Introduction
Glioblastoma is the most common type of malignancy
of the central nervous system, accounts for almost 80% of all
malignant primary brain tumors and is the second most commonly
reported type of brain tumor. It accounts for almost all gliomas in
the United States, with a 5-year survival rate of 3% (1). Despite ongoing efforts to develop
effective molecular targets and combination therapies, the
treatment of glioblastoma remains challenging due to several
factors, including high therapeutic resistance, redundancy in
aberrantly activated signaling pathways, and difficulties in
effective drug delivery (2).
Therefore, novel therapeutic strategies and additional research are
essential for enhancing the effectiveness and outcome of
glioblastoma treatment.
Tumor necrosis factor-related apoptosis-inducing
ligand (TRAIL) is a promising therapeutic treatment for cancer
(3). TRAIL is a cytokine that
induces apoptosis by binding to TRAIL-receptor 1 (DR4) and
TRAIL-receptor 2 (DR5) and forms an apoptosis-inducing signaling
complex by interacting with caspase-8 (4-6).
The extrinsic route that initiates TRAIL-induced apoptosis involves
proteolytic caspase-8 activation, followed by activation of
effector caspases, such as caspase-3 (7). TRAIL induces apoptosis in various
types of cancer cells but typically does not affect normal cells
(8). However, certain cancer
types, such as glioblastoma, colon (9) and breast cancer (10) and hepatocellular carcinoma
(11), exhibit resistance to
TRAIL-induced apoptosis. Therefore, TRAIL alone may be insufficient
for treating certain malignant tumors, highlighting the need to
sensitize these cancer cells to TRAIL-induced apoptosis for
effective therapy.
Chaetocin is a natural compound produced by members
of the fungal genus Chaetomium; it inhibits activity of the
histone methyltransferase SUV39H1 (12). Chaetocin serves as a potent and
selective anti-myeloma agent owing to its ability to induce
oxidative stress (13).
Additionally, chaetocin modulates SUV39H1 activity in a reactive
oxygen species (ROS)-dependent manner, resulting in DR-dependent
apoptosis (14). Liu et al
(15) reported that chaetocin
promotes endoplasmic reticulum (ER) stress and increases the
expression of ER stress markers, including activating transcription
factor 3 (ATF3) and C/EBP homologous protein (CHOP), ultimately
leading to DR5-dependent apoptosis. However, the precise mechanisms
underlying chaetocin-induced sensitization of tumors to TRAIL are
poorly understood. Therefore, the present study aimed to examine
the sensitizing effects and underlying mechanism of chaetocin in
TRAIL-induced apoptosis in human glioblastoma cells, in order to
evaluate its potential as a therapeutic agent for overcoming
treatment resistance in glioblastoma.
Materials and methods
Chemicals
Chaetocin was obtained from Enzo Life Sciences, Inc.
(cat. no. BML-GR349). Pan-caspase inhibitor z-VAD-fmk and
recombinant human TRAIL were purchased from R&D Systems, Inc.
(cat. nos. 375-TL and FMK001, respectively). N-acetylcysteine (NAC;
cat. no. 01810), cycloheximide (cat. no. A7250), MG132 (cat. no.
M8699), and thapsigargin (TG; ER stress inducer; cat. no. T9033)
were purchased from Sigma-Aldrich Dulbecco's modified eagle's
medium (DMEM), antibiotics, and fetal bovine serum (FBS; cat. no.
PK004-07) were obtained from Welgene, Inc.
Cell culture
Human glioblastoma U343MG, U87MG (glioblastoma of
unknown origin, HTB-14), U251MG, and T98G (CRL-1690) and embryonic
kidney (HEK293, CRL-1573) cells were purchased from American Type
Culture Collection and maintained in DMEM supplemented with 10% FBS
and 1% penicillin/streptomycin at 37°C in a humidified atmosphere
containing 5% CO2. Cells were cultured for 16 h until
60-70% confluency was reached, and then treated with chaetocin (500
nM) or TRAIL (50 ng/ml) for 6 h at 37°C.
Treatment
U87MG and T98G cells were seeded in 6-well plates at
a density of 25×104 cells/well. After 24 h, cells were
treated with 10 µM TG for 6 h at 37°C to induce ER
stress.
Small interfering (si)RNA-mediated RNA
interference
DR5-targeting and scrambled siRNAs were purchased
from Santa Cruz Biotechnology Co., Ltd. (cat. nos. sc-40237 and
sc-37007, respectively; sequences not available). U87MG and T98G
cells were seeded in 6-well plates at a density of
25×104 cells/well and transfected with 50 nM siRNA with
Lipofectamine 2000 (cat. no. 11668019; Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions at
37°C for 24 h. The medium was replaced with DMEM containing 10% FBS
without antibiotics, and the cells were treated as aforementioned.
Six hours after treatment, cells were collected for viability
assays and western blot analysis.
Cell viability assay and Morphological
Analysis
CellTiter 96® AQueous One Solution Cell
Proliferation Assay system (cat. no. 3580; Promega Corporation) was
used according to the manufacturer's instructions. Cells were
cultured in a 37°C CO2 incubator in 96-well cell culture
plates at a density of 2×104 cells/well for 1 day and
treated as aforementioned. Each well was loaded with MTS reagent
and incubated at 37°C for 20 min in a 5% CO2
environment. Absorbance at 450 nm was measured using a Synergy/HTX
spectrophotometer (BioTek Instruments, Inc.). Experiments were
carried out in triplicate. For cell morphology analysis, cells were
treated with chaetocin (500 nM) and TRAIL (50 ng/ml) for 6 h. Phase
contrast images were captured using a Leica light microscope (Leica
Microsystems), and image acquisition was performed with Leica LAS X
Core software (version 3.6).
Flow cytometry
Cells were fixed in 80% ethanol at 4°C for ≥1 h
before determining the DNA content using flow cytometry. The cells
were subsequently stained with propidium iodide for 30 min at 4°C
and analyzed using a BD FACSCanto™ II flow cytometer (BD
Biosciences). Quantification was performed with BD FACS Diva
software version 7.0 (BD Biosciences).
Western blotting
Cells were lysed with RIPA lysis buffer (cat. no.
89900, Thermo Fisher Scientific, Inc.) and supernatant was
collected. Bicinchoninic acid was used to measure the protein
content. Western blotting was performed as described previously
(16). Protein (50
µg/lane) was resolved using 13% SDS-PAGE and transferred to
nitrocellulose membranes. The membranes were blocked with 5%
non-fat milk for 1 h at room temperature. Membranes were incubated
with primary antibodies: anti-PARP (cat. no. #9542), anti-caspase-8
(cat. no. #9496; Cell Signaling Technology, Inc.),
anti-pro-caspase-3 (cat. no. #sc7148; Santa Cruz Biotechnology Co.,
Ltd.), anti-cleaved caspase-3 (cat. no. #9661S), anti-DR5 (cat. no.
#8074S; Cell Signaling Technology, Inc.), anti-DR4 (cat. no.
#sc8411), anti-78-kDa glucose-regulated protein (Bip) (cat. no.
#sc13968), anti-CHOP (cat. no. #sc7351), anti-p53 (1:1,000; cat.
no. #sc126; Santa Cruz Biotechnology Co., Ltd.) and anti-β-actin
(ACTB) (1:5,000; cat. no. A5441, Sigma-Aldrich). The membranes were
incubated overnight with the primary antibodies at 4°C. The
membranes were then incubated with anti-rabbit (cat. no.
111-035-045) and anti-mouse IgG (both 1:5,000; cat. no.
115-035-062; Jackson ImmunoResearch Laboratories, Inc.) at room
temperature for 1 h. Protein bands were identified using Immobilon
Western Chemiluminescent HRP Substrate (cat. no. WBKLS0500;
MilliporeSigma). The blots were quantified by densitometric
analysis using ImageJ v4.0 software (National Institutes of Health)
and the relative expression of each target protein was normalized
using ACTB.
