Introduction
Dipeptidyl peptidase 4 (DPP4; OMIM: 102720), also
named CD26, intestinal adenosine deaminase complexing protein 2
(ADCP2), is a type II transmembrane glycoprotein (1-3). One
of the lymphocyte membrane-bound proteins, DPP4 possesses serine
protease activity. DPP4 is located on human chromosome 2q24.2 and
encodes a protein composed of 766 amino acids with a predicted
molecular weight of 88,279 Da (4).
DPP4 is ubiquitously expressed in different cells, organs and body
fluids. It participates in diverse physiological and pathological
processes by mediating glucose metabolism (5,6), the
endocrine and cardiovascular systems, cell adhesion, apoptosis,
fibrosis, inflammation and immune function (1,2,7).
These effects are achieved by forming homodimers for the enzymatic
activity and/or protein-protein interactions. For example, DPP4 may
interact with the S1 domain of the S-protein (spike glycoprotein)
in severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) to
cause coronavirus disease 19 (COVID-19) disease (8). Its binding to caveolin-1 and
card-maguk protein 1 induces T-cell proliferation and NF-κB
activation (9). Its interaction
with adenosine deaminase (ADA) mediates epithelial and lymphocyte
cell adhesion (10). Thus, DPP4
plays an important role in the development of immune-mediated
disorders (7) and as a receptor of
ADA in lymphocytes. Furthermore, it has been shown that DPP4
promotes metastases of types of cancer (11).
In 2003, Conarello et al (6) concluded that DPP4 inhibition is a
viable therapeutic option for the treatment of metabolic diseases
related to diabetes and obesity. DPP4 inhibitors, including
hypoglycemic agents (e.g., sitagliptin, alogliptin and
linagliptin), have been approved by the U.S. Food and Drug
Administration (FDA) for the treatment of type 2 diabetes mellitus
(12,13). In addition, clinical trials and
preclinical studies have evaluated the efficacy and safety of DPP4
inhibitors in immune-mediated disorders and anti-cancer immune
responses (14). It is thought
that these inhibitors may act by regulating the balance of the T
helper 1/2 (Th1/Th2) phenotype and production of cytokines
(15,16). Over-expression of DPP4 in lung
diseases may increase the susceptibility to viral invasion. Further
evidence showed the role of DPP4 in the pathogenesis of lung
disorders, including lung cancer; thus, DPP4 may be a therapeutic
target in this setting (12).
DPP4 affects viral attacks in infectious respiratory
disorders, including Middle East respiratory syndrome (MERS)
(17,18) and non-infectious lung disorders,
including lung cancer (19),
chronic obstructive pulmonary disorder, pulmonary fibrosis and
asthma (20,21). It has been shown that ADA
competitively binds to DPP4, thereby preventing the binding of DPP4
with the MERS-CoV S1 domain. Thus, it is a potential agent for
blocking viral attacks (22).
During the COVID-19 pandemic, it was also shown that DPP4 is a
vital marker/receptor that might play a significant role in disease
progression (23,24) and susceptibility to COVID-19
(25). Decreased circulating DPP4
activity or expression is prognostic for severe outcomes of
COVID-19 (23,26). High DPP4 levels and other receptors
(such as CD147 and TMPRSS2) are correlated with the occurrence and
severity of COVID-19 (27). Acute
respiratory distress syndrome is a leading cause of mortality in
patients with COVID-19. DPP4 inhibitors may be a novel therapeutic
approach by decreasing the production of inflammatory factors, such
as IL-6, IL-1β and TNF-α (28).
Studies have documented a potential therapeutic effect of DPP4
inhibitors in patients with diabetes diagnosed with COVID-19 by
reducing cytokine production (29-31).
DPP4 inhibitors may reduce the mortality rate and improve outcomes
of COVID-19 patients with or without type1 and type2 diabetes
mellitus (32-37). Clinical trials of DPP4 inhibitors
for COVID-19 are currently being performed (38). Several inhibitors of DPP4, such as
alogliptin, linagliptin, sitagliptin, saxagliptin and vildagliptin,
have been approved worldwide, including by the European Medicines
Agency and the U.S. FDA (39). A
new monoclonal antibody interacting with angiotensin converting
enzyme 2 (ACE2) and DPP4 was recently developed; however, its use
in patients has yet to be assessed (40).
Evidence indicates the effects of COVID-19 on the
clinical outcome for patients with cancer; patients with cancer and
plus COVID-19 are at a high risk of mortality (41-44).
Therefore, patients with malignant cancer should be cautious during
the COVID-19 pandemic. The correlation between DPP4 and malignancy
is currently debatable and its role as a tumor suppressor or
promoter of carcinogenesis needs to be determined. Moreover, the
mechanisms underlying these processes are complex and warrant
further clarification (45).
The present study conducted comprehensive and
integrative analyses of DPP4 expression in healthy individuals and
patients in the pan-cancer setting using genomic, transcriptomic
and epigenomic data. The relationship between the expression of
DPP4 and immune cell infiltration was analyzed. The results of
analyses may reveal the susceptibility of different types of cancer
to SARS-CoV-2 attack and the importance of AD/C-X-C motif chemokine
ligand 3 (AD/CXCL3) signaling. By using N6, N6-dimethyladenosine
(m62A), cordycepin (CD) and adenosine (AD)
small molecules derived from natural products, the present study
may also identify potential therapeutic agents for SARS-CoV-2.
Materials and methods
Online databases
In GenBank, DPP4 in humans is coded as follows:
NP_001926.2 for protein and NM_001935.4 for the gene (https://www.ncbi.nlm.nih.gov/protein/NP_001926.2;
https://www.ncbi.nlm.nih.gov/nuccore/NM_001935.4?report=genbank&log$=seqview)
(46,47). Analysis of DPP4 expression in
healthy and cancer tissues was performed using data from the Human
Protein Atlas (HPA) (https://www.proteinatlas.org/ENSG00000197635-DPP4/tissue)
(48,49). The DPP4 expression in
different cancer tissues and matched healthy tissues, survival,
isoforms, distribution and structure of the domain were analyzed
through Gene Expression Profiling Interactive Analysis 2 (GEPIA2;
http://gepia2.cancer-pku.cn/#analysis) (50,51).
DNA methylation analysis of the DPP4 promoter in the pan-cancer
setting was performed using the DNA methylation interactive
visualization database (DNMIVD) (http://119.3.41.228/dnmivd/query_gene/?cancer=pancancer&gene=DPP4)
(52). Mutation analysis for the
DPP4 gene was performed through tumor genomics in cBioPortal
(https://www.cbioportal.org/results/cancerTypesSummary?case_set_id=all&gene_list=DPP4&cancer_study_list=5c8a7d55e4b046111fee2296)
(53). An integrated repository
portal, called tumor-immune system interactions database (TISIDB),
was utilized to analyze the relationship between the abundance of
tumor-infiltrating lymphocytes (TILs) and DPP4 expression
(http://cis.hku.hk/TISIDB/browse.php?gene=DPP4)
(54).
Reagents, cell lines and breast cancer
tissue collections
CD (cat no. B20196) was purchased from Must
Bio-Technology Co. Ltd. AD (cat no. A6218) and
m62A (cat no. N879945) were purchased from
Macklin Biochemical Technology Co., Ltd. Dimethyl sulfoxide (DMSO;
cat. no. D8148) was purchased from MilliporeSigma. The total RNA
extraction kit (cat no. DP419), buffer RZ solution (cat no. RK145)
and 2XTaq PCR Master Mix (cat no. KT211) were purchased from
TransGen Biotech Co Ltd. The reverse transcription (RT) PCR kit
(cat no. FSQ-201) was purchased from Toyobo Biotech Co., Ltd.
Sodium dodecyl sulfate (SDS; cat. no. L5750) and β-actin antibodies
(cat. no. A1978) were purchased from MilliporeSigma. The DPP4
antibody (cat. no. 67138S), mouse (cat. no. 7076S) and rabbit (cat.
no. 7074S) secondary antibodies were purchased from Cell Signaling
Technology, Inc. The lymphocyte activating 3 (LAG-3) polyclonal
antibody (cat no. 16616-1-AP) was purchased from Wuhan Sanying
Biotechnology. RPMI 1640 medium (cat no. C3010-0500) and Dulbecco's
modified Eagle's medium (DMEM; cat no. C3113-0500) were purchased
from Shanghai VivaCell Biosciences, Ltd. Fetal bovine serum (FBS;
cat. no. A6907) was purchased from Invigentech. Phosphate buffered
saline (PBS; cat no. P1010) was purchased from Beijing Solarbio
Science & Technology Co., Ltd. The
trypsin-ethylenediaminetetraacetic acid (Trypsin-EDTA) solution
(cat no. C0201) and penicillin/streptomycin solution (cat no.
C0222) were purchased from Beyotime Institute of Biotechnology. The
lung cancer cell lines H1975 and A549, breast cancer cell line
BT549 and prostate cancer cell lines PC3 and 22RV1 were purchased
from the American Type Culture Collection.
A total of 11 paired of breast cancer tissues and
the matched normal tissues from Chinese women were collected with
informed consent (the affiliated hospital of Southwest Medical
University; the date range was between January 2021 and September
2022). Total proteins were extracted for western blotting. All were
invasive breast cancers with patients' age range 40-64. The present
study was approved by the Ethical Committee of Southwest Medical
University (approval no. 20221117-049).
Animals
All animal experiments were performed in strict
accordance with international, national and institutional animal
care guidelines. The present study was reviewed and approved by the
Ethics Committee of Southwest Medical University, Sichuan, China
(approval no. 20221117-049). A total of six BALB/c female mice (20
weeks old, ~24 g) of specific-pathogen-free were purchased from
Chongqing Tengxin Biotechnology Co., Ltd. The mice were kept under
constant temperature at 22°C, 50-60% humidity and a 12 h light/dark
cycle (lights on from 7 a.m. to 7 p.m.) according to the feeding
standard. All mice were healthy and pathogen-free, with free access
to diet and water. After AD injection, the mice were observed every
12 h and showed no abnormalities. After 24 h, sodium pentobarbital
(200 mg/kg body weight) was injected intraperitoneally to sacrifice
the mice. Mortality was defined as no heartbeat following injection
and dilated pupils. The whole mouse experiment was conducted for 14
days.
Isolation of mouse lymphocytes
The six BALB/c female mice were divided into the
experimental and control groups (three mice per group). AD (0.006
mg/µl) solution (containing 20% DMSO, 30% polyethylene
glycol 400, 5% Tween 80 and 63% NaCl) was injected into each mouse
in the experimental group through the caudal vein for 24 h. Each
mouse in the experimental group received an injection with 25 mg/kg
AD. The mice were sacrificed and the spleens were isolated, ground
and filtered using 100 µm cell strainers (cat no. 15-1100;
Biologix Group Ltd.) in an ice bath. Each sample was collected in a
15 ml tube and centrifuged at 4°C, 400-600 g, 5 min. The
supernatant was collected, sterile 1Xred blood cell lysis solution
(150 mmol/l NH4Cl, 10 mmol/l KHCO3 and 0.1
mmol/l EDTA) (55) was added and
the mixture was thoroughly mixed and placed on ice for 5-8 min.
Following lysis, the reaction was terminated with sterile cold
1XPBS. The samples were centrifuged at 4°C, 400-600 g, 15 min, the
debris was discarded and the supernatant was mixed with cold 1XPBS.
Each sample was divided into two Eppendorf tubes for protein and
RNA extraction.
Immunohistochemistry assay (IHC)
IHC was performed using formalin-fixed,
paraffin-embedded sections of breast and lung cancer tissues
obtained from Chinese patients was performed as previously
described (47,56-60).
DPP4 antibody (1:100 dilution) was used in IHC. Histopathological
images were captured using a fluorescence microscope (cat no.
DM2500; Leica Microsystems GmbH).
Cell culture
RPMI 1640 medium containing 10% FBS and 1% double
antibiotics (penicillin-streptomycin) was used for H1975, A549, PC3
and 22RV1 cells; DMEM containing 10% FBS was used for BT549 and
293T-hACE2 cells (a gift from Professor Xianghui Fu of Sichuan
University) (61). When the cell
density reached ~50-70% or when the cells entered the logarithmic
growth phase 24 h later, CD, AD, or m62A were
added to the experimental group. Specifically, H1975 and BT549
cells were treated with CD, AD, or m62A at
different concentrations (0, 10, 20 and 40 µM) for 24 h in a
12-well plate (57). All cells
were cultured in an incubator at 37°C and 5% CO2.
Cell transfection and syncytia
formation
Syncytia formation is a hallmark cellular event in
SARS-CoV-2 infection (62).
293T-hACE2 cells were used to transfect the plasmid
pCDH-CMV-HnCoV-S-EF1-copGFP. A six-well plate was used as an
example, and the cell density was 40-50% for transfection.
Lipofectamine® 3000 (cat no. L3000001; Invitrogen;
Thermo Fisher Scientific Co., Ltd.) and P3000 were added to each
100 ng of plasmid at 0.3 µl. Then, two EP tubes were
prepared in each well, labeled tube 1 and tube 2. Opti-MEM (cat no.
31985070; Thermo Fisher Scientific Co., Ltd.), P3000 and plasmid
were added to tube 1, and Opti-MEM and Lipofectamine®
3000 (cat no. L3000001; Invitrogen; Thermo Fisher Scientific Co.,
Ltd.) were added to tube 2. The two tubes were mixed and left at
room temperature for 5 min. After rinsing twice with 1 ml 1XPBS, 1
ml Opti-MEM was added to each well. The mixture of tube 1 and tube
2 was added to the corresponding wells and the mixture was shaken
and mixed to make the mixture evenly cover the cells. The cells
were placed into a constant temperature incubator at 37°C and 5%
CO2 saturated humidity for another 6 h. After 6 h, the
mixture in the 6-well plate was removed and 2 ml of fresh complete
medium was added. Following continued culture for 18 h, the control
group was without CD treatment, the experimental group was treated
with 20 µM CD for 24 h. The syncytium formation of each
group was observed and analyzed using a ZOE Fluorescent Cell Imager
(Bio-Rad Laboratories, Inc.). The DPP4 protein and mRNA levels were
also measured.
