Introduction
First identified in mammalian cells in 1960, snoRNAs
are recognized as pivotal, naturally occurring guide RNAs.
Typically ranging from 60 to 300 nucleotides in length, they
function within ribonucleoprotein (RNP) complexes to precisely
target specific cellular RNAs. While the human genome harbors over
1,000 annotated snoRNA genes, a striking 80% of these remain
functionally enigmatic (1,2).
Accumulating evidence indicates that specific
snoRNAs exert direct control over gene expression by governing mRNA
stability, editing, and splicing-mechanisms that may operate
autonomously from canonical RNA modification pathways (3,4).
Notably, >50 snoRNAs have been implicated in dysregulation
across over 12 cancer types, although the precise mechanisms
underlying disease pathogenesis due to snoRNAs deletions or
mutations are still largely elusive, primarily owing to limitations
in current methodologies for comprehensive identification of snoRNA
targets and their transcriptome-wide modification profiles
(5).
In addition to their canonical forms, snoRNAs can
undergo specific processing to generate smaller fragments ranging
from 16 to 36 nucleotides, known as snoRNA-derived fragments
(sdRNAs). First identified in 2008, sdRNAs represent a relatively
unexplored class of small non-coding RNAs (ncRNAs), with their
functional roles and regulatory mechanisms still largely undefined
(6). These small fragments have
emerged as functional molecules with regulatory roles distinct from
their parental snoRNAs (7).
While several notable reviews have previously
explored the biology of snoRNAs, many have predominantly focused on
their canonical roles in ribosome biogenesis or provided
generalized observational overviews of their dysregulation in
specific cancer types. Furthermore, within existing literature,
sdRNAs are frequently relegated to brief mentions rather than being
critically evaluated as distinct, functionally significant
regulatory entities. The present review extended beyond these
previous works by applying a rigorous 'three-tier evidence
framework' to systematically evaluate the clinical readiness of
both snoRNAs and sdRNAs. Current knowledge was strictly stratified
into fundamental mechanistic drivers (tier 1), retrospective
tissue-based prognostic associations (tier 2), and highly
actionable translational applications, such as liquid biopsies and
nucleic acid therapeutics (NATs) (tier 3). By critically
synthesizing these distinct levels of evidence and placing an
unprecedented structural emphasis on the rapidly evolving sdRNA
landscape, the present review provided a uniquely structured,
translational roadmap for integrating these ncRNAs into precision
oncology.
The article is a narrative review intended to
provide an updated overview of the biogenesis, functions and
clinical relevance of snoRNAs and sdRNAs in cancer. Rather than
performing a formal systematic review or meta-analysis,
representative original studies and recent review articles were
qualitatively synthesized, with an emphasis on mechanistic
insights, biomarker potential, liquid biopsy applications and
therapeutic targeting. Furthermore, their emerging potential as
diagnostic biomarkers and therapeutic targets in cancer was
emphasized, providing novel perspectives for innovative strategies
in cancer diagnosis and treatment. By integrating current knowledge
and highlighting critical gaps in the field, the present review
sought to provide a foundation for future research directions in
snoRNA and sdRNA biology and their applications in oncology.
Biogenesis
snoRNAs, primarily originating from intronic
sequences, commonly reside within genes that encode proteins vital
for ribosome biogenesis and function. This genomic arrangement
facilitates the synchronized expression of ribosomal components
alongside their associated snoRNAs, thereby preserving cellular
homeostasis (8,9). Notably, a subset of snoRNAs
originates from introns of non-coding genomic regions, such as
growth arrest-specific 5 (GAS5) and snoRNA host gene (SNHG)1, which
produce distinct sets of eleven and nine snoRNAs, respectively
(10,11).
