metalloproteinase-9 (MMP-9) secretion (19) or other metalloproteases such as
the disintegrin and metalloprotease domain family member ADAM
metallopeptidase domain 17 (ADAM17) (20).
PDIA1 is mainly localized in the endoplasmic
reticulum, but cell surface and nuclear localization under certain
micro-environmental conditions have also been reported (21,22). Distinct PDIA1 subcellular
locations determine diverse PDIA1 signaling networks including the
regulation of the cellular redox state which in turn regulates its
function (2). Nuclear PDIA1 has
been shown to play a role in the regulation of the transcriptional
activity of redox responsive transcription factors (23,24). Indeed, PDIA1 has been shown to
directly interact and function as an estrogen receptor (ER)α
transcriptional cofactor (25,26). In addition, PDIA1 suppresses
nuclear factor (NF)-κB transcriptional activity (27,28) and regulates p53 protein stability
(29,30). These observations raise the
possibility that PDIA1 regulates the crosstalk between ERα and
NF-κB (31). Other
redox-responsive transcription factors associated with PDIA1
include HIF-1α (18) and nuclear
factor (erythroid-derived 2)-like 2 (NRF2) (32). PDIA1 is also implicated in
regulating the transcription of the gene targets of thyroid hormone
receptors α and β (33,34).
Several endoplasmic reticulum chaperones including
PDI have been detected in the mitochondria (35) and the mitochondrial and the
endoplasmic reticulum contact sites (MERC) (36). The function of the mitochondrial
PDIs has not been elucidated clearly but there is evidence to
suggest that PDIs regulate the exchange of Ca2+ between
the endoplasmic reticulum and mitochondria, mitochondrial energy
generation (37) and the immune
response to tumor cells (38),
suggesting that PDIs might affect carcinogenesis via multiple
pathways. Given that PDIA1 protein levels are breast cancer
subtype-specific (4,13) and taking into account the fact
that PDIA1 functions as a co-modulator for several transcription
factors including ERα (26), it
was hypothesized that PDIA1 would differentially affect
carcinogenesis in ERα-positive vs. ERα-negative breast cancer
cells.
In the present study, PDIA1 gene expression
was silenced in the ERα-positive MCF-7 and the ERα-negative
MDA-MB-231 breast cancer cells and the expression of genes under
conditions stimulating reactive oxygen species (ROS) generation
(treatment with interferon-γ (IFN-γ) (39) or etoposide (ETO) (40) was followed using RNA-seq.
Kaplan-Meier survival curves of the modulated genes detected by
RNA-seq were generated to identify whether high or low expression
of the upregulated or downregulated genes in the ERα-positive
(MCF-7) or triple-negative breast cancer (TNBC) cells (MDA-MB-231)
had positive or negative effect on the overall survival (OS) of the
ERα-positive or ERα-negative breast cancer patients, respectively.
The results indicated that PDIA1, in the ERα-positive breast cancer
patients, suppresses carcinogenesis by negatively regulating
catabolic processes of ROS. PDIA1 was found to induce
carcinogenesis by affecting cell cycle progression at the G2/M
checkpoint alongside spindle formation in the ERα-positive breast
cancer patients. In the ERα-negative breast cancer patients, PDIA1
was found to prevent tumor development by regulating the response
to NF-κB signaling, mitochondrial biogenesis, glycolysis and the
process of metastasis. PDIA1 induced breast cancer progression in
ERα-negative breast cancer patients by modifying the immune
response, cytokine signaling and calcium homeostasis. The findings
reported here shed new light on the differential pathways inducing
carcinogenesis in the ERα-positive and ERα-negative breast cancer
patients and could assist in the identification of selectively
beneficial therapeutic targets for ERα-positive or ERα-negative
patients.
Materials and methods
Cell culture
The human breast carcinoma cell lines MCF-7
(expressing ERα and wild-type p53) and MDA-MB-231 [ERα-negative,
bearing mutated p53 (R280K)] were obtained from the European
Collection of Cell Cultures (ECACC) and maintained in Dulbecco's
modified Eagle's medium (DMEM) (Sigma-Aldrich; Merck KGaA)
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) and 1% penicillin/streptomycin (Lonza Group,
Ltd.) at 37°C in a humidified atmosphere containing 5%
CO2 until they reached 70% confluency. Where indicated,
cells were treated with 10 ng/ml IFN-γ (Sigma-Aldrich; Merck KGaA)
for 24 h or 10 µM etoposide (ETO) for 24 h (Sigma-Aldrich;
Merck KGaA).
siRNA transfection
Concentration of 5 µM of the siGENOME PDIA1
siRNA and 5 µM of the siGENOME non-targeting siRNA pool was
added to each well containing 2×105 cells in DMEM and
incubated for 72 h according to the suppliers' instructions
(Dharmacon, UK) as described previously (4). The sequences of the siRNAs used are
indicated as follows: siGE-NOME PDIA1-targeting siRNA pool: ACA GGA
CGG UCA UUG AUU A, GGA CGG UCA UUG AUU ACA A, CCA AGAG UGU GUC UGA
CUA, and CAGAGAGGAUCACAGAGUU; siGENOME non-targeting (scramble)
siRNA pool: UAG CGA CUA AAC ACA UCA A, UAA GGC UAU GAA GAG AUA C,
AUG UAU UGG CCU GUA UUA G, and AUG AAC GUG AAU UGC UCA A.
Western blotting
Cellular extracts from MCF-7 and MDA-MB-231 and 30
µg of total protein per sample were loaded on a 20% precast
polyacrylamide gel and transferred to a PVDF membrane. The
membranes were then incubated in 5% milk in PBS-0.1% Tween-20 (v/v)
with anti-P4HB anti-body (Santa Cruz Biotechnology, sc-74551)
(dilution 1:500) or β-actin (Sigma-Aldrich; Merck KGaA; A1978)
(dilution 1:10,000) overnight at 4°C. After incubation with
secondary anti-mouse immunoglobulin G conjugated to horseradish
peroxidase (GE Healthcare) (dilution 1:1,000) in 2.5% milk in
PBS-0.1% Tween-20 (v/v) for 1 h at 25°C, the protein bands were
visualized using the ChemiDoc MP imaging system (Bio-Rad
Laboratories) The blots were quantified using ImageJ version 1.51
(National Institutes of Health).
RNA extraction
MCF-7 and MDA-MB-231 breast cancer cell lines were
seeded in 6-well plates at a concentration of 3×105
cells per well for 24 h after treatment. RNA was extracted
according to the RNeasy Mini Kit supplier instructions (Qiagen,
UK). After collecting RNA, the samples were maintained at −80°C
before sequencing carried out by the Next Generation Sequencing
Facility (University of Leeds, UK).
RNA sequencing
Initial alignment of RNA transcript counts, sequence
short reads, and normalization were carried out by the Next
Generation Sequencing Facility (University of Leeds, UK). Sequence
data in Fastq format were quality-checked using FastQC software
(http://www.bioinformatics.babraham.ac.uk/projects/fastqc/).
Cutadapt version 1.16 software (https://cutadapt.readthedocs.org/en/stable/) was used
to trim poor quality bases (Phred quality score <20) and
contaminating adapter sequences from raw reads. Reads trimmed to
fewer than 30 were discarded. Reads were aligned to a human (hg38)
genome reference sequences, obtained from the UCSC database
(41) using the splicing-aware
STAR aligner (42). STAR aligner
was run with known splice junctions supplied in GTF file format,
obtained from the UCSC database using Table Browser tool (43,44). The resulting alignments in BAM
file format were checked for quality using QualiMap software
(44) and Picard tools version
1.90 (http://picard.sourceforge.net).
Picard tools also used to mark PCR/Optical duplicate alignments.
BAM files were indexed using Samtools software (45) and visualised using IGV browser
(46) to check for genomic DNA
contamination and the presence of PCR duplicates. Bioconductor R
package RSubread (47) was used
to extract raw sequenced fragment counts per transcript using the
RefSeq hg38 annotation dataset used by STAR during alignment.
Multi-mapping read pairs were counted as a fraction of all
equivalent alignments. Read count data were generated with the
inclusion of reads marked as PCR/optical duplicates.
Filtration of the gene list
Significantly downregulated or upregulated genes
(Benjamini-Hochberg <0.01) in the PDIA1-silenced MCF-7
and MDA-MB-231 treated with IFN-γ or ETO compared to MCF-7 and
MDA-MB-231 cells expressing PDIA1 treated with IFN-γ
(39) or ETO (40) were selected. To visualize the
overall changes in gene expression between PDIA1-silenced
and scramble-transfected MCF-7 and MDA-MB-231 cells, the log value
of the change in gene expression was graphed against the normalized
read count for each transcript using the plotMA function of the
DeSeq2 R package (47). The read
count data required for each pairwise analysis was imported into
the R package DeSeq2 and the effect size of each sample's library
was defined and used to normalize the read count data.
Kaplan-Meier overall survival
analysis
To understand the clinical importance of the genes
identified and filtered from the RNA-seq data, the Kaplan-Meier
Plotter website (http://kmplot.com/analysis/) was utilized. For this
purpose, Kaplan-Meier (KM) plots for ERα-positive and ERα-negative
patients were generated for the genes identified to be upregulated
or downregulated in the RNA-seq data (48). The following parameters were
selected: Survival: OS, Split patients by: upper quartile,
Follow-up threshold: 240 months, Probe set option: only JetSet best
probe sets (49), ER status-IHC:
ER-positive (for the genes upregulated or down regulated in MCF-7
cells) or ER negative (for the genes upregulated or downregulated
in MDA-MB-231 cells) ER status-array: ER positive (for the genes
upregulated or downregulated in MCF-7 cells) and ER negative (for
the genes upregulated or down regulated in MDA-MB-231 cells). Log
rank P-values <0.05 for the KM plots of all genes was considered
statistically significant.
cBioPortal analysis
The Pearson correlation coefficient for the
comparison of PDIA1 mRNA levels with those of the genes
upregulated or downregulated in the PDIA1-silenced MCF-7
cells treated with etoposide (ETO) or interferon γ (IFN-γ) in
breast cancer patients was explored using the cBioPortal platform
(http://cbioportal.org) in combination with data
provided in the Breast Cancer (SMC 2018) database using the mRNA
expression z-scores relative to all sample (RNA seq TPM-168
samples) options.
