Tumors exist in constant interaction with components
of the tumor microenvironment (TME), which includes stromal cells
[such as cancer-associated fibroblasts (CAFs), endothelial cells
(ECs) and adipocytes], and immune cells [such as tumor-associated
macrophages (TAMs), cancer-associated neutrophils, dendritic cells
(DCs), natural killer (NK) cells, T lymphocytes and B lymphocytes],
as well as extracellular matrix (ECM) (1,2).
Cells in the TME are involved in reciprocal interaction with tumor
cells, which eventually leads to enhanced proliferation, metastasis
and chemoresistance (1,3,4).
TAMs are key innate immune cells involved in
regulation of primary tumor growth, antitumor adaptive immune
response, tumor angiogenesis, ECM remodeling and metastasis
(5,6). TAMs originate either from circulating
monocytes derived from bone marrow hematopoietic stem cells
(recruited) or yolk sac progenitors (resident) (7). These cells are attracted into the
primary tumor and activated by various signals in the TME (6).
TAMs are classified as M1 (classically activated
macrophages) and M2 (alternatively activated macrophages) (5). M1 TAMs secrete pro-inflammatory
cytokines IL-12, TNFα, C-X-C motif chemokine ligand (CXCL)-10 and
IFN-γ, and produce high levels of nitric oxide synthase (NOS),
while M2 TAMs secrete anti-inflammatory cytokines IL-10, IL-4 and
IL-13, and express scavenger receptors, including CD163, CD206,
CD204, macrophage receptor with collagenous structure (MARCO) and
stabilin-1 (5,6). TAMs are plastic cells that can change
their polarization from M1 to M2 and vice versa in response to
various stimuli from the TME, including cytokines, growth factors
and matrix remodeling proteins (8,9).
Cells in the TME are genetically more stable compared with tumor
cells, which makes them useful as therapeutic targets (10,11).
In the present review, data on the main mechanisms
of the TAM interaction with crucial components of the TME,
including CAFs, blood and lymphatic ECs (LECs), and diverse immune
cell infiltrates, were collected. It also discusses possible
therapeutic approaches that combine targeting of TAMs and other
stromal or immune cells.
CAFs are heterogenous stromal cells originating from
different precursor cells, including resident tissue fibroblasts
(via activation), bone marrow mesenchymal stem cells (via
differentiation), hematopoietic stem cells (via differentiation),
epithelial cells via epithelial-mesenchymal transition (EMT) and
ECs via endothelial-mesenchymal transition (EndoMT) (12,13).
Tumor cells activate EndoMT through production of cytokines and
growth factors, among which TGF-α and TGF-β are the most studied
(14,15). In some tumors, such as melanoma,
pancreatic cancer and lung cancer, EndoMT provides 30-40% of CAFs
(16,17).
In the TME, CAFs possess functions critical for
supporting cancer cell proliferation, angiogenesis,
lymphangiogenesis, invasion and metastasis. CAFs promote tumor
progression via various mechanisms, including cytokine and
chemokine secretion, as well as ECM remodeling that occurs due to
basement membrane degradation and interstitial stroma
disintegration (18-26).
Numerous studies have demonstrated that CAFs
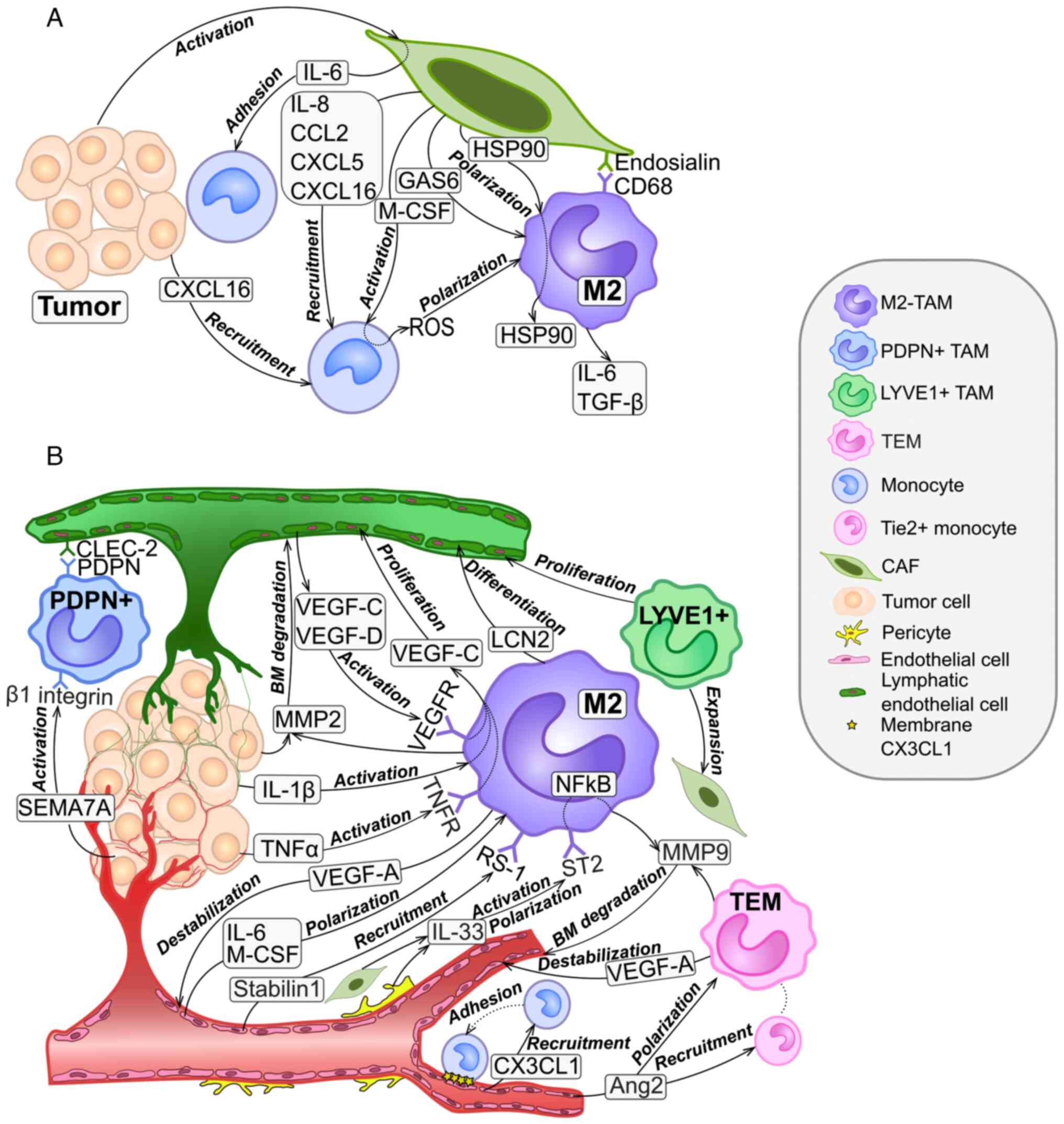
contribute to monocyte recruitment (Fig. 1A) (27-29).
In a Transwell migration assay, conditioned medium (CM) of colon
cancer-derived CAFs was more efficient in attracting monocytes
compared with CM of macrophages, differentiated from peripheral
blood mononuclear cells, or human HCT116 and DLD-1 colon cancer
cells (27). There are two key
factors promoting monocyte recruitment: Macrophage
colony-stimulating factor [M-CSF or colony stimulating factor 1
(CSF1)] and monocyte chemoattractant protein 1 [MCP-1; or C-C motif
chemokine ligand (CCL2)] (27,30,31).
Co-culture of CAFs with monocyte-derived macrophages (MDMs)
revealed high expression of CCL2 in MDMs, indicating CAFs to be one
of the major inducers of CCL2 production in macrophages (27). M-CSF is mainly produced by CAFs but
not by tumor cells or macrophages (27). Inhibition of M-CSF/M-CSF receptor
(M-CSFR) signaling results in decreased CCL2, CD163 and M-CSFR
expression (27). The role of
M-CSF in myeloid cell survival, differentiation and function has
been demonstrated in other studies (32,33).
IL-8 produced by CAFs derived from human colorectal cancer (CRC)
tissue has been reported to recruit monocytes, while IL-6 promoted
their adhesion to the tumor cells (28). CM from CAFs induced vascular cell
adhesion molecule 1 (VCAM-1) expression in CRC cells, which is
associated with enriched infiltration of TAMs in the TME (28). A Transwell assay revealed that
CAF-derived C-X-C motif chemokine receptor (CXCR)2 and IL-8
facilitated monocyte migration in a dose-dependent manner (28).
THP-1 human monocytes co-cultured with human oral
squamous cell carcinoma (OSCC)-derived CAFs acquired the M2
phenotype, which manifested in elevated expression levels of M2
markers IL-10 and TGFβ and increased adhesion, compared with
control propidium monoazide-treated macrophages (34). Triple co-culture of THP-1
monocytes, CAFs and OSCC cells revealed a more pronounced effect on
monocyte differentiation and further macrophage polarization
(34). Furthermore, tumor cells
from colon, breast and oral cancer stimulate CAFs to produce higher
levels of IL-6, CXCL5 and granulocyte M-CSF (GM-CSF/CSF-2)
(34). CAFs isolated from human
lung squamous cell carcinoma expressed monocyte- and
neutrophil-attracting chemokines, including CCL2, CCL7, CXCL1,
CXCL5 and CXCL8 (35). CAFs in
co-culture with macrophages exhibited higher expression levels of
M-CSF, IL-6, IL-8, hepatocyte growth factor and CCL2 (36). In this in vitro system,
macrophages were differentiated from peripheral blood monocytes of
healthy donors, and CAFs were obtained by mesenchymal stem cell
co-cultivation with esophageal squamous cell carcinoma cell lines
(36). CAFs obtained from human
breast cancer (BC) tissue induced the recruitment of monocytes via
CCL2 and stromal cell-derived factor-1 (SDF-1), and promoted M2
polarization (37).
In human hepatocellular carcinoma (HCC), CAFs
express endosialin, a transmembrane glycoprotein that is also
expressed in tumor cells (38).
Endosialin can interact with CD68, thereby recruiting macrophages
into the TME (38).
Endosialin-positive CAFs can promote M2 polarization of macrophages
through growth arrest specific 6 factor secretion (Fig. 1A) (38). In the TME, CAFs are the main
producers of CXCL12 and CXCL16, which are monocyte chemoattractants
that induce M2 polarization (29).
In TAMs differentiated from the THP-1 cell line, CXCL12 stimulated
the secretion of plasminogen activator inhibitor-1, inducing tumor
cell proliferation, angiogenesis, metastasis, EMT, cell apoptosis
inhibition and drug resistance (29). Immunohistochemistry (IHC) analysis
of primary triple-negative BC (TNBC) revealed that both tumor cells
and CAFs expressed CXCL16 (39).
CM of CAFs isolated from human tumors affected the migration of
human peripheral blood monocytes and, to a lesser extent, M2
macrophages (39). Monocyte
migration towards CXCL16 was more pronounced compared with that
towards CXCL12 (39). In
vitro, CAFs isolated from human prostate carcinoma tissue
induced the recruitment of macrophages, obtained from healthy donor
monocytes, towards tumor cells and macrophage M2 polarization via
SDF-1 (40). Other factors,
including CCL2 and CXCL14, mainly affect myeloid cell recruitment
to the tumor site; however, to the best of our knowledge, their
role in macrophage polarization has not yet been studied (40).
CAFs serve pivotal roles not only in monocyte
recruitment and differentiation but also in M2 polarization in
tumors (28,39). Cross-talk between CAFs and TAMs
mediated by different factors has been described in a number of
studies and in various tumor types, including esophageal and lung
cancer (35,36,41).
In vitro, CAFs derived from human CRC tissue induced
monocyte differentiation and M2 TAM polarization via the
upregulation of M2 markers CD206 and CD163 (28). CAFs promoted M2 recruitment in a
CRC xenograft mouse model by increasing VCAM-1 expression in tumor
cells (28). Flow cytometry
analysis of tumor tissue demonstrated that the expression levels of
VCAM-1 in CRC cells were associated with higher infiltration of
CD11b+CD68+ cells, specifically
CD163+CD206+ TAMs (28). A positive association between
CD163/CD206 and VCAM-1 expression in colon adenocarcinoma was
observed (28). TAMs induced by
CAFs promoted the migration of SW480 CRC cells and suppressed NK
cell functions in vitro (28). Increased tumor cell invasion has
been demonstrated in collagen gel co-cultures with TAMs and CAFs or
in triple co-culture with colon cancer cells (HCT116 and DLD-1)
(27). Human monocyte-derived TAMs
weakly induced invasion, whereas human colon cancer-derived CAFs
exhibited a more pronounced effect (27). CM derived from co-culture of CAFs
and TAMs had the most prominent effect on tumor cell migration
compared with CM from mono- and triple co-cultures (27).
CM obtained from CAFs isolated from human pancreatic
ductal adenocarcinoma (PDAC) tissue induced M2 polarization of
human peripheral blood monocytes in a co-culture system, as well as
in monoculture (42). Stimulation
of human peripheral blood monocytes by pancreatic CAF CM induced
CD206 expression and increased reactive oxygen species production.
M-CSF produced by CAFs is responsible for generating oxidative
stress resulting in M2 polarization (Fig. 1A) (42). In vitro, CAF-stimulated TAMs
enhanced pancreatic tumor cell proliferation, invasion and
migration (42). Another factor
involved in macrophage polarization is heat-shock protein 90 α
(HSP90α) (43). CAFs isolated from
murine PDAC tissue promoted M2 polarization in mouse RAW264.7
macrophages by secreting HSP90α, which induced a feedforward loop
of HSP90α secretion in macrophages (43). In CRC, EndoMT-derived CAFs also
exhibit high HSP90α expression (17).
In a transgenic mouse model of BC, genetic ablation
of chitinase 3 like 1 (CHI3L1) in CAFs attenuated tumor growth,
macrophage recruitment and M2 polarization (44). In vivo, injection of CHI3L1
facilitated macrophage infiltration and increased angiogenesis
(44). Several studies have
reported that overexpression of IL-33 in CAFs induced
repolarization of M1 macrophages into M2 macrophages (45,46).
Genome-wide expression profiling demonstrated upregulation of
M2-related genes [arginase 1 (Arg1), C-C motif chemokine receptor
(CCR)3, Cd163, Cdh1, F13a1, Hmox1, Il1r2, mannose receptor C-type 1
(MRC1), Pdcdl1g2, Serpinb2 and Stab1] in IL-33-stimulated
macrophages compared with non-stimulated macrophages (46). In a study in humans, as well as in
a mouse model, it has been demonstrated that IL-33-stimulated TAMs
actively promoted cancer metastasis (45,46).
The possible mechanism is associated with the
IL-33-NF-κB-MMP9-laminin axis (45,46).
CAFs express fibroblast-specific protease
fibroblast-activated protein (FAP), which is involved in metastasis
and modification of collagen matrices, as well as inflammation
(47). In vitro and in
vivo, class A scavenger receptor (SR-A)-expressing macrophages
exhibited higher adhesion to FAP-cleaved collagen (47). In a BC mouse model, CAFs increased
peritoneal macrophage adhesion to FAP-cleaved type I collagen
recognized by macrophages (47).
Macrophages with a lack of SR-A on their surface exhibited
decreased attachment compared with SR-A-expressing ones (47).
Macrophages are able to affect
'mesenchymal-mesenchymal transition' (MMT) in fibroblasts, thereby
activating their pro-tumor phenotype (48). MMT is the process during which
fibroblasts acquire the properties of mesenchymal cells after
undergoing transdifferentiation into myofibroblasts (48). In a human prostate cancer model, M2
macrophages activated healthy fibroblasts through MMT and converted
them into CAFs, which are traditionally referred to as
myofibroblasts (40,49). M1 macrophages are also able to
activate fibroblasts but to a lesser extent than M2 macrophages
(40). In a TNBC model,
macrophages actively expressed CD163 and the immunosuppressive
factors S100A9 and collagen VI, activating CAFs (39). Myeloid cells induced the functional
differentiation of fibroblasts into α-smooth muscle actin
(αSMA)+ CAFs. In turn, CAFs facilitated the migration of
monocytes via CXCL16 secretion (39).
In conclusion, the evidence indicates that CAFs
serve a major role in recruiting and pro-tumor polarization of
monocytes/macrophages, while TAMs can induce transdifferentiation
of CAFs.