Western blotting
After treatment with CD, AD, or
m62A, the cells were rinsed twice with
ice-cold 1XPBS and lysed using ice-cold 1XEBC buffer (containing 20
mM Tris-HCl pH8.0, 125 mM NaCl, 2 mM EDTA, 0.5% NP-40 and protease
inhibitors). Protein concentration was determined using Coomassie
brilliant blue staining, diluted to the same concentration. An
equal volume of loading buffer (2XSDS) was added to each sample for
protein extraction. The samples with the mass of protein ~50
µg per lane were subjected to 10% SDS-polyacrylamide gel
electrophoresis. Subsequently, proteins were transferred onto
polyvinylidene difluoride membranes. The membranes were blocked
with 1XPBST (1XPBS plus 0.05% Tween20) containing 5% skimmed milk
for 1-2 h at room temperature, followed by addition of 2% skimmed
milk solution. Next, the membranes were incubated with the primary
anti-DPP4 antibody (1:2,000 dilution) overnight at 4°C and washed
thrice with 1XPBST. Thereafter, the membranes were incubated with
secondary antibody (1:2,000 dilution) at room temperature for 2-4 h
on a decolorizing shaker and washed thrice with 1XPBST. Finally,
the membranes were soaked in fresh SuperSignal West Pico
Chemiluminescence Substrate (cat no. 34580; Thermo Fisher
Scientific Inc.) and visualized using a gel imaging system (Bio-Rad
Laboratories, Inc.). β-actin served as an internal control. The
protocols for western blotting analysis of breast cancer tissues
and matched healthy tissues obtained from Chinese patients has been
previously described (56,57).
Cycloheximide (CHX) chase assays
Chase assay for DPP4 was performed with 20
µg/ml cycloheximide (CHX; cat no. HY-12320; Med Chem Express
Technology Co., Ltd.) and AD using H1975 cells, as previously
described (57). All experiments
were performed in triplicate.
RNA extraction and RT
RNA extraction and cDNA synthesis were performed
according to the manufacture's protocols. Cell treated with CD, AD,
or m62A were rinsed twice with ice-cold 1XPBS
buffer. Next, buffer RZ solution was added at a concentration of 10
cm2/ml for 5 min. Subsequently, the cells were lysed
using an RNA extraction kit (cat no. DP419; Tiangen Biotech Co.,
Ltd.). The concentration and purity of the extracted total RNA were
determined using a Nanodrop spectrophotometer (Thermo Fisher
Scientific Inc.) at an optical density 260/280 nm of=1.8-2.0. RNA
integrity evaluated through 1% agarose gel electrophoresis.
Thereafter, the extracted RNA was reversely transcribed into cDNA
with reverse transcriptase. Total RNA (1 µg) and 5XRT Master
Mix (2 µl) were place in a tube and ddH2O was
added to a total volume of 10 µl. The RT reaction was
performed as follows: 37°C for 15 min; 50°C for 5 min; 98°C for 5
min; and maintenance at 4°C until use.
Semi-quantitative PCR
Each PCR tube (total volume: 10 µl) was
placed in an ice bath and contained the following: 2XTaq PCR Master
MIX (5 µl); primers (1 µl), cDNA (1 µl); and
ddH2O (3 µl). Following thorough mixing and
centrifugation, PCR was performed as follows: pre-denaturation at
95°C for 90 sec; denaturation at 95°C for 30 sec, annealing at 60°C
for 30 sec, extension at 72°C for 30 sec; 32 cycles for DPP4
and 23 cycles for β actin (ACTB), final extension at 72°C
for 5 min and maintenance at 16°C until use (63). The DPP4 primers used in
semi-quantitative RT-PCR were RT-DPP4-L: 5′-caaattgaagcagccagaca-3′
and RT-DPP4-R: 5′-cagggctttg-gagatctgag-3′. The size of the PCR
product for DPP4 was 354bp. The Lag-3 primers in mouse for
semi-quantitative RT-PCR included RT-mLag-3-L:
5′-gccatctcgttctcgttctc-3′ and RT-mLag-3-R:
5′-tttttgatgctgctgacagg-3′. PCR product size was 335 bp for Lag-3.
Of note, Actb served as an internal control. The Actb primers in
mouse for semi-quantitative RT-PCR included RT-ACTB-mL:
5′-TGT TAC CAA CTG GGA CGA CA-3′ and RT-ACTB-mR: 5′-TCT CAG
CTG TGG TGG TGA AG-3′. PCR product size was 392 bp for Actb. The
ACTB primers in human semi-quantitative RT-PCR included
RT-ACTB-5: 5′-CTC TTC CAG CCT TCC TTC CT-3′ and
RT-ACTB-3: 5′-CAC CTT CAC CGT TCC AGT TT-3′. The expected
product size was 510 bp. By using the nucleic acid dye GoldView
(cat no. G8142; Beijing Solarbio Science & Technology Co.,
Ltd.), PCR products were verified with 1.5% agarose gel
electrophoresis. PCR assays were performed in triplicate.
Statistical analysis
Clinical data from The Cancer Genome Atlas (TCGA)
database (64) for cancers were
used for overall survival (OS) analysis. The two group of patients
according to the upper quartile expression of DPP4 were compared
using Kaplan-Meier survival curves (https://kmplot.com/analysis/index.php?p=service).
Experimental statistical analysis was performed for six values
using independent samples t-test (two groups) for mouse experiments
and calculated P-values, expressed as mean ± standard deviation.
The mean grayscale values and fluorescence area of individual
fluorescence images were measured using ImageJ software (National
Institutes of Health). Spearman and Pearson were used to analyze
the correlations between gene expression levels and promoter
methylation levels in different tissues. All statistical analyses
were performed using SPSS version 13.0 (SPSS, Inc.) and GraphPad
Prism8 (Dotmatics) software. P<0.05 was considered to indicate a
statistically significant difference.
Results
DPP4 expression in human healthy
tissues
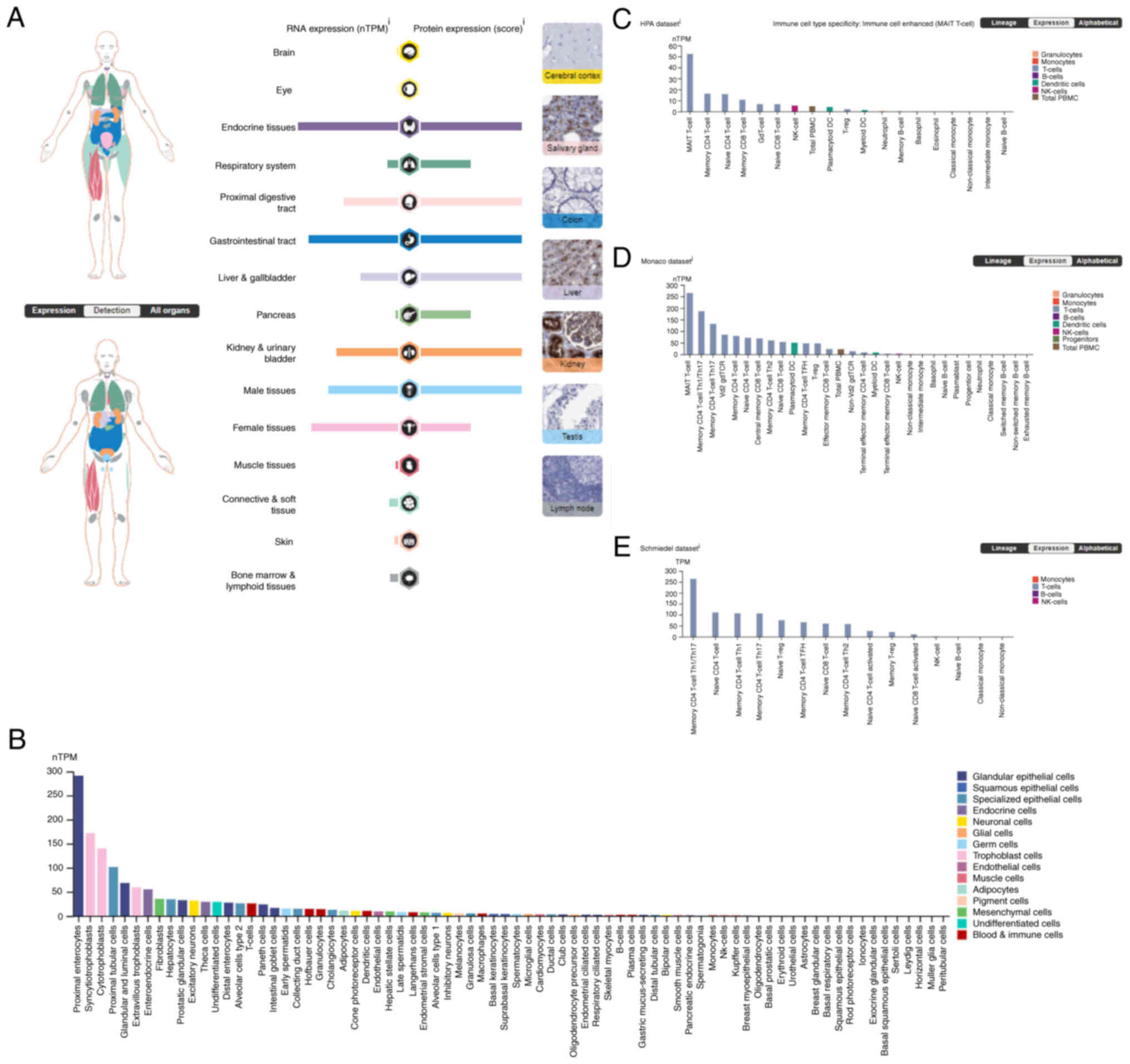
DPP4 mRNA expression is expressed in
endocrine tissues, the gastrointestinal tract, female tissue
(mainly in placenta), male tissue (mainly in prostate and seminal
vesicle), proximal digestive tract, kidneys, bladder, liver,
gallbladder and the respiratory system (Fig. 1A). DPP4 protein is expressed
endocrine tissues, the gastrointestinal tract, female tissue
(mainly in placenta), male tissue (mainly in prostate and seminal
vesicle), proximal digestive tract, kidneys, bladder, liver,
gallbladder, the respiratory system and the pancreas (Fig. 1A). Although the DPP4 mRNA
levels were low [lung, 16.0 normalized transcripts per kilobase per
million mapped reads (nTPM)], moderate expression of DPP4 protein
was observed (Fig. 1A),
demonstrating the role of DPP4 in viral invasion in the
lungs/bronchus/nasopharynx. DPP4 mRNA levels were high in
various cell types, including proximal enterocytes (glandular
epithelial cells, 291.1 nTPM), syncytiotrophoblasts (trophoblast
cells, 172.0 nTPM), cytotrophoblasts (trophoblast cells, 140.3
nTPM) and proximal tubular cells (squamous epithelial cells, 101.4
nTPM) (Fig. 1B).
The mRNA expression of DPP4 in immune cells
was analyzed and three datasets are presented (Fig. 1C-E). In the HPA dataset, which
includes 18 immune cell types and total peripheral blood
mononuclear cells (PBMC), the results showed that DPP4 mRNA
is mainly expressed in T cells [mucosal associated invariant T
(MAIT) cells, 53.3 nPTM), natural killer cells (5.3 nPTM) and total
PBMC (4.7 nPTM; Fig. 1C)]. In the
Monaco dataset, which includes 29 immune cell types and total PBMC,
the results indicated that DPP4 mRNA is mainly expressed in
T cells (MAIT, 264.4 nPTM, memory CD4 T-cell Th1/Th17, 186.0 nPTM),
dendritic cells (50.3 nPTM) and total PBMC (22.3 nPTM) (Fig. 1D). In the Schmiedel dataset, which
includes 15 immune cell types, the results indicated that
DPP4 mRNA is mainly expressed in T cells (memory CD4 T-cell
Th1/Th17, 263.5 nPTM) (Fig. 1E).
Collectively, these data demonstrated that DPP4 is highly expressed
in T cells.
DPP4 expression in human tumor tissues
from TCGA
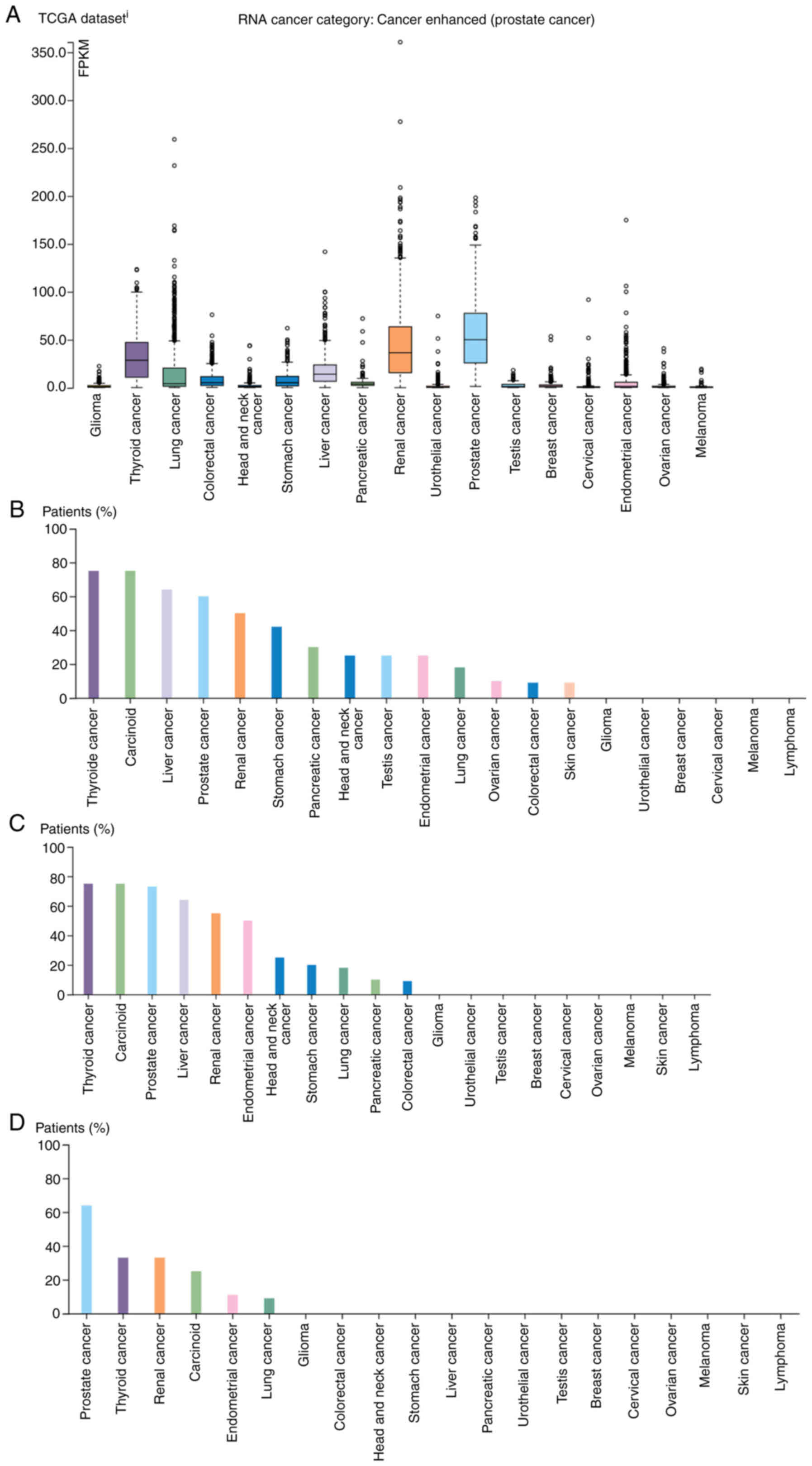
The DPP4 RNA expression was high in tumor
tissues, with the highest levels recorded for prostate cancer [50.1
fragments per kilobase per million mapped fragments (FPKM)], renal
cancers (36.5 FPKM), including kidney chromophobe (KICH), kidney
renal clear cell carcinoma (KIRC) and kidney renal papillary cell
carcinoma (KIRP) and thyroid cancer (28.6 FPKM; Fig. 2A).
