Multiple novel radiotherapies have emerged in recent
years, including brachytherapy, carbon ion radiotherapy and proton
therapy, which can be further subdivided into photothermal therapy
and photodynamic therapy (1-3). The
abscopal effect describes the shrinkage of unirradiated tumors that
occurs concurrently with irradiated tumors in patients with
multiple tumors. Since Mole's first proposal of this effect
(4), there had only been a few
cases reported, until the association with the immune system was
demonstrated by previous research that found the abscopal effect
was not observed in mice with immunodeficiency (5). With immunotherapy becoming a more
effective treatment for tumors (6,7),
clinical studies have revealed that a combination of radiotherapy
and immunotherapy, such as the immune checkpoint inhibitor
anti-cytotoxic T lymphocyte-associated antigen-4 (anti-CTLA-4),
anti-programmed death-1 (anti-PD-1) and anti-programmed cell death
1 ligand 1 (anti-PD-L1), produces the abscopal effect in the
treatment of lung cancer (8),
prostate cancer (9), melanoma
(10), breast cancer (11), liver cancer (12), type B3 thymoma (13) and glioblastoma (GBM). As a result,
the mechanism of the abscopal effect and how to produce it by
combining immunotherapy and radiotherapy has become a hotspot in
tumor research, which is also the focus of the present review.
The immunological mechanism of action behind the
abscopal effect remains unknown, although various studies have
shown that it is dependent on T cells (5,14,15)
and macrophages (7). Thus far, it
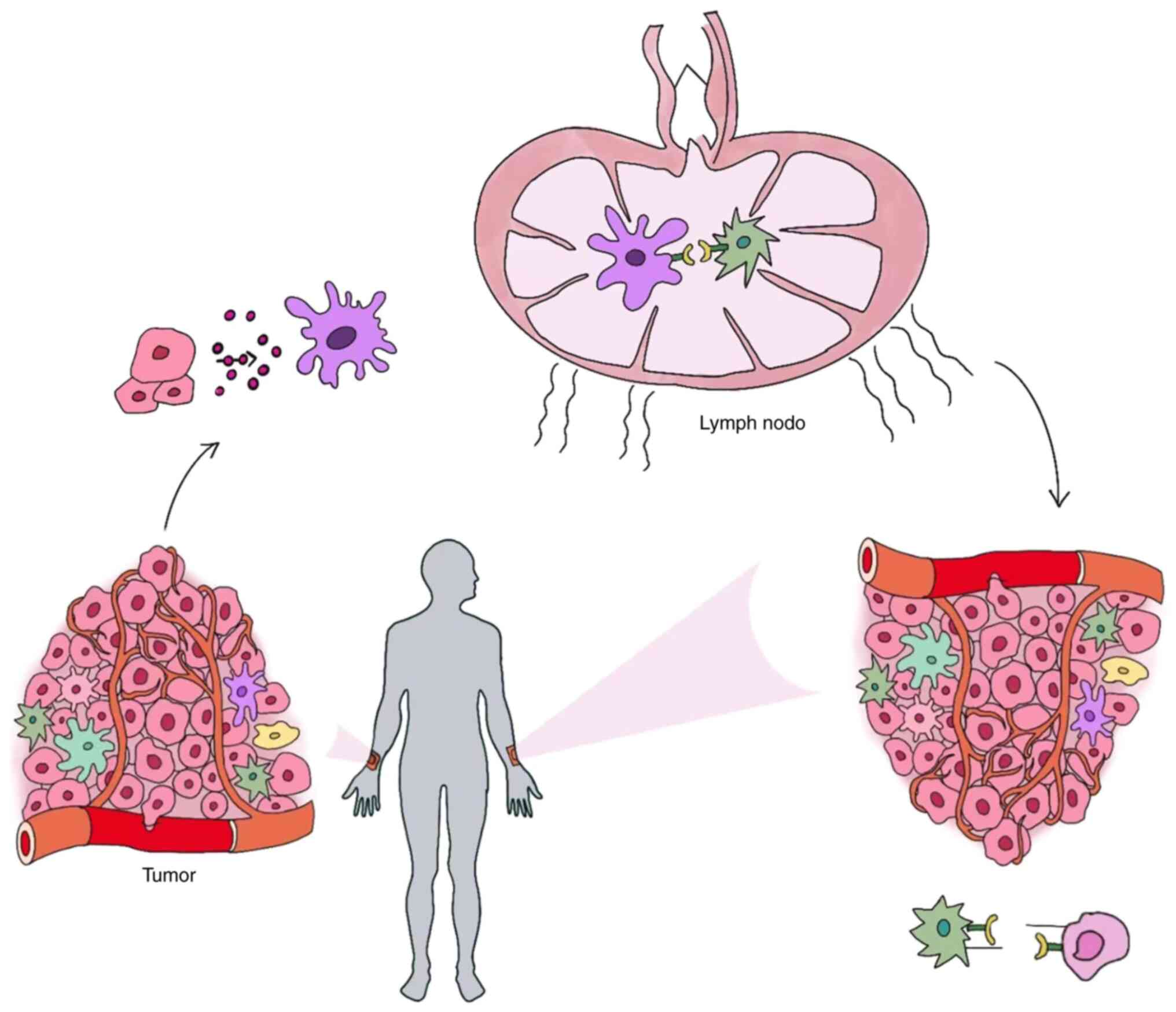
is known that tumor cells are triggered to produce tumor-associated
antigens (TAAs) when their DNA is damaged. TAAs are then
phagocytosed by antigen-presenting cells (APCs) before being
activated by major histocompatibility complex (MHC) molecules on
CD8+ T cells. Since CD8+ T cells not only
have a direct impact on primary tumors but also reach untreated
tumors via the blood and lymph circulation, they attach to tumor
cells and destroy them, thus exerting an antitumor effect (16).
Exosomes are secreted by cells for intercellular
signal transduction and information exchange. They carry nucleic
acid, proteins and lipids to the target cell by acting on its
surface or fusing with it (17).
Exosomes derived from various cells serve an immunosuppressive or
immunoenhancing role (18), and
participate in carcinogenesis, proliferation and metastasis
(19), thus playing distinct roles
in the abscopal effect. Furthermore, there are alterations in
exosome secretion following radiation (20,21).
The subsequent sections discuss the mechanisms by which distinct
cells produce exosomes after radiation to modify the abscopal
effect.
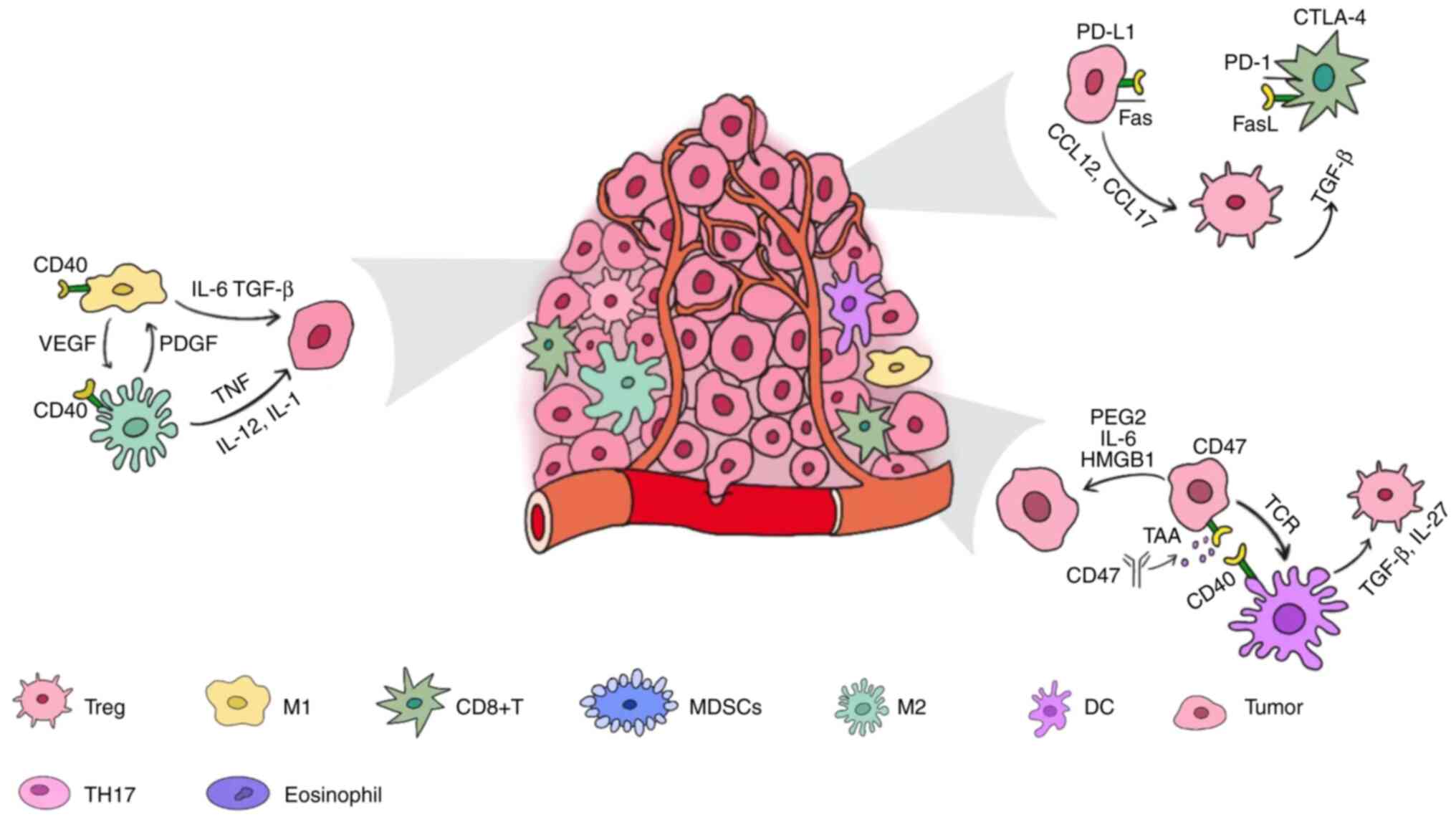
Tumor cells become more immunogenic after being
irradiated because their DNA is damaged, thus causing the
production of TAAs. The endoplasmic reticulum, which contains
calreticulin and the disulfide isomerase ERp57, migrates to the
plasma membrane and delivers an 'eat-me' signal to APCs, enhancing
their phagocytosis and abscopal action. On the other hand, due to
the formation of cytoplasmic double stranded DNA (dsDNA) induced by
radiation, GMP-AMP synthase (cGAS) and dsDNA initiate the formation
of cyclic guanosine monophosphate-adenosine monophosphate (cGAMP)
(22). The increased cGAMP level
combines with Stimulator of interferon genes (SINGs) to help
regulate the activity of downstream immune stimulating genes, and
ultimately promote the maturation and migration of dendritic cells
and the activity of CD8+ T cells, thus playing an
antitumor role (23,24). Certain exosomes produced by tumor
cells can enhance the antitumor effect. They are highly rich in
proteins, such as CD40L, which activates the CD40 signaling pathway
in dendritic cells (DCs), and induces DCs to mature and produce
IL-12, thus promoting anti-tumor immunity (25).
Previous research indicated that tumors with
neoepitopes, such as epidermal growth factor receptor (EGFR) vIII
in GBM, are more vulnerable to the abscopal effect (7), albeit the mechanism is unknown and
requires additional exploration. This tumor type also produces
exosomes with high EGFRvIII levels (26).
The radiation-induced damage to DNA, on the other
hand, can be repaired and therefore blocked by the DNA exonuclease
3 repair exonuclease 1 inside the tumor, and the degree to which
the calreticulin is exposed will be lowered by tumor cell
autophagy. Furthermore, CD47 on the plasma membrane can inhibit
phagocytosis of tumor cells by the immune system and emit a
'do-not-eat-me' signal (27) to
offset the impact of 'eat-me' signals (28). Furthermore, the expression of PD-L1
on the surface of tumor cells increases, and the combination of
PD-L1 and PD-1 on the surface of CD8+ T cells acts as an
immunological brake, weakening the action of effector T cells
(29-31), and therefore limiting the abscopal
effect (Fig. 1).
There are immunosuppressive exosomes as well as
immunoenhancing exosomes. Some of them produce cell membrane
proteins, including PD-L1 and MHC. According to a previous study,
PD-L1 immunosuppression on tumor cell-derived exosomes is
considerably greater than that on the membrane (32). These PD-L1-rich exosomes perform
the same function as PD-1 on the tumor surface, and their
immunosuppressive impact is amplified when they express both PD-L1
and MHC molecules (33).