The vascular system is a highly branched network
lined by ECs that supply tissues with oxygen and nutrients
(50). Lack of oxygen is a primary
stimulus for angiogenesis (50).
During this process, ECs require nutrients and energy to maintain
motility and biosynthesis of biomolecules, which are essential for
cell duplication (50). Tumor
cells produce several factors that induce angiogenesis, including
VEGF, the main angiogenesis activator, fibroblast growth factor
(FGF), which stimulates EC proliferation and induces
neovascularization, angiopoetin 2 (Ang2), responsible for
remodeling and stabilization of novel capillary tubes, IL-8, which
induces EC proliferation, and MMP2, which is a key factor for
basement membrane degradation and ECM remodeling (51). Tumor-associated vessels are
structurally abnormal; they are dilated, convoluted and
exceptionally permeant due to transcellular holes and the lack of
the basement membrane (51,52).
Additionally, tumor vessel walls may consist of both endothelial
and tumor cells (51-53). Tumor cells recruit ECs from
adjacent tissues to invade into the tumor stroma and form
neovessels (54). Tumor ECs (TECs)
exhibit higher expression levels of proangiogenic factors, such as
VEGFR-1, VEGFR-2, VEGFR-3, VEGF-D, angiopoietin receptor TEK
receptor tyrosine kinase (Tie2) and angiopoietin 1, compared with
normal ECs (53). TECs also
express adhesion molecules such as intercellular adhesion molecule
1 (ICAM-1), VCAM-1 and E-selectin, which are necessary for
interaction with stromal and tumor cells (53). TECs are highly proliferative and
self-sustaining (53).
Macrophages can contribute to tumor angiogenesis by
secreting basic FGF, macrophage migration inhibitory factor,
platelet-activating factor, prostaglandin E2, osteopontin,
adrenomedullin, VEGF-A, epidermal growth factor, placental growth
factor, TGF-β, TNFα, IL-1β, IL-8, CCL2, CXCL8 and CXCL12 (57-59).
A specific subpopulation of TAMs, which highly expresses immune
checkpoint molecule B7 homolog 3 protein, also known as CD276,
promoted angiogenesis and induced an immunosuppressive TME in a
mouse model of TNBC (60).
TAMs can induce vascular mimicry, a formation of new
blood vessels associated with an acquisition of vascular cell
features or functions by tumor cells of a non-vascular origin
(61). This phenomenon has been
described in BC, ovarian cancer, prostate cancer, lung cancer and
glioma (61,62). Macrophage-induced vascular mimicry
is an IL-6- and cyclooxygenase-2 (COX-2)-dependent process
(63). Vascular mimicry density is
positively associated with M2 polarization (63). This is explained by the presence of
hypoxia in the TME, which promotes macrophage infiltration and
vessel formation (63).
Monocytes and TAMs can be recruited to the
perivascular sites by ECs themselves (63). ECs express Ang2, stabilin-1 and
C-X3-C motif chemokine ligand 1 (CX3CL1), facilitating the
recruitment of macrophages (63).
In vitro, Tie2+ monocytes differentiated from THP-1 cells
actively migrated towards EC-derived Ang2 (Fig. 1B) (64). High Ang2 expression is associated
with increased MVD and poor prognosis in several cancer types,
including high grade serous ovarian, lung and gastric cancer
(63,65,66).
Ang2 expression is upregulated in response to anti-VEGF therapy,
resulting in treatment failure due to increased TAM infiltration
(64). Therefore, anti-VEGF and
anti-Ang2 combined therapy can improve the effectiveness of
anti-angiogenic therapy (67).
ECs are able to recruit monocytes and TAMs through
direct interaction via adhesion molecules (68). Stabilin-1 (Clever-1) is an adhesion
molecule that is expressed by ECs and involved in
monocyte/macrophage and regulatory T cell (Treg) recruitment
(68,69). Macrophages interact with stabilin-1
via the scavenger receptor stabilin-1 (Fig. 1B) (70). Stabilin-1 blockade in mice
decreased monocyte binding to tumor vessels by 70%, whereas no
inhibition of lymphocyte binding was observed (69).
In a mouse model of ovarian cancer, apoptosis
signal-regulating kinase 1, expressed by the vascular endothelium,
was responsible for EC activation and TAM recruitment, without
affecting lymphocyte recruitment (71). CX3CL1 expression in ECs is involved
in C-X3-C motif chemokine receptor 1 (CX3CR1)-dependent recruitment
of NK cells, CD8+ T cells and CX3CR1+
non-classical monocytes (63).
CX3CL1 exists both in a soluble form, acting as a monocyte
chemoattractant, and in a membrane form, promoting CX3CR1 monocyte
adhesion (Fig. 1B) (63).
In perivascular sites, macrophages are present in
two forms: Migratory TAMs and perivascular TAMs (PvTAMs) (72). Migratory TAMs are responsible for
tumor cell migration, thereby promoting cancer metastasis (73). An in vitro migration assay
demonstrated that rat mammary adenocarcinoma tumor cells migrating
in streams with TAMs, obtained from murine bone marrow, moved at
higher speeds, from greater distances towards HUVECs coated onto
Sephadex beads compared with tumor cells migrating without TAMs
(73). PvTAMs are a specific
subpopulation of TAMs residing close to blood vasculature
(<15-20 µm), which serves an important role in
proangiogenic niche formation and tumor metastasis (74,75).
PvTAMs are associated with increased tumor angiogenesis, distant
metastasis, poor prognosis and tumor recurrence after chemotherapy
(75). PvTAMs are found in
structures defined as the TME of metastasis (TMEM), and are
responsible for VEGF-A production, resulting in transient vascular
leakiness, as well as tumor cell extravasation and distant
metastasis (76). The TMEM
consists of PvTAMs, invasive tumor cells and ECs (76). In macrophage-depleted mice, the
CD68+ TAMs amount returned to normal levels in 4 days,
and TAMs were rarely found in tight proximity to CD31+
ECs (72). TAMs accumulated in
perivascular sites and developed PvTAM properties after 7 days
(72). It has been demonstrated
that PvTAMs are originated from CCR2+ monocytes and are
recruited to the perivascular space by perivascular CAFs (77). Perivascular CAFs secrete CXCL12,
IL-33 and SDF-1α in response to cancer cell activation (63). High infiltration of macrophages in
vascular sites can be responsible for pro-metastatic niche
formation (78). In a BC model,
perivascular macrophages were activated by the cancer cell-derived
ECM protein tenascin (TNC) (78).
PvTAMs contributed to vascular niche formation via nitric oxide and
TNF secretion, activating ECs (78). TNC deficiency decreased metastasis
formation in vivo (78). In
BC, PvTAMs are originated mainly from bone marrow-derived
interstitial macrophages, but not from resident macrophages, and
have a mixed M1/M2 phenotype (78). Immunofluorescence analysis of
metastatic lung tissue demonstrated that
VEGFR1+F4/80+ interstitial macrophages were
found in close proximity to CD31+ ECs (78). This suggested their pivotal role in
induction of the perivascular niche (78). PvTAMs in mouse mesentery mainly
express the M2 marker CD206 (79).
Co-culture of M1 and M2 TAMs with ECs demonstrated that M2 TAMs
exhibited greater binding to ECs compared with M1 TAMs (79). Immunofluorescence analysis of human
glioblastoma multiforme (GBM) tissues revealed that
CD68+CD206+ macrophages were localized in
proximity to CD31+ ECs (80). Similarly, in an orthotopic genetic
murine GBM model, F4/80+CD206+ macrophages
were localized near ECs (80).
GBM-derived ECs induced M2 polarization of macrophages via M-CSF
and IL-6 expression (80). IL-33
secreted by pericytes and CAFs promotes M2 polarization and induced
MMP9 expression in M2 TAMs facilitating metastasis development via
the IL-33-ST2-NF-κB-MMP9-laminin pathway in a mouse model of
pancreatic cancer (Fig. 1B)
(46). PvTAMs can serve an
immunosuppressive role in perivascular sites (75). They exhibit high secretion of
IL-10, which leads to cytotoxic T cells and Treg suppression in
perivascular sites (75).
Anticancer therapy causes vascular damage and
hypoxia, leading to increased recruitment of myeloid cells,
proliferation of ECs and migration of pericytes, which are involved
in reconstruction of the vessels in order to restore oxygen levels
and provide nutrients for tissue repair (81). The main role in this process is
assigned to the Ang-Tie2 pathway (63,75).
Tie2 is a receptor that is broadly expressed on vascular ECs and on
a subpopulation of macrophages [Tie2-expressing macrophages (TEMs)]
(75). Ang2 is a proangiogenic
factor and chemokine for TEMs, which also serves a role in
regulating the recruitment and/or activation of perivascular
macrophages in the TMEM (75).
TEMs are a critical subpopulation involved in tumor angiogenesis
and are able to adhere to ECs (81). TEMs are essential for vascular
anastomosis and formation of new vessels (75,81).
Ang2 acts as an inducer of M2 polarization and Tie2+
monocyte recruitment via the upregulation of M2 markers (IL-10 and
MRC1) and angiogenesis-related genes (cathepsin B and thymidine
phosphorylase) (63).
In a mouse model of fibrosarcoma, tumor relapse
after chemotherapy was associated with active vessel reconstruction
within 14 days (81). FACS
analysis revealed that the proportion of
CD11b+Tie2+ cells was increased compared with
that in untreated mice (81). Gene
expression profiling of TEMs isolated from mouse mammary tumors
revealed increased expression of pro-tumoral genes, including MMP9,
VEGFA, CXCL12, toll like receptor 4, neuropilin 1 (NRP1) and
platelet derived growth factor (PDGF) subunit B (75). Immunofluorescence analysis of the
TMEM obtained from tumor-bearing mice demonstrated that a major
fraction of PvTAMs was Tie2+/VEGFA+
macrophages (82). VEGFA
expression in TEMs was elevated compared with that in the adjacent
ECs and surrounding tumor tissue (82). TMEM-associated macrophages
exhibited expression of the TEM markers MRC1, CD11b and F4/80
(82). Anti-VEGFA treatment
decreased the amount of circulating tumor cells and vascular
permeability (82). In a mammary
mouse model, Tie2+ PvTAMs increased vessel permeability
via VEGF-A-dependent destabilization of EC junctions (Fig. 1B) (82).
Another specific subpopulation of PvTAMs exhibit
lymphatic vessel endothelial hyaluronan receptor 1 (LYVE1)
expression, which is considered to be a marker for lymphatic
endothelium (77). In a mouse
model of BC, LYVE1 was expressed by pro-angiogenic TAMs located
near the blood vasculature (77).
LYVE1+ PvTAMs formed nest structures in perivascular
niches in a CCR5-dependent manner (77). CCR5 depletion led to decreased
tumor growth in mice (77).
Impaired nest formation resulted in increased chemosensitivity in
tumor-bearing mice treated with fluorouracil (77). LYVE1+ PvTAMs are also
responsible for αSMA+ CAFs expansion within the
perivascular niche (Fig. 1B)
(74). PvTAMs are a major source
of PDGFC in the perivascular niche, which is the ligand for PDGF
receptor α expressed on αSMA+ CAFs (74). PDGF blockade led to decreased
migration of αSMA+ CAFs (74). This observation revealed reciprocal
interactions between PvTAMs and perivascular CAFs (PvCAFs), since
PvCAFs are responsible for monocyte/macrophage recruitment into
perivascular sites (77).
Several studies have demonstrated the important
role of TAMs in tumor lymphangiogenesis (83-87).
TAMs are positively associated with lymph node metastasis and
lymphatic vessel density (LVD) in numerous cancer types, including
lung cancer, tongue OSCC, pancreatic cancer, gastric cancer,
cervical cancer and ovarian cancer (88-95).
VEGF-C is a known lymphangiogenic factor that is
most essential for LEC proliferation and lymphatic vessel sprouting
(96). TAM expression of VEGF-C is
stimulated by different stimuli, such as TNFα/TNF receptor 1
(TNFR1) signaling (97-99), IL-1β secretion by tumor cells
(98), and via VEGFR activation by
endothelial cell-derived VEGF-C and VEGF-D (Fig. 1B) (99,100).
In a murine BC model, sphingolipid
sphingosine-1-phosphate receptor 1 (S1PR1) overexpression in TAMs
promoted pulmonary metastasis and tumor lymphangiogenesis in
vivo (101). The mechanism
included the upregulation of inflammasome-related gene NLR family
pyrin domain containing 3 in S1PR1+ TAMs followed by
IL-1β production (101).
Lung-derived LECs stimulated by IL-1β exhibit increased VEGF-C
expression and a high proliferation rate (101). M2 TAMs activated by tumor
cell-derived IL-1 promote lymphangiogenesis and lymph node
metastasis through the secretion of VEGF-A and VEGF-C (102). S1P protein derived from dying
breast tumor cells stimulated the expression of lipocalin 2 (LCN2)
in TAMs in mouse mammary tumor virus-polyoma middle tumor-antigen
mice (103). LCN2 promoted LEC
differentiation and tube formation via activation of VEGF-C
expression (103).
TNFα stimulates morphological changes and migration
of LECs in a VEGFR-3-dependent manner in vitro (97). Interaction of tumor cell-derived
TNFα with macrophage TNFR1 triggers VEGF-C secretion by
CD206+ TAMs, amplifying lymphangiogenesis and lymphatic
metastasis in a murine lung cancer model (97). TNFα and IL-1β secretion by tumor
cells stimulated VEGF-C expression in macrophages in a co-culture
of a melanoma cell line and murine peritoneal macrophages (98). Macrophages differentiated from
monocytes of healthy donors promoted BC cell adhesion to LECs in
vitro in an IL-1β-dependent manner, suggesting a possible
mechanism of lymphatic metastasis (104). It has been suggested that lymph
node metastasis associated transcript 1-induced CCL2 expression in
bladder cancer cells could promote TAM recruitment, which
contributed to lymphangiogenesis and lymphatic metastasis via
VEGF-C expression (105).
CM from co-culture of cervical cancer cell lines
and macrophages differentiated from THP-1 monocytes stimulated tube
formation of human LECs (93).
IL-1β, IL-8, VEGF-C and VEGF-A secretion was elevated in CM
(93). LECs isolated after a tube
formation assay exhibited increased expression levels of VEGFR-3
and podoplanin (PDPN) (93). In
vitro, CM from TAMs isolated from human epithelial ovarian
cancer (EOC) ascites induced LEC proliferation, migration and tube
formation (95).
The interaction between TAMs differentiated from
human peripheral monocytes and LECs isolated from metastatic lymph
nodes of human gastric cancer has been examined in co-culture
(106). Expression levels of MMP9
and adhesion molecules (CD44, neural cell adhesion molecule 1,
ICAM-1, VCAM-1, E-selectin and osteopontin) were upregulated in
capillary-forming LECs (106).
TAMs in contact co-culture with LECs exhibited increased expression
levels of various cytokines, including IL-1β, IL-6, IL-8, IL-10,
IL-12 and IL-18 (106). CXCL2
expression was upregulated, whereas CXCL6 and CXCR2 expression was
downregulated in macrophages in contact with LECs (106). In human papillary thyroid
carcinoma, both M2 TAMs and cancer cells secrete MMP2 responsible
for lymphatic vessel basement membrane degradation that could
further lead to lymphatic invasion (107).
In a murine BC 4T1 model, a specific subpopulation
of TAMs was localized near lymphatic vessels (83). TAMs isolated from 4T1 tumors
exhibited high expression levels of PDPN, a heavily O-glycosylated
small mucin-type transmembrane glycoprotein, and were referred to
as PDPN-expressing macrophages (PoEMs) (83). PoEMs interact with LECs via
PDPN-galectin 8 interaction and promote tumor lympangiogenesis
(83). PDPN knockout in mice led
to reduced tumor lymphatic growth and lymphatic metastasis
(83). In murine mammary tumors
with high SEMA7 expression, an elevated amount of PDPN+
TAMs was observed (84). In
vitro, treatment of murine macrophages with recombinant SEMA7A
increased PDPN expression as well as macrophage motility and
adherence to the human dermal LEC monolayer and promoted
lymphangiogenesis (84). The
adhesion of macrophages to the LEC is mediated by the upregulation
of β1 integrin and the interaction of PDPN with its receptor CLEC-2
(Fig. 1B) (84).
