Stress can be defined as a state of disrupted
homeostasis triggered by intrinsic or extrinsic stressors, which is
counteracted by a plethora of physiological and behavioural
adaptive responses aiming to re-establish the altered equilibrium
of the organism (1-3). The concept of a corporal 'steady
state' that was defined with the Greek-derived term
'homeostasis' in the beginning of the 20th century, was
initially described by ancient Greek natural philosophers, with the
words 'harmonious balance', and 'isonomia', later termed 'eucrasia'
by Hippocrates and 'eustatheia', by Epicurus, revealing an
intellectual understanding of this fundamental concept (1). Lifestyle in contemporary
civilizations has evolved and changed considerably from that of our
forefathers, and in combination with the lengthening of human life
expectancy, has allowed the currently high incidence of
'affluence-related' disorders (1,4).
External stressors of modern life, mainly chronic
psycho-socio-economic stress and protracted crises, such as the
circumstances encountered with the COVID-19 pandemic, economic
conditions and climate change, underscore the need to further
comprehend stress and its effects on humanity (5-10).
This is of particular importance, as uncontrolled chronic stress
can have unfavourable, potentially hazardous outcomes, as evidenced
by an ever-growing list of stress-related disorders, including
several forms of cancer (2,5,11).
Stress has been linked to cancer development and
incidence for a number of decades, if not millennia; however,
epidemiological studies and clinical trials have produced
contradictory results (3,11,12). Thus, psychological stressors have
been linked to the development of cancer since the 2nd century CE,
with the ancient Greek physician, Galen, noticing that tumours of
the reproductive organs were more frequent in women with
'melancholic natures' (12).
Other researchers have noted the importance of psychological
variables in the occurrence of cancer in women, such as those with
'greater sensitivity and frustration'; however, these studies were
based on limited observations and/or personal concerns (12,13).
It has been consistently demonstrated that the
immune system plays a critical role in inhibiting cancer
progression (12,14). Numerous preclinical and clinical
psychoneuroimmunological and neurobiological investigations have
been published over the past three decades, delving into the
processes behind the linkages between stress and cancer, and have
revealed molecular, cellular and endocrine processes that may be
implicated in these effects (12,15-18). Animal studies have revealed a
clearer link than clinical human studies, indicating that stress
can exacerbate the hallmarks of cancer, promoting tumour growth and
metastasis by directly altering the molecular properties of
malignant tissue, its microenvironment, its anti-host immune
reaction activity, and other indirect cancer progression modifiers,
as will be further analysed in the present review (12). Of note, discrepancies in
preclinical and clinical or epidemiological research observations
may be explained as follows: Firstly, preclinical studies link
stressful conditions or stress-relieving activities with phases of
cancer development on natural or transplanted tumours that are
particularly susceptible to the effects of stress; secondly,
theoretical and methodological challenges in carrying out human
studies that obscure the influence of stress on cancer development
(12).
The aim of the present review was to discuss stress
and its relation to cancer, examining various pathways that drive
carcinogenesis. The review initially summarizes epidemiological
data from human studies examining the risk of cancer development
and progression related to stress. Focus is then paid to the
mechanistic aspects of stress physiology and the discussion of the
mechanisms through which stress affects the molecular features of
malignant tissue, its microenvironment and its anti-host immune
reaction. Furthermore, other, indirect cancer progression
modifiers, that promote the growth and spread of numerous cancer
forms are also reviewed. The synthesis of the present review may
have practical clinical implications.
There is accumulating evidence to indicate that
stress increases the risk of developing cancer; nevertheless, not
all human studies on this topic are consistent (19). A meta-analysis of 12 European
cohort studies found no association between work-related stress and
overall cancer risk or, more specifically, colorectal, lung,
breast, or prostate cancer risk (20). However, another meta-analysis that
investigated the association between work-related stress and cancer
risk, and focused on colorectal, lung and oesophageal cancers,
found a significant association between stress and the risk of
cancer development in populations primarily from North America and
Europe (21). In addition,
another meta-analysis of 53 studies indicated that stress-related
psychosocial variables were linked to an increased cancer incidence
in healthy populations, a decreased survival time and an increased
mortality rate in patients with cancer (22). Stress-prone personalities,
unfavourable coping mechanisms, negative emotional responses and a
poor quality of life have also been linked to an increased cancer
incidence and mortality rate, as well as to a decreased survival
time (22). Of note, the same
meta-analysis found that there were publication biases and
methodological heterogeneity and potential errors in the studies
examined; the authors of that study suggested caution in
interpreting the findings and emphasized the need for further
investigations (11,23). Previous studies have linked
specific stressors, such as cold climates, bereavement, war and
depression, to a higher incidence of cancers (24-27). However, others studies have shown
no association between stress and ovarian or breast cancer
(28,29).
Examining stress and cancer progression (often by
evaluating the survival rates of patients with cancer) can be
relatively challenging. Stress, including life events, is often
assessed without regarding the time of cancer detection, while its
impact on cancer progression is not assessed (11). In addition, the majority of
patients with cancer experience some levels of distress, which may
influence cancer progression regardless of baseline stress levels;
this may mask the association between stress levels and cancer
progression, but could allow for the observation of the beneficial
effects of stress-reducing interventions (11,30). Emotional distress in patients with
cancer increases mental health issues, which, in turn, may affect
cancer prognosis and increase mortality rates (31,32). Psychological stress and discomfort
have also been linked to increased mortality rates (33). A recent meta-analysis demonstrated
that stressor-specific and cancer-specific effects on survival were
evident (11). Depression in
patients with breast cancer increases the risk of cancer-specific
mortality, while low social support in combination with depression
may increase the risk of cancer-related mortality (11,34-36).
The inconsistent effects of stress on cancer
development and incidence and heterogeneous approaches, preclude a
solid aetiological relation. The subjective stress perception of
patients with cancer is influenced by disease burden, resulting in
biased retrospective assessments (11). Malignant transformation in humans
is a prolonged process characterized by long-term 'dormancy' and a
high prevalence of subclinical cancer (37). Cancer incidence may be increased
by disease initiation, escape from dormancy, or a more rapid
progression to clinical manifestation (11). Despite the controversies
associated with human studies, data from animal models are more
consistent, as is presented below, in the pathophysiological
sections of the present review, following a brief description of
stress physiology.
The 'stress syndrome' corresponds to the
physiological adaptive response that coordinates homeostasis and
protects organisms during acute stress (1,46).
It involves the central nervous system (CNS) and peripheral organs
and tissues, and it facilitates adaptive functions, such as arousal
and cardiopulmonary function, and inhibits non-adaptive ones, such
as eating, growth and reproduction (47). Stress-related changes increase
oxygenation and nutrient supply to the brain, heart and skeletal
muscles, crucial for central coordination and the 'fight or flight'
reaction (1,48). The CNS retains basal homeostasis
and processes and integrate responses to various stimuli, including
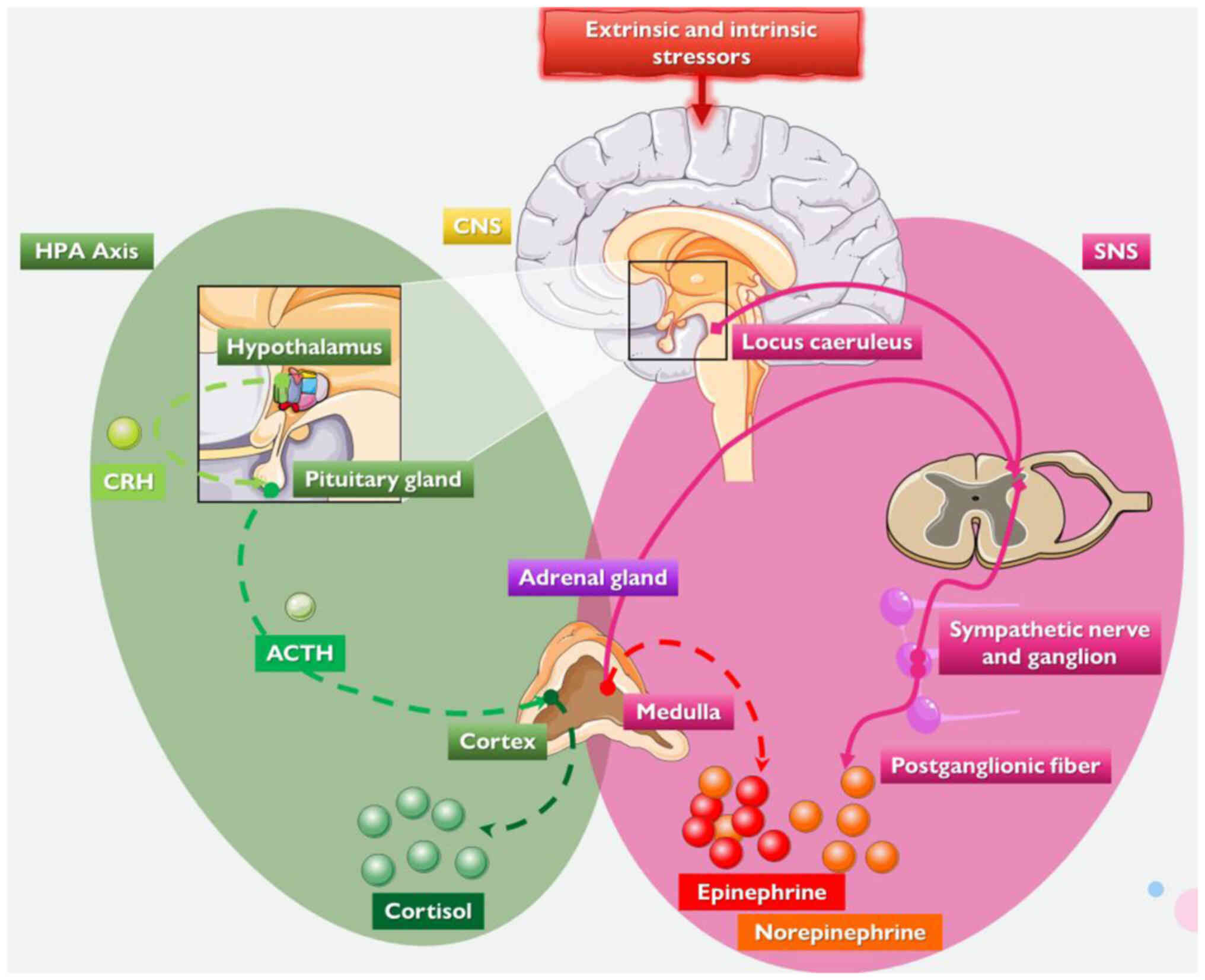
physiological reactions to external or internal stressors (1,3,42-44). The CNS orchestrates the complex
adaptation to stress by stimulating the sympathetic nervous system
(SNS) and the hypothalamic-pituitary-adrenal (HPA) axis, inducing
the secretion respectively of cortisol and adrenergic hormones,
primarily noradrenaline (norepinephrine) and adrenaline
(epinephrine), while it withdraws the activity of the
parasympathetic nervous system (3,42-46).
The HPA axis is activated or dysregulated by mental
health issues and/or behavioural changes, including depression and
social isolation, influencing pro-inflammatory and
anti-inflammatory stress-related immunomodulatory molecules and
pathways [including the cellular glucocorticoid signalling system,
interleukin (IL)-2 and interferon-γ] (1,11,43,49-52). The hypothalamus, part of the HPA
axis, secretes corticotropin-releasing hormone, stimulating
adrenocorticotropic hormone secretion by the anterior pituitary
gland, which stimulates the production of glucocorticoids by the
adrenal cortex, i.e., cortisol in primates and corticosterone in
rodents (1,11,43,53).