Furthermore, certain cancer types produce exosomes that increase
PD-1 expression on CD8+ T cells (34) and increase PD-L1 expression on the
surface of macrophages (35),
shielding tumor cells from CD8+ T cells. Exosomes rich
in TGF-β and IL-10 may increase tumor migration and invasion
(36), and tumor-derived exosomes
can cause tumor immune evasion via the T-cell immunoglobulin and
mucin domain 1 (TIM-1) signaling pathway (37). Furthermore, tumor-derived exosomes
are high in Fas-L (38), which
acts on the surface of T cells to trigger their death, thus
markedly reducing antitumor immunity. Meanwhile, these exosomes'
surfaces may be rich in integrin β 3 (ITGB3), which can activate
focal adhesion kinase and influence intracellular signaling
cascades, promoting tumor spread (39). The distinction is that tumor cells
from the original location secrete exosomes rich in integrin β-like
1 (ITGBL1) to act on the distant site, thus activating the
EVs-ITGBL1-CAFs-TNFAIP3-NF-κB signaling axis (40) to modify the tumor microenvironment
(TME) and accelerate tumor metastasis, severely reducing the
abscopal effect. A previous study has found that GBM-induced
exosomes contain CD274, DNA and other chemicals that influence the
transcriptional activator 3 STAT3 signaling pathway and promote
macrophage polarization to M2 cells (41). Finally, exosomes from lymphoma
cells include the inhibitor of apoptosis protein survivin, which
inhibits natural killer (NK) cell surveillance and cytotoxicity
(42).
Aside from proteins, nucleic acids in exosomes play
an important role in the abscopal action. DNA, for example, can
stimulate the STING signaling system, boost antitumor immunity and
promote the abscopal effect (43).
Furthermore, colorectal cancer-induced long non-coding RNA
(lncRNA)-rich exosomes operate on the TME to stimulate the
proliferation and differentiation of T helper (Th)17 cells and
enhance the antitumor action (44). Meanwhile, microRNAs (miRNAs or
miRs) in exosomes can influence IL-10 activation, resulting in an
immunosuppressive TME that inhibits the development of the abscopal
effect. Among these miRNAs, miR-212-3p has been shown to diminish
MHC-2 expression and induce immunological tolerance in DCs,
allowing tumors to escape immune surveillance (45). It has been reported that miR-934
(46), miR-301a-3p (47), miR-21-3p, miR-125b-5p and
miR-181d-5p (48) can cause
macrophages to polarize toward M2 cells, hence promoting tumor
spread. These tumor-derived exosomes can act not only on the local
TME, but also on the distant tumor site via the circulatory
system.
There are numerous strategies for reducing the
release of tumor-induced exosomes, which limit the abscopal effect.
The first is to target and block exosomes from being released via
routes such as endosomal sorting complex required for transport
(ESCRT), tumor susceptibility gene 101 protein and other ESCRT
proteins or ESCRT auxiliary molecules (49). The second is to influence the
exosome acceptance pathway. The third option is to disrupt the
functional pathway, such as by utilizing anti-PD-L1 or anti-PD-1
antibodies to prevent exosomes from acting on cells. In addition,
radiotherapy can reduce the secretion of exosomes that promote
tumor proliferation and metastasis by 25.8% (50). Furthermore, brachytherapy can boost
the release of particular exosomes rich in high mobility group to
induce macrophage polarization to M1 cells (51), hence considerably enhancing the
abscopal effect. A combination of radiotherapy and immunotherapy
can restrict the production and activity of immunosuppressive
exosomes, and it remains to be explored which combined option
should be utilized to do so while also increasing the release of
immunoenhancing exosomes.
Upon radiotherapy, tumor cells will release
damage-associated molecular patterns (DAMPs) such as ATP, high
mobility group box 1 protein (HMGB1), nucleic acid, prostaglandin
E2 (PGE2), sphingosine 1-phosphate, IL-6 and granulocyte macrophage
colony stimulating factor (GM-CSF) (29,52,53)
to exert their different biochemical effects.
On one hand, HMGB1 binds to receptor for advanced
glycation end product (RAGE), a self-receptor, to promote tumor
development and immunological tolerance while also inhibiting the
abscopal effect. When its oxidation sites are blocked, however,
HMGB1 increases immunity and enhances the abscopal effect (54,55).
The production of ATP at the start of radiation can
stimulate the activation of DCs and effector T cells, boosting the
abscopal effect (12,56-58).
However, a previous study found that high ATP release could cause
PD-1 overexpression in tumor cells as well as regulatory T cell
aggregation. It could also cause the decrease of the entire immune
process via APCs that promote immunological tolerance (59), thus substantially reducing the
abscopal effect. Meanwhile, radiotherapy-induced dying tumor cells
predominantly release ATP, which is converted to adenosine
monophosphate (AMP) and adenosine by ecto-5′-nucleotidase expressed
on macrophages, thus activating the A2a adenosine receptor on
macrophages and suppressing macrophage-mediated antitumor immunity
(60). In addition, ATP and
adenosine diphosphate (ADP) are cleaved into AMP by CD39, and then
CD73 converts AMP to adenosine, thereby inhibiting the stimulation
of CD8+ T cells, activating regulatory T cells (Tregs)
and promoting the differentiation of M2 macrophages to inhibit
antitumor immune responses (61).
CD39 and CD73 together play an important role in transforming an
ATP-mediated proinflammatory TME into an adenosine-mediated
immunosuppressive microenvironment (62). Furthermore, ATP and its metabolites
ADP and AMP all have immunosuppressive roles in inhibiting the
abscopal effect; thus, sedatives of several target adenosines are
being studied in the clinical practice to restore the abscopal
effect.
TGF-β affects the generation and activity of a range
of immune cells. It regulates acquired immunity by directly
stimulating Treg cell proliferation while suppressing the
production and function of CD8+ T cells and
antigen-presenting DCs. Similarly, TGF-β regulates the innate
immune system by decreasing NK cells and controlling the complex
activities of macrophages and neutrophils, as well as counteracting
the anti-CTLA-4 and anti-PD-1 effects of immunosuppressive
medications. As a result, research recently being conducted aimed
to diminish TGF-β's impact on immunological drugs and prevent the
reduction in acquired immunity by limiting its function, thus
improving the abscopal effect (63,64).
To summarize, tumor cells perform a dual role in the
abscopal effect. In general, tumors tend to block the abscopal
effect for self-protection as the disease advances. As a result,
efforts should be made to increase tumor cell immunogenicity and
immunostimulation while decreasing their immunosuppressive
effect.
DCs are the most effective APCs. Their Toll-like
receptor (TLR) receives radiotherapy-induced TAAs via a
TLR-signaling network (65), thus
allowing DCs to identify, phagocytose and process TAAs. After
swallowing TAAs, APCs can produce antigen peptides and
costimulatory signals that activate T cells and amplify the
abscopal effect. This boosting effect is smaller than that obtained
directly from APCs (66), but it
is sufficient to promote the abscopal effect. Thus, the tumor is
driven to create 'danger signals', which activate DCs and promote
the T cell response via MHC molecules. As a result, MHC-1 and
MHC-2-rich exosomes generated by mature DCs have been used in
clinical immunotherapy (67).
Furthermore, exosomes generated by DCs during irradiation can
strongly activate NK cells to destroy tumor cells (68). In comparison, despite their large
number, immunological DC-induced exosomes had a significantly
lesser effect than mature DC-induced microvesicles, which was
analogous to their role in the abscopal effect (69). Activated DCs significantly enhanced
the CD8+ T/Treg ratio in both primary (irradiated) and
secondary (unirradiated) tumors, thus amplifying the abscopal
effect (70). Furthermore, the
insensitivity of DCs to irradiation, in comparison to that of tumor
cells, ensures their survival and activity under high-dose
irradiation, thus facilitating that their relatively high
antigen-presenting effects are retained in the TME when high-dose
irradiation is required to ensure therapeutic outcomes (71).
Tumor cell-induced DCs, on the other hand, produce
TGF-β, IL-27 and other molecules that inhibit their own activity,
activate Tregs and limit antitumor immunity function, thus
decreasing the immune response and the abscopal effect (72,73).
Immature DCs severely impede the function of effector T cells and
suppress the immune response, resulting in the body's failure to
generate the abscopal effect (74). One of the most essential features
of solid tumors is a hypoxic TME, which substantially inhibits APC
activity and reduces the antitumor impact. To reinforce the
abscopal effect (75), it is
worthwhile examining methods to improve DCs' antitumor
immunological action while decreasing their immunosuppressive
role.
Tumor-associated macrophages (TAMs) phagocytose
tumor cells by recognizing their TAAs. TAMs can cause the abscopal
effect in two ways: i) By attaching to T cells to achieve the
antitumor immunological response or ii) By self-activation to reach
the distal tumor location and kill tumor cells. Previous research
has demonstrated that anti-PD-L1 can directly activate macrophages
to boost the abscopal effect (7),
and that it is a promising technique to promote the combination of
activated macrophages with targeted radiation to enhance tumor cell
damage and, thus, the abscopal effect (76).
TAMs are categorized into several subtypes, the most
common of which are M1 and M2. M1 cells primarily contribute to
antitumor immunity by directly phagocytosing tumor cells and
secreting cytokines such as TNF to macrophages of the M1 phenotype.
M1 macrophages can improve antitumor immunity by promoting the
activation of effector T cells and the maturation of DCs by
producing cytokines such as TNF-β, IL-6 and IL-23 (77). Meanwhile, M1 macrophages produce
exosomes that interfere with the NF-κB signaling pathway while
activating the caspase 3 signaling pathway, resulting in M1
macrophage polarization (78). M2
macrophages, on the other hand, predominantly suppress the immune
system through the production of angiogenesis factors, growth
factors and proteases, thus hastening the development of malignant
tumors. M2 macrophages release exosomal miR-590-3p, which passes
through the target LATS1 and activates YAP/β-catenin to decrease
the immunological response (79).
They also release integrin αMβ2-rich exosomes, which activate the
MMP-9 signaling pathway in receptor tumor cells, as well as
apolipoprotein E-rich exosomes, which promote tumor spread and
proliferation (80,81). When M2 macrophages produce
miRNA-21, it inhibits cell death, increases PI3K/AKT signaling
pathway activation by downregulating PTEN (82), and boosts drug resistance in
gastric cancer cells as well as tumor proliferation and metastasis,
thus decreasing the abscopal effect.
Furthermore, M1 and M2 cells can convert into each
other type, resulting in a shift in macrophage activity between
antitumor and pro-tumor effects, which has a marked impact on the
development of the abscopal effect.
The polarization of M1 and M2 cells is mutually
hostile and complex, and it is strongly associated with radiation.
According to a previous study, when a cumulative quantity of 10 Gy
is obtained based on a daily dosage of radiation of 2
Gy/fraction/day, the number of M1 cells increases while the number
of M2 cells declines (89). When
local low-dose irradiation is administered, M2 cells polarize back
to M1 cells, promoting CTL penetration into the TME (90), improving the therapeutic effect. On
the other hand, another study found that M2 cells were less
sensitive to radiation than M1 cells, so that the ratio of M2/M1
cells increased when the cumulative radiation dosage reached a
particular level (91). These
seemingly contradicting outcomes are strongly linked to the
radiation dose and method. In any case, M2 infiltration and
polarization during radiation has a significant impact on patients'
prognosis, and diminish their survival rate (92). Therefore, it is worth investigating
how to activate M1 cells while suppressing M2 cell activation, as
well as how to re-convert M2 cells into M1 cells to minimize the
M2/M1 cell ratio.
TAMs steadily increase the expression of PD-1 while
exerting antitumor immunological actions. Since PD-1 expression is
inversely correlated with macrophage phagocytic and
antigen-presenting capabilities (93), tumor escape can be inhibited by
reducing PD-1 expression, and anti-PD-1 treatment can boost
macrophage antitumor efficacy. TAMs, on the other hand, overexpress
indoleamine 2,3-dioxygenase (IDO) (94), which inhibits CTL activity.
Furthermore, CD40 on the surface of APCs will be highly expressed,
weakening their function; thus, currently, studies are using
multi-functional radiotherapy-associated biological materials to
inhibit the expression of CD40 by delivering an anti-CD40 antibody
to a tumor in situ, which allows the antitumor effects to be
maintained (95).
TAMs also cause exosomes containing miR-29a-3p and
miR-21-5p to inhibit STAT3, resulting in significant Treg
proliferation (96), whereas
exosomes containing miR-155 can control the TME, prompting
macrophages to polarize into M1 cells (97,98).
When TAMs produce lysyl oxidase like 4-rich exosomes, they promote
the production of their own PD-L1, resulting in an
immunosuppressive phenotype that inhibits the activity of
CD8+ T cells (99),
significantly decreasing antitumor immunity and the development of
the abscopal effect. Furthermore, macrophages can produce
GM-CSF-rich exosomes to enhance tumor angiogenesis (100), supply oxygen and nutrients to
tumors, and hence provide possibilities for tumor migration and
invasion.
Despite the several conditions that can restrict
APCs' function, it has been observed that when antigens are
available, boosting APCs function alone can induce a sufficiently
significant abscopal effect. As a result, targeted activation of
APCs contributes to the intensification of the abscopal effect.
There are various T cell types, and all play crucial
roles in the generation of the abscopal effect, with some of them
enhancing each other, while others antagonize each other.