IHC staining of human cervical squamous cell
carcinoma tissue demonstrated that, under hypoxic conditions,
lymphatic vessels were encapsulated by CD163+ TAMs, and
the LVD was higher compared with that under normoxic conditions
(108). These structures were
referred to as lymphatic vessels encapsulated by TAMs (LVEM)
(108). Hypoxic TAMs exhibited
high IL-10 expression (108). A
popliteal lymphatic metastasis mouse model revealed that IL-10
promoted lymphangiogenesis and LVEM formation (108). TAM-derived IL-10 activated LEC
production of CCL1, which resulted in further TAM recruitment and
sustained LVEM formation and lymphatic metastasis (108).
VEGF-C and VEGF-D-expressing TAMs form small
clusters close to lymphatic microvessels and tumor surfaces of
human cervical squamous carcinoma (99). All VEGF-C and VEGF-D-expressing
TAMs also express VEGFR-3, but in a granular intracellular pattern,
and not on the cell surface membrane (99). In human mammary carcinoma,
VEGF-C-expressing TAMs have been reported as morphologically
different from other macrophages (109). They were generally larger, oval
or irregular in shape and had abundant cytoplasm, and were located
mainly in peritumoral regions (109). Macrophage numbers and VEGF-C
expression were positively associated with LVD and LEC
proliferation (109).
Myeloid cells can contribute to lymphangiogenesis
by differentiating into myeloid-LEC progenitors (M-LECP) (110-112). In human BC, M-LECP have been
identified as cells co-expressing CD68 and LEC markers (LYVE1,
PDPN, VEGFR-3 and PROX-1) (113).
Human M-LECPs are usually identified by additional expression of a
marker of progenitor cells, such as CD133 (111). The density of M-LECP is
associated with LVD and lymphatic metastasis in human BC (113).
Th1 cytokines affect the polarization of
macrophages, and the latter, in turn, can change the
differentiation of Th1 and Th2 (117). The predominance of Th1 in the
tumor inhibits the maturation of tumor-infiltrating
CD11b+F4/80+ macrophages isolated from mouse
liver cancer tissue (117). It
has also been suggested that CD11b+F4/80+
macrophages, rather than Tregs, serve a major role in Th1
suppression (117). Fragile
site-associated tumor suppressor (Fats)−/− bone
marrow-derived macrophages (BMDMs) isolated from fat-deficient mice
stimulated CD4+ T lymphocyte differentiation to the Th1
phenotype in vitro (118).
In vivo, a similar effect was observed when
Fats−/− BMDMs were transferred to mice injected
subcutaneously with melanoma cells (118). CD4+ Th1 cytokines
activate CD8+ T cells, which display antitumor functions
in vivo (118). In
addition, CD4+ Th1 produce IFN-γ, which promotes
additional polarization of macrophages towards the M1 phenotype
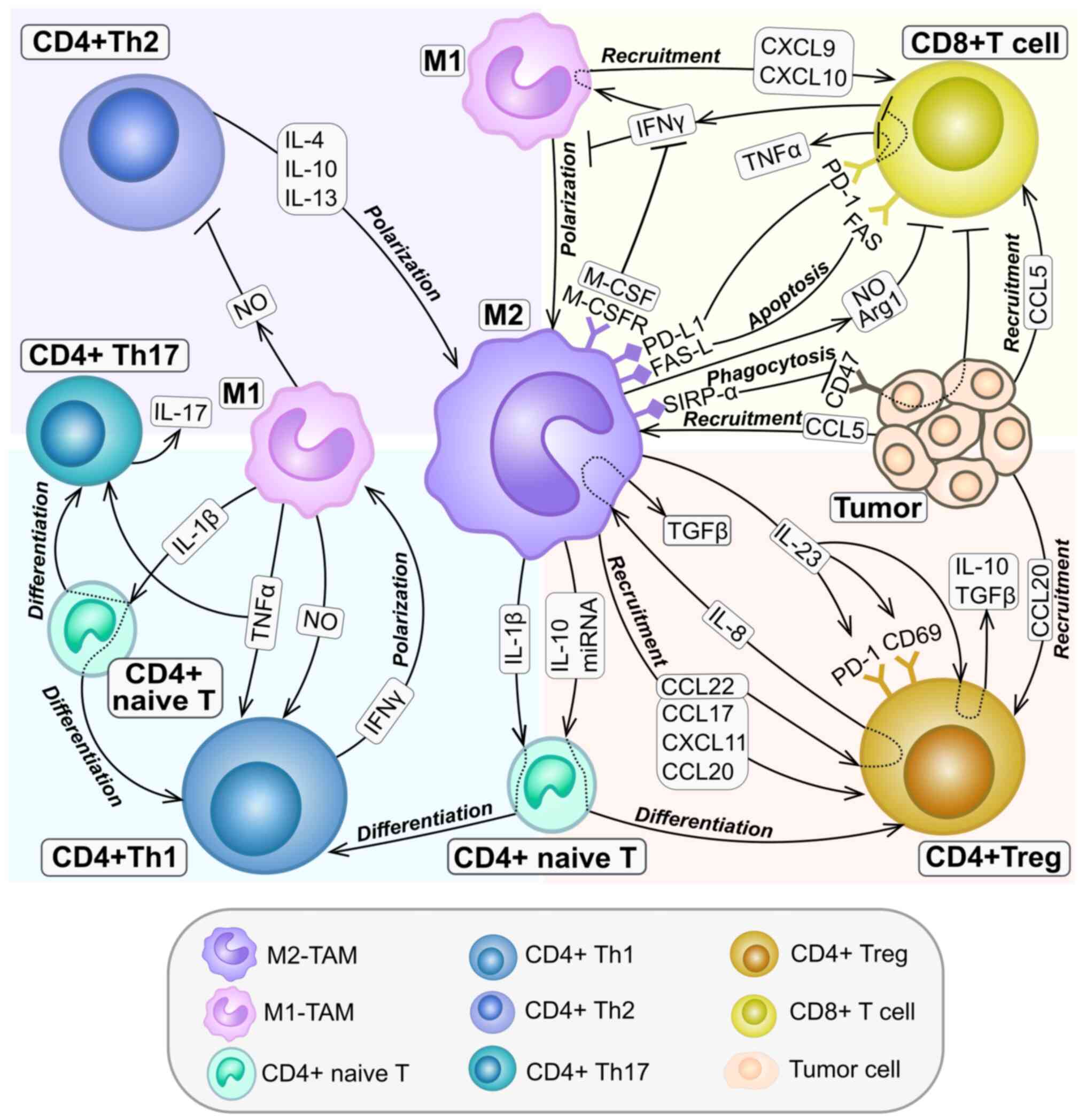
(Fig. 2) (118). Receptor-interacting
serine/threonine protein kinase 1 (RIP1) regulated macrophage
differentiation into a pro-tumor phenotype, and RIP1 inhibition in
TAMs altered the macrophage phenotype towards M1 in a mouse model
of PDAC (119). This induced Th
differentiation toward a mixed Th1/Th17 phenotype via a
TNFα-dependent mechanism (119).
The antitumor effect of gemcitabine in combination with
anti-programmed cell death protein 1 (PD-1) antibodies was mediated
by activation of Th1 and M1 macrophages in a mouse model of liver
metastasis of pancreatic cancer (120).
Th2, similarly to M2 macrophages, are pro-tumor
cells, and associated with tumor progression and metastasis
(121,122). Th2 cytokines are involved both in
repolarization of macrophages towards M2, and in the suppression of
Th1 and activation of tumor growth signaling pathways (121). In a mouse model of spontaneous
pancreatic islet carcinogenesis, tumor macrophages with altered
polarization after irradiation of the pancreatic region synthesized
an increased level of inducible NOS (iNOS; specific for M1
macrophages), which activated the cytokine profile of Th1 and
inhibited Th2 cytokines (121).
M2 pro-tumor macrophage activity was enhanced by CD4+
cells expressing higher levels of IL-4, IL-13 and IL-10 (specific
for Th2) compared with IFNγ (specific for Th1) or IL-17 (specific
for Th17) expression in a mouse BC model (Fig. 2) (123). In another study using a mouse
model of BC, it was demonstrated that depletion of TAMs in the
tumor resulted in a decrease in the number of CD4+ T
cells that express Th2 cytokines (122).
An increase in the number of TAMs along with Tregs
is observed during carcinogenesis and associated with unfavorable
prognosis in patients with colorectal (134), lung (135) and ovarian (136) cancer. In the tumor stroma of lung
cancer, MARCO-expressing macrophages have an immunosuppressive
phenotype, increasing the proliferation of Tregs, which is
associated with a negative prognosis (135). In liver cancer in mice,
intratumor triggering receptor expressed on myeloid cells 1
(TREM-1)+ macrophages suppress the activity of
CD8+ T cells by activating Tregs (137). The inhibition of
TREM-1+ macrophages does not eliminate the depletion of
CD8+ T cells, indicating that Tregs cause
CD8+ T cell dysfunction (137).
Direct interaction of TAMs with Tregs has been
demonstrated in numerous studies (137-141). TAMs synthesize a large amount of
chemoattractants that facilitate the recruitment of Tregs in tumors
(Fig. 2) (138). The key chemokines include CCL17
and CCL22 (138). Therapeutic use
of immunomodulator imiquimod reduced the production of CCL22 of
CD11b+ TAMs in a melanoma model, resulting in a decrease
in the population of Foxp3+ Tregs and an increase in the
antitumor response (139). In the
hypoxic tumor environment, TREM-1+ macrophages activated
the synthesis of the CCL20 ligand, which has an affinity to the
CCR6 receptors on Tregs, stimulating Treg recruitment,
immunosuppression and tumor growth in a mouse orthotopic liver
tumor model (137). Macrophages
isolated from malignant pleural effusions of patients with lung
cancer exhibit increased CCL22 expression (140). CCL22 induces the recruitment of
Tregs, which, through positive feedback, affect macrophages via the
production of IL-8 (140). IL-8
increases the production of TGF-β by macrophages, which suppresses
the immune system and induces carcinogenesis (140). TAMs from lung (3LL-R)
tumor-bearing mice were treated with agonists to liver X receptor
(a transcription factor from the nuclear receptor family), which
inhibited the secretion of CCL22 and CCL17 from TAMs (141). The macrophages were then
stimulated in vitro with GM-CSF or IL-4, and a decrease in
the synthesis of CCL17 and CCL22 by the M2 macrophages was observed
(141). This was associated with
a decrease in the number of Tregs in a mouse model of syngeneic
Lewis lung carcinoma (LLC) (141).
In a mouse CRC model, it was found that the homing
and migration of Tregs into the tumor depended on the interaction
of CCR6 receptor with ligand CCL20 produced by both tumor cells and
macrophages (142). Macrophages
isolated from the tumor tissue of patients with CRC express high
levels of CXCL11, which has an affinity to CXCR3 receptor expressed
on Foxp3+IL-17+ Tregs (143). The depletion of macrophages by
injection of diphtheria toxin into CD11b diphtheria toxin receptor
(CD11b-DTR) mice decreased CCL20 mRNA and protein expression in the
CRC tissue, indicating that TAMs are required for release of CCL20
from tumor cells and recruitment of Tregs (Fig. 2) (142). In mice injected with melanoma or
colon adenocarcinoma cells, Tregs inhibited the activity of
tumor-suppressing CD8+ T cells, which in turn led to an
increase in the M2 macrophage phenotype (144).
M2 macrophages derived from human peripheral blood
monocytes induce T cell differentiation towards Tregs via IL-10
secretion (136). In patients
with EOC, an increased number of TAMs and Tregs, and elevated
levels of IL-10 were found in tissue samples, and these were
associated with decreased survival time (136). In a mouse model of ovarian
cancer, TAMs secreted exosomes containing microRNAs, which affected
the differentiation of CD4+ T cells and increased the
number of immunosuppressive Tregs via the STAT3 mechanism (127). In vitro, recombinant human
IL-23 secreted predominantly by TAMs enhanced the immunosuppressive
activity of Tregs, as indicated by increased expression levels of
PD-1 and CD69, as well as the secretion of IL-10 and TGFβ (Fig. 2) (145).
CCL5 is predominantly expressed by tumor cells and
stimulates macrophages to synthesize CXCL9 (155). Tumor-infiltrating lymphocytes
express IFNγ, which in turn stimulates CXCL9 synthesis in
macrophages (Fig. 2) (156). In a mouse model of HCC, CXCL10
together with CXCL9 affected the cytotoxic index of CD8+
T cells (158). The decrease in
the expression of these cytokines was associated with an increase
in tumor infiltration by CD163+ TAMs (158).
M-CSF affects the proliferation and differentiation
of macrophages via M-CSFR receptor (162). The M-CSF/M-CSFR axis is able to
suppress the cytotoxicity of CD8+ T cells (163). The use of anti-bodies blocking
M-CSFR reduced the infiltration of TAMs and increased
CD8+ T cell activation in a mouse colon carcinoma model
(163). A decrease in M-CSF
expression increases the expression of IFN-γ by T cells (164). The IFNγ-IFNγ receptor 1 axis
between CD8+ T cells and TAMs reduces macrophage
polarization into the M2 phenotype (Fig. 2) (144).
In subcutaneous mouse breast and colon tumor
models, monoacylglycerol lipase deficiency resulted in lipid
accumulation in TAMs and their polarization into M2 phenotype,
resulting in reduced CD8+ T cell activity (165). The decreased expression of COX-2
increased the number of TAMs, which was associated with an increase
in CD8+ T cell infiltration and improved survival of
mice in a mammary carcinoma model (166). Blockage of COX-2 reduced the
expression of macrophage pro-tumor markers (arginase-1, IL-10 and
iNOS), which reversed the suppression of tumor-infiltrating
lymphocytes (166).
P65 is one of the proteins of the NF-κB family, and
its activation leads to increased expression of T-cell inhibitory
molecule B7x (also referred to as B7-H4 or B7S1) on the TAM surface
and a decrease in IL-10 in TAMs, suppressing the activation of
tumor-infiltrating CD8+ T lymphocytes during lung
tumorigenesis (167). However,
inactivation of macrophage PI(3)
kinase, which is an inhibitor of NF-κB, promoted CD8+ T
cell activation in tumors in mouse models of implanted human
papillomavirus (HPV)+ and HPV− head and neck
squamous cell, lung and breast carcinoma (168).
Several studies have demonstrated that anti-PD-1
therapy combined with macrophage inhibitors is more efficient in
terms of increasing the immune response and suppression of TAM
activity compared with mono-immunotherapy (150,159,163). In mouse models of glioblastoma
and ovarian cancer, anti-PD-1 therapy in combination with the
blockade of LIF (anti-IL-6 class cytokine that affects cell
proliferation) increased survival and promoted tumor regression
more effectively than mono-therapy (159). The use of a M-CSF inhibitor in
combination with anti-PD-1 therapy increased the migration and
infiltration of CD8+ T cells in a mouse BC model
(150). Another macrophage
inhibitor, lenvatinib, in combination with an anti-PD-1 agent,
increased the percentage of CD8+ T cells, followed by a
decrease in tumor growth and vascular invasion in a mouse colon
carcinoma model (163).
B cells are important components of the immune
defense against cancer. Opinions on the role of B cells in tumor
carcinogenesis vary based on collected evidence on both pro-tumor
and antitumor activity of B cells (177-181). For example, pro-tumor activity of
B cells can be mediated by the recruitment of TAMs via immune
complexes and Fcγ receptor interactions, whereas the beneficial
effect of B cells is related to the activation of antitumor
immunity through antigen presentation or direct cytotoxicity
towards tumor cells via secretion of granzyme B and TNF-related
apoptosis-inducing ligand (179,181). The complexity of studying B cells
in carcinogenesis is associated with difficulties in isolating and
purifying B cells (180).
Furthermore, there are several functional phenotypes depending on
the TME (177).
Evidence indicates that B cells and macrophages can
have a common bipotential progenitor in bone marrow (182). In the early stages of
development, pre-B cells co-expressing B-cell and myeloid markers
can differentiate into macrophages, due to the receptors of B cells
not being rearranged, which indicates the plasticity of B cells
(182).
When studying the cross-talk of Bruton's tyrosine
kinase (BTK)-expressing immune cells in a mouse model of pancreatic
carcinoma and in vitro, the important role of the
interaction between B cells and macrophages was noted (179). Activation of macrophage Fc
receptors initiates BTK signaling via the PI3K-dependent signaling
pathway and activates macrophage repolarization towards the M2
phenotype and promotes tumor development (179). Tumor B cells enhance the change
in macrophage phenotype in favor of type 2 by influencing
macrophage Fc receptors (179).