Caspase-3/7 activity assay
Cells (5×104/well) were seeded in a
96-well plate and treated as aforementioned. A total of 100
µl Caspase-Glo® 3/7 Assay reagent (cat. no.
G8090; Promega Corporation) was added and incubated at 37°C for 1 h
and luminescence was measured using a Synergy/HTX spectrophotometer
(BioTek instrument, Inc.).
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted from cells using TRIzol
(cat. no. 15596-026; Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions. RNA was reverse-transcribed into
cDNA using the reverse transcriptase premix (cat. no. EBT-1515;
Elpis Biotech, Inc.) at 40°C for 60 min, followed by enzyme
inactivation at 92°C for 5 min. RT-qPCR was performed using Blend
Taq DNA polymerase (cat. no. M8291; Promega Corporation) with
primers for DR5 and GAPDH. The sequences were follows: DR5
(forward: 5′-AAGACCCTTGTGCTCGTTGT-3′, reverse:
5′-GACACATTCGATGTCACTCCA-3′) and GAPDH (forward:
5′-CGTCTTCACCACCATGGAGA-3′, reverse: 5′-CGGCCATCACGCCCAGTTT-3′).
The qPCR thermocycling conditions were as follows: 95°C for 30 sec,
followed by 25 cycles of degradation at 94°C for 30 sec, annealing
at 56°C for 30 sec, extension at 72°C for 30 sec for DR5. Amplified
products were separated on a 2% agarose gel and bands were
visualized on a LAS-3000 (FujiFilm Wako Pure chemical corporation).
The relative gene expression levels were quantified using the
2−ΔΔCq method (17)
with normalization to the control.
Cell surface staining for DR5
Cells were washed three times in PBS and suspended
in 200 µl 2% FBS/PBS. The primary antibody (anti-DR5, 1:100,
cat. no. #8074S; Cell Signaling Technology, Inc.) was added at room
temperature for 30 min. The cells were washed twice with PBS,
resuspended in 200 µl 2% FBS/PBS, and incubated with
fluorescein isothiocyanate-conjugated secondary antibody (Alexa
Fluor® 488 Goat Anti-Rabbit IgG (H+L), 1:100, cat. no.
A11008; Invitrogen, Thermo Fisher Scientific, Inc.) for 30 min at
room temperature. The cells were centrifuged at 200 × g for 5 min
at room temperature to remove unbound secondary antibodies and
resuspended in 500 µl PBS. DR5 expression on the cell
surface was measured using flow cytometry as aforementioned.
Cycloheximide (CHX) chase assay
U87MG and T98G cells were seeded in 6-well plates at
a density of 25×104 cells/well and cultured for 16 h in
a 37°C CO2 incubator. The cells were then treated with
500 nM chaetocin for 6 h, followed by treatment with 50 µM
CHX or CHX + chaetocin for 1, 3, 5, and 7 h at 37°C. Protein
lysates were prepared using RIPA buffer (cat. no. 89900; Thermo
Fisher Scientific, Inc.). A total of 50 µg of lysates were
subjected to immunoblotting to assess the stability of DR5.
Anti-ACTB was used as a loading control. The blots were quantified
by densitometric analysis using ImageJ v4.0 software (National
Institutes of Health) and the relative expression level of DR5 was
normalized using ACTB.
Proteasome activity assay
Proteasome activity was measured using Suc-LLVY-AMC
(Biomol International). Cell lysates were prepared from
chaetocin-treated cells using RIPA lysis buffer (cat. no. 89900;
Thermo Fisher Scientific, Inc.). A mixture containing 5 µg
of protein in 100 mM Tris-HCl (pH 8.0), 10 mM MgCl2, and
2 mM ATP was incubated at 37°C for 30 min with 50 µM
Suc-LLVY-AMC. Chymotrypsin-like proteasome activity was measured
using a fluorometric plate reader with excitation and emission
wavelengths of 380 nm and 440 nm, respectively. MG132 (10
µM) was used as a positive control to inhibit proteasome
activity for 6 h at 37°C.
ROS production
The fluorescent probe 2′,7′
dichlorodihy-drofluorescein diacetate (H2DCFDA; Sigma-Aldrich;
Merck KGaA) was used to measure intracellular ROS production. U87MG
and T98G cells were pretreated with 500 nM chaetocin for 6 h at
37°C, after which 10 µM H2DCFDA was added for 30
min at room temperature. Following incubation, cells were
trypsinized, resuspended in PBS and transferred to
Falcon® FACS tubes. H2DCFDA fluorescence
intensity was evaluated with a BD FACS Canto™ II flow cytometer (BD
Biosciences). The excitation wavelength was 488 nm and the emission
wavelength was 520 nm. Data were analyzed using BD Biosciences FACS
Canto™ software (version 6.0).
Statistical analysis
Data are presented as the mean ± SD of three
independent experiments. Differences were assessed using one-way
ANOVA and Tukey's post-hoc test. All statistical analyses were
performed with SPSS 11.5 (SPSS, Inc.) software. P<0.05 was
considered to indicate a statistically significant difference.
Results
Chaetocin enhances TRAIL sensitivity in
human glioblastoma cells
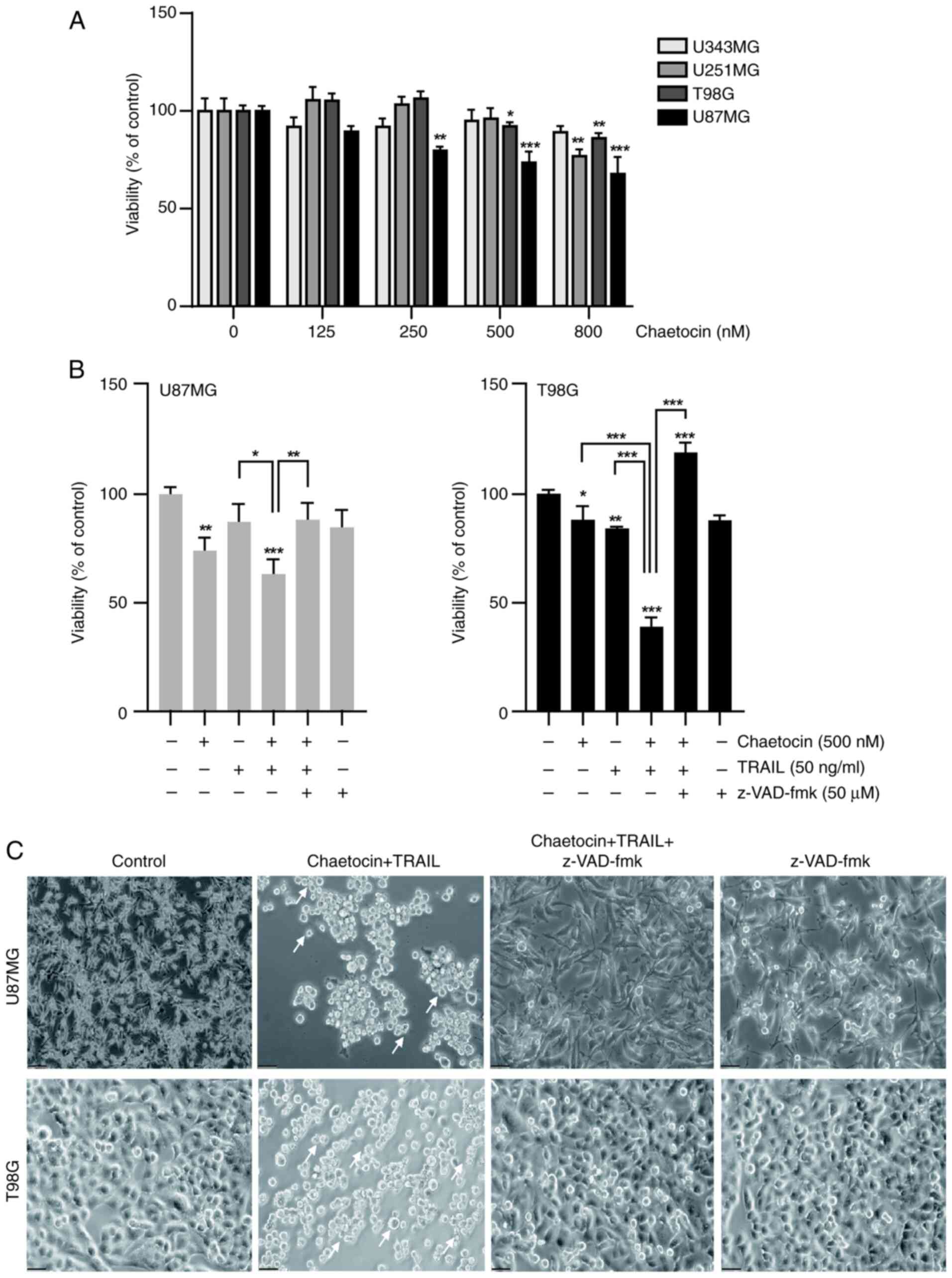
To assess the effect of chaetocin on cancer cell
sensitivity to TRAIL, the viability of human glioblastoma cell
lines U343MG, T98G, U251MG, and U87MG was evaluated after 6 h of
treatment with chaetocin. T98G and U87MG cells were the most
sensitive to chaetocin (Fig. 1A)
and were selected for the subsequent experiments. Chaetocin alone
significantly decreased the viability of the U87MG and T98G
glioblastoma cell lines (Fig.