Functional annotation
In order to understand the biological importance of
the gene lists generated from the previous section, the GeneCards
tool (50) was used to identify
linked genes to the genes listed in Tables I and II. After this, Metascape tool (51) was employed. Gene lists were
entered in the metascape tool using the Official Gene Symbol as the
identifier and Homo sapiens as the species.
 | Table IGenes upregulated or downregulated in
interferon γ (IFN-γ) or etoposide (ETO)-treated MCF-7 cells in
which PDIA1 gene expression was silenced compared to
PDIA1-expressing IFN-γ- or ETO-treated MCF-7 cells. |
Table I
Genes upregulated or downregulated in
interferon γ (IFN-γ) or etoposide (ETO)-treated MCF-7 cells in
which PDIA1 gene expression was silenced compared to
PDIA1-expressing IFN-γ- or ETO-treated MCF-7 cells.
MCF-7 cells
|
|---|
| Upregulated | Downregulated |
|---|
| L1CAM | AURKA |
| MAGED1 | BUB1 |
| MDK | CCNB2 |
| PHLDA2 | CDC25A |
| PRDX2 | CDC25C |
| PTPRS | CENPF |
| PYCR1 | CKAP5 |
| RHOD | E2F8 |
| SMAD6 | ESPL1 |
| TKT | FEN1 |
| UCP2 | GTSE1 |
| JUND |
| KIF11 |
| KIF20A |
| NEK2 |
| NUSAP1 |
| PKMYT1 |
| RACGAP1 |
| RANBP1 |
| S100PBP |
| TACC3 |
| TRIP13 |
| TUBB |
| UBE2C |
| Table IIGenes upregulated or downregulated in
interferon γ (IFN-γ) or etoposide (ETO)-treated MDA-MB-231 cells in
which PDIA1 gene expression was silenced compared to
PDIA1 expressing IFN-γ- or ETO-treated MDA-MB-231 cells. |
Table II
Genes upregulated or downregulated in
interferon γ (IFN-γ) or etoposide (ETO)-treated MDA-MB-231 cells in
which PDIA1 gene expression was silenced compared to
PDIA1 expressing IFN-γ- or ETO-treated MDA-MB-231 cells.
MDA-MB-231 cells
|
|---|
| Upregulated | Downregulated |
|---|
| HSP90B1 | CHCHD3 |
| IGFBP3 | PTPRJ |
| LAPTM5 | VMA21 |
| RNF213 | XIAP |
| SYNE1 | |
Identification of repurposed drugs
Following the identification of genes important for
breast cancer patient survival, the potential clinical targeting of
each gene was explored via DRUGSURV (52). Genes exerting oncogenic function
that were identified as statistically significant for patient OS
via KM Plotter were queried in turn, and the approved drugs that
targeted the genes directly or indirectly were recorded.
Results
Since PDIA1 regulates the transcriptional activity
of ERα, RNA-seq was employed to investigate whether this
oxidoreductase differentially affected gene expression in the
ERα-positive (MCF-7) vs. the ERα-negative (MDA-MB-231) breast
cancer cells. The differences in the gene expression of genes
involved in the oxidative stress pathways were explored in cells
treated with either IFN-γ or ETO, both of which are known to induce
ROS generation (39,40).
Genes upregulated and downregulated in
MCF-7 and MDA-MB-231 breast cancer cells in the absence of PDIA1
under oxidative stress conditions
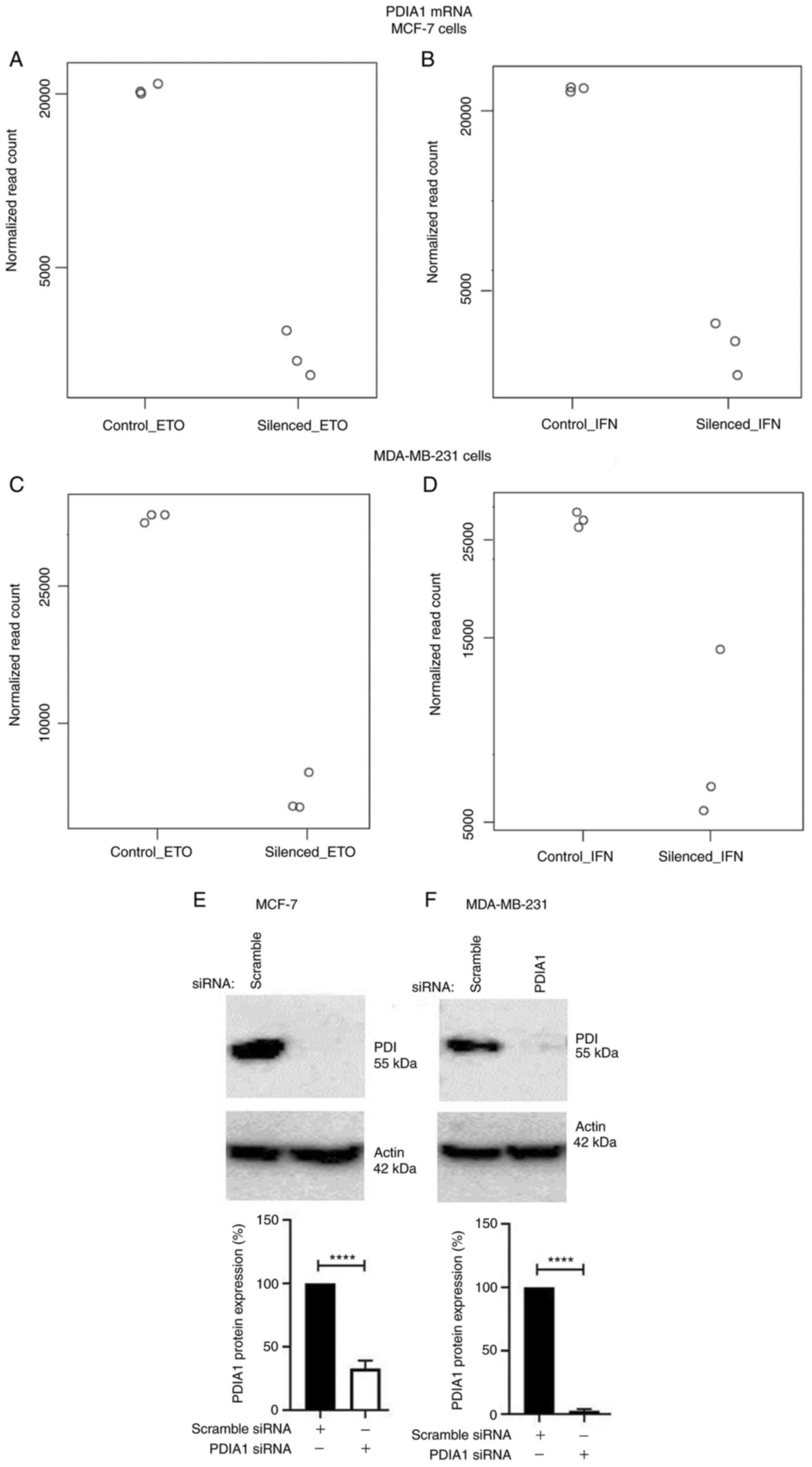
Total mRNA obtained from MCF-7 (Fig. 1A and B) or MDA-MB-231 (Fig. 1C and D) cells transfected with
either scramble or specific siRNA to silence PDIA1 gene
expression and treated with either ETO (Fig. 1A and C) or IFN-γ (Fig. 1B and D) was submitted to RNA-seq
analysis. The transfection efficiency for the PDIA1-siRNA
transfected MCF-7 cells compared to the transfection efficiency for
the scramble siRNA was 67.13% (Fig.
1E). The transfection efficiency for the PDIA1-siRNA
transfected MDA-MB-231 cells compared to the transfection
efficiency for the scramble siRNA was 96.67% (Fig. 1F).
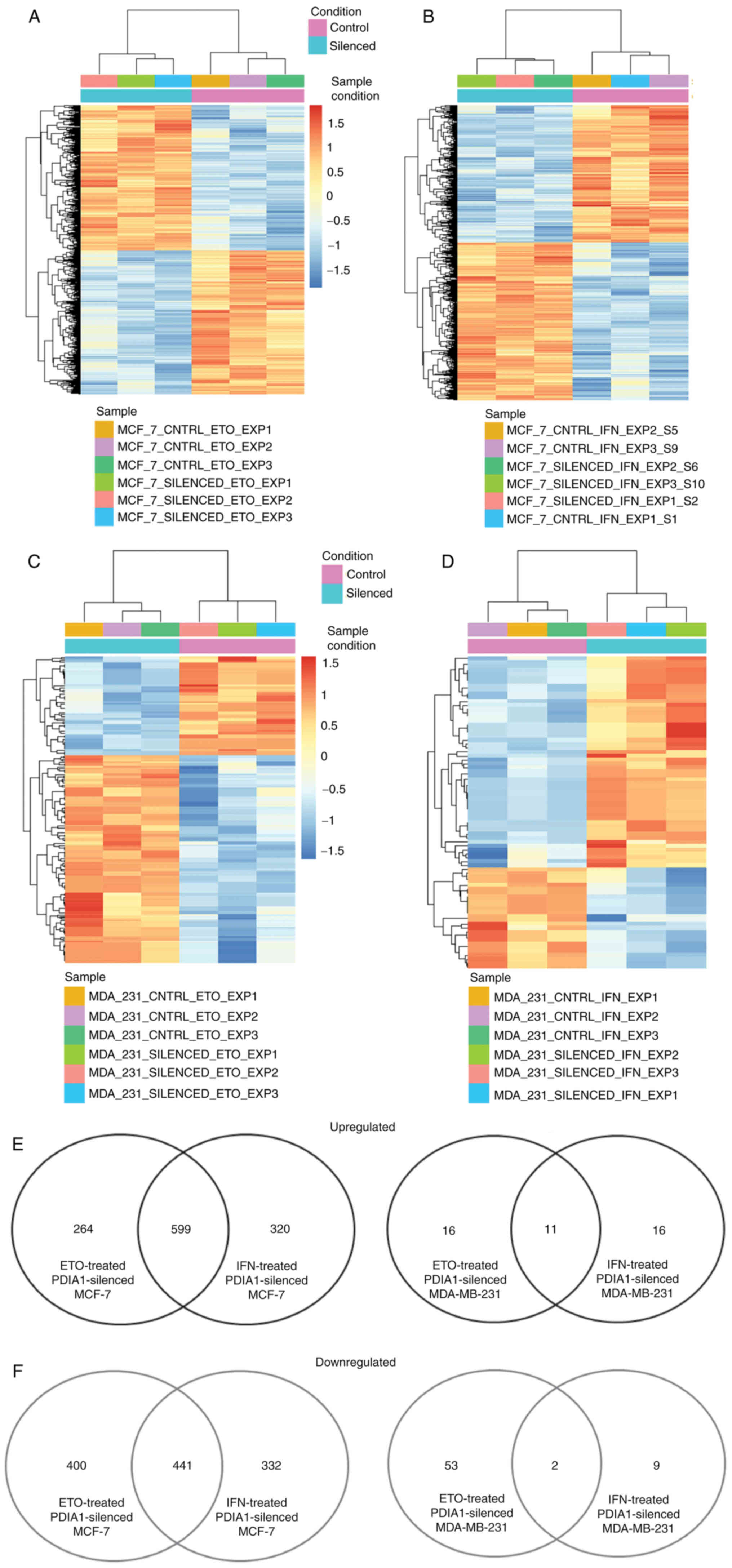
The total number of genes exhibiting statistically
significant downregulation in ETO-treated MCF-7 cells in which
PDIA1 gene expression had been silenced compared with
ETO-treated MCF-7 cells transfected with scramble siRNA was 841,
and the total number of genes upregulated under these conditions
was 863 (Fig. 2A). The total
number of genes exhibiting statistically significant
(P-adj<0.01) downregulation in IFN-γ-treated MCF-7 cells in
which PDIA1 gene expression had been silenced compared with
IFN-γ-treated MCF-7 cells transfected with scramble siRNA was 773,
and the total number of genes upregulated under these conditions
was 919 (Fig. 2B). The total
number of genes exhibiting statistically significant downregulation
in ETO-treated MDA-MB-231 cells in which PDIA1 gene
expression had been silenced compared with ETO-treated MDA-MB-231
cells transfected with scramble siRNA was 55, whereas the total
number of genes upregulated in these cells under the same
conditions was 27 (Fig. 2C). In
the IFN-γ-treated MDA-MB-231 cells in which PDIA1 gene
expression had been silenced compared with IFN-γ-treated MDA-MB-231
cells transfected with scramble siRNA the total number of the
downregulated genes was 11, whereas the total number of genes
upregulated in these cells under the same conditions was 27
(Fig. 2D). The numbers of
upregulated and downregulated genes are presented in Fig. 2E and F, respectively.