During stress, the SNS increases the release of the
catecholamines norepinephrine and epinephrine by the systemic
sympathetic system and the adrenal medulla (1,11,43). The neurohormones norepinephrine
and epinephrine cause arousal, increase the metabolic rate,
stimulate the cardiopulmonary system, enhance gluconeogenesis,
glycogenolysis, proteolysis and lipolysis, and increase catabolism
(1,3,11,42-44). Stress-induced hormones also affect
other key physiological and biochemical procedures, including
various brain networks (e.g., reward), and the water and
electrolyte equilibrium (1,3,11,42-44).
The dose-response curve for homeostatic processes,
including the stress system, has an inverted U shape (1,46,48,54). Basal, healthy homeostasis occurs
in the centre of the curve, the ideal region (1,46,48). Inadequate adaptation, known as low
allostasis or 'cacostasis', or an excessive response, known
as high allostasis, or 'cacostasis', may yield suboptimal
consequences (1). In high
allostasis, both the intensity and duration of stressors are key
predictors of their effects. Thus, both hypofunction and
hyperfunction of the homeostatic systems may have negative
consequences, including a decreased survival and higher morbidity
(1,46,48). When the stress exceeds the ability
of the individual to manage it, it becomes deleterious, and the
risk for illness increases by ~20% (11,55-57).
The interaction of homeostasis-disrupting stressors
and stressor-activated adaptive responses can result in one of
three outcomes: normal match, which yields the organism to its
basal homeostasis or 'eustasis'; defective match, which
results in 'cacostasis'; or improved match, which results in
a new, more resilient equilibrium, 'hyperstasis' (1,46,48). Patients are at a greater risk when
allostasis becomes demanding and the allostatic load exceeds
overload thresholds (11,55-57). The duration and intensity of the
response to stress vary significantly among individuals, and are
influenced by physiological factors, psychosocial characteristics
and previous stressful life events, such as childhood trauma
(11,55-57). As a result, patients may respond
differently to stressors such as cancer diagnosis, treatment and
survival.
Overall, the predominant effects of deleterious
stress can lead to the development of various chronic diseases and
comorbidities, including dysmetabolic conditions and
cardiometabolic diseases, which predispose to cancer development
via various mechanisms, indirect or direct (vide infra)
(2,58-65). The following sections focus on
cancer pathophysiology and its hallmarks, and thereafter, on the
direct impact of stress on these.
Multiple pathways are involved in the development of
cancer, including the upregulation of pro-oncogenes and the
suppression of onco-suppressor genes, as reviewed extensively
elsewhere (66-73). In oncogenesis, the shift from a
cell's original state to a malignant state is a process that
requires the cell to surmount its anti-oncogenic milestones
(74,75). Cancer is caused by various genetic
and epigenetic alterations in (stem) cells, primarily involving
mutations, deletions, inversions, amplifications and chromosome
translocations resulting, among others, in an altered oncogene
activity (66,76-78). Based on these characteristics, the
hallmarks of cancer have been compiled, as described Hanahan and
Weinberg (79,80). These features, which are
distinguishing characteristics with evolutionary benefits (80,81), include the capability of infinite
cell proliferation, persistent angiogenesis, resilience to cell
death, the potential of invasion and metastasis, the ability to
evade growth inhibitors, and self-sufficiency in growth factors
(74,79,82,83).
The dysregulation of metabolism, a mechanism that
plays a critical role in cellular stress signalling pathways and
procedures (including mitochondrial functions), and the avoidance
of the immune system are two additional characteristics of cancer
that have recently emerged (74,80,84). One of the most essential qualities
of tumour cells is their capacity to withstand environmental
stresses, such as hypoxia, nutritional deprivation and DNA-damaging
agents (74,83). Cellular stress is an extrinsic
element that influences cancer formation and development. It
consists of oxidative stress generated by reactive oxygen species,
metabolic stress owing to increased metabolic demands and genotoxic
stress, which involves DNA damage (74,85). Cellular stress generally initiates
the process of cell death (74,85). Nevertheless, cancer cells can
tolerate cellular stress by modifying gene expression, metabolism
and escaping growth inhibitory signals (74,80,81,83,86). Notably, pre-malignant or malignant
foci can be eliminated, become dormant or grow slowly, or progress
to clinical manifestation (37).
Some phases of this heterogeneous non-linear process
may theoretically be more important than others (11). Examples include activating the
angiogenic switch, allowing growth or escape from dormancy,
interacting with immune cells, circulating tumour cells passing
through capillaries, and the survival of tumour-associated
lymphocytes (87-92). During such potentially critical
times, the impact of stress may be amplified (11,74,85). Furthermore, whether stress
exacerbates or alleviates malignant processes may be affected by
the stage of malignant growth, unique tumour features and the range
of stress responses (11).
Additionally, immune-tumour interactions may either attenuate or
accelerate tumour development, and stress hormones can influence
both processes (15,51,93-96). Consequently, it is anticipated
that interactions between stress and cancer would be non-linear,
with the impact of stress potentially altering the anticipated
responses, depending on the stage of cancer progression (11).
Synchronized acute or chronic stress events with a
critical cancer phase may, in theory, exert a more prominent effect
on cancer growth than non-synchronized events (11). Animal models can focus on critical
times by using specific cancer types and stress paradigms,
optimizing the understanding of the effects of stress on cancer
cells (11). For instance,
stressing animals before and after tumour cell injection has been
shown to exacerbate the unfavourable effects on the ability of
marginating pulmonary natural killer (NK) cells to prevent
experimental lung metastasis (89,97,98). On the contrary, chronic stressors
do not affect initial breast cancer tumour formation in animal
models, but accelerate dissemination and metastasis. Social
isolation prior to inoculation does not affect primary tumour
development but, after already-palpable tumours were present, it
accelerates their growth (99-101). Of note, some of the
aforementioned key oncogenic pathophysiological phases may not be
identified in a clinical setting, whereas others, particularly
those associated with cancer therapeutic interventions, are known
to influence cancer progression and may be exploited to reduce the
effects of stress on cancer growth (11).
The sections that follow concentrate on various
hallmarks and stages of oncogenesis and cancer development, which
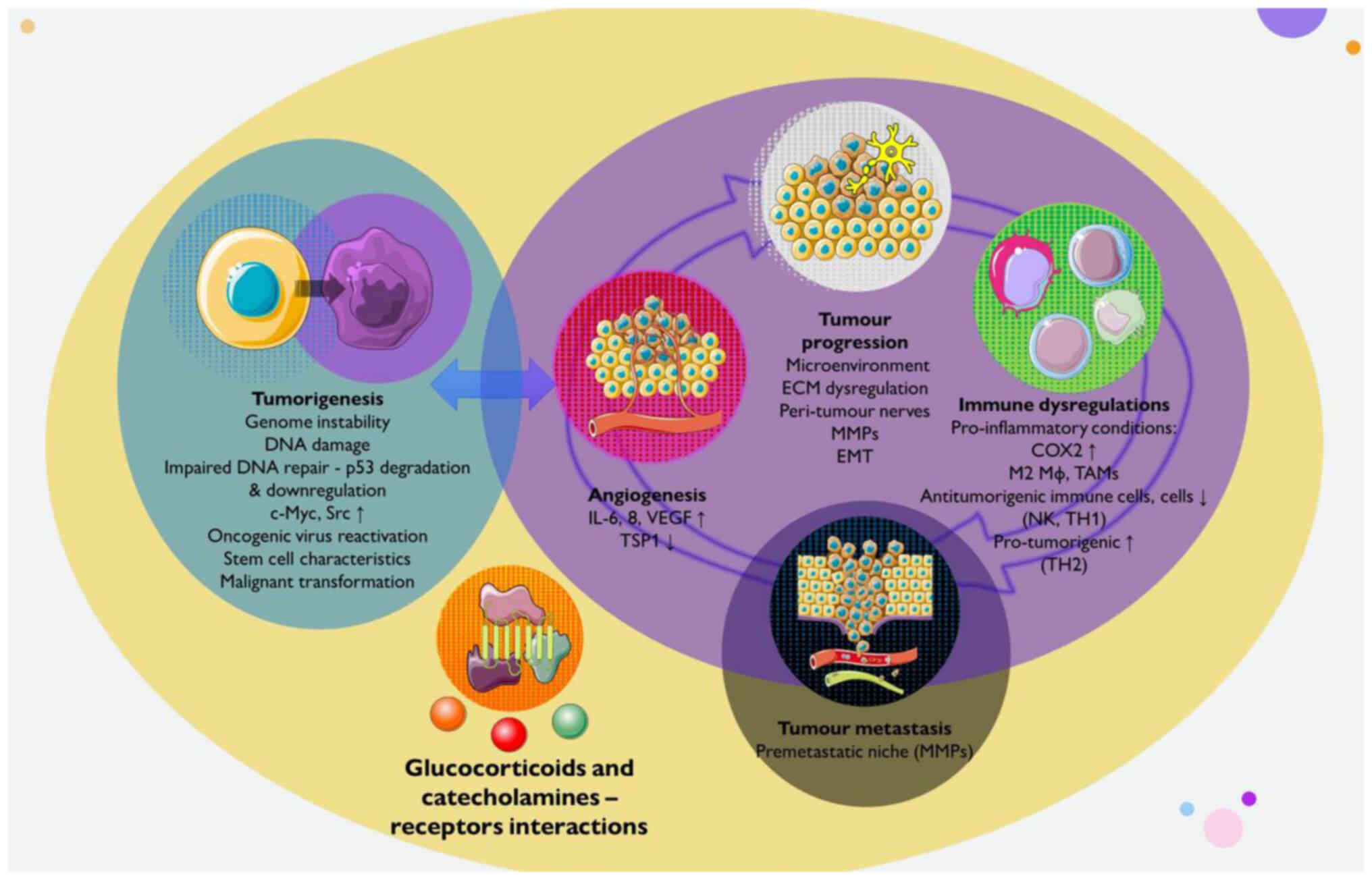
may be affected by stress. A summary of the effects of stress on
cancer initiation, progression, metastasis and selected underlying
mechanisms is illustrated in Fig.
2.
Stress-cancer interactions have been studied via
various strategies, including exposure to stressors in laboratory
animals and retrospective clinical studies in humans (12). A considerable body of evidence
suggests that stress can aggravate the majority of hallmarks of
cancer, including genome instability, tumour-promoting
inflammation, immune resistance, proliferative signalling,
resistance to cell death, angiogenesis, and invasion and metastasis
(11,12,80). Stress hormones, their receptor
systems and intracellular molecular pathways of adrenergic
receptors (ARs) and glucocorticoid receptors (GCRs) have been shown
to mediate these effects (102-105). Stress may contribute to cancer
initiation, progression and the development of metastasis (12). Herein, tumour initiation is
defined and referred to as the transition of non-malignant to
malignant tissue, as opposed to tumour progression, which arises
after this transformation, despite the fact that the majority of
cancer hallmarks may affect both the onset and progress of the
disease (11). Recently, it was
also shown that tumour-derived cytokines, immune mediators and
neurotransmitters can potentially control neuroendocrine centres
and modulating body homeostasis (106). These findings suggest a
bidirectional communication between local autonomic and sensory
nerves, and the tumour may affect the brain and HPA and other axes
(106).
Preclinical data published over the years have
highlighted the potential role of mediators of the neuroendocrine
stress response (particularly norepinephrine, epinephrine and
cortisol) in processes related to carcinogenesis which act directly
on cancer cells and promote tumour growth (5,12,102,111-113). Inflammation, angiogenesis,
genomic instability, metastasis and the expression of stem
cell-like genes are all facilitated by the binding of stress
hormones to their receptors (5,102,111). This occurs through the
epigenetic alteration or the activation of a variety of mechanisms.
