Liver cancer is ranked as the sixth most prevalent
malignancy worldwide, and was the third highest cause of
cancer-associated mortality worldwide in 2020 with ~905,677 new
cases and 830,180 cancer-associated mortalities annually (1). Hepatocellular carcinoma (HCC) is the
predominant subtype of hepatic carcinoma, and accounts for 75-85%
of all primary liver cancer cases (2). Infection with hepatitis B or C
viruses (HBV or HCV, respectively) causes chronic liver injury and
has recently been reported to play a pivotal place in the
carcinogenesis and development of HCC (3). Due to the vaccination against HBV,
the prevalence of HBV and the incidence rate of HCC have markedly
decreased in numerous high-risk regions, such as China (4). However, the current situation is far
from satisfactory in numerous low- and middle-income countries, due
to the shortage of HBV vaccines, and the lack of improved
sanitation and regular screening (5). Thus, the 5-year overall survival
rate of patients with HCC remains low (<25%), and large-scale
efforts are urgently required to elucidate the mechanisms
underlying the development of neoplasia and to improve the
preliminary diagnostic rate of HCC (6,7).
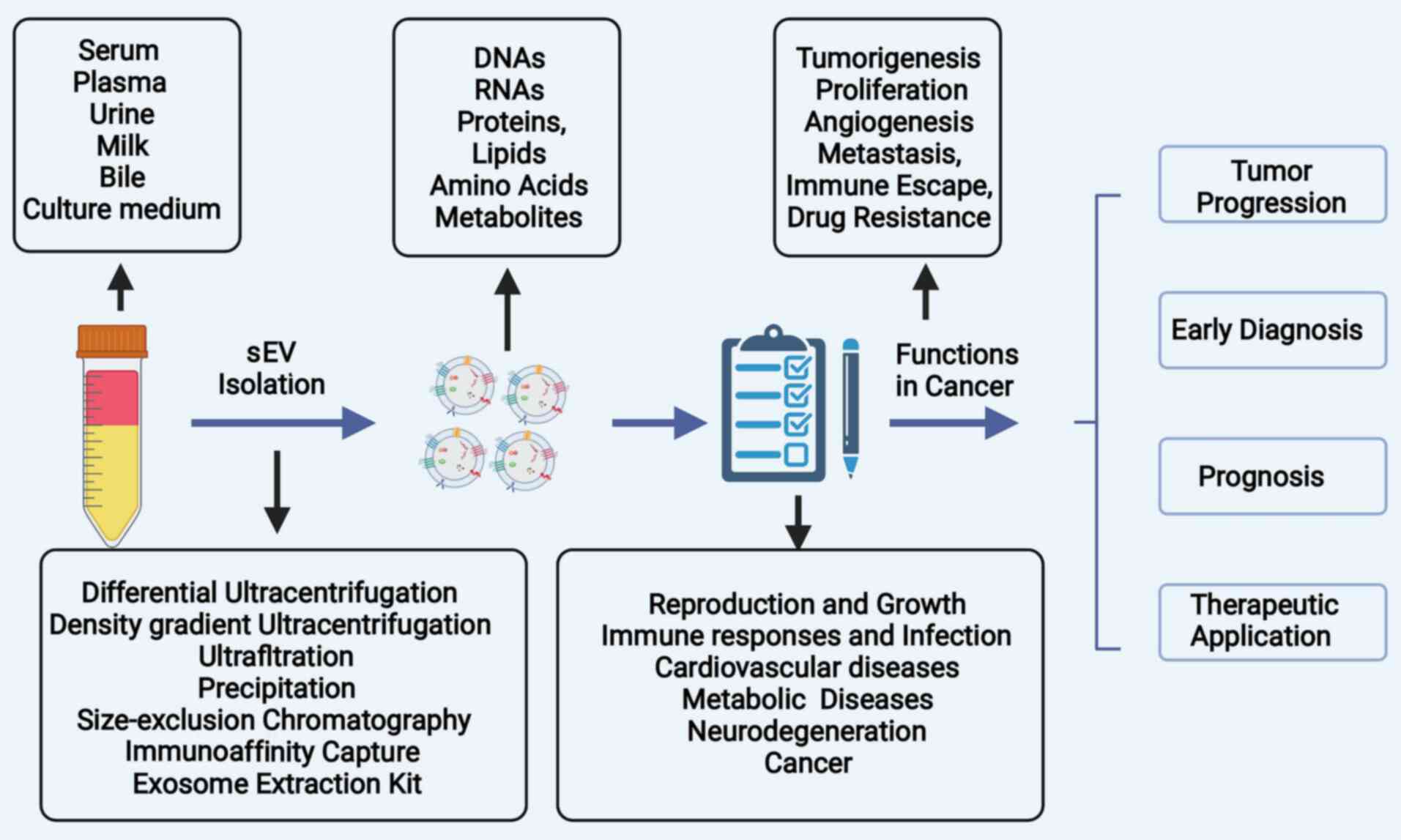
Small extracellular vesicles (sEVs), which were
previously known as exosomes, have a diameter of 30-200 nm and are
a subset of EVs that were first described by Johnstone et al
(8) in the 1980s. Following
several decades of research, it was observed that sEVs not only
function in cellular waste disposal, but also serve as an excellent
vehicle for cell-cell communications. sEVs contain complex and
diverse materials, including DNAs, RNAs, proteins, lipids and
metabolites, and they shuttle these bioactive molecules between
cells (9). The cargos of sEVs can
be internalized by recipient cells, thus mediating the metabolic
activities of recipient cells and consequently participating in
both normal physiology and acquired abnormalities, such as immune
responses, mammalian reproduction and development, central nervous
system-related diseases and cancers (10). In HCC, accumulated evidence has
indicated that sEVs play an essential role in carcinogenesis and in
the remodeling of the tumor microenvironment (TME), as well as in
proliferation, metastasis, angiogenesis and drug resistance
(11). The profiles of sEV cargos
are origin-specific, and the distinct expression of sEV cargos
between patients with HCC and healthy subjects renders sEVs a
potential diagnostic biomarker for HCC (12). Furthermore, certain sEV RNAs may
serve as molecular markers for the early detection, TNM staging,
prognostic evaluation and recurrence monitoring in HCC, which may
contribute more effective to diagnosis and treatment options
(13). Considering the intrinsic
property of sEVs of transferring information and altering the
biological response of recipient cells, recent studies have
highlighted their potential utility values in the therapeutic
fields of several diseases, including cardiovascular diseases and
cancers (14,15).
The present review article summarizes the biogenesis
of sEVs, as well as the role of sEVs in the tumorigenesis and
progression of HCC. In addition, the potential and emerging
clinical applications of sEVs in the diagnosis and treatment of HCC
are discussed (Fig. 1).
'EV' is a heterogeneous collective term for
phospholipid bilayer membrane-encapsulated nano or microvesicles.
Traditionally, EVs were broadly categorized into cytoplasmic
membrane-derived ectosomes and exosomes of endosome-origin
(16). However, without optimal
isolation methods and real-time imaging technologies to visualize
the process of release or specific markers of different subtypes of
EVs, the differentiation between exosomes and small ectosomes is
unlikely due to their analogous intrinsic properties and the
overlapping size. Thus, the latest guideline of Minimal Information
for Studies of Extracellular Vesicles 2018 (MISEV2018) proposed the
use of standard terminologies for EV subtypes followed by physical
characteristics, biochemical composition and the condition of
progenitor cells (17). In the
present review article, the term 'EV' encompasses a heterogeneous
population of both exosomes and nano-scaled ectosomes with a
diameter <200 nm.
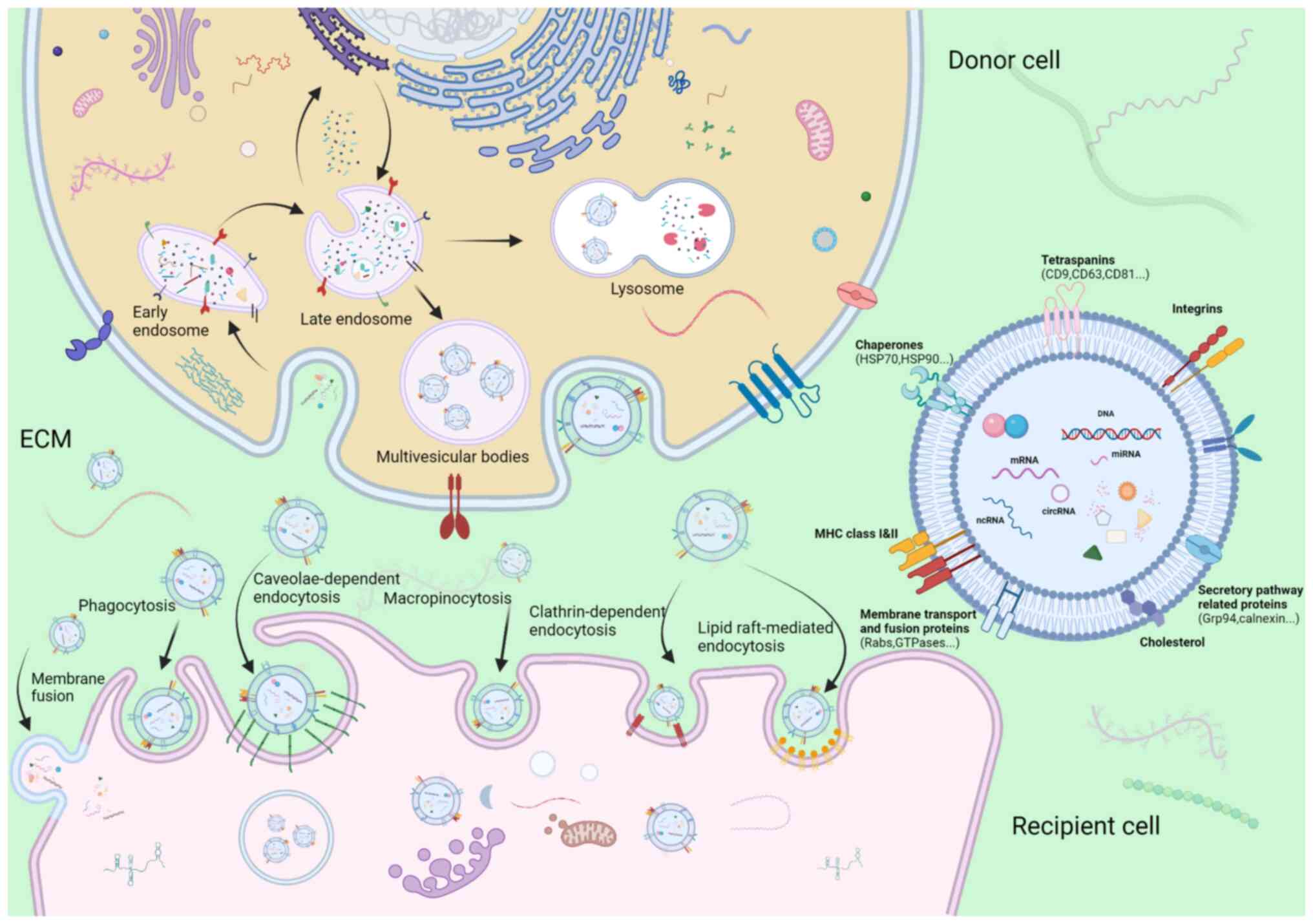
Ectosomes, which are microvesicles and
microparticles with a diameter ranging from 50 to 1,000 nm, are
vesicles produced directly by the outward budding of the plasma
membrane (18). By contrast, the
process of the synthesis and release of exosomes is a more
complicated and intricate sequence of multiple fusion events,
budding of the plasma membrane and releasing of specific payloads
(Fig. 2). The first inward
invagination of a lipid bilayers contributes to the generation of
early-sorting endosomes (ESEs) (19). ESEs can mature towards
late-sorting endosomes (LSEs), which is followed by the formation
of multivesicular bodies (MVBs) through the second intraluminal
budding of the endosomal membrane, during which, specific bioactive
compounds such as nucleic acids, proteins, and lipids are gradually
enriched in intraluminal vesicles (ILVs) (20). Although the mechanisms underlying
the formation of ILVs and specific bioactive compound sorting
system have not yet been well elucidated, the majority of
oncologists hypothesize that endosomal sorting complex required for
transport (ESCRT) facilitates exosome budding. Of note, an
ESCRT-independent mechanism may play a role in the biogenesis of
exosomes, since no notable decrease in the release of exosomes was
observed following the inhibition of ESCRT family activity
(21). These two pathways may not
be completely separated, although they function synergistically in
the synthesis of exosomes (22).