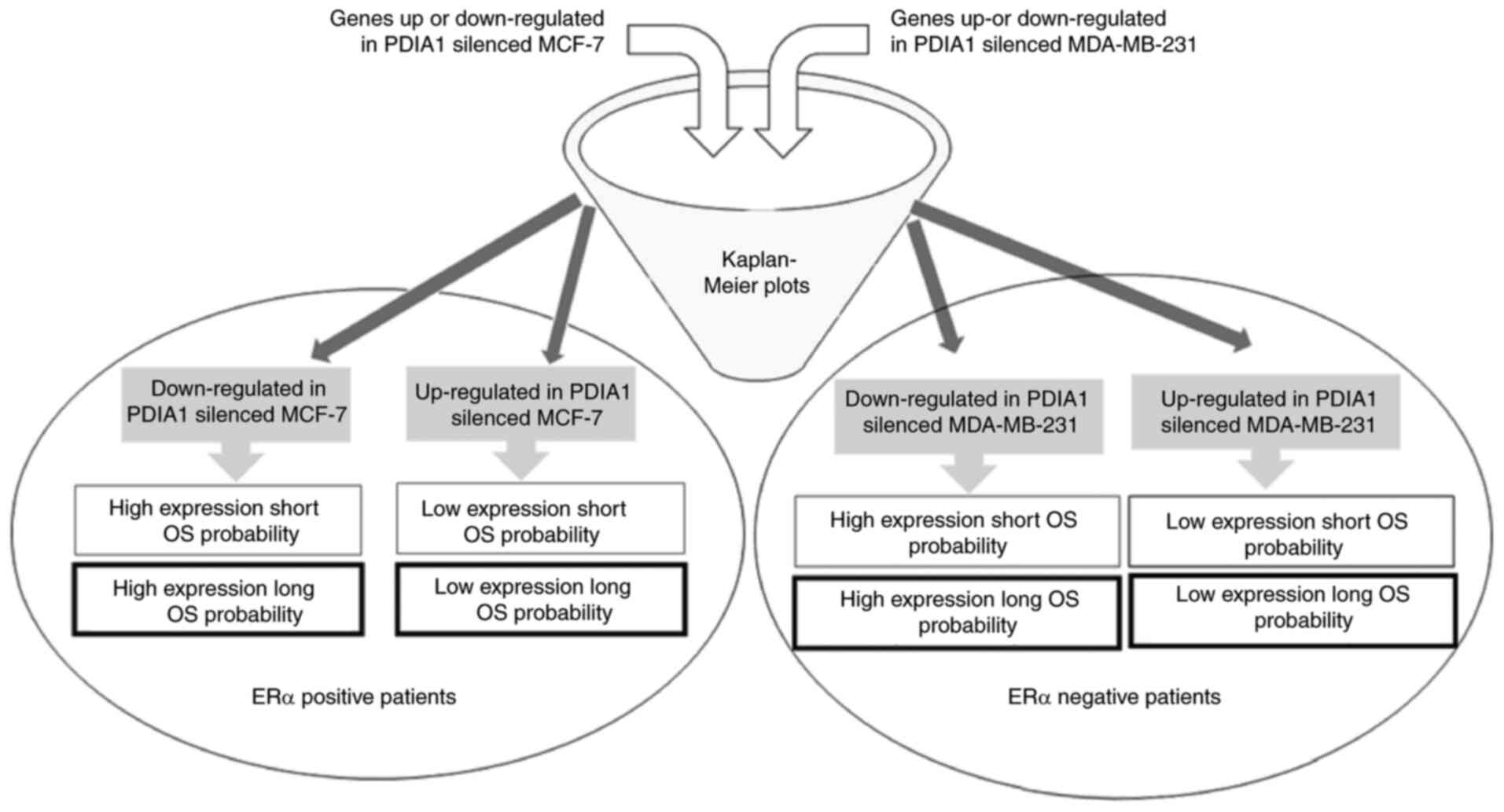
Clinical significance of the genes
modulated in IFN-γ- or ETO-treated and PDIA1-silenced MCF-7
cells
To shed light on the clinical significance of the
aforementioned findings, KM curves for the upregulated or
downregulated genes in the PDIA1-silenced compared with the
PDIA1-expressing and either IFN-γ- or ETO-treated MCF-7 and
MDA-MB-231 cells were plotted in the patients with ERα-positive and
ERα-negative breast cancer, respectively, as described in the
summary of the experimental design (Fig. 3). The upregulated or downregulated
genes in MCF-7 cells under the conditions described above that
exhibited statistically significant (P<0.05) KM results are
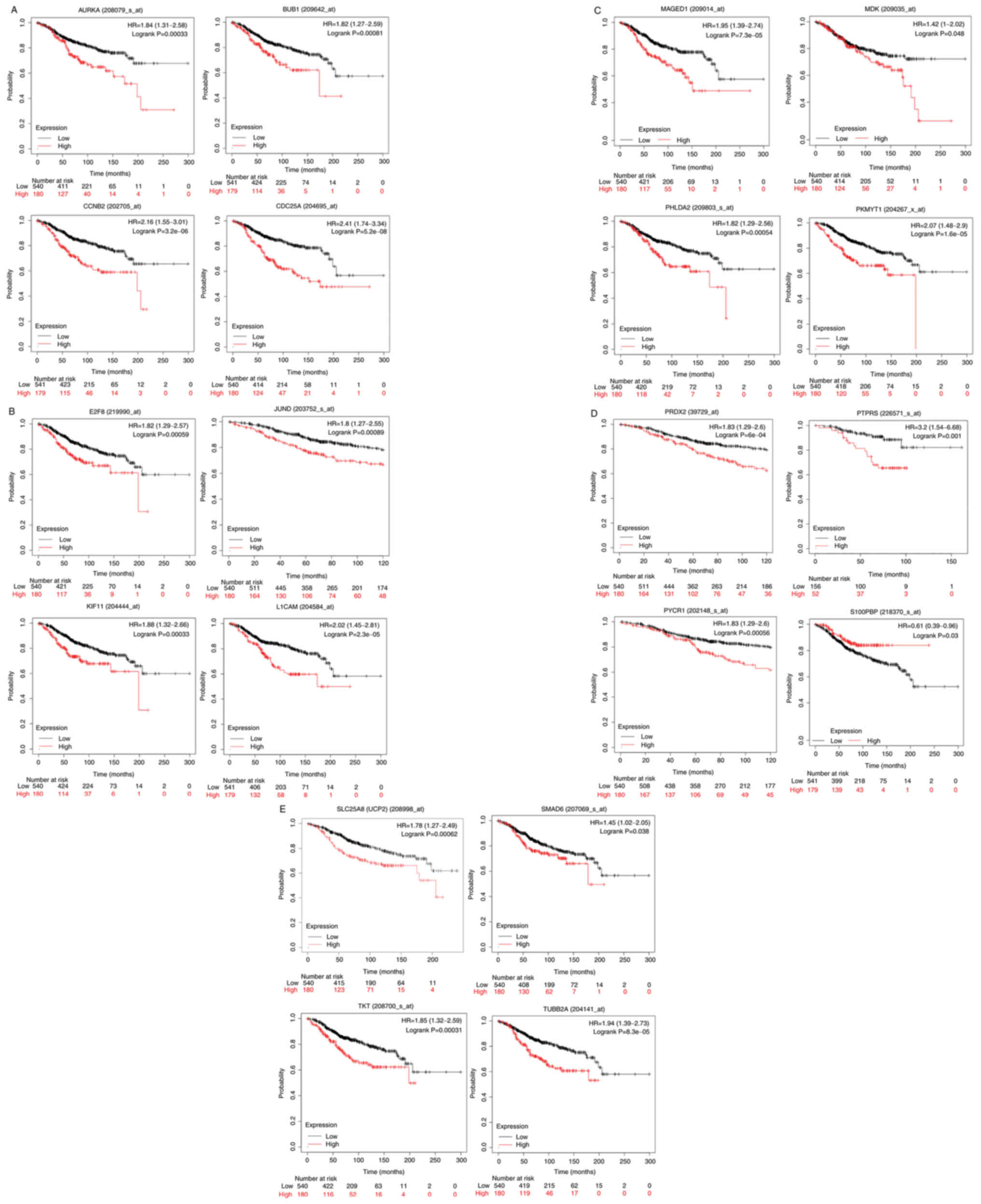
shown in Table I. The KM curves
plotted for these genes for ERα-positive patients are shown in
Fig. 4A-E. High expression of
aurora kinase-A (AURKA), mitotic checkpoint
serine/threonine-protein kinase BUB1 (BUB1), cyclin-B2
(CCNB2), cell division cycle 25C (CDC25A) (Fig. 4A), transcription factor E2F8
(E2F8), junD proto-oncogene (JUND), kinesin family
member 11 (KIF11), L1 cell adhesion molecule (L1CAM)
(Fig. 4B), melanoma antigen gene
D1 (MAGED1), midkine (MDK), pleckstrin homology-like
domain family A member 2 (PHLDA2), membrane-associated
tyrosine- and threonine kinase 1 (PKMYT1) (Fig. 4C), peroxiredoxin-2 (PRDX2),
protein tyrosine phosphatase receptor type S (PTPRS),
pyrroline-5-carboxylate reductase 1 (PYCR1) (Fig. 4D), uncoupling protein 2
(UCP2), SMAD family member 6 (SMAD6), transketolase
(TKT) and tubulin β 2A class IIa (TUBB2A) (Fig. 4E) genes was associated with
statistically significant (P<0.05) shorter OS probability in the
patients with ERα-positive breast cancer. On the other hand, high
expression of the S100P-binding protein (S100PBP) gene was
associated with statistically significant (P<0.05) longer OS
probability when comparing the 75th quartile with the 25th quartile
in KM plotter in patients with ERα-positive breast cancer (Fig. 4D).