The killing effect of CTLs can be activated when
they are targeted by the auto-specific antigen of certain tumors,
such as melanoma-associated antigen 1 (107) in melanoma and EGFRvIII (7) in GBM; therefore, these tumors are
more susceptible to the abscopal effect. Furthermore,
CD8+ T cells stimulate DCs by releasing exosomes rich in
cytoplasmic DNA, and DCs are activated via the cGAS/STING signaling
pathway (108). By creating
immunological synapses and suppressing apoptosis, DCs stimulate
CD8+ T cells, forming a powerful immune impact that can
boost the abscopal effect.
The presence of CD73 on the surface of Treg-derived
exosomes is required for Tregs to mediate immunosuppressive
effects, and its mechanism is similar to that of Th17 cells, which
facilitates the conversion of ATP to ADP and AMP (117,118). When the TCR on the surface of
Tregs is activated, it increases exosome secretion, inhibits the
tumor-killing activity of CTLs and inhibits the proliferation of
effector T cells (119). The
activated TCR promotes the release of IL-4 and IL-10, which is
linked to miR-150-rich exosomes generated by Tregs (120). Treg-secreted miR-146a-5p-rich
exosomes can play a role via suppressing STAT1 and interleukin 1
receptor associated kinase 2 (121).
Tregs serve an important function in preventing DC
maturation. They produce exosomes and deliver them to DCs for
intercellular communication with miRNAs in DCs (122), which significantly reduces
antitumor immunity. Let-7d miRNA is encased in exosomes and
preferentially acts on Th1 cells, reducing their proliferation and
immunity by inhibiting cyclooxygenase 2 (123). Certain Treg-derived exosomes have
a 25-100-fold higher IL-35 concentration in the cell surface, and
IL-35 acts on target cells to promote the expression of PD-1, Tim3
and Lag3 (124), suppressing
antitumor immunity. In summary, Tregs can suppress antitumor
immunity and the abscopal effect by secreting various exosomes.
After radiotherapy, CXCR4 antagonist can promote the
depletion of Tregs and enhance the antitumor and anti-metastasis
therapeutic effect (125,126). Depletion also encourages T cells
to mature into effector memory T cells (127), as well as inhibiting Treg
proliferation, resulting in a higher CD8+ T cell/Treg
ratio. Tregs are less vulnerable to radiation than CTLs, and their
increased activity can be maintained even at larger doses (128); therefore, the radiation dose can
be utilized to modify the CD8+ T cell/Treg ratio.
According to a previous study, hypo-fractionated stereotactic
radiation therapy can increase the number of CD8+ T
cells while decreasing the number of Tregs (129).
The role of Th17 cells in malignancies, on the other
hand, is complex, and their processes are unknown. Meanwhile, their
effects differ depending on the tumor type. Thus, further research
is required. To summarize, different types of T cells play diverse
roles in the abscopal effect. Only by further studying and
specifying the role and mechanism of distinct T cells, as well as
the connections between T cells and other cells, the human body's
antitumor immunity will be better understood and maintained.
MDSCs, which are immature myeloid cells produced and
secreted by the bone marrow, are recruited to the TME to control
the immune response and build an immunological-tolerant TME.
Meanwhile, additional MDSCs will be created by the bone marrow,
which is triggered by chronic inflammatory signals sent out by the
TME (138), thus creating a
feedback loop that interferes with tumor therapy. According to a
previous study, tumor cells in a model lacking MDSCs were quickly
removed by activated antitumor immune cells (139), indicating the critical role of
MDSCs in the immunosuppressive process. Exosomes secreted by MDSCs
play an important role in suppressing the abscopal effect and
promoting tumor immune escape, proliferation and migration. Thanks
to their contents (>4,000 types of protein), MDSCs-derived
exosomes and MDSCs support each other, strengthen the
immunosuppressive effect, and promote tumor proliferation and
survival (140).
It has been demonstrated that, even during
radiation, MDSCs suppress the abscopal effect, since they arrive at
the tumor site 10 days after radiotherapy (139), and decrease antitumor immunity by
releasing cytokines such as TGF-β and strongly expressing PD-L1
(141). Proliferation of MDSCs
can be aided in a variety of ways. VEGF has been shown to play a
significant role in promoting the proliferation of MDSCs (142), and an increase in MDSCs and a
decrease in effector T cells have been observed in tumor models
with high VEGF expression (143),
implying that VEGF inhibits the abscopal effect by promoting
MDSCs.
Moreover, the proliferation of MDSCs is promoted by
PGE2, which activates the p38MAPK/ERK signaling pathway to enhance
the release of TGF-β (150,151). HMGB1 is a high-content protein in
MDSCs-derived exosomes, which can induce the production and
accumulation of MDSCs. Other MDSCs-derived exosomes rich in TGF-β1,
IL-10 and IL-6 may be ingested by macrophages and T cells, causing
a significant increase in Tregs to play a stronger
immunosuppressive effect, a reduction in the proliferation of Th
cells, weakened CTL cytotoxic activity and a slight increase in the
lymphocyte apoptosis rate. Specifically, the content of TGF-β1
within these exosomes is 4.3-fold higher than that within the cell,
thus promoting tumor angiogenesis and metastasis (148,152,153). Moreover, it has been shown that
MDSC-derived exosomes are associated with the resistance of
chemotherapeutic drugs (152).
In summary, MDSCs can inhibit the generation of the
abscopal effect via a variety of mechanisms. Furthermore, the
greater the accumulation of MDSCs in the TME, the stronger its
immunosuppressive ability, the more conducive to tumor growth,
ultimately having a negative effect on the overall treatment of the
tumor. As a result, they may be utilized as a prognostic marker in
patients, and targeting MDSCs can help restore antitumor immunity
and boost the abscopal effect.
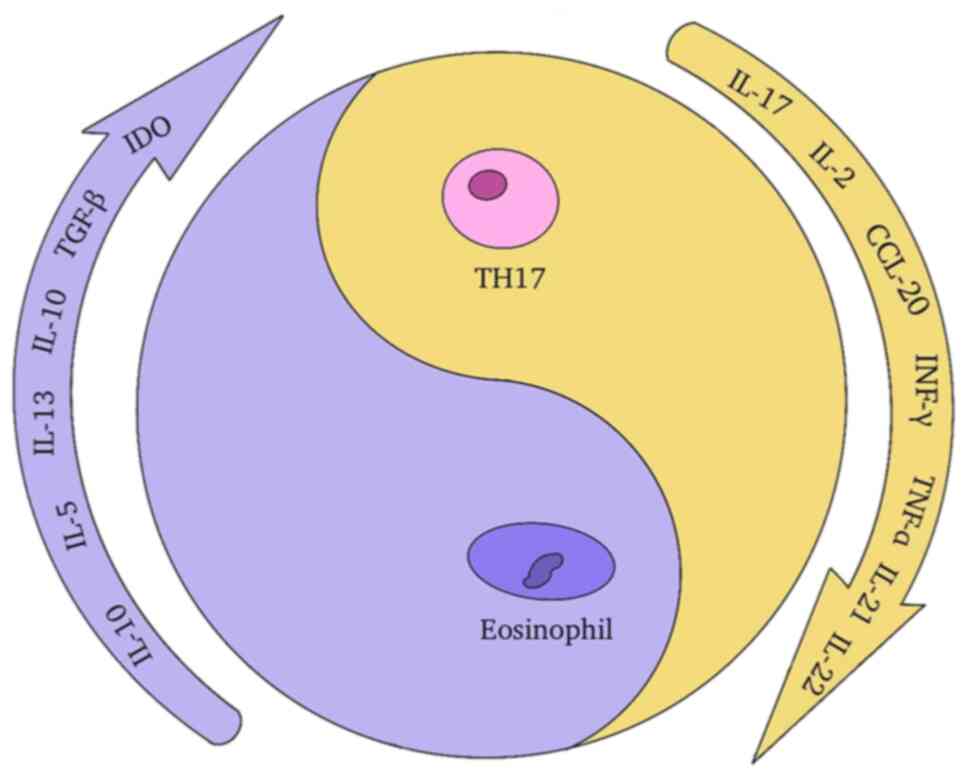
Eosinophils have been observed to infiltrate
numerous tumors, directly interact with tumor cells, govern tumor
formation by modifying the TME and contribute to antitumor immunity
via a number of pathways. Eosinophils have high levels of IL-1
receptor 1, TLR4 and RAGE expression. They also recruit to the
tumor site following radiotherapy-induced DAMPs such as HMGB1 and
IL-33 signals (154).
Eosinophils have a two-pronged effect on
malignancies. On one hand, they cause CD8+ T cells to
penetrate tumors, normalize blood vessels and drive macrophage
polarization to M1 cells, resulting in a marked antitumor action
(155). Although the underlying
mechanism is unknown, eosinophils can significantly boost the
abscopal effect. On the other hand, eosinophils play an inhibitory
function in immunity (156). When
triggered by thymic stromal lymphopoietin, they restrict DC
maturation and block tumor apoptosis by secreting a substantial
quantity of IL-10, IL-4, IL-5 and IL-13 (157), thus increasing tumor growth.
TGF-β (158) is also released by
eosinophils to impact the abscopal effect. Furthermore, they
produce IDO (159) to enhance
TME's immunosuppressive function, boost M2 polarization of
macrophages in the TME by releasing IL-13 and IL-4 (160), and decrease the antitumor effect.
As a result, inhibiting the tumor-promoting function of
eosinophils, so that they improve the antitumor action of
macrophages to promote the abscopal effect, is worth
investigating.
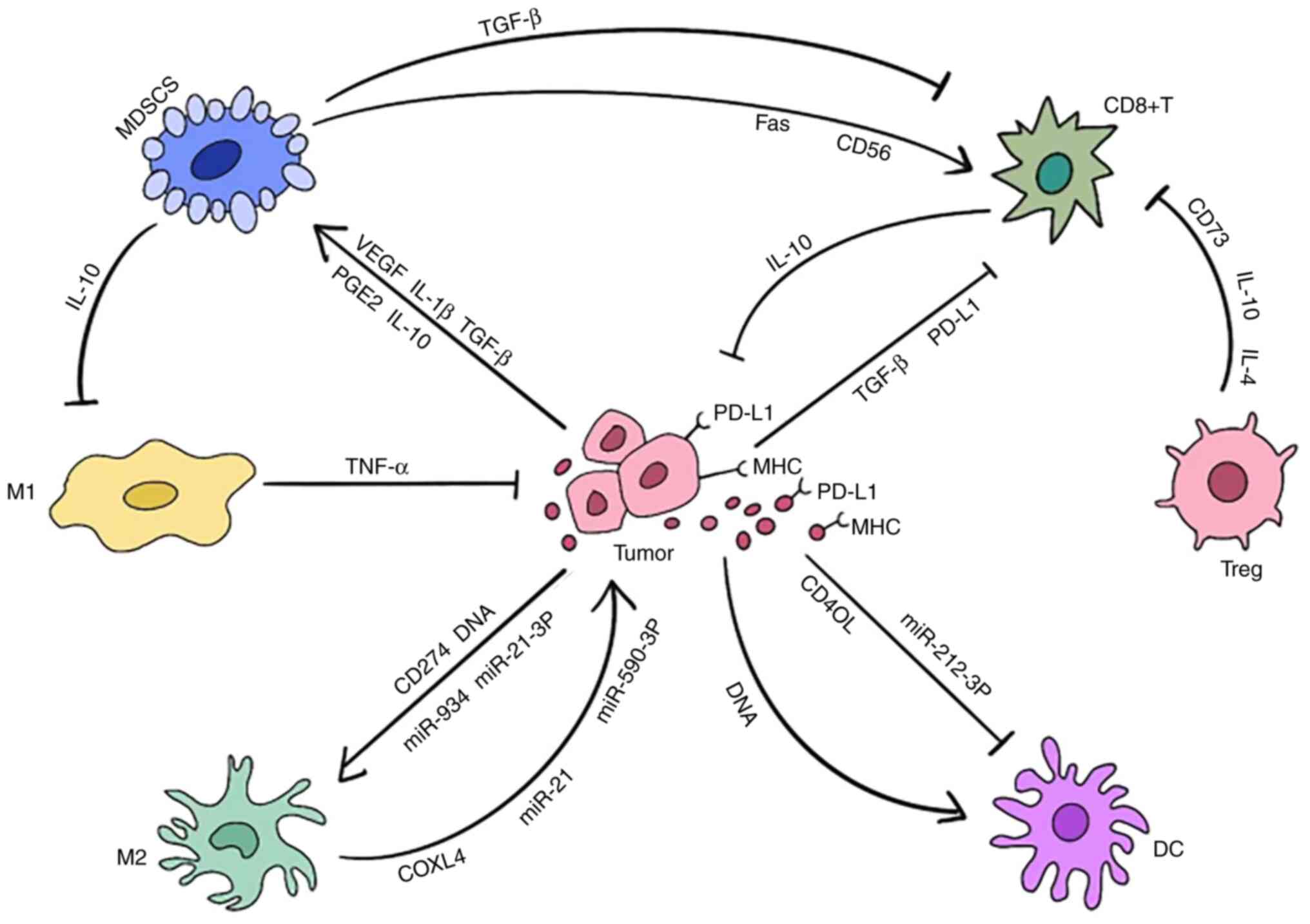
Immunosuppressive cells gradually win the fight to
suppress the abscopal effect later in the process. Despite the fact
that the current immunotherapy has overcome the disadvantages of
antitumor cells, the lack of knowledge of the underlying mechanism,
combined with the complexity of the aforementioned battles, hampers
the formation of the abscopal effect over time (Fig. 4).
