Introduction
An imbalance between cell proliferation and death is
the primary cause of cancer. The absence of apoptotic signals
inhibits cell death, leading to uncontrolled cell proliferation and
cancer (1,2). Cancer cells often express several
proteins that, when abnormally elevated, render tumor cells
resistant to apoptosis (3,4).
Stress-inducible heat shock protein 70 (Hsp70; also called Hsp72),
a member of the human Hsp70 family playing critical roles in cell
survival under stressful conditions, is often overexpressed in
cancer cells; the selective survival advantage that it confers may
contribute to tumor formation (5–7).
Interestingly, Hsp70 expression in certain cancer types has been
correlated with poor prognosis and resistance to chemotherapy
(8–10). Thus, reducing the levels of Hsp70
in cancer cells may be an effective means to prevent tumor
progression. Hsp70 has been targeted with pharmaceuticals, such as
triptolide, quercetin, and KNK437, which downregulate its
expression (11–13). Hesperetin, a flavanone found in
citrus fruits such as oranges and grapefruits, has various
biological and pharmacological activities, including
anti-inflammatory, antioxidant, antihypertension, and
lipid-lowering effects (14,15). Hesperetin has been extensively
studied for its anticancer properties associated with the
inhibition of cell proliferation, cell cycle progression, and
angiogenesis and the activation of apoptosis (16–18). Accumulated data have shown a
marked ability of hesperetin to induce apoptosis in cancer cells
through multiple target molecules and associated signaling
pathways, such as ASK1/JNK, p38/MAPK, Notch1, ROS, Bcl-2 family
members, and death receptors (19–24). The transduction of these
hesperetin-induced apoptotic signals eventually results in
mitochondrial outer membrane permeability, which is a key step in
intrinsic apoptosis.
The intrinsic mitochondria-mediated apoptosis
pathway is regulated by Bcl-2 family members, including
proapoptotic (Bax, Bak, Bad, Bid and Bim) and antiapoptotic (Bcl-2
and Bcl-XL) molecules. Although several lines of
evidence have indicated that hesperetin modulates the levels of Bax
and Bcl-2 in human cancer cell lines, leading to mitochondrial
membrane disruption and apoptosis (20,22,23), it is not clear how hesperetin
activates Bax and increases the Bax/Bcl-2 ratio. Targeting Hsp70,
which is critical regulator of Bcl-2 family members, and thereby
disposing cancer cells toward apoptosis have been described
previously in the literature. Previous studies have confirmed not
only that Hsp70 is cytoprotective, but also that it interferes
effectively with cell death induced by a wide variety of stimuli,
including several cancer-related stresses. In addition to the
function of Hsp70 as a potent inhibitor of the stress-activated
kinase pathway that apparently blocks apoptotic signals via
interactions with JNK, Ask1, and SEK1 (25–27), it has been well-documented that
Hsp70 is also a negative regulator of the mitochondrial pathway of
apoptosis. Studies on the antiapoptotic function of Hsp70 have
focused on events that occur after mitochondrial disruption. Hsp70
prevents the recruitment of procaspase-9 to the apoptosome and its
functional complex formation by direct interaction with apoptotic
protease-activating factor 1 (28,29). Conversely, several studies have
reported that Hsp70 can prevent apoptosis upstream of the
mitochondria by inhibiting events that ultimately permeabilize the
mitochondrial outer membrane, such as the activation of Bax. Hsp70
directly binds to Bax and prevents its translocation to the outer
mitochondrial membrane where it promotes cell death (30,31). Therefore, the hesperetin-induced
apoptosis via Bax prompted the investigation of whether or not
Hsp70 is a target in hesperetin-induced apoptosis in cancer
cells.
Materials and methods
Materials
Mouse monoclonal antibody against Hsp70 (cat. no.
SMC-100) was purchased from Stress Marq Biosciences, Inc. Goat
polyclonal antibody, anti-voltage-dependent anion channel (VDAC)
(product code ab37985), and rabbit polyclonal antibody
anti-STUB1/CHIP (product code ab134064) were obtained from Abcam.
Rabbit anti-Bax (product no. 5023), anti-active caspase-9 (product
no. 9502), anti-ubiquitin (product no. 3936), anti-MEK1/2 (product
no. 8727), and anti-poly (ADP-ribose) polymerase (PARP;
active-PARP) (product no. 9542) antibodies were purchased from Cell
Signaling Technology, Inc. The mouse monoclonal antibody against
β-actin (cat. no. 017-24551) and MG132 (cat. no. 139-18451) were
obtained from FUJIFILM Wako Pure Chemical Corporation. Hesperetin
(cat. no. H4125) and all other chemicals used in this study were
purchased from Sigma-Aldrich; Merck KGaA.
Cell culture and viability assay
A549 lung cancer cell line (JCRB no. JCRB0076) was
obtained from NIBIOHN. H358 lung cancer cell line (ATCC no.
CRL-5807) was from the American Type Culture Collection. Both
cancer cell lines were cultured in Dulbecco's modified Eagle's
medium containing 10% fetal bovine serum at 37°C. Hesperetin was
dissolved in dimethyl sulfoxide. Briefly A549 or H358 cells were
seeded at 5×105 cells/well in 12-well plates for 24 h at
37°C. A549 cells were treated with various concentrations of
hesperetin (0, 100, 200, 300, 400, 500, 600, 700, 800, 900, and
1,000 µM) for 48 h at 37°C. In addition, H358 cells were treated
with various concentrations of hesperetin (0, 200, 400 and 600 µM)
for 48 h at 37°C. Mitochondrial dehydrogenase
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetra-zolium bromide (MTT)
assay was used to assess cell death/survival. The cells were
incubated for 2 h with MTT solution at 37°C and then 50%
iso-propanol/10% SDS was added to dissolve the formazan. The
reaction product was measured by spectrophotometry at a wavelength
of 570 nm, and the relative viability of cells treated with
hesperetin vs. untreated cells was calculated. Cell viability was
used to calculate the half maximal inhibitory concentration
(IC50). In addition, to study the molecular mechanism of
Hsp70, A549 cells were pretreated with the proteasome inhibitor
MG132. MG132 (3 µM) was added to A549 cells for 2 h at 37°C, before
exposure to hesperetin.
Colony formation assay
A549 cells were plated in a 12-well plate at a
density of 1,000 cells/well for 24 h before treatment with various
concentrations of hesperetin (0, 200, 400, and 600 µM) for 48 h at
37°C. Following treatment, cells were incubated in fresh media for
7 days. Colonies were fixed with 4% paraformaldehyde for 20 min at
room temperature and visualized using 0.05% crystal violet solution
for 20 min at room temperature. A colony was defined as an
agglomeration of >50 cells. After washing, the ColonyArea
(version Dec 22, 2016) (https://b2share.eudat.eu/records/39fa39965b314f658e4a198a78d7f6b5),
a plugin for ImageJ software (version 1.53s; National Institutes of
Health), was used for quantification.
Cell cycle analysis
A549 cells were cultured in a 60-mm dish at a
density of 2×105 cells/dish. The cells treated with
various concentrations of hesperetin (0, 200, 400, and 600 µM) for
48 h at 37°C were fixed with 70% ethanol at −20°C for 2 h and
stained with a propidium iodide (PI) concentration of 0.05 mg/ml
for 30 min at 37°C. Fluorescence was measured using FACSCalibur (BD
Biosciences), and data were analyzed using CellQuest software
(version 3.1; BD Biosciences).