The antitumor or pro-tumor function of B cells can
be mediated by antibody synthesis (179). B-cell-expressed IgG binds to
macrophages and induces macrophage-mediated phagocytosis of tumor
cells through antibody-dependent cellular cytotoxicity in breast,
pancreatic and gastric cancer but not in clear cell renal cell
cancer, squamous cell carcinoma and non-small cell lung cancer
(179). IgG secretion by B cells
is associated with poor prognosis (183).
Despite these findings, to the best of our
knowledge, the direct mechanisms of the interaction of TAMs and B
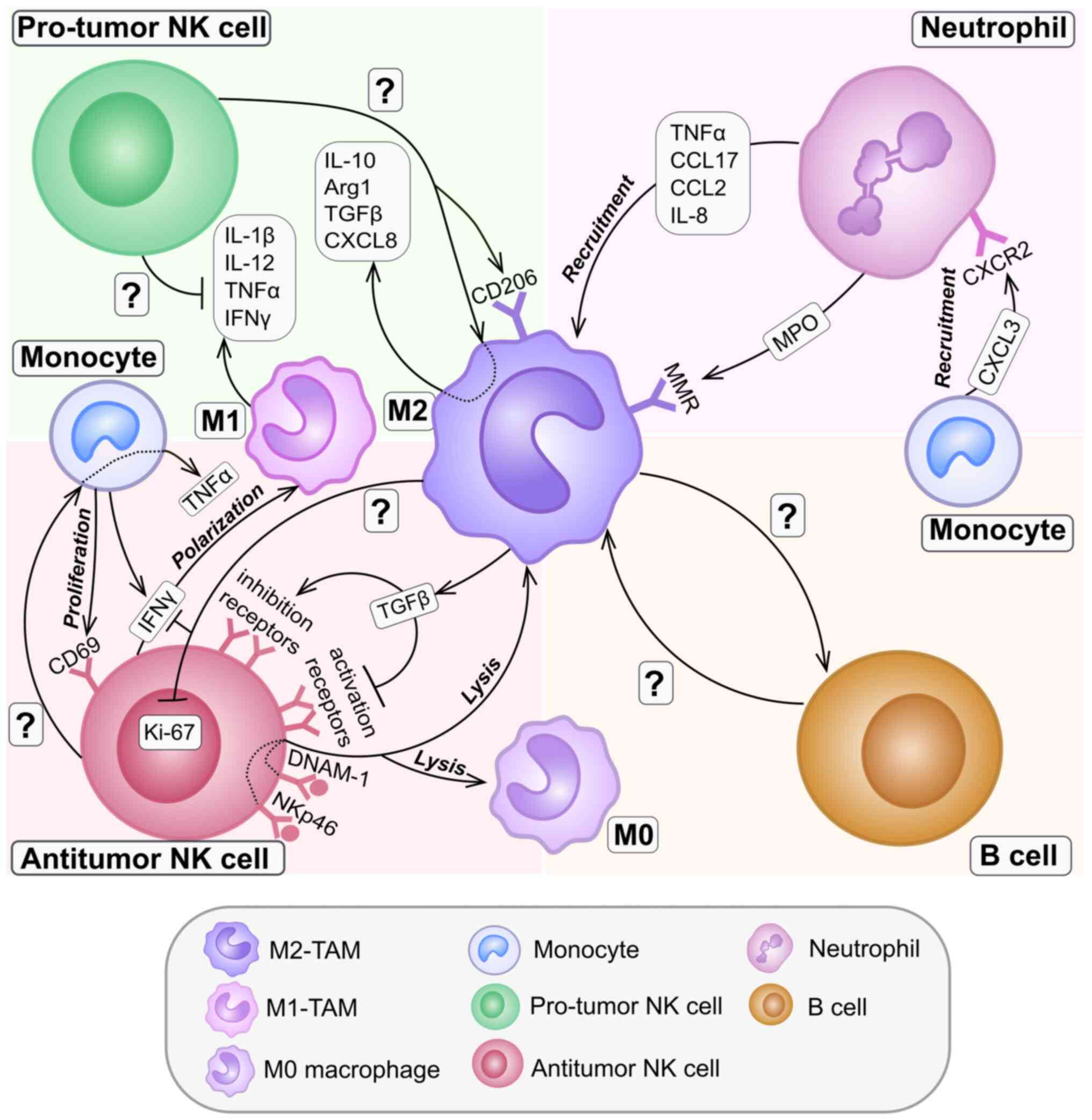
cells in the TME remain unknown (Fig.
3).
In turn, the TME can reduce the cytotoxic activity
of NK cells against tumor cells and inhibit the expression of NK
receptors responsible for the recognition and killing of tumor
cells (187,188). Macrophages are able to change the
expression of activating and inhibiting NK cell receptors (187,189-192). M2 TAMs suppress the cytotoxic
activity of NK cells (193).
Co-culture of NK cells isolated from the murine spleen and
peritoneal or bone marrow M2 macrophages revealed suppression of
the expression of CD27 activation marker, while co-culture with
M1-like macrophages under similar conditions did not suppress the
cytotoxicity of NK cells (187).
In patients with stage II and III esophageal
cancer, high tumor infiltration by TAMs and low infiltration by NK
cells are associated with poor prognosis (194). Macrophages/monocytes isolated
from autologous nontumor and tumor tissues of patients with gastric
cancer cultured together with allogeneic peripheral blood NK cells
reduced the expression of IFNγ and TNFα in NK cells co-cultured
with tumor-associated monocytes/macrophages (190). In addition, a decrease in the
expression of cell proliferation marker Ki-67 by NK cells was
observed, which together indicated a decrease in the functional
activity of NK cells (190).
MARCO-expressing TAMs exhibited immunosuppressive properties
against NK cells in a mouse melanoma model (195).
One of the mechanisms by which TAMs interact with
NK cells is the secretion of TGFβ1 by macrophages/monocytes, which,
in a gastric cancer ex vivo model, induced functional
impairment of NK cells (190).
In vitro, TGFβ1 affected the expression of activation
receptors, natural cytotoxicity triggering receptor 3 (NKp30) and
killer cell lectin like receptor K1 (NKG2D), in NK cells isolated
from human peripheral blood mononuclear cells (191). A decrease in the expression of
NKp30 and NKG2D suppresses the cytotoxic activity of NK cells
against a melanoma cell line (191). In peripheral blood NK cells from
patients with prostate cancer, TGFβ is able to increase the surface
expression of CD56 and CD9 receptors, which decrease the cytotoxic
activity of NK cells, and reduce the expression of NK cell
activation receptor NKG2D in vitro (192). Similarly, in a mouse lymphoma
model, M2-like macrophages suppressed the activity of NK cells by
secreting TGF-β, which decreased the expression of degranulation
factor CD107a by NK cells (187).
Activation of the natural cytotoxicity receptors 46
and DNAX accessory molecule-1 receptors on NK cells results in the
lysis of human peripheral blood monocyte-derived M2 macrophages
(196,197). Resting NK cells do not exhibit
cytolytic activity against macrophages; however, activated NK cells
lyse M0 and M2 macrophages (Fig.
3) (197). In co-culture,
monocytes obtained from peripheral blood and synovial fluid of
patients with inflammatory arthritis could activate NK cells via
increased expression of the proliferation marker CD69 and elevated
secretion of IFN-γ by NK cells (198). Additionally, NK cells increased
the secretion of the pro-inflammatory cytokine TNFα by monocytes
(198). IFN-γ secreted by NK
cells altered macrophage polarization to an M1 phenotype in a mouse
model of sarcoma (199).
In addition to antitumor activity, NK cells can be
involved in pro-tumor polarization of TAMs (192). Co-cultivation of human MDMs with
NK cells isolated from the blood of patients with prostate cancer
revealed a decrease in the expression of M1-like factors (IL-12,
TNFα, IFNγ and IL-1β) and an increase in the expression of M2-like
factors (CD206, Arg1, IL-10, TGFβ and CXCL8) (Fig. 3) (192).
Neutrophils are immune cells that serve an
important role in the development of inflammation (200). Similar to macrophages,
neutrophils are classified into N1 and N2: Antitumor and pro-tumor,
respectively (201). Activated
neutrophils are able to secrete IL-8 and TNFα, which recruit
macrophages into the inflammation site (202). The interaction of macrophages
with neutrophils likely occurs through the macrophage mannose
receptor highly expressed by macrophages and through
neutrophil-secreted myeloperoxidase (Fig. 3) (202). Neutrophil-derived CCL2 and CCL17
affected the migration of macrophages into the tumor in a mouse
model of HCC (203). In another
study, granulocyte colony-stimulating factor receptor-depleted mice
had neutropenia, which was associated with increased tumor
progression, compared with wild-type mice in a
3-methylcholanthrene-induced sarcoma model (200). An increase in neutropenia was
associated with an increase in the expression of the M2 macrophage
genes and was associated with poor prognosis (200). Tumor monocytes derived from
patients with non-small cell lung cancer are also able to affect
the migration of neutrophils via CXCL3, a ligand for the neutrophil
receptor CXCR2 (204).
The co-cultivation of tumor cells simultaneously
with macrophages and neutrophils resulted in increased
proliferation, invasion and colony formation of tumor cells
compared with those of tumor cells cultured with a monoculture of
macrophages or neutrophils (205). The data were confirmed in three
intrahepatic cholangiocarcinoma cell lines: SG231, RBE and HuCCT1
(205).
Anti-CAF and anti-TAM combined therapy can be a
promising option in antitumor immunotherapy (206-208). In murine models of lung cancer,
melanoma and colon cancer, immune modifying poly(lactic-co-glycolic
acid) nanoparticles (referred to as ONP-302) were used to target
CAFs and TAMs (209). The authors
of the previous study suggested that ONP-302 was able to alter the
polarization of TAMs and MDSCs from pro-tumor to antitumor state.
The same mechanism was described for CAFs (209).
Combined anti-VEGFA/Ang2 and anti-CD40 therapy led
to vascular pruning and normalization of blood vessels, myeloid
cell activation, including M1-like TAM skewing in colon cancer,
melanoma and BC models in vivo (210). Tie2 expression in TAMs and ECs
could be another possible target for combined therapy (211). The Tie2-targeting anti-tumor
hydrophobic peptide T4 was modified to generate self-assembly
nanoparticles (P-T4) (211). T4
interacted with Tie2 and inhibited signal transduction, cell
migration and angiogenesis (211). P-T4 exerted an inhibitory effect
on the viability of Tie2-positive RAW264.7 cells and ECs in
vitro, and reduced vessel density and distant metastasis
formation in vivo (211).
SAR131675 is a dose-dependent selective inhibitor of VEGFR-3
tyrosine kinase activity (212).
In a murine BC model, SAR131675 inhibited primary human lymphatic
cells in vitro and reduced the migration of cancer cells
into lymph nodes and lungs in tumor-bearing mice (212). IHC staining of tumor tissues
isolated from murine mammary tumors revealed reduced macrophage
infiltration, suggesting that reduction of tumor growth might be
associated with decreased TAM infiltration (212).
The use of immunotherapy against both macrophages
and Tregs can help improve the prognosis of patients with cancer,
since there is a positive association between TAMs and Tregs
(146). Mono-immunotherapy
against TAMs or Tregs was not efficient in a colon cancer in
vivo model (213). Deletion
of the CSF1 gene in colon adenocarcinoma cells (MC38 line)
led to a decrease in the population of M2 macrophages in
vivo but an increase in the number of Foxp3+ Tregs
followed by slight tumor growth in mice (213). In turn, depletion of
Foxp3+ Tregs in tumor-bearing mice upon administration
of diphtheria toxin resulted in an increase in colony-stimulating
factor 1 receptor (CSF1R)+ TAMs and slight tumor growth
(213). In a mouse model of head
and neck cancer, radiotherapy in combination with anti-CD25
depletion of Tregs resulted in reduced tumor growth and an increase
in the M1/M2 ratio (214).
Notably, the use of radiotherapy alone led to an increase in the
proportion of M2 macrophages (214).
In a mouse pancreatic cancer model, a CSF1R
blockade and PD-1/CTLA-4 antagonist combination therapy reduced
tumor progression more effectively than single therapy (215). In a mouse melanoma model, the use
of the CSF1R inhibitor in combination with the transfer of naive
CD8+ T cells activated with a synthetic peptide vaccine
resulted in a reduction in tumor growth (216). The use of regorafenib (an
inhibitor of protein kinases, including VEGFR, CSF1R and others) in
combination with anti-PD1 therapy inhibited tumor development due
to a decrease in M2 macrophages and Tregs, and an increase in
CD8+ T cells and M1 macrophages inside the tumor in a
mouse colon cancer model (217).
In mice with gastrointestinal stromal tumors, anti-CD40 therapy
activated TAMs against tumor cells and recruited more monocytes
from the bone marrow (218).
Additionally, the efficacy of treatment with the combination of
anti-CD40 with imatinib (affects the KIT mutation characteristic of
this type of cancer) was completely dependent on the presence of
macrophages in the tumor and partially on the presence of
CD8+ T cells (218).
During the development of methods of therapy for
pancreatic adenocarcinoma, it was revealed that the simultaneous
inhibition of CCR2+ macrophages and CXCR2+
neutrophils enhanced antitumor immunity (224).
The present review demonstrates the broad network
of TAM interactions with stromal and immune components of the TME
via cell-to-cell and extracellular mechanisms. The collected data
indicate that CAFs are an essential source in the TME for monocyte
recruitment and M2 TAM polarization. Despite the involvement of
TAMs in ECM remodeling, they have a minor impact on CAF activity,
only inducing MMT. After polarization, TAMs are able to interact
with naive fibroblasts and promote their differentiation into
functional CAFs, creating a self-sustaining loop.
TAMs are also involved in tumor angiogenesis and
lympangiogenesis via reciprocal interaction with blood ECs and
LECs, respectively. ECs are able to recruit TAMs to the tumor site,
where the latter are differentiated toward perivascular and
lymphatic vessel-associated macrophage subpopulations promoting
proliferation and migration of ECs, as well as growth of novel
vessels.
The crosstalk between TAMs and other immune cells
can be crucial for the balance between the pro-tumor and anti-tumor
immune state and regulation of immunosuppression.
Finally, combined targeting of TAMs and other cells
of the TME can be decisive to achieve advanced efficacy of
anticancer treatment. Most studies have focused on targeting
stromal or immune cells alone (225-228). However, it is critical to not
exclude cell-cell crosstalk within the TME and future studies
should aim to identify possible targets in these interactions in
order to increase the effectiveness of antitumor therapy. Combined
therapeutic approaches should be considered to prevent distant
metastasis and reduce the risk of tumor recurrence.
Not applicable.
The research project was designed, organized and
reviewed by IL, and reviewed and critiqued by JK. AK, TS, OK and IL
performed the literature research, wrote the first draft of
manuscript and prepared the figures. Data authentication is not
applicable. All authors read and approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
The authors would like to thank Ms. Militsa Rakina
(Laboratory of Translational Cellular and Molecular Biomedicine,
Tomsk State University, Tomsk, Russia) for the professional English
editing.
The present study was supported by Tomsk State University
Development Programme (Priority-2030; grant no. HY 2.1.1.22
MJI).
|
1
|
Yuan Y, Jiang YC, Sun CK and Chen QM: Role
of the tumor microenvironment in tumor progression and the clinical
applications (Review). Oncol Rep. 35:2499–2515. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Whiteside TL: The tumor microenvironment
and its role in promoting tumor growth. Oncogene. 27:5904–5912.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Werb Z and Lu P: The role of stroma in
tumor development. Cancer J. 21:250–253. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Baghban R, Roshangar L, Jahanban-Esfahlan
R, Seidi K, Ebrahimi-Kalan A, Jaymand M, Kolahian S, Javaheri T and
Zare P: Tumor microenvironment complexity and therapeutic
implications at a glance. Cell Commun Signal. 18:592020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Larionova I, Tuguzbaeva G, Ponomaryova A,
Stakheyeva M, Cherdyntseva N, Pavlov V, Choinzonov E and
Kzhyshkowska J: Tumor-associated macrophages in human breast,
colorectal, lung, ovarian and prostate cancers. Front Oncol.
10:5665112020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lin Y, Xu J and Lan H: Tumor-associated
macrophages in tumor metastasis: Biological roles and clinical
therapeutic applications. J Hematol Oncol. 12:762019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pan Y, Yu Y, Wang X and Zhang T:
Tumor-associated macrophages in tumor immunity. Front Immunol.
11:5830842020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Larionova I, Kazakova E, Patysheva M and
Kzhyshkowska J: Transcriptional, epigenetic and metabolic
programming of tumor-associated macrophages. Cancers (Basel).