MVBs mainly have two endings: i) These mature MVBs may be
incorporated into autophagosomes or lysosomes for hydrolysis of
vesicular contents; or ii) they can be incorporated into the
cellular plasma membrane and be subsequently expelled into the
extracellular space as exosomes (23). When arriving to their recipient
cells, sEVs are recognized and assimilated into cells via the
following mechanisms: donor-acceptor interaction, membrane fusion,
phagocytosis, and clathrin-independent and clathrin-dependent
endocytosis, depending on their physical and biological properties
(24,25). For example, angiopoietin-2
(ANGPT2)-bearing sEVs derived from HCC cells are transferred into
human umbilical vein endothelial cells (HUVECs) via endocytosis
(26). The processes of
formation, secretion and uptake of exosomes are depicted in
Fig. 2, as reported in a previous
study by the authors (27).
sEVs can be excreted by almost all cells types and
are abundant in the human body, existing in biological fluids, such
as plasma, urine, tears, plasma and breast milk (28). sEVs can be isolated from cell
culture conditioned media, multiple biofluids, or tissue using
several methods. The separation of sEVs principally involves five
approaches, including differential ultracentrifugation, sucrose and
iodixanol density ultracentrifugation, polyethylene glycol
precipitation, size exclusion chromatography (SEC) and
immunoaffinity capture (29).
However, it is extremely difficult to identify a single separation
strategy with both a high recovery rate and high specificity. The
present study aimed to systematically review the recent,
cutting-edge research on sEVs and HCC, focusing on high-quality
studies using differential ultracentrifugation or density gradient
centrifugation as the separation methods of sEVs, with an
intermediate recovery rate and purity according to MISEV2018
(17). Notably, all these
aforementioned approaches have their own advantages and
disadvantages; thus, a combined method, such as differential
ultracentrifugation followed by SEC is scalable for future
sEVs-based studies (30). Other
articles (31-68) discussing methods of isolation of
sEVs involving only ultracentrifugation or commercial kits are
cited Table SI. Further
investigations on a more effective and reproducible approach for
separating sEVs are urgently required.
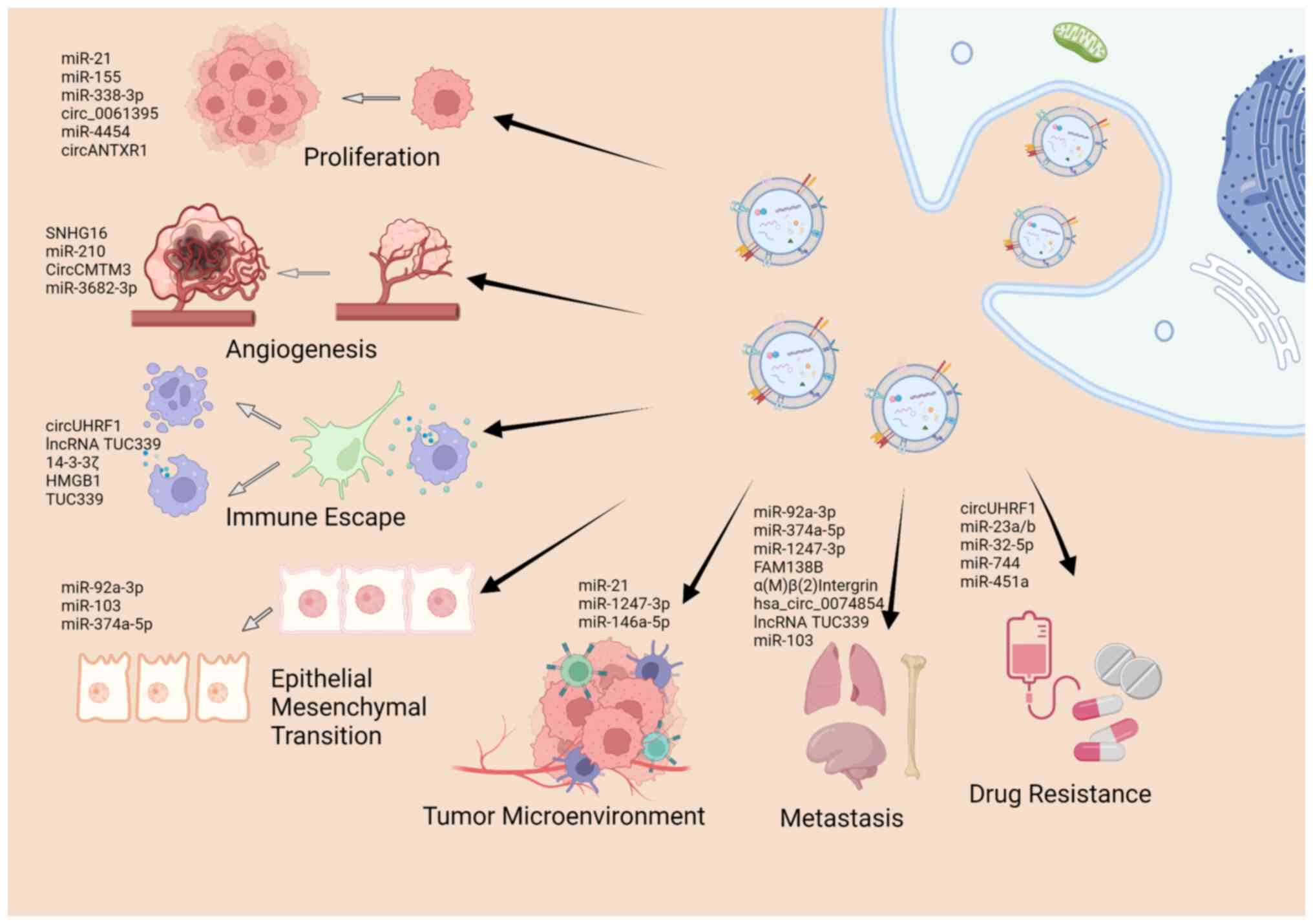
As aforementioned, sEVs encapsulate a series of
cargos, including nucleic acids, proteins and lipids, and
sEV-related research has mainly focused on the ability of sEVs to
exchange of these cargos between cells (69,70). Previous studies on the roles of
sEV cargos in cancer have demonstrated that sEVs are involved in
almost all hallmarks of cancers, including tumor initiation and
formation (71-75), in the remodeling of the TME
(76,77), apoptosis (50), angiogenesis (78,79), metastasis (80-83), immune escape (52), and drug resistance (84). The present review article
summarizes the literature that highlights the significance of sEV
cargos in the carcinogenesis and development of HCC (Fig. 3), as presented in Table I.
Tumorigenesis is not a single-step event, but a
consequence of long-term alteration of mutations of genes and
functional changes in the TME (85). Emerging evidence suggests that
sEVs participate in the initiation, formation and remodeling of the
TME in HCC (11). Chronic
hepatitis B (CHB) remains a main factor responsible for HCC
development, and sEVs are implicated in the spread, immune
regulation and antiviral response of HBV infections (86). For example, exosomes from
macrophages can deliver IFN-α-related microRNAs (miRNAs or miRs) to
HBV-infected hepatocytes, and activate the antiviral response to
suppress HBV replication and expression (87). The exosomal long non-coding RNA
(lncRNA) HOTTIP has been shown to play a role in mediating the
antiviral effect of tenofovir alafenamide following HBV infection
(88). It has been demonstrated
that sEVs from CD4+ T-cells can enhance B cell responses
and potentiate the efficacy of the hepatitis B surface antigen
vaccine (89). These studies
indicate that sEVs can mediate immune regulation and antiviral
response in HBV infection. A previous study also indicated that
sEVs may exert negative immune regulatory effects, and that they
are indispensable in the transformation from liver cirrhosis (LC)
to liver cancer (90). The
interplay between cancer cells and the TME is an essential activity
that supports or prevents tumor development and progression. In
HCC, tumor cells co-exist with other non-cancerous cells that
constitute the TME and enhance tumor growth via various mechanisms.
sEVs can exert an effect on the information and remodeling of the
TME. For instance, exosomal miR-21 derived from HCC has been found
to promote the conversion of hepatic stellate cells into
cancer-associated fibroblasts (CAFs), and to facilitate the
formation of the TME (91).
Exosomal-miR-1247-3p from HCC cells has been shown to reduce the
expression of B4GALT3 in CAFs and stabilize β1-integrin, leading to
the activation of fibroblasts via the NF-κB signaling pathway
(92). To summarize, sEVs play an
essential role in the pathogenesis of HBV-related hepatic diseases,
transformation from precancerous diseases to HCC and in the
formation of the TME to confer tumorigenesis in HCC.
The development of HCC can be partly attributed to
the rapid proliferation and uncontrolled expansion of tumor cells,
which also accounts for tumor progression and resistance to
therapy. sEVs mediate tumor growth and expansion by affecting the
cell cycle, proliferation rate and apoptosis of HCC cells (93-95). Cao et al (71) suggested that exosomal miR-21 can
influence HCC by altering the expression of the tumor suppressor
genes, PTEN and PTEN pseudogene 1. Sun et al (96) indicated that exosome-specific
miR-155 targeted PTEN and consequently stimulated the proliferation
of HCC cells. On the contrary, certain sEV-encapsulated cargos,
such as miR-338-3p can inhibit cell proliferation, induce cell
apoptosis and consequently repress the progression of HCC (97). In addition, another study
demonstrated that the proliferative and migratory abilities of HCC
cell lines were potentiated, while their apoptosis was counteracted
via the enforced expression of the exosomal lncRNA H19 (98). Furthermore, sEV constituents may
intervene in the cell cycle to regulate the progression of HCC. It
has been corroborated that circ_0061395 silencing can trigger cell
cycle arrest and apoptosis, and suppress the proliferation of HCC
in vitro, as well as inhibit tumor growth (56). Similarly, miR-4454
inhibitor-mediated exosomes can substantially exacerbate cycle
arrest, apoptosis and the formation of reactive oxygen species in
HCC (95). Of note, the
progression of HCC is a result of the accumulation of several
time-intersecting steps, including invasion, migration,
angiogenesis, immune escape and metastasis, and sEV cargos may also
function via several mechanisms. Huang et al (62) suggested that the silencing of
circANTXR1 can suppress HCC progression, not only by inhibiting the
proliferative ability of HCC, but also by hampering the migration,
invasion and metastasis of tumor cells. The roles of sEVs in other
hallmarks of tumor progression will be further discussed in the
following section.
Angiogenesis refers to the formation of new blood
vessels from pre-existing ones. It is a complex, multistep process
involving extracellular matrix remodeling, endothelial cell
migration and ultimately generation of microvessels. Angiogenesis
not only provides sufficient oxygen and nutrition for cancer cells,
but is also essential for HCC proliferation, local invasion and
distant metastasis. The significance of sEVs in cancer angiogenesis
has been widely explored and documented recently (98). In HCC, exosomal SNHG16 can sponge
miR-4500 and activate angiogenesis in HUVECs by regulating
polypeptide N-acetylgalactosaminyltransferase 1 via the
PI3K/Akt/mTOR pathway (99). Lin
et al (100) reported
that tumor-derived exosomes (TDEs) containing miR-210 could target
SMAD4 and STAT6 in endothelial cells, and thereby promote the
angiogenesis of HCC. The functions of these sEV cargos are
multifaceted, and they alter the gene expression of the recipient
cells, which become more aggressive and exhibit malignant
characteristics. Apart from regulating angiogenesis, they may also
control phenotypic changes, such as the proliferative or migratory
abilities of cancer cells. A previous study demonstrated that
circCMTM3-bearing sEVs can drive the angiogenesis of HUVECs, as
well as their viability, migration and invasion (58). Certain studies have found that
serval sEV cargos may play the opposite role and suppress
angiogenesis in HCC (34,101). For instance, HCC-derived
exosomes containing miR-3682-3p have been shown to attenuate
angiogenesis via targeting ANGPT1, which is dependent on the
RAS-MEK1/2-ERK1/2 pathway (101). Taken together, these results
indicate that angiogenesis is a complex process that is
orchestrated by multiple biological factors, and the treatment of
angiogenesis may provide a novel prospective therapeutic approach
for HCC.