For protein expression, moderate-to-strong
cytoplasmic or membranous positivity for the DPP4 antibody
HPA068778 was mainly observed in renal, liver, stomach and prostate
tissues; the highest levels were noted in thyroid, carcinoid,
liver, prostate, renal and other types of cancer (Fig. 2B). A moderate-to-strong cytoplasmic
and membranous positivity for the DPP4 antibody HPA071236 was
observed in carcinoid, endometrial, prostate, thyroid, renal and
liver tissues; the remaining cancers were essentially negative. The
highest levels were observed in thyroid, carcinoid, prostate,
liver, renal, endometrial and other types of cancer (Fig. 2C). As shown by the analysis using
the DPP4 antibody CAB045970, several cases of prostate cancer and a
few cases of renal cell carcinoma presented a moderate-to-strong
membranous and/or cytoplasmic positivity, whereas the remaining
types of cancer were negative. The highest in prostate cancer,
thyroid, renal, carcinoid, endometrial and lung cancer (Fig. 2D). Thus, DPP4 protein is mainly
expressed in thyroid, carcinoid, prostate, renal and liver cancer.
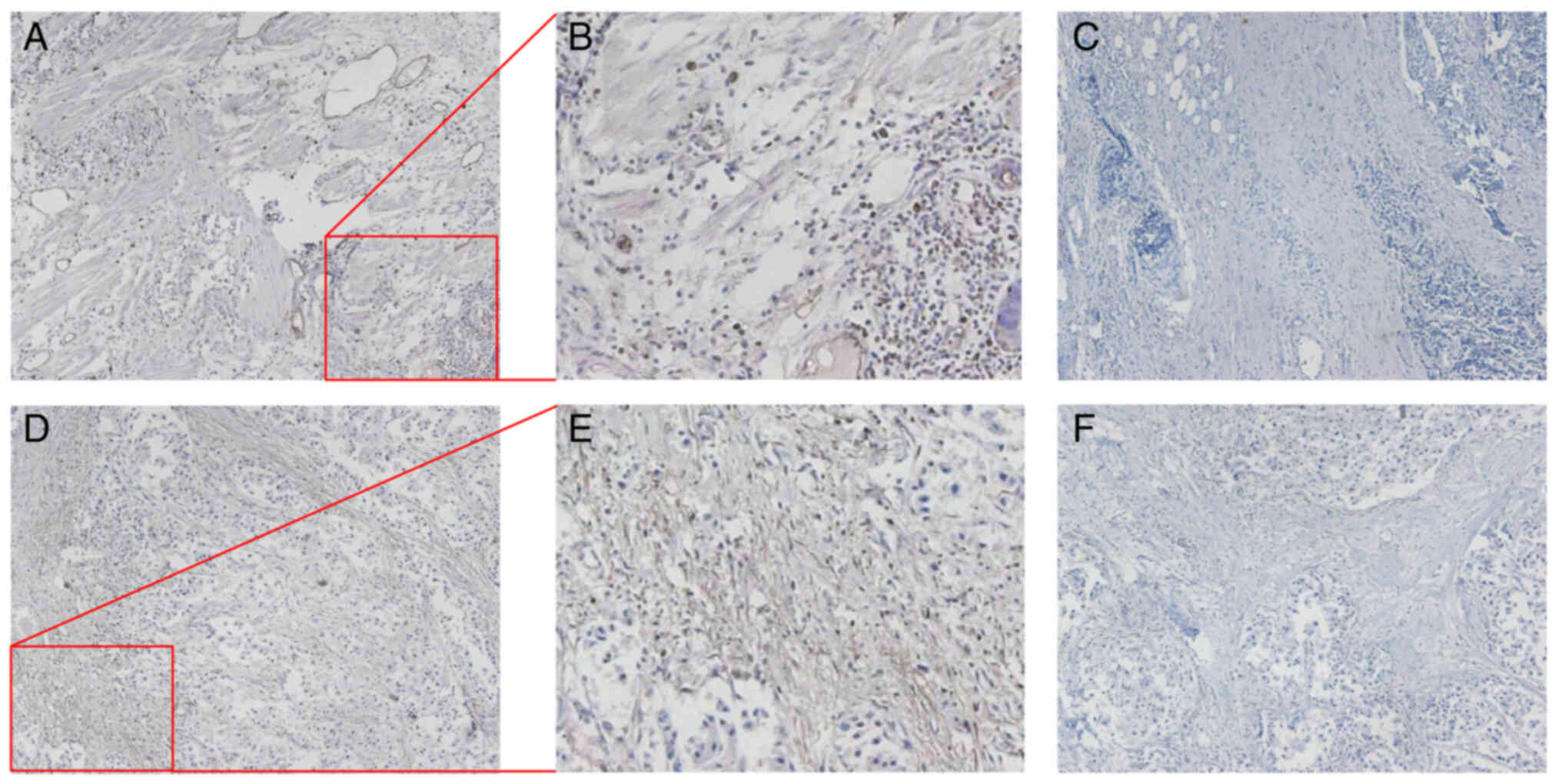
The present study also conducted IHC using tissues obtained from
patients with lung small cell carcinoma and breast cancer. The
localization of DPP4 protein in lung small cell carcinoma and
breast cancer is shown in Fig. 3.
A moderate-to-strong cytoplasmic and membranous positivity was
revealed, suggesting the involvement of DPP4 in viral invasion.
DPP4 expression and prognostic value in
various tumor tissues and matched healthy tissues
Patients with cancer are vulnerable to SARS-CoV-2
infection and the combination of these conditions is associated
with a high mortality rate (41-44).
Therefore, it is important to identify differences in the
expression levels of DPP4 between tumor tissues of various types of
cancer and matched healthy tissues. The results of the GEPIA2
analysis revealed that the levels of DPP4 are upregulated in
esophageal carcinoma (ESCA), KIRP, liver hepatocellular carcinoma
(LIHC), lung adenocarcinoma (LUAD), pancreatic adenocarcinoma
(PAAD), prostate adenocarcinoma (PRAD), stomach adenocarcinoma
(STAD), thyroid carcinoma (THCA) and thymoma (THYM) compared with
matched healthy tissues. However, they are downregulated in breast
invasive carcinoma (BRCA), KICH, lung squamous cell carcinoma
(LUSC) and skin cutaneous melanoma (SKCM; Fig. 4A-C). To verify these data, breast
cancer samples with predicted downregulation of DPP4 were collected
and western blotting was performed (Fig. 4D). The findings revealed that DPP4
protein levels were decreased in nine of 11 tumor tissues (81.8%)
compared with the corresponding healthy tissues. This observation
confirmed the results of the mRNA analysis from TCGA database for
patients with BRCA.
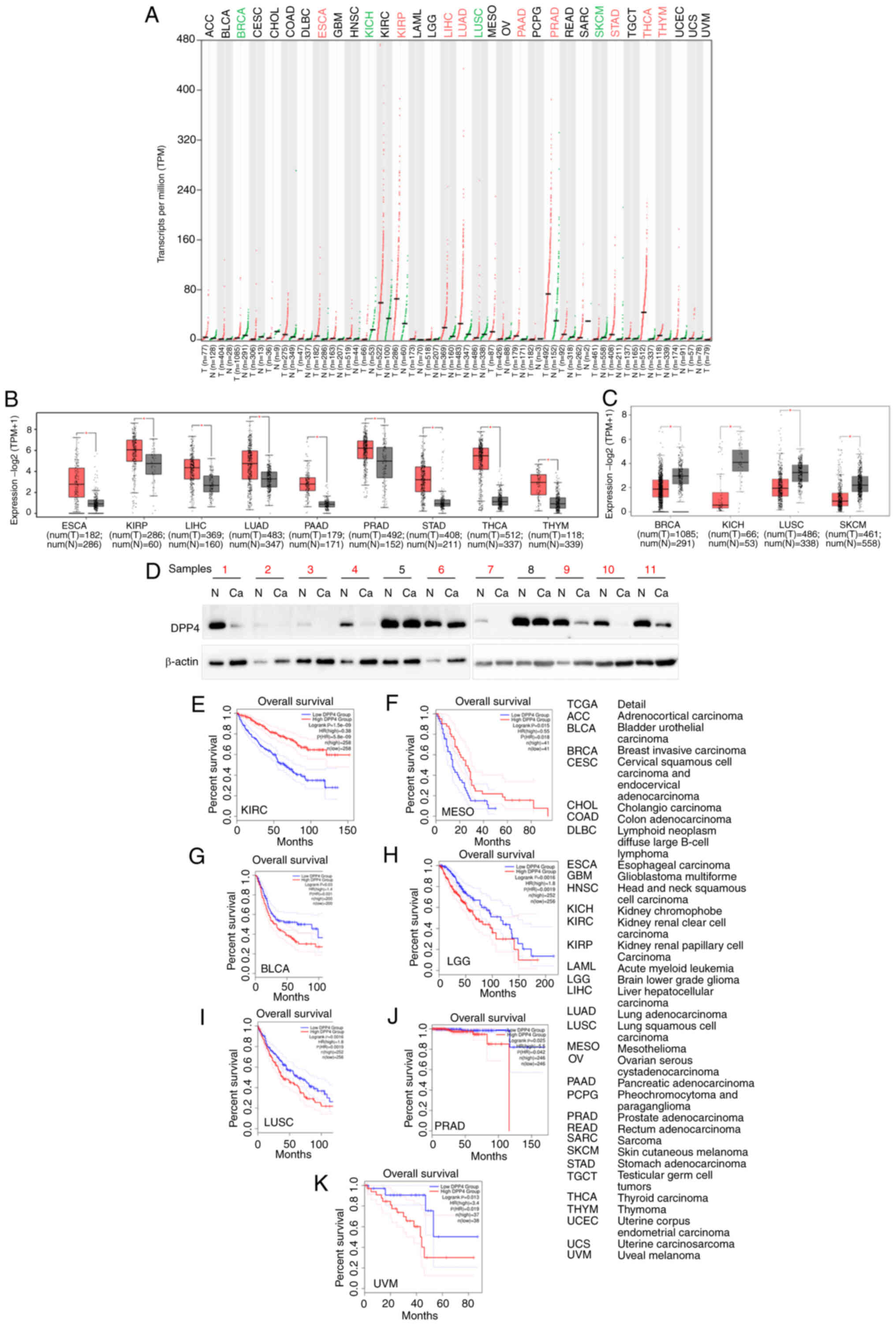 | Figure 4Comparison of DPP4 expression between
human tumor tissues and corresponding healthy tissues. (A)
comparison of the DPP4 expression profiles in different types of
human tumor tissues and corresponding healthy tissues. (B) Box
plots showing that DPP4 expression is increased in different types
of human tumor tissues compared with matched healthy tissues. (C)
Box plots showing that DPP4 expression is decreased in different
types of human tumor tissues compared with matched healthy tissues.
(D) Validation of the upregulation of DPP4 in breast cancer tissues
through western blotting. Overall survival analysis based on the
DPP4 expression and Kaplan-Meier curves for (E) KIRC, (F) MESO, (G)
BLCA, (H) LGG, (I) LUSC, (J) PRAD and (K) UVM, respectively. The
right panel provides the full description of all types of cancer.
The log rank P-value ≤0.5 was set as a difference and log rank
P-value ≤0.01 was set as a significant difference. DPP4, dipeptidyl
peptidase 4; KIRC, kidney renal clear cell carcinoma; MESO,
mesothelioma; BLCA, bladder urothelial carcinoma; LGG, brain lower
grade glioma; LUSC, lung squamous cell carcinoma; PRAD, prostate
adenocarcinoma; UVM, uveal melanoma; HR, hazard ratio. |
Further investigation of the prognostic value of
DPP4 demonstrated that higher expression levels were
associated with a long OS in KIRC and mesothelioma (MESO) (Fig. 4E-F), implying that DPP4 could be a
favorable marker. However, high expression was also linked to a
short OS in bladder urothelial carcinoma (BLCA), lower grade glioma
(LGG), LUSC, PRAD and UVM (Fig.
4G-K), suggesting that DPP4 could be an unfavorable marker.
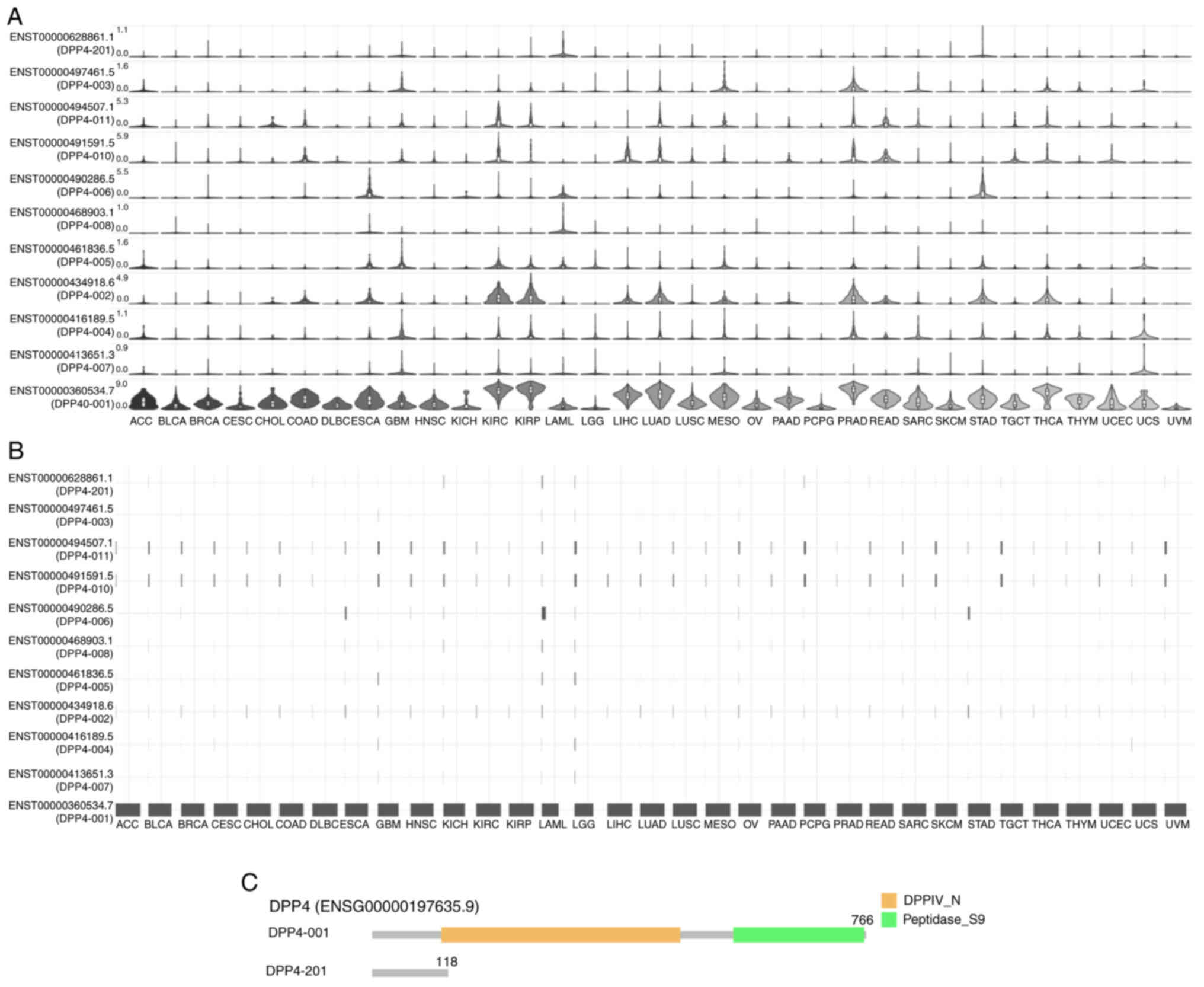
DPP4 isoform expression, distribution and
structure in different types of cancer
Various isoforms may play different roles in host
susceptibility to SARS-CoV-2 invasion (65,66).