12:14112020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gao J, Liang Y and Wang L: Shaping
polarization of tumor-associated macrophages in cancer
immunotherapy. Front Immunol. 13:8887132022. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Quail DF and Joyce JA: Microenvironmental
regulation of tumor progression and metastasis. Nat Med.
19:1423–1437. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Marusyk A, Almendro V and Polyak K:
Intra-tumour heterogeneity: A looking glass for cancer? Nat Rev
Cancer. 12:323–334. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gordillo CH, Sa ndova l P, Muñoz-Her
nández P, Pascual-Antón L, López-Cabrera M and Jiménez-Heffernan
JA: Mesothelial-to-mesenchymal transition contributes to the
generation of carcinoma-associated fibroblasts in locally advanced
primary colorectal carcinomas. Cancers (Basel). 12:4992020.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shiga K, Hara M, Nagasaki T, Sato T,
Takahashi H and Takeyama H: Cancer-associated fibroblasts: Their
characteristics and their roles in tumor growth. Cancers (Basel).
7:2443–2458. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Adjuto-Saccone M, Soubeyran P, Garcia J,
Audebert S, Camoin L, Rubis M, Roques J, Binétruy B, Iovanna JL and
Tournaire R: TNF-α induces endothelial-mesenchymal transition
promoting stromal development of pancreatic adenocarcinoma. Cell
Death Dis. 12:6492021. View Article : Google Scholar
|
|
15
|
Ciszewski WM, Sobierajska K, Wawro ME,
Klopocka W, Chefczyńska N, Muzyczuk A, Siekacz K, Wujkowska A and
Niewiarowska J: The ILK-MMP9-MRTF axis is crucial for EndMT
differentiation of endothelial cells in a tumor microenvironment.
Biochim Biophys Acta Mol Cell Res. 1864:2283–2296. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fan CS, Chen LL, Hsu TA, Chen CC, Chua KV,
Li CP and Huang TS: Endothelial-mesenchymal transition harnesses
HSP90α-secreting M2-macrophages to exacerbate pancreatic ductal
adenocarcinoma. J Hematol Oncol. 12:1382019. View Article : Google Scholar
|
|
17
|
Fan CS, Chen WS, Chen LL, Chen CC, Hsu YT,
Chua KV, Wang HD and Huang TS: Osteopontin-integrin engagement
induces HIF-1α-TCF12-mediated endothelial-mesenchymal transition to
exacerbate colorectal cancer. Oncotarget. 9:4998–5015. 2017.
View Article : Google Scholar
|
|
18
|
Chen C, Li WJ, Weng JJ, Chen ZJ, Wen YY,
Deng T, Le HB, Zhang YK and Zhang BJ: Cancer-associated
fibroblasts, matrix metalloproteinase-9 and lymphatic vessel
density are associated with progression from adenocarcinoma in situ
to invasive adenocarcinoma of the lung. Oncol Lett. 20:1302020.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xie J, Qi X, Wang Y, Yin X, Xu W, Han S,
Cai Y and Han W: Cancer-associated fibroblasts secrete
hypoxia-induced serglycin to promote head and neck squamous cell
carcinoma tumor cell growth in vitro and in vivo by activating the
Wnt/β-catenin pathway. Cell Oncol (Dordr). 44:661–671. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fullár A, Dudás J, Oláh L, Hollósi P, Papp
Z, Sobel G, Karászi K, Paku S, Baghy K and Kovalszky I: Remodeling
of extracellular matrix by normal and tumor-associated fibroblasts
promotes cervical cancer progression. BMC Cancer. 15:2562015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Erdogan B and Webb DJ: Cancer-associated
fibroblasts modulate growth factor signaling and extracellular
matrix remodeling to regulate tumor metastasis. Biochem Soc Trans.
45:229–236. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ren J, Smid M, Iaria J, Salvatori DCF, van
Dam H, Zhu HJ, Martens JWM and Ten Dijke P: Cancer-associated
fibroblast-derived Gremlin 1 promotes breast cancer progression.
Breast Cancer Res. 21:1092019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Takahashi M, Kobayashi H, Mizutani Y, Hara
A, Iida T, Miyai Y, Asai N and Enomoto A: Roles of the mesenchymal
stromal/stem cell marker meflin/Islr in cancer fibrosis. Front Cell
Dev Biol. 9:7499242021. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Attieh Y, Clark AG, Grass C, Richon S,
Pocard M, Mariani P, Elkhatib N, Betz T, Gurchenkov B and Vignjevic
DM: Cancer-associated fibroblasts lead tumor invasion through
integrin-β3-dependent fibronectin assembly. J Cell Biol.
216:3509–3520. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang J, Fu B, Li M and Mi S: Secretome of
activated fibroblasts induced by exosomes for the discovery of
biomarkers in non-small cell lung cancer. Small. 17:e20047502021.
View Article : Google Scholar
|
|
26
|
Glentis A, Oertle P, Mariani P, Chikina A,
El Marjou F, Attieh Y, Zaccarini F, Lae M, Loew D, Dingli F, et al:
Cancer-associated fibroblasts induce metalloprotease-independent
cancer cell invasion of the basement membrane. Nat Commun.
8:9242017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Stadler M, Pudelko K, Biermeier A,
Walterskirchen N, Gaigneaux A, Weindorfer C, Harrer N, Klett H,
Hengstschläger M, Schüler J, et al: Stromal fibroblasts shape the
myeloid phenotype in normal colon and colorectal cancer and induce
CD163 and CCL2 expression in macrophages. Cancer Lett. 520:184–200.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang R, Qi F, Zhao F, Li G, Shao S, Zhang
X, Yuan L and Feng Y: Cancer-associated fibroblasts enhance
tumor-associated macrophages enrichment and suppress NK cells
function in colorectal cancer. Cell Death Dis. 10:2732019.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen S, Morine Y, Tokuda K, Yamada S,
Saito Y, Nishi M, Ikemoto T and Shimada M: Cancer-associated
fibroblast-induced M2-polarized macrophages promote hepatocellular
carcinoma progression via the plasminogen activator inhibitor-1
pathway. Int J Oncol. 59:592021. View Article : Google Scholar :
|
|
30
|
Olingy CE, Dinh HQ and Hedrick CC:
Monocyte heterogeneity and functions in cancer. J Leukoc Biol.
106:309–322. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hao Q, Vadgama JV and Wang P: CCL2/CCR2
signaling in cancer pathogenesis. Cell Commun Signal. 18:822020.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gomez-Roca CA, Italiano A, Le Tourneau C,
Cassier PA, Toulmonde M, D'Angelo SP, Campone M, Weber KL, Loirat
D, Cannarile MA, et al: Phase I study of emactuzumab single agent
or in combination with paclitaxel in patients with
advanced/metastatic solid tumors reveals depletion of
immunosuppressive M2-like macrophages. Ann Oncol. 30:1381–1392.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cannarile MA, Weisser M, Jacob W, Jegg AM,
Ries CH and Rüttinger D: Colony-stimulating factor 1 receptor
(CSF1R) inhibitors in cancer therapy. J Immunother Cancer.
5:532017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cho H, Seo Y, Loke KM, Kim SW, Oh SM, Kim
JH, Soh J, Kim HS, Lee H, Kim J, et al: Cancer-stimulated CAFs
enhance monocyte differentiation and protumoral TAM Activation via
IL6 and GM-CSF secretion. Clin Cancer Res. 24:5407–5421. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xiang H, Ramil CP, Hai J, Zhang C, Wang H,
Watkins AA, Afshar R, Georgiev P, Sze MA, Song XS, et al:
Cancer-associated fibroblasts promote immunosuppression by inducing
ROS-generating monocytic MDSCs in lung squamous cell carcinoma.
Cancer Immunol Res. 8:436–450. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Higashino N, Koma YI, Hosono M, Takase N,
Okamoto M, Kodaira H, Nishio M, Shigeoka M, Kakeji Y and Yokozaki
H: Fibroblast activation protein-positive fibroblasts promote tumor
progression through secretion of CCL2 and interleukin-6 in
esophageal squamous cell carcinoma. Lab Invest. 99:777–792. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gok Yavuz B, Gunaydin G, Gedik ME,
Kosemehmetoglu K, Karakoc D, Ozgur F and Guc D: Cancer associated
fibroblasts sculpt tumour microenvironment by recruiting monocytes
and inducing immunosuppressive PD-1+ TAMs. Sci Rep.
9:31722019. View Article : Google Scholar
|
|
38
|
Yang F, Wei Y, Han D, Li Y, Shi S, Jiao D,
Wu J, Zhang Q, Shi C, Yang L, et al: Interaction with CD68 and
regulation of GAS6 expression by endosialin in fibroblasts drives
recruitment and polarization of macrophages in hepatocellular
carcinoma. Cancer Res. 80:3892–3905. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Allaoui R, Bergenfelz C, Mohlin S,
Hagerling C, Salari K, Werb Z, Anderson RL, Ethier SP, Jirström K,
Påhlman S, et al: Cancer-associated fibroblast-secreted CXCL16
attracts monocytes to promote stroma activation in triple-negative
breast cancers. Nat Commun. 7:130502016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Comito G, Giannoni E, Segura CP,
Barcellos-de-Souza P, Raspollini MR, Baroni G, Lanciotti M, Serni S
and Chiarugi P: Cancer-associated fibroblasts and M2-polarized
macrophages synergize during prostate carcinoma progression.
Oncogene. 33:2423–2431. 2014. View Article : Google Scholar
|
|
41
|
Hegab AE, Ozaki M, Kameyama N, Gao J,
Kagawa S, Yasuda H, Soejima K, Yin Y, Guzy RD, Nakamura Y, et al:
Effect of FGF/FGFR pathway blocking on lung adenocarcinoma and its
cancer-associated fibroblasts. J Pathol. 249:193–205. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang A, Qian Y, Ye Z, Chen H, Xie H, Zhou
L, Shen Y and Zheng S: Cancer-associated fibroblasts promote M2
polarization of macrophages in pancreatic ductal adenocarcinoma.
Cancer Med. 6:463–470. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chua KV, Fan CS, Chen CC, Chen LL, Hsieh
SC and Huang TS: Octyl gallate induces pancreatic ductal
adenocarcinoma cell apoptosis and suppresses
endothelial-mesenchymal transition-promoted M2-macrophages, HSP90α
secretion, and tumor growth. Cells. 9:912019. View Article : Google Scholar
|
|
44
|
Cohen N, Shani O, Raz Y, Sharon Y, Hoffman
D, Abramovitz L and Erez N: Fibroblasts drive an immunosuppressive
and growth-promoting microenvironment in breast cancer via
secretion of Chitinase 3-like 1. Oncogene. 36:4457–4468. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhou Q, Wu X, Wang X, Yu Z, Pan T, Li Z,
Chang X, Jin Z, Li J, Zhu Z, et al: The reciprocal interaction
between tumor cells and activated fibroblasts mediated by
TNF-α/IL-33/ST2L signaling promotes gastric cancer metastasis.
Oncogene. 39:1414–1428. 2020. View Article : Google Scholar
|
|
46
|
Andersson P, Yang Y, Hosaka K, Zhang Y,
Fischer C, Braun H, Liu S, Yu G, Liu S, Beyaert R, et al: Molecular
mechanisms of IL-33-mediated stromal interactions in cancer
metastasis. JCI Insight. 3:e1223752018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Mazur A, Holthoff E, Vadali S, Kelly T and
Post SR: Cleavage of type I collagen by fibroblast activation
protein-α enhances class A scavenger receptor mediated macrophage
adhesion. PLoS One. 11:e01502872016. View Article : Google Scholar
|
|
48
|
Cat B, Stuhlmann D, Steinbrenner H, Alili
L, Holtkötter O, Sies H and Brenneisen P: Enhancement of tumor
invasion depends on transdifferentiation of skin fibroblasts
mediated by reactive oxygen species. J Cell Sci. 119:2727–2738.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yang X, Lin Y, Shi Y, Li B, Liu W, Yin W,
Dang Y, Chu Y, Fan J and He R: FAP promotes immunosuppression by
cancer-associated fibroblasts in the tumor microenvironment via
STAT3-CCL2 signaling. Cancer Res. 76:4124–4135. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Potente M and Carmeliet P: The Link
between angiogenesis and endothelial metabolism. Annu Rev Physiol.
79:43–66. 2017. View Article : Google Scholar
|
|
51
|
Papetti M and Herman IM: Mechanisms of
normal and tumor-derived angiogenesis. Am J Physiol Cell Physiol.
282:C947–C970. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Hida K and Maishi N: Abnormalities of
tumor endothelial cells and cancer progression. Oral Sci Int.
15:1–6. 2018. View Article : Google Scholar
|
|
53
|
Hida K, Maishi N, Takeda R and Hida Y: The
roles of tumor endothelial cells in cancer metastasis. Metastasis
Brisbane (AU): Sergi CM: Exon Publications; 2022, View Article : Google Scholar
|
|
54
|
Wang WY, Lin D, Jarman EH, Polacheck WJ
and Baker BM: Functional angiogenesis requires microenvironmental
cues balancing endothelial cell migration and proliferation. Lab
Chip. 20:1153–1166. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Yang D, Guo P, He T and Powell CA: Role of
endothelial cells in tumor microenvironment. Clin Transl Med.
11:e4502021. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wei K, Ma Z, Yang F, Zhao X, Jiang W, Pan
C, Li Z, Pan X, He Z, Xu J, et al: M2 macrophage-derived exosomes
promote lung adenocarcinoma progression by delivering miR-942.
Cancer Lett. 526:205–216. 2022. View Article : Google Scholar
|
|
57
|
Baradaran A, Asadzadeh Z, Hemmat N,
Baghbanzadeh A, Shadbad MA, Khosravi N, Derakhshani A, Alemohammad
H, Afrashteh Nour M, Safarpour H, et al: The cross-talk between
tumor-associated macrophages and tumor endothelium: Recent advances
in macrophage-based cancer immunotherapy. Biomed Pharmacother.
146:1125882022. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Laoui D, Van Overmeire E, Di Conza G,
Aldeni C, Keirsse J, Morias Y, Movahedi K, Houbracken I, Schouppe
E, Elkrim Y, et al: Tumor hypoxia does not drive differentiation of
tumor-associated macrophages but rather fine-tunes the M2-like
macrophage population. Cancer Res. 74:24–30. 2014. View Article : Google Scholar
|
|
59
|
Hughes R, Qian BZ, Rowan C, Muthana M,
Keklikoglou I, Olson OC, Tazzyman S, Danson S, Addison C, Clemons
M, et al: Perivascular M2 macrophages stimulate tumor relapse after
chemotherapy. Cancer Res. 75:3479–3491. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Cheng N, Bei Y, Song Y, Zhang W, Xu L,
Zhang W, Yang N, Bai X, Shu Y and Shen P: B7-H3 augments the
pro-angiogenic function of tumor-associated macrophages and acts as
a novel adjuvant target for triple-negative breast cancer therapy.
Biochem Pharmacol. 183:1142982021. View Article : Google Scholar
|
|
61
|
Treps L, Faure S and Clere N: Vasculogenic
mimicry, a complex and devious process favoring
tumorigenesis-Interest in making it a therapeutic target. Pharmacol
Ther. 223:1078052021. View Article : Google Scholar
|
|
62
|
Zhang L, Xu Y, Sun J, Chen W, Zhao L, Ma
C, Wang Q, Sun J, Huang B, Zhang Y, et al: M2-like tumor-associated
macrophages drive vasculogenic mimicry through amplification of
IL-6 expression in glioma cells. Oncotarget. 8:819–832. 2017.
View Article : Google Scholar :
|
|
63
|
Delprat V and Michiels C: A bi-directional
dialog between vascular cells and monocytes/macrophages regulates
tumor progression. Cancer Metastasis Rev. 40:477–500. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Cortes-Santiago N, Hossain MB,
Gabrusiewicz K, Fan X, Gumin J, Marini FC, Alonso MM, Lang F, Yung
WK, Fueyo J, et al: Soluble Tie2 overrides the heightened invasion
induced by anti-angiogenesis therapies in gliomas. Oncotarget.
7:16146–16157. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Sopo M, Sallinen H, Hämäläinen K, Kivelä
A, Ylä-Herttuala S, Kosma VM, Keski-Nisula L and Anttila M: High
expression of Tie-2 predicts poor prognosis in primary high grade
serous ovarian cancer. PLoS One. 15:e02414842020. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Dong Z, Chen J, Yang X, Zheng W, Wang L,
Fang M, Wu M, Yao M and Yao D: Ang-2 promotes lung cancer
metastasis by increasing epithelial-mesenchymal transition.