Moreover, chronic stress-induced tumour cells become resistant to
apoptosis and cancer therapy (5,103).
Cancer is primarily caused by random mutations in
non-cancerous stem cells during DNA replication, with environmental
factors and inherited predispositions accounting for only a third
of variation among tissues (74,76,85,114-116). Notably, carcinogenic infections
account for 13-15% of human cancers, and stress may increase the
risk of developing cancer by promoting the spread or reactivation
of latent oncogenic viruses (68,115,117-122). Stress hormones can reactivate
oncogenic viruses, such as human papilloma virus, Epstein-Barr
virus, Kaposi sarcoma-associated herpesvirus, and hepatitis B and C
viruses, stimulate oncogenes in infected cells, reduce interferon
production and impair antiviral immunity (123-126). Psychosocial stress is an
environmental factor that contributes to cancer induction by
increasing somatic DNA mutation frequency and sensitizing cells to
environmental carcinogens (127). Research has demonstrated that
exposure to swim stress, noise, or foot shock increases chromosomal
aberrations and sister chromatid exchanges in bone marrow cells
(128). In healthy female
workers, poor stress-coping behaviours have been shown to result in
increased levels of cancer-related oxidative DNA damage in
leukocytes, and these biomarkers of DNA damage are associated with
average working hours, self-blame coping strategies and recent
family loss (128). Highly
distressed individuals have a reduced DNA repair in lymphocytes
exposed to γ-irradiation, while chronically stressed individuals
are more sensitive to DNA damage induction by external factors,
such as H2O2 and γ-radiation (129,130). These findings suggest that
psychological stress may increase susceptibility to environmental
mutagenic agents (130).
Studies using mechanistic approaches have been
carried out in order to obtain an understanding of the processes
and pathways linking stress and mutation, including AR and GCR
signalling pathways and the ATR-p21 pathway (11,12). It has been demonstrated that some
stressors cause DNA damage and hinder DNA repair, which may favour
malignant transformation (11).
Specifically, it has been shown that serum from stressed animals or
adrenaline, noradrenaline and cortisol (each component alone and
synergistically when combined) enhances DNA damage and/or inhibits
DNA repair following UV irradiation (131). In non-cancerous murine and human
cell lines, adrenaline receptor AR-mediated reactive oxygen species
production and arrestin-MDM2-dependent p53 degradation aggravated
DNA damage and hindered DNA repair (132).
Chronic stress induces these two AR-mediated
processes, and glucocorticoid-mediated responses can also boost
MDM2-dependent p53 downregulation and increase apoptosis resistance
in response to ionising radiation (133,134). In mice expressing c-Myc, a
proto-oncogene, the HPA axis was investigated in cancer induction
(135,136). Mice developed intraepithelial
prostate neoplasia (PIN) and this progressed to invasive
adenocarcinoma (135). Chemical
sympathectomy, administered at a young age, reduced the incidence
of PIN by 83%. However, the effect of sympathectomy was not
observed in adult individuals, as the PIN had already developed in
adult mice (135). β2- and
β3-ARs play a primary role in prostate adenocarcinoma development,
and the deletion of genes for these receptors leads to a
significant reduction in the incidence of adenocarcinoma in
genetically modified mice (135).
Norepinephrine, epinephrine and cortisol increase
the formation of oxygen radicals, causing DNA damage and reducing
cell repair processes (12). It
has been shown that the short-term exposure to physiological
concentrations of these substances induces at least a 5-fold
increase in DNA damage in treated murine 3T3 cells (113,131,132). Pre-treatment with an antagonist
of ARs or GCRs protected these cells from DNA damage (131). Norepinephrine and cortisol
reduced DNA repair in 3T3 cells exposed to UV radiation (131). The infusion of isoproterenol, an
β-AR agonist, induces the degradation of the tumour suppressor
protein, p53, in mouse thymus cells, while propranolol increases
the gene expression of p53 in primary melanoma-derived and
metastasis-derived tumours in mice (132,137). Chronic restraint stress
attenuates p53 functions and promotes ionizing radiation-induced
tumorigenesis in p53+/− mice, with glucocorticoids
playing a central role in these processes (134). β-adrenergic signalling also
participates in the activation of oncogenes, with the stimulation
of ovarian cancer cells with norepinephrine significantly activates
oncogene Src (138). A similar
effect of adrenergic signalling on the gene expression of Her2 has
also been described (139).
Rather than focusing on intermediate signs, such as
DNA damage or the reactivation of carcinogenic viruses, several
in vivo animal investigations on the influence of stress on
carcinogenesis have been conducted (11). Restraint stress, social isolation
and a cold ambient temperature, all lead to carcinogen-induced
cancer development (134,140-142).
Repetitive restraint stress enhances pancreatic carcinogenesis in
transgenic models of spontaneous cancer via AR signalling (143), although sympathetic denervation
results in a reduction in carcinogenesis in models of prostate
cancer (135). Models based on
accelerated cancer induction struggle to differentiate the effects
of stress on tumour initiation and development due to the overlap
between the time course of stress and the initiation and
progression periods (134,143-145).
The neuroendocrine system modifies the TME in a
manner that favours cancer progression (5,111). The TME is composed of immune
cells and other stromal cells, such as fibroblasts, adipocytes and
endothelial cells, in addition to extracellular components
(extracellular matrix) (5,147).
These components pertain to the circulatory system, lymphatic
system and peripheral nerves, and they support cancer cells
(148). Chronic stress-related
hormones influence tumour development, including interactions
between cancer cells, and invading immune and stromal cell
populations in the TME; recent research indicates that tumours can
attract nerves into the TME and form peri-tumour nerves, which
influence carcinogenesis, angiogenesis, invasion and metastasis
(5,149,150).
Pro-inflammatory molecules released by immune cells
can lead to mutagenic processes, transforming normal cells into
cancer cells (164). Stress can
affect inflammation-related mutagenic processes by activating the
sympathetic nervous system (165). A previous study on diethyl
nitrosamine-induced hepatocarcinogenesis in rats demonstrated that
sympathectomy with 6-hydroxydopamine reduced the development of
hepatocellular carcinoma (165).
However, prazosin, an α-AR antagonist, led to carcinoma development
in up to 64% of rats. Sympathetic innervation is crucial for
maintaining the liver inflammatory microenvironment, which
initiates hepatocellular carcinoma development (165). Stress-induced-adrenergic
signalling increases inflammation in malignant cells and
tumour-associated macrophages (TAMs), leading to an increased
expression of cyclooxygenase 2, prostaglandin secretion and
increase levels of pro-inflammatory cytokines such as IL-6, which
in turn increases macrophage recruitment and
M2-pro-inflammatory-polarization (99,151,153,161,166-169). Social isolation and depression
are linked to an increased M2 polarization in breast tumours, while
higher levels of depression and an increased expression of genes
encoding AR and prostaglandins predict a decreased survival rate of
cancer patients (166,170).
According to animal and human research, stress
affects numerous anti- and pro-cancer immune system components
(12). Chronic stress in humans
has been shown to suppress host resistance to metastasis, causing
lung tumour retention and metastasis (98). An acute stressor, such as swimming
stress, suppresses NK cell activity and increases lung tumour
retention (98). This effect can
be attenuated or prevented by reducing the release of
norepinephrine and epinephrine or using β1- and β2-blockers
(98). However, the
administration of epinephrine or other agonists of β-AR can also
suppress NK cell activity and increase lung tumour retention
(17). This suggests that acute
stress suppresses NK cell activity and compromises resistance to
NK-sensitive metastasis (17,98).
Stress-induced-adrenergic signalling or agonists
can inhibit NK cell activity against tumour cells, leading to
increased lung metastases (17,89,97,98,171,172). A lower NK cytotoxicity in
patients with ovarian cancer is linked to less social support and
depression (173). Stress
induces a shift from T-helper 1 cell (TH1) to T-helper 2 cell (TH2)
cytokine production, increasing tumour growth in colorectal and
squamous cell carcinoma mouse models (140,174,175). A depressive and worried mood are
related to a decreased TH1 cell/TH2 cell-type cytokine ratio in
patients with ovarian cancer (176). The stress-induced-adrenergic
response promotes tumour growth by upregulating suppressive immune
cells, such as myeloid-derived suppressor cells (MDSCs) and
regulatory T-cells (140,158,163,175,177).
Higher levels of stress are associated with a decrease in the
number of circulating MDSCs (178). Additionally, chronic sound
stress has been shown to increase colon cancer progression, plasma
norepinephrine and corticosterone levels, and to induce a shift in
the TH1 to TH2 response (158,174). Similar effects of stress on
immunological functions have been observed in clinical
investigations, including a reduction in protective immunity, the
exacerbation of chronic inflammation, and the enhancement of
immunosuppressive processes (15).
Stress hormones, which can be generated
systemically or even secreted locally in the TME by infiltrating
sympathetic nerve endings, immune cells, or tumour cells per se can
have direct effects on cancer cells and boosting their malignant
properties (139,179-182). Both noradrenaline and adrenaline
enhance cancer cell proliferation, as well as survival (through
anti-apoptosis), cell migration and invasion,
epithelial-mesenchymal transition (EMT), the production of
prostaglandins and the activation of MMPs (100,152,166,183-190). In animal models, psychological
or physiological stressors (such as social confrontation, restraint
and surgery) have been shown to enhance tumour development and
metastasis by activating tumour the AR, as demonstrated by
pharmacological or molecular blockage, or genetic knockout
(18,99,175,183,188,191-193).
A recent study demonstrated that tumour innervation
led to the advancement of cancer (194). Tumours can generate neural
growth agents that promote sympathetic tumour innervation. During
stress-induced sympathetic activation, higher tumour noradrenaline
levels establish a feedforward loop that promotes cancer
progression (143). Multiple
cancer forms express AR, and higher levels of tumour noradrenaline
and/or plasma adrenaline are associated with larger tumour size,
advanced stage, lymph node metastases and/or a shorter survival
(143,152,166,167,183,184,187,189,190,195).
Chronic behavioural stress can increase tissue
catecholamine levels and promote the growth and invasiveness of
ovarian carcinoma cells, primarily through the activation of β2-AR
(18). The effect of stress on
cancer cell resistance to death supports previous findings, such as
the inhibition of the apoptosis of prostate and breast cancer cells
by epinephrine, chemical sympathectomy increasing the gene
expression of apoptotic factors in mouse melanoma tumours, and
selective antagonists of β1- and β2-AR on colorectal cancer cell
growth (16,201,202). Psychologic stress can inhibit
apoptosis, but this effect is prevented by administration of a
selective β2-AR antagonist (16).
Numerous of the aforementioned and other
stress-induced processes contribute to metastasis, in addition to
promoting initiation and progression. Stress significantly
increases the development of metastases, as demonstrated by studies
on colon carcinoma, nasopharyngeal carcinoma, and ovarian cancer
cells (203-205). As previously demonstrated,
norepinephrine treatment increases the locomotor activity of colon
carcinoma cells; however, this effect is blocked by propranolol
(203). MMP-2 and MMP-9 levels
have also been shown to be increased in the cell culture
supernatant, whereas this effect is blocked by propranolol
(206). Other examples include
studies in mice, demonstrating that stress-induced AR activation
promotes circulating tumour cell migration to the bones via the
increased expression of RANKL by bone marrow-derived stem cells or
to the lungs via the CC-chemokine ligand 2-CC-chemokine receptor
2-mediated attraction of macrophages, thereby forming
pre-metastatic niches and increasing organ-specific metastasis
(191,207). Additionally, stress increases
tumour and stromal cell MMP production, tumour cell anoikis
resistance, and cancer cell EMT, thus boosting malignant cell
detachment, invasiveness and circulation survival (99,100,157,183, 186,188-190,208).