Widespread metastasis in patients with HCC remains a
major challenge for treatment, and a main reason for treatment
failure, as well as one of the leading-causes of cancer-associated
mortality (102). Metastasis is
a multistep process involving EMT, invasion into vessels,
intravascular transport and organ-specific seeding. The most common
mode of metastasis in HCC is intrahepatic metastasis, followed by
lymphatic metastasis and distant metastasis to the lungs. sEVs are
involved in multiple steps of HCC metastasis, and the importance of
sEVs in HCC metastasis has recently been widely reported. Firstly,
sEVs contribute to the EMT of HCC cells. Yang et al
(103) demonstrated that
exosomal miR-92a-3p from high-metastatic HCC cell lines can
potentiate EMT and metastasis by inactivating PTEN and activating
Akt/Snail signaling. Similarly, Chen et al (104) suggested that TDEs from HCC cells
can accelerate EMT, and induce HCC progression and recurrence by
activating the MAPK/ERK signaling pathway; however, those studies
did not clarify the specific sEVs-carrying cargo that is involved
in this process. sEVs exacerbate the migratory and invasive
abilities of HCC, which may promote the metastasis of HCC. It has
been observed that miR-374a-5p in exosomes potentiates the
migration and invasion of HCC by regulating growth arrest and DNA
damage inducible alpha (105).
In addition, sEVs can orchestrate the organotropic metastasis of
HCC by converting the pre-metastatic microenvironment into a tumor
cell-friendly site. A previous study demonstrated that
Exo-miR-1247-3p derived from HCC can trigger β1-integrin-NF-κB
signaling in fibroblasts in the lungs, and is positively associated
with several pro-inflammatory cytokines, such as IL-8 and IL-6,
which promote the lung metastasis of HCC (92). Recent studies have identified
numerous sEV cargos that are involved in the metastasis of HCC,
including FAM138B (106), α(M)
β(2) integrin (107), hsa_circ_0074854 (108) and lncRNA TUC339 (109). It should be noted that the
aforementioned sEV cargos may not only participate in one step of
metastasis, but may play multifaceted roles in the whole process of
metastasis. For example, Fang et al (42) indicated that, apart from promoting
EMT and enhancing tumor motility in vitro, HCC cells can
secrete sEV-encapsulated miR-103, which also potentiates vascular
permeability and lung metastasis in mouse models.
The immune system plays a paramount role in
recognizing and eliminating malignant cells and foreign invaders.
In the processes of tumor initiation and progression, aberrant
proliferation and gene alteration in cancer cells can generate
abnormally expressed antigens, which should be adequately
presented, recognized and eliminated by the immune system (110). During their fight against the
immune system, cancer cells also evolve, and may acquire the
ability to evade immunosurveillance via various mechanisms. Among
the immune escape effects, that contribute to cancer progression
and drug resistance, engagement of the attenuation or abrogation of
immunocytes is worth mentioning, and sEVs play a pivotal role in
this process (111). Recent
studies have suggested that HCC-derived sEVs can impair the
function of natural killer (NK) and T-cells, as well as activate
immuno-suppressive cells such as M2 macrophages. For instance,
exosomal circUHRF1 from HCC triggers the exhaustion of NK cells and
subsequently induces resistance to therapy (52). Exo-lncRNA TUC339 has also been
shown to be internalized by macrophages, and to modulate M1/M2
polarization and suppress the antitumor immune response in HCC
(109). Apart from diminishing
the activity of the innate immune system, previous studies have
indicated that TDEs from HCC can also impede the activation and
function of specific immunocytes such as T- and B-cells (32,46,52). Wang et al (112) found that exosomal 14-3-3ζ
released by HCC cells suppressed the antineoplastic characteristics
of tumor-infiltrating T-lymphocytes. Tumor-derived exosomal HMGB1
can enhance the expansion the T-cell Ig and mucin domain (TIM)-1
(+) regulatory B-cells and facilitate HCC immune evasion (113). Collectively, sEVs play multiple
roles in the communication of HCC and immune cells, and are
critical for the immune escape of HCC cells and tumor progression.
Thus, sEVs may serve as ideal therapeutic targets for HCC, although
further investigations into this matter are warranted.
HCC is one of the most aggressive cancer types, and
hepatic resection remains the gold standard of treatment for HCC if
the patients can withstand surgery. For patients who experience HCC
recurrence or cannot tolerate surgery, targeted therapies involving
the use of sorafenib, a multi-kinase inhibitor compound, and
chemotherapy including paclitaxel and 5-fluorouracil (5-FU) are
first-line treatments. Resistance to drugs remains a main obstacle
to the effective treatment of these patients. The mechanisms
underlying drug resistance remain complex and elusive; however, but
the roles of sEVs in this process is emerging and have captured the
interest of researchers. For instance, a previous study found that
transfection with GRP78 small interfering RNA into bone
marrow-derived mesenchymal stem cells could yield sEVs containing
siGRP78, thus mediating targeted RNA silencing, which increased the
sensitivity of drug-resistant cancer cells to sorafenib and
improved the drug resistance reversion (114). Furthermore, as previously
demonstrated, sEV-encapsulated miR-23a/b derived from adipocytes
was transferred to neighboring HCC cells, which enhanced their
chemoresistance to 5-FU by targeting the VHL/HIF axis (115). The upregulation of
miR-32-5p-bearing sEVs has been shown to induce multidrug
resistance by potentiating EMT and angiogenesis via targeting the
PI3K/PTEN/Akt signaling pathway (34). Another study demonstrated that
sEVs secreted from cancer stem cells induced regorafenib
insensitivity by upregulating Nanog expression (116). These findings highlight the
significance of sEVs in the drug resistance of HCC, which results
from sEVs directly suppressing drug efficacy against tumor cells,
or from sEVs regulating the gene expression of recipient cells to
facilitate cancer survival. Since sEVs can alter the drug
sensitivity of HCC cells, it is tempting to engineer a sEV-derived
vehicle to deliver specific agents for HCC treatment (117). Studies on this topic are
currently underway, and the preliminary results are promising. Wang
et al (118) indicated
that the downregulation of miR-744 in HCC tissue and cell lines was
implicated in the chemoresistance to sorafenib, and HCC cell lines
treated with miR-744-upregulating sEVs were more sensitive to
sorafenib, which provides a potential approach to reduce the
occurrence of drug resistance.
Despite the advances in the diagnosis of HCC, the
number of new cases and cancer mortalities associated with HCC
remain high (1). The diagnosis of
HCC relies heavily on imaging analyses, such as magnetic resonance
imaging or computed tomography; however, the diagnosis of the
majority of patients is confirmed at an advanced stage, and thus
these patients miss the optimal treatment period (119). Immense efforts have been made as
regards the early diagnosis of HCC, although success has been
limited. Alpha-fetoprotein (AFP) is a traditional HCC marker with a
low specificity, which has a limited value in the differential
diagnosis between HCC and other liver diseases (120,121). As regards other biomarkers, such
as golgi glycoprotein 73, AFP-L3, phosphatidylinositol proteoglycan
3 and decarboxylated prothrombin, they do not provide any obvious
advantage in the early diagnosis of HCC compared with AFP (122,123). Therefore, a non-invasive method
with a high diagnostic sensitivity and specificity is urgently
required. Recently, liquid biopsies and particularly circulating
tumor cells, have attracted extensive interest for the diagnosis
and monitoring of HCC (124).
Studies on other tumor-derived components, such as circulating
tumor DNA, sEVs and serum miRNAs are also increasing (125-127).
sEVs have the following advantages: i) Due to being
protected by the sEV membrane, sEV cargos have a high stability and
cannot easily degraded by lysosomes; ii) since the secretion of
sEVs is a normal physiological event for tumor cells, sEVs can be
detected in the majority of fluids, and their extraction is
relatively non-invasive; iii) compared with plasma biomarkers,
bioactive molecules from sEVs contain less interference of plasma;
and iv) markedly, cargos of sEVs have extensive homology with
recipient cells, which can confer sEVs superior sensitivity and
specificity than traditional methods (128-130). Differential ultracentrifugation
is the most common method used to separate sEVs from the cell
culture medium. However, each biological fluid presents specific
biophysical and chemical characteristics that render it different
from culture conditioned medium. Despite current mainstream
commercial kits are based on precipitation, which may result in EV
populations bound to or mixed with introduced components, such as
antibodies, beads or polymers; the majority of studies on the
potential clinical applications of sEVs use this method as it is
user-friendly, cost-effective and has potential for scale-up
production (17). Furthermore,
the efficiency and repeatability of sEVs separated using the
ExoQuick™ kit have been demonstrated to be comparable
with those of differential ultracentrifugation (131). Therefore, the articles cited in
the current section include those that using commercial kits to
isolate sEVs.
Numerous studies on the role of sEVs as HCC
promising biomarkers have been conducted (132-136). The importance of sEVs as HCC
biomarkers is reflected in numerous aspects, including the fact
that sEV cargos may serve as biomarkers for the early detection of
HCC; among these sEV cargos, miRNAs are the most extensively
investigated ones. For instance, the expression of miR-21 and
miR-10b in sEVs is markedly increased in patients with HCC compared
with that of healthy individuals and patients with CHB, indicating
that sEVs-carrying miR-21 and miR-10b may be used as early
diagnostic biomarkers for HCC (80). Similarly, by comparison with that
of patients with LC, the expression of miR-221, miR-192 and
miR-146a in exosomes was increased in patients with HCC, and Fründt
et al (137) indicated
that sEVs carrying miR-146a could distinguish patients with HCC
from patients with LC with an area under the curve value of
0.80±0.14 in a logistic regression model, and miR-96, miR-122,
miR-200a had similar effect (138-140). Other sEV cargos, such as
proteins and other non-coding RNAs (ncRNAs), including lncRNAs and
circRNAs, may also play a role in the preliminary diagnosis of HCC,
and it has been observed that LINC00161, circRNA 0006602, LDHC,
sphingosines, dilysocardiolipins, lysophosphatidylserines, and
(O-acyl)-1-hydroxy fatty acids are early diagnostic biomarker
candidates (48,121,141-143).
Apart from their early diagnostic value, sEV cargos
may be involved in the prediction of tumor staging and metastasis
(144-147). Exo-miR-1307-5p expression in
plasma has been found to be positively associated with tumor stage
and progression, while sEVs carrying miR-125b have been shown to
possess anti-metastatic features and are indicators of early
metastasis in HCC (148-150). Other sEV cargos can play a
similar role in tumor staging or metastasis prediction, and the
function of lncRNA ATB, hnRNPH1 and ASMTL-AS1 in this regard has
been reported (151-153). In addition, certain sEV cargos
may serve as prognostic indicators and may predict the prognosis of
patients with HCC. It has been corroborated that miR-638,
miR-150-3p, lncRNA CRNDE and circAKT3 in the sEVs of patients with
HCC are implicated in overall survival and disease-free survival
and may serve as independent indicators of a poor prognosis
(31,154-157). It should be noted that the
abnormal expression of certain sEV cargos, such as miR-718,
miR-125b and miR-92b is not only an effective tool to evaluate
survival, but is also a potential marker to predict the recurrence
of HCC (32,149,158). Notably, sEVs-carrying miR-122,
hsa-circRNA-G004213 and DANCR are also potential markers to
evaluate the efficacy of HCC surgical and interventional treatment
(153,159-161). In addition to the above, a panel
of tumor specific biochemical indicators has been proposed for a
higher sensitivity and specificity compared with single one
(162-166). For example, Sorop et al
(167) established exosomal miR
HCC Score including serum AFP and the level of plasma sEVs-carrying
miR-21-5p and miR-92a-3p with a great diagnostic ability of HCC
(AUC=0.85). Taken together, previous studies have demonstrated that
certain sEV cargos, including ncRNAs, mRNAs, lipids and proteins,
may serve as potential HCC diagnostic and prognostic biomarkers.