1B). In particular, combined treatment with chaetocin and TRAIL
significantly decreased cell viability and induced morphological
changes, such as cell shrinkage and blebbing, in U87MG and T98G
cells (Fig. 1B and C). To
investigate the role of caspases in cell death, the present study
assessed the impact of the pan-caspase inhibitor z-VAD-fmk on cell
viability. z-VAD-fmk inhibited cell death induced by chaetocin +
TRAIL (Fig. 1B). Additionally,
phase contrast microscopy showed that co-treatment with chaetocin
and TRAIL increased the number of apoptotic bodies compared with
the untreated group (Fig. 1C).
Moreover, chaetocin improved the sensitivity of U343MG and U251MG
cells to TRAIL-mediated cell death (Fig. S1A) without affecting normal cells
(Fig. S1B). Collectively, these
data suggest that chaetocin enhances TRAIL-induced
caspase-dependent cell death.
Chaetocin enhances TRAIL-mediated
apoptosis in human glioblastoma cells
Since chaetocin promotes TRAIL-mediated cell death
through caspase activation, the present study investigated whether
the cell death induced by the combination of chaetocin and TRAIL
was attributable to apoptosis. U87MG and T98G cells were treated
with chaetocin and TRAIL for 6 h and harvested for cell cycle
analysis. The combination of chaetocin and TRAIL increased the
percentage of U87MG and T98G cells in the sub-G1 phase to 12.5% and
18.4%, respectively, compared with 2.8% and 1.8% in the control
group (Fig. 2A). Furthermore, the
present study confirmed the effect of chaetocin on TRAIL-mediated
apoptosis by examining caspase-3/7 activity. Notably, chaetocin
significantly enhanced TRAIL-induced activation of caspase-3/7 both
in U87MG and T98G cells (Fig.
2B). However, treatment with pan-caspase inhibitor z-VAD-fmk
almost completely blocked apoptosis and caspase-3/7 activity,
indicating that apoptosis induced by chaetocin and TRAIL was
caspase-dependent (Fig. 2A and
B). Additionally, combined treatment with TRAIL significantly
induced the cleavage of apoptosis markers PARP, caspase-8, and
caspase-3 in U87MG and T98G cells (Fig. 2C). Treatment with z-VAD-fmk
inhibited the cleavage of PARP and caspase-3 (Figs. 2C and S2). Collectively, these findings
indicate that chaetocin enhances TRAIL-induced apoptosis in
glioblastoma cells.
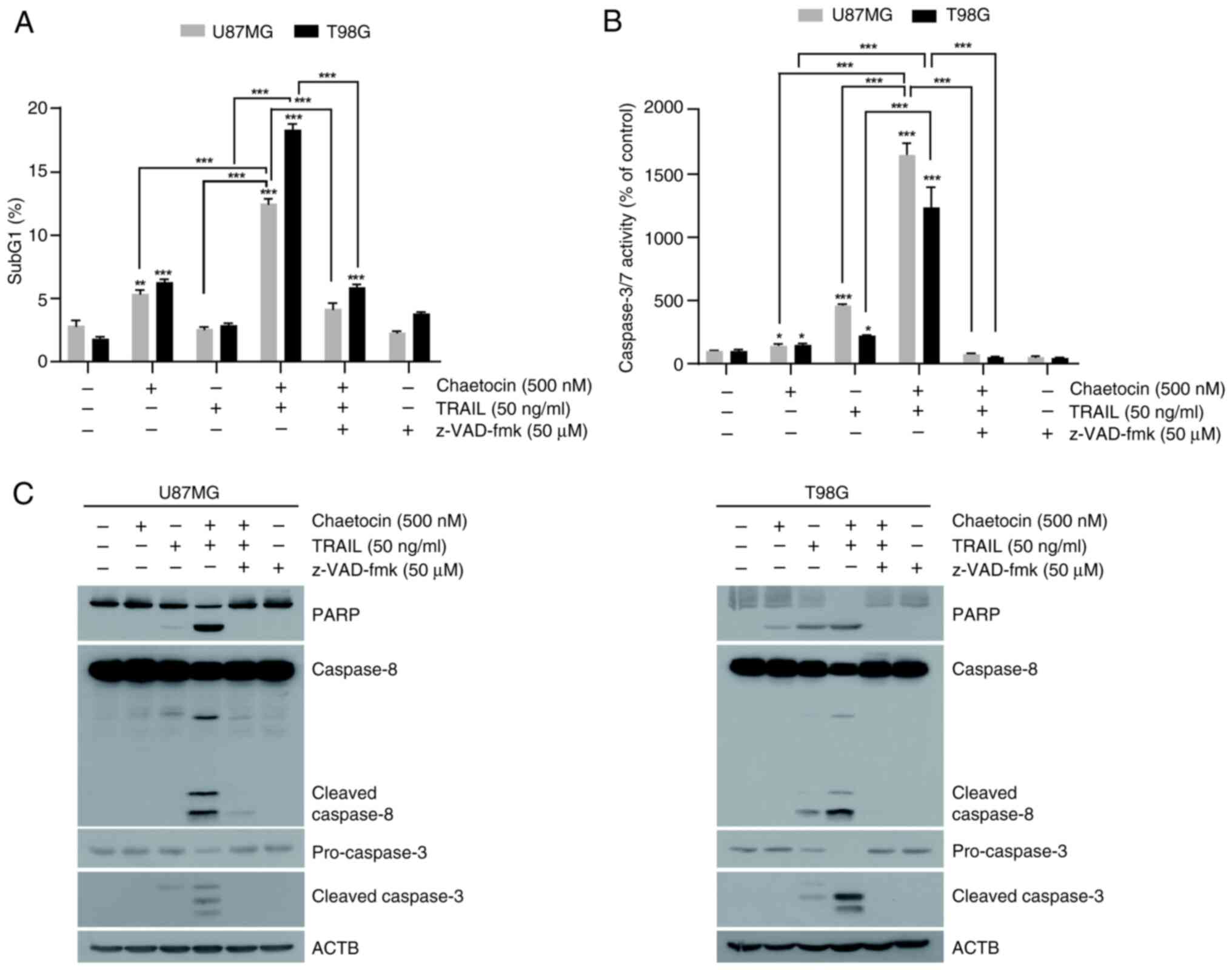 | Figure 2Chaetocin enhances TRAIL-mediated
apoptosis in human glioblastoma cell lines. (A) Flow cytometric
analysis of U87MF and T98G cells treated with chaetocin alone or in
combination with TRAIL and z-VAD-FMK for 6 h to determine the
sub-G1 population. (B) Caspase-3/7 activity in U87MG and T98G
cells. (C) Western blot analysis of PARP, caspase-8, pro-caspase-3,
and cleaved caspase-3 expression in U87MG and T98G cells treated
with chaetocin and TRAIL, with or without z-VAD-fmk, for 6 h.