To explore whether PDIA1 gene expression in
patients with ERα-positive breast cancer was correlated with the
upregulated or downregulated genes in either IFN-γ- or ETO-treated
PDIA1-silenced MCF-7 cells, the cBioPortal bioinformatics tool was
used to investigate the Pearson correlation coefficient between
PDIA1 mRNA levels and that of the genes shown in Table I. This investigation was carried
out by analyzing the data provided in the Breast Cancer (SMC 2018)
database using the mRNA expression z-scores relative to all samples
(RNA seq TPM, 168 samples) option. Positive correlations were
observed between PDIA1 gene expression and that of AURKA,
CDC25A, centromere protein F (CENPF),
cytoskeleton-associated protein 5 (CKAP5), E2F8, extra
spindle pole bodies like 1, separase (ESPL1), flap
structure-specific endonuclease 1 (FEN1), G2 and S
phase-expressed protein 1 (GTSE1), JUND, KIF11, kinesin-like
protein KIF20A (KIF20A), MAGED1, MDK, PKMYT1, PRDX2,
PTPRS, PYCR1, RAN binding protein 1 (RANBP1),
S100PBP, SMAD6, transforming acidic coiled-coil-containing
protein 3 (TACC3), TKT, thyroid hormone receptor
interactor 13 (TRIP13), tubulin b class 1 (TUBB),
ubiquitin-conjugating enzyme E2 C (UBE2C) and UCP2
genes. In other words, in the presence of PDIA1, the expression of
these genes would be upregulated, whereas in the absence of PDIA1,
the expression of these genes would be downregulated in patients
with ERα-positive breast cancer. Among these genes, the correlation
between PDIA1 (shown as P4HB) gene expression and
that of the CKAP5, FEN1, PYCR1, TACC3, TKT and TUBB genes
exhibited higher Pearson correlation coefficients (Fig. 5).
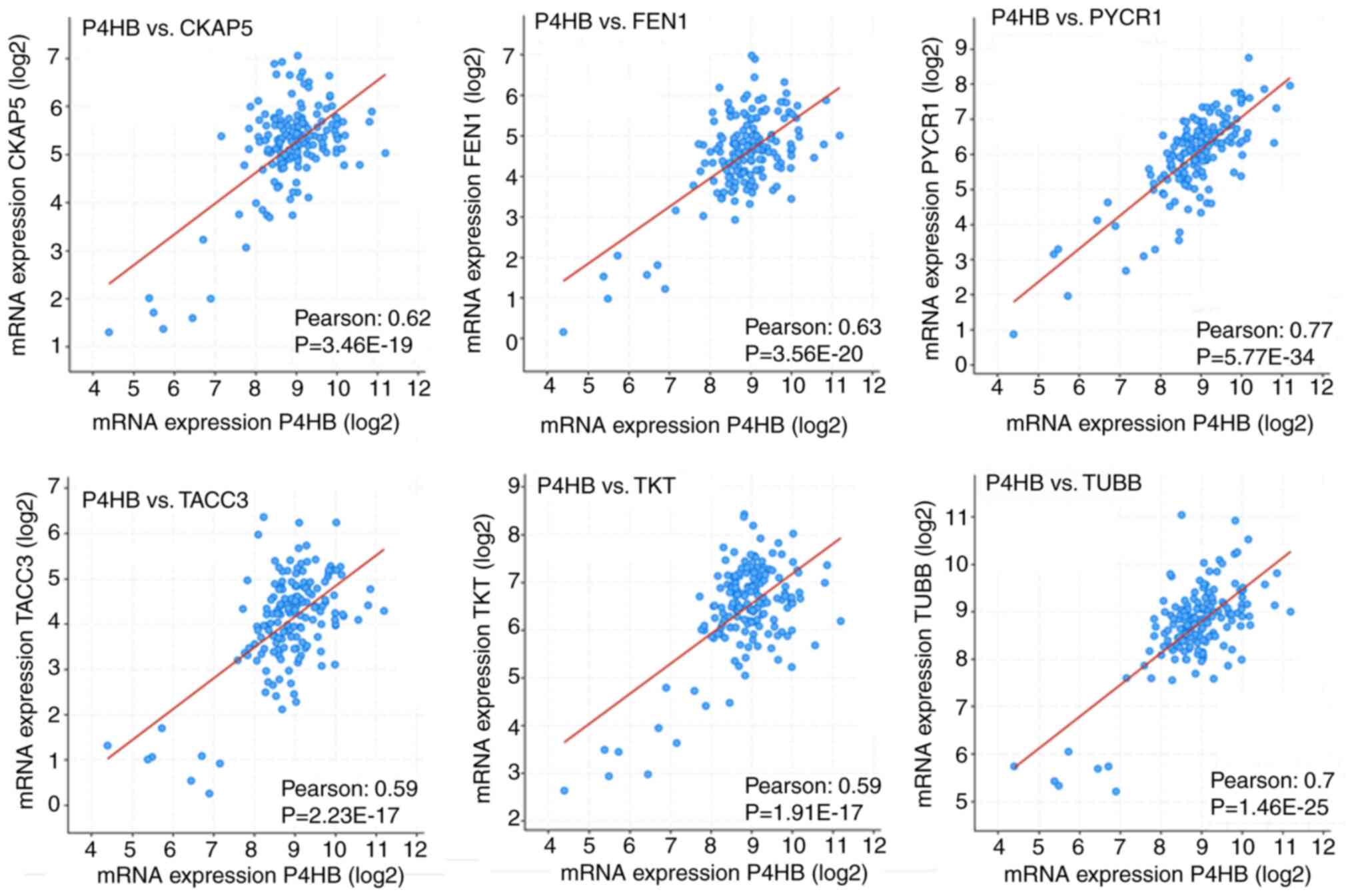 | Figure 5Pearson correlation coefficient
between PDIA1 (P4HB) mRNA levels and that of CKAP5, FEN1,
PYCR1, TACC3, TKT and TUBB genes. CKAP5, cytoskeleton
associated protein 5; FEN1, flap structure-specific endonuclease 1;
PYCR1, pyrroline-5-carboxylate reductase 1; TACC3, transforming
acidic coiled-coil containing protein 3; TKT, transketolase; TUBB,
tubulin β class I. |
Positive correlations between PDIA1 gene
expression and that of MAGED1, MDK, PRDX2, PTPRS, PYCR1, SMAD6,
TKT and UCP2 genes were found, which were demonstrated
to be upregulated in the PDIA1-silenced MCF-7 cells (Table I), thus indicating that in the
presence of PDIA1 in patients with ERα-positive breast cancer these
genes would be downregulated. According to the KM plots, the
downregulation of MAGED1, MDK, PRDX2, PTPRS, PYCR1, SMAD6,
TKT and UCP2 mRNA levels was associated with higher OS
probability in the patients with ERα-positive breast cancer
(Fig. 4). This suggested that
PDIA1 can associate with MAGED1, MDK, PRDX2, PTPRS, PYCR1,
SMAD6, TKT and UCP2 to induce anti-oncogenic effects in
patients with ERα-positive breast cancer. To explore the pathways
involved in the PDIA1-mediated anti-oncogenic effects in patients
with ERα-positive breast cancer, the Metascape bioinformatics tool
was employed (51). The results
of this analysis showed that the pathways by which PDIA1 exerts
tumor-suppressive effects include cell-substrate adhesion,
regulation of growth and response to ROS (Table III).
| Table IIIGene Ontology pathways and gene
networks through which PDIA1 confers anti-carcinogenic effects to
patients with ERα-positive breast cancer. |
Table III
Gene Ontology pathways and gene
networks through which PDIA1 confers anti-carcinogenic effects to
patients with ERα-positive breast cancer.
| Term | Description | Hits | Log P | Log (q-value) |
|---|
| 1 | GO:0031589 | Cell-substrate
adhesion | SMAD6 | −6.61 | −2.26 |
| 2 | GO:0040008 | Regulation of
growth | PTPRS,
TKT | −3.93 | −0.48 |
| 3 | GO:0000302 | Response to
reactive oxygen species | PYCR1, PRDX2,
UCP2 | −3.98 | −0.48 |
Direct correlations between PDIA1 gene
expression and that of the AURKA, CDC25A, CENPF, CKAP5, E2F8,
ESPL1, FEN1, GTSE1, JUND, KIF11, KIF20A, PKMYT1, RANBP1, S100PBP,
TACC3, TRIP13, TUBB and UBE2C genes were also observed,
which were found to be downregulated in the PDIA1-silenced MCF-7
cells (Table I). This suggested
that in the presence of PDIA1 in patients with ERα-positive breast
cancer these genes would be upregulated. The KM plots indicated
that high levels of these genes were associated with shorter OS in
patients with ERα-breast cancer (Fig.