Perceived stress, depressive symptoms, or social
isolation have been shown to be associated with the increased
expression of EMT-related genes in tumours in patients with breast
and ovarian cancer, as well as higher levels of MMP-9 in tumour
cells and/or TAMs (99,206). In numerous mouse models, AR
inhibition significantly reduces experimental and spontaneous
metastases (89,99,161,183,191,207,209). Similar to how tumour AR
expression levels have been associated with lymph node metastasis
in patients with gastric and lung cancer, incidental AR-blocker use
has been linked to a lower risk of developing metastasis or
recurrence in patients with breast and ovarian cancer, as well as
to the improved survival of patients with melanoma and breast
cancer, but not lung and ovarian cancer (161,183,210-216).
It has been demonstrated that chronic stress in
cancer patients with elevated depressive symptoms and low social
support leads to a 30-fold increase in metastasis to distant
tissues (99). Stress-induced
lymphatic vessel rearrangement may also contribute to cancer cell
dissemination (161). β2-AR
activation reduces deformability in metastatic human breast cancer
cells, rendering them more invasive (217). Randomized controlled trials are
required to evaluate the effects of AR-blockers on long-term cancer
outcomes due to the discrepancies in the analysed indices (such as
metastasis vs. survival), the diversity of cancer types and the
uncontrolled settings of correlational research (10).
The present review indicates that stress has been
linked to cancer development and incidence for a number of decades.
Psychological stressors have been linked to cancer development,
with the immune system playing a critical role in inhibiting cancer
progression. Recent research has advanced the current understanding
of the role of stress in cancer induction, growth and metastasis
development, with the SNS and HPA axis playing crucial roles.
Animal studies have revealed a clearer link than clinical human
studies, suggesting that stress factors can exacerbate cancer
hallmarks and promote growth and metastasis by directly affecting
malignant tissue, its microenvironment, antitumor immune activity
and other indirect cancer progression modifiers. Stress-related
hormones can influence tumour development, migration, invasion, and
cancer cell proliferation. The therapeutic potential of these
pathophysiological mechanisms is shown by the discovery of numerous
procedures that are triggered by stress in patients with cancer;
however, these are beyond the scope of this review and can be
further investigated in the future. Randomized controlled trials
are required to evaluate the effects of stress on long-term cancer
outcomes. Psychological therapies can potentially target stress and
benefit individuals. To minimize cancer recurrence and associated
mortality, chronic stress-management interventions need to be
tested during critical periods, accompanied by pharmacological
approaches, and include individualized modules.
Not applicable.
DAS and VEG conceptualized the study. IGL, VEG, PP,
GPC and DAS made a substantial contribution to the interpretation
and analysis of data to be included in the review, and wrote and
prepared the draft of the manuscript. DAS and GPC analysed the data
from studies for inclusion in the review and provided critical
revisions. All authors contributed to manuscript revision, and have
read and approved the final version of the manuscript. Data
authentication is not applicable.
Not applicable.
Not applicable.
DAS is the Editor-in-Chief for the journal, but had
no personal involvement in the reviewing process, or any influence
in terms of adjudicating on the final decision, for this article.
The other authors declare that they have no competing
interests.
Not applicable.
No funding was received.
|
1
|
Chrousos GP: Stress and disorders of the
stress system. Nat Rev Endocrinol. 5:374–381. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Agorastos A and Chrousos GP: The
neuroendocrinology of stress: The stress-related continuum of
chronic disease development. Mol Psychiatry. 27:502–513. 2022.
View Article : Google Scholar
|
|
3
|
Tsigos C, Kyrou I, Kassi E and Chrousos
GP: Stress: Endocrine Physiology and Pathophysiology. Endotext.
Feingold KR, Anawalt B, Blackman MR, Boyce A, Chrousos G, Corpas E,
de Herder WW, Dhatariya K, Dungan K, Hofland J, et al: MDTextcom,
Inc; South Dartmouth, MA: 2000
|
|
4
|
Chrousos GP: The glucocorticoid receptor
gene, longevity, and the complex disorders of Western societies. Am
J Med. 117:204–207. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yan J, Chen Y, Luo M, Hu X, Li H, Liu Q
and Zou Z: Chronic stress in solid tumor development: From
mechanisms to interventions. J Biomed Sci. 30:82023. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lempesis IG, Karlafti E, Papalexis P,
Fotakopoulos G, Tarantinos K, Lekakis V, Papadakos SP, Cholongitas
E and Georgakopoulou VE: COVID-19 and liver injury in individuals
with obesity. World J Gastroenterol. 29:908–916. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Georgakopoulou VE, Lembessis P, Skarlis C,
Gkoufa A, Sipsas NV and Mavragani CP: Hematological abnormalities
in COVID-19 disease: Association with type I interferon pathway
activation and disease outcomes. Front Med (Lausanne).
9:8504722022. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tsamakis K, Gavriatopoulou M, Schizas D,
Stravodimou A, Mougkou A, Tsiptsios D, Sioulas V, Spartalis E,
Sioulas AD, Tsamakis C, et al: Oncology during the COVID-19
pandemic: Challenges, dilemmas and the psychosocial impact on
cancer patients. Oncol Lett. 20:441–447. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Georgakopoulou VE, Makrodimitri S,
Triantafyllou M, Samara S, Voutsinas PM, Anastasopoulou A,
Papageorgiou CV, Spandidos DA, Gkoufa A, Papalexis P, et al:
Immature granulocytes: Innovative biomarker for SARS-CoV-2
infection. Mol Med Rep. 26:2172022. View Article : Google Scholar :
|
|
10
|
Lempesis IG and Georgakopoulou VE:
Implications of obesity and adiposopathy on respiratory infections;
focus on emerging challenges. World J Clin Cases. 11:29252023.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Eckerling A, Ricon-Becker I, Sorski L,
Sandbank E and Ben-Eliyahu S: Stress and cancer: Mechanisms,
significance and future directions. Nat Rev Cancer. 21:767–785.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mravec B, Tibensky M and Horvathova L:
Stress and cancer. Part I: Mechanisms mediating the effect of
stressors on cancer. J Neuroimmunol. 346:5773112020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Deshaies-Gendron M: Enquiries Into the
Nature, Knowledge, and Cure of Cancers. Mr Gendron Deshaies : Done
out of French. Gale Ecco, Print Editions. April 22–2018.
|
|
14
|
Selye H: A syndrome produced by diverse
nocuous agents. Nature. 138:32. 1936. View Article : Google Scholar
|
|
15
|
Antoni MH and Dhabhar FS: The impact of
psychosocial stress and stress management on immune responses in
patients with cancer. Cancer. 125:1417–1431. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hassan S, Karpova Y, Baiz D, Yancey D,
Pullikuth A, Flores A, Register T, Cline JM, D'Agostino R Jr,
Danial N, et al: Behavioral stress accelerates prostate cancer
development in mice. J Clin Invest. 123:874–886. 2013.PubMed/NCBI
|
|
17
|
Inbar S, Neeman E, Avraham R, Benish M,
Rosenne E and Ben-Eliyahu S: Do stress responses promote leukemia
progression? An animal study suggesting a role for epinephrine and
prostaglandin-E2 through reduced NK activity. PLoS One.
6:e192462011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Thaker PH, Han LY, Kamat AA, Arevalo JM,
Takahashi R, Lu C, Jennings NB, Armaiz-Pena G, Bankson JA, Ravoori
M, et al: Chronic stress promotes tumor growth and angiogenesis in
a mouse model of ovarian carcinoma. Nat Med. 12:939–944. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wu Y, Zhou L, Zhang X, Yang X, Niedermann
G and Xue J: Psychological distress and eustress in cancer and
cancer treatment: Advances and perspectives. Sci Adv.
8:eabq79822022. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Heikkilä K, Nyberg ST, Theorell T,
Fransson EI, Alfredsson L, Bjorner JB, Bonenfant S, Borritz M,
Bouillon K, Burr H, et al: Work stress and risk of cancer:
Meta-analysis of 5700 incident cancer events in 116 000 European
men and women. BMJ. 346:f1652013. View Article : Google Scholar
|
|
21
|
Yang T, Qiao Y, Xiang S, Li W, Gan Y and
Chen Y: Work stress and the risk of cancer: A meta-analysis of
observational studies. Int J Cancer. 144:2390–2400. 2019.
View Article : Google Scholar
|
|
22
|
Chida Y, Hamer M, Wardle J and Steptoe A:
Do stress-related psychosocial factors contribute to cancer
incidence and survival? Nat Clin Pract Oncol. 5:466–475. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Coyne JC, Ranchor AV and Palmer SC:
Meta-analysis of stress-related factors in cancer. Nat Rev Clin
Oncol. 72010. View Article : Google Scholar
|
|
24
|
Mravec B and Tibensky M: Increased cancer
incidence in 'cold' countries: An (un) sympathetic connection? J
Therm Biol. 89:1025382020. View Article : Google Scholar
|
|
25
|
Keinan-Boker L, Vin-Raviv N, Liphshitz I,
Linn S and Barchana M: Cancer incidence in Israeli jewish survivors
of World War II. J Natl Cancer Inst. 101:1489–1500. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Huang T, Poole EM, Okereke OI, Kubzansky
LD, Eliassen AH, Sood AK, Wang M and Tworoger SS: Depression and
risk of epithelial ovarian cancer: Results from two large
prospective cohort studies. Gynecol Oncol. 139:481–486. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fang F, Fall K, Sparén P, Adami HO,
Valdimarsdóttir HB, Lambe M and Valdimarsdóttir U: Risk of
infection-related cancers after the loss of a child: A follow-up
study in Sweden. Cancer Res. 71:116–122. 2011. View Article : Google Scholar
|
|
28
|
Schoemaker MJ, Jones ME, Wright LB,
Griffin J, McFadden E, Ashworth A and Swerdlow AJ: Psychological
stress, adverse life events and breast cancer incidence: A cohort
investigation in 106,000 Women in the United Kingdom. Breast Cancer
Res. 18:722016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Trudel-Fitzgerald C, Poole EM, Idahl A,
Lundin E, Sood AK, Kawachi I, Kubzansky LD and Tworoger SS: The
association of work characteristics with ovarian cancer risk and
mortality. Psychosom Med. 79:1059–1067. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Carlson L, Angen M, Cullum J, Goodey E,
Koopmans J, Lamont L, MacRae JH, Martin M, Pelletier G, Robinson J,
et al: High levels of untreated distress and fatigue in cancer
patients. Br J Cancer. 90:2297–2304. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mitchell AJ, Chan M, Bhatti H, Halton M,
Grassi L, Johansen C and Meader N: Prevalence of depression,
anxiety, and adjustment disorder in oncological, haematological,
and palliative-care settings: A meta-analysis of 94 interview-based
studies. Lancet Oncol. 12:160–174. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lu D, Andersson TM, Fall K, Hultman CM,
Czene K, Valdimarsdóttir U and Fang F: Clinical diagnosis of mental
disorders immediately before and after cancer diagnosis: A
nationwide matched cohort study in Sweden. JAMA Oncol. 2:1188–1196.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Batty GD, Russ TC, Stamatakis E and
Kivimäki M: Psychological distress in relation to site specific
cancer mortality: Pooling of unpublished data from 16 prospective
cohort studies. BMJ. 356:1082017. View Article : Google Scholar
|
|
34
|
Wang X, Wang N, Zhong L, Wang S, Zheng Y,
Yang B, Zhang J, Lin Y and Wang Z: Prognostic value of depression
and anxiety on breast cancer recurrence and mortality: A systematic
review and meta-analysis of 282,203 patients. Mol Psychiatry.