Thus, analysis of the prevalence and structures of DPP4 isoforms in
the pan-cancer setting was performed. According to the results, 11
isoforms exhibited different DPP4 expression levels (Fig. 5A). The expression of isoform
ENST00000360534.7(DPP4-001) was high in all 33 types of cancer;
nevertheless, the expression of other isoforms was very low or no
detectable (Fig. 5A). Identical
results were obtained regarding the utilization of isoforms
(Fig. 5B). The genomic structures
of DPP4 isoforms in 33 types of cancer are shown in Fig. 5C. The isoform DPP4-001 has 766
amino acids and includes the DPPIV_N domain and Peptidase_S9
domain. However, the DPP4-201 isoform has 118 amino acids and lacks
functional domains. Information on the other nine isoforms is
missing. These results indicated that ENST00000360534.7(DPP4-001)
may be the functional isoform for tumorigenesis and SARS-CoV-2
entry in patients in the pan-cancer setting.
DNA methylation of the DPP4 promoter
region may regulate DPP4 expression in some types of cancer
DNA methylation could be a mechanism underlying the
regulation of DPP4 gene expression. Thus, the present study
investigated the types of cancer in which DPP4 is regulated through
DNA methylation. For the purpose, it analyzed data from the DNMIVD
database to determine the degree of methylation of the DPP4
promoter and changes in expression in 23 different types of cancer.
It was found that the mRNA levels of DPP4 were decreased in
two types of cancer (BRCA and colon adenocarcinoma (COAD)) compared
with matched healthy tissues; however DPP4 methylation levels were
increased (Fig. 6A vs. B, E vs. F,
respectively). The mRNA levels of DPP4 were increased in
THCA samples and the promoter regions of DPP4 were decreased in
cancer tissues compared with matched healthy tissues (Fig. 6I vs. J). Pearson and Spearman
correlation analyses showed the inverse correlations between
promoter methylation and DPP4 expression in BRCA, COAD and
THCA compared with matched healthy tissues (Fig. 6C vs. D, G vs. H, K vs. L,
respectively). However, P>0.05 in BRCA and COAD patients.
Nevertheless, these results indicated that DNA methylation may be
the mechanism regulating DPP4 expression in BRCA, COAD and THCA.
Other mechanisms may be involved in the regulation of DPP4
expression in other types of cancer.
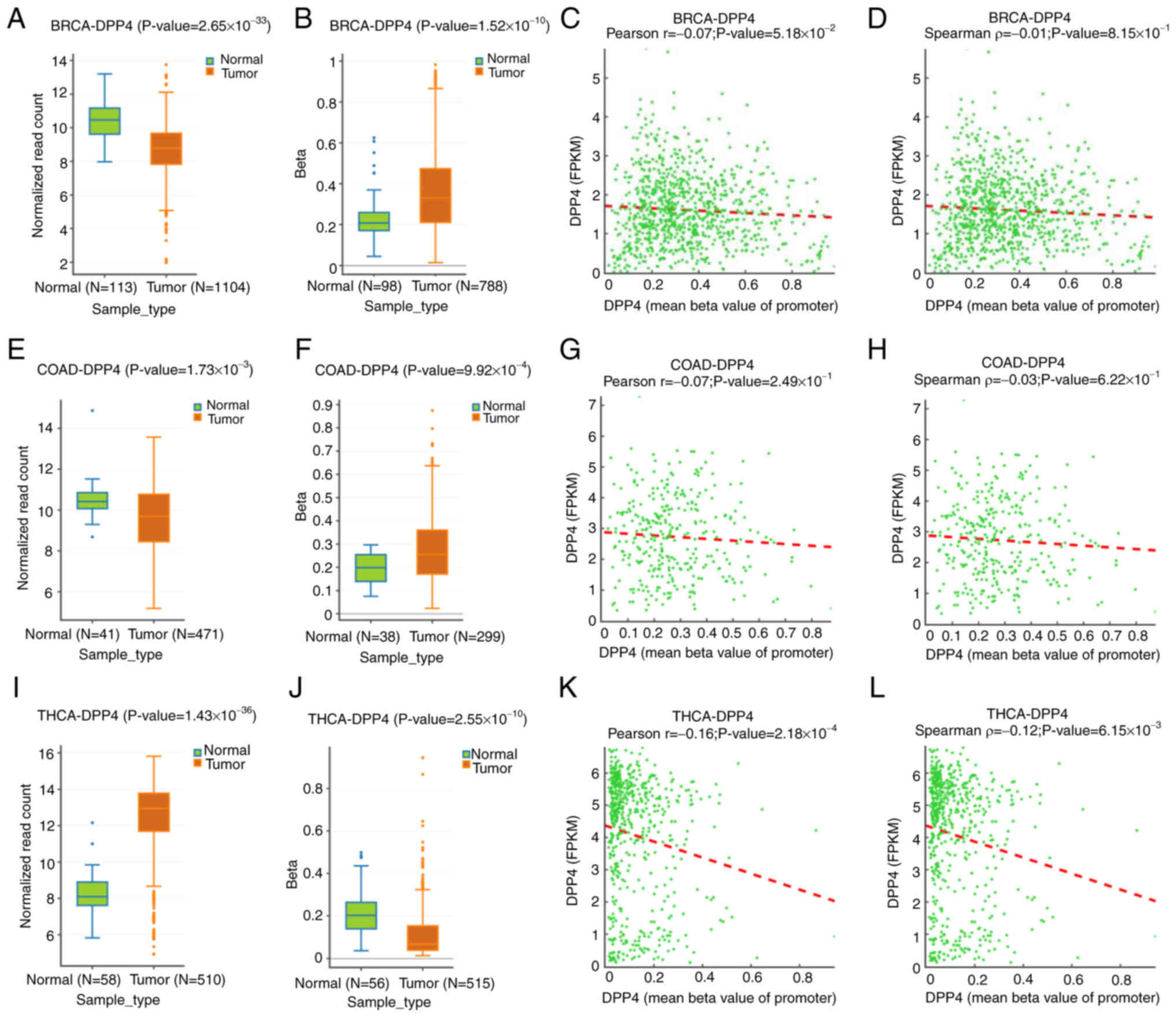 | Figure 6Methylation status of the DPP4
promoter in tumor tissues and corresponding healthy tissues. (A-D)
Expression levels, methylation status at the promoter region,
Pearson correlation and Spearman correlation for DPP4 in BRCA
tissues and corresponding healthy tissues, respectively. (E-H)
Expression levels, methylation status at the promoter region,
Pearson correlation and Spearman correlation for DPP4 in COAD
tissues and matched healthy tissues, respectively. (I-L) Expression
levels, methylation status at the promoter region, Pearson
correlation and Spearman correlation for DPP4 in THCA tissues and
corresponding healthy tissues, respectively. DPP4, dipeptidyl
peptidase 4; BRCA, breast invasive carcinoma; COAD, colon
adenocarcinoma; THCA, thyroid carcinoma. |
Altered DNA profiles of DPP4 in different
types of cancer
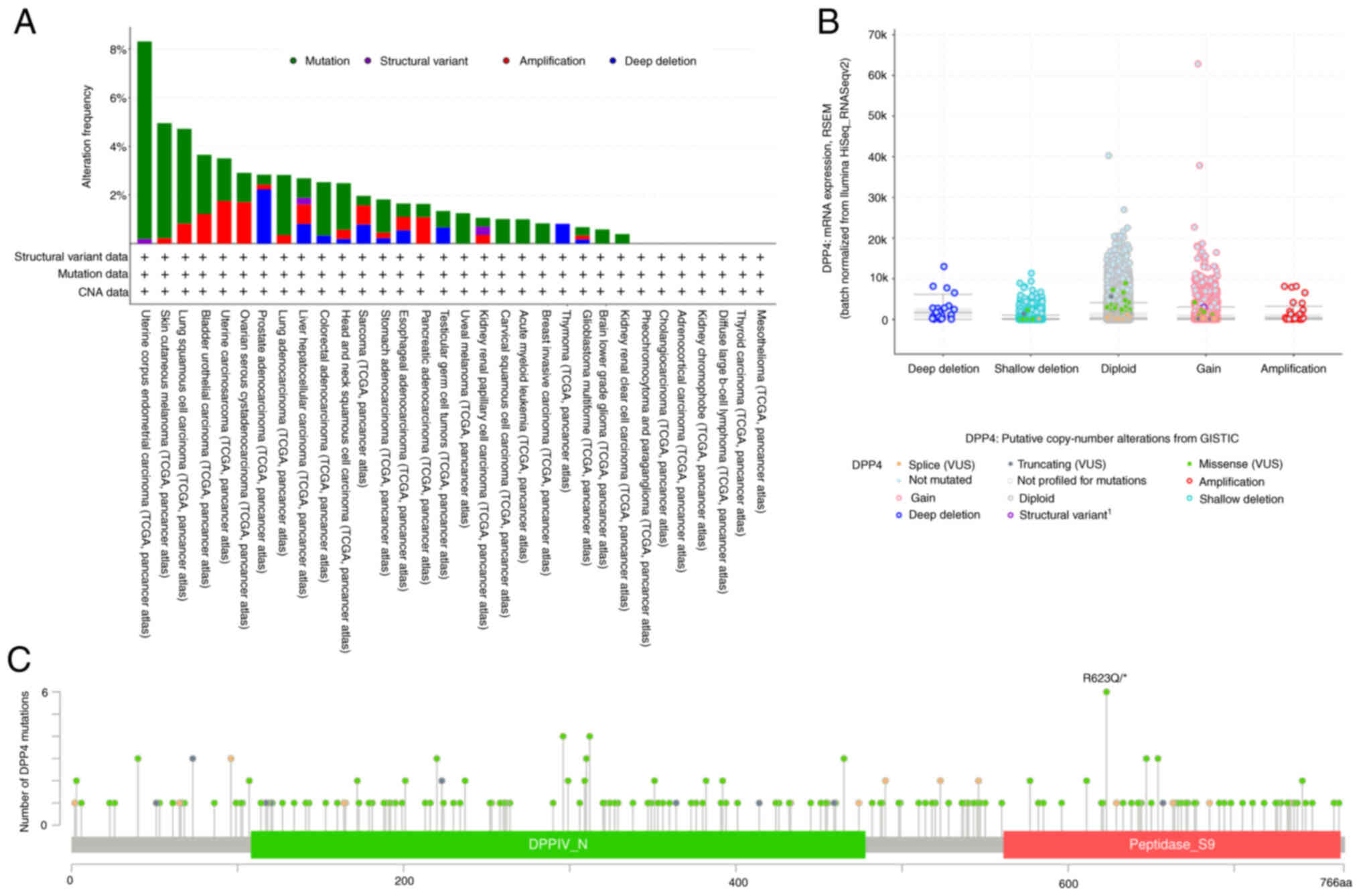
It was recently reported that DPP4 gene
polymorphisms are associated with clinicopathological
characteristics in oral cancer (67). The DPP4 rs3788979 polymorphism may
be associated with severe COVID-19 (68). The present study analyzed the
DPP4 mutation profile in 32 different types of cancer to
determine relationships with the development of malignancy,
recurrence and therapeutic resistance. By analyzing DPP4
mutations, it was found that uterine corpus endometrial carcinoma
(UCEC) has the highest frequency of mutations (8.32% of 529 cases),
followed by SKCM (4.95% of 444 cases); KIRC has the lowest
frequency of mutations (0.39% of 511 cases; Fig. 7A). Fig. 7B shows structural variations,
mutations, amplifications and deep deletions with mutations being
the dominant type of alteration (Fig.
7A and B). There were no DPP4 mutations found in the
other seven types of cancer, which are presented in Fig. 7A. The detailed landscape included
missense mutation, splicing, truncation and structural
variation/fusion along the whole DPP4 gene, with missense
mutation being the dominant type of alteration (Fig. 7C).