4A-C and E), apart from S100PBP (Fig. 4D). Therefore, the association of
PDIA1 with these genes is a potential route by which PDIA1 exerts
oncogenic effects in patients with ERα-positive breast cancer. The
pathways affected by the networks formed by these genes were
investigated using the Metascape bioinformatics tool and they
included control of cell cycle transition to the mitotic phase as
well as regulation of the cell cycle checkpoints (Table IV).
| Table IVGene Ontology pathways and gene
networks through which PDIA1 confers pro-oncogenic effects
to ERα-positive breast cancer patients. |
Table IV
Gene Ontology pathways and gene
networks through which PDIA1 confers pro-oncogenic effects
to ERα-positive breast cancer patients.
| Term | Description | Hits | Log P | Log (q-value) |
|---|
| 1 | GO:0010564 | Regulation of cell
cycle process | AURKA, CDC25A,
CENPF, CKAP5, E2F8, ESPL1, FEN1, GTSE1, KIF11, KIF20A, PKMYT1,
TACC3, TRIP13, TUBB, UBE2C | −42.08 | −37.73 |
| 2 | GO:0044770 | Cell cycle phase
transition | AURKA, CDC25A,
CENPF, CKAP5, E2F8, ESPL1, GTSE1, PKMYT1, TACC3, TRIP13,
TUBB | −37.03 | −33.15 |
| 3 | GO:0007346 | Regulation of
mitotic cell cycle | AURKA, CDC25A,
CENPF, CKAP5, E2F8, ESPL1, GTSE1, KIF11, PKMYT1, RANBP1, TACC3,
TRIP13, TUBB, UBE2C | −25.16 | −21.80 |
| 4 | GO:0000075 | Cell cycle
checkpoint | AURKA, CENPF,
E2F8, GTSE1, TRIP13 | −16.50 | −13.66 |
| 5 | GO:0044839 | Cell cycle G2/M
phase transition | AURKA, CDC25A,
CENPF, CKAP5, GTSE1, PKMYT1, TUBB | −18.57 | −15.61 |
Clinical significance of the genes
modulated in IFN-γ- or ETO-treated and PDIA1-silenced MDA-MB-231
cells
Similar analysis to that described for the MCF-7
cells was carried out for the upregulated or downregulated genes in
the IFN-γ- or ETO-treated MDA-MB-231 cells with PDIA1 silenced
compared with IFN-γ- or ETO-treated cells transfected with scramble
siRNA that exhibited statistically significant (P<0.05) KM
results are shown in Table II,
and the KM curves plotted for these genes in ERα-negative patients
are presented in Fig. 6. KM
survival curves were plotted for the upregulated or downregulated
genes in MDA-MB-231 cells under various oxidative stress conditions
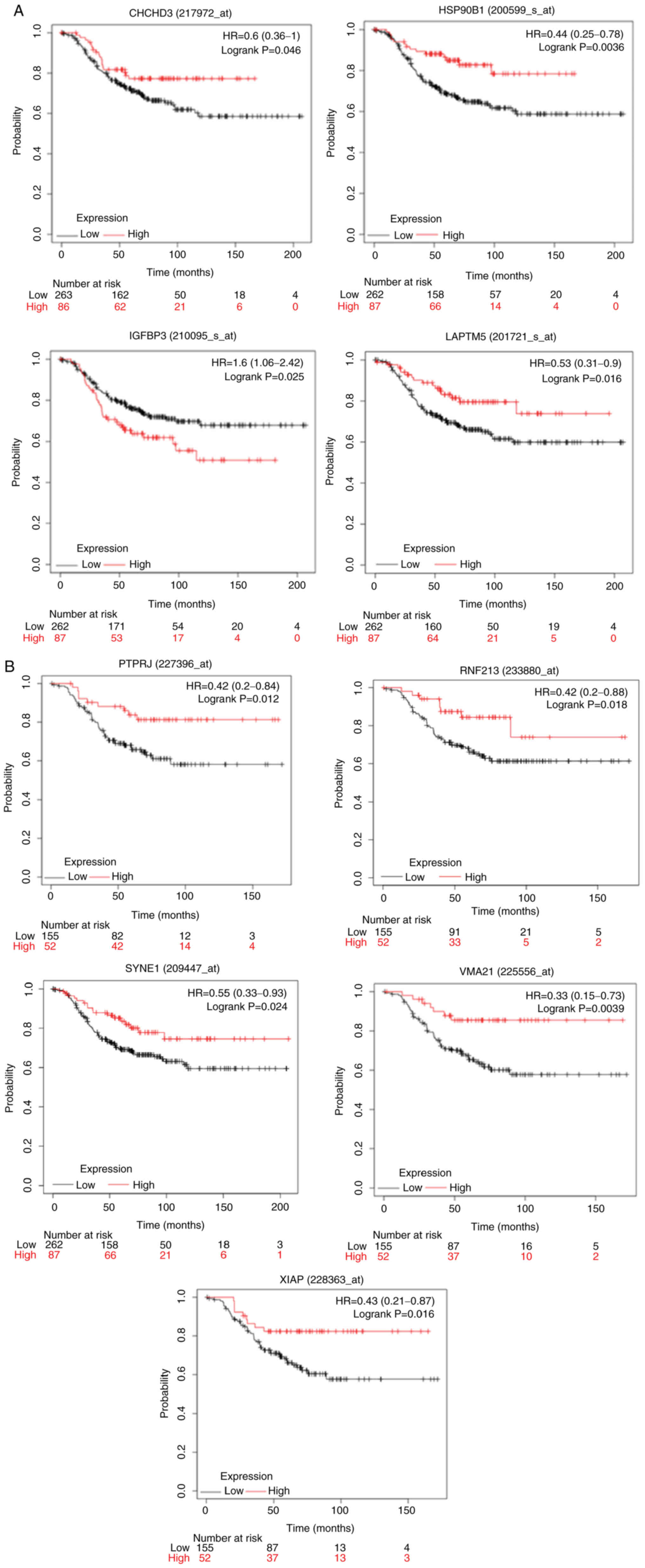
in patients with ERα-negative breast cancer (Fig. 6A and B). High expression of the
coiled-coil-helix -coiled-coil-helix domain containing 3
(CHCHD3), heat shock protein 90 β family member 1
(HSP90B1), lysosomal-associated transmembrane protein 5
(LAPTM5) (Fig. 6A),
protein tyrosine phosphatase receptor type J (PTPRJ), ring
finger protein 213 (RNF213), spectrin repeat containing
nuclear envelope protein 1 (SYNE1), vacuolar ATPase assembly
factor VMA21 (VMA21) and X-linked inhibitor of apoptosis
(XIAP) (Fig. 6B) genes was
associated with statistically significant (P<0.05) longer OS in
the patients with ERα-negative breast cancer. On the other hand,
high gene expression of insulin-like growth factor-binding protein
3 (IGFBP3) was associated with lower OS probability in these
patients (Fig. 6A).
The Pearson correlation coefficients between
PDIA1 gene expression and the upregulate or downregulated
genes in the PDIA1-silenced MDA-MB-231 cells under oxidative stress
conditions (Table II) were
investigated using the cBioPortal platform. To carry out this
analysis, only samples originating from patients with ERα-negative
breast cancer were included, but no statistical significance or
data were available to manually calculate the correlation between
PDIA1 gene expression and these genes in the patients with
ERα-negative breast cancer. Thus, it was hypothesized that the
Pearson correlation coefficients between PDIA1 gene
expression with that of the genes shown in Table II were positive, meaning that in
the presence of PDIA1, HSP90B1, IGFBP3, LAPTM5, RNF213 and
SYNE1 genes would be downregulated, while CHCHD3, PTPRJ,
VMA21 and XIAP would be upregulated, in patients with
ERα-negative breast cancer.
Direct associations between PDIA1 and
CHCHD3, PTPRJ, VMA21 and XIAP indicated that these
genes can induce tumor-suppressive effects in patients with
ERα-negative breast cancer since the KM plots in these patients
showed that the high expression of these genes was associated with
longer OS (Fig. 6A and B). PDIA1
would mediate tumor-suppressive effects in patients with
ERα-negative breast cancer patients via IGFBP3 as well since
this gene in the presence of PDIA1 is downregulated (Table II) and the KM plot in these
patients showed that low expression of IGFBP3 was associated
with longer OS (Fig. 6A). In
addition, direct association between PDIA1 and HSP90B1, LAPTM5,
RNF213 and SYNE1 gene expression suggested that these
genes confer oncogenic effects to patients with ERα-negative breast
cancer since the KM plots for these patients indicated that low
expression was associated with lower OS probability (Fig. 6A and B).
The Metascape bioinformatics tool was used to
identify the pathways affected by the genes modulated by PDIA1 to
confer anti- or pro-carcinogenic effects in patients with
ERα-negative breast cancer. This analysis revealed that PDIA1 may
exert tumor-suppressive effects in patients with ERα-negative
breast cancer by interfering with the NF-kB signaling pathway
(XIAP), glycolysis (IGFBP3), mitochondrial biogenesis
(CHCHD3) and negative regulation of cell migration
(IGFBP3, P4HB, PTPRJ) (Table
V). Metascape analysis also showed that PDIA1 may trigger
pro-oncogenic effects by forming signaling networks with
calmodulin-3 (CALM3), HSP90B1, LAPTM5 and RNF213
genes, which could affect pathways such as the adaptive immune
system and cytokine signalling in the immune system, as well as
activating signal transduction in the immune response (Table VI).
| Table VGene Ontology pathways and gene
networks through which PDIA1 confers anti-oncogenic effects
to ERα-negative breast cancer patients. |
Table V
Gene Ontology pathways and gene
networks through which PDIA1 confers anti-oncogenic effects
to ERα-negative breast cancer patients.
| Term | Description | Hits | LogP | Log (q-value) |
|---|
| 1 | hsa04064 | NF-κB signaling
pathway | XIAP | −16.64 | −3.35 |
| 2 | GO:0006006 | Glucose metabolic
process | IGFBP3 | −8.6 | −8.6 |
| 3 | R-HSA-1592230 | Mitochondrial
biogenesis | CHCHD3 | −2.71 | −2.71 |
| 4 | GO:0030336 | Negative regulation
of cell migration | IGFBP3,
PTPRJ | −1.90 | −0.96 |
| 5 | GO:0030155 | Regulation of cell
adhesion | P4HB,
PTPRJ | −8.96 | −1.03 |
| Table VIGene Ontology pathways and gene
networks through which PDIA1 confers pro-oncogenic effects
to ERα-negative breast cancer patients. |
Table VI
Gene Ontology pathways and gene
networks through which PDIA1 confers pro-oncogenic effects
to ERα-negative breast cancer patients.