The stability of intronically encoded snoRNAs is
maintained through their cotranscriptional association with RNPs,
which protect them from exonucleolytic degradation (12). These RNP complexes, along with
auxiliary factors including nuclear assembly factor 1, sno/sca
RNA-associated H/ACA assembly factor 1 homolog, and nuclear
factor-like protein, ensure proper processing, stability, and
nucleolar localization of snoRNAs (13). Following synthesis, snoRNPs are
transported to Cajal bodies for further maturation before
localizing to nucleoli, where they modify ncRNAs (14). Notably, the transcriptional
regulation of snoRNAs varies across species. In plants and yeast,
certain snoRNAs are transcribed in a polycistronic manner, whereas
in the case of GAS5, snoRNAs are interspersed with exons.
Furthermore, yeast exhibit a unique mechanism in which some snoRNAs
are transcribed by RNA polymerase II, highlighting the evolutionary
diversity in snoRNA transcription and processing mechanisms among
different organisms (15).
snoRNA classes and functions
snoRNA classes and canonical
function
snoRNAs play a central role in post-transcriptional
modifications, primarily guiding 2'-O-ribose methylation and
pseudouridylation of ribosomal RNAs (rRNAs) and small nuclear RNAs
(snRNAs). These modifications enhance molecular interactions,
promote rRNA folding, and ultimately optimize ribosome and
spliceosome function (16,17).
snoRNAs function as guide RNAs within snoRNP complexes, which
include core proteins that provide catalytic activity and ensure
target specificity via complementary base pairing (18-20).
snoRNAs are classified into 2 main groups: C/D box
snoRNAs (SNORDs) and H/ACA box snoRNAs (SNORAs), which direct
methylation and pseudouridylation, respectively. In addition to
these, a third category, small Cajal body-specific RNAs (scaRNAs),
exhibits hybrid features of both SNORDs and SNORAs, combining their
functional properties to perform specialized roles in RNA
modification. This diversity in snoRNA classes underscores their
versatility and importance in RNA processing and cellular function