The potential HCC biomarkers are summarized in Table II and the efficacy of these
candidates warrants further validation.
Liver cancer is ranked as the third leading cause of
cancer-associated mortality worldwide due to resistance to
traditional drugs, as well as diagnosis in the late stage (1,168). The identification of novel drugs
for targeted therapy is imperative for patients with HCC (169). As aforementioned, the
distinctive property of sEVs in delivering functional molecules and
altering the biological behavior of recipient cells highlights
their potential application as ideal therapeutic vehicles in cancer
therapy, both at the theoretical and practical level. The
development of engineered sEVs, with a purpose of acting as
alternatives to chemotherapeutic and targeted agents, is currently
ongoing. sEVs have several advantages compared with previous drug
carriers, such as liposomes: First, sEVs achieve highly efficient
drug delivery due to their facility of penetrating biological
barriers. In addition, the cellular origin of sEVs makes them well
tolerated, and they can easily escape immune clearance, which also
reduces drug dose and toxicity (111,170,171); Furthermore, the heterogeneity of
proteins on the sEVs membrane facilitates the targeting abilities
of sEVs. In addition, sEVs are more biocompatible, safe and stable
than liposomes (172,173). Taken together, sEVs have a great
potential to serve as nano-carriers in the treatment field. The
present section mainly focuses on the advancements made in sEV
research as regards their application in therapy.
To achieve a better understanding of sEVs as
vehicles for therapeutic agents, methods of sEV preparation, the
engineering of sEVs and the selection of cargos are under
investigation, and the preliminary results are promising. Multiple
therapeutic agents, including chemotherapeutic drugs and nucleic
acids or their inhibitors, can be loaded. For example, in a
previous study, the subcutaneous injection of sEVs containing
miR-let-7a into a breast cancer mouse model exhibited an antitumor
ability by targeting EGFR (174). In HCC, recent research has
validated the importance of sEVs in the delivery of EV-packaged
drugs (175). A previous study
provided a prospective approach for generating sEV-associated
adeno-associated virus containing inducible caspase 9 (iCasp9)
suicide gene (Vexo-AAV6-iCasp9). The engineered sEVs possessed a
low immunogenicity and toxicity, and were readily absorbed by HCC
cells, consequently increasing HCC regression in an in vivo
xenograft model (176). Another
study encapsulated erastin (a typical ferroptosis inducer) and rose
bengal (RB, a well-known photosensitizer) into sEVs and engineered
CD47 on the surface of sEVs to protect the designed sEVs from
phagocytosis by macrophages. The sEVs induced obvious ferroptosis
in HCC, with minimized toxicity in the liver and kidneys (177). Apart from packing antitumor
payloads into sEVs, previous studies have developed nanoparticles
targeting specific adhesion or receptor proteins on the surface of
sEVs membranes for targeted delivery. Tian et al (80) designed a nano-drug based on the
PDCM system by targeting sEVs shuttling miR-21 and miR-10b, which
markedly decreased HCC growth and the numbers of metastatic lung
nodules. TDEs not only assist in exacerbating tumor progression,
but also increase the resistance of cancer cells to antitumor
treatments (178). Sorafenib and
transarterial chemoembolization have been considered optional
treatments for terminal-stage HCC for numerous years. Due to
acquired drug resistance to commonly used chemotherapeutic agents,
the clinical outcome and overall survival of patients with HCC
remain unsatisfactory (179).
The expression of programmed cell death protein 1 (PD-1) in HCC
tissues from patients with HCC who accepted sorafenib treatment was
upregulated and induced T-cell apoptosis (180). Therefore, immune checkpoint
inhibitors, such as commonly used PD-1 antibodies and PD-L1
anti-bodies have been introduced into medical practice as part of
the HCC regimen; however, the efficacy of the combination of
sorafenib and immunotherapy has not yet been fully elucidated. Shi
et al (180) treated
mouse models of HCC with the triple treatment of sorafenib, PD-1
antibodies and DC-derived exosomes (DEXs), which markedly prolonged
the survival time mice with HCC by comparison with the mice treated
with sorafenib alone, DEXs alone, or the combination of DEXs and
sorafenib. Taken together, these preclinical studies offer
encouragement for the application of sEVs as vehicles for HCC
treatment.
Despite the rapid development of advanced
techniques, major limitations remain in the current understanding
of sEVs vs. ideal treatment scenarios: i) For sEV technology to
play a role as a drug delivery vector, it is necessary to ensure
high purity and adequate production. There are still several
obstacles which hamper the efficacy of current methods, such as
being time consuming, having a high cost and generating polluting
by-products. Oncologists increase the total yield of sEVs by
intracellular calcium production, external stress, cytoskeletal
blocking, drug stimulation and the induction of gene expression
factors. Furthermore, sEVs usually represent heterogeneous
populations from different cell sources, and no standard separation
process has been established to date to achieve product consistency
(181); ii) the efficient
incorporation of external antitumor agents and molecules is another
demanding challenge that needs to be optimized. The high
drug-loading content in sEVs must be sufficient to obtain a
therapeutic response. Several approaches, including transfection,
electroporation and sonication (182,183), can be applied to upload the
desired biomolecules into sEVs. However, it is difficult to ensure
the integrity and biostability of the plasma membrane and the
function of sEVs; iii) the current evidence for the ability of sEVs
to deliver specific messages derives from cell culture studies. The
biodistribution and tissue or cellular tropism in vivo will
determine the application of therapeutic sEVs in clinical practice.
At present, there is insufficient evidence for in vivo and
clinical applications, which is a critical topic for future
research in this area. Due to being subjected to elimination by the
mononuclear phagocyte system, the half-life of sEVs in the systemic
circulation is relatively short (184). Thus, further studies on the
balance between prolonged circulation time and increased risk of
toxicity on major organs are warranted; and iv) currently, sEVs
need to be stored under-20 and -80°C in phosphate buffer saline
(185). Therefore, identifying a
suitable storage method is one of the barriers to be overcome.
In summary, while the application of sEVs as a
therapeutic drug delivery system remains in its infancy, the deeper
understanding of the aforementioned obstacles will provide a new
orientation for cancer nanomedicine and immunotherapy.
sEVs can trigger the alteration of gene expression
and induce aggressive behaviors in HCC cells; however, whether such
observations can be replicated in vivo needs to be further
investigated, since the precise isolation and high concentration of
cell culture-derived sEVs could not be achieved in the majority of
in vivo studies published thus far. Paradigm-shifting
findings in the field of HCC diagnosis have resulted in new avenues
for research on HCC biomarkers. Their easy availability,
vesicle-tethering stability and high donor-homology confer sEVs an
unparalleled advantage as HCC biomarkers compared with traditional
biomarkers. However, the clinical value of sEVs as HCC biomarkers
is still limited due to the absence of clinical research with large
sample sizes. Extensive efforts are currently being made to
identify sEVs biomarkers with high specificity and sensitivity, and
apply them to clinical practice. The role of sEVs in cancer therapy
has been studied extensively in recent years; however, research on
sEVs for HCC remains limited. Before applying them in clinical
practice, it is important to validate the purity, safety and
effectiveness of sEVs-encapsulated agents. Further research is
warranted to guarantee the homogeneity of sEVs, improve the
efficiency of their isolation methods and reduce the associated
side-effects. The targeting of sEVs is another issue that needs to
be resolved. Surface modification is a typical approach to harvest
targeted sEVs by modifying the proteins or peptides that
specifically expressed on the cell membrane through gene
transfection. Engineered sEVs can be selectively delivered to
target cells and reach the standard in terms of yield and targeted
therapy. However, the safety, mutagenesis and time-consuming limit
their clinical applications. Currently, aptamers, also known as
chemical antibodies, have attracted the attention of oncologists.
The majority of aptamers have been utilized to guide nanoparticles,
therapeutic and imaging agents to target locations in several
promising anticancer preclinical studies, whereby they are able to
modulate tumor retention and biodistribution. However, all these
issues cannot be solved in a short period of time, and as the
number of clinical studies increases, more patients will gain
clinical benefit from research in sEVs.
Not applicable.
GZ and PL conceptualized the study. SY and JW were
involved in the writing and preparation of the original draft. SW
and AZ were involved in the writing, reviewing and editing of the
manuscript. All authors have read and approved the final
manuscript. Data authentication is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
The present review article was funded by the National Natural
Science Foundation of China (grant no. 82070575), the Digestive
Medical Coordinated Development Center of Beijing Municipal
Administration of Hospitals (grant no. XXZ0205) and the Beijing
Municipal Science and Technology Commission (grant no.
Z191100006619080).
|
1
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Petrick JL, Florio AA, Znaor A, Ruggieri
D, Laversanne M, Alvarez CS, Ferlay J, Valery PC, Bray F and
McGlynn KA: International trends in hepatocellular carcinoma
incidence, 1978-2012. Int J Cancer. 147:317–330. 2020. View Article : Google Scholar :
|
|
3
|
Ringelhan M, McKeating JA and Protzer U:
Viral hepatitis and liver cancer. Philos Trans R Soc Lond B Biol
Sci. 372:201602742017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen SQ, Li J, Wang D, Fung H, Wong LY and
Zhao L: The hepatitis B epidemic in China should receive more
attention. Lancet. 391:15722018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sarin SK, Kumar M, Eslam M, George J, Al
Mahtab M, Akbar SMF, Jia J, Tian Q, Aggarwal R, Muljono DH, et al:
Liver diseases in the Asia-pacific region: A lancet
gastroenterology & hepatology commission. Lancet Gastroenterol
Hepatol. 5:167–228. 2020. View Article : Google Scholar :
|
|
6
|
Zhao C, Xing F, Yeo YH, Jin M, Le R, Le M,
Jin M, Henry L, Cheung R and Nguyen MH: Only one-third of
hepatocellular carcinoma cases are diagnosed via screening or
surveillance: A systematic review and meta-analysis. Eur J
Gastroenterol Hepatol. 32:406–419. 2020. View Article : Google Scholar
|
|
7
|
Wu G, Wu J, Wang B, Zhu X, Shi X and Ding
Y: Importance of tumor size at diagnosis as a prognostic factor for
hepatocellular carcinoma survival: A population-based study. Cancer
Manag Res. 10:4401–4410. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Johnstone RM, Adam M, Hammond JR, Orr L
and Turbide C: Vesicle formation during reticulocyte maturation.
Association of plasma membrane activities with released vesicles
(exosomes). J Biol Chem. 262:9412–9420. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen W, Mao Y, Liu C, Wu H and Chen S:
Exosome in hepatocellular carcinoma: An update. J Cancer.
12:2526–2536. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chang W, Xiao D, Fang X and Wang J:
Phospholipids in small extracellular vesicles: Emerging regulators
of neurodegenerative diseases and cancer. Cytotherapy. 24:93–100.
2022. View Article : Google Scholar
|
|
11
|
Sun F, Wang JZ, Luo JJ, Wang YQ and Pan Q:
Exosomes in the oncobiology, diagnosis, and therapy of hepatic
carcinoma: A new player of an old game. Biomed Res Int.
2018:27474612018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wen SW, Lima LG, Lobb RJ, Norris EL,
Hastie ML, Krumeich S and Möller A: Breast cancer-derived exosomes
reflect the cell-of-origin phenotype. Proteomics. 19:e18001802019.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang S, Xu M, Li X, Su X, Xiao X, Keating
A and Zhao RC: Exosomes released by hepatocarcinoma cells endow
adipocytes with tumor-promoting properties. J Hematol Oncol.
11:822018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zheng X, Hermann DM, Bähr M and Doeppner
TR: The role of small extracellular vesicles in cerebral and
myocardial ischemia-molecular signals, treatment targets, and
future clinical translation. Stem Cells. 39:403–413. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang L, He F, Gao L, Cong M, Sun J, Xu J,
Wang Y, Hu Y, Asghar S, Hu L and Qiao H: Engineering exosome-like
nanovesicles derived from asparagus cochinchinensis can inhibit the
proliferation of hepatocellular carcinoma cells with better safety
profile. Int J Nanomedicine. 16:1575–1586. 2021. View Article : Google Scholar :
|
|
16
|
Kim OY, Lee J and Gho YS: Extracellular
vesicle mimetics: Novel alternatives to extracellular vesicle-based
theranostics, drug delivery, and vaccines. Semin Cell Dev Biol.