Oncotarget. 9:12705–12717. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Hidalgo M, Martinez-Garcia M, Le Tourneau
C, Massard C, Garralda E, Boni V, Taus A, Albanell J, Sablin MP,
Alt M, et al: First-in-human phase I study of single-agent
vanucizumab, a first-in-class bispecific
anti-angiopoietin-2/anti-VEGF-A antibody, in adult patients with
advanced solid tumors. Clin Cancer Res. 24:1536–1545. 2018.
View Article : Google Scholar
|
|
68
|
Karikoski M, Irjala H, Maksimow M,
Miiluniemi M, Granfors K, Hernesniemi S, Elima K, Moldenhauer G,
Schledzewski K, Kzhyshkowska J, et al: Clever-1/stabilin-1
regulates lymphocyte migration within lymphatics and leukocyte
entrance to sites of inflammation. Eur J Immunol. 39:3477–3487.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Karikoski M, Marttila-Ichihara F, Elima K,
Rantakari P, Hollmén M, Kelkka T, Gerke H, Huovinen V, Irjala H,
Holmdahl R, et al: Clever-1/stabilin-1 controls cancer growth and
metastasis. Clin Cancer Res. 20:6452–6464. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Riabov V, Yin S, Song B, Avdic A,
Schledzewski K, Ovsiy I, Gratchev A, Llopis Verdiell M, Sticht C,
Schmuttermaier C, et al: Stabilin-1 is expressed in human breast
cancer and supports tumor growth in mammary adenocarcinoma mouse
model. Oncotarget. 7:31097–31110. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Yin M, Zhou HJ, Zhang J, Lin C, Li H, Li
X, Li Y, Zhang H, Breckenridge DG, Ji W and Min W: ASK1-dependent
endothelial cell activation is critical in ovarian cancer growth
and metastasis. JCI Insight. 2:e918282017. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Arwert EN, Harney AS, Entenberg D, Wang Y,
Sahai E, Pollard JW and Condeelis JS: A unidirectional transition
from migratory to perivascular macrophage is required for tumor
cell intravasation. Cell Rep. 23:1239–1248. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Leung E, Xue A, Wang Y, Rougerie P, Sharma
VP, Eddy R, Cox D and Condeelis J: Blood vessel
endothelium-directed tumor cell streaming in breast tumors requires
the HGF/C-Met signaling pathway. Oncogene. 36:2680–2692. 2017.
View Article : Google Scholar :
|
|
74
|
Opzoomer JW, Anstee JE, Dean I, Hill EJ,
Bouybayoune I, Caron J, Muliaditan T, Gordon P, Sosnowska D, Nuamah
R, et al: Macrophages orchestrate the expansion of a proangiogenic
perivascular niche during cancer progression. Sci Adv.
7:eabg95182021. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Lewis CE, Harney AS and Pollard JW: The
multifaceted role of perivascular macrophages in tumors. Cancer
Cell. 30:3652016. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Sparano JA, Gray R, Oktay MH, Entenberg D,
Rohan T, Xue X, Donovan M, Peterson M, Shuber A, Hamilton DA, et
al: A metastasis biomarker (MetaSite Breast™ Score) is associated
with distant recurrence in hormone receptor-positive, HER2-negative
early-stage breast cancer. NPJ Breast Cancer. 3:422017. View Article : Google Scholar
|
|
77
|
Anstee JE, Opzoomer JW, Dean I, Muller HP,
Bahri M, Liakath-Ali K, Liu Z, Choy D, Caron J, Sosnowska D, et al:
Perivascular macrophages collaborate to facilitate chemotherapy
resistance in cancer. bioRxiv. 2022.2002.2003.478952. 2022.
|
|
78
|
Hongu T, Pein M, Insua-Rodríguez J,
Gutjahr E, Mattavelli G, Meier J, Decker K, Descot A, Bozza M,
Harbottle R, et al: Perivascular tenascin C triggers sequential
activation of macrophages and endothelial cells to generate a
pro-metastatic vascular niche in the lungs. Nat Cancer. 3:486–504.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
He H, Mack JJ, Güç E, Warren CM, Squadrito
ML, Kilarski WW, Baer C, Freshman RD, McDonald AI, Ziyad S, et al:
Perivascular macrophages limit permeability. Arterioscler Thromb
Vasc Biol. 36:2203–2212. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Wang Q, He Z, Huang M, Liu T, Wang Y, Xu
H, Duan H, Ma P, Zhang L, Zamvil SS, et al: Vascular niche IL-6
induces alternative macrophage activation in glioblastoma through
HIF-2α. Nat Commun. 9:5592018. View Article : Google Scholar
|
|
81
|
Chen L, Li J, Wang F, Dai C, Wu F, Liu X,
Li T, Glauben R, Zhang Y, Nie G, et al: Tie2 expression on
macrophages is required for blood vessel reconstruction and tumor
relapse after chemotherapy. Cancer Res. 76:6828–6838. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Harney AS, Arwert EN, Entenberg D, Wang Y,
Guo P, Qian BZ, Oktay MH, Pollard JW, Jones JG and Condeelis JS:
Real-time imaging reveals local, transient vascular permeability,
and tumor cell intravasation stimulated by TIE2hi
macrophage-derived VEGFA. Cancer Discov. 5:932–943. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Bieniasz-Krzywiec P, Martín-Pérez R,
Ehling M, García-Caballero M, Pinioti S, Pretto S, Kroes R, Aldeni
C, Di Matteo M, Prenen H, et al: Podoplanin-expressing macrophages
promote lymphangiogenesis and lymphoinvasion in breast cancer. Cell
Metab. 30:917–936.e10. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Elder AM, Tamburini BAJ, Crump LS, Black
SA, Wessells VM, Schedin PJ, Borges VF and Lyons TR: Semaphorin 7A
promotes macrophage-mediated lymphatic remodeling during postpartum
mammary gland involution and in breast cancer. Cancer Res.
78:6473–6485. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Evans R, Flores-Borja F, Nassiri S,
Miranda E, Lawler K, Grigoriadis A, Monypenny J, Gillet C, Owen J,
Gordon P, et al: Integrin-mediated macrophage adhesion promotes
lymphovascular dissemination in breast cancer. Cell Rep.
27:1967–1978.e4. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Bron S, Henry L, Faes-Van't Hull E,
Turrini R, Vanhecke D, Guex N, Ifticene-Treboux A, Marina Iancu E,
Semilietof A, Rufer N, et al: TIE-2-expressing monocytes are
lymphangiogenic and associate specifically with lymphatics of human
breast cancer. Oncoimmunology. 5:e10738822015. View Article : Google Scholar
|
|
87
|
Moussai D, Mitsui H, Pettersen JS, Pierson
KC, Shah KR, Suárez-Fariñas M, Cardinale IR, Bluth MJ, Krueger JG
and Carucci JA: The human cutaneous squamous cell carcinoma
microenvironment is characterized by increased lymphatic density
and enhanced expression of macrophage-derived VEGF-C. J Invest
Dermatol. 131:229–236. 2011. View Article : Google Scholar
|
|
88
|
Zhang BC, Gao J, Wang J, Rao ZG, Wang BC
and Gao JF: Tumor-associated macrophages infiltration is associated
with peritumoral lymphangiogenesis and poor prognosis in lung
adenocarcinoma. Med Oncol. 28:1447–1452. 2011. View Article : Google Scholar
|
|
89
|
Zhang B, Yao G, Zhang Y and Gao J, Yang B,
Rao Z and Gao J: M2-polarized tumor-associated macrophages are
associated with poor prognoses resulting from accelerated
lymphangiogenesis in lung adenocarcinoma. Clinics (Sao Paulo).
66:1879–1886. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Yamagata Y, Tomioka H, Sakamoto K, Sato K,
Harada H, Ikeda T and Kayamori K: CD163-positive macrophages within
the tumor stroma are associated with lymphangiogenesis and lymph
node metastasis in oral squamous cell carcinoma. J Oral Maxillofac
Surg. 75:2144–2153. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Kurahara H, Takao S, Maemura K, Mataki Y,
Kuwahata T, Maeda K, Sakoda M, Iino S, Ishigami S, Ueno S, et al:
M2-polarized tumor-associated macrophage infiltration of regional
lymph nodes is associated with nodal lymphangiogenesis and occult
nodal involvement in pN0 pancreatic cancer. Pancreas. 42:155–159.
2013. View Article : Google Scholar
|
|
92
|
Wu H, Xu JB, He YL, Peng JJ, Zhang XH,
Chen CQ, Li W and Cai SR: Tumor-associated macrophages promote
angiogenesis and lymphangiogenesis of gastric cancer. J Surg Oncol.
106:462–468. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Ding H, Cai J, Mao M, Fang Y, Huang Z, Jia
J, Li T, Xu L, Wang J, Zhou J, et al: Tumor-associated macrophages
induce lymphangiogenesis in cervical cancer via interaction with
tumor cells. APMIS. 122:1059–1069. 2014.PubMed/NCBI
|
|
94
|
Utrera-Barillas D, Castro-Manrreza M,
Castellanos E, Gutiérrez-Rodríguez M, Arciniega-Ruíz de Esparza O,
García-Cebada J, Velazquez JR, Flores-Reséndiz D,
Hernández-Hernández D and Benítez-Bribiesca L: The role of
macrophages and mast cells in lymphangiogenesis and angiogenesis in
cervical carcinogenesis. Exp Mol Pathol. 89:190–196. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Zhang W, Tian J and Hao Q: HMGB1 combining
with tumor-associated macrophages enhanced lymphangiogenesis in
human epithelial ovarian cancer. Tumour Biol. 35:2175–2186. 2014.
View Article : Google Scholar
|
|
96
|
Sweat RS, Sloas DC and Murfee WL: VEGF-C
induces lymphangiogenesis and angiogenesis in the rat mesentery
culture model. Microcirculation. 21:532–540. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Ji H, Cao R, Yang Y, Zhang Y, Iwamoto H,
Lim S, Nakamura M, Andersson P, Wang J, Sun Y, et al: TNFR1
mediates TNF-α-induced tumour lymphangiogenesis and metastasis by
modulating VEGF-C-VEGFR3 signalling. Nat Commun. 5:49442014.
View Article : Google Scholar
|
|
98
|
Peppicelli S, Bianchini F and Calorini L:
Inflammatory cytokines induce vascular endothelial growth factor-C
expression in melanoma-associated macrophages and stimulate
melanoma lymph node metastasis. Oncol Lett. 8:1133–1138. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Schoppmann SF, Birner P, Stöckl J, Kalt R,
Ullrich R, Caucig C, Kriehuber E, Nagy K, Alitalo K and Kerjaschki
D: Tumor-associated macrophages express lymphatic endothelial
growth factors and are related to peritumoral lymphangiogenesis. Am
J Pathol. 161:947–956. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Mazzone M and Bergers G: Regulation of
blood and lymphatic vessels by immune cells in tumors and
metastasis. Annu Rev Physiol. 81:535–560. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Weichand B, Popp R, Dziumbla S, Mora J,
Strack E, Elwakeel E, Frank AC, Scholich K, Pierre S, Syed SN, et
al: S1PR1 on tumor-associated macrophages promotes
lymphangiogenesis and metastasis via NLRP3/IL-1β. J Exp Med.
214:2695–2713. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Watari K, Shibata T, Kawahara A, Sata K,
Nabeshima H, Shinoda A, Abe H, Azuma K, Murakami Y, Izumi H, et al:
Tumor-derived interleukin-1 promotes lymphangiogenesis and lymph
node metastasis through M2-type macrophages. PLoS One.
9:e995682014. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Jung M, Ören B, Mora J, Mertens C,
Dziumbla S, Popp R, Weigert A, Grossmann N, Fleming I and Brüne B:
Lipocalin 2 from macrophages stimulated by tumor cell-derived
sphingosine 1-phosphate promotes lymphangiogenesis and tumor
metastasis. Sci Signal. 9:ra642016. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Storr SJ, Safuan S, Ahmad N, El-Refaee M,
Jackson AM and Martin SG: Macrophage-derived interleukin-1beta
promotes human breast cancer cell migration and lymphatic adhesion
in vitro. Cancer Immunol Immunother. 66:1287–1294. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Chen C, He W, Huang J, Wang B, Li H, Cai
Q, Su F, Bi J, Liu H, Zhang B, et al: LNMAT1 promotes lymphatic
metastasis of bladder cancer via CCL2 dependent macrophage
recruitment. Nat Commun. 9:38262018. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Tauchi Y, Tanaka H, Kumamoto K, Tokumoto
M, Sakimura C, Sakurai K, Kimura K, Toyokawa T, Amano R, Kubo N, et
al: Tumor-associated macrophages induce capillary morphogenesis of
lymphatic endothelial cells derived from human gastric cancer.
Cancer Sci. 107:1101–1109. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Kabasawa T, Ohe R, Aung NY, Urano Y,
Kitaoka T, Tamazawa N, Utsunomiya A and Yamakawa M: Potential role
of M2 TAMs around lymphatic vessels during lymphatic invasion in
papillary thyroid carcinoma. Sci Rep. 11:11502021. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Chen XJ, Wei WF, Wang ZC, Wang N, Guo CH,
Zhou CF, Liang LJ, Wu S, Liang L and Wang W: A novel lymphatic
pattern promotes metastasis of cervical cancer in a hypoxic
tumour-associated macrophage-dependent manner. Angiogenesis.
24:549–565. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Ding M, Fu X, Tan H, Wang R, Chen Z and
Ding S: The effect of vascular endothelial growth factor C
expression in tumor-associated macrophages on lymphangiogenesis and
lymphatic metastasis in breast cancer. Mol Med Rep. 6:1023–1029.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Ran S and Wilber A: Novel role of immature
myeloid cells in formation of new lymphatic vessels associated with
inflammation and tumors. J Leukoc Biol. 102:253–263. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Ran S and Volk-Draper L: Lymphatic
endothelial cell progenitors in the tumor microenvironment. Adv Exp
Med Biol. 1234:87–105. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Ran S and Montgomery KE:
Macrophage-mediated lymphangiogenesis: The emerging role of
macrophages as lymphatic endothelial progenitors. Cancers (Basel).
4:618–657. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Volk-Draper L, Patel R, Bhattarai N, Yang
J, Wilber A, DeNardo D and Ran S: Myeloid-derived lymphatic
endothelial cell progenitors significantly contribute to lymphatic
metastasis in clinical breast cancer. Am J Pathol. 189:2269–2292.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Gordon EJ, Rao S, Pollard JW, Nutt SL,
Lang RA and Harvey NL: Macrophages define dermal lymphatic vessel
calibre during development by regulating lymphatic endothelial cell
proliferation. Development. 137:3899–3910. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Dannenmann SR, Thielicke J, Stöckli M,
Matter C, von Boehmer L, Cecconi V, Hermanns T, Hefermehl L,
Schraml P, Moch H, et al: Tumor-associated macrophages subvert
T-cell function and correlate with reduced survival in clear cell
renal cell carcinoma. Oncoimmunology. 2:e235622013. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Gordon S and Martinez FO: Alternative
activation of macrophages: Mechanism and functions. Immunity.
32:593–604. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
He Z and Zhang S: Tumor-associated
macrophages and their functional transformation in the hypoxic
tumor microenvironment. Front Immunol. 12:7413052021. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Zhang L, Zhang K, Zhang J, Zhu J, Xi Q,
Wang H, Zhang Z, Cheng Y, Yang G, Liu H, et al: Loss of fragile
site-associated tumor suppressor promotes antitumor immunity via
macrophage polarization. Nat Commun. 12:43002021. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Wang W, Marinis JM, Beal AM, Savadkar S,
Wu Y, Khan M, Taunk PS, Wu N, Su W, Wu J, et al: RIP1 kinase drives
macrophage-mediated adaptive immune tolerance in pancreatic cancer.
Cancer Cell. 34:757–774.e7. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Ho TTB, Nasti A, Seki A, Komura T, Inui H,
Kozaka T, Kitamura Y, Shiba K, Yamashita T, Yamashita T, et al:
Combination of gemcitabine and anti-PD-1 antibody enhances the
anticancer effect of M1 macrophages and the Th1 response in a
murine model of pancreatic cancer liver metastasis. J Immunother
Cancer. 8:e0013672020. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Klug F, Prakash H, Huber PE, Seibel T,
Bender N, Halama N, Pfirschke C, Voss RH, Timke C, Umansky L, et
al: Low-dose irradiation programs macrophage differentiation to an
iNOS+/M1 phenotype that orchestrates effective T cell
immunotherapy. Cancer Cell. 24:589–602. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Shiao SL, Ruffell B, DeNardo DG, Faddegon
BA, Park CC and Coussens LM: TH2-polarized CD4(+) T cells and
macrophages limit efficacy of radiotherapy. Cancer Immunol Res.