*P<0.05, **P<0.01,
***P<0.001 vs. control. TRAIL, tumor necrosis
factor-related apoptosis-inducing ligand; ACTB, β-actin; z-VAD-fmk,
carbobenzoxy-valyl-alanyl-aspartyl-[O-methyl]-fluoromethylketone. |
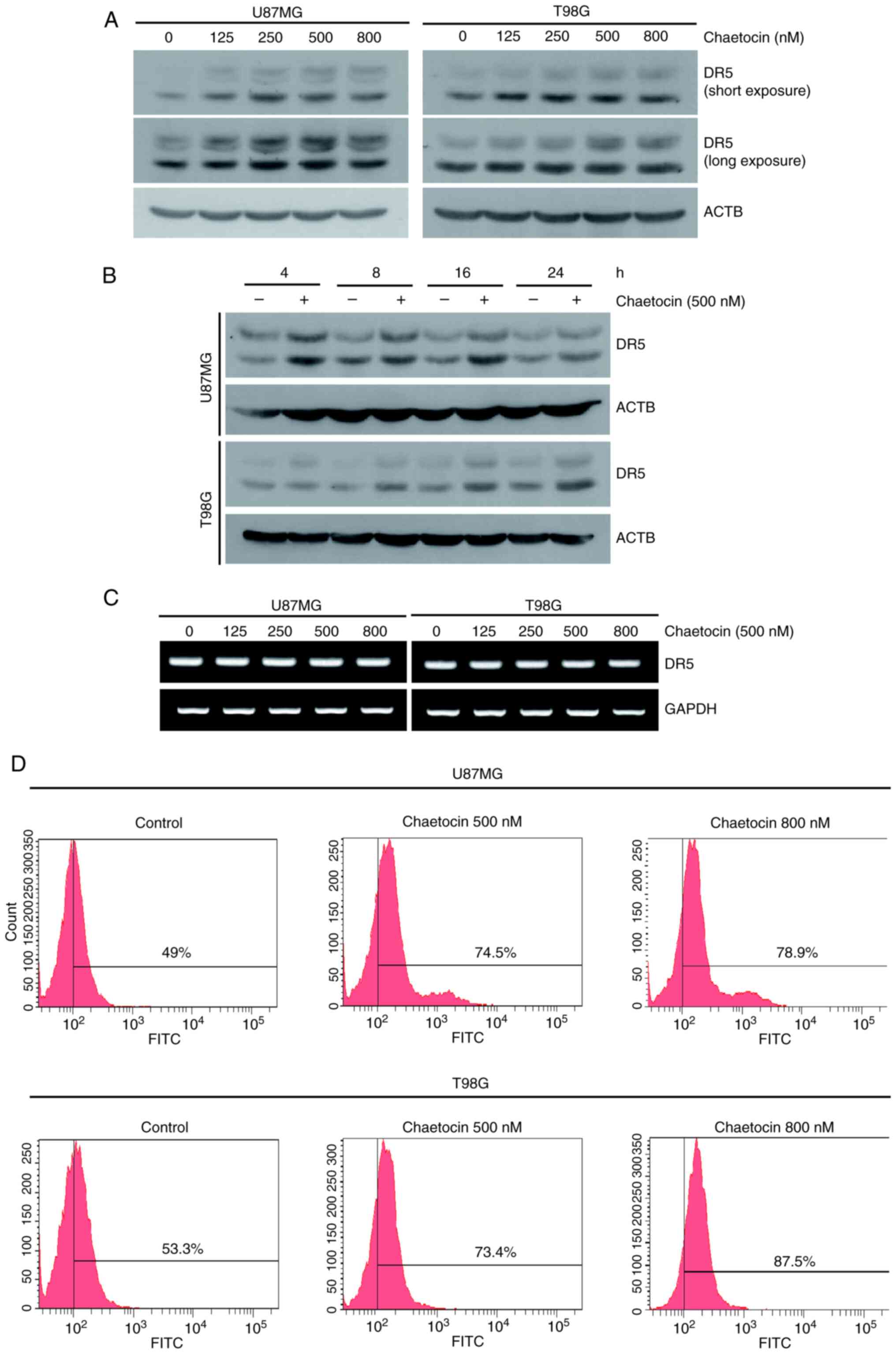
Chaetocin upregulates DR5 expression in
human glioblastoma cells by stabilizing the protein and inhibiting
proteasome activity
To clarify the molecular mechanisms of
chaetocin-mediated TRAIL sensitization, the present study
investigated the expression of the TRAIL receptor DR5. Chaetocin
treatment markedly upregulated DR5 expression in a dose- and
time-dependent manner (Figs. 3A and
B, and S3A and B), whereas
DR4 expression remained unchanged (Fig. S3C). Chaetocin did not
significantly affect DR5 mRNA expression, suggesting that chaetocin
upregulated DR5 expression post-transcriptionally (Fig. 3C). As increased DR5 surface
expression is essential for TRAIL-mediated cell death (18), the present study examined whether
chaetocin increased DR5 surface expression. Consistent with the
western blotting results, chaetocin increased DR5 protein levels on
the surface of U87MG and T98G cells (Fig. 3D). Taken together, these results
indicate that chaetocin upregulates DR5 expression.