Quantification of apoptosis by flow
cytometry
A549 cells were seeded at 5×105
cells/dish in a 60-mm dish. The cells were washed with Annexin V
staining buffer [10 mM HEPES (pH 7.4), 150 mM NaCl, 5 mM KCl, 1 mM
MgCl2, and 1.8 mM CaCl2] and incubated with CF488A-Annexin V and PI
(cat. no. 15342-54; Nacalai Tesque, Inc.) in a staining buffer for
30 min at 37°C in the dark. Fluorescence was measured using
FACSCalibur, and data were analyzed using CellQuest software
(version 3.1).
Immunoblotting and cell
fractionation
A549 cells were cultured in a 60-mm dish at a
density of 2×105 cells/dish. The cells were lysed using
immunoprecipitation assay buffer [50 mM Tris-HCl (pH 7.5), 150 mM
NaCl, 1 mM EDTA, and 0.1% NP-40] containing Protease Inhibitor
Cocktail Set (Nacalai Tesque, Inc.). The cell lysates were resolved
in Laemmli sample buffer. The concentration of total protein was
measured using a BCA assay. The samples (20 µg protein/lane) were
subjected to 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE), transferred onto a polyvinylidene
difluoride membrane. The membranes were blocked with 10% skim milk
in TBS-0.1% Tween-20 buffer for 2 h at room temperature, and then
reacted with the respective antibodies: Hsp70 (1:1,000), VDAC
(1:500), CHIP (1:5,000), Bax (1:1,000), caspase-9 (1:1,000),
ubiquitin (1:1,000), MEK1/2 (1:1,000), PARP (1:1,000) and β-actin
(1:5,000) overnight at 4°C. The membranes were treated with the
secondary antibodies conjugated with horseradish peroxidase (HRP)
(1:4,000; cat. nos. HAF007, HAF109, and HAF008; R&D systems)
for 1 h at room temperature and detected with an ECL
chemiluminescence detection kit (GE Healthcare; Cytiva). A549 cells
(2×105 cells/dish in 100 mm dish) lysed in 20 mM
HEPES-KOH (pH 7.5) buffer containing 0.25 M sucrose were
homogenized in a Dounce homogenizer and centrifuged at 1,000 × g
for 30 min at 4°C to separate nuclei and unbroken cells. The
supernatants were centrifuged at 10,000 × g for 15 min at 4°C, and
the pellets were collected as the heavy membrane/mitochondrial
fraction. The cytosolic fractions were isolated according to the
manufacturer's protocol, using the Mitochondria Isolation Kit for
Cultured Cells (cat. no. 89874) from Thermo Fisher Scientific,
Inc.
Immunoprecipitation
For the immunoprecipitation, the cell lysates were
incubated with 5 µg Hsp70 antibody (cat. no. SPC-103; Stress Marq
Bioscience, Inc.) overnight at 4°C. Subsequently, 20 µl of Protein
G-sepharose beads (cat. no. P3296; Sigma-Aldrich; Merck KGaA) were
added to collect the immunocomplexes for an additional 1 h of
incubation at 4°C. The pellets were washed three times with assay
buffer by centrifugation at 1,000 × g for 5 min at 4°C. The
supernatant was discarded, and the beads were resuspended with 40
µl of electrophoresis buffer and analyzed by SDS-PAGE and
immunoblotting using an anti-ubiquitin (cat. no. 3936; Cell
Signaling technology, Inc.) or anti-Hsp70 (cat. no. SMC-100; Stress
Marq Bioscience, Inc.) antibody.
Overexpression of Hsp70
A549 cells (2×105 cells/dish in a 60-mm
dish) were seeded into 12-well plates for 24 h before transfection.
The expression plasmid of pCMV-Tag3B myc-Hsp70 was supplied by Dr
Tohru Ichimura (Laboratory of Cell Molecular Biology, Department of
Applied Chemistry, National Defense Academy, Yokosuka, Japan). The
negative control plasmid, pCMV (an empty vector), was supplied from
RIKEN BioResource Center. Transfection was performed with 8 µg
vector using Lipofectamine 3000 (Thermo Fisher Scientific, Inc.) in
accordance with the manufacturer's instructions. The cells were
incubated for 12 h at 37°C, and subsequently treated with
hesperetin for 48 h.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA from cells was extracted using QIAshredder
and RNeasy Mini Kit (Qiagen GmbH) according to manufacturer's
protocol. The cDNA synthesis was performed by High-Capacity
RNA-to-cDNA™ Kit (Thermo Fisher Scientific Inc.) and
qPCR was subsequently performed on QuantStudio® 5
Real-Time PCR (Thermo Fisher Scientific Inc.) system using
PowerUp™ SYBR™ Green Master Mix (Thermo
Fisher Scientific, Inc.). The reaction was carried out according to
the following amplification protocol: 95°C for 2 min, 40 cycles at
95°C for 15 sec, 60°C for 1 min and 95°C for 15 sec. Gene
expression was analyzed according to the 2−ΔΔCq method
(32). The following sequences of
the primers were used, respectively: Hsp70 forward primer (F),
5′-GCCGAGAAGGACGAGTTTGA-3′ and reverse primer (R),
5′-TCCGCTGATGATGGGGTTAC-3′; β-actin (F),
5′-ATGTGGCCGAGGACTTTGATT-3′ and (R),
5′-AGTGGGGTGGCTTTTAGGATG-3′.
Knockdown of CHIP
The specific small interfering RNA (siRNA) for CHIP
was obtained from Ajinomoto Bio-Pharma: siCHIP sequence of the
sense strand (5′-AGGCCAAGCACGACAAGUAdTdT-3′). The negative control
siRNA (MISSION siRNA Universal Negative Control #1; cat. no.
SIC001) was purchased from Merck KGaA. A549 cells were transfected
with siRNA (10 nmol/l) using Lipofectamine RNAi MAX (Thermo Fisher
Scientific). The cells were then incubated for 24 h at 37°C, and
subsequently treated with 600 µM of hesperetin and grown for 48 h
at 37°C to allow an effective decrease in the expression of the
respective target molecules. For the MTT assay, A549 cells were
treated with various concentrations of hesperetin (0, 200, 400 and
600 µM) for 48 h at 37°C.
JC-1 staining
Mitochondrial permeability transition was determined
by staining the cells with
5,5′,6,6′-tetrachloro-1,1,3,3′-tetraethyl-benzimidazolyl-carbocyanin
iodide (JC-1; Molecular Probes; Invitrogen; Thermo Fisher
Scientific, Inc.) for 2 h at 37°C in the dark. The cells were
subsequently washed with assay buffer in accordance with the
manufacturer's protocol and immediately imaged using a fluorescence
microscope (Keyence Corporation) with the red (λexcitation: 560±40
nm band pass filter, λdetection: 630±60 nm band pass filter) and
green (λexcitation: 470±40 nm band pass filter, λdetection: 535±50
nm band pass filter) fluorescence channels.