| Term | Description | Hits | Log P | Log (q-value) |
|---|
| 1 | R-HSA-1280218 | Adaptive immune
system | CALM3,
RNF213 | −4.22 | −0.94 |
| 2 | R-HSA-1280215 | Cytokine signaling
in immune system | HSP90B1 | −2.91 | −0.16 |
| 3 | GO:0006874 | Cellular calcium
ion homeostasis | CALM3,
HSP90B1 | −3.52 | −0.50 |
| 4 | GO:0002757 | Immune
response-activating signal transduction | HSP90B1,
LAPTM5 | −2.86 | −0.25 |
Drugs targeting genes involved in
pathways conferring oncogenic activity in breast cancer
patients
To investigate whether approved or experimental
drugs targeting the genes involved in the pathways conferring
oncogenic activity in the patients with ERα-positive (Table VII) or ERα-negative (Table VIII) breast cancer, the genes
associated with PDIA1 and its oncogenic effects were investigated
in each subgroup of patients with breast cancer using the DRUGSURV
(52) bioinformatics tool. This
investigation revealed that in the patients with ERα-positive
breast cancer, drugs targeting the function of the AURKA, FEN1,
KIF11, PKMYT1 and TUBB genes, which are involved in the
regulation of cell cycle progression, would be selectively
effective for this group of patients. On the other hand, drugs
targeting the function of the HSP90B1 gene, which is
involved in the regulation of toll-like receptor translocation
through the endoplasmic reticulum to the plasma membrane (53), would be preferential for the
patients with ERα-negative breast cancer.
| Table VIIApproved drugs directly targeting
genes associated with PDIA1 that confer pro-oncogenic effects to
ERα-positive breast cancer patients. |
Table VII
Approved drugs directly targeting
genes associated with PDIA1 that confer pro-oncogenic effects to
ERα-positive breast cancer patients.
| Gene symbol | Drugs targeting
genes associated with PDIA1 with potential anti-carcinogenic
activity in ERα-positive breast cancer patients. |
|---|
| AURKA | Phosphonothreonine,
alisertib, cenisertib, enzastaurin, fostamatinib |
| FEN1 | Idarubicin,
quinacrine, masoprocol, mitoxantrone |
| KIF11 | Monastrol |
| PKMYT1 | Dasatinib |
| TUBB | Colchicine,
vinblastine, albendazole, podofilox, vinorelbine, vincristine |
| Table VIIIApproved drugs targeting directly or
indirectly genes associated with PDIA1 that confer
pro-oncogenic effects to ERα-negative breast cancer patients. |
Table VIII
Approved drugs targeting directly or
indirectly genes associated with PDIA1 that confer
pro-oncogenic effects to ERα-negative breast cancer patients.
| Gene symbol | Drugs targeting
genes associated with PDIA1 with potential anti-carcinogenic
activity in ERα-negative patients |
|---|
| HSP90B1 | Rifabutin |
Discussion
In our previous study, we described the effect of
protein disulfide isomerase A1 (PDIA1) silencing on reactive oxygen
species (ROS) generation, glutathione (GSH) levels, mitochondrial
membrane potential, adenosine triphosphate (ATP) production and
human leukocyte antigen G (HLA-G) surface levels in MCF-7 and
MDA-MB-231 cells treated with etoposide or interferon (IFN)-γ in
the presence or absence of PDIA1 (4). We and others reported that PDIA1
exerts differential effects in estrogen receptor (ER)α-positive vs.
triple-negative breast cancer (TNBC) cells (4,10).
These effects in the ERα-positive cells are mediated possibly
through the physical interaction between PDIA1 and the ERα and the
regulation of the transcriptional activity of the receptor
(26) whereas the impact of the
PDIA1 in the ERα-negative cells is mediated through
estrogen-independent pathways. To solidify our previous
observations indicating the differential PDIA1 effects in a type
and genetic background tumor-dependent mode (4) and taking into account the fact that
PDIA1 exerts these differential effects functioning as a
transcriptional co-modulator (27,33,54), we silenced PDIA1 gene expression
in ERα-positive MCF-7 and TNBC MDA-MB-231 cells and submitted total
mRNA isolated from these cells to RNA-seq analysis. The RNAseq
experiments were performed in PDIA1 siRNA or scramble transfected
MCF-7 and MDA-MB-231 cells treated with either IFN-γ or etoposide.
A limitation of this study is that the PDIA1 mRNA expression
levels in untreated MCF-7 and MDA-MB-231 transfected with scramble
or PDIA1 siRNA were not analyzed. The results indicated that the
number of genes that were upregulated or downregulated in the
PDIA1-silenced MCF-7 cells was higher compared to that identified
in the MDA-MB-231 cells suggesting that the main effects of
PDIA1 on gene expression were mediated by the activity of
the ERα transcription factor whereas in the TNBC cells through
other transcription factors associated with PDIA1 such as
hypoxia-inducible factor 1α (HIF-1α) (18), nuclear factor (erythroid-derived
2)-like 2 (NRF2) (32) and
nuclear factor (NF)-κB activity (27).
To overcome the limitation of the lack of patient
recruitment in the present study, the clinical significance of the
modified genes was investigated by plotting KM survival curves to
identify whether high or low expression of each one of these genes
affected the overall survival (OS) probability in ERα-positive or
ERα-negative patients. Furthermore, the pathways that the modulated
genes were involved in to confer anti-carcinogenic or pro-oncogenic
effects were identified using bioinformatic tools and grouped into
four categories. The first group included the genes that were
downregulated in the PDIA1-silenced MCF-7 breast cancer cells and
therefore upregulated in the presence of PDIA1 in the ERα-positive
breast cancer patients and their high expression was found to be
associated with short survival probability; or downregulated in the
PDIA1-silenced MCF-7 breast cancer cells and therefore upregulated
in the presence of PDIA1 in breast cancer patients and their high
expression was found to be associated with long OS probability. The
genes that were downregulated in the ERα-positive breast cancer
patients (upregulated in the PDIA1-silenced MCF-7 cells) and their
low expression was associated with reduced OS; or downregulated in
the ERα-positive breast cancer patients (upregulated in the
PDIA1-silenced MCF-7 cells) and their low expression was associated
with long OS probability formed the second category. The same
categories forming the third and fourth groups were created for the
genes exerting negative or positive effects on the OS of
ERα-negative patients. Although several genes were found to be
upregulated or downregulated in both the MCF-7 and MDA-MB-231
cells, the genes whose high expression resulted in statistically
significant probability of short or long OS in the ERα-positive or
ERα-negative patients were different.
To investigate the possibility that different gene
networks in ERα-positive vs. ERα-negative breast cancer patients
are associated with tumor-suppressive or oncogenic effects, we
employed the Metascape bioinformatic tool (51). The results of this analysis
indicated that the pathways induced to confer tumor-suppressive
effects in the ERα-positive patients are mainly cell-substrate
adhesion (SMAD6) (55-57) regulation of growth (PTPRS,
TKT) (58,59) and the response to reactive oxygen
species (PYCR1, PRDX2, UCP2) (60). Tumor-suppressive effects in the
ERα-negative breast cancer patients were found to be mediated by
pathways related to positive regulation of cell death
(IGFBP3) (61), NF-κB
signaling pathway (XIAP) (62), mitochondrial biogenesis
(CHCHD3) (63) and
negative regulation of cell migration (PTPRJ) (64).
The gene networks with which PDIA1 was associated
to induce tumorigenesis in the ERα-positive breast cancer patients
regulate pathways controlling cell cycle progression (65). Aurora kinase-A (AURKA) gene
amplification or mutations are common aberrations in breast cancer,
especially in ERα-positive breast carcinomas (66). Cyclin A2 (CCNA2) is overexpressed
in various types of cancer including breast cancer reducing the OS
of ERα-positive breast cancer patients and induces resistance to
tamoxifen treatment (67). The
CDC25C phosphatase participates in the regulation of the cell cycle
progression from the G2 to M phase by dephosphorylating and
activating the cyclin B1/CDK1 complex. Overexpression of Cdc25A in
breast carcinoma patients is associated with poor survival
(68). The KIF11 gene is
involved in the control of cell cycle progression by promoting
centrosome separation and its high expression is associated with
poor prognosis of breast cancer patients (69). Tubulin α1a (TUBA1A) is one
of three alpha-tubulin genes which are the major components of
microtubules. Analysis of the expression profile of breast cancer
tumors has indicated that TUBA1A is upregulated in tumor tissues
compared to tumor-adjacent normal breast tissues (70).
The oncogenic activities of PDIA1 in ERα-negative
patients are exerted through its association with genes involved in
the regulation of the immune response (HSP90B1, LAPTM5
RNF213) (71-73). Aberrant regulation of gene
expression of genes involved in modulating the immune response in
breast tumors could lead to a low level of tumor infiltrating
lymphocytes (TILs) and hence immune response evasion and breast
cancer aggressiveness (74,75). It should be noted that analysis of
the correlation between PDIA1 gene expression and that of
the genes upregulated or downregulated in the PDIA1-silenced
MDA-MB-231 cells was not carried out because data for these genes
were not available in the ERα-negative patients or the Pearson
correlation coefficient for those that data were available was not
statistically significant.
To further investigate the possibility of
differential targeting of the oncogenic pathways in ERα-positive
vs. ERα-negative breast cancer patients, the existence of drugs
targeting the genes implicated in oncogenic pathways were
investigated using the DRUGSURV tool (52). The drugs targeting oncogenic
pathways in the ERα-positive patients include those that target
cell cycle progression such as cenisertib, enzastaurin, and
rostamatinib, whereas drugs targeting genes involved in immune
response pathways such as rifabutin could be used for the treatment
of ERα-negative patients.
In conclusion, PDIA1 functions as a transcriptional
cofactor regulating the transcriptional activity of the ER in
ERα-positive breast cancers and HIF-1α, NRF2 and NF-κB activity in
ERα-negative breast cancers. The consequence of this is that
diverse oncogenic pathways are induced by PDIA1 in the ERα-positive
vs. the ERα-negative breast cancers allowing the hypothesis that
these two types of breast cancer can be treated differentially by
drugs targeting cell cycle progression (ERα-positive breast
cancers) or evasion of immunosurveillance (ERα-negative breast
cancers).