25:3186–3197. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pinquart M and Duberstein P: Depression
and cancer mortality: A meta-analysis. Psychol Med. 40:1797–1810.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Pinquart M and Duberstein PR: Associations
of social networks with cancer mortality: A meta-analysis. Crit Rev
Oncol Hematol. 75:122–137. 2010. View Article : Google Scholar :
|
|
37
|
Manjili MH: Tumor dormancy and relapse:
From a natural byproduct of evolution to a disease state. Cancer
Res. 77:2564–2569. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cannon WB: The wisdom of the body. 2nd.
Norton and Co; New York, NY: 1939
|
|
39
|
Cannon WB: Bodily changes in pain, hunger,
fear, and rage An account of recent researches into the function of
emotional excitement. D. Appleton and Company; 1915, View Article : Google Scholar
|
|
40
|
Selye H: The stress of life. McGraw-Hill
Education; New York, NY: 1956
|
|
41
|
Chrousos GP: Hans Selye memorial lecture:
Stressors, stress and neuroendocrine integration of the adaptive
response. Ann NY Acad Sci. 851:311–335. 1997. View Article : Google Scholar
|
|
42
|
Lim CT and Khoo B: Normal Physiology of
ACTH and GH Release in the Hypothalamus and Anterior Pituitary in
Man. Endotext. Feingold KR, Anawalt B, Blackman MR, Boyce A,
Chrousos G, Corpas E, de Herder WW, Dhatariya K, Dungan K, Hofland
J, et al: MDTextcom, Inc; South Dartmouth, MA: 2000
|
|
43
|
Gassen NC, Chrousos GP, Binder EB and
Zannas AS: Life stress, glucocorticoid signaling, and the aging
epigenome: Implications for aging-related diseases. Neurosci
Biobehav Rev. 74:356–365. 2017. View Article : Google Scholar
|
|
44
|
Nicolaides NC, Chrousos G and Kino T:
Glucocorticoid Receptor. Endotext. Feingold KR, Anawalt B, Blackman
MR, Boyce A, Chrousos G, Corpas E, de Herder WW, Dhatariya K,
Dungan K, Hofland J, et al: MDText.com, Inc.; South Dartmouth MA:
2000
|
|
45
|
Chrousos GP, McCarty R, Pacak K, Cizza G,
Sternberg G, Gold P and Květňanský R: Stress: Basic mechanisms and
clinical implications. 1995.
|
|
46
|
Chrousos GP and Gold PW: The concepts of
stress and stress system disorders: Overview of physical and
behavioral homeostasis. JAMA. 267:1244–1252. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Chrousos GP: Organization and integration
of the endocrine system: The arousal and sleep perspective. Sleep
Med Clin. 2:125–145. 2007. View Article : Google Scholar
|
|
48
|
Charmandari E, Tsigos C and Chrousos G:
Neuroendocrinology of stress. Ann Rev Physiol. 67:259–284. 2005.
View Article : Google Scholar
|
|
49
|
Gold PW, Goodwin FK and Chrousos GP:
Clinical and biochemical manifestations of depression. N Engl J
Med. 319:413–420. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Chrousos GP and Kino T: Glucocorticoid
action networks and complex psychiatric and/or somatic disorders.
Stress. 10:213–219. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Franchimont D, Kino T, Galon J, Meduri GU
and Chrousos G: Glucocorticoids and Inflammation Revisited: The
State of the ArtNIH clinical staff conference.
Neuroimmunomodulation. 10:247–260. 2003. View Article : Google Scholar
|
|
52
|
Chrousos GP: The stress response and
immune function: Clinical implications. Ann NY Acad Sci. 917:38–67.
2000. View Article : Google Scholar
|
|
53
|
Karalis K, Sano H, Redwine J, Listwak S,
Wilder RL and Chrousos GP: Autocrine or paracrine inflammatory
actions of corticotropin-releasing hormone in vivo. Science.
254:421–423. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Sapolsky RM: Stress and the brain:
Individual variability and the inverted-U. Nat Neurosci.
18:1344–1346. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
McEwen BS: Neurobiological and systemic
effects of chronic stress. Chronic Stress (Thousand Oaks).
1:24705470176923282017.PubMed/NCBI
|
|
56
|
McEwen BS and Gianaros PJ: Central role of
the brain in stress and adaptation: Links to socioeconomic status,
health, and disease. Ann N Y Acad Sci. 1186:190–222. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
McEwen BS and Stellar E: Stress and the
individual: Mechanisms leading to disease. Arch Intern Med.
153:2093–2101. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Stefanaki C, Pervanidou P, Boschiero D and
Chrousos GP: Chronic stress and body composition disorders:
Implications for health and disease. Hormones (Athens). 17:33–43.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Chrousos GP and Tsigos C: Stress, obesity,
and metabolic syndrome. Blackwell Pub on behalf of the New York
Academy of Sciences; 2006
|
|
60
|
Chrousos G: The role of stress and the
hypothalamic-pituitary-adrenal axis in the pathogenesis of the
metabolic syndrome: Neuro-endocrine and target tissue-related
causes. Int J Obes Relat Metab Disord. 24(Suppl 2): S50–S55. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Lempesis IG, Hoebers N, Essers Y, Jocken
JWE, Dineen R, Blaak EE, Manolopoulos KN and Goossens GH: Distinct
inflammatory signatures of upper and lower body adipose tissue and
adipocytes in women with normal weight or obesity. Front Endocrinol
(Lausanne). 14:16002023. View Article : Google Scholar
|
|
62
|
Lempesis IG, Tsilingiris D, Liu J and
Dalamaga M: Of mice and men: Considerations on adipose tissue
physiology in animal models of obesity and human studies. Metabol
Open. 15:1002082022. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Lempesis IG and Georgakopoulou VE:
Physiopathological mechanisms related to inflammation in obesity
and type 2 diabetes mellitus. World J Exp Med. 13:7–16. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Lempesis IG, Varrias D, Sagris M, Attaran
RR, Altin ES, Bakoyiannis C, Palaiodimos L, Dalamaga M and
Kokkinidis DG: Obesity and peripheral artery disease: Current
evidence and controversies. Curr Obes Rep. May 27–2023.Epub ahead
of print. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Lempesis IG, van Meijel RLJ, Manolopoulos
KN and Goossens GH: Oxygenation of adipose tissue: A human
perspective. Acta Physiol (Oxf). 228:e132982020. View Article : Google Scholar
|
|
66
|
Spandidos DA: A unified theory for the
development of cancer. Biosci Rep. 6:691–708. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Spandidos DA and Anderson ML: Oncogenes
and onco-suppressor genes: Their involvement in cancer. J Pathol.
157:1–10. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Simatou A, Simatos G, Goulielmaki M,
Spandidos DA, Baliou S and Zoumpourlis V: Historical retrospective
of the SRC oncogene and new perspectives (Review). Mol Clin Oncol.
13:212020.PubMed/NCBI
|
|
69
|
Nitulescu GM, Van De Venter M, Nitulescu
G, Ungurianu A, Juzenas P, Peng Q, Olaru OT, Grădinaru D, Tsatsakis
A, Tsoukalas D, et al: The Akt pathway in oncology therapy and
beyond (Review). Int J Oncol. 53:2319–2331. 2018.PubMed/NCBI
|
|
70
|
Fotakopoulos G, Georgakopoulou VE,
Spandidos DA, Papalexis P, Angelopoulou E, Aravantinou-Fatorou A,
Trakas N, Trakas I and Brotis AG: Role of miR-200 family in brain
metastases: A systematic review. Mol Clin Oncol. 18:152023.
View Article : Google Scholar
|
|
71
|
Damaskos C, Garmpis N, Dimitroulis D,
Garmpi A, Diamantis E, Sarantis P, Georgakopoulou VE, Patsouras A,
Despotidis M, Prevezanos D, et al: The Role of SNHG15 in the
pathogenesis of hepatocellular carcinoma. J Pers Med. 12:7532022.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Garmpis N, Damaskos C, Garmpi A, Nonni A,
Georgakopoulou VE, Antoniou E, Schizas D, Sarantis P, Patsouras A,
Syllaios A, et al: Histone deacetylases and their inhibitors in
colorectal cancer therapy: Current evidence and future
considerations. Curr Med Chem. 29:2979–2994. 2022. View Article : Google Scholar
|
|
73
|
Garmpis N, Damaskos C, Angelou A, Garmpi
A, Georgakopoulou VE, Valsami S, Schizas D, Voutyritsa E, Syllaios
A, Diamantis E, et al: Animal models for the calculation of
circulating tumor cells for experimental demonstration. Anticancer
Res. 40:6599–6607. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Tsatsakis A, Oikonomopoulou T,
Nikolouzakis TK, Vakonaki E, Tzatzarakis M, Flamourakis M, Renieri
E, Fragkiadaki P, Iliaki E, Bachlitzanaki M, et al: Role of
telomere length in human carcinogenesis (Review). Int J Oncol.
63:782023. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Anderson ML and Spandidos DA:
Onco-suppressor genes and their involvement in cancer (review).
Anticancer Res. 8:873–879. 1988.PubMed/NCBI
|
|
76
|
Lang JC and Spandidos DA: The structure
and function of eukaryotic enhancer elements and their role in
oncogenesis. Anticancer Res. 6:437–449. 1986.PubMed/NCBI
|
|
77
|
Field JK and Spandidos DA: Expression of
oncogenes in human tumours with special reference to the head and
neck region. J Oral Pathol. 16:97–107. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Spandidos DA and Anderson ML: A study of
mechanisms of carcinogenesis by gene transfer of oncogenes into
mammalian cells. Mutat Res. 185:271–291. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Fouad YA and Aanei C: Revisiting the
hallmarks of cancer. Am J Cancer Res. 7:10162017.PubMed/NCBI
|
|
82
|
Spandidos DA and Anderson ML: A role of
ras oncogenes in carcinogenesis and differentiation. Adv Exp Med
Biol. 265:127–131. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Gonos ES and Spandidos DA: Oncogenes in
cellular immortalisation and differentiation (review). Anticancer
Res. 13:1117–1122. 1993.PubMed/NCBI
|
|
84
|
Neagu M, Constantin C, Popescu ID, Zipeto
D, Tzanakakis G, Nikitovic D, Fenga C, Stratakis CA, Spandidos DA
and Tsatsakis AM: Inflammation and metabolism in cancer
cell-mitochondria key player. Front Oncol. 9:3482019. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Pierouli K, Papakonstantinou E,
Papageorgiou L, Diakou I, Mitsis T, Dragoumani K, Spandidos DA,
Bacopoulou F, Chrousos GP, Goulielmos GN, et al: Long non-coding
RNAs and microRNAs as regulators of stress in cancer (Review). Mol
Med Rep. 26:3612022. View Article : Google Scholar
|
|
86
|
Connerty P, Lock RB and De Bock CE: Long
non-coding RNAs: Major regulators of cell stress in cancer. Front
Oncol. 10:2852020. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Bergers G and Benjamin LE: Tumorigenesis
and the angiogenic switch. Nat Rev Cancer. 3:401–410. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Patidar A, Selvaraj S, Sarode A, Chauhan
P, Chattopadhyay D and Saha B: DAMP-TLR-cytokine axis dictates the
fate of tumor. Cytokine. 104:114–123. 2018. View Article : Google Scholar
|
|
89
|
Melamed R, Rosenne E, Shakhar K, Schwartz
Y, Abudarham N and Ben-Eliyahu S: Marginating pulmonary-NK activity
and resistance to experimental tumor metastasis: Suppression by
surgery and the prophylactic use of a beta-adrenergic antagonist
and a prostaglandin synthesis inhibitor. Brain Behav Immun.