67:74–82. 2017. View Article : Google Scholar
|
|
17
|
Théry C, Witwer KW, Aikawa E, Alcaraz MJ,
Anderson JD, Andriantsitohaina R, Antoniou A, Arab T, Archer F,
Atkin-Smith GK, et al: Minimal information for studies of
extracellular vesicles 2018 (MISEV2018): A position statement of
the international society for extracellular vesicles and update of
the MISEV2014 guidelines. J Extracell Vesicles. 7:15357502018.
View Article : Google Scholar
|
|
18
|
Murphy DE, de Jong OG, Brouwer M, Wood MJ,
Lavieu G, Schiffelers RM and Vader P: Extracellular vesicle-based
therapeutics: Natural versus engineered targeting and trafficking.
Exp Mol Med. 51:1–12. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Aslan C, Maralbashi S, Salari F, Kahroba
H, Sigaroodi F, Kazemi T and Kharaziha P: Tumor-derived exosomes:
Implication in angiogenesis and antiangiogenesis cancer therapy. J
Cell Physiol. 234:16885–16903. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Antimisiaris SG, Mourtas S and Marazioti
A: Exosomes and exosome-inspired vesicles for targeted drug
delivery. Pharmaceutics. 10:2182018. View Article : Google Scholar
|
|
21
|
Tian X, Shen H, Li Z, Wang T and Wang S:
Tumor-derived exosomes, myeloid-derived suppressor cells, and tumor
microenvironment. J Hematol Oncol. 12:842019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hessvik NP and Llorente A: Current
knowledge on exosome biogenesis and release. Cell Mol Life Sci.
75:193–208. 2018. View Article : Google Scholar :
|
|
23
|
Doyle LM and Wang MZ: Overview of
extracellular vesicles, their origin, composition, purpose, and
methods for exosome isolation and analysis. Cells. 8:7272019.
View Article : Google Scholar :
|
|
24
|
Farooqi AA, Desai NN, Qureshi MZ,
Librelotto DRN, Gasparri ML, Bishayee A, Nabavi SM, Curti V and
Daglia M: Exosome biogenesis, bioactivities and functions as new
delivery systems of natural compounds. Biotechnol Adv. 36:328–334.
2018. View Article : Google Scholar
|
|
25
|
Li X, Wang Y, Wang Q, Liu Y, Bao W and Wu
S: Exosomes in cancer: Small transporters with big functions.
Cancer Lett. 435:55–65. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xie JY, Wei JX, Lv LH, Han QF, Yang WB, Li
GL, Wang PX, Wu SB, Duan JX, Zhuo WF, et al: Angiopoietin-2 induces
angiogenesis via exosomes in human hepatocellular carcinoma. Cell
Commun Signal. 18:462020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhao Z, Yang S, Zhou A, Li X, Fang R,
Zhang S, Zhao G and Li P: Small extracellular vesicles in the
development, diagnosis, and possible therapeutic application of
esophageal squamous cell carcinoma. Front Oncol. 11:7327022021.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yu S, Cao H, Shen B and Feng J:
Tumor-derived exosomes in cancer progression and treatment failure.
Oncotarget. 6:37151–37168. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Greening DW, Xu R, Ji H, Tauro BJ and
Simpson RJ: A protocol for exosome isolation and characterization:
Evaluation of ultra-centrifugation, density-gradient separation,
and immunoaffinity capture methods. Methods Mol Biol. 1295:179–209.
2015. View Article : Google Scholar
|
|
30
|
Vaswani K, Koh YQ, Almughlliq FB, Peiris
HN and Mitchell MD: A method for the isolation and enrichment of
purified bovine milk exosomes. Reprod Biol. 17:341–348. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yugawa K, Yoshizumi T, Mano Y, Itoh S,
Harada N, Ikegami T, Kohashi K, Oda Y and Mori M: Cancer-associated
fibroblasts promote hepatocellular carcinoma progression through
downregulation of exosomal miR-150-3p. Eur J Surg Oncol.
47:384–393. 2021. View Article : Google Scholar
|
|
32
|
Nakano T, Chen IH, Wang CC, Chen PJ, Tseng
HP, Huang KT, Hu TH, Li LC, Goto S, Cheng YF, et al: Circulating
exosomal miR-92b: Its role for cancer immunoediting and clinical
value for prediction of posttransplant hepatocellular carcinoma
recurrence. Am J Transplant. 19:3250–3262. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Moh-Moh-Aung A, Fujisawa M, Ito S,
Katayama H, Ohara T, Ota Y, Yoshimura T and Matsukawa A: Decreased
miR-200b-3p in cancer cells leads to angiogenesis in HCC by
enhancing endothelial ERG expression. Sci Rep. 10:104182020.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fu X, Liu M, Qu S, Ma J, Zhang Y, Shi T,
Wen H, Yang Y, Wang S, Wang J, et al: Exosomal microRNA-32-5p
induces multidrug resistance in hepatocellular carcinoma via the
PI3K/Akt pathway. J Exp Clin Cancer Res. 37:522018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tang J, Li Y, Liu K, Zhu Q, Yang WH, Xiong
LK and Guo DL: Exosomal miR-9-3p suppresses HBGF-5 expression and
is a functional biomarker in hepatocellular carcinoma. Minerva Med.
109:15–23. 2018.
|
|
36
|
Xue X, Wang X, Zhao Y, Hu R and Qin L:
Exosomal miR-93 promotes proliferation and invasion in
hepatocellular carcinoma by directly inhibiting
TIMP2/TP53INP1/CDKN1A. Biochem Biophys Res Commun. 502:515–521.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang Q, Wang G, Niu L, Zhao S, Li J, Zhang
Z, Jiang H, Zhang Q, Wang H, Sun P, et al: Exosomal MiR-1290
promotes angiogenesis of hepatocellular carcinoma via targeting
SMEK1. J Oncol. 2021:66177002021.PubMed/NCBI
|
|
38
|
Ouyang Y, Tang Y, Fu L, Peng S, Wu W, Tan
D and Fu X: Exosomes secreted by chronic hepatitis B patients with
PNALT and liver inflammation grade ≥ A2 promoted the progression of
liver cancer by transferring miR-25-3p to inhibit the co-expression
of TCF21 and HHIP. Cell Prolif. 53:e128332020. View Article : Google Scholar
|
|
39
|
Shi Y, Yang X, Xue X, Sun D, Cai P, Song
Q, Zhang B and Qin L: HANR promotes lymphangiogenesis of
hepatocellular carcinoma via secreting miR-296 exosome and
regulating EAG1/VEGFA signaling in HDLEC cells. J Cell Biochem.
120:17699–17708. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Xiong L, Zhen S, Yu Q and Gong Z: HCV-E2
inhibits hepatocellular carcinoma metastasis by stimulating mast
cells to secrete exosomal shuttle microRNAs. Oncol Lett.
14:2141–2146. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chen W, Huang L, Liang J, Ye Y, He S and
Niu J: Hepatocellular carcinoma cells-derived exosomal
microRNA-378b enhances hepatocellular carcinoma angiogenesis. Life
Sci. 273:1191842021. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Fang JH, Zhang ZJ, Shang LR, Luo YW, Lin
YF, Yuan Y and Zhuang SM: Hepatoma cell-secreted exosomal
microRNA-103 increases vascular permeability and promotes
metastasis by targeting junction proteins. Hepatology.
68:1459–1475. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhang Z, Li X, Sun W, Yue S, Yang J, Li J,
Ma B, Wang J, Yang X, Pu M, et al: Loss of exosomal miR-320a from
cancer-associated fibroblasts contributes to HCC proliferation and
metastasis. Cancer Lett. 397:33–42. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Bai ZZ, Li HY, Li CH, Sheng CL and Zhao
XN: M1 macrophage-derived exosomal MicroRNA-326 suppresses
hepatocellular carcinoma cell progression via mediating NF-κB
signaling pathway. Nanoscale Res Lett. 15:2212020. View Article : Google Scholar
|
|
45
|
Chen QL, Xie CF, Feng KL, Cui DY, Sun SL,
Zhang JC, Xiong CM, Huang JH and Chong Z: microRNAs carried by
exosomes promote epithelial-mesenchymal transition and metastasis
of liver cancer cells. Am J Transl Res. 12:6811–6826.
2020.PubMed/NCBI
|
|
46
|
Fan F, Chen K, Lu X, Li A, Liu C and Wu B:
Dual targeting of PD-L1 and PD-L2 by PCED1B-AS1 via sponging
hsa-miR-194-5p induces immunosuppression in hepatocellular
carcinoma. Hepatol Int. 15:444–458. 2021. View Article : Google Scholar
|
|
47
|
Wang LP, Lin J, Ma XQ, Xu DY, Shi CF, Wang
W and Jiang XJ: Exosomal DLX6-AS1 from hepatocellular carcinoma
cells induces M2 macrophage polarization to promote migration and
invasion in hepatocellular carcinoma through microRNA-15a-5p/CXCL17
axis. J Exp Clin Cancer Res. 40:1772021. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sun L, Su Y, Liu X, Xu M, Chen X, Zhu Y,
Guo Z, Bai T, Dong L, Wei C, et al: Serum and exosome long non
coding RNAs as potential biomarkers for hepatocellular carcinoma. J
Cancer. 9:2631–2639. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang J, Pu J, Zhang Y, Yao T, Luo Z, Li W,
Xu G, Liu J, Wei W and Deng Y: Exosome-transmitted long non-coding
RNA SENP3-EIF4A1 suppresses the progression of hepatocellular
carcinoma. Aging (Albany NY). 12:11550–11567. 2020. View Article : Google Scholar
|
|
50
|
Li X and Li N: LINC ROR from
hepatocarcinoma cell-derived exosomes modulates inflammation in
human macrophages. Sichuan Da Xue Xue Bao Yi Xue Ban. 50:177–181.
2019.In Chinese. PubMed/NCBI
|
|
51
|
Li B, Mao R, Liu C, Zhang W, Tang Y and
Guo Z: LncRNA FAL1 promotes cell proliferation and migration by
acting as a CeRNA of miR-1236 in hepatocellular carcinoma cells.
Life Sci. 197:122–129. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhang PF, Gao C, Huang XY, Lu JC, Guo XJ,
Shi GM, Cai JB and Ke AW: Cancer cell-derived exosomal circUHRF1
induces natural killer cell exhaustion and may cause resistance to
anti-PD1 therapy in hepatocellular carcinoma. Mol Cancer.
19:1102020. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Su Y, Lv X, Yin W, Zhou L, Hu Y, Zhou A
and Qi F: CircRNA Cdr1as functions as a competitive endogenous RNA
to promote hepatocellular carcinoma progression. Aging (Albany NY).
11:8183–8203. 2019. View Article : Google Scholar
|
|
54
|
Xu J, Ji L, Liang Y, Wan Z, Zheng W, Song
X, Gorshkov K, Sun Q, Lin H, Zheng X, et al: CircRNA-SORE mediates
sorafenib resistance in hepatocellular carcinoma by stabilizing
YBX1. Signal Transduct Target Ther. 5:2982020. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zhu C, Su Y, Liu L, Wang S, Liu Y and Wu
J: Circular RNA hsa_circ_0004277 stimulates malignant phenotype of
hepatocellular carcinoma and epithelial-mesenchymal transition of
peripheral cells. Front Cell Dev Biol. 8:5855652021. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Yu Y, Bian L, Liu R, Wang Y and Xiao X:
Circular RNA hsa_circ_0061395 accelerates hepatocellular carcinoma
progression via regulation of the miR-877-5p/PIK3R3 axis. Cancer
Cell Int. 21:102021. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Li Y, Zang H, Zhang X and Huang G:
Exosomal circ-ZNF652 promotes cell proliferation, migration,
invasion and glycolysis in hepatocellular carcinoma via
miR-29a-3p/GUCD1 axis. Cancer Manag Res. 12:7739–7751. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Hu K, Li NF, Li JR, Chen ZG, Wang JH and
Sheng LQ: Exosome circCMTM3 promotes angiogenesis and tumorigenesis
of hepatocellular carcinoma through miR-3619-5p/SOX9. Hepatol Res.