Radiotherapy is an important cancer treatment that
can boost the abscopal effect. Radiotherapy was previously only
utilized to treat local malignancies due to technological
limitations and its harmful effects on normal tissues. In addition
to its cell-killing impact, radiation modulates immunomodulation.
Radiotherapy, for example, renders tumor cells more vulnerable to T
cell attack. After each 10-25 Gy low-fraction radiation session,
the expression of MHC-1 molecules on the surface of human melanoma
cells was increased (161), which
enhances the presentation of antigens, making it easier for these
tumor cells to be destroyed and removed by T lymphocytes.
Furthermore, various immune cells respond differently to radiation;
for example, a radiation dosage of 0.94 Gy strongly inhibits Treg
proliferation (162). Due to the
lack of studies on different tumor radiotherapy doses and the
susceptibility of different cells to radiotherapy, it is difficult
to utilize radiotherapy alone to overcome the inhibitory impact of
the TME, which is why the abscopal effect was uncommon in the
past.
However, immunotherapy compensates for this rarity.
Multiple clinical and pre-clinical studies (Table I) have shown that, compared with
the effect of radiotherapy or immunotherapy alone, the combination
of radiotherapy and immunotherapy can significantly increase the
incidence and intensity of the abscopal effect (7,13,163-166). Commonly used immune checkpoint
inhibitors are anti-CTLA-4, anti-PD-1 and anti-PD-L1 antibodies,
all of which strengthen the effect of T cells on tumor cells and
can be combined with radiotherapy to enhance the incidence rate of
the abscopal effect. A recent study showed that the combination of
8 Gy x3F radiotherapy and anti-PD-L1 monoclonal antibody can
enhance the abscopal effect, significantly reduce MDSCs and promote
CD8+ T cell infiltration (167). Another clinical study indicated
that radiotherapy combined with immune adjuvant GM-CSF treatment
can trigger the abscopal effect in 30% of patients with cancer
(168). Similarly, anti-CD40
antibody could maintain the antitumor effect of APCs (95), and FMS-like tyrosine kinase 3
ligand could recruit and stimulate APCs (169), so both of them can enhance the
effect of APCs, and increase the incidence of the abscopal effect
when combined with radiotherapy. In addition, when combined with
tumor cells, both anti-CD47 and anti-CD73 antibodies can promote
APCs to phagocytose tumor cells (170-172), and further exert their antitumor
role.
However, immunotherapy also has defects such as
cross-reactions. Specifically, while anti-CD47 eliminates tumor
cells, it may accidentally injure the red blood cells that also
carry CD47 on their surface (173), leading to anemia. Besides, the
overlap and systemic toxicity of a combination of radiotherapy and
immunotherapy are still difficult to deal with, although a
reasonable combination can overcome immunosuppression and promote
the generation of the abscopal effect (16). Moreover, immune-related adverse
reactions are worrying. Even if radiotherapy can overcome the drug
resistance of anti-PD-1 antibody, it does not have a long-lasting
therapeutic effect on ~80% of patients with non-small cell lung
cancer (NSCLC), due to tumor oxidative metabolism obstacles
(174). The dosage and sequence
of the combination of radiotherapy and immunotherapy for different
tumors, or different combination options from different tumor
radiotherapy and immunotherapy methods are currently unclear;
therefore, it is necessary to further explore immunotherapy options
and timings for different tumor types to find the best timing,
dosage and sequence of the combination of radiotherapy and
immunotherapy. Emerging methods and technologies may help to
understand how to generate the abscopal effect and promote its
incidence rate, which is beneficial to the treatment and prognosis
of patients. These benefits are subsequently described in the
present review.
New radiotherapy technologies are conducive to the
abscopal effect. The development of radiotherapy technology has
made a great progress. Compared with traditional radiotherapy
methods, new technologies may be more conducive to the generation
of the abscopal effect. Stereotactic body radiation therapy
combined with immunotherapy is well tolerated and relatively safe,
and there were cases of lung cancer as well as head and neck
squamous cell carcinoma where the abscopal effect was generated by
such a combination (163,166). In addition, technologies such as
intensity-modulated radiation therapy, stereotactic ablative
radiotherapy or proton therapy can change the range of radiotherapy
according to the tumor size and greatly reduce radiation toxicity
(3), thus overcoming the toxicity
caused by combined therapy. Besides, high-dose radiation (HDR)
brachytherapy can protect adjacent healthy tissues by bringing the
emission source into the tumor tissue, thereby reducing
radiotherapy-induced toxicity (175). Research has shown that a
combination of HDR brachytherapy with anti-PD-1 or anti-CD137
antibodies can produce the abscopal effect (176). When these new radiotherapy
approaches are combined with different immunotherapy methods, the
best combination option and timing may be found, which may overcome
the limitations caused by the toxicity of radiotherapy in the past
and help to improve the current type of combination of radiotherapy
and immunotherapy.
As aforementioned, the combination of radiotherapy
and immunotherapy can achieve in an improved way the antitumor
effect and reduce the drug resistance to immunotherapy; however,
such combination also has a limited effect on the generation of an
abscopal effect sufficiently strong in certain tumors such as
NSCLC. The underlying mechanism of such limitation may be the
regulation of the ERK signaling pathway to act on Src homology
region 2-containing protein tyrosine phosphatase 2 (SHP2), which
regulates tumor cell proliferation (177) and is the main effector mediating
the downstream signal transduction of PD-1 in T cells (178). Previous research has shown that
the triple therapy of SHP2 inhibitor, anti-PD-L1 antibody and
radiotherapy can increase the ratio of M1/M2 cells and CTL/Treg
lymphocytes to stimulate antitumor immunity (174). In addition, the oxidative
phosphorylation (OXPHOS) of tumor mitochondria may be another cause
of the aforementioned poor effect of the combination of
radiotherapy and immunotherapy (179). The triple therapy of IACS-010759,
an OXPHOS inhibitor, combined with anti-PD-1 antibody and
radiotherapy can promote the abscopal effect (180) and resolve the problem of
anti-PD-1 resistance in NSCLC. Other triple or quadruple therapies
could be used to overcome the disadvantages of the traditional
combination of radiotherapy and immunotherapy. For example, genetic
ablation of the TGF-β signaling pathway components added to the
conventional radioimmunotherapy could trigger a powerful antitumor
response (63), as well as a
combination of anti-PD-1 treatment after radiotherapy (181) and targeted suppression of
antitumor immunity. Similarly, exosomes within the TME can also
enhance the therapeutic effect, and ultimately promote the abscopal
effect to prolong the survival time of the patient.
In oncolytic immunotherapy, an oncolytic virus is
often injected locally into tumors, which has a tropism for
malignant tumor cells and can replicate in tumor cells to
eventually promote their lysis (182). Oncolytic virus replication can
induce the death of tumor immunogenic cells, send out immunological
danger signals, promote tumors to produce TNF-α, and induce the
body to produce strong immune effects (183), thereby enhancing the occurrence
of the abscopal effect. The shrinkage of distal tumors after the
local injection of an oncolytic virus has been reported, and the
mechanism is similar to that of the abscopal effect (184). When oncolytic viruses are used in
combination with immune checkpoint inhibitors and radiotherapy
their effects can be enhanced. Specifically, the oncolytic
adenovirus is currently one of the most promising oncolytic viruses
(185). Recently, a patient with
Hodgkin's lymphoma infected with the new coronavirus experienced
systemic tumor regression. The reason may be that the coronavirus
triggered antitumor immunity in his body (186); therefore, this novel coronavirus
may also have the potential to be developed as an oncolytic virus
to promote the abscopal effect.
Smart material technology is divided into
nanoparticles and intelligent radiotherapy biomaterials (187). Within nanoparticles,
nanoparticle-delivered drugs have great potential for improving the
antitumor immune effect. Nano-immunotherapy, which is the
combination of nanoparticle-delivered drugs and immunotherapy, can
be achieved in three different ways, and these nano-drugs are used
to target cancer cells and the TME (58). When targeting cancer cells,
nanoparticle-delivered drugs cause the immunogenic death of tumor
cells and can be combined with immunotherapy to greatly promote
antitumor immunity (188).
Moreover, when combined with photodynamic radiotherapy to treat
primary tumors, nanodrugs can promote the occurrence of abscopal
effects (189). Calcium carbonate
nanoparticles with anti-CD47 activity have been developed (190). When targeting the TME, nanodrugs
such as antigen capture nanoparticles can capture TAAs to activate
DCs, and thus promote the abscopal effect (191). Certain nanodrugs can also act on
immunosuppressive molecules, such as IDO, TGF-β and IL-2 (192) to reshape the TME, which is
beneficial for antitumor immunity.
The second type of smart material technology, smart
radiotherapy biomaterials, also promotes the abscopal effect. For
example, a hydrogel formed by alginate can capture the drug formed
by the combination of 131I-labeled catalase and the
immune adjuvant CpG, and the immune checkpoint inhibitor of the
combination of the hydrogel and the drug can produce powerful
antitumor immunity and the abscopal effect, which has been observed
in experimental mice (193).
Compared with traditional technologies, these new technologies have
improved treatment methods to reduce toxicity towards normal
tissues and/or lymphocytes, have improved targeting ability, are
beneficial to patients, and can reduce the cost of treatment for
patients.
Tumor metastasis has caused the suffering and
mortality of >90% of patients with cancer. The abscopal effect
can be used to combat tumor metastasis. The biggest advantage of
this abscopal effect is the inhibition and elimination of distant
and metastatic tumors. Therefore, by further studying the
underlying mechanism and improved using of new technologies and
methods to enhance the abscopal effect, an improved treatment plan
for patients with cancer could be developed. Furthermore, new
radiotherapy and immunotherapy approaches based on cells and
exosomes that play a role in the abscopal effect are beneficial to
increase the incidence of abscopal effects in clinical practice.
Besides, its needs to be taken into consideration how to reduce the
toxicity caused by treatment, relieve the suffering of patients,
and reduce the cost of treatment. It is worth noting that microwave
ablation has been found to induce the abscopal effect in clinical
practice (194), and the effect
of oncolytic viruses is similar to that of the abscopal effect.
Therefore, investigating the mechanism of oncolytic viruses may
help to find another way to promote the abscopal effect.
Not applicable.
JW and GK wrote the manuscript. ZW and CL created
the figures. JW and JL revised the manuscript. All authors read and
approved the final version of the manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
The present study was supported by the National Students'
Innovation and Entrepreneurship Training Program (grant nos.
S202210632275, S202210632129 and 202110632052).
|
1
|
Chargari C, Deutsch E, Blanchard P, Gouy
S, Martelli H, Guérin F, Dumas I, Bossi A, Morice P, Viswanathan AN
and Haie-Meder C: Brachytherapy: An overview for clinicians. CA
Cancer J Clin. 69:386–401. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mohamad O, Tabuchi T, Nitta Y, Nomoto A,
Sato A, Kasuya G, Makishima H, Choy H, Yamada S, Morishima T, et
al: Risk of subsequent primary cancers after carbon ion
radiotherapy, photon radiotherapy, or surgery for localised
prostate cancer: A propensity score-weighted, retrospective, cohort
study. Lancet Oncol. 20:674–685. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kahalley LS, Peterson R, Ris MD, Janzen L,
Okcu MF, Grosshans DR, Ramaswamy V, Paulino AC, Hodgson D, Mahajan
A, et al: Superior intellectual outcomes after proton radiotherapy
compared with photon radiotherapy for pediatric medulloblastoma. J
Clin Oncol. 38:454–461. 2020. View Article : Google Scholar :
|
|
4
|
Mole RH: Whole body irradiation;
radiobiology or medicine? Br J Radiol. 26:234–241. 1953. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Demaria S, Ng B, Devitt ML, Babb JS,
Kawashima N, Liebes L and Formenti SC: Ionizing radiation
inhibition of distant untreated tumors (abscopal effect) is immune
mediated. Int J Radiat Oncol Biol Phys. 58:862–870. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tan AC, Ashley DM, López GY, Malinzak M,
Friedman HS and Khasraw M: Management of glioblastoma: State of the
art and future directions. CA Cancer J Clin. 70:299–312. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ene CI, Kreuser SA, Jung M, Zhang H, Arora
S, White Moyes K, Szulzewsky F, Barber J, Cimino PJ, Wirsching HG,
et al: Anti-PD-L1 antibody direct activation of macrophages
contributes to a radiation-induced abscopal response in
glioblastoma. Neuro Oncol. 22:639–651. 2020. View Article : Google Scholar
|
|
8
|
Lheureux S, Braunstein M and Oza AM:
Epithelial ovarian cancer: Evolution of management in the era of
precision medicine. CA Cancer J Clin. 69:280–304. 2019.PubMed/NCBI
|
|
9
|
Formenti SC, Rudqvist NP, Golden E, Cooper
B, Wennerberg E, Lhuillier C, Vanpouille-Box C, Friedman K, Ferrari
de Andrade L, Wucherpfennig KW, et al: Radiotherapy induces
responses of lung cancer to CTLA-4 blockade. Nat Med. 24:1845–1851.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang W, Zhang F, Deng H, Lin L, Wang S,
Kang F, Yu G, Lau J, Tian R, Zhang M, et al: Smart
nanovesicle-mediated immunogenic cell death through tumor
microenvironment modulation for effective photodynamic
immunotherapy. ACS Nano. 14:620–631. 2020. View Article : Google Scholar
|
|
11
|
Hu ZI, McArthur HL and Ho AY: The abscopal
effect of radiation therapy: What is it and how can we use it in
breast cancer? Curr Breast Cancer Rep. 9:45–51. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Beyls C, Haustermans K, Deroose CM, Pans
S, Vanbeckevoort D, Verslype C and Dekervel J: Could autoimmune
disease contribute to the abscopal effect in metastatic
hepatocellular carcinoma? Hepatology. 72:1152–1154. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Guan S, Wang H, Qi XH, Guo Q, Zhang HY,
Liu H and Zhu BJ: Abscopal effect of local irradiation treatment
for thymoma: A case report. Am J Transl Res. 12:2234–2240.