(21). Characterization,
biogenesis and canonical function of snoRNAs are shown in Fig. 1.
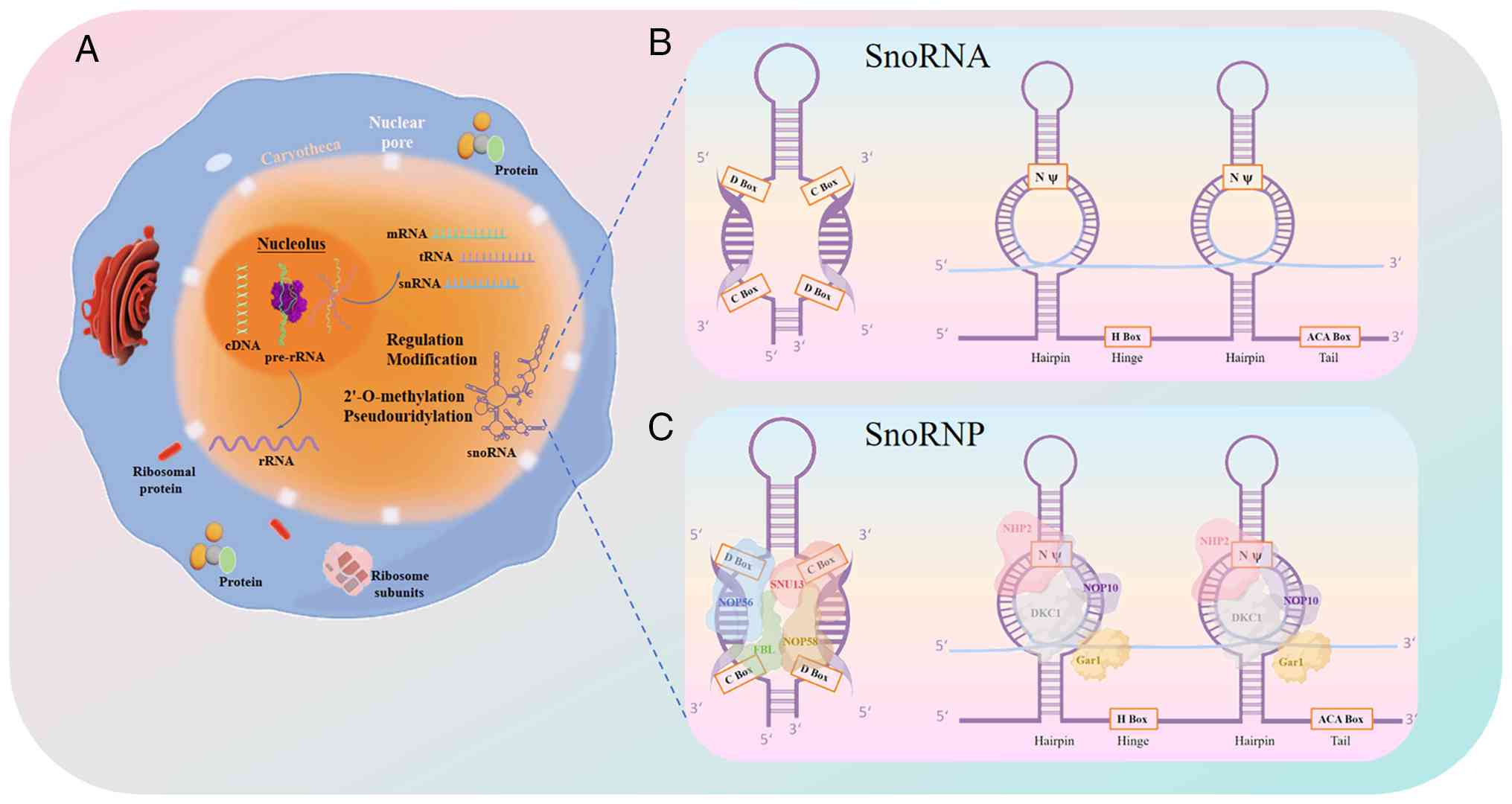 | Figure 1Characterization, biogenesis and
canonical function of snoRNAs. (A) Genomic organization of snoRNAs:
Intronic snoRNAs are processed from host gene transcripts, while
some snoRNAs are transcribed from independent promoters. (B)
Structural features of C/D box and H/ACA box snoRNAs and canonical
functions: Showing conserved motifs (C/D boxes, H/ACA domains) and
core binding proteins (fibrillarin and dyskerin). C/D box snoRNAs
guide 2'-O-methylation of rRNA and snRNA; H/ACA box snoRNAs guide
pseudouridylation. (C) Biogenesis pathway: snoRNAs associate with
core proteins to form snoRNPs, are transported to Cajal bodies for
maturation, and localize to nucleoli. snoRNA, small nucleolar RNA;
rRNA, ribosomal RNA; snRNA, small nuclear RNA; snoRNP, small
nucleolar ribonucleoprotein; mRNA, messenger RNA; tRNA transfer
RNA. |
SNORDs
SNORDs are characterized by conserved C (RUGAUGA)
and D (CUGA) motifs near their termini, forming a kink-turn
structure. They also contain internal C' and D' boxes. These
structural elements scaffold proteins such as nucleolar protein
(NOP)56, NOP58, 15.5 kilodalton protein, and fibrillarin, which are
essential for the stability, localization, and functional integrity
of snoRNPs.
The target specificity of SNORDs is mediated by a
short guide region, typically 10-21 nucleotides in length, located
upstream of the D or D' boxes. This region, referred to as the
antisense element, plays a critical role in recognizing and binding
to complementary sequences on target RNAs. Fibrillarin, a core
component of the snoRNP complex, serves as the catalytic force for
methylating the fifth nucleotide upstream of the D or D' box motif,
thus fulfilling the essential modification role of snoRNP. This
precise targeting mechanism ensures the accurate and site-specific
2'-O-methylation of rRNAs and snRNAs, highlighting the
sophisticated regulatory role of SNORDs in RNA processing and
modification (22-24).