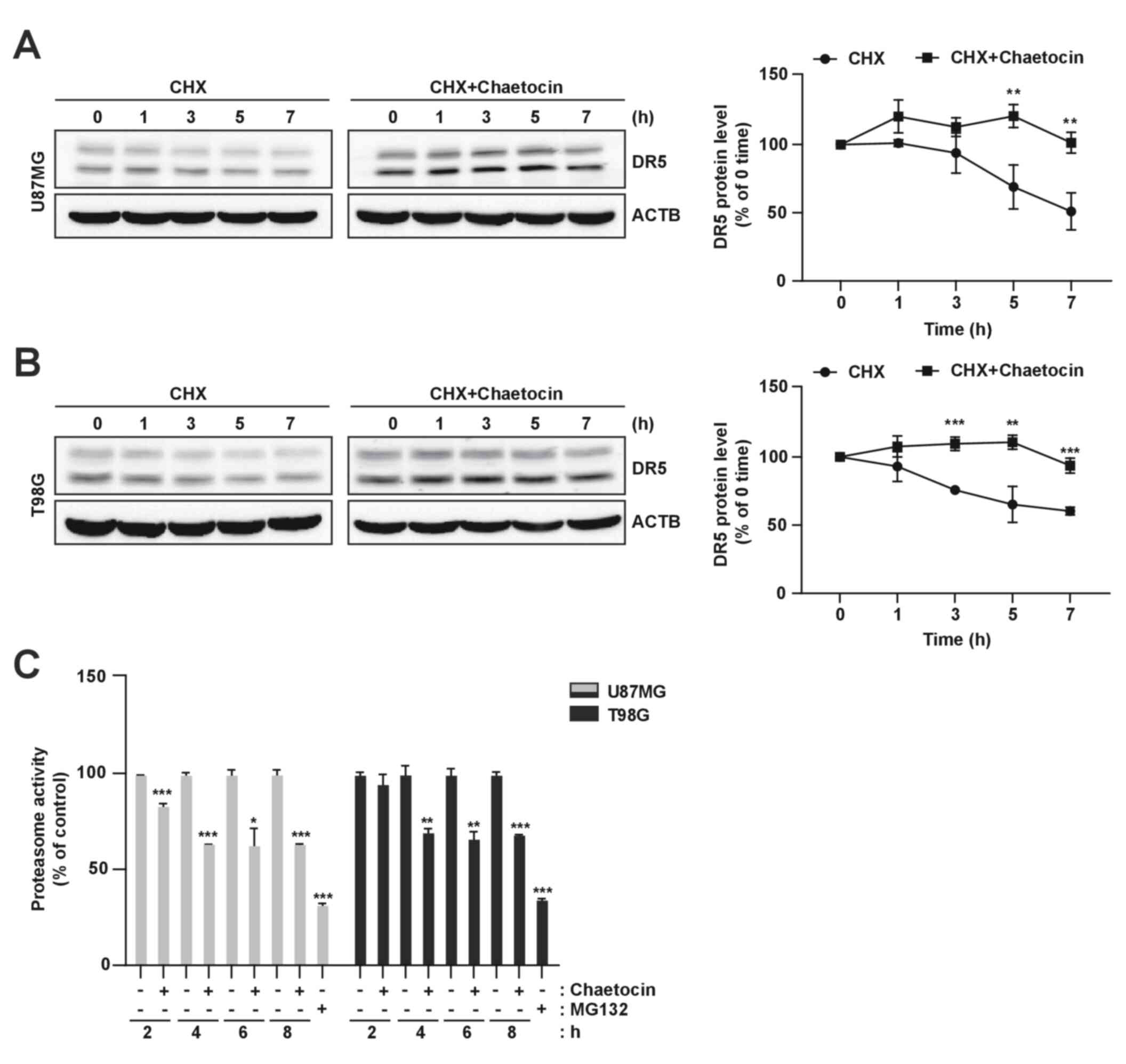
To determine the mechanism underlying
chaetocin-mediated DR5 upregulation, the present study investigated
the effect of chaetocin on DR5 protein stability in U87MG and T98G
cells. Following chaetocin treatment for 4 h, the cells were
exposed to cycloheximide (CHX), a de novo protein synthesis
inhibitor, in the presence or absence of chaetocin for 1-7 h. DR5
protein levels were determined using western blot analysis.
Although CHX resulted in a gradual decrease in DR5 expression, CHX
and chaetocin co-treatment sustained DR5 expression (Fig. 4A and B). Additionally, this study
investigated the involvement of the proteasome in chaetocin-induced
DR5 regulation and found that chaetocin reduced proteasome activity
(Fig. 4C). Overall, these results
indicated that chaetocin upregulates DR5 expression by increasing
the stability of DR5 protein.
Chaetocin enhances TRAIL-induced
apoptosis via DR5 upregulation
Glioblastoma cells were transfected with siRNA
targeting DR5 to investigate its functional role in
chaetocin/TRAIL-induced apoptosis. siRNA-mediated knockdown of DR5
effectively inhibited apoptosis induced by the combined treatment
of chaetocin and TRAIL in U87MG and T98G cells (Fig. 5A). Although chaetocin + TRAIL
treatment reduced the viability of cells transfected with scramble
by 36%, DR5 knockdown significantly restored cell viability to 87%
(Fig. 5B). DR5 knockdown reduced
the cleavage of caspase-3 and PARP induced by chaetocin + TRAIL
treatment in both U87MG and T98G cells (Figs. 5C and S4). Collectively, these findings
indicate that DR5 plays a crucial role in chaetocin/TRAIL-mediated
apoptosis in human glioblastoma cells.
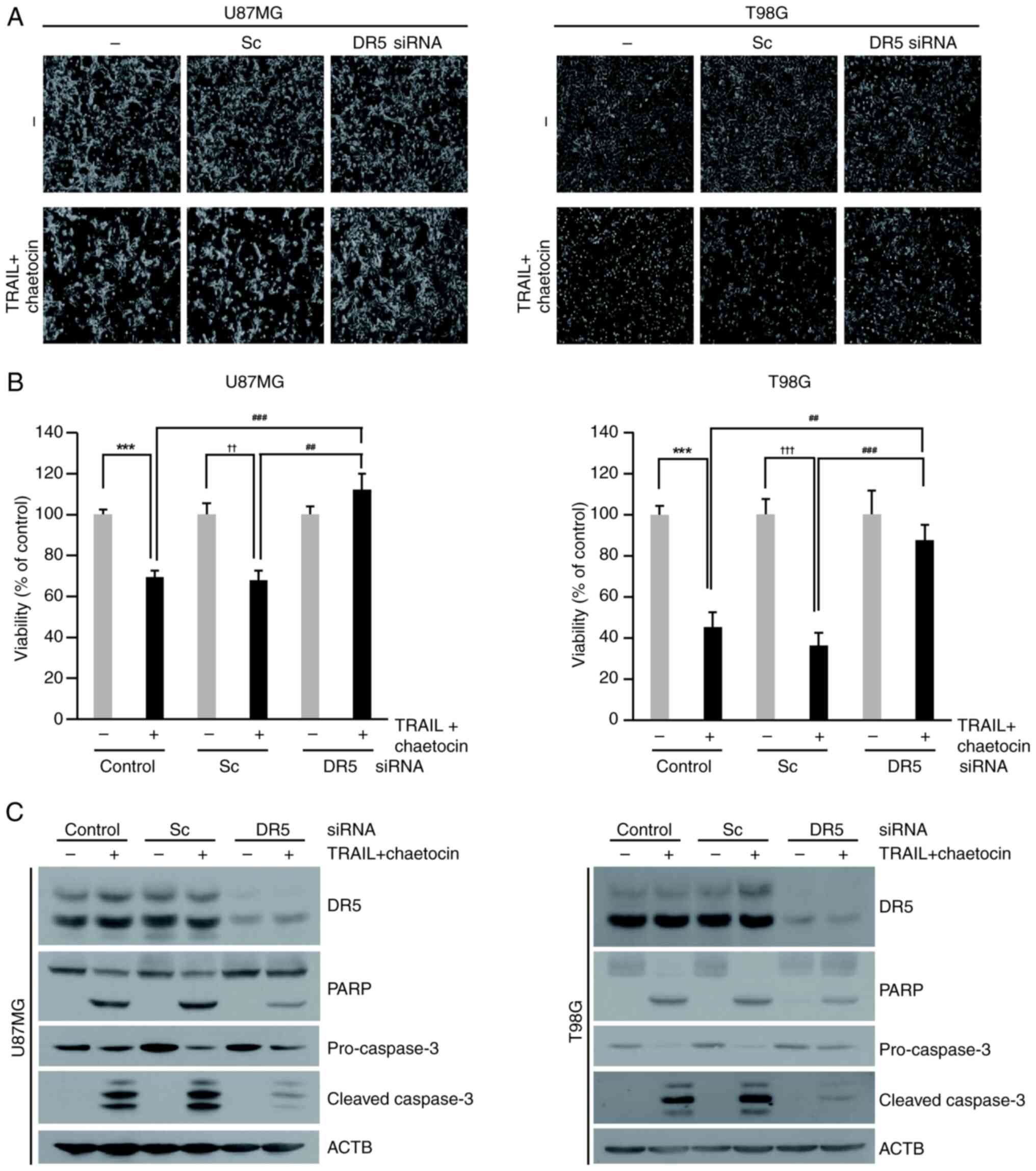 | Figure 5Blocking DR5 expression inhibits the
effect of chaetocin on TRAIL-mediated apoptosis. (A-C) Cells were
transfected with DR5 or Sc siRNA (50 nM) for 24 h and then treated
with chaetocin (500 nM) and TRAIL (50 ng/ml) for 6 h. (A) Cell
morphology was examined using interference light microscopy
(magnification, ×200). (B) U87MG and T98G cell viability. (C)
Western blot analysis was performed to determine the expression of
DR5 and apoptotic markers (PARP, cleaved caspase-3, and
pro-caspase-3). ***P<0.001 vs. control,
††P<0.05, †††P<0.001 vs. Sc,
##P<0.05, ###P<0.001 vs. TRAIL +
chaetocin. DR, death receptor; TRAIL, tumor necrosis factor-related
apoptosis-inducing ligand; Sc, scrambled control; si, small
interfering; ACTB, β-actin. |
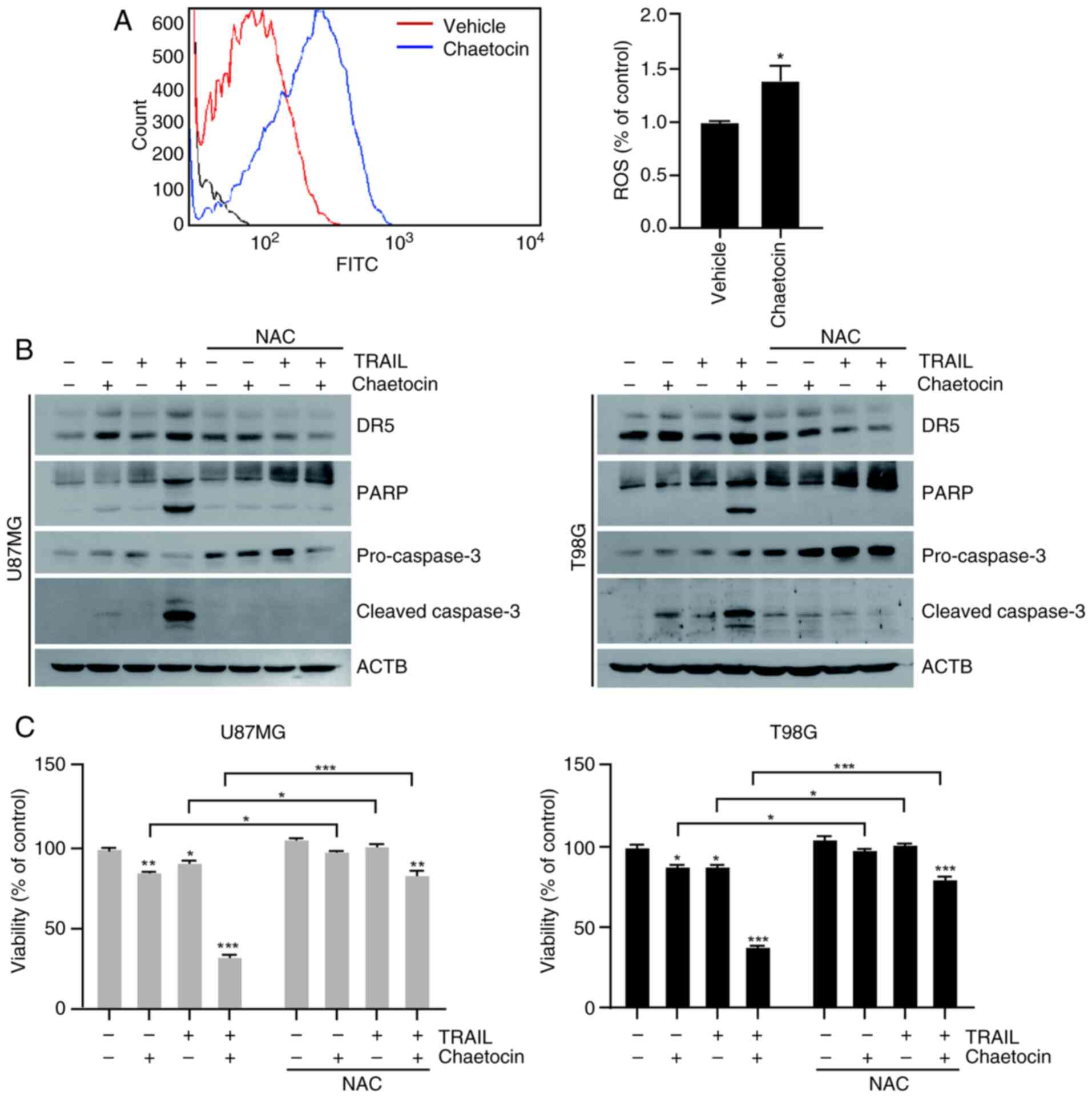
Chaetocin enhances TRAIL-mediated
apoptosis via ROS generation
ROS are key activators of DR5 upregulation, which
leads to TRAIL-mediated apoptosis (19,20). To investigate whether ROS are
required for chaetocin/TRAIL-induced apoptosis in U87MG and T98G
cells, the present study assessed the impact of chaetocin treatment
on ROS production. Chaetocin treatment for 10 min markedly
increased H2DCFDA fluorescence intensity in U87MG cells
(Fig. 6A), suggesting increased
ROS production. To confirm the role of ROS in
chaetocin/TRAIL-induced apoptosis, cells were pretreated with 5 mM