2020.PubMed/NCBI
|
|
14
|
Chakravarty PK, Alfieri A, Thomas EK, Beri
V, Tanaka KE, Vikram B and Guha C: Flt3-ligand administration after
radiation therapy prolongs survival in a murine model of metastatic
lung cancer. Cancer Res. 59:6028–6032. 1999.
|
|
15
|
Camphausen K, Moses MA, Ménard C, Sproull
M, Beecken WD, Folkman J and O'Reilly MS: Radiation abscopal
antitumor effect is mediated through p53. Cancer Res. 63:1990–1993.
2003.PubMed/NCBI
|
|
16
|
Ngwa W, Irabor OC, Schoenfeld JD, Hesser
J, Demaria S and Formenti SC: Using immunotherapy to boost the
abscopal effect. Nat Rev Cancer. 18:313–322. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sexton RE, Mpilla G, Kim S, Philip PA and
Azmi AS: Ras and exosome signaling. Semin Cancer Biol. 54:131–137.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Möller A and Lobb RJ: The evolving
translational potential of small extracellular vesicles in cancer.
Nat Rev Cancer. 20:697–709. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kalluri R and LeBleu VS: The biology,
function, and biomedical applications of exosomes. Science.
367:eaau69772020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
He C, Li L, Wang L, Meng W, Hao Y and Zhu
G: Exosome-mediated cellular crosstalk within the tumor
microenvironment upon irradiation. Cancer Biol Med. 18:21–33. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hu X, Qiu Y, Zeng X and Wang H: Exosomes
reveal the dual nature of radiotherapy in tumor immunology. Cancer
Sci. 113:1105–1112. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Vanpouille-Box C, Alard A, Aryankalayil
MJ, Sarfraz Y, Diamond JM, Schneider RJ, Inghirami G, Coleman CN,
Formenti SC and Demaria S: DNA exonuclease Trex1 regulates
radiotherapy-induced tumour immunogenicity. Nat Commun.
8:156182017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Craig DJ, Nanavaty NS, Devanaboyina M,
Stanbery L, Hamouda D, Edelman G, Dworkin L and Nemunaitis JJ: The
abscopal effect of radiation therapy. Future Oncol. 17:1683–1694.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhao X, Hu S, Zeng L, Liu X, Song Y, Zhang
Y, Chen Q, Bai Y, Zhang J, Zhang H, et al: Irradiation combined
with PD-L1(-/-) and autophagy inhibition enhances the antitumor
effect of lung cancer via cGAS-STING-mediated T cell activation.
iScience. 25:1046902022. View Article : Google Scholar
|
|
25
|
Wang J, Wang L, Lin Z, Tao L and Chen M:
More efficient induction of antitumor T cell immunity by exosomes
from CD40L gene-modified lung tumor cells. Mol Med Rep. 9:125–131.
2014. View Article : Google Scholar
|
|
26
|
Choi D, Montermini L, Kim DK, Meehan B,
Roth FP and Rak J: The impact of oncogenic EGFRvIII on the proteome
of extracellular vesicles released from glioblastoma cells. Mol
Cell Proteomics. 17:1948–1964. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hsieh RC, Krishnan S, Wu RC, Boda AR, Liu
A, Winkler M, Hsu WH, Lin SH, Hung MC, Chan LC, et al: ATR-mediated
CD47 and PD-L1 up-regulation restricts radiotherapy-induced immune
priming and abscopal responses in colorectal cancer. Sci Immunol.
7:eabl93302022. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Feng M, Jiang W, Kim BYS, Zhang CC, Fu YX
and Weissman IL: Phagocytosis checkpoints as new targets for cancer
immunotherapy. Nat Rev Cancer. 19:568–586. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
He S, Cheng J, Sun L, Wang Y, Wang C, Liu
X, Zhang Z, Zhao M, Luo Y, Tian L, et al: HMGB1 released by
irradiated tumor cells promotes living tumor cell proliferation via
paracrine effect. Cell Death Dis. 9:6482018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Leonardi GC, Falzone L, Salemi R, Zanghì
A, Spandidos DA, Mccubrey JA, Candido S and Libra M: Cutaneous
melanoma: From pathogenesis to therapy (Review). Int J Oncol.
52:1071–1080. 2018.PubMed/NCBI
|
|
31
|
Minton K: Predicting the anti-PD1
response. Nat Rev Immunol. 19:414–415. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Daassi D, Mahoney KM and Freeman GJ: The
importance of exosomal PDL1 in tumour immune evasion. Nat Rev
Immunol. 20:209–215. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bennett F, Luxenberg D, Ling V, Wang IM,
Marquette K, Lowe D, Khan N, Veldman G, Jacobs KA, Valge-Archer VE,
et al: Program death-1 engagement upon TCR activation has distinct
effects on costimulation and cytokine-driven proliferation:
Attenuation of ICOS, IL-4, and IL-21, but not CD28, IL-7, and IL-15
responses. J Immunol. 170:711–718. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yuan Y, Wang L, Ge D, Tan L, Cao B, Fan H
and Xue L: Exosomal O-GlcNAc transferase from esophageal carcinoma
stem cell promotes cancer immunosuppression through up-regulation
of PD-1 in CD8(+) T cells. Cancer Lett. 500:98–106. 2021.
View Article : Google Scholar
|
|
35
|
Liu J, Fan L, Yu H, Zhang J, He Y, Feng D,
Wang F, Li X, Liu Q, Li Y, et al: Endoplasmic reticulum stress
causes liver cancer cells to release exosomal miR-23a-3p and
Up-regulate programmed death ligand 1 expression in macrophages.
Hepatology. 70:241–258. 2019.PubMed/NCBI
|
|
36
|
Ye L, Zhang Q, Cheng Y, Chen X, Wang G,
Shi M, Zhang T, Cao Y, Pan H, Zhang L, et al: Tumor-derived
exosomal HMGB1 fosters hepatocellular carcinoma immune evasion by
promoting TIM-1(+) regulatory B cell expansion. J Immunother
Cancer. 6:1452018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang Y, Yi J, Chen X, Zhang Y, Xu M and
Yang Z: The regulation of cancer cell migration by lung cancer
cell-derived exosomes through TGF-β and IL-10. Oncol Lett.
11:1527–1530. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Andreola G, Rivoltini L, Castelli C, Huber
V, Perego P, Deho P, Squarcina P, Accornero P, Lozupone F, Lugini
L, et al: Induction of lymphocyte apoptosis by tumor cell secretion
of FasL-bearing microvesicles. J Exp Med. 195:1303–1316. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Fuentes P, Sesé M, Guijar ro PJ, Emperador
M, Sánchez-Redondo S, Peinado H, Hümmer S and Cajal SRY: Publisher
Correction: ITGB3-mediated uptake of small extracellular vesicles
facilitates intercellular communication in breast cancer cells. Nat
Commun. 11:47302020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ji Q, Zhou L, Sui H, Yang L, Wu X, Song Q,
Jia R, Li R, Sun J, Wang Z, et al: Primary tumors release
ITGBL1-rich extracellular vesicles to promote distal metastatic
tumor growth through fibroblast-niche formation. Nat Commun.
11:12112020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Gabrusiewicz K, Li X, Wei J, Hashimoto Y,
Marisetty AL, Ott M, Wang F, Hawke D, Yu J, Healy LM, et al:
Glioblastoma stem cell-derived exosomes induce M2 macrophages and
PD-L1 expression on human monocytes. Oncoimmunology.
7:e14129092018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ferguson Bennit HR, Gonda A, Kabagwira J,
Oppegard L, Chi D, Licero Campbell J, De Leon M and Wall NR:
Natural killer cell phenotype and functionality affected by
exposure to extracellular survivin and lymphoma-derived exosomes.
Int J Mol Sci. 22:12552021. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kitai Y, Kawasaki T, Sueyoshi T, Kobiyama
K, Ishii KJ, Zou J, Akira S, Matsuda T and Kawai T: DNA-containing
exosomes derived from cancer cells treated with topotecan activate
a STING-Dependent pathway and reinforce antitumor immunity. J
Immunol. 198:1649–1659. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sun J, Jia H, Bao X, Wu Y, Zhu T, Li R and
Zhao H: Tumor exosome promotes Th17 cell differentiation by
transmitting the lncRNA CRNDE-h in colorectal cancer. Cell Death
Dis. 12:1232021. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ding G, Zhou L, Qian Y, Fu M, Chen J, Chen
J, Xiang J, Wu Z, Jiang G and Cao L: Pancreatic cancer-derived
exosomes transfer miRNAs to dendritic cells and inhibit RFXAP
expression via miR-212-3p. Oncotarget. 6:29877–29888. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhao S, Mi Y, Guan B, Zheng B, Wei P, Gu
Y, Zhang Z, Cai S, Xu Y, Li X, et al: Tumor-derived exosomal
miR-934 induces macrophage M2 polarization to promote liver
metastasis of colorectal cancer. J Hematol Oncol. 13:1562020.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wang X, Luo G, Zhang K, Cao J, Huang C,
Jiang T, Liu B, Su L and Qiu Z: Hypoxic tumor-derived exosomal
miR-301a Mediates M2 macrophage polarization via PTEN/PI3Kγ to
promote pancreatic cancer metastasis. Cancer Res. 78:4586–4598.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Chen X, Zhou J, Li X and Wang X, Lin Y and
Wang X: Exosomes derived from hypoxic epithelial ovarian cancer
cells deliver microRNAs to macrophages and elicit a tumor-promoted
phenotype. Cancer Lett. 435:80–91. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Hurley JH: ESCRTs are everywhere. EMBO J.
34:2398–2407. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
de Araujo Farias V, O'Valle F,
Serrano-Saenz S, Anderson P, Andrés E, López-Peñalver J, Tovar I,
Nieto A, Santos A, Martín F, et al: Exosomes derived from
mesenchymal stem cells enhance radiotherapy-induced cell death in
tumor and metastatic tumor foci. Mol Cancer. 17:1222018. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Stary V, Wolf B, Unterleuthner D, List J,
Talic M, Laengle J, Beer A, Strobl J, Stary G, Dolznig H and
Bergmann M: Short-course radiotherapy promotes pro-inflammatory
macrophages via extracellular vesicles in human rectal cancer. J
Immunother Cancer. 8:e0006672020. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ahn J, Xia T, Rabasa Capote A, Betancourt
D and Barber GN: Extrinsic phagocyte-dependent STING signaling
dictates the immunogenicity of dying cells. Cancer Cell.
33:862–873.e5. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Jiang MJ, Chen YY, Dai JJ, Gu DN, Mei Z,
Liu FR, Huang Q and Tian L: Dying tumor cell-derived exosomal
miR-194-5p potentiates survival and repopulation of tumor
repopulating cells upon radiotherapy in pancreatic cancer. Mol
Cancer. 19:682020. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Khambu B, Huda N, Chen X, Antoine DJ, Li
Y, Dai G, Köhler UA, Zong WX, Waguri S, Werner S, et al: HMGB1
promotes ductular reaction and tumorigenesis in autophagy-deficient
livers. J Clin Invest. 128:2419–2435. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Kazama H, Ricci JE, Herndon JM, Hoppe G,
Green DR and Ferguson TA: Induction of immunological tolerance by
apoptotic cells requires caspase-dependent oxidation of
high-mobility group box-1 protein. Immunity. 29:21–32. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Golden EB, Frances D, Pellicciotta I,
Demaria S, Helen Barcellos-Hoff M and Formenti SC: Radiation
fosters dose-dependent and chemotherapy-induced immunogenic cell
death. Oncoimmunology. 3:e285182014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Michaud M, Martins I, Sukkurwala AQ,
Adjemian S, Ma Y, Pellegatti P, Shen S, Kepp O, Scoazec M, Mignot
G, et al: Autophagy-dependent anticancer immune responses induced
by chemotherapeutic agents in mice. Science. 334:1573–1577. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Shi Y and Lammers T: Combining
nanomedicine and immunotherapy. Acc Chem Res. 52:1543–1554. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Lecciso M, Ocadlikova D, Sangaletti S,
Trabanelli S, De Marchi E, Orioli E, Pegoraro A, Portararo P,
Jandus C, Bontadini A, et al: ATP release from chemotherapy-treated
dying leukemia cells elicits an immune suppressive effect by
increasing regulatory T cells and tolerogenic dendritic cells.