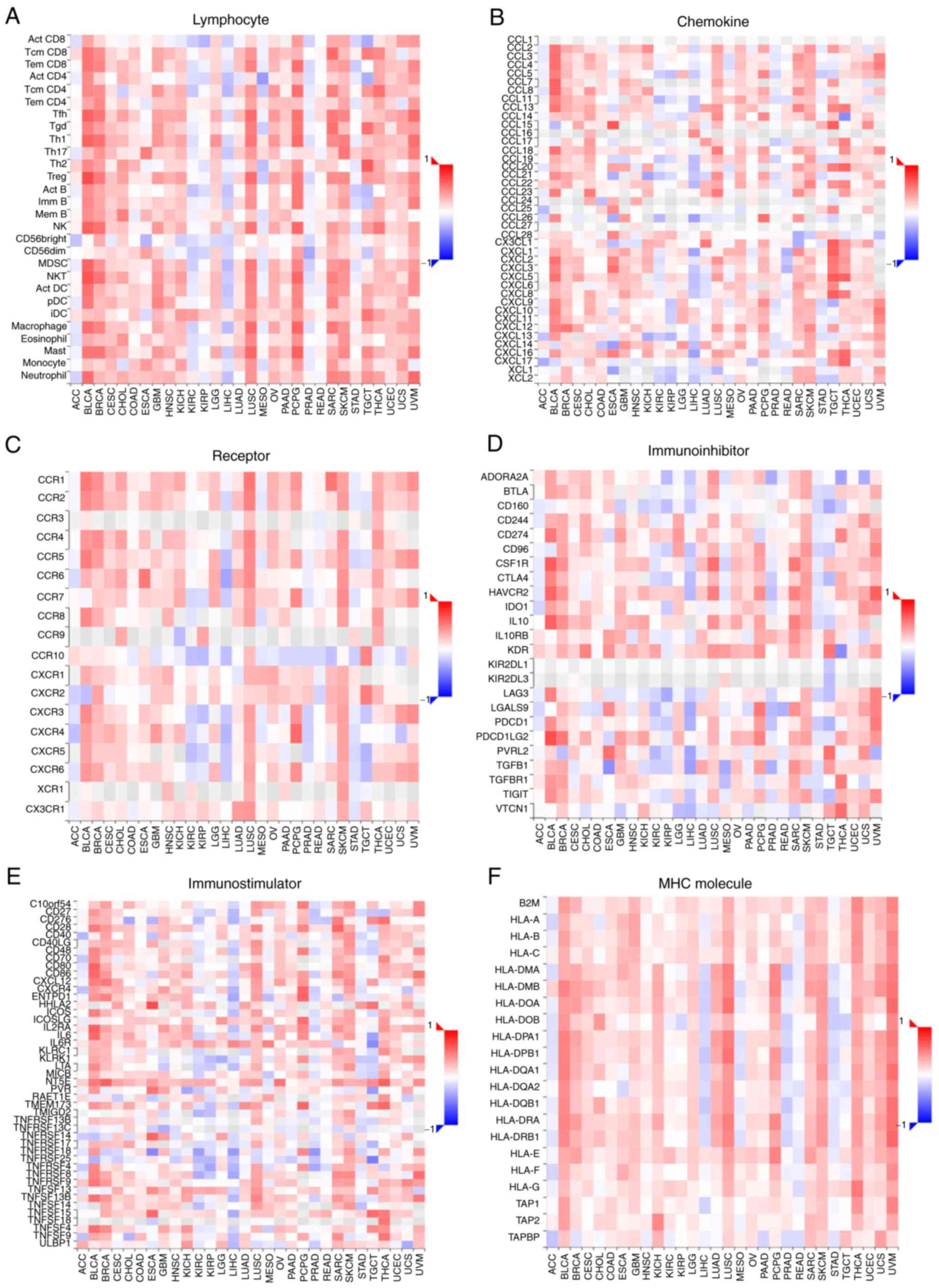
Association studies for DPP4 expression
with tumor-immune systems in different types of cancer
The activity of DPP4 regulates numerous chemokines,
cytokines and immunosuppressant/immunostimulants. DPP4 is also
involved in cancer immunology (14). Immune system response plays
indispensable roles in anti-viral and anti-cancer processes.
Therefore, the present study conducted association studies to
examine the relationship between DPP4 mRNA expression and
the extent of immune infiltration in different types of cancer.
Notably, it found correlations between DPP4 mRNA expression
and immune lymphocytes (Fig. 8A),
chemokines (Fig. 8B), receptors
(Fig. 8C), immunosuppressants
(Fig. 8D), immunostimulants
(Fig. 8E) and major
histocompatibility complex (MHC) molecules (Fig. 8F) in almost all types of cancer.
These findings suggest therapeutic and preventive roles of DPP4 in
cancer and SARS-CoV-2 invasion, respectively.
CD inhibits DPP4 expression in H1975,
A549 and BT549 cancer cells
Small molecules or active substances derived from
natural products can manipulate gene expression. The present study
sought to determine whether these substances would target DPP4
expression. The effect of CD, a nucleoside derivative (adenosine
derivative), on the expression of DPP4 was tested in a
triple-negative breast cancer cell line (BT549) and two lung cancer
cell lines (A549 and H1975). The results demonstrated that CD
inhibited both DPP4 protein and mRNA expression in a
dosage-dependent manner in H1975 (Fig.
9A and D), A549 (Fig. 9B and
E) and BT549 (Fig. 9C and F)
cells.
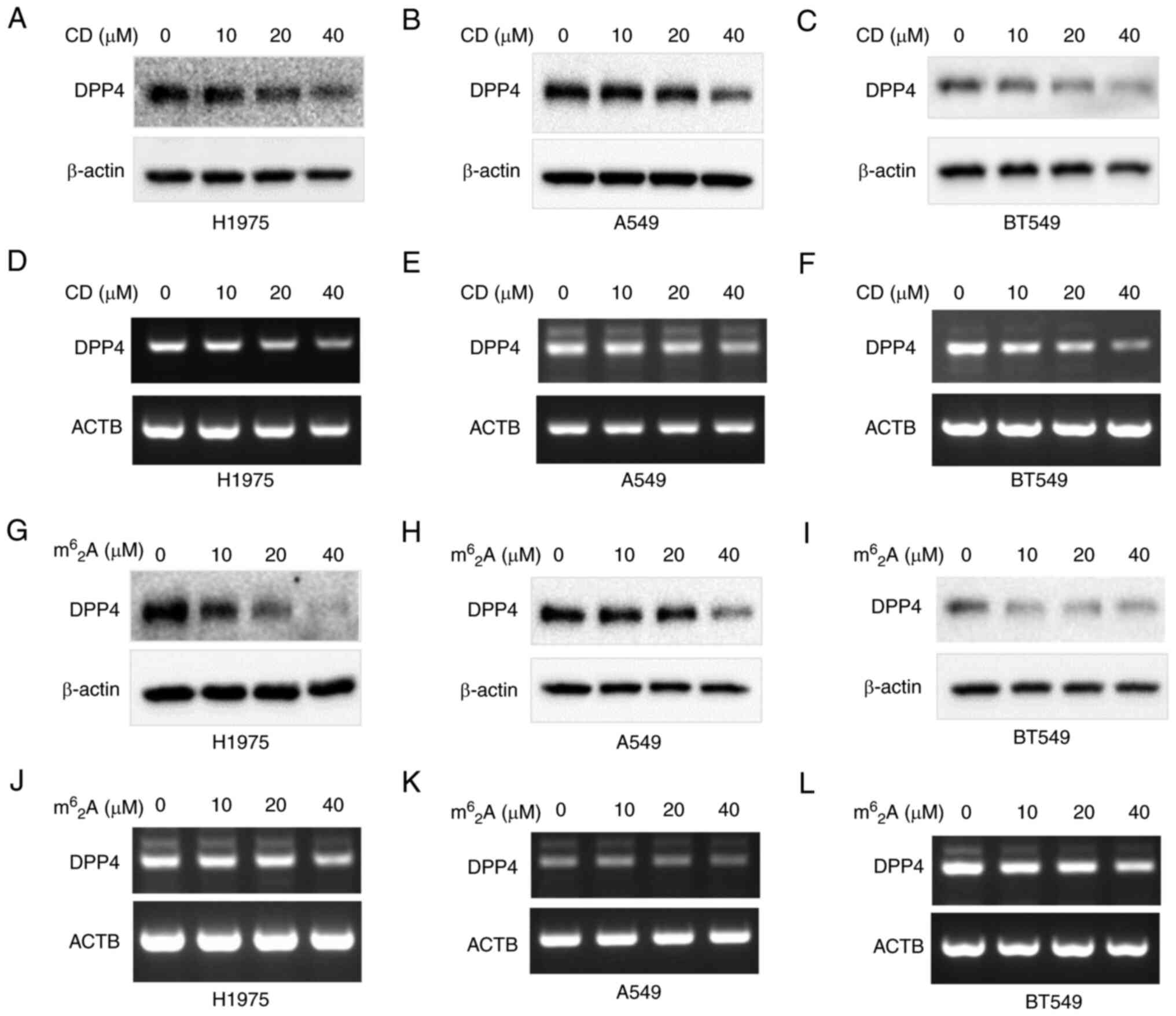 | Figure 9CD and m62A
inhibit DPP4 expressions of both protein and mRNA in various cancer
cells. CD decreases DPP4 expression in (A and D) H1975 lung cancer
cells, (B and E) A549 lung cancer cells, (C and F) BT549 breast
cancer cells. m62A decreases DPP4 expression
in (G and J) H1975 lung cancer cells, (H and K) A549 lung cancer
cells and (I and L) BT549 breast cancer cells. Panels A, B, C, G, H
and I are protein expressions while panels D, E, F, J, K and L are
mRNA expressions. CD, cordycepin; DPP4, dipeptidyl peptidase 4. |
m62A inhibits DPP4
expression in H1975, A549 and BT549 cancer cells
The effect of m62A, another
adenosine derivative, on DPP4 expression was also tested in BT549,
A549 and H1975 cells. The results indicated that
m62A also inhibited both DPP4 protein and
mRNA expression in a dosage-dependent manner in H1975 (Fig. 9G and J), A549 (Fig. 9H and K) and BT549 (Fig. 9I and L) cells.
The findings of the present study suggested that
both the adenosine derivatives CD and m62A
may have potential therapeutic value as anti-SARS-CoV-2 molecules
through inhibition of DPP4 expression in cancer.
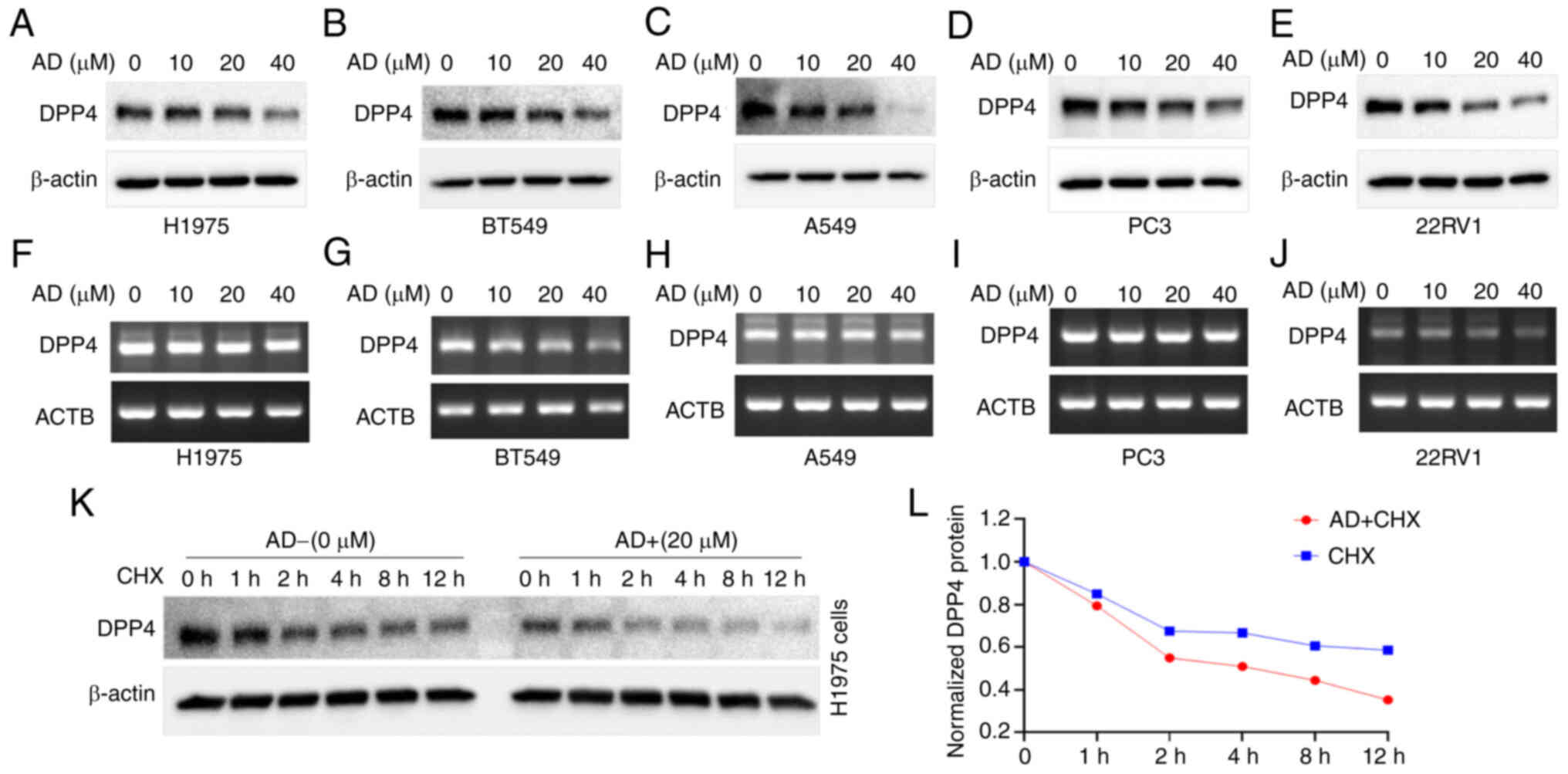
AD inhibits DPP4 expression in H1975,
A549, BT549 and other cancer cells
The effect of AD on DPP4 expression was also
investigated in H1975, A549, BT549 and other cancer cells. The
results indicated that AD also inhibited DPP4 protein expression in
a dosage-dependent manner in H1975 (Fig. 10A), BT549 (Fig. 10B) and A549 (Fig. 10C) cells. Moreover, we found that
AD inhibited DPP4 protein expression in a dosage-dependent manner
in the PC3 (Fig. 10D) and 22RV1
(Fig. 10E) prostate cancer cell
line. Surprisingly, unlike CD and m62A, AD
did not induce significant changes in DPP4 mRNA expression
in these cell lines, except 22RV1 (Fig. 10F-J, respectively). Nevertheless,
these results suggest that AD itself might exert therapeutic
effects as an anti-SARS-CoV-2 agent through inhibition of DPP4
expression in cancer.
Based on the lack of change in the mRNA levels, the
present study sought to investigate whether protein stability
affects DPP4 protein expression. Thus, western blotting analysis
was performed after CHX treatment with or without AD in H1975
cells. The results are shown in Fig.
10K, while the quantitated protein levels are shown in Fig. 10L. Treatment with AD decreased the
protein stability of DPP4 compared with control; the half-life of
the protein was decreased from >12 h to ~4 h (Fig. 10K and L). These results indicate
that treatment with AD could decrease the stability of DPP4
protein.