Statistical analysis
The data were expressed as the mean ± standard
deviation (SD) for three independent experiments. Comparisons
between quantitative variables were assessed using one-way ANOVA
followed Tukey's post hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
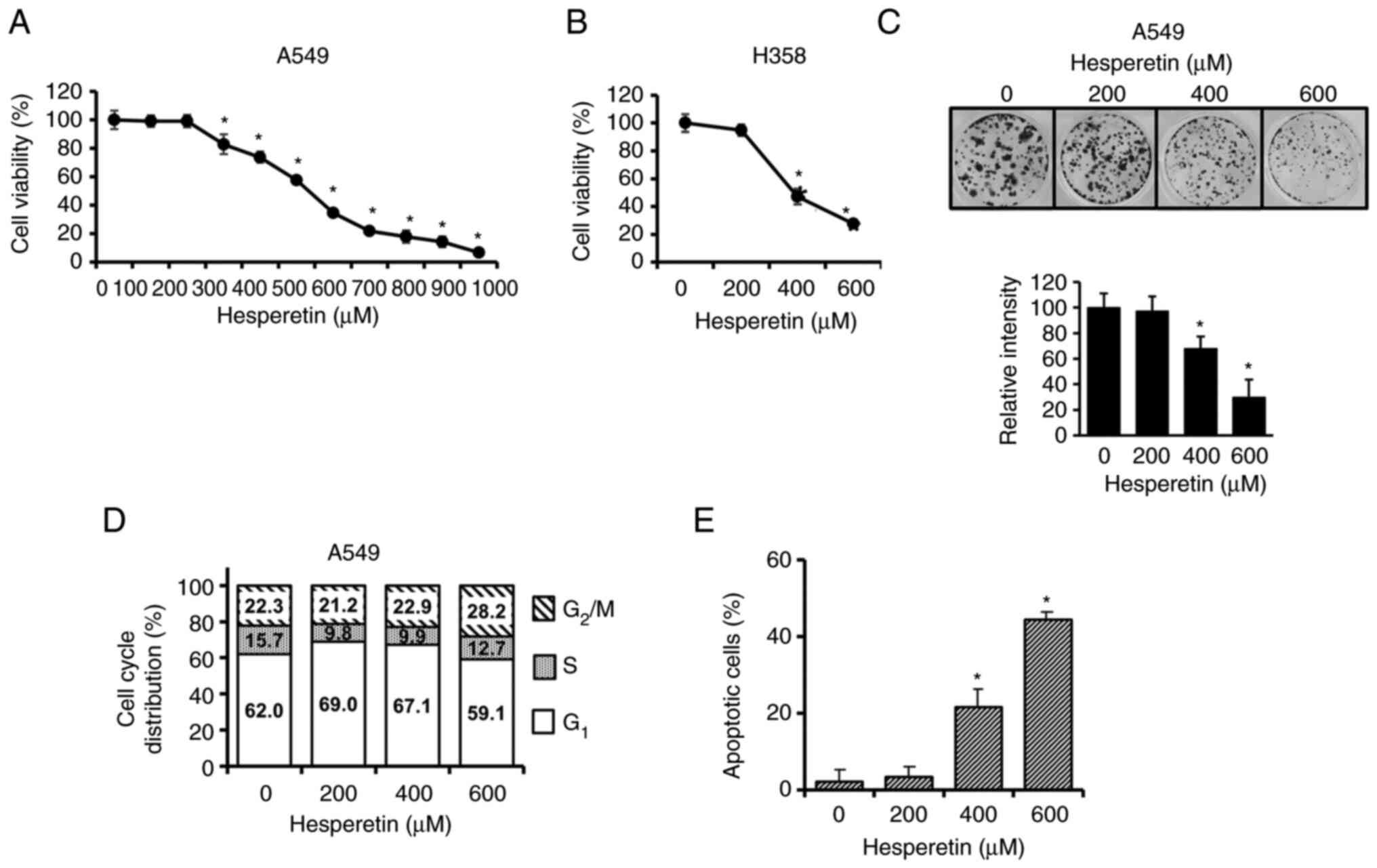
Hesperetin inhibits proliferation and
induces apoptosis in lung cancer cell lines
The survival of lung cancer cells treated with
hesperetin was assessed using an MTT assay. Following treatment
with hesperetin, the viability of A549 (Fig. 1A) and H358 (Fig. 1B) lung cancer cell lines decreased
in a concentration-dependent manner. The IC50 value of
the potential cytotoxic effects of hesperetin against A549 was 520
µM (Fig. 1A). To further examine
the effect of hesperetin on A549 cell proliferation, a colony
formation assay was performed. Hesperetin markedly reduced the
colonigenic potential of A549 cells in a dose-dependent manner
(Fig. 1C). The colony formation
measured by percent area of the colonies with hesperetin treatments
of 400 and 600 µM were 68 and 30%, respectively, when that of the
control without hesperetin was set at 100%. These results were not
significantly different from those of the MTT assay. Cell cycle
analysis was performed with a flow cytometer to examine the
inhibitory effect of hesperetin on lung cancer cells. As shown in
Fig. 1D, the cell cycle
progression of A549 cells revealed no significant change after
treatment with various concentrations of hesperetin. Considering
that hesperetin did not affect cell cycle progression, it was then
investigated whether or not the inhibitory effects of hesperetin on
A549 cell viability are induced via apoptotic cell death. Apoptotic
cell death was assessed using CF488A-Annexin V through flow
cytometry. The increase in the number of apoptotic cells in
hesperetin-treated cultures indicated that hesperetin decreased
cell viability through apoptosis (Fig. 1E).
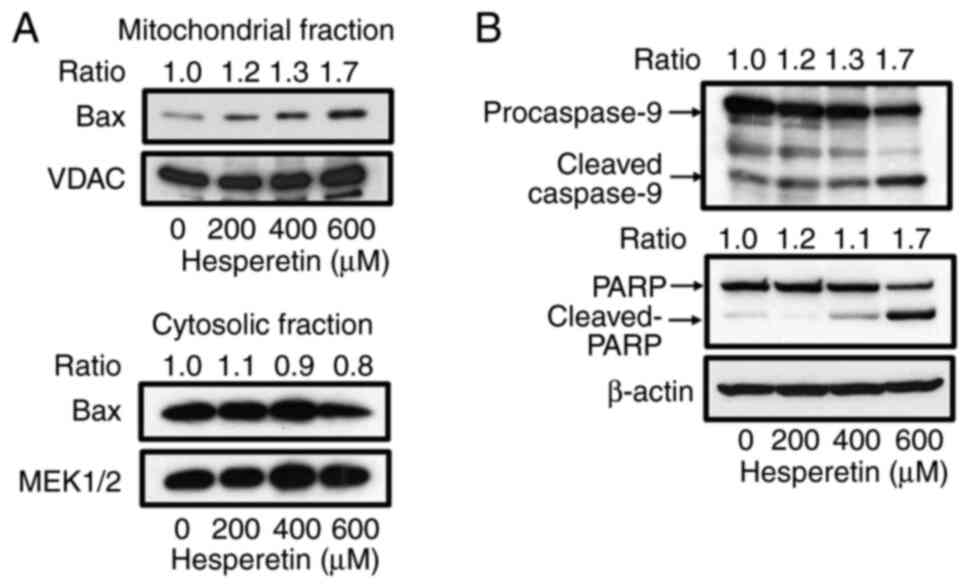
Hesperetin induces
mitochondria-dependent apoptosis associated with the activation of
Bax in A549 cells
Several studies have shown that mitochondria and
pro-apoptotic molecule, Bax, may be a direct and important target
of hesperetin in sensitive cells (19,20,22,23,33). It was examined whether or not
hesperetin promotes mitochondria-dependent apoptosis by inducing
Bax translocation in A549 cells. Mitochondrial fractions were
prepared from A549 cells incubated with or without hesperetin and
immunoblotted for Bax in these fractions. In the presence of
hesperetin, Bax was localized to the mitochondria in a
dose-dependent manner, whereas hesperetin slightly reduced the
amount of cytosolic Bax (Fig.