SNORAs
SNORAs are characterized by a 3' ACA tail and 2
hairpin structures connected by a hinge region containing a
conserved (ANANNA) motif, known as the H motif or BOX H. Each
hairpin contains an internal loop, termed the pseudouridylation
pocket, which base-pairs with target RNAs. Typically, the uridine
residue in the target RNA, positioned 14-15 nucleotides upstream of
the H or ACA box, undergoes pseudouridylation. This reaction is
catalyzed by dyskerin, a pseudouridine synthase within the
H/ACA-associated snRNP complex, which also includes nuclear H/ACA
ribonucleoprotein 2 (NHP2), NOP10, and glycine-argine rich protein
1 (25-27).
scaRNAs
scaRNAs are distinguished by their localization
within Cajal bodies, specialized sub-nuclear compartments, mediated
by a unique UGAG motif. These RNAs play critical roles in
spliceosome and ribosome biogenesis, facilitating the precise
methylation and pseudouridylation of snRNAs and rRNAs. By guiding
these modifications, scaRNAs ensure the structural and functional
integrity of these essential molecular complexes, underscoring
their importance in cellular processes (28,29).
Noncanonical snoRNA functions
snoRNAs constitute a diverse and functionally
versatile class of RNA molecules with multifaceted roles in
cellular processes. Notably, approximately half of human snoRNAs
are 'orphans' with uncharacterized functions (30). Recent studies have revealed
diverse noncanonical roles beyond rRNA/snRNA modification,
expanding our understanding of their roles beyond traditional
paradigms and revealing previously unexplored biological activities
(31,32).
Additional modifications of rRNAs and
snRNAs
Emerging research indicates that snoRNAs may possess
an expanded functional repertoire beyond prior recognition,
encompassing diverse modification types and a broader spectrum of
RNA targets. For instance, the orphan snoRNAs snR4 and snR45
facilitate the acetylation of two N4 cytidine residues in 18S rRNA
by guiding the cytidine acetyltransferase Kre33 to its target
sites, a process critical for maintaining the accuracy of protein
synthesis (33). Advanced
techniques such as chimeric enhanced crosslinking and
immunoprecipitation have uncovered novel snoRNA-rRNA/snRNA
interactions in an RNA-binding protein-dependent manner, revealing
novel interactions and regulatory roles in RNA biogenesis (34).
Interaction with transfer RNAs
(tRNAs)
Post-transcriptional modifications of tRNAs,
especially at the anticodon wobble position and the 3' end, play a
vital role in stabilizing mRNA-tRNA interactions. For example, the
methylation of cytidine at the wobble position, guided by SNORD97,
targets the anticodon loop of elongator tRNAMet through sequence
complementarity, enhancing tRNA functionality and protecting it
from cleavage by stress-induced endonucleases such as angiogenin
(35). Although recent studies
have illuminated specific snoRNA-tRNA interactions (36,37), earlier co-immunoprecipitation
experiments had already revealed physical associations between
snoRNAs and tRNAs (38,39). This suggests that numerous
additional interactions remain to be discovered, highlighting the
profound complexity of snoRNA-mediated regulatory networks.
Interaction with mRNAs
Beyond their well-documented roles in ncRNA
modification, high-throughput sequencing of ligated RNAs has
uncovered surprising interactions between snoRNAs and mRNAs. A
prominent example is SNORD83B, an orphan snoRNA that bolsters the
stability of mRNAs-including NOP14, ribosomal protein S5, and
serine/arginine-rich splicing factor 3 via direct base-pairing
interactions (40). Research
reveals that SNORA73 forms stable secondary structures with target
mRNAs via non-canonical RNA binding sequences. Additionally,
SNORA73 engages with 7SL RNA, a crucial element of the signal
recognition particle (SRP), assembling a 'mRNA-snoRNA-7SL RNA'
ternary complex. This complex significantly boosts the binding
affinity of target mRNAs to the SRP, facilitating the efficient
translocation of the translational complex to the endoplasmic
reticulum. Thereby, this interaction promotes the effective
translocation and release of secretory proteins, revealing a novel
role for snoRNAs in orchestrating protein trafficking and secretion
(41). These findings highlight
the wide-ranging and multifaceted roles of snoRNAs in cellular
processes, extending well beyond their canonical functions in RNA
modification.