51:1139–1152. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Liu D, Kang H, Gao M, Jin L, Zhang F, Chen
D, Li M and Xiao L: Exosome-transmitted circ_MMP2 promotes
hepatocellular carcinoma metastasis by upregulating MMP2. Mol
Oncol. 14:1365–1380. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Wang G, Liu W, Zou Y, Wang G, Deng Y, Luo
J, Zhang Y, Li H, Zhang Q, Yang Y and Chen G: Three isoforms of
exosomal circPTGR1 promote hepatocellular carcinoma metastasis via
the miR449a-MET pathway. EBioMedicine. 40:432–445. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Chen W, Quan Y, Fan S, Wang H, Liang J,
Huang L, Chen L, Liu Q, He P and Ye Y: Exosome-transmitted circular
RNA hsa_circ_0051443 suppresses hepatocellular carcinoma
progression. Cancer Lett. 475:119–128. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Huang C, Yu W, Wang Q, Huang T and Ding Y:
CircANTXR1 contributes to the malignant progression of
hepatocellular carcinoma by promoting proliferation and metastasis.
J Hepatocell Carcinoma. 8:1339–1353. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Dai W, Wang Y, Yang T, Wang J, Wu W and Gu
J: Downregulation of exosomal CLEC3B in hepatocellular carcinoma
promotes metastasis and angiogenesis via AMPK and VEGF signals.
Cell Commun Signal. 17:1132019. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Huang A, Dong J, Li S, Wang C, Ding H, Li
H, Su X, Ge X, Sun L, Bai C, et al: Exosomal transfer of vasorin
expressed in hepatocellular carcinoma cells promotes migration of
human umbilical vein endothelial cells. Int J Biol Sci. 11:961–969.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Cheng Z, Lei Z, Yang P, Si A, Xiang D,
Tang X, Guo G, Zhou J and Hüser N: Exosome-transmitted p120-catenin
suppresses hepatocellular carcinoma progression via STAT3 pathways.
Mol Carcinog. 58:1389–1399. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Li M, Lu Y, Xu Y, Wang J, Zhang C, Du Y,
Wang L, Li L, Wang B, Shen J, et al: Horizontal transfer of
exosomal CXCR4 promotes murine hepatocarcinoma cell migration,
invasion and lymphangiogenesis. Gene. 676:101–109. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Xia Y, Zhen L, Li H, Wang S, Chen S, Wang
C and Yang X: MIRLET7BHG promotes hepatocellular carcinoma
progression by activating hepatic stellate cells through exosomal
SMO to trigger Hedgehog pathway. Cell Death Dis. 12:3262021.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Fu Q, Zhang Q, Lou Y, Yang J, Nie G, Chen
Q, Chen Y, Zhang J, Wang J, Wei T, et al: Primary tumor-derived
exosomes facilitate metastasis by regulating adhesion of
circulating tumor cells via SMAD3 in liver cancer. Oncogene.
37:6105–6118. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Zhou Y, Zhang Y, Gong H, Luo S and Cui Y:
The role of exosomes and their applications in cancer. Int J Mol
Sci. 22:122042021. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Wang H, Lu Z and Zhao X: Tumorigenesis,
diagnosis, and therapeutic potential of exosomes in liver cancer. J
Hematol Oncol. 12:1332019. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Cao LQ, Yang XW, Chen YB, Zhang DW, Jiang
XF and Xue P: Exosomal miR-21 regulates the TETs/PTENp1/PTEN
pathway to promote hepatocellular carcinoma growth. Mol Cancer.
18:1482019. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Fu X, Tang Y, Wu W, Ouyang Y, Tan D and
Huang Y: Exosomal microRNA-25 released from cancer cells targets
SIK1 to promote hepatocellular carcinoma tumorigenesis. Dig Liver
Dis. S1590-8658(21): 00385–6. 2021.Epub ahead of print.
|
|
73
|
Huang X, Sun L, Wen S, Deng D, Wan F, He
X, Tian L, Liang L, Wei C, Gao K, et al: RNA sequencing of plasma
exosomes revealed novel functional long noncoding RNAs in
hepatocellular carcinoma. Cancer Sci. 111:3338–3349. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Jiang K, Dong C, Yin Z, Li R, Mao J, Wang
C, Zhang J, Gao Z, Liang R, Wang Q and Wang L: Exosome-derived ENO1
regulates integrin α6β4 expression and promotes hepatocellular
carcinoma growth and metastasis. Cell Death Dis. 11:9722020.
View Article : Google Scholar
|
|
75
|
Lai Z, Wei T, Li Q, Wang X, Zhang Y and
Zhang S: Exosomal circFBLIM1 promotes hepatocellular carcinoma
progression and glycolysis by regulating the miR-338/LRP6 axis.
Cancer Biother Radiopharm. Sep 9–2020.Epub ahead of print.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Ma YS, Liu JB, Lin L, Zhang H, Wu JJ, Shi
Y, Jia CY, Zhang DD, Yu F, Wang HM, et al: Exosomal microRNA-15a
from mesenchymal stem cells impedes hepatocellular carcinoma
progression via downregulation of SALL4. Cell Death Discov.
7:2242021. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Yin C, Han Q, Xu D, Zheng B, Zhao X and
Zhang J: SALL4-mediated upregulation of exosomal miR-146a-5p drives
T-cell exhaustion by M2 tumor-associated macrophages in HCC.
Oncoimmunology. 8:16014792019. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Conigliaro A, Costa V, Lo Dico A, Saieva
L, Buccheri S, Dieli F, Manno M, Raccosta S, Mancone C, Tripodi M,
et al: CD90+ liver cancer cells modulate endothelial cell phenotype
through the release of exosomes containing H19 lncRNA. Mol Cancer.
14:1552015. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Li R, Wang Y, Zhang X, Feng M, Ma J, Li J,
Yang X, Fang F, Xia Q, Zhang Z, et al: Exosome-mediated secretion
of LOXL4 promotes hepatocellular carcinoma cell invasion and
metastasis. Mol Cancer. 18:182019. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Tian XP, Wang CY, Jin XH, Li M, Wang FW,
Huang WJ, Yun JP, Xu RH, Cai QQ and Xie D: Acidic microenvironment
up-regulates exosomal miR-21 and miR-10b in early-stage
hepatocellular carcinoma to promote cancer cell proliferation and
metastasis. Theranostics. 9:1965–1979. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Liu H, Chen W, Zhi X, Chen EJ, Wei T,
Zhang J, Shen J, Hu LQ, Zhao B, Feng XH, et al: Tumor-derived
exosomes promote tumor self-seeding in hepatocellular carcinoma by
transferring miRNA-25-5p to enhance cell motility. Oncogene.
37:4964–4978. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Liu G, Ouyang X, Sun Y, Xiao Y, You B, Gao
Y, Yeh S, Li Y and Chang C: The miR-92a-2-5p in exosomes from
macrophages increases liver cancer cells invasion via altering the
AR/PHLPP/p-AKT/β-catenin signaling. Cell Death Differ.
27:3258–3272. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Liu C, Zhou X, Long Q, Zeng H, Sun Q, Chen
Y, Wu D and Liu L: Small extracellular vesicles containing
miR-30a-3p attenuate the migration and invasion of hepatocellular
carcinoma by targeting SNAP23 gene. Oncogene. 40:233–245. 2021.
View Article : Google Scholar
|
|
84
|
Zeng Y, Yao X, Liu X, He X, Li L, Liu X,
Yan Z, Wu J and Fu BM: Anti-angiogenesis triggers exosomes release
from endothelial cells to promote tumor vasculogenesis. J Extracell
Vesicles. 8:16298652019. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Wu Q, Zhou L, Lv D, Zhu X and Tang H:
Exosome-mediated communication in the tumor microenvironment
contributes to hepatocellular carcinoma development and
progression. J Hematol Oncol. 12:532019. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Yang Y, Han Q, Hou Z, Zhang C, Tian Z and
Zhang J: Exosomes mediate hepatitis B virus (HBV) transmission and
NK-cell dysfunction. Cell Mol Immunol. 14:465–475. 2017. View Article : Google Scholar :
|
|
87
|
Wu W, Wu D, Yan W, Wang Y, You J, Wan X,
Xi D, Luo X, Han M and Ning Q: Interferon-induced
macrophage-derived exosomes mediate antiviral activity against
hepatitis B virus through miR-574-5p. J Infect Dis. 223:686–698.
2021. View Article : Google Scholar
|
|
88
|
Liu QM, He YY, Liu LL and Wang LK:
Exosomal lncRNA HOTTIP mediates antiviral effect of tenofovir
alafenamide (TAF) on HBV infection. J Inflamm Res. 14:5489–5500.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Lu J, Wu J, Xie F, Tian J, Tang X, Guo H,
Ma J, Xu P, Mao L, Xu H and Wang S: CD4+ T cell-released
extracellular vesicles potentiate the efficacy of the HBsAg vaccine
by enhancing B cell responses. Adv Sci (Weinh). 6:18022192019.
View Article : Google Scholar
|
|
90
|
Wei Y, Tang X, Ren Y, Yang Y, Song F, Fu
J, Liu S, Yu M, Chen L, Wang S, et al: An RNA-RNA crosstalk network
involving HMGB1 and RICTOR facilitates hepatocellular carcinoma
tumorigenesis by promoting glutamine metabolism and impedes
immunotherapy by PD-L1+ exosomes activity. Signal Transduct Target
Ther. 6:4212021. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Zhou Y, Ren H, Dai B, Li J, Shang L, Huang
J and Shi X: Hepatocellular carcinoma-derived exosomal miRNA-21
contributes to tumor progression by converting hepatocyte stellate
cells to cancer-associated fibroblasts. J Exp Clin Cancer Res.
37:3242018. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Fang T, Lv H, Lv G, Li T, Wang C, Han Q,
Yu L, Su B, Guo L, Huang S, et al: Tumor-derived exosomal
miR-1247-3p induces cancer-associated fibroblast activation to
foster lung metastasis of liver cancer. Nat Commun. 9:1912018.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Lou G, Song X, Yang F, Wu S, Wang J, Chen
Z and Liu Y: Exosomes derived from miR-122-modified adipose
tissue-derived MSCs increase chemosensitivity of hepatocellular
carcinoma. J Hematol Oncol. 8:1222015. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Lin M, Liao W, Dong M, Zhu R, Xiao J, Sun
T, Chen Z, Wu B and Jin J: Exosomal neutral sphingomyelinase 1
suppresses hepatocellular carcinoma via decreasing the ratio of
sphingomyelin/ceramide. FEBS J. 285:3835–3848. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Lin H, Zhang R, Wu W and Lei L: miR-4454
promotes hepatic carcinoma progression by targeting Vps4A and
Rab27A. Oxid Med Cell Longev. 2021:92304352021. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Sun JF, Zhang D, Gao CJ, Zhang YW and Dai
QS: Exosome-mediated MiR-155 transfer contributes to hepatocellular
carcinoma cell proliferation by targeting PTEN. Med Sci Monit Basic
Res. 25:218–228. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Li YH, Lv MF, Lu MS and Bi JP: Bone marrow
mesenchymal stem cell-derived exosomal MiR-338-3p represses
progression of hepatocellular carcinoma by targeting ETS1. J Biol
Regul Homeost Agents. 35:617–627. 2021.PubMed/NCBI
|
|
98
|
Wang D, Xing N, Yang T, Liu J, Zhao H, He
J, Ai Y and Yang J: Exosomal lncRNA H19 promotes the progression of
hepatocellular carcinoma treated with Propofol via
miR-520a-3p/LIMK1 axis. Cancer Med. 9:7218–7230. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Li S, Qi Y, Huang Y, Guo Y, Huang T and
Jia L: Exosome-derived SNHG16 sponging miR-4500 activates HUVEC
angiogenesis by targeting GALNT1 via PI3K/Akt/mTOR pathway in
hepatocellular carcinoma. J Physiol Biochem. 77:667–682. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Lin XJ, Fang JH, Yang XJ, Zhang C, Yuan Y,
Zheng L and Zhuang SM: Hepatocellular carcinoma cell-secreted
exosomal MicroRNA-210-promotes angiogenesis in vitro and in vivo.