2A). Apoptotic signal transmission to the mitochondria leads to
the activation of caspase-9, which then converts procaspase-3 into
active caspase-3, resulting in PARP cleavage and apoptosis
(34,35). The activation of caspase-9 and
cleavage of PARP in A549 cells treated with hesperetin was assessed
to examine whether or not hesperetin influences downstream
mitochondrial-related apoptotic events. As shown in Fig. 2B, hesperetin induced the cleavage
of caspase-9 in a concentration-dependent manner in A549 cells.
Hesperetin also caused an increase in the level of cleaved PARP
(Fig. 2B).
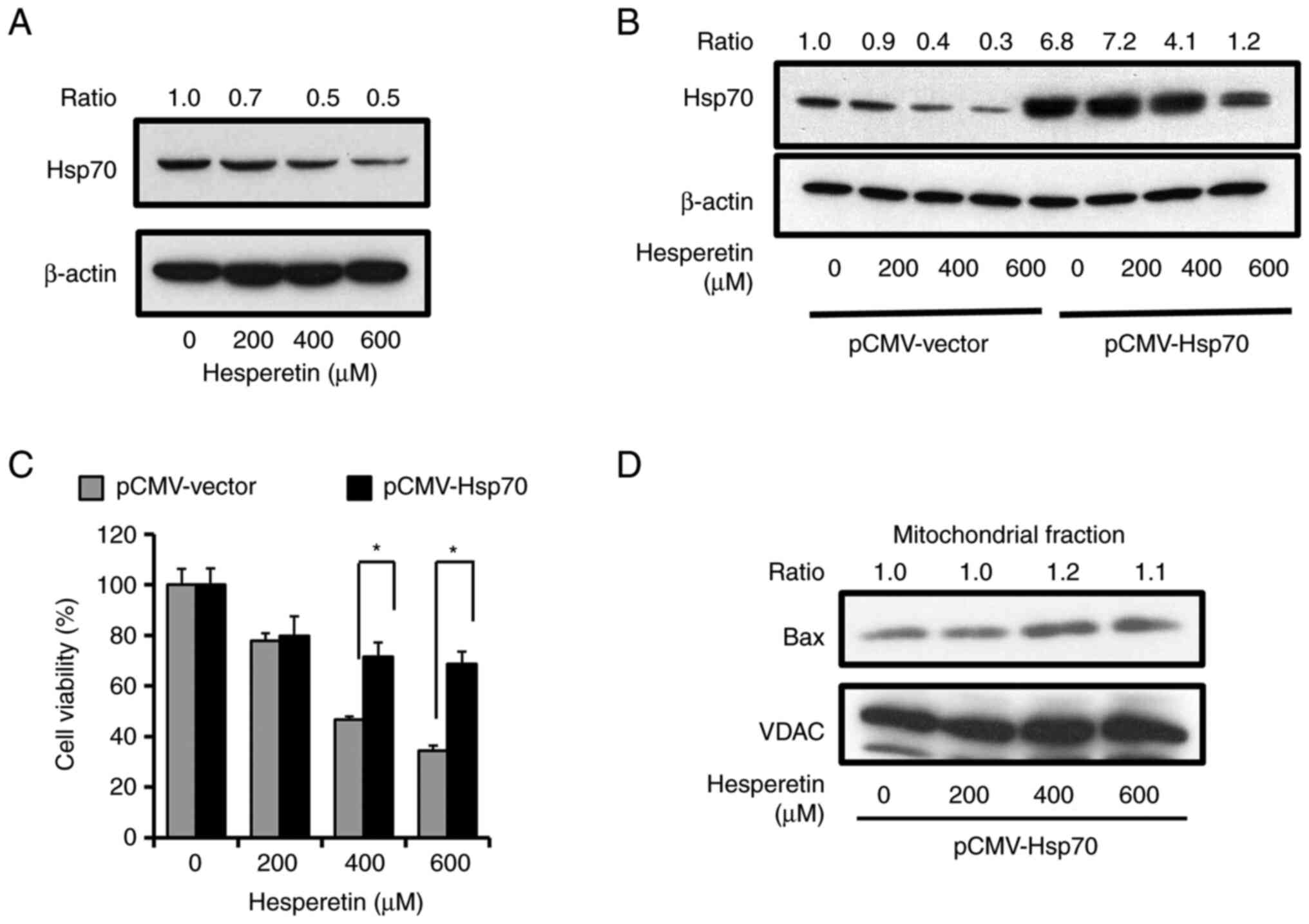
Hesperetin induces apoptosis by
suppressing Hsp70 in A549 cells
Hsp70 was then investigated, which generally
interacts with Bax to prevent its translocation to the mitochondria
in living cells and inhibits cell death, to clarify the molecular
mechanism that underlies the activation of Bax in
hesperetin-induced apoptosis (30,31). The expression of Hsp70 after the
exposure of A549 cells to various concentrations of hesperetin was
analyzed by immunoblotting to examine the effects of hesperetin on
Hsp70. Hesperetin significantly decreased the expression level of
Hsp70 in A549 cells (Fig. 3A).
The expression of Hsp70 was overexpressed as a myc-tagged protein
in A549 cells (Fig. 3B) and its
effect on the decrease of cell viability induced by hesperetin was
assessed to ascertain the effects of Hsp70 expression on cell death
while excluding all the effects of hesperetin unrelated to Hsp70.
As revealed in Fig. 3C, Hsp70
overexpression rescued A549 cells from hesperetin-induced apoptosis
compared to cells transfected with the empty vector. It was next
examined whether or not the forced expression of Hsp70 prevents the
translocation of Bax to mitochondria caused by hesperetin. In A549
cells with exogenously overexpressed Hsp70, Bax localization to the
mitochondria in the presence of increasing doses of hesperetin
significantly decreased as compared to that in the cells with
endogenous expression of Hsp70 (Figs.
2A and 3D). These
observations indicated that hesperetin promoted the apoptotic
signal at the Bax activation step of the mitochondrial apoptotic
pathway via Hsp70 suppression, potentially contributing to the
ability of hesperetin to inhibit A549 cell proliferation.