19:114–126. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Melamed R, Rosenne E, Benish M, Goldfarb
Y, Levi B and Ben-Eliyahu S: The marginating-pulmonary immune
compartment in rats: Characteristics of continuous inflammation and
activated NK cells. J Immunother. 33:16–29. 2010. View Article : Google Scholar
|
|
91
|
Sorski L, Melamed R, Levi B, Matzner P,
Lavon H, Rosenne E, Shaashua L, Ricon I, Sandbank E, Benbenishty A
and Ben-Eliyahu S: Prevention of liver metastases through
perioperative acute CpG-C immune stimulation. Cancer Immunol
Immunother. 69:2021–2031. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Strilic B and Offermanns S: Intravascular
survival and extravasation of tumor cells. Cancer Cell. 32:282–293.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Gonzalez H, Hagerling C and Werb Z: Roles
of the immune system in cancer: From tumor initiation to metastatic
progression. Genes Dev. 32:1267–1284. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Neeman E and Ben-Eliyahu S: Surgery and
stress promote cancer metastasis: New outlooks on perioperative
mediating mechanisms and immune involvement. Brain Behav Immun.
30(Suppl): S32–S40. 2013. View Article : Google Scholar
|
|
95
|
Agorastos A, Nicolaides NC, Bozikas VP,
Chrousos GP and Pervanidou P: Multilevel interactions of stress and
circadian system: Implications for traumatic stress. Front
Psychiatry. 10:10032020. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Chrousos GP: The
hypothalamic-pituitary-adrenal axis and immune-mediated
inflammation. N Engl J Med. 332:1351–1363. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Rosenne E, Shakhar G, Melamed R, Schwartz
Y, Erdreich-Epstein A and Ben-Eliyahu S: Inducing a mode of
NK-resistance to suppression by stress and surgery: A potential
approach based on low dose of poly I-C to reduce postoperative
cancer metastasis. Brain Behav Immun. 21:395–408. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Ben-Eliyahu S, Shakhar G, Page GG,
Stefanski V and Shakhar K: Suppression of NK cell activity and of
resistance to metastasis by stress: A role for adrenal
catecholamines and β-adrenoceptors. Neuroimmunomodulation.
8:154–164. 2000. View Article : Google Scholar
|
|
99
|
Sloan EK, Priceman SJ, Cox BF, Yu S,
Pimentel MA, Tangkanangnukul V, Arevalo JM, Morizono K, Karanikolas
BD, Wu L, et al: The sympathetic nervous system induces a
metastatic switch in primary breast cancer. Cancer Res.
70:7042–7052. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Du P, Zeng H, Xiao Y, Zhao Y, Zheng B,
Deng Y, Liu J, Huang B, Zhang X, Yang K, et al: Chronic stress
promotes EMT-mediated metastasis through activation of STAT3
signaling pathway by miR-337-3p in breast cancer. Cell Death Dis.
11:7612020. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Madden KS, Szpunar MJ and Brown EB: Early
impact of social isolation and breast tumor progression in mice.
Brain Behav Immun. 30(Suppl): S135–S141. 2013. View Article : Google Scholar
|
|
102
|
Cole SW and Sood AK: Molecular pathways:
Beta-adrenergic signaling in cancer. Clin Cancer Res. 18:1201–1206.
2012. View Article : Google Scholar :
|
|
103
|
Eng JWL, Kokolus KM, Reed CB, Hylander BL,
Ma WW and Repasky EA: A nervous tumor microenvironment: The impact
of adrenergic stress on cancer cells, immunosuppression, and
immunotherapeutic response. Cancer Immunol Immunother.
63:1115–1128. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Cole SW, Nagaraja AS, Lutgendorf SK, Green
PA and Sood AK: Sympathetic nervous system regulation of the tumour
microenvironment. Nat Rev Cancer. 15:563–572. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Volden PA and Conzen SD: The influence of
glucocorticoid signaling on tumor progression. Brain Behav Immun.
30(Suppl): S26–S31. 2013. View Article : Google Scholar
|
|
106
|
Slominski RM, Raman C, Chen JY and
Slominski AT: How cancer hijacks the Body's homeostasis through the
neuroendocrine system. Trends Neurosci. 46:263–275. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Rosch PJ: Stress and cancer: Disorders of
communication, control, and civilization. Handbook of stress,
medicine, and health. Cooper CL: CRC Press/Routledge/Taylor and
Francis Group; Oxfordshire: pp. 27–60. 1996
|
|
108
|
Slawikowski GJ: Tumor development in
adrenalectomized rats given inoculations of aged tumor cells after
surgical stress. Cancer Res. 20:316–320. 1960.PubMed/NCBI
|
|
109
|
Helgesson Ö, Cabrera C, Lapidus L,
Bengtsson C and Lissner L: Self-reported stress levels predict
subsequent breast cancer in a cohort of Swedish women. Eur J Cancer
Prev. 377–381. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Nielsen NR, Zhang ZF, Kristensen TS,
Netterstr B, Schnohr P and Gr M: Self reported stress and risk of
breast cancer: Prospective cohort study. BMJ. 331:5482005.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Cui B, Peng F, Lu J, He B, Su Q, Luo H,
Deng Z, Jiang T, Su K, Huang Y, et al: Cancer and stress: NextGen
strategies. Brain Behav Immun. 93:368–383. 2021. View Article : Google Scholar
|
|
112
|
Yasuda MT, Sakakibara H and Shimoi K:
Estrogen-and stress-induced DNA damage in breast cancer and
chemoprevention with dietary flavonoid. Genes Environ. 39:102017.
View Article : Google Scholar
|
|
113
|
Jenkins FJ, Van Houten B and Bovbjerg DH:
Effects on DNA damage and/or repair processes as biological
mechanisms linking psychological stress to cancer risk. J Appl
Biobehav Res. 19:3–23. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Tomasetti C and Vogelstein B: Variation in
cancer risk among tissues can be explained by the number of stem
cell divisions. Science. 347:78–81. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Chalkia AK, Spandidos DA and Detorakis ET:
Viral involvement in the pathogenesis and clinical features of
ophthalmic pterygium. Int J Mol Med. 32:539–543. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Filippakis H, Spandidos DA and Sourvinos
G: Herpesviruses: Hijacking the Ras signaling pathway. Biochim
Biophys Acta. 1803:777–785. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Plummer M, de Martel C, Vignat J, Ferlay
J, Bray F and Franceschi S: Global burden of cancers attributable
to infections in 2012: A synthetic analysis. Lancet Glob Health.
4:e609–e616. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
de Martel C, Georges D, Bray F, Ferlay J
and Clifford GM: Global burden of cancer attributable to infections
in 2018: A worldwide incidence analysis. Lancet Glob Health.
8:e180–e190. 2020. View Article : Google Scholar
|
|
119
|
Cacioppo JT, Kiecolt-Glaser JK, Malarkey
WB, Laskowski BF, Rozlog LA, Poehlmann KM, Burleson MH and Glaser
R: Autonomic and glucocorticoid associations with the steady-state
expression of latent Epstein-Barr virus. Horm Behav. 42:32–41.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Glaser R, Friedman SB, Smyth J, Ader R,
Bijur P, Brunell P, Cohen N, Krilov LR, Lifrak ST, Stone A and
Toffler P: The differential impact of training stress and final
examination stress on herpesvirus latency at the United States
Military Academy at West Point. Brain Behav Immun. 13:240–251.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Stamatiou DP, Derdas SP, Zoras OL and
Spandidos DA: Herpes and polyoma family viruses in thyroid cancer.
Oncol Lett. 11:1635–1644. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Mammas IN, Sourvinos G, Giannoudis A and
Spandidos DA: Human papilloma virus (HPV) and host cellular
interactions. Pathol Oncol Res. 14:345–354. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Antoni MH, Lutgendorf SK, Cole SW, Dhabhar
FS, Sephton SE, McDonald PG, Stefanek M and Sood AK: The influence
of bio-behavioural factors on tumour biology: Pathways and
mechanisms. Nat Rev Cancer. 6:240–248. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Irwin MR and Cole SW: Reciprocal
regulation of the neural and innate immune systems. Nat Rev
Immunol. 11:625–632. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Collado-Hidalgo A, Sung C and Cole S:
Adrenergic inhibition of innate anti-viral response: PKA blockade
of Type I interferon gene transcription mediates catecholamine
support for HIV-1 replication. Brain Behav Immun. 20:552–563. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Fang CY, Miller SM, Bovbjerg DH, Bergman
C, Edelson MI, Rosenblum NG, Bove BA, Godwin AK, Campbell DE and
Douglas SD: Perceived stress is associated with impaired T-cell
response to HPV16 in women with cervical dysplasia. Ann Behav Med.
35:87–96. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Fischman HK, Pero RW and Kelly DD:
Psychogenic stress induces chromosomal and DNA damage. Int J
Neuroscience. 84:219–227. 1996. View Article : Google Scholar
|
|
128
|
Irie M, Asami S, Nagata S, Ikeda M, Miyata
M and Kasai H: Psychosocial factors as a potential trigger of
oxidative DNA damage in human leukocytes. Japanese J Cancer Res.
92:367–376. 2001. View Article : Google Scholar
|
|
129
|
Kiecolt-Glaser JK, Stephens RE, Lipetz PD,
Speicher CE and Glaser R: Distress and DNA repair in human
lymphocytes. J Behavioral Med. 8:311–320. 1985. View Article : Google Scholar
|
|
130
|
Dimitroglou E, Zafiropoulou M,
Messini-Nikolaki N, Doudounakis S, Tsilimigaki S and Piperakis SM:
DNA damage in a human population affected by chronic psychogenic
stress. Int J Hyg Environ Health. 206:39–44. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Flint MS, Baum A, Chambers WH and Jenkins
FJ: Induction of DNA damage, alteration of DNA repair and
transcriptional activation by stress hormones.
Psychoneuroendocrinology. 32:470–479. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Hara MR, Kovacs JJ, Whalen EJ, Rajagopal
S, Strachan RT, Grant W, Towers AJ, Williams B, Lam CM, Xiao K, et
al: A stress response pathway regulates DNA damage through
β2-adrenoreceptors and β-arrestin-1. Nature. 477:349–353. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Hara MR, Sachs BD, Caron MG and Lefkowitz
RJ: Pharmacological blockade of a β2AR-β-arrestin-1 signaling
cascade prevents the accumulation of DNA damage in a behavioral
stress model. Cell Cycle. 12:219–224. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Feng Z, Liu L, Zhang C, Zheng T, Wang J,
Lin M, Zhao Y, Wang X, Levine AJ and Hu W: Chronic restraint stress
attenuates p53 function and promotes tumorigenesis. Proc Natl Acad
Sci USA. 109:7013–7018. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Magnon C, Hall SJ, Lin J, Xue X, Gerber L,
Freedland SJ and Frenette PS: Autonomic nerve development
contributes to prostate cancer progression. Science.
341:12363612013. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Zachos G, Varras M, Koffa M, Ergazaki M
and Spandidos DA: The association of the H-ras oncogene and steroid
hormone receptors in gynecological cancer. J Exp Ther Oncol.