Mol Ther Nucleic Acids. 11:243–252. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Dong SS, Dong DD, Yang ZF, Zhu GQ, Gao DM,
Chen J, Zhao Y and Liu BB: Exosomal miR-3682-3p suppresses
angiogenesis by targeting ANGPT1 via the RAS-MEK1/2ERK1/2 pathway
in hepatocellular carcinoma. Front Cell Dev Biol. 9:6333582021.
View Article : Google Scholar
|
|
102
|
Huang XY, Huang ZL, Huang J, Xu B, Huang
XY, Xu YH, Zhou J and Tang ZY: Exosomal circRNA-100338 promotes
hepatocellular carcinoma metastasis via enhancing invasiveness and
angiogenesis. J Exp Clin Cancer Res. 39:202020. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Yang B, Feng X, Liu H, Tong R, Wu J, Li C,
Yu H, Chen Y, Cheng Q, Chen J, et al: High-metastatic cancer cells
derived exosomal miR92a-3p promotes epithelial-mesenchymal
transition and metastasis of low-metastatic cancer cells by
regulating PTEN/Akt pathway in hepatocellular carcinoma. Oncogene.
39:6529–6543. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Chen L, Guo P, He Y, Chen Z, Chen L, Luo
Y, Qi L, Liu Y, Wu Q, Cui Y, et al: HCC-derived exosomes elicit HCC
progression and recurrence by epithelial-mesenchymal transition
through MAPK/ERK signalling pathway. Cell Death Dis. 9:5132018.
View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Lin Q, Zhou CR, Bai MJ, Zhu D, Chen JW,
Wang HF, Li MA, Wu C, Li ZR and Huang MS: Exosome-mediated miRNA
delivery promotes liver cancer EMT and metastasis. Am J Transl Res.
12:1080–1095. 2020.PubMed/NCBI
|
|
106
|
Zhuo C, Yi T, Pu J, Cen X, Zhou Y, Feng S,
Wei C, Chen P, Wang W, Bao C, et al: Exosomal linc-FAM138B from
cancer cells alleviates hepatocellular carcinoma progression via
regulating miR-765. Aging (Albany NY). 12:26236–26247. 2020.
View Article : Google Scholar
|
|
107
|
Wu J, Gao W, Tang Q, Yu Y, You W, Wu Z,
Fan Y, Zhang L, Wu C, Han G, et al: M2 macrophage-derived exosomes
facilitate HCC metastasis by transferring αM β2 integrin to tumor
cells. Hepatology. 73:1365–1380. 2021. View Article : Google Scholar
|
|
108
|
Wang Y, Gao R, Li J, Tang S and Li S, Tong
Q and Li S: Downregulation of hsa_circ_0074854 suppresses the
migration and invasion in hepatocellular carcinoma via interacting
with HuR and via suppressing exosomes-mediated macrophage M2
polarization. Int J Nanomedicine. 16:2803–2818. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Li X, Lei Y, Wu M and Li N: Regulation of
macrophage activation and polarization by HCC-derived exosomal
lncRNA TUC339. Int J Mol Sci. 19:29582018. View Article : Google Scholar :
|
|
110
|
Ozga AJ, Chow MT and Luster AD: Chemokines
and the immune response to cancer. Immunity. 54:859–874. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Syn NL, Wang L, Chow EK, Lim CT and Goh
BC: Exosomes in cancer nanomedicine and immunotherapy: Prospects
and challenges. Trends Biotechnol. 35:665–676. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Wang X, Shen H, Zhangyuan G, Huang R,
Zhang W, He Q, Jin K, Zhuo H, Zhang Z, Wang J, et al: 14-3-3ζ
delivered by hepatocellular carcinoma-derived exosomes impaired
anti-tumor function of tumor-infiltrating T lymphocytes. Cell Death
Dis. 9:1592018. View Article : Google Scholar
|
|
113
|
Ye L, Zhang Q, Cheng Y, Chen X, Wang G,
Shi M, Zhang T, Cao Y, Pan H, Zhang L, et al: Tumor-derived
exosomal HMGB1 fosters hepatocellular carcinoma immune evasion by
promoting TIM-1+ regulatory B cell expansion. J
Immunother Cancer. 6:1452018. View Article : Google Scholar
|
|
114
|
Li H, Yang C, Shi Y and Zhao L: Exosomes
derived from siRNA against GRP78 modified bone-marrow-derived
mesenchymal stem cells suppress Sorafenib resistance in
hepatocellular carcinoma. J Nanobiotechnology. 16:1032018.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Liu Y, Tan J, Ou S, Chen J and Chen L:
Adipose-derived exosomes deliver miR-23a/b to regulate tumor growth
in hepatocellular cancer by targeting the VHL/HIF axis. J Physiol
Biochem. 75:391–401. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Huang H, Hou J, Liu K, Liu Q, Shen L, Liu
B, Lu Q, Zhang N, Che L, Li J, et al: RAB27A-dependent release of
exosomes by liver cancer stem cells induces Nanog expression in
their differentiated progenies and confers regorafenib resistance.
J Gastroenterol Hepatol. 36:3429–3437. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Xu Y, Lai Y, Cao L, Li Y, Chen G, Chen L,
Weng H, Chen T, Wang L and Ye Y: Human umbilical cord mesenchymal
stem cells-derived exosomal microRNA-451a represses
epithelial-mesenchymal transition of hepatocellular carcinoma cells
by inhibiting ADAM10. RNA Biol. 18:14232021. View Article : Google Scholar
|
|
118
|
Wang G, Zhao W, Wang H, Qiu G, Jiang Z,
Wei G and Li X: Exosomal MiR-744 inhibits proliferation and
sorafenib chemoresistance in hepatocellular carcinoma by targeting
PAX2. Med Sci Monit. 25:7209–7217. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Pinyol R, Montal R, Bassaganyas L, Sia D,
Takayama T, Chau GY, Mazzaferro V, Roayaie S, Lee HC, Kokudo N, et
al: Molecular predictors of prevention of recurrence in HCC with
sorafenib as adjuvant treatment and prognostic factors in the phase
3 STORM trial. Gut. 68:1065–1075. 2019. View Article : Google Scholar
|
|
120
|
Zhang Y, Xi H, Nie X, Zhang P, Lan N, Lu
Y, Liu J and Yuan W: Assessment of miR-212 and other biomarkers in
the diagnosis and treatment of HBV-infection-related liver
diseases. Curr Drug Metab. 20:785–798. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Guo S, Hu C, Zhai X and Sun D: Circular
RNA 0006602 in plasma exosomes: A new potential diagnostic
biomarker for hepatocellular carcinoma. Am J Transl Res.
13:6001–6015. 2021.PubMed/NCBI
|
|
122
|
De Stefano F, Chacon E, Turcios L, Marti F
and Gedaly R: Novel biomarkers in hepatocellular carcinoma. Dig
Liver Dis. 50:1115–1123. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Galle PR, Foerster F, Kudo M, Chan SL,
Llovet JM, Qin S, Schelman WR, Chintharlapalli S, Abada PB, Sherman
M and Zhu AX: Biology and significance of alpha-fetoprotein in
hepatocellular carcinoma. Liver Int. 39:2214–2229. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Ahn JC, Teng PC, Chen PJ, Posadas E, Tseng
HR, Lu SC and Yang JD: Detection of circulating tumor cells and
their implications as a biomarker for diagnosis, prognostication,
and therapeutic monitoring in hepatocellular carcinoma. Hepatology.
73:422–436. 2021. View Article : Google Scholar
|
|
125
|
von Felden J, Garcia-Lezana T, Schulze K,
Losic B and Villanueva A: Liquid biopsy in the clinical management
of hepatocellular carcinoma. Gut. 69:2025–2034. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Mjelle R, Dima SO, Bacalbasa N, Chawla K,
Sorop A, Cucu D, Herlea V, Sætrom P and Popescu I: Comprehensive
transcriptomic analyses of tissue, serum, and serum exosomes from
hepatocellular carcinoma patients. BMC Cancer. 19:10072019.
View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Sheng LQ, Li JR, Qin H, Liu L, Zhang DD,
Zhang Q, Huang ML, Li XL, Xu XY, Wei YN, et al: Blood exosomal
micro ribonucleic acid profiling reveals the complexity of
hepatocellular carcinoma and identifies potential biomarkers for
differential diagnosis. World J Gastrointest Oncol. 12:1195–1208.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Thind A and Wilson C: Exosomal miRNAs as
cancer biomarkers and therapeutic targets. J Extracell Vesicles.
5:312922016. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Salehi M and Sharifi M: Exosomal miRNAs as
novel cancer biomarkers: Challenges and opportunities. J Cell
Physiol. 233:6370–6380. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Fitts CA, Ji N, Li Y and Tan C: Exploiting
exosomes in cancer liquid biopsies and drug delivery. Adv Healthc
Mater. 8:e18012682019. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Caradec J, Kharmate G, Hosseini-Beheshti
E, Adomat H, Gleave M and Guns E: Reproducibility and efficiency of
serum-derived exosome extraction methods. Clin Biochem.
47:1286–1292. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Ge Y, Mu W, Ba Q, Li J, Jiang Y, Xia Q and
Wang H: Hepatocellular carcinoma-derived exosomes in organotropic
metastasis, recurrence and early diagnosis application. Cancer
Lett. 477:41–48. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Zhang C, Yang X, Qi Q, Gao Y, Wei Q and
Han S: lncRNA-HEIH in serum and exosomes as a potential biomarker
in the HCV-related hepatocellular carcinoma. Cancer Biomark.
21:651–659. 2018. View Article : Google Scholar
|
|
134
|
Lyu L, Yang W, Yao J, Wang H, Zhu J, Jin
A, Liu T, Wang B, Zhou J, Fan J, et al: The diagnostic value of
plasma exosomal hsa_circ_0070396 for hepatocellular carcinoma.
Biomark Med. 15:359–371. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Cho HJ, Baek GO, Seo CW, Ahn HR, Sung S,
Son JA, Kim SS, Cho SW, Jang JW, Nam SW, et al: Exosomal
microRNA-4661-5p-based serum panel as a potential diagnostic
biomarker for early-stage hepatocellular carcinoma. Cancer Med.
9:5459–5472. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Cui Y, Xu HF, Liu MY, Xu YJ, He JC, Zhou Y
and Cang SD: Mechanism of exosomal microRNA-224 in development of
hepatocellular carcinoma and its diagnostic and prognostic value.
World J Gastroenterol. 25:1890–1898. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Fründt T, Krause L, Hussey E, Steinbach B,
Köhler D, von Felden J, Schulze K, Lohse AW, Wege H and
Schwarzenbach H: Diagnostic and prognostic value of miR-16,
miR-146a, miR-192 and miR-221 in exosomes of hepatocellular
carcinoma and liver cirrhosis patients. Cancers (Basel).
13:24842021. View Article : Google Scholar
|
|
138
|
Wang S, Yang Y, Sun L, Qiao G, Song Y and
Liu B: Exosomal MicroRNAs as liquid biopsy biomarkers in
hepatocellular carcinoma. Onco Targets Ther. 13:2021–2030. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Liu WH, Ren LN, Wang X, Wang T, Zhang N,
Gao Y, Luo H, Navarro-Alvarez N and Tang LJ: Combination of
exosomes and circulating microRNAs may serve as a promising tumor
marker complementary to alpha-fetoprotein for early-stage
hepatocellular carcinoma diagnosis in rats. J Cancer Res Clin
Oncol. 141:1767–1778. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Wang Y, Zhang C, Zhang P, Guo G, Jiang T,
Zhao X, Jiang J, Huang X, Tong H and Tian Y: Serum exosomal
microRNAs combined with alpha-fetoprotein as diagnostic markers of
hepatocellular carcinoma. Cancer Med. 7:1670–1679. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Cui Z, Li Y, Gao Y, Kong L, Lin Y and Chen
Y: Cancer-testis antigen lactate dehydrogenase C4 in hepatocellular
carcinoma: A promising biomarker for early diagnosis, efficacy
evaluation and prognosis prediction. Aging (Albany NY).