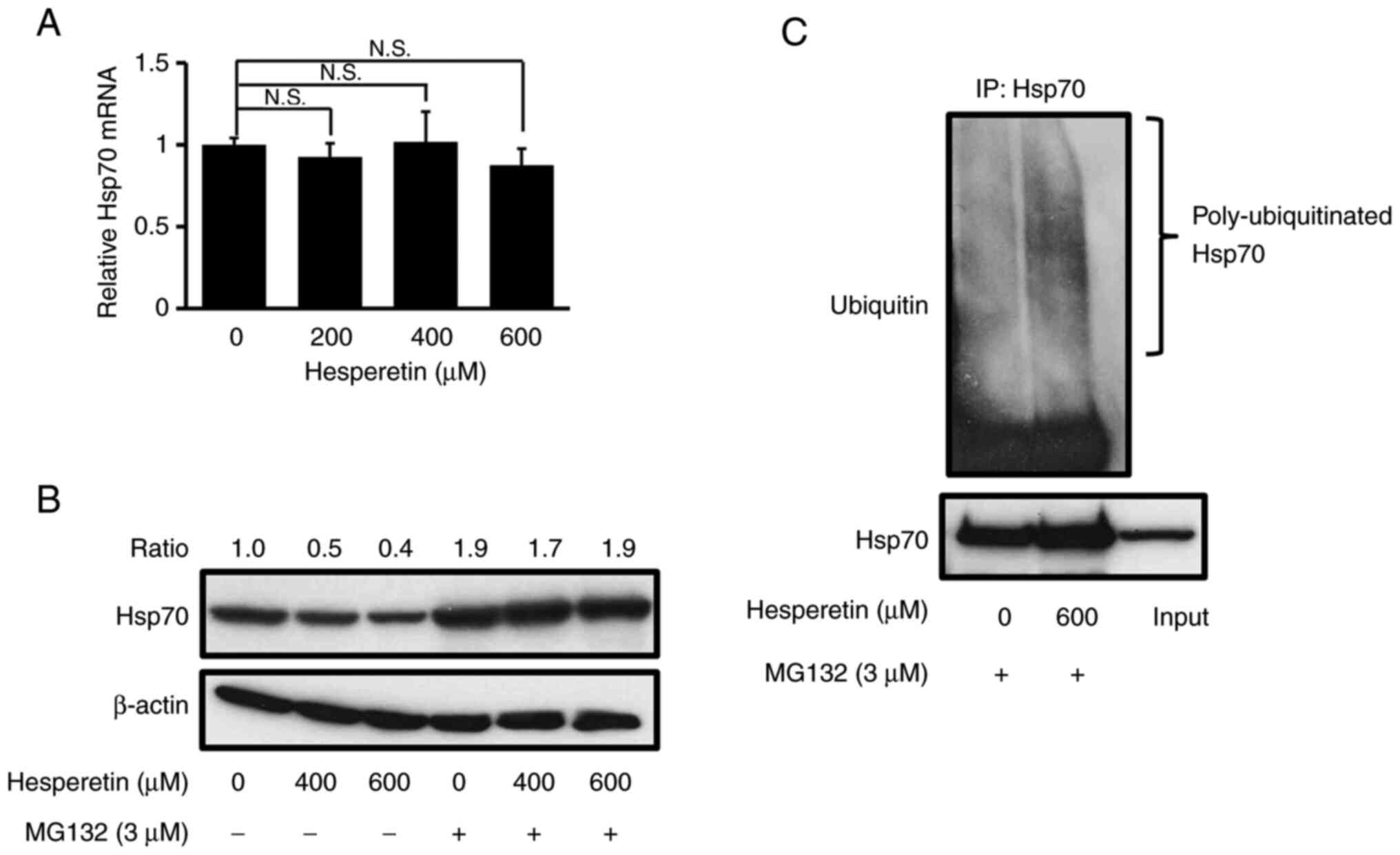
Hesperetin decreases Hsp70 expression
by inducing a proteasome-mediated degradation
The mechanisms by which hesperetin decreases Hsp70
expression are poorly known. The expression of Hsp70 was examined
at the mRNA level in A549 cells by RT-qPCR after treatment with
hesperetin (Fig. 4A). The absence
of a decrease in the mRNA level of Hsp70 following hesperetin
treatment suggests that hesperetin causes its degradation in A549
cells. It is noteworthy that Hsp70 can often serve as a proteasome
substrate and that it is stabilized by inhibition of the proteasome
(36,37). To study the machinery that turns
over Hsp70, A549 cells were pretreated with the proteasome
inhibitor MG132 before exposure to hesperetin. The total amount of
Hsp70 was nearly the same in the absence of hesperetin, regardless
of cell exposure to MG132. MG132 prominently blocked the
hesperetin-dependent decrease in Hsp70 expression in A549 cells
(Fig. 4B). These observations
indicated that hesperetin induced Hsp70 degradation by the
proteasome. Based these observations, the ubiquitination of Hsp70
was further assessed. In the presence of MG132, hesperetin
substantially increased the level of Hsp70 ubiquitination compared
with control cells (Fig. 4C).
Overall, these data clearly indicated that hesperetin promoted the
proteasome-mediated degradation of Hsp70 in A549 cells.
Hesperetin induces the
proteasome-mediated degradation of Hsp70 by increasing CHIP,
leading to A549 cell apoptosis
CHIP has been demonstrated, in vitro and
in vivo, to be associated with Hsp70 and ubiquitinate it,
targeting it to the proteasome for degradation (36,37). The effect of hesperetin on CHIP in
A549 cells was first investigated to clarify the mechanisms of CHIP
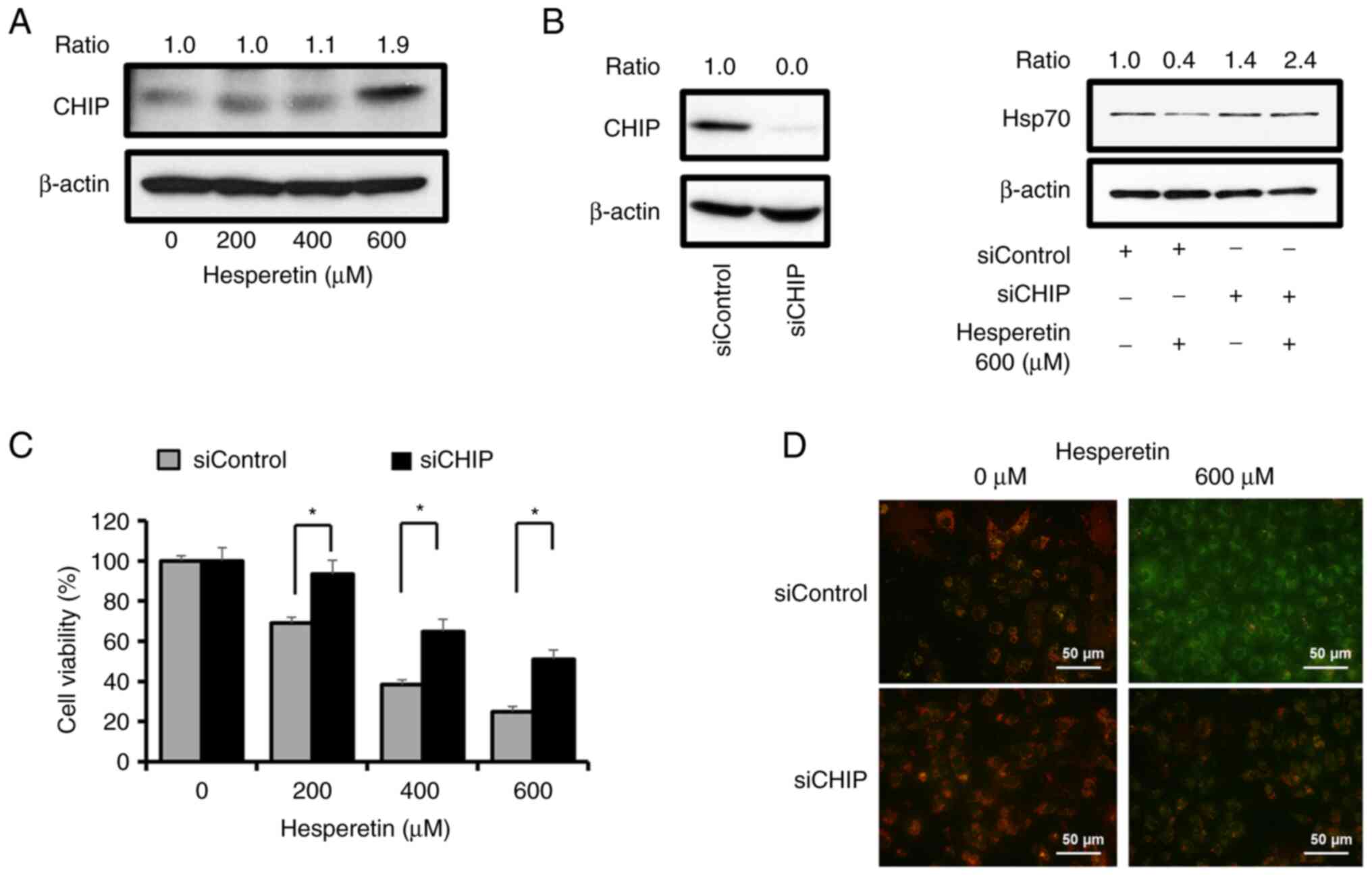
responsible for the degradation of Hsp70. Interestingly, the level
of CHIP markedly increased after hesperetin treatment in a
concentration-dependent manner (Fig.