Front Immunol. 8:19182017. View Article : Google Scholar
|
|
60
|
Yamaguchi H, Maruyama T, Urade Y and
Nagata S: Immunosuppression via adenosine receptor activation by
adenosine monophosphate released from apoptotic cells. Elife.
3:e021722014. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Allard D, Allard B and Stagg J: On the
mechanism of anti-CD39 immune checkpoint therapy. J Immunother
Cancer. 8:e0001862020. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Baghbani E, Noorolyai S, Shanehbandi D,
Mokhtarzadeh A, Aghebati-Maleki L, Shahgoli VK, Brunetti O, Rahmani
S, Shadbad MA, Baghbanzadeh A, et al: Regulation of immune
responses through CD39 and CD73 in cancer: Novel checkpoints. Life
Sci. 282:1198262021. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Batlle E and Massagué J: Transforming
growth factor-β signaling in immunity and cancer. Immunity.
50:924–940. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Formenti SC, Lee P, Adams S, Goldberg JD,
Li X, Xie MW, Ratikan JA, Felix C, Hwang L, Faull KF, et al: Focal
irradiation and systemic TGFβ blockade in metastatic breast cancer.
Clin Cancer Res. 24:2493–2504. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Karapetyan L, Luke JJ and Davar D:
Toll-Like Receptor 9 Agonists in Cancer. Onco Targets Ther.
13:10039–10060. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Vincent-Schneider H, Stumptner-Cuvelette
P, Lankar D, Pain S, Raposo G, Benaroch P and Bonnerot C: Exosomes
bearing HLA-DR1 molecules need dendritic cells to efficiently
stimulate specific T cells. Int Immunol. 14:713–722. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Xie F, Zhou X, Fang M, Li H, Su P, Tu Y,
Zhang L and Zhou F: Extracellular vesicles in cancer immune
microenvironment and cancer immunotherapy. Adv Sci (Weinh).
6:19017792019. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Pitt JM, André F, Amigorena S, Soria JC,
Eggermont A, Kroemer G and Zitvogel L: Dendritic cell-derived
exosomes for cancer therapy. J Clin Invest. 126:1224–1232. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Quah BJ and O'Neill HC: Maturation of
function in dendritic cells for tolerance and immunity. J Cell Mol
Med. 9:643–654. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Pang G, Chen C, Liu Y, Jiang T, Yu H, Wu
Y, Wang Y, Wang FJ, Liu Z and Zhang LW: Bioactive polysaccharide
nanoparticles improve radiation-induced abscopal effect through
manipulation of dendritic cells. ACS Appl Mater Interfaces.
11:42661–42670. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Accogli T, Bruchard M and Végran F:
Modulation of CD4 T cell response according to tumor cytokine
microenvironment. Cancers (Basel). 13:3732021. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Shiokawa A, Kotaki R, Takano T,
Nakajima-Adachi H and Hachimura S: Mesenteric lymph node CD11b(−)
CD103(+) PD-L1High dendritic cells highly induce
regulatory T cells. Immunology. 152:52–64. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Zhou F, Zhang GX and Rostami A: Distinct
Role of IL-27 in Immature and LPS-Induced mature dendritic
cell-mediated development of CD4(+) CD127(+)3G11(+) regulatory T
cell subset. Front Immunol. 9:25622018. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Shirasawa M, Yoshida T, Matsumoto Y,
Shinno Y, Okuma Y, Goto Y, Horinouchi H, Yamamoto N, Watanabe SI,
Ohe Y and Motoi N: Impact of chemoradiotherapy on the
immune-related tumour microenvironment and efficacy of anti-PD-(L)1
therapy for recurrences after chemoradiotherapy in patients with
unresectable locally advanced non-small cell lung cancer. Eur J
Cancer. 140:28–36. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Tang C, Wang X, Soh H, Seyedin S, Cortez
MA, Krishnan S, Massarelli E, Hong D, Naing A, Diab A, et al:
Combining radiation and immunotherapy: A new systemic therapy for
solid tumors? Cancer Immunol Res. 2:831–838. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Cassetta L and Pollard JW: Targeting
macrophages: Therapeutic approaches in cancer. Nat Rev Drug Discov.
17:887–904. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Wang J, Deng Z, Wang Z, Wu J, Gu T, Jiang
Y and Li G: MicroRNA-155 in exosomes secreted from helicobacter
pylori infection macrophages immunomodulates inflammatory response.
Am J Transl Res. 8:3700–3709. 2016.PubMed/NCBI
|
|
78
|
Wang P, Wang H, Huang Q, Peng C, Yao L,
Chen H, Qiu Z, Wu Y, Wang L and Chen W: Exosomes from M1-Polarized
macrophages enhance paclitaxel antitumor activity by activating
macrophages-mediated inflammation. Theranostics. 9:1714–1727. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Deng F, Yan J, Lu J, Luo M, Xia P, Liu S,
Wang X, Zhi F and Liu D: M2 Macrophage-Derived Exosomal miR-590-3p
Attenuates DSS-Induced mucosal damage and promotes epithelial
repair via the LATS1/YAP/β-Catenin Signalling Axis. J Crohns
Colitis. 15:665–677. 2021. View Article : Google Scholar
|
|
80
|
Wu J, Gao W, Tang Q, Yu Y, You W, Wu Z,
Fan Y, Zhang L, Wu C, Han G, et al: M2 Macrophage-derived exosomes
facilitate HCC metastasis by transferring αM
β2 integrin to tumor cells. Hepatology. 73:1365–1380.
2021. View Article : Google Scholar
|
|
81
|
Tavazoie MF, Pollack I, Tanqueco R,
Ostendorf BN, Reis BS, Gonsalves FC, Kurth I, Andreu-Agullo C,
Derbyshire ML, Posada J, et al: LXR/ApoE Activation restricts
innate immune suppression in cancer. Cell. 172:825–840.e18. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Zheng P, Chen L, Yuan X, Luo Q, Liu Y, Xie
G, Ma Y and Shen L: Exosomal transfer of tumor-associated
macrophage-derived miR-21 confers cisplatin resistance in gastric
cancer cells. J Exp Clin Cancer Res. 36:532017. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Hao Y, Yasmin-Karim S, Moreau M, Sinha N,
Sajo E and Ngwa W: Enhancing radiotherapy for lung cancer using
immunoadjuvants delivered in situ from new design radiotherapy
biomaterials: A preclinical study. Phys Med Biol. 61:N697–n707.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Qian M, Wang S, Guo X, Wang J, Zhang Z,
Qiu W, Gao X, Chen Z, Xu J, Zhao R, et al: Hypoxic glioma-derived
exosomes deliver microRNA-1246 to induce M2 macrophage polarization
by targeting TERF2IP via the STAT3 and NF-κB pathways. Oncogene.
39:428–442. 2020. View Article : Google Scholar
|
|
85
|
Kelly A, Gunaltay S, McEntee CP,
Shuttleworth EE, Smedley C, Houston SA, Fenton TM, Levison S, Mann
ER and Travis MA: Human monocytes and macrophages regulate immune
tolerance via integrin αvβ8-mediated TGFβ activation. J Exp Med.
215:2725–2736. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Veremeyko T, Yung AWY, Dukhinova M,
Kuznetsova IS, Pomytkin I, Lyundup A, Strekalova T, Barteneva NS
and Ponomarev ED: Cyclic AMP pathway suppress autoimmune
neuroinflammation by inhibiting functions of encephalitogenic CD4 T
cells and enhancing M2 macrophage polarization at the site of
inflammation. Front Immunol. 9:502018. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Su B, Han H, Gong Y, Li X, Ji C, Yao J,
Yang J, Hu W, Zhao W, Li J, et al: Let-7d inhibits intratumoral
macrophage M2 polarization and subsequent tumor angiogenesis by
targeting IL-13 and IL-10. Cancer Immunol Immunother. 70:1619–1634.
2021. View Article : Google Scholar
|
|
88
|
Ivashkiv LB: IFNγ: Signalling, epigenetics
and roles in immunity, metabolism, disease and cancer
immunotherapy. Nat Rev Immunol. 18:545–558. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Teresa Pinto A, Laranjeiro Pinto M,
Patrícia Cardoso A, Monteiro C, Teixeira Pinto M, Filipe Maia A,
Castro P, Figueira R, Monteiro A, Marques M, et al: Ionizing
radiation modulates human macrophages towards a pro-inflammatory
phenotype preserving their pro-invasive and pro-angiogenic
capacities. Sci Rep. 6:187652016. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Klug F, Prakash H, Huber PE, Seibel T,
Bender N, Halama N, Pfirschke C, Voss RH, Timke C, Umansky L, et
al: Low-dose irradiation programs macrophage differentiation to an
iNOS+/M1 phenotype that orchestrates effective T cell
immunotherapy. Cancer Cell. 24:589–602. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Leblond MM, Pérès EA, Helaine C, Gérault
AN, Moulin D, Anfray C, Divoux D, Petit E, Bernaudin M and Valable
S: M2 macrophages are more resistant than M1 macrophages following
radiation therapy in the context of glioblastoma. Oncotarget.
8:72597–72612. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Proctor DT, Huang J, Lama S, Albakr A, Van
Marle G and Sutherland GR: Tumor-associated macrophage infiltration
in meningioma. Neurooncol Adv. 1:vdz0182019.
|
|
93
|
Gordon SR, Maute RL, Dulken BW, Hutter G,
George BM, McCracken MN, Gupta R, Tsai JM, Sinha R, Corey D, et al:
PD-1 expression by tumour-associated macrophages inhibits
phagocytosis and tumour immunity. Nature. 545:495–499. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Zhao Y, Tao F, Jiang J, Chen L, Du J,
Cheng X, He Q, Zhong S, Chen W, Wu X, et al: Tryptophan 2,
3-dioxygenase promotes proliferation, migration and invasion of
ovarian cancer cells. Mol Med Rep. 23:4452021. View Article : Google Scholar :
|
|
95
|
Suek N, Campesato LF, Merghoub T and
Khalil DN: Targeted APC activation in cancer immunotherapy to
enhance the abscopal effect. Front Immunol. 10:6042019. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Zhou J, Li X, Wu X, Zhang T, Zhu Q and
Wang X, Wang H, Wang K, Lin Y and Wang X: Exosomes Released from
Tumor-Associated Macrophages Transfer miRNAs that induce a
Treg/Th17 cell imbalance in epithelial ovarian cancer. Cancer
Immunol Res. 6:1578–1592. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Jiang M, Liu X, Zhang D, Wang Y, Hu X, Xu
F, Jin M, Cao F and Xu L: Celastrol treatment protects against
acute ischemic stroke-induced brain injury by promoting an
IL-33/ST2 axis-mediated microglia/macrophage M2 polarization. J
Neuroinflammation. 15:782018. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Wang C, Zhang C, Liu L, A X, Chen B, Li Y
and Du J: Macrophage-Derived mir-155-Containing exosomes suppress
fibroblast proliferation and promote fibroblast inflammation during
cardiac injury. Mol Ther. 25:192–204. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Tan HY, Wang N, Zhang C, Chan YT, Yuen MF
and Feng Y: Lysyl Oxidase-Like 4 fosters an immunosuppressive
microenvironment during hepatocarcinogenesis. Hepatology.
73:2326–2341. 2021. View Article : Google Scholar
|
|
100
|
Chen X, Zhang L, Zhang IY, Liang J, Wang
H, Ouyang M, Wu S, da Fonseca ACC, Weng L, Yamamoto Y, et al: RAGE
expression in tumor-associated macrophages promotes angiogenesis in
glioma. Cancer Res. 74:7285–7297. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Barnes TA and Amir E: HYPE or HOPE: The
prognostic value of infiltrating immune cells in cancer. Br J
Cancer. 118:e52018. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Wang X, Shen H, Zhangyuan G, Huang R,
Zhang W, He Q, Jin K, Zhuo H, Zhang Z, Wang J, et al: 14-3-3ζ
delivered by hepatocellular carcinoma-derived exosomes impaired
anti-tumor function of tumor-infiltrating T lymphocytes. Cell Death
Dis. 9:1592018. View Article : Google Scholar
|
|
103
|
Golstein P and Griffiths GM: An early
history of T cell-mediated cytotoxicity. Nat Rev Immunol.
18:527–535. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Alonso R, Rodríguez MC, Pindado J, Merino
E, Mérida I and Izquierdo M: Diacylglycerol kinase alpha regulates
the secretion of lethal exosomes bearing Fas ligand during
activation-induced cell death of T lymphocytes. J Biol Chem.