3:518–525. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
DeNardo DG, Barreto JB, Andreu P, Vasquez
L, Tawfik D, Kolhatkar N and Coussens LM: CD4(+) T cells regulate
pulmonary metastasis of mammary carcinomas by enhancing protumor
properties of macrophages. Cancer Cell. 16:91–102. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Yasuda K, Takeuchi Y and Hirota K: The
pathogenicity of Th17 cells in autoimmune diseases. Semin
Immunopathol. 41:283–297. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Chang SH: T helper 17 (Th17) cells and
interleukin-17 (IL-17) in cancer. Arch Pharm Res. 42:549–559. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Kryczek I, Banerjee M, Cheng P, Vatan L,
Szeliga W, Wei S, Huang E, Finlayson E, Simeone D, Welling TH, et
al: Phenotype, distribution, generation, and functional and
clinical relevance of Th17 cells in the human tumor environments.
Blood. 114:1141–1149. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Zhou J, Li X, Wu X, Zhang T, Zhu Q and
Wang X, Wang H, Wang K, Lin Y and Wang X: Exosomes released from
tumor-associated macrophages transfer miRNAs that induce a
Treg/Th17 cell imbalance in epithelial ovarian cancer. Cancer
Immunol Res. 6:1578–1592. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Yang JY, Jie Z, Mathews A, Zhou X, Li Y,
Gu M, Xie X, Ko CJ, Cheng X, Qi Y, et al: Intestinal epithelial
tbk1 prevents differentiation of T-helper 17 cells and
tumorigenesis in mice. Gastroenterology. 159:1793–1806. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Maruyama T, Kono K, Mizukami Y, Kawaguchi
Y, Mimura K, Watanabe M, Izawa S and Fujii H: Distribution of Th17
cells and FoxP3(+) regulatory T cells in tumor-infiltrating
lymphocytes, tumor-draining lymph nodes and peripheral blood
lymphocytes in patients with gastric cancer. Cancer Sci.
101:1947–1954. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Knochelmann HM, Dwyer CJ, Bailey SR, Amaya
SM, Elston DM, Mazza-McCrann JM and Paulos CM: When worlds collide:
Th17 and Treg cells in cancer and autoimmunity. Cell Mol Immunol.
15:458–469. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Ohue Y and Nishikawa H: Regulatory T
(Treg) cells in cancer: Can Treg cells be a new therapeutic target?
Cancer Sci. 110:2080–2089. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Whiteside TL: What are regulatory T cells
(Treg) regulating in cancer and why? Semin Cancer Biol. 22:327–334.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Tanaka A and Sakaguchi S: Regulatory T
cells in cancer immunotherapy. Cell Res. 27:109–118. 2017.
View Article : Google Scholar :
|
|
134
|
Waniczek D, Lorenc Z, Śnietura M, Wesecki
M, Kopec A and Muc-Wierzgoń M: Tumor-associated macrophages and
regulatory T cells infiltration and the clinical outcome in
colorectal cancer. Arch Immunol Ther Exp (Warsz). 65:445–454. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
135
|
La Fleur L, Botling J, He F, Pelicano C,
Zhou C, He C, Palano G, Mezheyeuski A, Micke P, Ravetch JV, et al:
Targeting MARCO and IL37R on immunosuppressive macrophages in lung
cancer blocks regulatory T cells and supports cytotoxic lymphocyte
function. Cancer Res. 81:956–967. 2021. View Article : Google Scholar
|
|
136
|
Zhu Q, Wu X, Wu Y and Wang X: Interaction
between Treg cells and tumor-associated macrophages in the tumor
microenvironment of epithelial ovarian cancer. Oncol Rep.
36:3472–3478. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Wu Q, Zhou W, Yin S, Zhou Y, Chen T, Qian
J, Su R, Hong L, Lu H, Zhang F, et al: Blocking triggering receptor
expressed on myeloid cells-1-positive tumor-associated macrophages
induced by hypoxia reverses immunosuppression and anti-programmed
cell death ligand 1 resistance in liver cancer. Hepatology.
70:198–214. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Fujimura T, Kambayashi Y, Fujisawa Y,
Hidaka T and Aiba S: Tumor-associated macrophages: Therapeutic
targets for skin cancer. Front Oncol. 8:32018. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Furudate S, Fujimura T, Kambayashi Y,
Kakizaki A, Hidaka T and Aiba S: Immunomodulatory effect of
imiquimod through CCL22 produced by tumor-associated macrophages in
B16F10 melanomas. Anticancer Res. 37:3461–3471. 2017.PubMed/NCBI
|
|
140
|
Wang D, Yang L, Yue D, Cao L, Li L, Wang
D, Ping Y, Shen Z, Zheng Y, Wang L and Zhang Y: Macrophage-derived
CCL22 promotes an immunosuppressive tumor microenvironment via IL-8
in malignant pleural effusion. Cancer Lett. 452:244–253. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Carbó JM, León TE, Font-Díaz J, De la Rosa
JV, Castrillo A, Picard FR, Staudenraus D, Huber M, Cedó L,
Escolà-Gil JC, et al: Pharmacologic activation of LXR alters the
expression profile of tumor-associated macrophages and the
abundance of regulatory T cells in the tumor microenvironment.
Cancer Res. 81:968–985. 2021. View Article : Google Scholar
|
|
142
|
Liu J, Zhang N, Li Q, Zhang W, Ke F, Leng
Q and Wang H, Chen J and Wang H: Tumor-associated macrophages
recruit CCR6+ regulatory T cells and promote the development of
colorectal cancer via enhancing CCL20 production in mice. PLoS One.
6:e194952011. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Yang S, Wang B, Guan C, Wu B, Cai C, Wang
M, Zhang B, Liu T and Yang P: Foxp3+IL-17+ T cells promote
development of cancer-initiating cells in colorectal cancer. J
Leukoc Biol. 89:85–91. 2011. View Article : Google Scholar
|
|
144
|
Liu C, Chikina M, Deshpande R, Menk AV,
Wang T, Tabib T, Brunazzi EA, Vignali KM, Sun M, Stolz DB, et al:
Treg cells promote the SREBP1-dependent metabolic fitness of
tumor-promoting macrophages via repression of CD8+ T
cell-derived interferon-γ. Immunity. 51:381–397.e6. 2019.
View Article : Google Scholar
|
|
145
|
Fu Q, Xu L, Wang Y, Jiang Q, Liu Z, Zhang
J, Zhou Q, Zeng H, Tong S, Wang T, et al: Tumor-associated
macrophage-derived Interleukin-23 interlinks kidney cancer
glutamine addiction with immune evasion. Eur Urol. 75:752–763.
2019. View Article : Google Scholar
|
|
146
|
Sun W, Wei FQ, Li WJ, Wei JW, Zhong H, Wen
YH, Lei WB, Chen L, Li H, Lin HQ, et al: A positive-feedback loop
between tumour infiltrating activated Treg cells and type 2-skewed
macrophages is essential for progression of laryngeal squamous cell
carcinoma. Br J Cancer. 117:1631–1643. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Campesato LF, Budhu S, Tchaicha J, Weng
CH, Gigoux M, Cohen IJ, Redmond D, Mangarin L, Pourpe S, Liu C, et
al: Blockade of the AHR restricts a Treg-macrophage suppressive
axis induced by L-Kynurenine. Nat Commun. 11:40112020. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Raskov H, Orhan A, Christensen JP and
Gögenur I: Cytotoxic CD8+ T cells in cancer and cancer
immunotherapy. Br J Cancer. 124:359–367. 2021. View Article : Google Scholar
|
|
149
|
Reiser J and Banerjee A: Effector, memory,
and dysfunctional CD8(+) T cell fates in the antitumor immune
response. J Immunol Res. 2016:89412602016. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Peranzoni E, Lemoine J, Vimeux L, Feuillet
V, Barrin S, Kantari-Mimoun C, Bercovici N, Guérin M, Biton J,
Ouakrim H, et al: Macrophages impede CD8 T cells from reaching
tumor cells and limit the efficacy of anti-PD-1 treatment. Proc
Natl Acad Sci USA. 115:E4041–E4050. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Garrido-Martin EM, Mellows TWP, Clarke J,
Ganesan AP, Wood O, Cazaly A, Seumois G, Chee SJ, Alzetani A, King
EV, et al: M1hot tumor-associated macrophages boost
tissue-resident memory T cells infiltration and survival in human
lung cancer. J Immunother Cancer. 8:e0007782020. View Article : Google Scholar
|
|
152
|
Dai X, Lu L, Deng S, Meng J, Wan C, Huang
J, Sun Y, Hu Y, Wu B, Wu G, et al: USP7 targeting modulates
anti-tumor immune response by reprogramming tumor-associated
macrophages in lung cancer. Theranostics. 10:9332–9347. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Casanova-Acebes M, Dalla E, Leader AM,
LeBerichel J, Nikolic J, Morales BM, Brown M, Chang C, Troncoso L,
Chen ST, et al: Tissue-resident macrophages provide a
pro-tumorigenic niche to early NSCLC cells. Nature. 595:578–584.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Yao W, Ba Q, Li X, Li H, Zhang S, Yuan Y,
Wang F, Duan X, Li J, Zhang W and Wang H: A natural CCR2 antagonist
relieves tumor-associated macrophage-mediated immunosuppression to
produce a therapeutic effect for liver cancer. EBioMedicine.
22:58–67. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Dolina JS, Van Braeckel-Budimir N, Thomas
GD and Salek-Ardakani S: CD8+ T cell exhaustion in
cancer. Front Immunol. 12:7152342021. View Article : Google Scholar
|
|
156
|
Dangaj D, Bruand M, Grimm AJ, Ronet C,
Barras D, Duttagupta PA, Lanitis E, Duraiswamy J, Tanyi JL,
Benencia F, et al: Cooperation between constitutive and inducible
chemokines enables T cell engraftment and immune attack in solid
tumors. Cancer Cell. 35:885–900.e10. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
House IG, Savas P, Lai J, Chen AXY, Oliver
AJ, Teo ZL, Todd KL, Henderson MA, Giuffrida L, Petley EV, et al:
Macrophage-derived CXCL9 and CXCL10 are required for antitumor
immune responses following immune checkpoint blockade. Clin Cancer
Res. 26:487–504. 2020. View Article : Google Scholar
|
|
158
|
Petty AJ, Li A, Wang X, Dai R, Heyman B,
Hsu D, Huang X and Yang Y: Hedgehog signaling promotes
tumor-associated macrophage polarization to suppress intratumoral
CD8+ T cell recruitment. J Clin Invest. 129:5151–5162. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Pascual-García M, Bonfill-Teixidor E,
Planas-Rigol E, Rubio-Perez C, Iurlaro R, Arias A, Cuartas I,
Sala-Hojman A, Escudero L, Martínez-Ricarte F, et al: LIF regulates
CXCL9 in tumor-associated macrophages and prevents CD8+
T cell tumor-infiltration impairing anti-PD1 therapy. Nat Commun.
10:24162019. View Article : Google Scholar
|
|
160
|
Yu J, Green MD, Li S, Sun Y, Journey SN,
Choi JE, Rizvi SM, Qin A, Waninger JJ, Lang X, et al: Liver
metastasis restrains immunotherapy efficacy via macrophage-mediated
T cell elimination. Nat Med. 27:152–164. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Kusmartsev S and Gabrilovich DI: STAT1
signaling regulates tumor-associated macrophage-mediated T cell
deletion. J Immunol. 174:4880–4891. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Stanley ER and Chitu V: CSF-1 receptor
signaling in myeloid cells. Cold Spring Harb Perspect Biol.
6:a0218572014. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Kato Y, Tabata K, Kimura T,
Yachie-Kinoshita A, Ozawa Y, Yamada K, Ito J, Tachino S, Hori Y,
Matsuki M, et al: Lenvatinib plus anti-PD-1 antibody combination
treatment activates CD8+ T cells through reduction of
tumor-associated macrophage and activation of the interferon
pathway. PLoS One. 14:e02125132019. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Mehta AK, Cheney EM, Hartl CA, Pantelidou
C, Oliwa M, Castrillon JA, Lin JR, Hurst KE, de Oliveira Taveira M,
Johnson NT, et al: Targeting immunosuppressive macrophages
overcomes PARP inhibitor resistance in BRCA1-associated
triple-negative breast cancer. Nat Cancer. 2:66–82. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Xiang W, Shi R, Kang X, Zhang X, Chen P,
Zhang L, Hou A, Wang R, Zhao Y, Zhao K, et al: Monoacylglycerol
lipase regulates cannabinoid receptor 2-dependent macrophage
activation and cancer progression. Nat Commun. 9:25742018.
View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Chen EP, Markosyan N, Connolly E, Lawson
JA, Li X, Grant GR, Grosser T, FitzGerald GA and Smyth EM: Myeloid
cell COX-2 deletion reduces mammary tumor growth through enhanced
cytotoxic T-lymphocyte function. Carcinogenesis. 35:1788–1797.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Li L, Han L, Sun F, Zhou J, Ohaegbulam KC,
Tang X, Zang X, Steinbrecher KA, Qu Z and Xiao G: NF-κB RelA
renders tumor-associated macrophages resistant to and capable of
directly suppressing CD8+ T cells for tumor promotion.
Oncoimmunology. 7:e14352502018. View Article : Google Scholar
|
|
168
|
Kaneda MM, Messer KS, Ralainirina N, Li H,
Leem CJ, Gorjestani S, Woo G, Nguyen AV, Figueiredo CC, Foubert P,
et al: PI3Kγ is a molecular switch that controls immune
suppression. Nature. 539:437–442. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
McIntire RH and Hunt JS: Antigen
presenting cells and HLA-G-a review. Placenta. 26(Suppl A):
S104–S109. 2005. View Article : Google Scholar
|
|
170
|
Nixon BG, Kuo F, Liu M, Capistrano K, Do
M, Franklin RA, Wu X, Kansler ER, Srivastava RM, Purohit TA, et al:
IRF8 governs tumor-associated macrophage control of T cell
exhaustion. bioRxiv. 2020.2003.2012.989731. 2020.
|
|
171
|
Tseng D, Volkmer JP, Willingham SB,
Contreras-Trujillo H, Fathman JW, Fernhoff NB, Seita J, Inlay MA,
Weiskopf K, Miyanishi M and Weissman IL: Anti-CD47
antibody-mediated phagocytosis of cancer by macrophages primes an
effective antitumor T-cell response. Proc Natl Acad Sci USA.
110:11103–11108. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Ruffell B, Chang-Strachan D, Chan V,
Rosenbusch A, Ho CM, Pryer N, Daniel D, Hwang ES, Rugo HS and
Coussens LM: Macrophage IL-10 blocks CD8+ T cell-dependent
responses to chemotherapy by suppressing IL-12 expression in
intratumoral dendritic cells. Cancer Cell. 26:623–637. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Fang W, Zhou T, Shi H, Yao M, Zhang D,
Qian H, Zeng Q, Wang Y, Jin F, Chai C and Chen T: Progranulin
induces immune escape in breast cancer via up-regulating PD-L1
expression on tumor-associated macrophages (TAMs) and promoting
CD8+ T cell exclusion. J Exp Clin Cancer Res. 40:42021.
View Article : Google Scholar
|
|
174
|
Lin H, Wei S, Hurt EM, Green MD, Zhao L,
Vatan L, Szeliga W, Herbst R, Harms PW, Fecher LA, et al: Host
expression of PD-L1 determines efficacy of PD-L1 pathway
blockade-mediated tumor regression. J Clin Invest. 128:805–815.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Chow MT, Ozga AJ, Servis RL, Frederick DT,
Lo JA, Fisher DE, Freeman GJ, Boland GM and Luster AD: Intratumoral
activity of the CXCR3 chemokine system is required for the efficacy
of anti-PD-1 therapy. Immunity. 50:1498–1512.e5. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Lee AH, Sun L, Mochizuki AY, Reynoso JG,
Orpilla J, Chow F, Kienzler JC, Everson RG, Nathanson DA, Bensinger
SJ, et al: Neoadjuvant PD-1 blockade induces T cell and cDC1
activation but fails to overcome the immunosuppressive tumor
associated macrophages in recurrent glioblastoma. Nat Commun.