5A). Blockage of Hsp70 inhibition by the treatment of A549
cells with MG132 and the upregulation of CHIP in the presence of
hesperetin suggested that Hsp70 was targeted by hesperetin for
degradation by the proteasome. To further determine the functional
importance of CHIP in the proteasome-mediated degradation of Hsp70
by hesperetin, the amount of endogenous CHIP present in A549 cells
was attenuated with RNAi and its effects on Hsp70 expression were
studied. Treatment with CHIP dsRNA did not cause the decrease in
Hsp70 levels mediated by hesperetin compared with that measured in
cells treated with scrambled siRNA (Fig. 5B). It was then examined how a
selective knockdown of CHIP affects the survival of A549 cells
exposed to increasing concentrations of hesperetin. As revealed in
Fig. 5C, an MTT assay indicated
that the inhibitory effect of hesperetin on the viability of A549
cells significantly decreased after CHIP silencing. This finding
was consistent with the result of JC-1 staining used to determine
the mitochondria-dependent apoptosis of A549 cells after treatment
with hesperetin (Fig. 5D). These
results clearly demonstrated the critical role of CHIP in the
induction of hesperetin-mediated apoptosis in A549 cells.
Discussion
Epidemiological studies have indicated an inverse
association between the dietary consumption of flavonoids and the
risk of various illnesses, such as cardiovascular diseases,
metabolic diseases, and cancer (15,38,39). Recently, more attention has been
paid to the anticancer effects of hesperetin. Hesperetin was
demonstrated to inhibit carcinogenesis in preclinical experiments
using various cancer cells, including breast, lung, gastric, colon,
and hepatic cancer cells (19,22–24,33). The most attractive feature of
hesperetin is its broad application for a new promising class of
anticancer therapeutic agents from functional food with few side
effects, since it is toxic to cancer cells but not to normal cells
as compared with currently available cytotoxic drugs (40,41). One widely known anticancer effect
of hesperetin on cancer cells is the activation of the apoptosis
pathway. Although the induction of apoptosis by hesperetin is well
established, the detailed mechanisms and pathways by which this
phenomenon occurs remain unknown. To link this hesperetin-dependent
event mechanistically, the action of Hsp70 in apoptosis induced by
hesperetin was explored. Hesperetin significantly decreased Hsp70
expression in lung adenocarcinoma cell lines and induced the
apoptosis of A549 cells. This finding is fundamentally consistent
with previous studies, which revealed that functionally related
small molecules that inhibit Hsp70 decrease the viability of
colorectal or pancreatic cancer cells by promoting apoptosis via
the downregulation of Hsp70 (6,12,13). To the best of our knowledge, the
present study is the first to demonstrate the involvement of Hsp70
in hesperetin-induced cell death. This observation is important
because targeting the expression or function of Hsp70 is a
potential treatment strategy for several types of cancer,
considering the hypothesis that a high level of Hsp70 protects
against cell death and increases the survival of cells exposed to
apoptotic stimuli (27,42). Previous research identified Hsp70
as a putative oncogene whose upregulation is common and driven by
its genomic amplification in cancer cells (5–7),
and increased Hsp70 expression is positively associated with the
advanced pathologic stage and grade of cancers and related to poor
outcomes of patients (8–10). These notions offer an attractive
opportunity for therapeutic approaches that inhibit Hsp70. The
inhibitory effects of hesperetin on Hsp70 may be supported by
previous research which demonstrated that the flavonoid quercetin
decreases Hsp70 expression in human cancer cell lines (11).
Several studies have indicated that activation of
the mitochondrial pathway is a critical event in apoptosis induced
by hesperetin (19,20,22,23,33), whereas another study reported that
hesperetin induces apoptosis in cancer cells via a
mitochondria-independent pathway (24). This discrepancy may be due to the
different experimental systems used to evaluate apoptosis or
reflects the variability of apoptotic pathways among different cell
lines. In the present study, hesperetin facilitated the
translocation of proapoptotic Bax to the mitochondria, and the
events occurred up- and downstream of mitochondrial disruption in
A549 cells. The key regulators of apoptosis, including members of
the antiapoptotic and proapoptotic Bcl-2 family, affect the outer
membrane permeability of the mitochondria (43). In fact, reduced expression of
death-inducing members (Bax) vs. increased expression of
death-inhibitory members (Bcl-2, Bcl-xL) has been reported in
various cancer types and malignancies (44). Bax generally plays an important
role in inhibiting the progression of cancer and promoting the
apoptosis of some types of cancers. Interestingly, earlier studies
have shown that hesperetin increases Bax expression in cancer cells
(20,22,33), although the specific mechanisms
modulating the activation of Bax remain unknown. In the present
study, overexpression of Hsp70, which actively sequestrates Bax in
the cytosol to interfere with its translocation to the
mitochondria, rescued A549 cells from hesperetin. This result
suggests that Hsp70 acts as an inhibitory molecule for the
activation of Bax in hesperetin-mediated apoptosis. To clarify the
mechanisms underlying the hesperetin-induced downregulation of
Hsp70, it was further demonstrated that the degradation of Hsp70,
associated with the upregulation of CHIP in the ubiquitin
proteasome pathway, is a critical event in apoptosis induced by
hesperetin. Hsp70 becomes very stable and accumulates at very high
levels in cancer cells, which is a feature of the gain-of-function
in tumorigenesis and cancer progression (5–7).