280:28439–28450. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Cai Z, Yang F, Yu L, Yu Z, Jiang L, Wang
Q, Yang Y, Wang L, Cao X and Wang J: Activated T cell exosomes
promote tumor invasion via Fas signaling pathway. J Immunol.
188:5954–5961. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Lugini L, Cecchetti S, Huber V, Luciani F,
Macchia G, Spadaro F, Paris L, Abalsamo L, Colone M, Molinari A, et
al: Immune surveillance properties of human NK cell-derived
exosomes. J Immunol. 189:2833–2842. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
van der Bruggen P, Traversari C, Chomez P,
Lurquin C, De Plaen E, Van den Eynde BJ, Knuth A and Boon T: A gene
encoding an antigen recognized by cytolytic T lymphocytes on a
human melanoma. J Immunol. 178:2617–2621. 2007.PubMed/NCBI
|
|
108
|
Torralba D, Baixauli F, Villarroya-Beltri
C, Fernández-Delgado I, Latorre-Pellicer A, Acín-Pérez R,
Martín-Cófreces NB, Jaso-Tamame ÁL, Iborra S, Jorge I, et al:
Priming of dendritic cells by DNA-containing extracellular vesicles
from activated T cells through antigen-driven contacts. Nat Commun.
9:26582018. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Wang X, Shen H, He Q, Tian W, Xia A and Lu
XJ: Exosomes derived from exhausted CD8+ T cells impaired the
anticancer function of normal CD8+ T cells. J Med Genet. 56:29–31.
2019. View Article : Google Scholar
|
|
110
|
Hui E, Cheung J, Zhu J, Su X, Taylor MJ,
Wallweber HA, Sasmal DK, Huang J, Kim JM, Mellman I and Vale RD: T
cell costimulatory receptor CD28 is a primary target for
PD-1-mediated inhibition. Science. 355:1428–1433. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Anderson AC, Joller N and Kuchroo VK:
Lag-3, Tim-3, and TIGIT: Co-inhibitory receptors with specialized
functions in immune regulation. Immunity. 44:989–1004. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Dimitrijević M, Arsenović-Ranin N, Kosec
D, Bufan B, Nacka-Aleksić M, Pilipović I and Leposavić G: Sexual
dimorphism in Th17/Treg axis in lymph nodes draining inflamed
joints in rats with collagen-induced arthritis. Brain Behav Immun.
76:198–214. 2019. View Article : Google Scholar
|
|
113
|
Asano T, Meguri Y, Yoshioka T, Kishi Y,
Iwamoto M, Nakamura M, Sando Y, Yagita H, Koreth J, Kim HT, et al:
PD-1 modulates regulatory T-cell homeostasis during low-dose
interleukin-2 therapy. Blood. 129:2186–2197. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Abbas AK, Trotta E, R Simeonov D, Marson A
and Bluestone JA: Revisiting IL-2: Biology and therapeutic
prospects. Sci Immunol. 3:eaat14822018. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Jukić T, Jurin Martić A, Ivanković S,
Antica M, Pavan Jukić D, Rotim C and Jurin M: The role of
regulatory T lymphocytes in immune control of MC-2 fibrosarcoma.
Acta Clin Croat. 59:351–358. 2020.
|
|
116
|
Mailloux AW and Young MR: Regulatory
T-cell trafficking: From thymic development to tumor-induced immune
suppression. Crit Rev Immunol. 30:435–447. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Hammami A, Allard D, Allard B and Stagg J:
Targeting the adenosine pathway for cancer immunotherapy. Semin
Immunol. 42:1013042019. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Smyth LA, Ratnasothy K, Tsang JY, Boardman
D, Warley A, Lechler R and Lombardi G: CD73 expression on
extracellular vesicles derived from CD4+ CD25+ Foxp3+ T cells
contributes to their regulatory function. Eur J Immunol.
43:2430–2440. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Aiello S, Rocchetta F, Longaretti L,
Faravelli S, Todeschini M, Cassis L, Pezzuto F, Tomasoni S,
Azzollini N, Mister M, et al: Extracellular vesicles derived from T
regulatory cells suppress T cell proliferation and prolong
allograft survival. Sci Rep. 7:115182017. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Tung SL, Fanelli G, Matthews RI, Bazoer J,
Letizia M, Vizcay-Barrena G, Faruqu FN, Philippeos C, Hannen R,
Al-Jamal KT, et al: Regulatory T cell extracellular vesicles modify
t-effector cell cytokine production and protect against human skin
allograft damage. Front Cell Dev Biol. 8:3172020. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Torri A, Carpi D, Bulgheroni E, Crosti MC,
Moro M, Gruarin P, Rossi RL, Rossetti G, Di Vizio D, Hoxha M, et
al: Extracellular MicroRNA signature of human helper T cell subsets
in health and autoimmunity. J Biol Chem. 292:2903–2915. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Tung SL, Boardman DA, Sen M, Letizia M,
Peng Q, Cianci N, Dioni L, Carlin LM, Lechler R, Bollati V, et al:
Regulatory T cell-derived extracellular vesicles modify dendritic
cell function. Sci Rep. 8:60652018. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Okoye IS, Coomes SM, Pelly VS, Czieso S,
Papayannopoulos V, Tolmachova T, Seabra MC and Wilson MS:
MicroRNA-containing T-regulatory-cell-derived exosomes suppress
pathogenic T helper 1 cells. Immunity. 41:89–103. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Sullivan JA, Tomita Y, Jankowska-Gan E,
Lema DA, Arvedson MP, Nair A, Bracamonte-Baran W, Zhou Y, Meyer KK,
Zhong W, et al: Treg-Cell-Derived IL-35-Coated extracellular
vesicles promote infectious tolerance. Cell Rep. 30:1039–1051.e5.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Eckert F, Schilbach K, Klumpp L, Bardoscia
L, Sezgin EC, Schwab M, Zips D and Huber SM: Potential Role of
CXCR4 targeting in the context of radiotherapy and immunotherapy of
cancer. Front Immunol. 9:30182018. View Article : Google Scholar
|
|
126
|
Zhou M, Luo C, Zhou Z, Li L and Huang Y:
Improving anti-PD-L1 therapy in triple negative breast cancer by
polymer-enhanced immunogenic cell death and CXCR4 blockade. J
Control Release. 334:248–262. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Kohno M, Murakami J, Wu L, Chan ML, Yun Z,
Cho BCJ and de Perrot M: Foxp3(+) Regulatory T cell depletion after
nonablative oligofractionated irradiation boosts the abscopal
effects in murine malignant mesothelioma. J Immunol. 205:2519–2531.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Anderson BE, McNiff JM, Matte C,
Athanasiadis I, Shlomchik WD and Shlomchik MJ: Recipient CD4+ T
cells that survive irradiation regulate chronic graft-versus-host
disease. Blood. 104:1565–1573. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Zhang T, Yu H, Ni C, Zhang T, Liu L, Lv Q,
Zhang Z, Wang Z, Wu D, Wu P, et al: Hypofractionated stereotactic
radiation therapy activates the peripheral immune response in
operable stage I non-small-cell lung cancer. Sci Rep. 7:48662017.
View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Harrington LE, Hatton RD, Mangan PR,
Turner H, Murphy TL, Murphy KM and Weaver CT: Interleukin
17-producing CD4+ effector T cells develop via a lineage distinct
from the T helper type 1 and 2 lineages. Nat Immunol. 6:1123–1132.
2005. View
Article : Google Scholar : PubMed/NCBI
|
|
131
|
Ye ZJ, Zhou Q, Gu YY, Qin SM, Ma WL, Xin
JB, Tao XN and Shi HZ: Generation and differentiation of
IL-17-producing CD4+ T cells in malignant pleural effusion. J
Immunol. 185:6348–6354. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Zou W and Restifo NP: T(H)17 cells in
tumour immunity and immunotherapy. Nat Rev Immunol. 10:248–256.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Berkhout L, Barikbin R, Schiller B,
Ravichandran G, Krech T, Neumann K, Sass G and Tiegs G: Deletion of
tumour necrosis factor α receptor 1 elicits an increased TH17
immune response in the chronically inflamed liver. Sci Rep.
9:42322019. View Article : Google Scholar
|
|
134
|
Campanati A, Orciani M, Lazzarini R,
Ganzetti G, Consales V, Sorgentoni G, Di Primio R and Offidani A:
TNF-α inhibitors reduce the pathological
Th1-Th17/Th2 imbalance in
cutaneous mesenchymal stem cells of psoriasis patients. Exp
Dermatol. 26:319–324. 2017. View Article : Google Scholar
|
|
135
|
Nalbant A: IL-17, IL-21, and IL-22
Cytokines of T Helper 17 cells in cancer. J Interferon Cytokine
Res. 39:56–60. 2019. View Article : Google Scholar
|
|
136
|
Chang SH: T helper 17 (Th17) cells and
interleukin-17 (IL-17) in cancer. Arch Pharm Res. 42:549–559. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Papadopoulou E, Nicolatou-Galitis O,
Papassotiriou I, Linardou H, Karagianni A, Tsixlakis K, Tarampikou
A, Michalakakou K, Vardas E and Bafaloukos D: The use of crevicular
fluid to assess markers of inflammation and angiogenesis, IL-17 and
VEGF, in patients with solid tumors receiving zoledronic acid
and/or bevacizumab. Support Care Cancer. 28:177–184. 2020.
View Article : Google Scholar
|
|
138
|
Messmer MN, Netherby CS, Banik D and
Abrams SI: Tumor-induced myeloid dysfunction and its implications
for cancer immunotherapy. Cancer Immunol Immunother. 64:1–13. 2015.
View Article : Google Scholar :
|
|
139
|
Waigel S, Rendon BE, Lamont G, Richie J,
Mitchell RA and Yaddanapudi K: MIF inhibition reverts the gene
expression profile of human melanoma cell line-induced MDSCs to
normal monocytes. Genom Data. 7:240–242. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Fenselau C and Ostrand-Rosenberg S:
Molecular cargo in myeloid-derived suppressor cells and their
exosomes. Cell Immunol. 359:1042582021. View Article : Google Scholar :
|
|
141
|
Marvel D and Gabrilovich DI:
Myeloid-derived suppressor cells in the tumor microenvironment:
Expect the unexpected. J Clin Invest. 125:3356–3364. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Gabrilovich DI, Chen HL, Girgis KR,
Cunningham HT, Meny GM, Nadaf S, Kavanaugh D and Carbone DP:
Production of vascular endothelial growth factor by human tumors
inhibits the functional maturation of dendritic cells. Nat Med.
2:1096–1103. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Horikawa N, Abiko K, Matsumura N,
Hamanishi J, Baba T, Yamaguchi K, Yoshioka Y, Koshiyama M and
Konishi I: Expression of vascular endothelial growth factor in
ovarian cancer inhibits tumor immunity through the accumulation of
myeloid-derived suppressor cells. Clin Cancer Res. 23:587–599.
2017. View Article : Google Scholar
|
|
144
|
Hsieh CC, Hung CH, Chiang M, Tsai YC and
He JT: Hepatic stellate cells enhance liver cancer progression by
inducing myeloid-derived suppressor cells through interleukin-6
signaling. Int J Mol Sci. 20:50792019. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Li L, Zhang J, Diao W, Wang D, Wei Y,
Zhang CY and Zen K: MicroRNA-155 and MicroRNA-21 promote the
expansion of functional myeloid-derived suppressor cells. J
Immunol. 192:1034–1043. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Meng G, Wei J, Wang Y, Qu D and Zhang J:
miR-21 regulates immunosuppression mediated by myeloid-derived
suppressor cells by impairing RUNX1-YAP interaction in lung cancer.
Cancer Cell Int. 20:4952020. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Safarzadeh E, Asadzadeh Z, Safaei S,
Hatefi A, Derakhshani A, Giovannelli F, Brunetti O, Silvestris N
and Baradaran B: MicroRNAs and lncRNAs-A new layer of
myeloid-derived suppressor cells regulation. Front Immunol.
11:5723232020. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Geis-Asteggiante L, Belew AT, Clements VK,
Edwards NJ, Ostrand-Rosenberg S, El-Sayed NM and Fenselau C:
Differential Content of Proteins, mRNAs, and miRNAs Suggests that
MDSC and their exosomes may mediate distinct immune suppressive
functions. J Proteome Res. 17:486–498. 2018. View Article : Google Scholar :
|
|
149
|
Li T, Li M, Xu C, Xu X, Ding J, Cheng L
and Ou R: miR-146a regulates the function of Th17 cell
differentiation to modulate cervical cancer cell growth and
apoptosis through NF-κB signaling by targeting TRAF6. Oncol Rep.
41:2897–2908. 2019.PubMed/NCBI
|
|
150
|
Tian S, Song X, Wang Y, Wang X, Mou Y,
Chen Q, Zhao H, Ma K, Wu Z, Yu H, et al: Chinese herbal medicine
Baoyuan Jiedu decoction inhibits the accumulation of myeloid
derived suppressor cells in pre-metastatic niche of lung via
TGF-β/CCL9 pathway. Biomed Pharmacother. 129:1103802020. View Article : Google Scholar
|
|
151
|
Mao Y, Sarhan D, Steven A, Seliger B,
Kiessling R and Lundqvist A: Inhibition of tumor-derived
prostaglandin-e2 blocks the induction of myeloid-derived suppressor
cells and recovers natural killer cell activity. Clin Cancer Res.