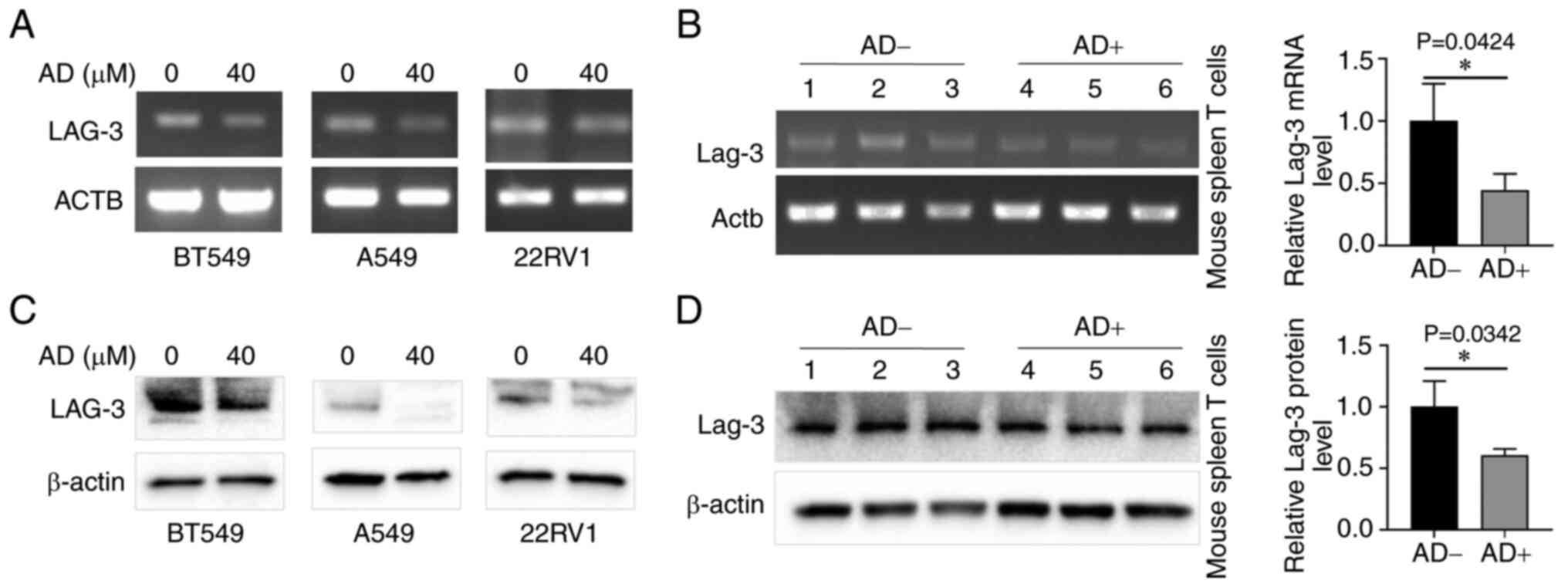
Role of AD in immune molecules and DPP4
expression associated genes
The TISIDB database analysis revealed that DPP4
regulates chemokines, cytokines and
immunosuppressants/immunostimulants; the involvement of immune
molecules from AD/A2AR signaling has been reported in the
literature (69-72). The present study investigated which
immune molecules are affected by AD and are associated with DPP4
expression. Cancer cell lines were treated with or without AD and
semi-quantitative RT-PCR was conducted to examine the expression of
14 candidate genes (Table I). The
results are shown in Fig. 11; AD
downregulated the expression of LAG-3 in BT549, A549 and 22RV1
cells at both the mRNA (Fig. 11A)
and protein (Fig. 11C) levels;
however, it did not alter the levels of the other 13 genes (data
not shown).
 | Table ISelected genes and their primer
sequences for PCR. |
Table I
Selected genes and their primer
sequences for PCR.
| Gene | Primers | Sequence (from
5′-3′) | GenBank |
|---|
| IL6 | RT-IL6-L |
AGACAGCCACTCACCTCTTCA | NM_000600.5 |
| RT-IL6-R |
TAAAGCTGCGCAGAATGAGAT | NM_000600.5 |
| CD28 | RT-CD28-L |
TGTGAAAGGGAAACACCTTTG | NM_006139.4 |
| RT-CD28-R |
TGAGATGTGCAGGTGAGTGAG | NM_006139.4 |
| CD80 | RT-CD80-L |
CACCCTCCAATCTCTGTGTGT | NM_005191.4 |
| RT-CD80-R |
TCCCCAGACATCATAGTCAGC | NM_005191.4 |
| CD86 | RT-CD86-L |
GGGTGAAAGCTTTGCTTCTCT | NM_175862.4 |
| RT-CD86-R |
GTCCAACTGTCCGAATCAAAA | NM_175862.4 |
| IL10 | RT-IL10-L |
GAGTCCTTGCTGGAGGACTTT | NM_000572.3 |
| RT-IL10-R |
GATGCCTTTCTCTTGGAGCTT | NM_000572.3 |
| CTLA-4 | RT-CTLA-4-L |
CAACCTACATGATGGGGAATG | NM_005214.5 |
| RT-CTLA-4-R |
TGCTTTTCACATTCTGGCTCT | NM_005214.5 |
| LAG-3 | RT-LAG-3-L |
CAGAGATGGCTTCAACGTCTC | NM_002286.6 |
| RT-LAG-3-R |
CTGGCTCACATCCTCTAGTCG | NM_002286.6 |
| CXCL1 | RT-CXCL1-L |
CCCAAGAACATCCAAAGTGTG | NM_001511.4 |
| RT-CXCL1-R |
CCTCTGCAGCTGTGTCTCTCT | NM_001511.4 |
| CXCL2 | RT-CXCL2-L |
GGAATTCACCTCAAGAACATCC | NM_002089.4 |
| RT-CXCL2-R |
CCTCTGCAGCTGTGTCTCTCT | NM_002089.4 |
| CXCL3 | RT-CXCL3-L |
TGGGAAGAAAGCTTGTCTCAA | NM_002090.3 |
| RT-CXCL3-R |
GTTCCCCACCCTGTCATTTAT | NM_002090.3 |
| CXCL5 | RT-CXCL5-L |
AATCTTCGCTCCTCCAATCTC | NM_002994.5 |
| RT-CXCL5-R |
CAAATTTCCTTCCCGTTCTTC | NM_002994.5 |
| CXCL6 | RT-CXCL6-L |
ACCCCAAAACGATTGGTAAAC | NM_002993.4 |
| RT-CXCL6-R |
TCTTACTGGGTCCAGGGATCT | NM_002993.4 |
| CXCL8 | RT-CXCL8-L |
TTTGCCAAGGAGTGCTAAAGA | NM_000584.4 |
| RT-CXCL8-R |
TATTGCATCTGGCAACCCTAC | NM_000584.4 |
| A2AR | RT-A2AR-L |
TCAACAGCAACCTGCAGAAC | NM_000675.6 |
| RT-A2AR-R |
TCCAACCTAGCATGGGAGTC | NM_000675.6 |
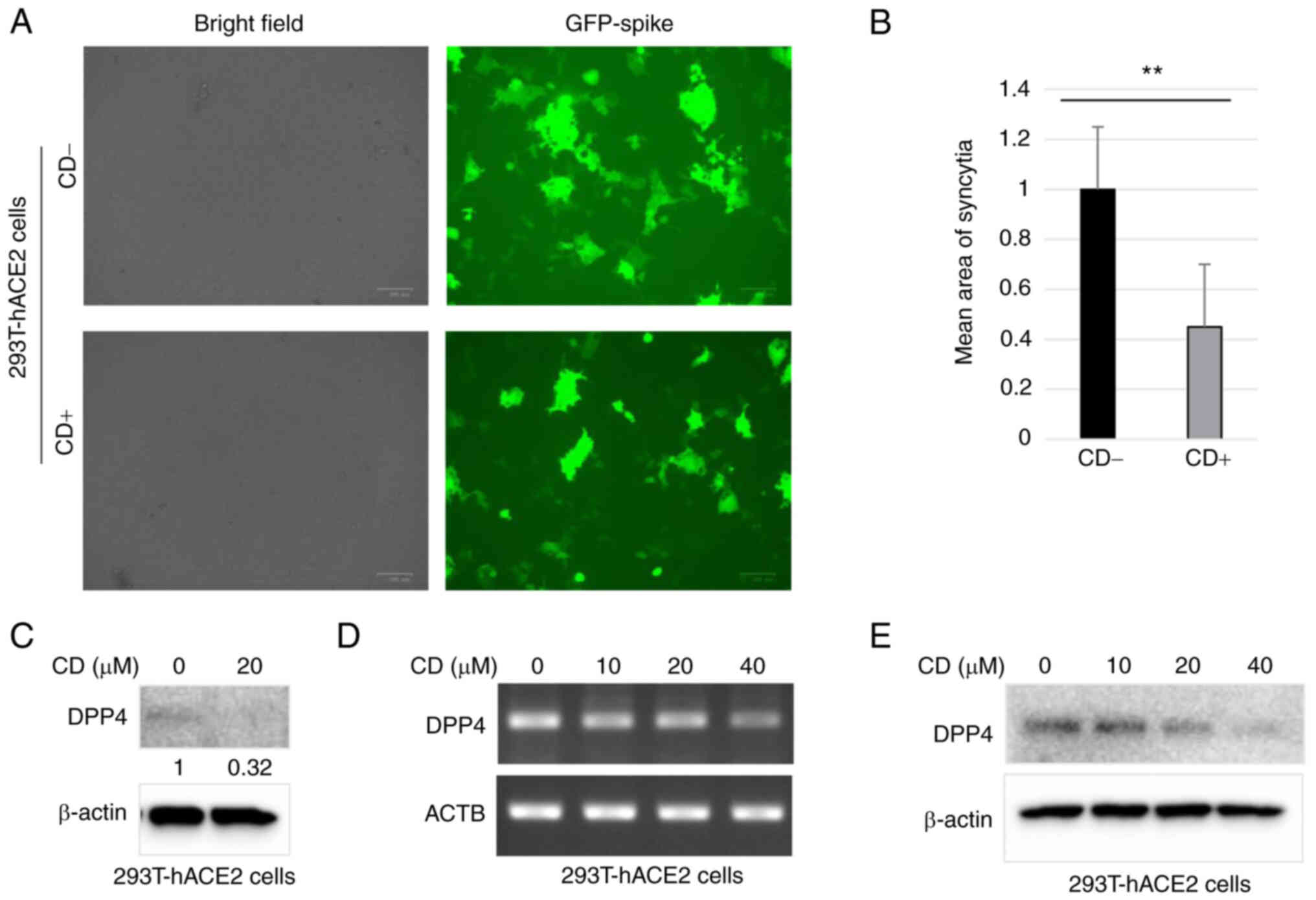
CD inhibits syncytial formation likely
through DPP4
Evidence indicates that a large number of
multinucleated cells characteristic of syncytial pathology are
present in patients with COVID-19 (73). This is a pathological hallmark of
SARS-CoV-2 infection. Syncytium formation is required for the
participation of SARS-CoV-2 Spike protein and host cell having the
human ACE2 gene (62). In
the present study, after treatment with CD, 293T-hACE2 cells were
transfected with SARS-CoV-2-Spike plasmids with GFP fluorescence.
Numerous large syncytia with GFP green fluorescence were observed
in control cells, indicating SARS-CoV-2 cell invasion (Fig. 12A; upper panels). Notably,
treatment with CD significantly decreased the area of fluorescence
of GFP-positive syncytia (Fig.
12A; bottom panels). The quantitative results are shown in
Fig. 12B; treatment with CD
significantly reduced the mean fluorescence area of syncytia
compared with the control group. The present study also
investigated whether treatment with CD reduced syncytia formation,
at least partially, through DPP4. Western blotting revealed that
the levels of DPP4 protein were significantly decreased in
293T-hACE2 cells following treatment with CD compared with control
(Fig. 12C). As expected, CD
inhibited DPP4 expression in 293T-hACE2 cells at both the protein
and mRNA levels in a dosage-dependent manner (Fig. 12D and E). Thus, CD may inhibit the
formation of syncytia through DPP4.
Discussion
The present study found that, in healthy tissues,
DPP4 was mainly expressed in in endocrine tissues, the
gastrointestinal tract, female tissue (mainly in placenta), male
tissue (mainly in prostate and seminal vesicle), proximal digestive
tract, kidneys, bladder, liver, gallbladder and the respiratory
system. Although the DPP4 RNA levels were low (lung; 16.0
nTPM), the DPP4 protein expression was moderate, demonstrating its
role in viral invasion in the lungs/bronchus/nasopharynx. Among 15
immune cell types, DPP4 mRNA was mainly expressed in T cells
(memory CD4 T-cell Th1/Th17). Compared with matched healthy
tissues, the levels of DPP4 were upregulated in ESCA, KIRP,
LIHC, LUAD, PAAD, PRAD, STAD, THCA and THYM. In contrast, they were
downregulated in BRCA, KICH, LUSC and SKCM. These findings
indicated the roles of DPP4 in viral invasion in most types of
cancer. Higher expression levels were associated with a long OS in
KIRC and MESO, implying that DPP4 could be a favorable marker.
However, high expression was also linked to a short OS in BLCA,
LGG, LUSC, PRAD and UVM, suggesting that DPP4 could be an
unfavorable marker. The isoform DPP4-001 includes both the DPPIV_N
domain and Peptidase_S9 domain with 766 amino acids, indicating its
function in tumorigenesis and SARS-CoV-2 invasion in patients with
different types of cancer. DNA methylation in BRCA, COAD and THCA
may be the mechanism regulating DPP4 expression. UCEC had the
highest mutation frequency (8.32%), followed by SKCM (4.95%); KIRC
had the lowest mutation frequency (0.39%). DPP4 expression
in tumor-immune systems revealed correlations between DPP4
expression and immune lymphocytes, receptors, chemokines,
immunosuppressants, immunostimulants and MHC molecules in almost
all types of cancer. These results suggested a therapeutic role of
DPP4 in cancer and SARS-CoV-2. The levels of soluble DPP4 (sDPP4)
levels were increased in patients with acute and chronic viral
infections; thus, the concentration of sDPP4 might be useful as a
biomarker for these diseases (74). DPP4 inhibitors (e.g., gliptins)
could be beneficial for patients with COVID-19, probably through
interference with viral invasion (74) and activation of inflammatory
pathways (75).
CD is a natural active substance derived from the
traditional Chinese medicine fungus Cordyceps militaris,
which possesses anticancer activity (76-78).
m62A is a modified ribonucleoside in tRNA
derived from Mycobacterium bovis Bacille Calmette-Guérin
(79). CD and
m62A are both derivatives of AD that suppress
cathepsin L (CTSL) expression in cancer cells. CTSL is another
receptor for SARS-CoV-2 (57).
Additionally, CD suppresses the expression of other SARS-CoV-2
receptors (e.g., FURIN and transmembrane serine protease 2), in
different cancer cells (47,80).
Evidence indicates that a large number of multinucleated cells
characteristic of syncytial pathology are present in patients with
COVID-19 (73). This is a
pathological hallmark of SARS-CoV-2 infection (62). Syncytium formation is required for
the participation of SARS-CoV-2 Spike protein and host cell having
human the ACE2 gene (62).