Interaction with long ncRNAs
(lncRNAs)
Research has revealed that SNORD80, an
intron-encoded SNORD originating from the GAS5 gene, orchestrates
the 2'-O-methylation of GAS5 lncRNA. This critical modification
bolsters the stability of the lncRNA, triggering its subsequent
upregulation and ultimately fine-tuning cellular stress response
mechanisms (42).
Interaction with proteins
Some snoRNAs function by binding proteins. SNORA13
departs from canonical snoRNA function by regulating cellular
senescence independently of ribosomal interactions. Rather than
modulating ribosomal translation, it directly binds MDM2, thereby
activating the p53 pathway and inducing senescence (30). By contrast, SNORD46 governs
metabolism and immune function through its specific interaction
with interleukin (IL)-15. While wild-type SNORD46 binds IL-15 via
G11, the G11A mutation significantly enhances this binding affinity
and promotes obesity in knockin mice. Mechanistically, SNORD46
inhibits IL-15-dependent phosphorylation of CD36 and monoglyceride
lipase in adipocytes-mediated by feline sarcoma-related kinase,
effectively suppressing lipolysis and thermogenic browning.
Concurrently, in natural killer (NK) cells, SNORD46 disrupts
IL-15-driven autophagy, reducing NK cell viability during obesity.
Notably, SNORD46 inhibitors effectively counteract these effects,
alleviating obesity and restoring critical NK cell functions,
including the antitumor efficacy of chimeric antigen receptor-NK
therapy (43). Similarly, the
snoRNA snR107 plays a distinct regulatory role in meiosis by
interacting with the RNA-binding proteins meiosis mitotic inhibitor
1 and meiosis 2. This interaction establishes a reciprocal
inhibition mechanism that tightly controls meiotic gene expression
throughout sexual differentiation (44). SNORA73 exhibits a unique mechanism
involving 5' end non-canonical structural binding to
poly(ADP-ribose) polymerase 1 (PARP1), inhibition of PARP1
auto-PARylation, and formation of a specialized snoRNP complex with
dyskerin pseudouridine synthase 1/NHP2 to modulate cancer genome
stability (45).
Regulation of alternative splicing
snoRNAs have surfaced as pivotal regulators of
alternative splicing, critically influencing the processing of key
mRNAs such as 5-hydroxytryptamine serotonin receptor 2c (HTR2C).
This intricate regulation is orchestrated by the orphan snoRNA
SNORD115, which harbors an 18-nucleotide sequence perfectly
complementary to the alternative exon 5b within HTR2C mRNA.
SNORD115 actively promotes the inclusion of exon 5b, yielding a
long, functional receptor isoform; conversely, its exclusion
produces a non-functional variant. Beyond its splicing role,
SNORD115 directly competes with adenosine deaminases acting on RNA
(ADAR) enzymes, which catalyze crucial adenosine-to-inosine editing
within this same region. Such editing can markedly alter three
amino acids in exon 5, profoundly impacting G-protein coupling and
downstream signaling cascades of the receptor. The influence of
SNORD115 extends to at least five additional alternative splicing
targets, powerfully underscoring its notable regulatory versatility
(46). Similarly, SNORD116,
residing at the same chromosomal locus as SNORD115, is implicated
in splicing regulation, with numerous predicted binding sites
precisely mapping to exon junctions (47). Other snoRNAs, such as SNORD27,
demonstrably direct the splicing of the E2F7 transcription factor
(4), while SNORD88C regulates the
alternative splicing of fibroblast growth factor receptor 3 (FGFR3)
by effectively masking cryptic splice sites, thereby preventing
aberrant exon inclusion (48).
These compelling findings illuminate the rapidly expanding role of
snoRNAs in meticulously fine-tuning mRNA splicing and their
profound potential impact on gene expression and cellular
function.
Involvement in 3'-end processing
Beyond their well-established roles in
post-transcriptional modifications and alternative splicing
regulation, snoRNAs are gaining growing recognition for their
involvement in mRNA 3'-end processing. Previous research reveals
that a specific subset of snoRNAs interacts directly with the
mammalian 3'-end processing complex, illuminating their function in
poly(A) site selection. These snoRNAs specifically associate with
factor interacting with poly(A) polymerase 1 (Fip1), a key cleavage
and polyadenylation specificity factor component. Among them,
SNORD50A has emerged as a pivotal regulator, inhibiting 3' mRNA
processing for specific transcripts by occupying the interaction
site between Fip1 and the poly(A) signal (3).