In addition, Hsp70 is generally regulated mainly and maintained at
basal levels under normal physiological conditions (45,46). Hsp70 stability is regulated by
proteasome-mediated degradation through the E3 ubiquitin ligase
CHIP. Previous studies have indicated the importance of
CHIP-mediated Hsp70 turnover for the regulatory mechanism of Hsp70
(33,34). The lack of an inhibitory effect on
Hsp70 expression and an anti-proliferative effect by hesperetin in
A549 cells treated with CHIP dsRNA clearly indicate that CHIP is a
notable target for the induction of hesperetin-mediated apoptosis.
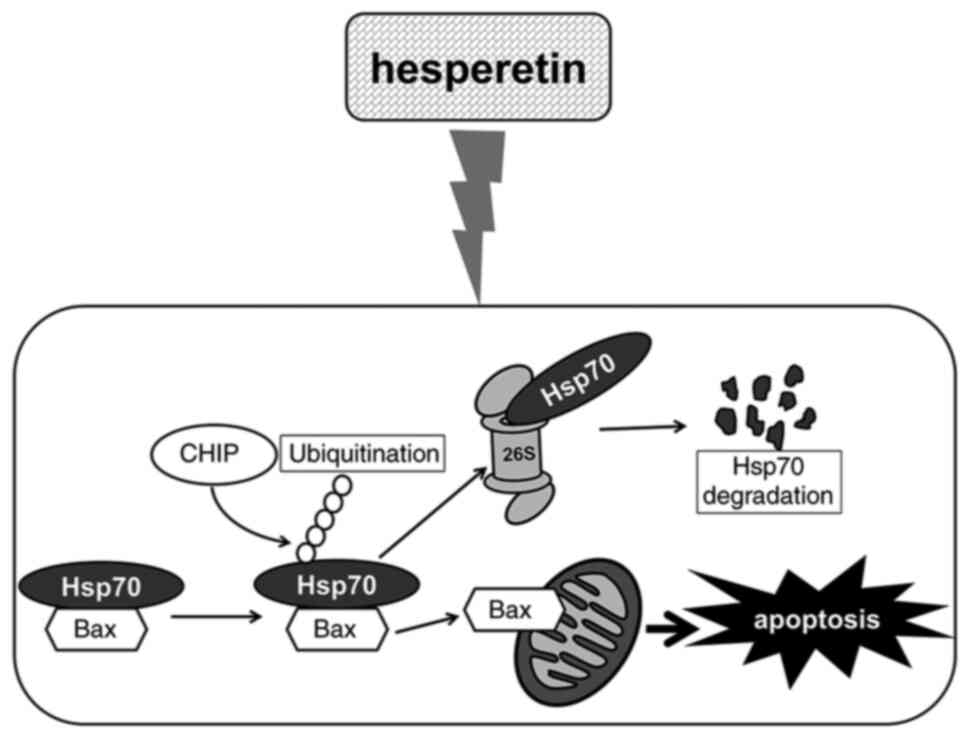
Based on these findings, a mechanism model for hesperetin-mediated
apoptosis is proposed, as summarized in Fig. 6. Collectively, the present study
provided the first evidence that Hsp70 is critical for determining
the cellular decision to either initiate or suppress apoptosis
mediated by hesperetin. These findings may contribute toward the
development of new and less toxic chemotherapies that use natural
compounds against cancer, and they also highlight the putative
merits of developing anticancer treatments targeting Hsp70.
Acknowledgements
We would like to thank Dr Tohru Ichimura (Laboratory
of Cell Molecular Biology, Department of Applied Chemistry,
National Defense Academy, Japan) for providing the myc-Hsp70
construct.
Funding
The present study was supported by the JSPS Grants-in Aid for
Scientific Research (grant nos. 19K059932 and 18K05536).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
MY and HE designed the research and experiments, and
supervised the experimental analysis. MT contributed to the design
and implementation of the research. MT and KS carried out the
experiments. MT performed the cell viability assay and prepared the
samples and characterized them by flow cytometry. KS performed
almost all of the other experiments. MT, HE, KS and MY analyzed the
data of this study. HE supervised MT and KS who investigated the
findings of this study. MY and HE confirm the authenticity of all
raw data. MY wrote the paper with input from all the authors. All
authors discussed the results. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nakano K and Vousden KH: PUMA, a novel
proapoptotic gene, is induced by p53. Mol Cell. 7:683–694. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Igney FH and Krammer PH: Death and
anti-death: Tumour resistance to apoptosis. Nat Rev Cancer.
2:277–288. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Endo H, Inoue I, Masunaka K, Tanaka M and
Yano M: Curcumin induces apoptosis in lung cancer cells by 14-3-3
protein-mediated activation of Bad. Biosci Biotechnol Biochem.
84:2440–2447. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jäättelä M: Escaping cell death: Survival
proteins in cancer. Exp cell Res. 248:30–43. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Aghdassi A, Phillips P, Dudeja V,
Dhaulakhandi D, Sharif R, Dawra R, Lerch MM and Saluja A: Heat
shock protein 70 increases tumorigenicity and inhibits apoptosis in
pancreatic adenocarcinoma. Cancer Res. 67:616–625. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Endo H, Yano M, Okumura Y and Kido H:
Ibuprofen enhances the anticancer activity of cisplatin in lung
cancer cells by inhibiting the heat shock protein 70. Cell Death
Dis. 5:e10272014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ciocca DR, Clark GM, Tandon AK, Fuqua SA,
Welch WJ and McGuire WL: Heat shock protein hsp70 in patients with
axillary lymph node-negative breast cancer: Prognostic
implications. J Natl Cancer Res. 85:570–574. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cornford PA, Dodson AR, Parsons KF,
Desmond AD, Woolfenden A, Fordham M, Neoptolemos JP, Ke Y and
Foster CS: Heat shock protein expression independently predicts
clinical outcome in prostate cancer. Cancer Res. 60:7099–7105.
2000.PubMed/NCBI
|
|
10
|
Vargas-Roig LM, Gago FE, Tello O, Aznar JC
and Ciocca DR: Heat shock protein expression and drug resistance in
breast cancer patients treated with induction chemotherapy. Int J
Cancer. 79:468–475. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hosokawa N, Hirayoshi K, Nakai A, Hosokawa
Y, Marui N, Yoshida M, Sakai T, Nishino H, Aoike A, Kawai K, et al:
Flavonoids inhibit the expression of heat shock proteins. Cell
Struct Funct. 15:393–401. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yokota S, Kitahara M and Nagata K:
Benzylidene lactam compound, KNK437, a novel inhibitor of
acquisition of thermotolerance and heat shock protein induction in
human colon carcinoma cells. Cancer Res. 60:2942–2948.
2000.PubMed/NCBI
|
|
13
|
Westerheide SD, Kawahara TL, Orton K and
Morimoto RI: Triptolide, an inhibitor of the human heat shock
response that enhances stress-induced cell death. J Biol Chem.