The present study demonstrated that CD inhibits syncytia formation
likely through DPP4. As expected, CD and m62A
inhibit DPP4 expression in cancer cells, suggesting anti-SARS-CoV-2
and anti-cancer role through suppression of DPP4 expression.
AD is regarded as a mainly metabolic and immune
checkpoint regulator in the tumor microenvironment, implicated in
tumor escape from the host immune system (81). Targeting the AD pathway could be
useful for cancer immunotherapy (82,83).
Markedly, AD inhibited DPP4 expression in five types of cancer
cells; this suggested its therapeutic potential as an
anti-SARS-CoV-2 agent by inhibiting DPP4 expression in cancer.
LAG-3 belongs to a novel class of immune checkpoint
receptors. It is highly expressed in TILs of various solid tumors,
such as colon cancer, hepatocellular carcinoma, head and neck
cancer, non-small cell lung cancer and pancreatic cancer. In
pathological states, LAG-3 is highly expressed at the TILs
surfaces, which positively correlates with the development and
occurrence of cancer (84).
Similar to programmed cell death protein 1 (PD-1) and cytotoxic
T-lymphocyte antigen 4, LAG-3 is considered a vital next-generation
immune checkpoint molecule (85).
Numerous LAG-3 inhibitors have been reported in 108 clinical trials
(https://clinicaltrials.gov/) (86). Encouragingly, the first anti-LAG-3
inhibitor (monoclonal antibody), relatlimab (also called
BMS-986016), has been approved for clinical use by the U.S. FDA.
Moreover, in March 2022, relatlimab in combination with nivolumab
(PD-1 inhibitor) was approved by the U.S. FDA under the name
opdualag. This is the first antibody approved for the treatment of
adult and pediatric patients with unresectable or metastatic
melanoma. This therapeutic regimen triggered synergistic immune
responses to increase the progression-free survival and reduce the
number of unresponsive patients. Nivolumab is an important
anti-PD-1 antibody for the treatment of different types of cancer
(87,88); it has been approved for clinical
use in more than 65 countries, including the USA and China
(87). By analyzing the predicted
genes for both DPP4 regulation and AD/A2AR signaling (69-72),
the present study showed that AD downregulated LAG-3 expression in
cancer cell lines, thereby demonstrating the anti-cancer
synergistic effects of DPP4 and AD regulation. However, further
research is warranted to elucidate the mechanism underlying DPP4
regulation and SARS-CoV-2 infection.
The present study showed that CD can inhibit viral
entry by pseudovirus-SARS-CoV-2 (Spike protein) experiments shown
in Fig. 12. Mouse experiments
involving AD treatments followed western blotting and RT-PCR,
showed in Fig. 11B and D. The two
may demonstrate the anti-SARS-CoV-2 or immune response. However,
the present study does not have data to show directly anti-cancer
effect in vitro and in vivo. This is a limitation
which may be addressed in the near future.
In conclusion, the present study revealed the
importance of DPP4 in different types of cancer, susceptibility to
SARS-CoV-2 attack and possible DPP4/AD/LAG-3 signaling. These data
also indicated potential immunotherapy options for SARS-CoV-2 by
targeting DPP4 using small molecules derived from natural products,
such as m62A, CD and AD. Further mechanism of
DPP4 regulation and SARS-CoV-2 infection by these small molecules
needs to be clarified in more detail.
Availability of data and materials
All data generated or analyzed during this study
are included in this published article.
Authors' contributions
JD, JiF, WZ, LZ and JC performed experimental
studies, data acquisition, data analysis and sample collection. JuF
and HC designed the project and analyzed the data. JuF and TH and
supervised the project. JuF wrote and edited the manuscript. JD and
JuF confirm the authenticity of all the raw data. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
The present study was reviewed and approved by the
Ethics Committee of Southwest Medical University, Sichuan, China
(approval no. 20221117-049). Informed consent was obtained from
patients for tissues samples.
Patient consent for publication
Not applicable
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The authors thank Dr Kai Wang and Mrs. Jiayue He
from the Research Center for Preclinical Medicine, Southwest
Medical University for their help in the syncytial formation
experiment.
Funding
The present study was supported by the Foundation of Science and
Technology Department of Sichuan Province (grant no.
2022NSFSC0737), the Joint Innovation Special Project of Science and
Technology Plan of Sichuan Province (grant no. 2022YFS0623-C3), in
part by the Research Foundation of Luzhou City (grant no.
2021-SYF-37) and the National Natural Science Foundation of China
(grant nos. 81672887, 82073263).
References
|
1
|
Daddona PE and Kelley WN: Human adenosine
deaminase. Stoichiometry of the adenosine deaminase-binding protein
complex. Biochim Biophys Acta. 580:302–311. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kameoka J, Tanaka T, Nojima Y, Schlossman
SF and Morimoto C: Direct association of adenosine deaminase with a
T cell activation antigen, CD26. Science. 261:466–469. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Morrison ME, Vijayasaradhi S, Engelstein
D, Albino AP and Houghton AN: A marker for neoplastic progression
of human melanocytes is a cell surface ectopeptidase. J Exp Med.
177:1135–1143. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Abbott CA, Baker E, Sutherland GR and
McCaughan GW: Genomic organization, exact localization, and tissue
expression of the human CD26 (dipeptidyl peptidase IV) gene.
Immunogenetics. 40:331–338. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Marguet D, Baggio L, Kobayashi T, Bernard
AM, Pierres M, Nielsen PF, Ribel U, Watanabe T, Drucker DJ and
Wagtmann N: Enhanced insulin secretion and improved glucose
tolerance in mice lacking CD26. Proc Natl Acad Sci USA.
97:6874–6879. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Conarello SL, Li Z, Ronan J, Roy RS, Zhu
L, Jiang G, Liu F, Woods J, Zycband E, Moller DE, et al: Mice
lacking dipeptidyl peptidase IV are protected against obesity and
insulin resistance. Proc Natl Acad Sci USA. 100:6825–6830. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Klemann C, Wagner L, Stephan M and von
Horsten S: Cut to the chase: A review of CD26/dipeptidyl
peptidase-4′s (DPP4) entanglement in the immune system. Clin Exp
Immunol. 185:1–21. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Vankadari N and Wilce JA: Emerging WuHan
(COVID-19) coronavirus: Glycan shield and structure prediction of
spike glycoprotein and its interaction with human CD26. Emerg
Microbes Infect. 9:601–604. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ohnuma K, Uchiyama M, Yamochi T,
Nishibashi K, Hosono O, Takahashi N, Kina S, Tanaka H, Lin X, Dang
NH and Morimoto C: Caveolin-1 triggers T-cell activation via CD26
in association with CARMA1. J Biol Chem. 282:10117–10131. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gines S, Marino M, Mallol J, Canela EI,
Morimoto C, Callebaut C, Hovanessian A, Casadó V, Lluis C and
Franco R: Regulation of epithelial and lymphocyte cell adhesion by
adenosine deaminase-CD26 interaction. Biochem J. 361:203–209. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ohnuma K, Hatano R, Komiya E, Otsuka H,
Itoh T, Iwao N, Kaneko Y, Yamada T, Dang NH and Morimoto C: A novel
role for CD26/dipeptidyl peptidase IV as a therapeutic target.
Front Biosci (Landmark Ed). 23:1754–1779. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang T, Tong X, Zhang S, Wang D, Wang L,
Wang Q and Fan H: The roles of dipeptidyl peptidase 4 (DPP4) and
DPP4 inhibitors in different lung diseases: New evidence. Front
Pharmacol. 12:7314532021. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
da Cruz Freire JE, Junior JEM, Pinheiro
DP, da Cruz Paiva Lima GE, do Amaral CL, Veras VR, Madeira MP,
Freire EBL, Ozório RG, Fernandes VO, et al: Evaluation of the
anti-diabetic drug sitagliptin as a novel attenuate to SARS-CoV-2
evidence-based in silico: Molecular docking and molecular dynamics.
3 Biotech. 12:3442022. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Scheen AJ: Cardiovascular effects of
dipeptidyl peptidase-4 inhibitors: From risk factors to clinical
outcomes. Postgrad Med. 125:7–20. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hu X, Wang X and Xue X: Therapeutic
perspectives of CD26 inhibitors in imune-mediated diseases.
Molecules. 27:44982022. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Thompson MA, Ohnuma K, Abe M, Morimoto C
and Dang NH: CD26/dipeptidyl peptidase IV as a novel therapeutic
target for cancer and immune disorders. Mini Rev Med Chem.
7:253–273. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Alkharsah KR, Aljaroodi SA, Rahman JU,
Alnafie AN, Al Dossary R, Aljindan RY, Alnimr AM and Hussen J: Low
levels of soluble DPP4 among Saudis may have constituted a risk
factor for MERS endemicity. PLoS One. 17:e02666032022. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang N, Shi X, Jiang L, Zhang S, Wang D,
Tong P, Guo D, Fu L, Cui Y, Liu X, et al: Structure of MERS-CoV
spike receptor-binding domain complexed with human receptor DPP4.
Cell Res. 23:986–993. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sedo A, Krepela E, Kasafirek E, Kraml J
and Kadlecova L: Dipeptidyl peptidase IV in the human lung and
spinocellular lung cancer. Physiol Res. 40:359–362. 1991.PubMed/NCBI
|
|
20
|
Bishnoi R, Hong YR, Shah C, Ali A, Skelton
WP IV, Huo J, Dang NH and Dang LH: Dipeptidyl peptidase 4
inhibitors as novel agents in improving survival in diabetic
patients with colorectal cancer and lung cancer: A surveillance
epidemiology and endpoint research medicare study. Cancer Med.
8:3918–3927. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Colice G, Price D, Gerhardsson de Verdier
M, Rabon-Stith K, Ambrose C, Cappell K, Irwin DE, Juneau P and
Vlahiotis A: The effect of DPP-4 inhibitors on asthma control: An
administrative database study to evaluate a potential
pathophysiological relationship. Pragmat Obs Res. 8:231–240. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Raj VS, Smits SL, Provacia LB, van den
Brand JM, Wiersma L, Ouwendijk WJ, Bestebroer TM, Spronken MI, van
Amerongen G, Rottier PJ, et al: Adenosine deaminase acts as a
natural antagonist for dipeptidyl peptidase 4-mediated entry of the
Middle East respiratory syndrome coronavirus. J Virol.
88:1834–1838. 2014. View Article : Google Scholar :
|
|
23
|
deKay JT, May TL, Riker RR, Rud J, Gagnon
DJ, Sawyer DB, Seder DB and Ryzhov S: The number of circulating
CD26 expressing cells is decreased in critical COVID-19 illness.
Cytometry A. Mar 16–2022.Epub ahead of print.
|
|
24
|
Cameron K, Rozano L, Falasca M and Mancera
RL: Does the SARS-CoV-2 spike protein receptor binding domain
interact effectively with the DPP4 (CD26) Receptor? A Molecular
Docking Study. Int J Mol Sci. 22:70012021. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Govender Y, Shalekoff S, Ebrahim O, Waja
Z, Chaisson RE, Martinson N and Tiemessen CT: Systemic DPP4/CD26 is
associated with natural HIV-1 control: Implications for COVID-19
susceptibility. Clin Immunol. 230:1088242021. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nadasdi A, Sinkovits G, Bobek I, Lakatos
B, Förhécz Z, Prohászka ZZ, Réti M, Arató M, Cseh G, Masszi T, et
al: Decreased circulating dipeptidyl peptidase-4 enzyme activity is
prognostic for severe outcomes in COVID-19 inpatients. Biomark Med.
16:317–330. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Radzikowska U, Ding M, Tan G, Zhakparov D,
Peng Y, Wawrzyniak P, Wang M, Li S, Morita H, Altunbulakli C, et
al: Distribution of ACE2, CD147, CD26, and other SARS-CoV-2
associated molecules in tissues and immune cells in health and in
asthma, COPD, obesity, hypertension, and COVID-19 risk factors.
Allergy. 75:2829–2845. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kawasaki T, Chen W, Htwe YM, Tatsumi K and
Dudek SM: DPP4 inhibition by sitagliptin attenuates LPS-induced
lung injury in mice. Am J Physiol Lung Cell Mol Physiol.
315:L834–L845. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kifle ZD, Woldeyohanin AE and Demeke CA:
SARS-CoV-2 and diabetes: A potential therapeutic effect of
dipeptidyl peptidase 4 inhibitors in diabetic patients diagnosed
with COVID-19. Metabol Open. 12:1001342021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nitulescu GM, Paunescu H, Moschos SA,
Petrakis D, Nitulescu G, Ion GND, Spandidos DA, Nikolouzakis TK,
Drakoulis N and Tsatsakis A: Comprehensive analysis of drugs to
treat SARSCoV2 infection: Mechanistic insights into current COVID19
therapies (Review). Int J Mol Med. 46:467–488. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gordon DE, Jang GM, Bouhaddou M, Xu J,
Obernier K, White KM, O'Meara MJ, Rezelj VV, Guo JZ, Swaney DL, et
al: A SARS-CoV-2 protein interaction map reveals targets for drug
repurposing. Nature. 583:459–468. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pal R, Banerjee M, Mukherjee S, Bhogal RS,
Kaur A and Bhadada SK: Dipeptidyl peptidase-4 inhibitor use and
mortality in COVID-19 patients with diabetes mellitus: An updated
systematic review and meta-analysis. Ther Adv Endocrinol Metab.
12:20420188219964822021. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Han T, Ma S, Sun C, Zhang H, Qu G, Chen Y,
Cheng C, Chen EL, Ayaz Ahmed M, Kim KY, et al: Association between
anti-diabetic agents and clinical outcomes of COVID-19 in patients
with diabetes: A systematic review and meta-analysis. Arch Med Res.
53:186–195. 2022. View Article : Google Scholar
|
|
34
|
Zein A and Raffaello WM: Dipeptidyl
peptidase-4 (DPP-IV) inhibitor was associated with mortality
reduction in COVID-19-A systematic review and meta-analysis. Prim
Care Diabetes. 16:162–167. 2022. View Article : Google Scholar
|
|
35
|
Rakhmat II, Kusmala YY, Handayani DR,
Juliastuti H, Nawangsih EN, Wibowo A, Lim MA and Pranata R:
Dipeptidyl peptidase-4 (DPP-4) inhibitor and mortality in
coronavirus disease 2019 (COVID-19)-A systematic review,
meta-analysis, and meta-regression. Diabetes Metab Syndr.