NAC, a ROS scavenger, prior to treatment with chaetocin and TRAIL
for 6 h. NAC pretreatment significantly decreased DR5 and cleaved
caspase-3 expression (Figs. 6B
and S5). Treatment with
chaetocin + TRAIL promoted cell death, while pretreatment with NAC
notably reduced this effect (Fig.
6C). Furthermore, p53 (21,22), CHOP (23,24), and NF-κB (25) may play a role in the upregulation
of DR5. Therefore, the present study investigated whether chaetocin
upregulated DR5 expression by increasing p53 and CHOP expression.
Chaetocin treatment did not significantly affect p53, CHOP, and Bip
expression in U87MG and T98G cells (Figs. S6A and B). Collectively, these
findings suggest that chaetocin-induced ROS generation was
essential for DR5 expression and contributed to sensitizing cells
to TRAIL-induced apoptosis.
Discussion
Our data demonstrate that the co-treatment of
chaetocin and TRAIL induces apoptosis in U87MG and T98G human
glioblastoma cells via chaetocin-induced ROS-dependent upregulation
of DR5. Notably, the present findings also highlight that chaetocin
plays a pivotal role in sustaining DR5 stability in these
cells.
Chaetocin, a fungal metabolite derived from species
of the genus Chaetomium, has a thiodioxopiperazine structure
(26). It exhibits potent
antiproliferative activity against solid tumors via apoptosis
induction. Chaetocin has been shown to induce ROS production,
JNK/c-Jun pathway activation and macrophage phagocytosis to trigger
caspase-dependent apoptosis in colorectal cancer cells (27). Additionally, chaetocin increases
apoptosis by enhancing ROS levels in OVCAR-3 ovarian cancer cells
(28). Han et al (13) reported that chaetocin induces
ROS-mediated apoptosis in human melanoma cells and suppresses the
growth of melanoma tumors in nude mice. Moreover, chaetocin
triggers not only apoptosis but also autophagy; it exerts
anticancer effects in hepatoma cell lines by inducing autophagy
(29). Despite these findings,
the anticancer effects of chaetocin in human glioblastoma and the
underlying mechanisms have not been fully elucidated.
TRAIL, a member of the TNF family, induces apoptosis
in various types of cancer cell but not in normal cells (3,30).
Notably, although TRAIL is regarded as a promising anticancer agent
due to its ability to selectively target cancer cells, several
cancer types including colon, breast, and hepatocellular carcinoma
- exhibit resistance to TRAIL-induced apoptosis (9-11).
Therefore, novel therapeutic strategies are required to sensitize
cancer cells to TRAIL-induced apoptosis. TRAIL functions by binding
to cell surface DRs, primarily DR4 and DR5, to initiate apoptotic
signaling (31). Additionally,
the strength and duration of TRAIL-induced apoptotic signaling are
influenced by expression of DRs (32). Notably, DR5 is upregulated by
silibinin (33), mitoxantrone
(34), and lovastatin (35) during TRAIL-mediated apoptosis in
glioblastoma cells. In the present study, chaetocin effectively
upregulated DR5 expression and the knockdown of DR5 significantly
inhibited chaetocin/TRAIL-induced apoptosis in both U87MG and T98G
cells.