281:9616–9622. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Parhiz H, Roohbakhsh A, Soltani F, Rezaee
R and Iranshahi M: Antioxidant and anti-inflammatory properties of
the citrus flavonoids hesperidin and hesperetin: An updated review
of their molecular mechanisms and experimental models. Phytother
Res. 29:323–331. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Scoditti E: Neuroinflammation and
neurodegeneration: The promising protective role of the citrus
flavanone hesperetin. Nutrients. 12:23362020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Aranganathan S and Nalini N:
Antiproliferative efficacy of hesperetin (citrus flavanoid) in
1,2-dimethylhydrazine-induced colon cancer. Phytother Res.
27:999–1005. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Choi EJ: Hesperetin induced G1-phase cell
cycle arrest in human breast cancer MCF-7 cells: Involvement of
CDK4 and p21. Nutr Cancer. 59:115–119. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nalini N, Aranganathan S and Kabalimurthy
J: Chemopreventive efficacy of hesperetin (citrus flavonone)
against 1,2-dimethylhydrazine-induced rat colon carcinogenesis.
Toxicol Mech Methods. 22:397–408. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Palit S, Kar S, Sharma G and Das PK:
Hesperetin induces apoptosis in breast carcinoma by triggering
accumulation of ROS and activation of ASK1/JNK pathway. J Cell
Physiol. 230:1729–1739. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li Q, Miao Z, Wang R, Yang J and Zhang D:
Hesperetin induces apoptosis in human glioblastoma cells via p38
MAPK activation. Nutr Cancer. 72:538–545. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Patel PN, Yu XM, Jaskula-Sztul R and Chen
H: Hesperetin activates the Notch1 signaling cascade, causes
apoptosis, and induces cellular differentiation in anaplastic
thyroid cancer. Ann Surg Oncol. 21 (Suppl 4):S497–S504. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang J, Song J, Wu D, Wang J and Dong W:
Hesperetin induces the apoptosis of hepatocellular carcinoma cells
via mitochondrial pathway mediated by the increased intracellular
reactive oxygen species, ATP and calcium. Med Oncol. 32:1012015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Roohbakhsh A, Parhiz H, Soltani F, Rezaee
R and Iranshari M: Molecular mechanisms behind the biological
effects of hesperidin and hesperetin for the prevention of cancer
and cardiovascular diseases. Life Sci. 124:64–74. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Elango R, Athinarayanan J, Subbarayan VP,
Lei DKY and Alshatwi AA: Hesperetin induces an apoptosis-triggered
extrinsic pathway and a p53-independent pathway in human lung
cancer H522 cells. J Asian Nat Prod Res. 20:559–569. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gabai VL, Meriin AB, Mosser DD, Caron AW,
Rits S, Shifrin VI and Sherman MY: Hsp70 prevents activation of
stress kinases. A novel pathway of cellular thermotolerance. J Biol
Chem. 272:18033–18037. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Park HS, Cho SG, Kim CK, Hwang HS, Noh KT,
Kim MS, Huh SH, Kim MJ, Ryoo K, Kim EK, et al: Heat shock protein
hsp72 is a negative regulator of apoptosis signal-regulating kinase
1. Mol Cell Biol. 22:7721–7730. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mosser DD and Morimoto RI: Molecular
chaperones and the stress of oncogenesis. Oncogene. 23:2907–2918.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Beere HM, Wolf BB, Cain K, Mosser DD,
Mahboubi A, Kuwana T, Tailor P, Morimoto RI, Cohen GM and Green DR:
Heat-shock protein 70 inhibits apoptosis by preventing recruitment
of procaspase-9 to the Apaf-1 apoptosome. Nat Cell Biol. 2:469–475.
2000. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Saleh A, Srinivasula SM, Balkir L, Robbins
PD and Alnemri ES: Negative regulation of the Apaf-1 apoptosome by
Hsp70. Nat Cell Biol. 2:476–483. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gotoh T, Terada K, Oyadomari S and Mori M:
hsp70-DnaJ chaperone pair prevents nitric oxide- and CHOP-induced
apoptosis by inhibiting translocation of Bax to mitochondria. Cell
Death Differ. 11:390–402. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Stankiewicz AR, Lachapelle G, Foo CZ,
Radicioni SM and Mosser DD: Hsp70 inhibits heat-induced apoptosis
upstream of mitochondria by preventing Bax translocation. J Biol
Chem. 280:38729–38739. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang J, Wu D, Vikash, Song J, Wang J, Yi
J and Dong W: Hesperetin induces the apoptosis of gastric cancer
cells via activating mitochondrial pathway by increasing reactive
oxygen species. Dig Dis Sci. 60:2985–2995. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Parry N, Wheadon H and Copland M: The
application of BH3 mimetics in myeloid leukemias. Cell Death Dis.
12:2222021. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Los M, Mozoluk M, Ferrari D, Stepczynska
A, Stroh C, Renz A, Herceg Z, Wang ZQ and Schulze-Osthoff K:
Activation and caspase-mediated inhibition of PARP: A molecular
switch between fibroblast necrosis and apoptosis in death receptor
signaling. Mol Biol Cell. 13:978–988. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Qian SB, McDonough H, Boellmann F, Cyr DM
and Patterson C: CHIP-mediated stress recovery by sequential
ubiquitination of substrates and Hsp70. Nature. 440:551–555. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kundrat L and Regan L: Identification of
residues on Hsp70 and Hsp90 ubiquitinated by the cochaperone CHIP.
J Mol Biol. 395:587–594. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tresserra-Rimbau A, Medina-Remón A,
Pérez-Jiménez J, Martínez-González MA, Covas MI, Corella D,
Salas-Salvadó J, Gómez-Gracia E, Lapetra J, Arós F, et al: Dietary
intake and major food sources of polyphenols in a Spanish
population at high cardiovascular risk: The PREDIMED study. Nutr
Metab Cardiovasc Dis. 23:953–959. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chun OK, Chung SJ and Song WO: Estimated
dietary flavonoid intake and major food sources of U.S. adults. J
Nutr. 137:1244–1252. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen X, Wei W, Li Y, Huang J and Ci X:
Hesperetin relieves cisplatin-induced acute kidney injury by
mitigating oxidative stress, inflammation and apoptosis. Chem Biol
Interact. 308:269–278. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
He P, Ma J, Liu Y, Deng H and Dong W:
Hesperetin promotes cisplatin-induced apoptosis of gastric cancer
in vitro and in vivo by upregulating PTEN expression. Front
Pharmacol. 11:13262020. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Schmitt E, Gehrmann M, Brunet M, Multhoff
G and Garrido C: Intracellular and extracellular functions of heat
shock proteins: Repercussions in cancer therapy. J Leu Biol.
81:15–27. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yang J, Liu X, Bhalla K, Kim CN, Ibrado
AM, Cai J, Peng TI, Jones DP and Wnag X: Prevention of apoptosis by
Bcl-2: Release of cytochrome c from mitochondria blocked. Science.
275:1129–1132. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ismail NI, Othman I, Abas F, H Lajis N and
Naidu R: Mechanism of apoptosis induced by curcumin in colorectal
cancer. Int J Mol Sci. 20:24542019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Feder JH, Rossi JM, Solomon J, Solomon N
and Lindquist S: The consequences of expressing hsp70 in Drosophila
cells at normal temperatures. Genes Dev. 6:1402–1413. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Volloch VZ and Sherman MY: Oncogenic
potential of Hsp72. Oncogene. 18:3648–3651. 1999. View Article : Google Scholar : PubMed/NCBI
|