20:4096–4106. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Deng Z, Rong Y, Teng Y, Zhuang X,
Samykutty A, Mu J, Zhang L, Cao P, Yan J, Miller D and Zhang HG:
Exosomes miR-126a released from MDSC induced by DOX treatment
promotes lung metastasis. Oncogene. 36:639–651. 2017. View Article : Google Scholar :
|
|
153
|
Zöller M, Zhao K, Kutlu N, Bauer N,
Provaznik J, Hackert T and Schnölzer M: Immunoregulatory effects of
myeloid-derived suppressor cell exosomes in mouse model of
autoimmune alopecia areata. Front Immunol. 9:12792018. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Grisaru-Tal S, Itan M, Klion AD and Munitz
A: A new dawn for eosinophils in the tumour microenvironment. Nat
Rev Cancer. 20:594–607. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Poggio M, Hu T, Pai CC, Chu B, Belair CD,
Chang A, Montabana E, Lang UE, Fu Q, Fong L and Blelloch R:
Suppression of Exosomal PD-L1 induces systemic anti-tumor immunity
and memory. Cell. 177:414–427.e13. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Mesnil C, Raulier S, Paulissen G, Xiao X,
Birrell MA, Pirottin D, Janss T, Starkl P, Ramery E, Henket M, et
al: Lung-resident eosinophils represent a distinct regulatory
eosinophil subset. J Clin Invest. 126:3279–3295. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Choi Y, Kim YM, Lee HR, Mun J, Sim S, Lee
DH, Pham DL, Kim SH, Shin YS, Lee SW and Park HS: Eosinophil
extracellular traps activate type 2 innate lymphoid cells through
stimulating airway epithelium in severe asthma. Allergy. 75:95–103.
2020. View Article : Google Scholar
|
|
158
|
da Silva JM, Moreira Dos Santos TP, Sobral
LM, Queiroz-Junior CM, Rachid MA, Proudfoot AEI, Garlet GP, Batista
AC, Teixeira MM, Leopoldino AM, et al: Relevance of CCL3/CCR5 axis
in oral carcinogenesis. Oncotarget. 8:51024–51036. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Hu Y, Chen Z, Jin L, Wang M and Liao W:
Decreased expression of indolamine 2,3-dioxygenase in childhood
allergic asthma and its inverse correlation with fractional
concentration of exhaled nitric oxide. Ann Allergy Asthma Immunol.
119:429–434. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Kratochvill F, Neale G, Haverkamp JM, Van
de Velde LA, Smith AM, Kawauchi D, McEvoy J, Roussel MF, Dyer MA,
Qualls JE and Murray PJ: TNF counterbalances the emergence of M2
tumor macrophages. Cell Rep. 12:1902–1914. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Herrera FG, Bourhis J and Coukos G:
Radiotherapy combination opportunities leveraging immunity for the
next oncology practice. CA Cancer J Clin. 67:65–85. 2017.
View Article : Google Scholar
|
|
162
|
Cao M, Cabrera R, Xu Y, Liu C and Nelson
D: Different radio-sensitivity of CD4(+)CD25(+) regulatory T cells
and effector T cells to low dose gamma irradiation in vitro. Int J
Radiat Biol. 87:71–80. 2011. View Article : Google Scholar
|
|
163
|
Kareff SA, Lischalk JW, Krochmal R and Kim
C: Abscopal effect in pulmonary carcinoid tumor following ablative
stereotactic body radiation therapy: A case report. J Med Case Rep.
14:1772020. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Ohmatsu K, Hashimoto Y, Kawanishi M, Ishii
Y, Kono S, Kuribayashi S, Ariizumi S and Karasawa K: Abscopal
complete regression of hepatocellular carcinoma with multiple
pleural metastases. Int Cancer Conf J. 10:54–58. 2020. View Article : Google Scholar
|
|
165
|
Hotta T, Okuno T, Nakao M, Amano Y, Isobe
T and Tsubata Y: Reproducible abscopal effect in a patient with
lung cancer who underwent whole-brain irradiation and atezolizumab
administration. Thorac Cancer. 12:985–988. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Choi JS, Sansoni ER, Lovin BD, Lindquist
NR, Phan J, Mayo LL, Ferrarotto R and Su SY: Abscopal effect
following immunotherapy and combined stereotactic body radiation
therapy in recurrent metastatic head and neck squamous cell
carcinoma: A report of two cases and literature review. Ann Otol
Rhinol Laryngol. 129:517–522. 2020. View Article : Google Scholar
|
|
167
|
Wang H, Lin X, Luo Y, Sun S, Tian X, Sun
Y, Zhang S, Chen J, Zhang J, Liu X, et al: α-PD-L1 mAb enhances the
abscopal effect of hypo-fractionated radiation by attenuating PD-L1
expression and inducing CD8(+) T-cell infiltration. Immunotherapy.
11:101–118. 2019. View Article : Google Scholar
|
|
168
|
Golden EB, Chhabra A, Chachoua A, Adams S,
Donach M, Fenton-Kerimian M, Friedman K, Ponzo F, Babb JS, Goldberg
J, et al: Local radiotherapy and granulocyte-macrophage
colony-stimulating factor to generate abscopal responses in
patients with metastatic solid tumours: A proof-of-principle trial.
Lancet Oncol. 16:795–803. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Thorne AH, Malo KN, Wong AJ, Nguyen TT,
Cooch N, Reed C, Yan J, Broderick KE, Smith TRF, Masteller EL and
Humeau L: Adjuvant screen identifies synthetic DNA-Encoding Flt3L
and CD80 immunotherapeutics as candidates for enhancing anti-tumor
T cell responses. Front Immunol. 11:3272020. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Wennerberg E, Spada S, Rudqvist NP,
Lhuillier C, Gruber S, Chen Q, Zhang F, Zhou XK, Gross SS, Formenti
SC and Demaria S: CD73 blockade promotes dendritic cell
infiltration of irradiated tumors and tumor rejection. Cancer
Immunol Res. 8:465–478. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Peluso MO, Adam A, Armet CM, Zhang L,
O'Connor RW, Lee BH, Lake AC, Normant E, Chappel SC, Hill JA, et
al: The Fully human anti-CD47 antibody SRF231 exerts dual-mechanism
antitumor activity via engagement of the activating receptor CD32a.
J Immunother Cancer. 8:e0004132020. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Tsukui H, Horie H, Koinuma K, Ohzawa H,
Sakuma Y, Hosoya Y, Yamaguchi H, Yoshimura K, Lefor AK, Sata N and
Kitayama J: CD73 blockade enhances the local and abscopal effects
of radiotherapy in a murine rectal cancer model. BMC Cancer.
20:4112020. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Puro RJ, Bouchlaka MN, Hiebsch RR,
Capoccia BJ, Donio MJ, Manning PT, Frazier WA, Karr RW and Pereira
DS: Development of AO-176, a Next-Generation Humanized Anti-CD47
antibody with novel anticancer properties and negligible red blood
cell binding. Mol Cancer Ther. 19:835–846. 2020. View Article : Google Scholar
|
|
174
|
Chen D, Barsoumian HB, Yang L, Younes AI,
Verma V, Hu Y, Menon H, Wasley M, Masropour F, Mosaffa S, et al:
SHP-2 and PD-L1 inhibition combined with radiotherapy enhances
systemic antitumor effects in an Anti-PD-1-Resistant model of
non-small cell lung cancer. Cancer Immunol Res. 8:883–894. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Muenkel J, Xu Z, Traughber BJ, Baig T, Xu
K, Langmack C, Harris E and Podder TK: Feasibility of improving
patient's safety with in vivo dose tracking in intracavitary and
interstitial HDR brachytherapy. Brachytherapy. 20:353–360. 2021.
View Article : Google Scholar
|
|
176
|
Liu Y, Dong Y, Kong L, Shi F, Zhu H and Yu
J: Abscopal effect of radiotherapy combined with immune checkpoint
inhibitors. J Hematol Oncol. 11:1042018. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Ahmed TA, Adamopoulos C, Karoulia Z, Wu X,
Sachidanandam R, Aaronson SA and Poulikakos PI: SHP2 drives
adaptive resistance to ERK signaling inhibition in molecularly
defined subsets of ERK-Dependent tumors. Cell Rep. 26:65–78.e5.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Strazza M, Adam K, Lerrer S, Straube J,
Sandigursky S, Ueberheide B and Mor A: SHP2 Targets ITK Downstream
of PD-1 to Inhibit T cell function. Inflammation. 44:1529–1539.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Pustylnikov S, Costabile F, Beghi S and
Facciabene A: Targeting mitochondria in cancer: Current concepts
and immunotherapy approaches. Transl Res. 202:35–51. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Chen D, Barsoumian HB, Fischer G, Yang L,
Verma V, Younes AI, Hu Y, Masropour F, Klein K, Vellano C, et al:
Combination treatment with radiotherapy and a novel oxidative
phosphorylation inhibitor overcomes PD-1 resistance and enhances
antitumor immunity. J Immunother Cancer. 8:e0002892020. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Vanpouille-Box C, Diamond JM, Pilones KA,
Zavadil J, Babb JS, Formenti SC, Barcellos-Hoff MH and Demaria S:
TGFβ Is a master regulator of radiation therapy-induced antitumor
immunity. Cancer Res. 75:2232–2242. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Liang Y, He J and Zhao Y: Modification of
oncolytic adenovirus and its application in cancer therapy. Discov
Med. 30:129–144. 2020.
|
|
183
|
Havunen R, Santos JM, Sorsa S, Rantapero
T, Lumen D, Siurala M, Airaksinen AJ, Cervera-Carrascon V, Tähtinen
S, Kanerva A and Hemminki A: Abscopal effect in Non-injected tumors
achieved with cytokine-armed oncolytic adenovirus. Mol Ther
Oncolytics. 11:109–121. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Kaufman HL, Amatruda T, Reid T, Gonzalez
R, Glaspy J, Whitman E, Harrington K, Nemunaitis J, Zloza A, Wolf M
and Senzer NN: Systemic versus local responses in melanoma patients
treated with talimogene laherparepvec from a multi-institutional
phase II study. J Immunother Cancer. 4:122016. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Ono R, Takayama K, Sakurai F and Mizuguchi
H: Efficient antitumor effects of a novel oncolytic adenovirus
fully composed of species B adenovirus serotype 35. Mol Ther
Oncolytics. 20:399–409. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Challenor S and Tucker D:
SARS-CoV-2-induced remission of Hodgkin lymphoma. Br J Haematol.
192:4152021. View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Ngwa W, Boateng F, Kumar R, Irvine DJ,
Formenti S, Ngoma T, Herskind C, Veldwijk MR, Hildenbrand GL,
Hausmann M, et al: Smart Radiation Therapy Biomaterials. Int J
Radiat Oncol Biol Phys. 97:624–637. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
188
|
Zhao X, Yang K, Zhao R, Ji T, Wang X, Yang
X, Zhang Y, Cheng K, Liu S, Hao J, et al: Inducing enhanced
immunogenic cell death with nanocarrier-based drug delivery systems
for pancreatic cancer therapy. Biomaterials. 102:187–197. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Duan X, Chan C, Guo N, Han W, Weichselbaum
RR and Lin W: Photodynamic therapy mediated by nontoxic core-shell
nanoparticles synergizes with immune checkpoint blockade to elicit
antitumor immunity and antimetastatic effect on breast cancer. J Am
Chem Soc. 138:16686–16695. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Chen Q, Wang C, Zhang X, Chen G, Hu Q, Li
H, Wang J, Wen D, Zhang Y, Lu Y, et al: In situ sprayed
bioresponsive immunotherapeutic gel for post-surgical cancer
treatment. Nat Nanotechnol. 14:89–97. 2019. View Article : Google Scholar
|
|
191
|
Min Y, Roche KC, Tian S, Eblan MJ,
McKinnon KP, Caster JM, Chai S, Herring LE, Zhang L, Zhang T, et
al: Antigen-capturing nanoparticles improve the abscopal effect and
cancer immunotherapy. Nat Nanotechnol. 12:877–882. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Zheng Y, Tang L, Mabardi L, Kumari S and
Irvine DJ: Enhancing adoptive cell therapy of cancer through
targeted delivery of small-molecule immunomodulators to
internalizing or noninternalizing receptors. ACS Nano.
11:3089–3100. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Chao Y, Xu L, Liang C, Feng L, Xu J, Dong
Z, Tian L, Yi X, Yang K and Liu Z: Combined local immunostimulatory
radioisotope therapy and systemic immune checkpoint blockade
imparts potent antitumour responses. Nat Biomed Eng. 2:611–621.
2018. View Article : Google Scholar
|
|
194
|
Xu H, Sun W, Kong Y, Huang Y, Wei Z, Zhang
L, Liang J and Ye X: Immune abscopal effect of microwave ablation
for lung metastases of endometrial carcinoma. J Cancer Res Ther.
16:1718–1721. 2020.
|