Availability of data and materials
RNA-seq data are available and can be accessed at
the link: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE188914.
Authors' contributions
Conceptualization of the study concept was carried
out by MKD, CD and RA. Data curation was conducted by EYB, CD, and
RA. Formal analysis and confirmation of the data were conducted by
EYB and RA. Funding acquisition was achieved by CD and RA.
Investigations, including bioinformatics database search, data
collection and data analysis, were the responsibility of EYB and
RA. Research methodology was the responsibility of EYB, CD, and RA.
Project administration was conducted by CD and RA. Resources were
the responsibility of CD and RA. Supervision was undertaken by MF,
MKD, and CD. Visualization was conducted by EYB, RA, MKD and CD.
Writing of the original draft was carried out by EYB, MKD, CD, and
RA. Writing, review and editing were carried out by EYB, MF, MKD,
CD, and RA. All authors read and agreed to the published version of
the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Acknowledgments
We thank Dr T. Geladopoulos for reading the
manuscript and providing constructive suggestions.
Funding
No funding was received.
References
|
1
|
Riemer J, Appenzeller-Herzog C, Johansson
L, Bodenmiller B, Hartmann-Petersen R and Ellgaard L: A luminal
flavoprotein in endoplasmic reticulum-associated degradation. Proc
Natl Acad Sci USA. 106:14831–14836. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Moretti AI and Laurindo FR: Protein
disulfide isomerases: Redox connections in and out of the
endoplasmic reticulum. Arch Biochem Biophys. 617:106–119. 2017.
View Article : Google Scholar
|
|
3
|
Hudson DA, Gannon SA and Thorpe C:
Oxidative protein folding: From thiol-disulfide exchange reactions
to the redox poise of the endoplasmic reticulum. Free Radic Biol
Med. 80:171–182. 2015. View Article : Google Scholar
|
|
4
|
Alhammad R, Khunchai S, Tongmuang N,
Limjindaporn T, Yenchitsomanus PT, Mutti L, Krstic-Demonacos M and
Demonacos C: Protein disulfide isomerase A1 regulates breast cancer
cell immunorecognition in a manner dependent on redox state. Oncol
Rep. 44:2406–2418. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Grek C and Townsend DM: Protein disulfide
isomerase superfamily in disease and the regulation of apoptosis.
Endoplasmic Reticulum Stress Dis. 1:4–17. 2014.PubMed/NCBI
|
|
6
|
Jang I, Pottekat A, Poothong J, Yong J,
Lagunas-Acosta J, Charbono A, Chen Z, Scheuner DL, Liu M,
Itkin-Ansari P, et al: PDIA1/P4HB is required for efficient
proinsulin maturation and ß cell health in response to diet induced
obesity. ELife. 8:e445282019. View Article : Google Scholar
|
|
7
|
Kukita K, Tamura Y, Tanaka T, Kajiwara T,
Kutomi G, Saito K, Okuya K, Takaya A, Kanaseki T, Tsukahara T, et
al: Cancer-associated oxidase ERO1-α regulates the expression of
MHC class I molecule via oxidative folding. J Immunol.
194:4988–4996. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Park B, Lee S, Kim E, Cho K, Riddell SR,
Cho S and Ahn K: Redox regulation facilitates optimal peptide
selection by MHC class I during antigen processing. Cell.
127:369–382. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tanaka T, Kutomi G, Kajiwara T, Kukita K,
Kochin V, Kanaseki T, Tsukahara T, Hirohashi Y, Torigoe T, Okamoto
Y, et al: Cancer-associated oxidoreductase ERO1-α promotes immune
escape through up-regulation of PD-L1 in human breast cancer.
Oncotarget. 8:24706–24718. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee E and Lee DH: Emerging roles of
protein disulfide isomerase in cancer. BMB Rep. 50:401–410. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xu S, Sankar S and Neamati N: Protein
disulfide isomerase: A promising target for cancer therapy. Drug
Discov Today. 19:222–240. 2014. View Article : Google Scholar
|
|
12
|
Wise R, Duhachek-Muggy S, Qi Y, Zolkiewski
M and Zolkiewska A: Protein disulfide isomerases in the endoplasmic
reticulum promote anchorage-independent growth of breast cancer
cells. Breast Cancer Res Treat. 157:241–252. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Thongwatchara P, Promwikorn W, Srisomsap
C, Chokchaichamnankit D, Boonyaphiphat P and Thongsuksai P:
Differential protein expression in primary breast cancer and
matched axillary node metastasis. Oncol Rep. 26:185–191.
2011.PubMed/NCBI
|
|
14
|
Zou H, Wen C, Peng Z, Shao YY, Hu L, Li S,
Li C and Zhou HH: P4HB and PDIA3 are associated with tumor
progression and therapeutic outcome of diffuse gliomas. Oncol Rep.
39:501–510. 2018.
|
|
15
|
Ma X, Wang J, Zhuang J, Ma X, Zheng N,
Song Y and Xia W: P4HB modulates epithelial-mesenchymal transition
and the β-catenin/snail pathway influencing chemoresistance in
liver cancer cells. Oncol Lett. 20:257–265. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xia W, Zhuang J, Wang G, Ni J, Wang J and
Ye Y: P4HB promotes HCC tumorigenesis through downregulation of
GRP78 and subsequent upregulation of epithelial-to-mesenchymal
transition. Oncotarget. 8:8512–8521. 2017. View Article : Google Scholar :
|
|
17
|
Zhang Y, Li T, Zhang L, Shangguan F, Shi
G, Wu X, Cui Y, Wang X, Wang X, Liu Y, et al: Targeting the
functional interplay between endoplasmic reticulum oxidoreductin-1α
and protein disulfide isomerase suppresses the progression of
cervical cancer. EBioMedicine. 41:408–419. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang J, Wu Y, Lin YH, Guo S, Ning PF,
Zheng ZC, Wang Y and Zhao Y: Prognostic value of hypoxia-inducible
factor-1 alpha and prolyl 4-hydroxylase beta polypeptide
overexpression in gastric cancer. World J Gastroenterol.
24:2381–2391. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yousef EM, Tahir MR, St-Pierre Y and
Gaboury LA: MMP-9 expression varies according to molecular subtypes
of breast cancer. BMC Cancer. 14:6092014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bartels AK, Göttert S, Desel C, Schäfer M,
Krossa S, Scheidig AJ, Grötzinger J and Lorenzen I: KDEL receptor 1
contributes to cell surface association of protein disulfide
isomerases. Cell Physiol Biochem. 52:850–868. 2019.PubMed/NCBI
|
|
21
|
Khan HA and Mutus B: Protein disulfide
isomerase a multifunctional protein with multiple physiological
roles. Front Chem. 2:702014.
|
|
22
|
Wiersma VR, Michalak M, Abdullah TM,
Bremer E and Eggleton P: Mechanisms of translocation of ER
chaperones to the cell surface and immunomodulatory roles in cancer
and autoimmunity. Front Oncol. 5:72015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Brigelius-Flohé R and Flohé L: Basic
principles and emerging concepts in the redox control of
transcription factors. Antioxid Redox Signal. 15:2335–2381. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang K: Endoplasmic reticulum stress
response and transcriptional reprogramming. Front Genet. 5:4602015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fu X, Wang P and Zhu BT: Protein disulfide
isomerase is a multi-functional regulator of estrogenic status in
target cells. J Steroid Biochem Mol Biol. 112:127–137. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Schultz-Norton JR, McDonald WH, Yates JR
and Nardulli AM: Protein disulfide isomerase serves as a molecular
chaperone to maintain estrogen receptor α structure and function.
Mol Endocrinol. 20:1982–1995. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Higuchi T, Watanabe Y and Waga I: Protein
disulfide isomerase suppresses the transcriptional activity of
NF-κB. Biochem Biophys Res Commun. 318:46–52. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xiao Y, Li C, Gu M, Wang H, Chen W, Luo G,
Yang G, Zhang Z, Zhang Y, Xian G, et al: Protein disulfide
isomerase silence inhibits inflammatory functions of macrophages by
suppressing reactive oxygen species and NF-κB pathway.
Inflammation. 41:614–625. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kranz P, Neumann F, Wolf A, Classen F,
Pompsch M, Ocklenburg T, Baumann J, Janke K, Baumann M, Goepelt K,
et al: PDI is an essential redox-sensitive activator of PERK during
the unfolded protein response (UPR). Cell Death Dis. 8:e2986. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhou Y, Yang J, Zhang Q, Xu Q, Lu L, Wang
J and Xia W: P4HB knockdown induces human HT29 colon cancer cell
apoptosis through the generation of reactive oxygen species and
inactivation of STAT3 signaling. Mol Med Rep. 19:231–237. 2019.
|
|
31
|
Frasor J, Weaver A, Pradhan M, Dai Y,
Miller LD, Lin CY and Stanculescu A: Positive cross-talk between
estrogen receptor and NF-kappaB in breast cancer. Cancer Res.
69:8918–8925. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee LC, Weng YT, Wu YR, Soong BW, Tseng
YC, Chen CM and Lee-Chen GJ: Downregulation of proteins involved in
the endoplasmic reticulum stress response and Nrf2-ARE signaling in
lymphoblastoid cells of spinocerebellar ataxia type 17. J Neural
Transm (Vienna). 121:601–610. 2014. View Article : Google Scholar
|
|
33
|
Campos JLO, Doratioto TR, Videira NB,
Filho HVR, Batista FA, Fattori J, de C Indolfo N, Nakahira M,
Bajgelman MC, Cvoro A, et al: Protein disulfide isomerase modulates
the activation of thyroid hormone receptors. Front Endocrinol
(Lausanne). 9:7842019. View Article : Google Scholar
|
|
34
|
Hashimoto S and Imaoka S:
Protein-disulfide isomerase regulates the thyroid hormone
receptor-mediated gene expression via redox factor-1 through thiol
reduction-oxidation. J Biol Chem. 288:1706–1716. 2013. View Article : Google Scholar :
|
|
35
|
Kimura T, Horibe T, Sakamoto C, Shitara Y,
Fujiwara F, Komiya T, Yamamoto A, Hayano T, Takahashi N and Kikuch
M: Evidence for mitochondrial localization of P5, a member of the
protein disulphide isomerase family. J Biochem. 144:187–196. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Fan Y and Simmen T: Mechanistic
connections between endoplasmic reticulum (ER) redox control and
mitochondrial metabolism. Cells. 8:10712019. View Article : Google Scholar :
|
|
37
|
Kaufman RJ and Malhotra JD: Calcium
trafficking integrates endoplasmic reticulum function with
mitochondrial bioenergetics. Biochim Biophys Acta. 1843:2233–2239.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Luo B and Lee AS: The critical roles of
endoplasmic reticulum chaperones and unfolded protein response in
tumorigenesis and anticancer therapies. Oncogene. 32:805–818. 2013.