12:69382021. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Tsou P, Katayama H, Ostrin EJ and Hanash
SM: The emerging role of B cells in tumor immunity. Cancer Res.
76:5597–5601. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Nelson BH: CD20+ B cells: The other
tumor-infiltrating lymphocytes. J Immunol. 185:4977–4982. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Gunderson AJ, Kaneda MM, Tsujikawa T,
Nguyen AV, Affara NI, Ruffell B, Gorjestani S, Liudahl SM, Truitt
M, Olson P, et al: Bruton tyrosine kinase-dependent immune cell
cross-talk drives pancreas cancer. Cancer Discov. 6:270–285. 2016.
View Article : Google Scholar
|
|
180
|
Fremd C, Schuetz F, Sohn C, Beckhove P and
Domschke C: B cell-regulated immune responses in tumor models and
cancer patients. Oncoimmunology. 2:e254432013. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Wang SS, Liu W, Ly D, Xu H, Qu L and Zhang
L: Tumor-infiltrating B cells: Their role and application in
anti-tumor immunity in lung cancer. Cell Mol Immunol. 16:6–18.
2019. View Article : Google Scholar :
|
|
182
|
Audzevich T, Bashford-Rogers R, Mabbott
NA, Frampton D, Freeman TC, Potocnik A, Kellam P and Gilroy DW:
Pre/pro-B cells generate macrophage populations during homeostasis
and inflammation. Proc Natl Acad Sci USA. 114:E3954–E3963. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Fridman WH, Petitprez F, Meylan M, Chen
TW, Sun CM, Roumenina LT and Sautès-Fridman C: B cells and cancer:
To B or not to B? J Exp Med. 218:e202008512021. View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Gonzalez H, Hagerling C and Werb Z: Roles
of the immune system in cancer: From tumor initiation to metastatic
progression. Genes Dev. 32:1267–1284. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Michel T, Hentges F and Zimmer J:
Consequences of the cross-talk between monocytes/macrophages and
natural killer cells. Front Immunol. 3:4032013. View Article : Google Scholar
|
|
186
|
Sivori S, Pende D, Quatrini L, Pietra G,
Della Chiesa M, Vacca P, Tumino N, Moretta F, Mingari MC, Locatelli
F and Moretta L: NK cells and ILCs in tumor immunotherapy. Mol
Aspects Med. 80:1008702021. View Article : Google Scholar
|
|
187
|
Krneta T, Gillgrass A, Poznanski S, Chew
M, Lee AJ, Kolb M and Ashkar AA: M2-polarized and tumor-associated
macrophages alter NK cell phenotype and function in a
contact-dependent manner. J Leukoc Biol. 101:285–295. 2017.
View Article : Google Scholar
|
|
188
|
Platonova S, Cherfils-Vicini J, Damotte D,
Crozet L, Vieillard V, Validire P, André P, Dieu-Nosjean MC,
Alifano M, Régnard JF, et al: Profound coordinated alterations of
intratumoral NK cell phenotype and function in lung carcinoma.
Cancer Res. 71:5412–5422. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Kloss M, Decker P, Baltz KM, Baessler T,
Jung G, Rammensee HG, Steinle A, Krusch M and Salih HR: Interaction
of monocytes with NK cells upon Toll-like receptor-induced
expression of the NKG2D ligand MICA. J Immunol. 181:6711–6719.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Peng LS, Zhang JY, Teng YS, Zhao YL, Wang
TT, Mao FY, Lv YP, Cheng P, Li WH, Chen N, et al: Tumor-associated
monocytes/macrophages impair NK-cell function via TGFβ1 in human
gastric cancer. Cancer Immunol Res. 5:248–256. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Castriconi R, Cantoni C, Della Chiesa M,
Vitale M, Marcenaro E, Conte R, Biassoni R, Bottino C, Moretta L
and Moretta A: Transforming growth factor beta 1 inhibits
expression of NKp30 and NKG2D receptors: Consequences for the
NK-mediated killing of dendritic cells. Proc Natl Acad Sci USA.
100:4120–4125. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Gallazzi M, Baci D, Mortara L, Bosi A,
Buono G, Naselli A, Guarneri A, Dehò F, Capogrosso P, Albini A, et
al: Prostate cancer peripheral blood NK cells show enhanced CD9,
CD49a, CXCR4, CXCL8, MMP-9 production and secrete
monocyte-recruiting and polarizing factors. Front Immunol.
11:5861262021. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Malekghasemi S, Majidi J, Baghbanzadeh A,
Abdolalizadeh J, Baradaran B and Aghebati-Maleki L:
Tumor-associated macrophages: Protumoral macrophages in
inflammatory tumor microenvironment. Adv Pharm Bull. 10:556–565.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Xu B, Chen L, Li J, Zheng X, Shi L, Wu C
and Jiang J: Prognostic value of tumor infiltrating NK cells and
macrophages in stage II+III esophageal cancer patients. Oncotarget.
7:74904–74916. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Eisinger S, Sarhan D, Boura VF,
Ibarlucea-Benitez I, Tyystjärvi S, Oliynyk G, Arsenian-Henriksson
M, Lane D, Wikström SL, Kiessling R, et al: Targeting a scavenger
receptor on tumor-associated macrophages activates tumor cell
killing by natural killer cells. Proc Natl Acad Sci USA.
117:32005–32016. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
196
|
Bellora F, Castriconi R, Dondero A,
Pessino A, Nencioni A, Liggieri G, Moretta L, Mantovani A, Moretta
A and Bottino C: TLR activation of tumor-associated macrophages
from ovarian cancer patients triggers cytolytic activity of NK
cells. Eur J Immunol. 44:1814–1822. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
197
|
Bellora F, Castriconi R, Dondero A,
Reggiardo G, Moretta L, Mantovani A, Moretta A and Bottino C: The
interaction of human natural killer cells with either unpolarized
or polarized macrophages results in different functional outcomes.
Proc Natl Acad Sci USA. 107:21659–21664. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
198
|
Dalbeth N, Gundle R, Davies RJO, Lee YCG,
McMichael AJ and Callan MFC: CD56bright NK cells are enriched at
inflammatory sites and can engage with monocytes in a reciprocal
program of activation. J Immunol. 173:6418–6426. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
199
|
O'Sullivan T, Saddawi-Konefka R, Vermi W,
Koebel CM, Arthur C, White JM, Uppaluri R, Andrews DM, Ngiow SF,
Teng MW, et al: Cancer immunoediting by the innate immune system in
the absence of adaptive immunity. J Exp Med. 209:1869–1882. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
200
|
Ponzetta A, Carriero R, Carnevale S,
Barbagallo M, Molgora M, Perucchini C, Magrini E, Gianni F,
Kunderfranco P, Polentarutti N, et al: Neutrophils driving
unconventional T cells mediate resistance against murine sarcomas
and selected human tumors. Cell. 178:346–360.e24. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
201
|
Wu L and Zhang XH: Tumor-associated
neutrophils and macrophages-heterogenous but not chaotic. Front
Immunol. 11:5539672020. View Article : Google Scholar : PubMed/NCBI
|
|
202
|
Kim J and Bae JS: Tumor-associated
macrophages and neutrophils in tumor microenvironment. Mediators
Inflamm. 2016:60581472016. View Article : Google Scholar : PubMed/NCBI
|
|
203
|
Zhou SL, Zhou ZJ, Hu ZQ, Huang XW, Wang Z,
Chen EB, Fan J, Cao Y, Dai Z and Zhou J: Tumor-associated
neutrophils recruit macrophages and T-regulatory cells to promote
progression of hepatocellular carcinoma and resistance to
sorafenib. Gastroenterology. 150:1646–1658.e17. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
204
|
Zilionis R, Engblom C, Pfirschke C, Savova
V, Zemmour D, Saatcioglu HD, Krishnan I, Maroni G, Meyerovitz CV,
Kerwin CM, et al: Single-cell transcriptomics of human and mouse
lung cancers reveals conserved myeloid populations across
individuals and species. Immunity. 50:1317–1334.e10. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
205
|
Zhou Z, Wang P, Sun R, Li J, Hu Z, Xin H,
Luo C, Zhou J, Fan J and Zhou S: Tumor-associated neutrophils and
macrophages interaction contributes to intrahepatic
cholangiocarcinoma progression by activating STAT3. J Immunother
Cancer. 9:e0019462021. View Article : Google Scholar : PubMed/NCBI
|
|
206
|
Gunaydin G: CAFs interacting with TAMs in
tumor microenvironment to enhance tumorigenesis and immune evasion.
Front Oncol. 11:6683492021. View Article : Google Scholar : PubMed/NCBI
|
|
207
|
Sweeny L, Liu Z, Lancaster W, Hart J,
Hartman YE and Rosenthal EL: Inhibition of fibroblasts reduced head
and neck cancer growth by targeting fibroblast growth factor
receptor. Laryngoscope. 122:1539–1544. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
208
|
Vu HN, Miller WJ, O'Connor SA, He M,
Schafer PH, Payvandi F, Muller GW, Stirling DI and Libutti SK:
CC-5079: A small molecule with MKP1, antiangiogenic, and antitumor
activity. J Surg Res. 164:116–125. 2010. View Article : Google Scholar
|
|
209
|
Donthireddy L, Vonteddu P, Murthy T, Kwak
T, Eraslan RN, Podojil JR, Elhofy A, Boyne MT II, Puisis JJ, Veglia
F, et al: ONP-302 nanoparticles inhibit tumor growth by altering
tumor-associated macrophages and cancer-associated fibroblasts. J
Cancer. 13:1933–1944. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
210
|
Kashyap AS, Schmittnaegel M, Rigamonti N,
Pais-Ferreira D, Mueller P, Buchi M, Ooi CH, Kreuzaler M,
Hirschmann P, Guichard A, et al: Optimized antiangiogenic
reprogramming of the tumor microenvironment potentiates CD40
immunotherapy. Proc Natl Acad Sci USA. 117:541–551. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
211
|
Zhang L, Qi Y, Min H, Ni C, Wang F, Wang
B, Qin H, Zhang Y, Liu G, Qin Y, et al: Cooperatively responsive
peptide nanotherapeutic that regulates angiopoietin receptor Tie2
activity in tumor microenvironment to prevent breast tumor relapse
after chemotherapy. ACS Nano. 13:5091–5102. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
212
|
Alam A, Blanc I, Gueguen-Dorbes G, Duclos
O, Bonnin J, Barron P, Laplace MC, Morin G, Gaujarengues F, Dol F,
et al: SAR131675, a potent and selective VEGFR-3-TK inhibitor with
antilymphangiogenic, antitumoral, and antimetastatic activities.
Mol Cancer Ther. 11:1637–1649. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
213
|
Gyori D, Lim EL, Grant FM, Spensberger D,
Roychoudhuri R, Shuttleworth SJ, Okkenhaug K, Stephens LR and
Hawkins PT: Compensation between CSF1R+ macrophages and Foxp3+ Treg
cells drives resistance to tumor immunotherapy. JCI Insight.
3:e1206312018. View Article : Google Scholar : PubMed/NCBI
|
|
214
|
Oweida AJ, Darragh L, Phan A, Binder D,
Bhatia S, Mueller A, Court BV, Milner D, Raben D, Woessner R, et
al: STAT3 modulation of regulatory T cells in response to radiation
therapy in head and neck cancer. J Natl Cancer Inst. 111:1339–1349.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
215
|
Zhu Y, Knolhoff BL, Meyer MA, Nywening TM,
West BL, Luo J, Wang-Gillam A, Goedegebuure SP, Linehan DC and
DeNardo DG: CSF1/CSF1R blockade reprograms tumor-infiltrating
macrophages and improves response to T-cell checkpoint
immunotherapy in pancreatic cancer models. Cancer Res.
74:5057–5069. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
216
|
Sluijter M, van der Sluis TC, van der
Velden PA, Versluis M, West BL, van der Burg SH and van Hall T:
Inhibition of CSF-1R supports T-cell mediated melanoma therapy.
PLoS One. 9:e1042302014. View Article : Google Scholar : PubMed/NCBI
|
|
217
|
Doleschel D, Hoff S, Koletnik S, Rix A,
Zopf D, Kiessling F and Lederle W: Regorafenib enhances anti-PD1
immunotherapy efficacy in murine colorectal cancers and their
combination prevents tumor regrowth. J Exp Clin Cancer Res.
40:2882021. View Article : Google Scholar : PubMed/NCBI
|
|
218
|
Zhang JQ, Zeng S, Vitiello GA, Seifert AM,
Medina BD, Beckman MJ, Loo JK, Santamaria-Barria J, Maltbaek JH,
Param NJ, et al: Macrophages and CD8+ T cells mediate
the antitumor efficacy of combined CD40 ligation and imatinib
therapy in gastrointestinal stromal tumors. Cancer Immunol Res.
6:434–447. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
219
|
Zhang M, Wen B, Anton OM, Yao Z, Dubois S,
Ju W, Sato N, DiLillo DJ, Bamford RN, Ravetch JV and Waldmann TA:
IL-15 enhanced antibody-dependent cellular cytotoxicity mediated by
NK cells and macrophages. Proc Natl Acad Sci USA.
115:E10915–E10924. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
220
|
Jarosz-Biej M, Kamińska N, Matuszczak S,
Cichoń T, Pamuła-Piłat J, Czapla J, Smolarczyk R, Skwarzyńska D,
Kulik K and Szala S: M1-like macrophages change tumor blood vessels
and microenvironment in murine melanoma. PLoS One. 13:e01910122018.
View Article : Google Scholar : PubMed/NCBI
|
|
221
|
Gaggero S, Witt K, Carlsten M and Mitra S:
Cytokines orchestrating the natural killer-myeloid cell crosstalk
in the tumor microenvironment: implications for natural killer
cell-based cancer immunotherapy. Front Immunol. 11:6212252021.
View Article : Google Scholar : PubMed/NCBI
|
|
222
|
Zhou Z, Zhang C, Zhang J and Tian Z:
Macrophages help NK cells to attack tumor cells by stimulatory
NKG2D ligand but protect themselves from NK killing by inhibitory
ligand Qa-1. PLoS One. 7:e369282012. View Article : Google Scholar : PubMed/NCBI
|
|
223
|
Wang C, Cui A, Bukenya M, Aung A, Pradhan
D, Whittaker CA, Agarwal Y, Thomas A, Liang S, Amlashi P, et al:
Reprogramming NK cells and macrophages via combined antibody and
cytokine therapy primes tumors for elimination by checkpoint
blockade. Cell Rep. 37:1100212021. View Article : Google Scholar : PubMed/NCBI
|
|
224
|
Nywening TM, Belt BA, Cullinan DR, Panni
RZ, Han BJ, Sanford DE, Jacobs RC, Ye J, Patel AA, Gillanders WE,
et al: Targeting both tumour-associated CXCR2+
neutrophils and CCR2+ macrophages disrupts myeloid
recruitment and improves chemotherapeutic responses in pancreatic
ductal adenocarcinoma. Gut. 67:1112–1123. 2018. View Article : Google Scholar
|
|
225
|
Li X, Yao W, Yuan Y, Chen P, Li B, Li J,
Chu R, Song H, Xie D, Jiang X and Wang H: Targeting of
tumour-infiltrating macrophages via CCL2/CCR2 signalling as a
therapeutic strategy against hepatocellular carcinoma. Gut.
66:157–167. 2017. View Article : Google Scholar
|
|
226
|
Veillette A and Tang Z: Signaling
regulatory protein (SIRP) α-CD47 blockade joins the ranks of immune
checkpoint inhibition. J Clin Oncol. 37:1012–1014. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
227
|
Wesolowski R, Sharma N, Reebel L, Rodal
MB, Peck A, West BL, Marimuthu A, Severson P, Karlin DA, Dowlati A,
et al: Phase Ib study of the combination of pexidartinib (PLX3397),
a CSF-1R inhibitor, and paclitaxel in patients with advanced solid
tumors. Ther Adv Med Oncol. 11:17588359198542382019. View Article : Google Scholar : PubMed/NCBI
|
|
228
|
Wu F, Yang J, Liu J, Wang Y, Mu J, Zeng Q,
Deng S and Zhou H: Signaling pathways in cancer-associated
fibroblasts and targeted therapy for cancer. Signal Transduct
Target Ther. 6:2182021. View Article : Google Scholar : PubMed/NCBI
|