15:777–782. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Carrasco-Sanchez FJ, Carretero-Anibarro E,
Gargallo MÁ, Gómez-Huelgas R, Merino-Torres JF, Orozco-Beltrán D,
Pines Corrales PJ and Ruiz Quintero MA: Executive Summary from
Expert consensus on effectiveness and safety of iDPP-4 in the
treatment of patients with diabetes and COVID-19. Endocrinol
Diabetes Nutr (Engl Ed). 69:209–218. 2022.PubMed/NCBI
|
|
37
|
Shestakova MV, Vikulova OK, Elfimova AR,
Deviatkin AA, Dedov II and Mokrysheva NG: Risk factors for COVID-19
case fatality rate in people with type 1 and type 2 diabetes
mellitus: A nationwide retrospective cohort study of 235,248
patients in the Russian Federation. Front Endocrinol (Lausanne).
13:9098742022. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Solerte SB, Di Sabatino A, Galli M and
Fiorina P: Dipeptidyl peptidase-4 (DPP4) inhibition in COVID-19.
Acta Diabetol. 57:779–783. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sebastian-Martin A, Sanchez BG,
Mora-Rodriguez JM, Bort A and Diaz-Laviada I: Role of dipeptidyl
Peptidase-4 (DPP4) on COVID-19 physiopathology. Biomedicines.
10:20262022. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ojha R, Gurjar K, Ratnakar TS, Mishra A
and Prajapati VK: Designing of a bispecific antibody against
SARS-CoV-2 spike glycoprotein targeting human entry receptors DPP4
and ACE2. Hum Immunol. 83:346–355. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Elkrief A, Hennessy C, Kuderer NM,
Rubinstein SM, Wulff-Burchfield E, Rosovsky RP, Vega-Luna K,
Thompson MA, Panagiotou OA, Desai A, et al: Geriatric risk factors
for serious COVID-19 outcomes among older adults with cancer: A
cohort study from the COVID-19 and Cancer Consortium. Lancet
Healthy Longev. 3:e143–e152. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Desai A, Gupta R, Advani S, Ouellette L,
Kuderer NM, Lyman GH and Li A: Mortality in hospitalized patients
with cancer and coronavirus disease 2019: A systematic review and
meta-analysis of cohort studies. Cancer. 127:1459–1468. 2021.
View Article : Google Scholar
|
|
43
|
Grivas P, Khaki AR, Wise-Draper TM, French
B, Hennessy C, Hsu CY, Shyr Y, Li X, Choueiri TK, Painter CA, et
al: Association of clinical factors and recent anticancer therapy
with COVID-19 severity among patients with cancer: A report from
the COVID-19 and Cancer Consortium. Ann Oncol. 32:787–800. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Fu C, Stoeckle JH, Masri L, Pandey A, Cao
M, Littman D, Rybstein M, Saith SE, Yarta K, Rohatgi A, et al:
COVID-19 outcomes in hospitalized patients with active cancer:
Experiences from a major New York City health care system. Cancer.
127:3466–3475. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Beckenkamp A, Davies S, Willig JB and
Buffon A: DPPIV/CD26: A tumor suppressor or a marker of malignancy?
Tumour Biol. 37:7059–7073. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Fu J, Liao L, Balaji KS, Wei C, Kim J and
Peng J: Epigenetic modification and a role for the E3 ligase RNF40
in cancer development and metastasis. Oncogene. 40:465–474. 2021.
View Article : Google Scholar :
|
|
47
|
Li D, Liu X, Zhang L, He J, Chen X, Liu S,
Fu J, Fu S, Chen H, Fu J and Cheng J: COVID-19 disease and
malignant cancers: The impact for the furin gene expression in
susceptibility to SARS-CoV-2. Int J Biol Sci. 17:3954–3967. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Uhlen M, Fagerberg L, Hallstrom BM,
Lindskog C, Oksvold P, Mardinoglu A, Sivertsson Å, Kampf C,
Sjöstedt E, Asplund A, et al: Proteomics. Tissue-based map of the
human proteome. Science. 347:12604192015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Uhlen M, Zhang C, Lee S, Sjöstedt E,
Fagerberg L, Bidkhori G, Benfeitas R, Arif M, Liu Z, Edfors F, et
al: A pathology atlas of the human cancer transcriptome. Science.
357:eaan25072017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res. 45:W98–W102.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Tang Z, Kang B, Li C, Chen T and Zhang Z:
GEPIA2: An enhanced web server for large-scale expression profiling
and interactive analysis. Nucleic Acids Res. 47:W556–W560. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ding W, Chen J, Feng G, Chen G, Wu J, Guo
Y, Ni X and Shi T: DNMIVD: DNA methylation interactive
visualization database. Nucleic Acids Res. 48:D856–D862. 2020.
View Article : Google Scholar :
|
|
53
|
Cerami E, Gao J, Dogrusoz U, Gross BE,
Sumer SO, Aksoy BA, Jacobsen A, Byrne CJ, Heuer ML, Larsson E, et
al: The cBio cancer genomics portal: An open platform for exploring
multidimensional cancer genomics data. Cancer Discov. 2:401–404.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ru B, Wong CN, Tong Y, Zhong JY, Zhong
SSW, Wu WC, Chu KC, Wong CY, Lau CY, Chen I, et al: TISIDB: An
integrated repository portal for tumor-immune system interactions.
Bioinformatics. 35:4200–4202. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Fu J, Li L and Lu G: Relationship between
microdeletion on Y chromosome and patients with idiopathic
azoospermia and severe oligozoospermia in the Chinese. Chin Med J
(Engl). 115:72–75. 2002.PubMed/NCBI
|
|
56
|
Zhang L, Yang M, Gan L, He T, Xiao X,
Stewart MD, Liu X, Yang L, Zhang T, Zhao Y and Fu J: DLX4
upregulates TWIST and enhances tumor migration, invasion and
metastasis. Int J Biol Sci. 8:1178–1187. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhang L, Wei C, Li D, He J, Liu S, Deng H,
Cheng J, Du J, Liu X, Chen H, et al: COVID-19 receptor and
malignant cancers: Association of CTSL expression with
susceptibility to SARS-CoV-2. Int J Biol Sci. 18:2362–2371. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Wang K, Deng H, Song B, He J, Liu S, Fu J,
Zhang L, Li D, Balaji KS, Mei Z, et al: The correlation between
immune invasion and SARS-COV-2 entry protein ADAM17 in cancer
patients by bioinformatic analysis. Front Immunol. 13:9235162022.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Wei C, Liu Y, Liu X, Cheng J and Fu J,
Xiao X, Moses RE, Li X and Fu J: The speckle-type POZ protein
(SPOP) inhibits breast cancer malignancy by destabilizing TWIST1.
Cell Death Discov. 8:3892022. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Fu J, Song B, Du J, Liu S, He J, Xiao T,
Zhou B, Li D, Liu X, He T, et al: Impact of BSG/CD147 gene
expression on diagnostic, prognostic and therapeutic strategies
towards malignant cancers and possible susceptibility to
SARS-CoV-2. Mol Biol Rep. 1–13. 2022. View Article : Google Scholar : Epub ahead of
print. PubMed/NCBI
|
|
61
|
Liu G, Du W, Sang X, Tong Q, Wang Y, Chen
G, Yuan Y, Jiang L, Cheng W, Liu D, et al: RNA G-quadruplex in
TMPRSS2 reduces SARS-CoV-2 infection. Nat Commun. 13:14442022.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Jocher G, Grass V, Tschirner SK, Riepler
L, Breimann S, Kaya T, Oelsner M, Hamad MS, Hofmann LI, Blobel CP,
et al: ADAM10 and ADAM17 promote SARS-CoV-2 cell entry and spike
protein-mediated lung cell fusion. EMBO Rep. 23:e543052022.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Fu J, Zhou B, Zhang L, Balaji KS, Wei C,
Liu X, Chen H, Peng J and Fu J: Expressions and significances of
the angiotensin-converting enzyme 2 gene, the receptor of
SARS-CoV-2 for COVID-19. Mol Biol Rep. 47:4383–4392. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Györffy B, Lanczky A, Eklund AC, Denkert
C, Budczies J, Li Q and Szallasi Z: An online survival analysis
tool to rapidly assess the effect of 22,277 genes on breast cancer
prognosis using microarray data of 1,809 patients. Breast Cancer
Res Treat. 123:725–731. 2010. View Article : Google Scholar
|
|
65
|
Blume C, Jackson CL, Spalluto CM, Legebeke
J, Nazlamova L, Conforti F, Perotin JM, Frank M, Butler J, Crispin
M, et al: A novel ACE2 isoform is expressed in human respiratory
epithelia and is upregulated in response to interferons and RNA
respiratory virus infection. Nat Genet. 53:205–214. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Onabajo OO, Banday AR, Stanifer ML, Yan W,
Obajemu A, Santer DM, Florez-Vargas O, Piontkivska H, Vargas JM,
Ring TJ, et al: Interferons and viruses induce a novel truncated
ACE2 isoform and not the full-length SARS-CoV-2 receptor. Nat
Genet. 52:1283–1293. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Chen PJ, Lu HJ, Nassef Y, Lin CW, Chuang
CY, Lee CY, Chiu YW, Yang SF and Yang WE: Association of dipeptidyl
peptidase IV polymorphism with clinicopathological characters of
oral cancer. J Oral Pathol Med. 51:730–737. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Posadas-Sanchez R, Sanchez-Munoz F,
Guzman-Martin CA, Hernández-Díaz Couder A, Rojas-Velasco G, Fragoso
JM and Vargas-Alarcón G: Dipeptidylpeptidase-4 levels and DPP4 gene
polymorphisms in patients with COVID-19. Association with disease
and with severity. Life Sci. 276:1194102021. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Cekic C and Linden J: Purinergic
regulation of the immune system. Nat Rev Immunol. 16:177–192. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Fong L, Hotson A, Powderly JD, Sznol M,
Heist RS, Choueiri TK, George S, Hughes BGM, Hellmann MD, Shepard
DR, et al: Adenosine 2A receptor blockade as an immunotherapy for
treatment-refractory renal cell cancer. Cancer Discov. 10:40–53.
2020. View Article : Google Scholar
|
|
71
|
Novitskiy SV, Ryzhov S, Zaynagetdinov R,
Goldstein AE, Huang Y, Tikhomirov OY, Blackburn MR, Biaggioni I,
Carbone DP, Feoktistov I and Dikov MM: Adenosine receptors in
regulation of dendritic cell differentiation and function. Blood.
112:1822–1831. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Liu J, Shi Y, Liu X, Zhang D, Bai Y, Xu Y
and Wang M: Blocking Adenosine/A2AR pathway for cancer therapy.
Zhongguo Fei Ai Za Zhi. 25:460–467. 2022.In Chinese. PubMed/NCBI
|
|
73
|
Braga L, Ali H, Secco I, Chiavacci E,
Neves G, Goldhill D, Penn R, Jimenez-Guardeño JM, Ortega-Prieto AM,
Bussani R, et al: Drugs that inhibit TMEM16 proteins block
SARS-CoV-2 spike-induced syncytia. Nature. 594:88–93. 2021.
View Article : Google Scholar :
|
|
74
|
Krejner-Bienias A, Grzela K and Grzela T:
DPP4 inhibitors and COVID-19-holy grail or another dead end? Arch
Immunol Ther Exp (Warsz). 69:12021. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Alomair BM, Al-Kuraishy HM, Al-Buhadily
AK, Al-Gareeb AI, De Waard M, Elekhnawy E and Batiha GE: Is
sitagliptin effective for SARS-CoV-2 infection: False or true
prophecy? Inflammopharmacology. 30:2411–2415. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Radhi M, Ashraf S, Lawrence S, Tranholm
AA, Wellham PAD, Hafeez A, Khamis AS, Thomas R, McWilliams D and de
Moor CH: A systematic review of the biological effects of
cordycepin. Molecules. 26:58862021. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Tima S, Tapingkae T, To-Anun C, Noireung
P, Intaparn P, Chaiyana W, Sirithunyalug J, Panyajai P,
Viriyaadhammaa N, Nirachonkul W, et al: Antileukaemic cell
proliferation and cytotoxic activity of edible golden cordyceps
(Cordyceps militaris) extracts. Evid Based Complement Alternat Med.
2022:53477182022. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Wei C, Khan MA, Du J, Cheng J, Tania M,
Leung EL and Fu J: Cordycepin Inhibits triple-negative breast
cancer cell migration and invasion by regulating EMT-TFs SLUG,
TWIST1, SNAIL1, and ZEB1. Front Oncol. 12:8985832022. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Chan CT, Chionh YH, Ho CH, Lim KS, Babu
IR, Ang E, Wenwei L, Alonso S and Dedon PC: Identification of N6,
N6-dimethyladenosine in transfer RNA from Mycobacterium bovis
Bacille Calmette-Guerin. Molecules. 16:5168–5181. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Fu J, Liu S, Tan Q, Liu Z, Qian J, Li T,
Du J, Song B, Li D, Zhang L, et al: Impact of TMPRSS2 Expression,
Mutation Prognostics, and Small Molecule (CD, AD, TQ, and TQFL12)
Inhibition on Pan-Cancer Tumors and Susceptibility to SARS-CoV-2.
Molecules. 27:74132022. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Boison D and Yegutkin GG: Adenosine
metabolism: Emerging concepts for cancer therapy. Cancer Cell.
36:582–596. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Hammami A, Allard D, Allard B and Stagg J:
Targeting the adenosine pathway for cancer immunotherapy. Semin
Immunol. 42:1013042019. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Vijayan D, Young A, Teng MWL and Smyth MJ:
Targeting immunosuppressive adenosine in cancer. Nat Rev Cancer.
17:709–724. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Huo JL, Wang YT, Fu WJ, Lu N and Liu ZS:
The promising immune checkpoint LAG-3 in cancer immunotherapy: From
basic research to clinical application. Front Immunol.
13:9560902022. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Chocarro L, Blanco E, Zuazo M, Arasanz H,
Bocanegra A, Fernández-Rubio L, Morente P, Fernández-Hinojal G,
Echaide M, Garnica M, et al: Understanding LAG-3 Signaling. Int J
Mol Sci. 22:52822021. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Chocarro L, Bocanegra A, Blanco E,
Fernández-Rubio L, Arasanz H, Echaide M, Garnica M, Ramos P,
Piñeiro-Hermida S, Vera R, et al: Cutting-Edge: Preclinical and
clinical development of the first approved Lag-3 inhibitor. Cells.
11:23512022. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Robert C, Long GV, Brady B, Dutriaux C,
Maio M, Mortier L, Hassel JC, Rutkowski P, McNeil C,
Kalinka-Warzocha E, et al: Nivolumab in previously untreated
melanoma without BRAF mutation. N Engl J Med. 372:320–330. 2015.
View Article : Google Scholar
|
|
88
|
Ansell SM, Lesokhin AM, Borrello I,
Halwani A, Scott EC, Gutierrez M, Schuster SJ, Millenson MM, Cattry
D, Freeman GJ, et al: PD-1 blockade with nivolumab in relapsed or
refractory Hodgkin's lymphoma. N Engl J Med. 372:311–319. 2015.
View Article : Google Scholar :
|