The roles of DR4 and DR5 are critical for the
cytotoxic effects of TRAIL and other chemotherapeutic agents
(20,33,36,37). In the present study, chaetocin
treatment led to DR5 upregulation in a dose- and time-dependent
manner and increased DR5 surface expression on cells. Overall,
these results suggest that chaetocin-induced upregulation of DR5
may play an essential role in chaetocin- and TRAIL-induced
apoptosis. Although a previous study demonstrated that chaetocin
serves as an apoptosis-inducing agent and induces DR5-mediated cell
death in TRAIL-treated glioblastoma cells (38), the present study found that
chaetocin regulated expression and stability of DR5 in U87MG and
T98G cells. Additionally, further experiments using protein
synthesis inhibitor CHX showed that chaetocin did not affect the
mRNA expression of DR5 but affected its protein levels.
Chaetocin regulates SUV39H1 activity in an
ROS-dependent manner, influencing the expression of DR-related
genes and leading to DR-dependent apoptosis (14). Additionally, chaetocin promotes
ROS generation and upregulates antioxidant genes, such as heme
oxygenase 1, NAD(P)H quinone dehydrogenase 1, glutamate-cysteine
ligase modifier subunit and thioredoxin reductase 1 (39,40). In the present study, chaetocin
induced ROS generation, whereas NAC pretreatment inhibited
chaetocin-induced ROS production. Collectively, these results
revealed the key role of ROS in chaetocin-induced DR5 expression.
Furthermore, the present data emphasize the interaction between
TRAIL-mediated signaling and oxidative stress response, with
previous studies demonstrating that ROS production enhances DR5
expression in human carcinoma cell lines (41-43). Notably, compounds such as
baicalein (42) and vitisin A
(43) sensitize prostate cancer
cells to TRAIL by promoting ROS generation and upregulating
DR5.
The combination of chaetocin and TRAIL has clinical
relevance for glioblastoma treatment by addressing key challenges
in current therapeutic strategies. This approach selectively
induces apoptosis in glioblastoma cells while sparing normal cells,
highlighting its potential safety and specificity. Mechanistically,
chaetocin enhances TRAIL sensitivity through ROS-mediated DR5
stabilization, offering a basis for personalized therapy using
biomarkers including ROS or DR5. In future, in vivo
experiments will be essential to validate the findings of in
vitro study and assess the effects of chaetocin and TRAIL
co-treatment on glioblastoma tumor growth to determine whether
chaetocin enhances TRAIL-mediated apoptosis and tumor suppression
in glioblastoma, further highlighting its potential as a
therapeutic agent for glioblastoma treatment. Its integration with
standard treatments such as radiotherapy and temozolomide may
provide synergistic benefits, particularly in treatment-resistant
cases. Although further preclinical studies are needed, the dual
role of chaetocin as an epigenetic modifier and apoptosis
sensitizer positions it as a promising candidate for clinical
trials. In conclusion, chaetocin enhances TRAIL-induced apoptosis
in human glioblastoma cells by increasing DR5 stability, promoting
ROS production and upregulating DR5 expression. Overall, the
present study clarified the mechanism by which chaetocin enhances
cancer cell sensitivity to TRAIL and offers potential therapeutic
strategies for cancer treatment.
Supplementary Data
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
HJJ wrote the manuscript, performed experiments and
analyzed data. JKK wrote the manuscript and performed experiments.
SIS performed the literature review. WKB conceived and supervised
the study. HJJ and WKB confirm the authenticity of all the raw
data. All authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
This work was supported by a National Research Foundation of
Korea (NRF) Sejong Science fellowship (NRF-2022R1C1C2010228) and by
NRF grants funded by the Korea government [MSIP] (2016R1A2B101205
and 2018R1A2B5A01023660).
References
|
1
|
Hanif F, Muzaffar K, Perveen K, Malhi SM
and Simjee ShU: Glioblastoma multiforme: A review of its
epidemiology and pathogenesis through clinical presentation and
treatment. Asian Pac J Cancer Prev. 18:3–9. 2017.PubMed/NCBI
|
|
2
|
Shergalis A, Bankhead A III, Luesakul U,
Muangsin N and Neamati N: Current challenges and opportunities in
treating glioblastoma. Pharmacol Rev. 70:412–445. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dai X, Zhang J, Arfuso F, Chinnathambi A,
Zayed ME, Alharbi SA, Kumar AP, Ahn KS and Sethi G: Targeting
TNF-related apoptosis-inducing ligand (TRAIL) receptor by natural
products as a potential therapeutic approach for cancer therapy.
Exp Biol Med (Maywood). 240:760–773. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bin L, Thorburn J, Thomas LR, Clark PE,
Humphreys R and Thorburn A: Tumor-derived mutations in the TRAIL
receptor DR5 inhibit TRAIL signaling through the DR4 receptor by
competing for ligand binding. J Biol Chem. 282:28189–28194. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pimentel JM, Zhou JY and Wu GS: The role
of TRAIL in apoptosis and immunosurveillance in cancer. Cancers
(Basel). 15:27522023. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wachmann K, Pop C, van Raam BJ, Drag M,
Mace PD, Snipas SJ, Zmasek C, Schwarzenbacher R, Salvesen GS and
Riedl SJ: Activation and specificity of human caspase-10.
Biochemistry. 49:8307–8315. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Deng Y, Lin Y and Wu X: TRAIL-induced
apoptosis requires Bax-dependent mitochondrial release of
Smac/Diablo. Genes Dev. 16:33–45. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Meurette O, Rebillard A, Huc L, Le Moigne
G, Merino D, Micheau O, Lagadic-Gossmann D and Dimanche-Boitrel MT:
TRAIL induces receptor-interacting protein 1-dependent and
caspase-dependent necrosis-like cell death under acidic
extracellular conditions. Cancer Res. 67:218–226. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kim B, Seo JH, Lee KY and Park B: Icariin
sensitizes human colon cancer cells to TRAIL-induced apoptosis via
ERK-mediated upregulation of death receptors. Int J Oncol.
56:821–834. 2020.PubMed/NCBI
|
|
10
|
Manouchehri JM, Turner KA and Kalafatis M:
TRAIL-induced apoptosis in TRAIL-resistant breast carcinoma through
quercetin cotreatment. Breast Cancer (Auckl).
12:11782234177498552018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu S, Qiu J, He G, He W, Liu C, Cai D and
Pan H: TRAIL promotes hepatocellular carcinoma apoptosis and
inhibits proliferation and migration via interacting with IER3.
Cancer Cell Int. 21:632021. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Greiner D, Bonaldi T, Eskeland R, Roemer E
and Imhof A: Identification of a specific inhibitor of the histone
methyltransferase SU(VAR)3-9. Nat Chem Biol. 1:143–145. 2005.
View Article : Google Scholar
|
|
13
|
Han X, Han Y, Zheng Y, Sun Q, Ma T, Zhang
J and Xu L: Chaetocin induces apoptosis in human melanoma cells
through the generation of reactive oxygen species and the intrinsic
mitochondrial pathway, and exerts its anti-tumor activity in vivo.
PLoS One. 12:e01759502017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chaib H, Nebbioso A, Prebet T, Castellano
R, Garbit S, Restouin A, Vey N, Altucci L and Collette Y:
Anti-leukemia activity of chaetocin via death receptor-dependent
apoptosis and dual modulation of the histone methyl-transferase
SUV39H1. Leukemia. 26:662–674. 2012. View Article : Google Scholar
|
|
15
|
Liu X, Guo S, Liu X and Su L: Chaetocin
induces endoplasmic reticulum stress response and leads to death
receptor 5-dependent apoptosis in human non-small cell lung cancer
cells. Apoptosis. 20:1499–1507. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jung HJ, Seo I, Jha BK, Suh SI and Baek
WK: Miconazole induces autophagic death in glioblastoma cells via
reactive oxygen species-mediated endoplasmic reticulum stress.