View Article : Google Scholar
|
|
39
|
Spencer NG, Schilling T, Miralles F and
Eder C: Mechanisms underlying interferon-γ-induced priming of
microglial reactive oxygen species production. PLoS One.
11:e0162497. 2016. View Article : Google Scholar
|
|
40
|
Shin HJ, Kwon HK, Lee JH, Anwar MA and
Choi S: Etoposide induced cytotoxicity mediated by ROS and ERK in
human kidney proximal tubule cells. Sci Rep. 6:340642016.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kuhn RM, Haussler D and Kent WJ: The UCSC
genome browser and associated tools. Brief Bioinform. 14:144–161.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Dobin A, Davis CA, Schlesinger F, Drenkow
J, Zaleski C, Jha S, Batut P, Chaisson M and Gingeras TR: STAR:
Ultrafast universal RNA-seq aligner. Bioinformatics. 29:15–21.
2013. View Article : Google Scholar
|
|
43
|
Karolchik D, Baertsch R, Diekhans M, Furey
TS, Hinrichs A, Lu YT, Roskin KM, Schwartz M, Sugnet CW, Thomas DJ,
et al: The UCSC genome browser database. Nucleic Acids Res.
31:51–54. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Okonechnikov K, Conesa A and
García-Alcalde F: Qualimap 2: Advanced multi-sample quality control
for high-throughput sequencing data. Bioinformatics. 32:292–294.
2016.
|
|
45
|
Li H, Handsaker B, Wysoker A, Fennell T,
Ruan J, Homer N, Marth G, Abecasis G and Durbin R; 1000 Genome
Project Data Processing Subgroup: The sequence alignment/map format
and SAMtools. Bioinformatics. 25:2078–2079. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Robinson JT, Thorvaldsdóttir H, Winckler
W, Guttman M, Lander ES, Getz G and Mesirov JP: Integrative
genomics viewer. Nat Biotechnol. 29:24–26. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Liao Y, Smyth GK and Shi W: The R package
Rsubread is easier, faster, cheaper and better for alignment and
quantification of RNA sequencing reads. Nucleic Acids Res.
47:e472019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Stel VS, Dekker FW, Tripepi G, Zoccali C
and Jager KJ: Survival analysis I: The kaplan-meier method. Nephron
Clin Pract. 119. pp. c83–c88. 2011, View Article : Google Scholar
|
|
49
|
Li Q, Birkbak NJ, Gyorffy B, Szallasi Z
and Eklund AC: Jetset: Selecting the optimal microarray probe set
to represent a gene. BMC Bioinformatics. 12:4742011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Stelzer G, Rosen N, Plaschkes I, Zimmerman
S, Twik M, Fishilevich S, Stein TI, Nudel R, Lieder I, Mazor Y, et
al: The GeneCards suite: From gene data mining to disease genome
sequence analyses. Curr Protoc Bioinformatics. 54:1.30.31–31.30.33.
2016. View
Article : Google Scholar
|
|
51
|
Zhou Y, Zhou B, Pache L, Chang M,
Khodabakhshi AH, Tanaseichuk O, Benner C and Chanda SK: Metascape
provides a biologist-oriented resource for the analysis of
systems-level datasets. Nat Commun. 10:15232019. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Amelio I, Gostev M, Knight RA, Willis AE,
Melino G and Antonov AV: DRUGSURV: A resource for repositioning of
approved and experimental drugs in oncology based on patient
survival information. Cell Death Dis. 5:e10512014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Kawasaki T and Kawai T: Toll-like receptor
signaling pathways. Front Immunol. 5:4612014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ebersole JL, Novak MJ, Orraca L,
Martinez-Gonzalez J, Kirakodu S, Chen KC, Stromberg A and Gonzalez
OA: Hypoxia-inducible transcription factors, HIF1A and HIF2A,
increase in aging mucosal tissues. Immunology. 154:452–464. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Kiefel H, Bondong S, Hazin J, Ridinger J,
Schirmer U, Riedle S and Altevogt P: L1CAM: A major driver for
tumor cell invasion and motility. Cell Adh Migr. 6:374–384. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Blom M, Reis K, Heldin J, Kreuger J and
Aspenström P: The atypical Rho GTPase RhoD is a regulator of actin
cytoskeleton dynamics and directed cell migration. Exp Cell Res.
352:255–264. 2017. View Article : Google Scholar
|
|
57
|
Filippou PS, Karagiannis GS and
Constantinidou A: Midkine (MDK) growth factor: A key player in
cancer progression and a promising therapeutic target. Oncogene.
39:2040–2054. 2020. View Article : Google Scholar
|
|
58
|
Wang ZC, Gao Q, Shi JY, Guo WJ, Yang LX,
Liu XY, Liu LZ, Ma LJ, Duan M, Zhao YJ, et al: Protein tyrosine
phosphatase receptor S acts as a metastatic suppressor in
hepatocellular carcinoma by control of epithermal growth factor
receptor-induced epithelialmesenchymal transition. Hepatology.
62:1201–1214. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Tseng CW, Kuo WH, Chan SH, Chan HL, Chang
KJ and Wang LH: Transketolase regulates the metabolic switch to
control breast cancer cell metastasis via the α-ketoglutarate
signaling pathway. Cancer Res. 78:2799–2812. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yi JW, Park JY, Sung JY, Kwak SH, Yu J,
Chang JH, Kim JH, Ha SY, Paik EK, Lee WS, et al: Genomic evidence
of reactive oxygen species elevation in papillary thyroid carcinoma
with Hashimoto thyroiditis. Endocr J. 62:857–877. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Wang SH, Chen YL, Hsiao JR, Tsai FY, Jiang
SS, Lee AYL, Tsai HJ and Chen YW: Insulin-like growth factor
binding protein 3 promotes radiosensitivity of oral squamous cell
carcinoma cells via positive feedback on NF-κB/IL-6/ROS signaling.
J Exp Clin Cancer Res. 40:952021. View Article : Google Scholar
|
|
62
|
Hofer-Warbinek R, Schmid JA, Stehlik C,
Binder BR, Lipp J and de Martin R: Activation of NF-kappa B by
XIAP, the X chromosome-linked inhibitor of apoptosis, in
endothelial cells involves TAK1. J Biol Chem. 275:22064–22068.
2000. View Article : Google Scholar
|
|
63
|
Darshi M, Mendiola VL, Mackey MR, Murphy
AN, Koller A, Perkins GA, Ellisman MH and Taylor SS: ChChd3, an
inner mitochondrial membrane protein, is essential for maintaining
crista integrity and mitochondrial function. J Biol Chem.
286:2918–2932. 2011. View Article : Google Scholar :
|
|
64
|
Schneble N, Müller J, Kliche S, Bauer R,
Wetzker R, Böhmer FD, Wang ZQ and Müller JP: The protein-tyrosine
phosphatase DEP-1 promotes migration and phagocytic activity of
microglial cells in part through negative regulation of fyn
tyrosine kinase. Glia. 65:416–428. 2017. View Article : Google Scholar
|
|
65
|
Tabach Y, Milyavsky M, Shats I, Brosh R,
Zuk O, Yitzhaky A, Mantovani R, Domany E, Rotter V and Pilpel Y:
The promoters of human cell cycle genes integrate signals from two
tumor suppressive pathways during cellular transformation. Mol Syst
Biol. 1:2005.00222005. View Article : Google Scholar
|
|
66
|
Fu J, Bian M, Jiang Q and Zhang C: Roles
of Aurora kinases in mitosis and tumorigenesis. Mol Cancer Res.
5:1–10. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Gao T, Han Y, Yu L, Ao S, Li Z and Ji J:
CCNA2 is a prognostic biomarker for ER+ breast cancer and tamoxifen
resistance. PLoS One. 9:e917712014. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Cangi MG, Cukor B, Soung P, Signoretti S,
Moreira G Jr, Ranashinge M, Cady B, Pagano M and Loda M: Role of
the Cdc25A phosphatase in human breast cancer. J Clin Invest.
106:753–761. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Rapley J, Nicolàs M, Groen A, Regué L,
Bertran MT, Caelles C, Avruch J and Roig J: The NIMA-family kinase
Nek6 phosphorylates the kinesin Eg5 at a novel site necessary for
mitotic spindle formation. J Cell Sci. 121:3912–3921. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Nami B and Wang Z: Genetics and expression
profile of the tubulin gene superfamily in breast cancer subtypes
and its relation to taxane resistance. Cancers (Basel). 10:2742018.
View Article : Google Scholar
|
|
71
|
Lin T, Qiu Y, Peng W and Peng L: Heat
shock protein 90 family isoforms as prognostic biomarkers and their
correlations with immune infiltration in breast cancer. Biomed Res
Int. 2020:21482532020. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Glowacka WK, Alberts P, Ouchida R, Wang JY
and Rotin D: LAPTM5 protein is a positive regulator of
proinflammatory signaling pathways in macrophages. J Biol Chem.
287:27691–27702. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Sharma P, Barlow WE, Godwin AK, Parkes EE,
Knight LA, Walker SM, Kennedy RD, Harkin DP, Logan GE, Steele CJ,
et al: Validation of the DNA damage immune response signature in
patients with triple-negative breast cancer from the SWOG 9313c
trial. J Clin Oncol. 37:3484–3492. 2019. View Article : Google Scholar
|
|
74
|
Leung KL, Verma D, Azam YJ and Bakker E:
The use of multi-omics data and approaches in breast cancer
immunotherapy: A review. Future Oncol. 16:2101–2119. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Sabbatino F, Liguori L, Polcaro G, Salvato
I, Caramori G, Salzano FA, Casolaro V, Stellato C, Col JD and Pepe
S: Role of human leukocyte antigen system as a predictive biomarker
for checkpoint-based immunotherapy in cancer patients. Int J Mol
Sci. 21:72952020. View Article : Google Scholar :
|