12:19455–19467. 2020.Epub ahead of print. View Article : Google Scholar
|
|
142
|
Kim SS, Baek GO, Ahn HR, Sung S, Seo CW,
Cho HJ, Nam SW, Cheong JY and Eun JW: Serum small extracellular
vesicle-derived LINC00853 as a novel diagnostic marker for early
hepatocellular carcinoma. Mol Oncol. 14:2646–2659. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Sanchez JI, Jiao J, Kwan SY, Veillon L,
Warmoes MO, Tan L, Odewole M, Rich NE, Wei P, Lorenzi PL, et al:
Lipidomic profiles of plasma exosomes identify candidate biomarkers
for early detection of hepatocellular carcinoma in patients with
cirrhosis. Cancer Prev Res (Phila). 14:955–962. 2021. View Article : Google Scholar
|
|
144
|
Li W, Ding X, Wang S, Xu L, Yin T, Han S,
Geng J and Sun W: Downregulation of serum exosomal miR-320d
predicts poor prognosis in hepatocellular carcinoma. J Clin Lab
Anal. 34:e232392020.PubMed/NCBI
|
|
145
|
Qu Z, Wu J, Wu J, Ji A, Qiang G, Jiang Y,
Jiang C and Ding Y: Exosomal miR-665 as a novel minimally invasive
biomarker for hepatocellular carcinoma diagnosis and prognosis.
Oncotarget. 8:80666–80678. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Xu H, Chen Y, Dong X and Wang X: Serum
exosomal long noncoding RNAs ENSG00000258332.1 and LINC00635 for
the diagnosis and prognosis of hepatocellular carcinoma. Cancer
Epidemiol Biomarkers Prev. 27:710–716. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Cho HJ, Baek GO, Yoon MG, Ahn HR, Son JA,
Kim SS, Cheong JY and Eun JW: Overexpressed proteins in HCC
cell-derived exosomes, CCT8, and cofilin-1 are potential biomarkers
for patients with HCC. Diagnostics (Basel). 11:12212021. View Article : Google Scholar
|
|
148
|
Eun JW, Seo CW, Baek GO, Yoon MG, Ahn HR,
Son JA, Sung S, Kim DW, Kim SS, Cho HJ and Cheong JY: Circulating
exosomal MicroRNA-1307-5p as a predictor for metastasis in patients
with hepatocellular carcinoma. Cancers (Basel). 12:38192020.
View Article : Google Scholar
|
|
149
|
Liu W, Hu J, Zhou K, Chen F, Wang Z, Liao
B, Dai Z, Cao Y, Fan J and Zhou J: Serum exosomal miR-125b is a
novel prognostic marker for hepatocellular carcinoma. Onco Targets
Ther. 10:3843–3851. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Kim HS, Kim JS, Park NR, Nam H, Sung PS,
Bae SH, Choi JY, Yoon SK, Hur W and Jang JW: Exosomal miR-125b
exerts anti-metastatic properties and predicts early metastasis of
hepatocellular carcinoma. Front Oncol. 11:6372472021. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Lee YR, Kim G, Tak WY, Jang SY, Kweon YO,
Park JG, Lee HW, Han YS, Chun JM, Park SY and Hur K: Circulating
exosomal noncoding RNAs as prognostic biomarkers in human
hepatocellular carcinoma. Int J Cancer. 144:1444–1452. 2019.
View Article : Google Scholar
|
|
152
|
Xu H, Dong X, Chen Y and Wang X: Serum
exosomal hnRNPH1 mRNA as a novel marker for hepatocellular
carcinoma. Clin Chem Lab Med. 56:479–484. 2018. View Article : Google Scholar
|
|
153
|
Ma D, Gao X, Liu Z, Lu X, Ju H and Zhang
N: Exosome-transferred long non-coding RNA ASMTL-AS1 contributes to
malignant phenotypes in residual hepatocellular carcinoma after
insufficient radiofrequency ablation. Cell Prolif. 53:e127952020.
View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Shi M, Jiang Y, Yang L, Yan S, Wang YG and
Lu XJ: Decreased levels of serum exosomal miR-638 predict poor
prognosis in hepatocellular carcinoma. J Cell Biochem.
119:4711–4716. 2018. View Article : Google Scholar
|
|
155
|
Wang T, Zhu H, Xiao M and Zhou S: Serum
exosomal long noncoding RNA CRNDE as a prognostic biomarker for
hepatocellular carcinoma. J Clin Lab Anal. 35:e239592021.
View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Luo Y, Liu F and Gui R: High expression of
circulating exosomal circAKT3 is associated with higher recurrence
in HCC patients undergoing surgical treatment. Surg Oncol.
33:276–281. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Yokota Y, Noda T, Okumura Y, Kobayashi S,
Iwagami Y, Yamada D, Tomimaru Y, Akita H, Gotoh K, Takeda Y, et al:
Serum exosomal miR-638 is a prognostic marker of HCC via
downregulation of VE-cadherin and ZO-1 of endothelial cells. Cancer
Sci. 112:1275–1288. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Sugimachi K, Matsumura T, Hirata H, Uchi
R, Ueda M, Ueo H, Shinden Y, Iguchi T, Eguchi H, Shirabe K, et al:
Identification of a bona fide microRNA biomarker in serum exosomes
that predicts hepatocellular carcinoma recurrence after liver
transplantation. Br J Cancer. 112:532–538. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Suehiro T, Miyaaki H, Kanda Y, Shibata H,
Honda T, Ozawa E, Miuma S, Taura N and Nakao K: Serum exosomal
microRNA-122 and microRNA-21 as predictive biomarkers in
transarterial chemoembolization-treated hepatocellular carcinoma
patients. Oncol Lett. 16:3267–3273. 2018.PubMed/NCBI
|
|
160
|
Wang SC, Li CY, Chang WT, Cheng WC, Yen
CH, Tu WY, Lin ZY, Lin CC, Yeh ML, Huang CF, et al: Exosome-derived
differentiation antagonizing non-protein coding RNA with risk of
hepatitis C virus-related hepatocellular carcinoma recurrence.
Liver Int. 41:956–968. 2021. View Article : Google Scholar
|
|
161
|
Qin L, Zhan Z, Wei C, Li X, Zhang T and Li
J: Has-circRNA-G004213 promotes cisplatin sensitivity by regulating
miR-513b-5p/PRPF39 in liver cancer. Mol Med Rep. 23:4212021.
View Article : Google Scholar
|
|
162
|
Ghosh S, Bhowmik S, Majumdar S, Goswami A,
Chakraborty J, Gupta S, Aggarwal S, Ray S, Chatterjee R,
Bhattacharyya S, et al: The exosome encapsulated microRNAs as
circulating diagnostic marker for hepatocellular carcinoma with low
alpha-fetoprotein. Int J Cancer. 147:2934–2947. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Sun XH, Wang YT, Li GF, Zhang N and Fan L:
Serum-derived three-circRNA signature as a diagnostic biomarker for
hepatocellular carcinoma. Cancer Cell Int. 20:2262020. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Kim SS, Baek GO, Son JA, Ahn HR, Yoon MK,
Cho HJ, Yoon JH, Nam SW, Cheong JY and Eun JW: Early detection of
hepatocellular carcinoma via liquid biopsy: Panel of small
extracellular vesicle-derived long noncoding RNAs identified as
markers. Mol Oncol. 15:2715–2731. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Huang C, Tang S, Shen D, Li X, Liang L,
Ding Y and Xu B: Circulating plasma exosomal miRNA profiles serve
as potential metastasis-related biomarkers for hepatocellular
carcinoma. Oncol Lett. 21:1682021. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Itami-Matsumoto S, Hayakawa M,
Uchida-Kobayashi S, Enomoto M, Tamori A, Mizuno K, Toyoda H, Tamura
T, Akutsu T, Ochiya T, et al: Circulating exosomal miRNA profiles
predict the occurrence and recurrence of hepatocellular carcinoma
in patients with direct-acting antiviral-induced sustained viral
response. Biomedicines. 7:872019. View Article : Google Scholar
|
|
167
|
Sorop A, Iacob R, Iacob S, Constantinescu
D, Chitoiu L, Fertig TE, Dinischiotu A, Chivu-Economescu M,
Bacalbasa N, Savu L, et al: Plasma small extracellular vesicles
derived miR-21-5p and miR-92a-3p as potential biomarkers for
hepatocellular carcinoma screening. Front Genet. 11:7122020.
View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Xu LX, He MH, Dai ZH, Yu J, Wang JG, Li
XC, Jiang BB, Ke ZF, Su TH, Peng ZW, et al: Genomic and
transcriptional heterogeneity of multifocal hepatocellular
carcinoma. Ann Oncol. 30:990–997. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Huang A, Yang XR, Chung WY, Dennison AR
and Zhou J: Targeted therapy for hepatocellular carcinoma. Signal
Transduct Target Ther. 5:1462020. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Yang Y, Hong Y, Cho E, Kim GB and Kim IS:
Extracellular vesicles as a platform for membrane-associated
therapeutic protein delivery. J Extracell Vesicles. 7:14401312018.
View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Ha D, Yang N and Nadithe V: Exosomes as
therapeutic drug carriers and delivery vehicles across biological
membranes: Current perspectives and future challenges. Acta Pharm
Sin B. 6:287–296. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Ding J, Wang J and Chen J: Exosomes as
therapeutic vehicles in liver diseases. Ann Transl Med. 9:7352021.
View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Tai YL, Chen KC, Hsieh JT and Shen TL:
Exosomes in cancer development and clinical applications. Cancer
Sci. 109:2364–2374. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Lowry MC, Gallagher WM and O'Driscoll L:
The role of exosomes in breast cancer. Clin Chem. 61:1457–1465.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Xue D, Han J, Liu Y, Tuo H and Peng Y:
Current perspectives on exosomes in the diagnosis and treatment of
hepatocellular carcinoma (review). Cancer Biol Ther. 22:279–290.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Khan N, Maurya S, Bammidi S and
Jayandharan GR: AAV6 vexosomes mediate robust suicide gene delivery
in a murine model of hepatocellular carcinoma. Mol Ther Methods
Clin Dev. 17:497–504. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Du J, Wan Z, Wang C, Lu F, Wei M, Wang D
and Hao Q: Designer exosomes for targeted and efficient ferroptosis
induction in cancer via chemo-photodynamic therapy. Theranostics.
11:8185–8196. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Li I and Nabet BY: Exosomes in the tumor
microenvironment as mediators of cancer therapy resistance. Mol
Cancer. 18:322019. View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Le Grazie M, Biagini MR, Tarocchi M,
Polvani S and Galli A: Chemotherapy for hepatocellular carcinoma:
The present and the future. World J Hepatol. 9:907–920. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Shi S, Rao Q, Zhang C, Zhang X, Qin Y and
Niu Z: Dendritic cells pulsed with exosomes in combination with
PD-1 antibody increase the efficacy of sorafenib in hepatocellular
carcinoma model. Transl Oncol. 11:250–258. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Loch-Neckel G, Matos AT, Vaz AR and Brites
D: Challenges in the development of drug delivery systems based on
small extracellular vesicles for therapy of brain diseases. Front
Pharmacol. 13:8397902022. View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Pospichalova V, Svoboda J, Dave Z,
Kotrbova A, Kaiser K, Klemova D, Ilkovics L, Hampl A, Crha I,
Jandakova E, et al: Simplified protocol for flow cytometry analysis
of fluorescently labeled exosomes and microvesicles using dedicated
flow cytometer. J Extracell Vesicles. 4:255302015. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Zarovni N, Corrado A, Guazzi P, Zocco D,
Lari E, Radano G, Muhhina J, Fondelli C, Gavrilova J and Chiesi A:
Integrated isolation and quantitative analysis of exosome shuttled
proteins and nucleic acids using immunocapture approaches. Methods.
87:46–58. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Bala S, Csak T, Momen-Heravi F, Lippai D,
Kodys K, Catalano D, Satishchandran A, Ambros V and Szabo G:
Biodistribution and function of extracellular miRNA-155 in mice.
Sci Rep. 5:107212015. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Zhou H, Yuen PS, Pisitkun T, Gonzales PA,
Yasuda H, Dear JW, Gross P, Knepper MA and Star RA: Collection,
storage, preservation, and normalization of human urinary exosomes
for biomarker discovery. Kidney Int. 69:1471–1476. 2006. View Article : Google Scholar : PubMed/NCBI
|