Oncol Lett. 21:3352021. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
18
|
Yamamoto K, Makino M, Watanapokasin R,
Tashiro E and Imoto M: Inostamycin enhanced TRAIL-induced apoptosis
through DR5 upregulation on the cell surface. J Antibiot (Tokyo).
65:295–300. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dilshara MG, Jayasooriya RGPT, Molagoda
IMN, Jeong JW, Lee S, Park SR, Kim GY and Choi YH: Silibinin
sensitizes TRAIL-mediated apoptosis by upregulating DR5 through
ROS-induced endoplasmic reticulum stress-Ca2+-CaMKII-Sp1
pathway. Oncotarget. 9:10324–10342. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yodkeeree S, Sung B, Limtrakul P and
Aggarwal BB: Zerumbone enhances TRAIL-induced apoptosis through the
induction of death receptors in human colon cancer cells: Evidence
for an essential role of reactive oxygen species. Cancer Res.
69:6581–6589. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hori T, Kondo T, Kanamori M, Tabuchi Y,
Ogawa R, Zhao QL, Ahmed K, Yasuda T, Seki S, Suzuki K and Kimura T:
Nutlin-3 enhances tumor necrosis factor-related apoptosis-inducing
ligand (TRAIL)-induced apoptosis through up-regulation of death
receptor 5 (DR5) in human sarcoma HOS cells and human colon cancer
HCT116 cells. Cancer Lett. 287:98–108. 2010. View Article : Google Scholar
|
|
22
|
Wang W and El-Deiry WS: Requirement of p53
targets in chemosensitization of colonic carcinoma to death ligand
therapy. Proc Natl Acad Sci USA. 9:15095–15100. 2003. View Article : Google Scholar
|
|
23
|
Lu M, Lawrence DA, Marsters S,
Acosta-Alvear D, Kimmig P, Mendez AS, Paton AW, Paton JC, Walter P
and Ashkenazi A: Opposing unfolded-protein-response signals
converge on death receptor 5 to control apoptosis. Science.
345:98–101. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yue D and Sun X: Ixazomib promotes
CHOP-dependent DR5 induction and apoptosis in colorectal cancer
cells. Cancer Biol Ther. 20:284–294. 2019. View Article : Google Scholar :
|
|
25
|
Kong F, You H, Zhao J, Liu W, Hu L, Luo W,
Hu W, Tang R and Zheng K: The enhanced expression of death receptor
5 (DR5) mediated by HBV X protein through NF-kappaB pathway is
associated with cell apoptosis induced by (TNF-α related apoptosis
inducing ligand) TRAIL in hepatoma cells. Virol J. 12:1922015.
View Article : Google Scholar
|
|
26
|
Sekita S, Yoshihira K, Natori S, Udagawa
S, Muroi T, Sugiyama Y, Kurata H and Umeda M: Mycotoxin production
by Chaetomium spp and related fungi. Can J Microbiol. 27:766–772.
1981. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang H, Wen C, Chen S, Li W, Qin Q, He L,
Wang F, Chen J, Ye W, Li W, et al: ROS/JNK/C-jun pathway is
involved in chaetocin induced colorectal cancer cells apoptosis and
macrophage phagocytosis enhancement. Front Pharmacol.
12:7293672021. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li Z, Huang L, Wei L, Hou Z, Ye W and
Huang S: Chaetocin induces caspase-dependent apoptosis in ovarian
cancer cells via the generation of reactive oxygen species. Oncol
Lett. 18:1915–1921. 2019.PubMed/NCBI
|
|
29
|
Jung HJ, Seo I, Casciello F, Jacquelin S,
Lane SW, Suh SI, Suh MH, Lee JS and Baek WK: The anticancer effect
of chaetocin is enhanced by inhibition of autophagy. Cell Death
Dis. 7:e20982016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Holoch PA and Griffith TS: TNF-related
apoptosis-inducing ligand (TRAIL): A new path to anti-cancer
therapies. Eur J Pharmacol. 625:63–72. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
LeBlanc HN and Ashkenazi A: Apo2L/TRAIL
and its death and decoy receptors. Cell Death Differ. 10:66–75.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang S and El-Deiry WS: TRAIL and
apoptosis induction by TNF-family death receptors. Oncogene.
22:8628–8633. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Son YG, Kim EH, Kim JY, Kim SU, Kwon TK,
Yoon AR, Yun CO and Choi KS: Silibinin sensitizes human glioma
cells to TRAIL-mediated apoptosis via DR5 up-regulation and
down-regulation of c-FLIP and survivin. Cancer Res. 67:8274–8284.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Senbabaoglu F, Cingoz A, Kaya E,
Kazancioglu S, Lack NA, Acilan C and Bagci-Onder T: Identification
of mitoxantrone as a TRAIL-sensitizing agent for glioblastoma
multiforme. Cancer Biol Ther. 17:546–557. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu PC, Lu G, Deng Y, Wang CD, Su XW, Zhou
JY, Chan TM, Hu X and Poon WS: Inhibition of NF-κB pathway and
modulation of MAPK signaling pathways in glioblastoma and
implications for lovastatin and tumor necrosis factor-related
apoptosis inducing ligand (TRAIL) combination therapy. PLoS One.
12:e01711572017. View Article : Google Scholar
|
|
36
|
Аrtykov АА, Belov DA, Shipunova VO,
Trushina DB, Deyev SM, Dolgikh DA, Kirpichnikov MP and Gasparian
ME: Chemotherapeutic agents sensitize resistant cancer cells to the
DR5-specific variant DR5-B more efficiently than to TRAIL by
modulating the surface expression of death and decoy receptors.
Cancers (Basel). 12:11292020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gibson SB, Oyer R, Spalding AC, Anderson
SM and Johnson GL: Increased expression of death receptors 4 and 5
synergizes the apoptosis response to combined treatment with
etoposide and TRAIL. Mol Cell Biol. 20:205–212. 2000. View Article : Google Scholar
|
|
38
|
Ozyerli-Goknar E, Sur-Erdem I, Seker F,
Cingöz A, Kayabolen A, Kahya-Yesil Z, Uyulur F, Gezen M, Tolay N,
Erman B, et al: The fungal metabolite chaetocin is a sensitizer for
pro-apoptotic therapies in glioblastoma. Cell Death Dis.
10:8942019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mandal PK, Schneider M, Kölle P,
Kuhlencordt P, Förster H, Beck H, Bornkamm GW and Conrad M: Loss of
thioredoxin reductase 1 renders tumors highly susceptible to
pharmacologic glutathione deprivation. Cancer Res. 70:9505–9514.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sporn MB and Liby KT: NRF2 and cancer: The
good, the bad and the importance of context. Nat Rev Cancer.
12:564–571. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yamaguchi H and Wang HG: CHOP is involved
in endoplasmic reticulum stress-induced apoptosis by enhancing DR5
expression in human carcinoma cells. J Biol Chem. 279:45495–45502.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Taniguchi H, Yoshida T, Horinaka M, Yasuda
T, Goda AE, Konishi M, Wakada M, Kataoka K, Yoshikawa T and Sakai
T: Baicalein overcomes tumor necrosis factor-related
apoptosis-inducing ligand resistance via two different
cell-specific pathways in cancer cells but not in normal cells.
Cancer Res. 68:8918–8927. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Shin D, Kwon HY, Sohn EJ, Nam MS, Kim JH,
Lee JC, Ryu SY, Park B and Kim SH: Upregulation of death receptor 5
and production of reactive oxygen species mediate sensitization of
PC-3 prostate cancer cells to TRAIL induced apoptosis by vitisin A.
Cell Physiol Biochem. 36:1151–1162. 2015. View Article : Google Scholar : PubMed/NCBI
|