1:335–341. 1996.PubMed/NCBI
|
|
137
|
Wrobel LJ and Le Gal FA: Inhibition of
human melanoma growth by a non-cardioselective β-blocker. J
Investigative Dermatol. 135:525–531. 2015. View Article : Google Scholar
|
|
138
|
Armaiz-Pena GN, Allen JK, Cruz A, Stone
RL, Nick AM, Lin YG, Han LY, Mangala LS, Villares GJ, Vivas-Mejia
P, et al: Src activation by β-adrenoreceptors is a key switch for
tumour metastasis. Nat Commun. 4:14032013. View Article : Google Scholar
|
|
139
|
Shi M, Liu D, Duan H, Qian L, Wang L, Niu
L, Zhang H, Yong Z, Gong Z, Song L, et al: The β2-adrenergic
receptor and Her2 comprise a positive feedback loop in human breast
cancer cells. Br Cancer Res Treat. 125:351–362. 2011. View Article : Google Scholar
|
|
140
|
Saul AN, Oberyszyn TM, Daugherty C,
Kusewitt D, Jones S, Jewell S, Malarkey WB, Lehman A, Lemeshow S
and Dhabhar FS: Chronic stress and susceptibility to skin cancer. J
Natl Cancer Inst. 97:1760–1767. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Sumis A, Cook KL, Andrade FO, Hu R, Kidney
E, Zhang X, Kim D, Carney E, Nguyen N, Yu W, et al: Social
isolation induces autophagy in the mouse mammary gland: Link to
increased mammary cancer risk. Endocr Relat Cancer. 23:839–856.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Kokolus KM, Capitano ML, Lee CT, Eng JW,
Waight JD, Hylander BL, Sexton S, Hong CC, Gordon CJ, Abrams SI and
Repasky EA: Baseline tumor growth and immune control in laboratory
mice are significantly influenced by subthermoneutral housing
temperature. Proc Natl Acad Sci USA. 110:20176–20181. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Renz BW, Takahashi R, Tanaka T, Macchini
M, Hayakawa Y, Dantes Z, Maurer HC, Chen X, Jiang Z, Westphalen CB,
et al: β2 adrenergic-neurotrophin feedforward loop promotes
pancreatic cancer. Cancer Cell. 33:75–90.e7. 2018. View Article : Google Scholar
|
|
144
|
Hermes GL, Delgado B, Tretiakova M,
Cavigelli SA, Krausz T, Conzen SD and McClintock MK: Social
isolation dysregulates endocrine and behavioral stress while
increasing malignant burden of spontaneous mammary tumors. Proc
Natl Acad Sci USA. 106:22393–22398. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Hasen NS, O'Leary KA, Auger AP and Schuler
LA: Social isolation reduces mammary development, tumor incidence,
and expression of epigenetic regulators in wild-type and
p53-heterozygotic mice. Cancer Prev Res. 3:620–629. 2010.
View Article : Google Scholar
|
|
146
|
Riley V: Psychoneuroendocrine influences
on immunocompetence and neoplasia. Science. 212:1100–1109. 1981.
View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Leonardi GC, Candido S, Cervello M,
Nicolosi D, Raiti F, Travali S, Spandidos DA and Libra M: The tumor
microenvironment in hepatocellular carcinoma (review). Int J Oncol.
40:1733–1747. 2012.PubMed/NCBI
|
|
148
|
Wu T and Dai Y: Tumor microenvironment and
therapeutic response. Cancer Lett. 387:61–68. 2017. View Article : Google Scholar
|
|
149
|
Gysler SM and Drapkin R: Tumor
innervation: Peripheral nerves take control of the tumor
microenvironment. J Clin Invest. 131:e1472762021. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Monje M, Borniger JC, D'Silva NJ, Deneen
B, Dirks PB, Fattahi F, Frenette PS, Garzia L, Gutmann DH, Hanahan
D, et al: Roadmap for the emerging field of cancer neuroscience.
Cell. 181:219–222. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Yang EV, Kim SJ, Donovan EL, Chen M, Gross
AC, Webster Marketon JI, Barsky SH and Glaser R: Norepinephrine
upregulates VEGF, IL-8, and IL-6 expression in human melanoma tumor
cell lines: Implications for stress-related enhancement of tumor
progression. Brain Behav Immun. 23:267–275. 2009. View Article : Google Scholar :
|
|
152
|
Moretti S, Massi D, Farini V, Baroni G,
Parri M, Innocenti S, Cecchi R and Chiarugi P: β-adrenoceptors are
upregulated in human melanoma and their activation releases
pro-tumorigenic cytokines and metalloproteases in melanoma cell
lines. Lab Invest. 93:279–290. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Madden KS, Szpunar MJ and Brown EB:
β-Adrenergic receptors (β-AR) regulate VEGF and IL-6 production by
divergent pathways in high β-AR-expressing breast cancer cell
lines. Br Cancer Res Treat. 130:747–758. 2011. View Article : Google Scholar
|
|
154
|
Lutgendorf SK, Cole S, Costanzo E, Bradley
S, Coffin J, Jabbari S, Rainwater K, Ritchie JM, Yang M and Sood
AK: Stress-related mediators stimulate vascular endothelial growth
factor secretion by two ovarian cancer cell lines. Clin Cancer Res.
9:4514–4521. 2003.PubMed/NCBI
|
|
155
|
Chen H, Liu D, Yang Z, Sun L, Deng Q, Yang
S, Qian L, Guo L, Yu M, Hu M, et al: Adrenergic signaling promotes
angiogenesis through endothelial cell-tumor cell crosstalk. Endocr
Relat Cancer. 21:783–795. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Medeiros PJ and Jackson DN: Neuropeptide Y
Y5-receptor activation on breast cancer cells acts as a paracrine
system that stimulates VEGF expression and secretion to promote
angiogenesis. Peptides. 48:106–113. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Shan T, Ma J, Ma Q, Guo K, Guo J, Li X, Li
W, Liu J, Huang C, Wang F and Wu E: β2-AR-HIF-1α: A novel
regulatory axis for stress-induced pancreatic tumor growth and
angiogenesis. Curr Mol Med. 13:1023–1034. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Xu P, He H, Gu Y, Wang Y, Sun Z, Yang L
and Miao C: Surgical trauma contributes to progression of colon
cancer by downregulating CXCL4 and recruiting MDSCs. Exp Cell Res.
370:692–698. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Budiu RA, Vlad AM, Nazario L, Bathula C,
Cooper KL, Edmed J, Thaker PH, Urban J, Kalinski P, Lee AV, et al:
Restraint and social isolation stressors differentially regulate
adaptive immunity and tumor angiogenesis in a breast cancer mouse
model. Cancer Clin Oncol. 6:12–24. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Hulsurkar M, Li Z, Zhang Y, Li X, Zheng D
and Li W: Beta-adrenergic signaling promotes tumor angiogenesis and
prostate cancer progression through HDAC2-mediated suppression of
thrombospondin-1. Oncogene. 36:1525–1536. 2017. View Article : Google Scholar
|
|
161
|
Le CP, Nowell CJ, Kim‑Fuchs C, Botteri E,
Hiller JG, Ismail H, Pimentel MA, Chai MG, Karnezis T, Rotmensz N,
et al: Chronic stress in mice remodels lymph vasculature to promote
tumour cell dissemination. Nat Commun. 7:106342016. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Colon-Echevarria CB, Lamboy-Caraballo R,
Aquino-Acevedo AN and Armaiz-Pena GN: Neuroendocrine regulation of
tumor-associated immune cells. Front Oncol. 9:10772019. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Qiao G, Chen M, Bucsek MJ, Repasky EA and
Hylander BL: Adrenergic signaling: A targetable checkpoint limiting
development of the antitumor immune response. Front Immunol.
9:1642018. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Grivennikov SI, Greten FR and Karin M:
Immunity, inflammation, and cancer. Cell. 140:883–899. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Huan HB, Wen XD, Chen XJ, Wu L, Wu LL,
Zhang L, Yang DP, Zhang X, Bie P, Qian C and Xia F: Sympathetic
nervous system promotes hepatocarcinogenesis by modulating
inflammation through activation of alpha1-adrenergic receptors of
Kupffer cells. Brain Behav Immun. 59:118–134. 2017. View Article : Google Scholar
|
|
166
|
Nagaraja AS, Dorniak PL, Sadaoui NC, Kang
Y, Lin T, Armaiz-Pena G, Wu SY, Rupaimoole R, Allen JK, Gharpure
KM, et al: Sustained adrenergic signaling leads to increased
metastasis in ovarian cancer via increased PGE2 synthesis.
Oncogene. 35:2390–2397. 2016. View Article : Google Scholar :
|
|
167
|
Qin JF, Jin FJ, Li N, Guan HT, Lan L, Ni H
and Wang Y: Adrenergic receptor β2 activation by stress promotes
breast cancer progression through macrophages M2 polarization in
tumor microenvironment. BMB Rep. 48:2952015. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Armaiz-Pena GN, Gonzalez-Villasana V,
Nagaraja AS, Rodriguez-Aguayo C, Sadaoui NC, Stone RL, Matsuo K,
Dalton HJ, Previs RA, Jennings NB, et al: Adrenergic regulation of
monocyte chemotactic protein 1 leads to enhanced macrophage
recruitment and ovarian carcinoma growth. Oncotarget. 6:4266–4273.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Lamkin DM, Ho H-Y, Ong TH, Kawanishi CK,
Stoffers VL, Ahlawat N, Ma JCY, Arevalo JMG, Cole SW and Sloan EK:
β-Adrenergic-stimulated macrophages: Comprehensive localization in
the M1-M2 spectrum. Brain Behav Immun. 57:338–346. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Bower JE, Shiao SL, Sullivan P, Lamkin DM,
Atienza R, Mercado F, Arevalo J, Asher A, Ganz PA and Cole SW:
Prometastatic molecular profiles in breast tumors from socially
isolated women. JNCI Cancer Spectr. 2:pky0292018. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Shakhar G and Ben-Eliyahu S: In vivo
β-adrenergic stimulation suppresses natural killer activity and
compromises resistance to tumor metastasis in rats. J Immunol.
160:3251–3258. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Rosenne E, Sorski L, Shaashua L, Neeman E,
Matzner P, Levi B and Ben-Eliyahu S: In vivo suppression of NK cell
cytotoxicity by stress and surgery: Glucocorticoids have a minor
role compared to catecholamines and prostaglandins. Brain Behav
Immun. 37:207–219. 2014. View Article : Google Scholar
|
|
173
|
Lutgendorf SK, Sood AK, Anderson B, McGinn
S, Maiseri H, Dao M, Sorosky JI, De Geest K, Ritchie J and Lubaroff
DM: Social support, psychological distress, and natural killer cell
activity in ovarian cancer. J Clin Oncol. 23:7105–7113. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Hou N, Zhang X, Zhao L, Zhao X, Li Z, Song
T and Huang C: A novel chronic stress-induced shift in the Th1 to
Th2 response promotes colon cancer growth. Biochem Biophys Res
Commun. 439:471–476. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Bucsek MJ, Qiao G, MacDonald CR,
Giridharan T, Evans L, Niedzwecki B, Liu H, Kokolus KM, Eng JW,
Messmer MN, et al: β-Adrenergic signaling in mice housed at
standard temperatures suppresses an effector phenotype in
CD8+ T cells and undermines checkpoint inhibitor
therapy. Cancer Res. 77:5639–5651. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Lutgendorf SK, Lamkin DM, DeGeest K,
Anderson B, Dao M, McGinn S, Zimmerman B, Maiseri H, Sood AK and
Lubaroff DM: Depressed and anxious mood and T-cell cytokine
expressing populations in ovarian cancer patients. Brain Behav
Immun. 22:890–900. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Mohammadpour H, MacDonald CR, Qiao G, Chen
M, Dong B, Hylander BL, McCarthy PL, Abrams SI and Repasky EA: β2
adrenergic receptor-mediated signaling regulates the
immunosuppressive potential of myeloid-derived suppressor cells. J
Clin Invest. 129:5537–5552. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Mundy-Bosse BL, Thornton LM, Yang HC,
Andersen BL and Carson WE: Psychological stress is associated with
altered levels of myeloid-derived suppressor cells in breast cancer
patients. Cell Immunol. 270:80–87. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Nguyen KD, Qiu Y, Cui X, Goh YP, Mwangi J,
David T, Mukundan L, Brombacher F, Locksley RM and Chawla A:
Alternatively activated macrophages produce catecholamines to
sustain adaptive thermogenesis. Nature. 480:104–108. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Flierl MA, Rittirsch D, Nadeau BA, Chen
AJ, Sarma JV, Zetoune FS, McGuire SR, List RP, Day DE, Hoesel LM,
et al: Phagocyte-derived catecholamines enhance acute inflammatory
injury. Nature. 449:721–725. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Wong HPS, Yu L, Lam EKY, Tai EKK, Wu WKK
and Cho CH: Nicotine promotes cell proliferation via α7-nicotinic
acetylcholine receptor and catecholamine-synthesizing
enzymes-mediated pathway in human colon adenocarcinoma HT-29 cells.
Toxicol Appl Pharmacol. 221:261–267. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Amaro F, Silva D, Reguengo H, Oliveira JC,
Quintas C, Vale N, Gonçalves J and Fresco P: β-adrenoceptor
activation in breast MCF-10A cells induces a pattern of
catecholamine production similar to that of tumorigenic MCF-7
cells. Int J Mol Sci. 21:79682020. View Article : Google Scholar
|
|
183
|
Zhang X, Zhang Y, He Z, Yin K, Li B, Zhang
L and Xu Z: Chronic stress promotes gastric cancer progression and
metastasis: An essential role for ADRB2. Cell Death Dis.
10:7882019. View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Zhi X, Li B, Li Z, Zhang J, Yu J, Zhang L
and Xu Z: Adrenergic modulation of AMPK-dependent autophagy by
chronic stress enhances cell proliferation and survival in gastric
cancer. Int J Oncol. 54:1625–1638. 2019.PubMed/NCBI
|
|
185
|
Wong HP, Ho JW, Koo MW, Yu L, Wu WK, Lam
EK, Tai EK, Ko JK, Shin VY, Chu KM and Cho CH: Effects of
adrenaline in human colon adenocarcinoma HT-29 cells. Life Sci.
88:1108–1112. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Sood AK, Armaiz-Pena GN, Halder J, Nick
AM, Stone RL, Hu W, Carroll AR, Spannuth WA, Deavers MT, Allen JK,
et al: Adrenergic modulation of focal adhesion kinase protects
human ovarian cancer cells from anoikis. J Clin Invest.
120:1515–1523. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Liu H, Wang C, Xie N, Zhuang Z, Liu X, Hou
J and Huang H: Activation of adrenergic receptor β2 promotes tumor
progression and epithelial mesenchymal transition in tongue
squamous cell carcinoma. Int J Mol Med. 41:147–154. 2018.
|
|
188
|
Kim-Fuchs C, Le CP, Pimentel MA,
Shackleford D, Ferrari D, Angst E, Hollande F and Sloan EK: Chronic
stress accelerates pancreatic cancer growth and invasion: A
critical role for beta-adrenergic signaling in the pancreatic
microenvironment. Brain Behav Immun. 40:40–47. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Pu J, Zhang X, Luo H, Xu L, Lu X and Lu J:
Adrenaline promotes epithelial-to-mesenchymal transition via
HuR-TGFβ regulatory axis in pancreatic cancer cells and the
implication in cancer prognosis. Biochem Biophys Res Commun.
493:1273–1279. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Liu J, Qu L, Wan C, Xiao M, Ni W, Jiang F,
Fan Y, Lu C and Ni R: A novel β2-AR/YB-1/β-catenin axis mediates
chronic stress-associated metastasis in hepatocellular carcinoma.
Oncogenesis. 9:842020. View Article : Google Scholar
|
|
191
|
Chen H, Liu D, Guo L, Cheng X, Guo N and
Shi M: Chronic psychological stress promotes lung metastatic
colonization of circulating breast cancer cells by decorating a
pre-metastatic niche through activating β-adrenergic signaling. J
Pathol. 244:49–60. 2018. View Article : Google Scholar
|
|
192
|
Lamkin DM, Sloan EK, Patel AJ, Chiang BS,
Pimentel MA, Ma JC, Arevalo JM, Morizono K and Cole SW: Chronic
stress enhances progression of acute lymphoblastic leukemia via
β-adrenergic signaling. Brain Behav Immun. 26:635–641. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Chang A, Le CP, Walker AK, Creed SJ, Pon
CK, Albold S, Carroll D, Halls ML, Lane JR, Riedel B, et al:
β2-Adrenoceptors on tumor cells play a critical role in
stress-enhanced metastasis in a mouse model of breast cancer. Brain
Behav Immun. 57:106–115. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Zahalka AH and Frenette PS: Nerves in
cancer. Nat Rev Cancer. 20:143–157. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Lutgendorf SK, DeGeest K, Dahmoush L,
Farley D, Penedo F, Bender D, Goodheart M, Buekers TE, Mendez L,
Krueger G, et al: Social isolation is associated with elevated
tumor norepinephrine in ovarian carcinoma patients. Brain Behav
Immun. 25:250–255. 2011. View Article : Google Scholar :
|
|
196
|
Schuller HM and Cole B: Regulation of cell
proliferation by β-adrenergjc receptors in a human lung
adenocarcinoma cell line. Carcinogenesis. 10:1753–1755. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
197
|
Huang XY, Wang HC, Yuan Z, Huang J and
Zheng Q: Norepinephrine stimulates pancreatic cancer cell
proliferation, migration and invasion via β-adrenergic
receptor-dependent activation of P38/MAPK pathway.
Hepatogastroenterology. 59:889–893. 2012.
|
|
198
|
Lackovicova L, Banovska L, Bundzikova J,
Janega P, Bizik J, Kiss A and Mravec B: Chemical sympathectomy
suppresses fibrosarcoma development and improves survival of
tumor-bearing rats. Neoplasma. 58:424–429. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
199
|
Simon WE, Albrecht M, Trams G, Dietel M
and Hölzel F: In vitro growth promotion of human mammary carcinoma
cells by steroid hormones, tamoxifen, and prolactin. J Natl Cancer
Inst. 73:313–321. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
200
|
Zhao XY, Malloy PJ, Krishnan AV, Swami S,
Navone NM, Peehl DM and Feldman D: Glucocorticoids can promote
androgen-independent growth of prostate cancer cells through a
mutated androgen receptor. Nat Med. 6:703–706. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
201
|
Horvathova L, Padova A, Tillinger A,
Osacka J, Bizik J and Mravec B: Sympathectomy reduces tumor weight
and affects expression of tumor-related genes in melanoma tissue in
the mouse. Stress. 19:528–534. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
202
|
Chin CC, Li JM, Lee KF, Huang YC, Wang KC,
Lai HC, Cheng CC, Kuo YH and Shi CS: Selective β2-AR blockage
suppresses colorectal cancer growth through regulation of
EGFR-Akt/ERK1/2 signaling, G1-phase arrest, and apoptosis. J Cell
Physiol. 231:459–472. 2016. View Article : Google Scholar
|
|
203
|
Masur K, Niggemann B, Zanker KS and
Entschladen F: Norepinephrine-induced migration of SW 480 colon
carcinoma cells is inhibited by β-blockers. Cancer Res.
61:2866–2869. 2001.PubMed/NCBI
|
|
204
|
Yang EV, Sood AK, Chen M, Li Y, Eubank TD,
Marsh CB, Jewell S, Flavahan NA, Morrison C, Yeh PE, et al:
Norepinephrine up-regulates the expression of vascular endothelial
growth factor, matrix metalloproteinase (MMP)-2, and MMP-9 in
nasopharyngeal carcinoma tumor cells. Cancer Res. 66:10357–10364.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
205
|
Sood AK, Bhatty R, Kamat AA, Landen CN,
Han L, Thaker PH, Li Y, Gershenson DM, Lutgendorf S and Cole SW:
Stress hormone-mediated invasion of ovarian cancer cells. Clin
Cancer Res. 12:369–375. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
206
|
Lutgendorf SK, Lamkin DM, Jennings NB,
Arevalo JM, Penedo F, DeGeest K, Langley RR, Lucci JA III, Cole SW,
Lubaroff DM and Sood AK: Biobehavioral influences on matrix
metalloproteinase expression in ovarian carcinoma. Clin Cancer Res.
14:6839–6846. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
207
|
Campbell JP, Karolak MR, Ma Y, Per rien
DS, Masood-Campbell SK, Penner NL, Munoz SA, Zijlstra A, Yang X,
Sterling JA and Elefteriou F: Stimulation of host bone marrow
stromal cells by sympathetic nerves promotes breast cancer bone
metastasis in mice. PLoS Biol. 10:e10013632012. View Article : Google Scholar : PubMed/NCBI
|
|
208
|
Simpson CD, Anyiwe K and Schimmer AD:
Anoikis resistance and tumor metastasis. Cancer Lett. 272:177–185.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
209
|
Benish M, Bartal I, Goldfarb Y, Levi B,
Avraham R, Raz A and Ben-Eliyahu S: Perioperative use of
beta-blockers and COX-2 inhibitors may improve immune competence
and reduce the risk of tumor metastasis. Ann Surg Oncol.
15:2042–2052. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
210
|
Kaira K, Kamiyoshihara M, Kawashima O,
Endoh H, Imaizumi K, Sugano M, Tanaka S, Fujita A, Kogure Y,
Shimizu A, et al: Prognostic impact of β2 adrenergic receptor
expression in surgically resected pulmonary pleomorphic carcinoma.
Anticancer Res. 39:395–403. 2019. View Article : Google Scholar
|
|
211
|
Choy C, Raytis JL, Smith DD, Duenas M,
Neman J, Jandial R and Lew MW: Inhibition of β2-adrenergic receptor
reduces triple-negative breast cancer brain metastases: The
potential benefit of perioperative β-blockade. Oncol Rep.
35:3135–3142. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
212
|
Al-Niaimi A, Dickson EL, Albertin C,
Karnowski J, Niemi C, Spencer R, Shahzad MM, Uppal S, Saha S, Rice
L and Nally AM: The impact of perioperative β blocker use on
patient outcomes after primary cytoreductive surgery in high-grade
epithelial ovarian carcinoma. Gynecol Oncol. 143:521–525. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
213
|
Barron TI, Connolly RM, Sharp L, Bennett K
and Visvanathan K: Beta blockers and breast cancer mortality: A
population-based study. J Clin Oncol. 29:2635–2644. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
214
|
Lemeshow S, Sørensen HT, Phillips G, Yang
EV, Antonsen S, Riis AH, Lesinski GB, Jackson R and Glaser R:
β-Blockers and survival among Danish patients with malignant
melanoma: A population-based cohort study. Cancer Epidemiol
Biomarkers Prev. 20:2273–2279. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
215
|
Cata JP, Villarreal J, Keerty D, Thakar
DR, Liu DD, Sood AK and Gottumukkala V: Perioperative beta-blocker
use and survival in lung cancer patients. J Clin Anesth.
26:106–117. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
216
|
Heitz F, Hengsbach A, Harter P, Traut A,
Ataseven B, Schneider S, Prader S, Kurzeder C, Sporkmann M and du
Bois A: Intake of selective beta blockers has no impact on survival
in patients with epithelial ovarian cancer. Gynecol Oncol.
144:181–186. 2017. View Article : Google Scholar
|
|
217
|
Kim TH, Gill NK, Nyberg KD, Nguyen AV,
Hohlbauch SV, Geisse NA, Nowell CJ, Sloan EK and Rowat AC: Cancer
cells become less deformable and more invasive with activation of
β-adrenergic signaling. J Cell Sci. 129:4563–4575. 2016.PubMed/NCBI
|