Extranuclear functions
While snoRNAs are traditionally linked to nucleolar
functions, accumulating evidence indicates they also exert
biological roles beyond the nuclear compartment. Under cellular
stress conditions, specific snoRNAs, including SNORD32A, SNORD33,
and SNORD35A, have been observed to accumulate in the cytosol,
suggesting potential extranuclear functional activities (49).
sdRNAs: Biogenesis and function
Biogenesis of sdRNAs
The precise mechanisms generating smaller fragments
from longer snoRNAs remain elusive. Current research confirms that
certain snoRNA-derived miRNAs, including those from H/ACA scaRNA15
(ACA45), require the ribonuclease III Dicer (DICER) enzyme for
their biogenesis. Comparative deep sequencing analysis of wildtype
vs. DICER/DiGeorge syndrome critical region 8 (DGCR8) knockout
embryonic stem cells revealed profound alterations in the length
distribution patterns of SNORA-sdRNAs. These findings compellingly
suggest the microprocessor complex (DICER/DGCR8) plays a
substantial role in a process analogous to miRNA biogenesis.
Conversely, the production of smaller fragments from SNORD appears
to operate through a distinct mechanism, evidenced by their
continued generation despite the absence of functional DICER/DGCR8
components. However, comprehensive cross-linking studies of
cellular RNAs with DGCR8 demonstrate extensive interactions between
snoRNAs and DGCR8. Crucially, DGCR8-mediated cleavage of snoRNAs
occurs independently of DICER, indicating the potential existence
of an alternative sdRNA biogenesis pathway. This alternative route
may involve other enzymatic components associated with DGCR8
(50,51). Based on their biogenesis and
structural features, sdRNAs can be broadly classified into three
categories: i) H/ACA-derived sdRNAs, typically 20-24 nt fragments
originating from the 3' terminus; ii) C/D-derived sdRNAs, which
include both longer (>26 nt) and shorter (17-19 nt) fragments
primarily derived from the 5' end; and iii) sno-derived
P-element-induced wimpy testis (PIWI)-interacting RNAs (piRNAs),
which are associated with PIWI proteins and range from 26-31 nt
(52,53). These distinct classes exhibit
diverse functional properties, as discussed below.
Function of sdRNAs
High-throughput sequencing studies have revealed
that snoRNAs undergo specific cleavage to produce smaller
functional fragments, rather than simply suffering random
degradation. This conclusion is bolstered by several key
observations: First, the expression patterns of sdRNAs exhibit
striking conservation across diverse species; second, an inverse
relationship often exists between snoRNA abundance and sdRNA
production, suggesting that highly abundant sdRNAs may originate
from relatively low-expressed snoRNAs; and third, the selective
presence of stable fragments from specific snoRNAs regions, as
opposed to random degradation products from other parts of the same
molecule, strongly implies the existence of a regulated
stabilization mechanism (48).
From a structural and functional perspective, sdRNAs
can be classified into distinct categories based on their
biogenesis and physical characteristics. SNORAs primarily generate
20-24 nucleotide fragments preferentially derived from their 3'
termini. By contrast, SNORDs produce two distinct size classes:
Longer fragments over 26 nucleotides and shorter fragments ranging
from 17-19 nucleotides, with the latter primarily originating from
the 5' end. This phenomenon of sdRNA production appears pervasive,
with experimental evidence demonstrating that >50% of all
snoRNAs are capable of generating these smaller fragments (54,55). While the biological functions of
numerous sdRNAs remain incompletely characterized, their widespread
occurrence and evolutionary conservation strongly suggest diverse
regulatory roles, some of which are increasingly being revealed
through ongoing research. The biogenesis and function of sdRNAs are
shown in Fig. 2.
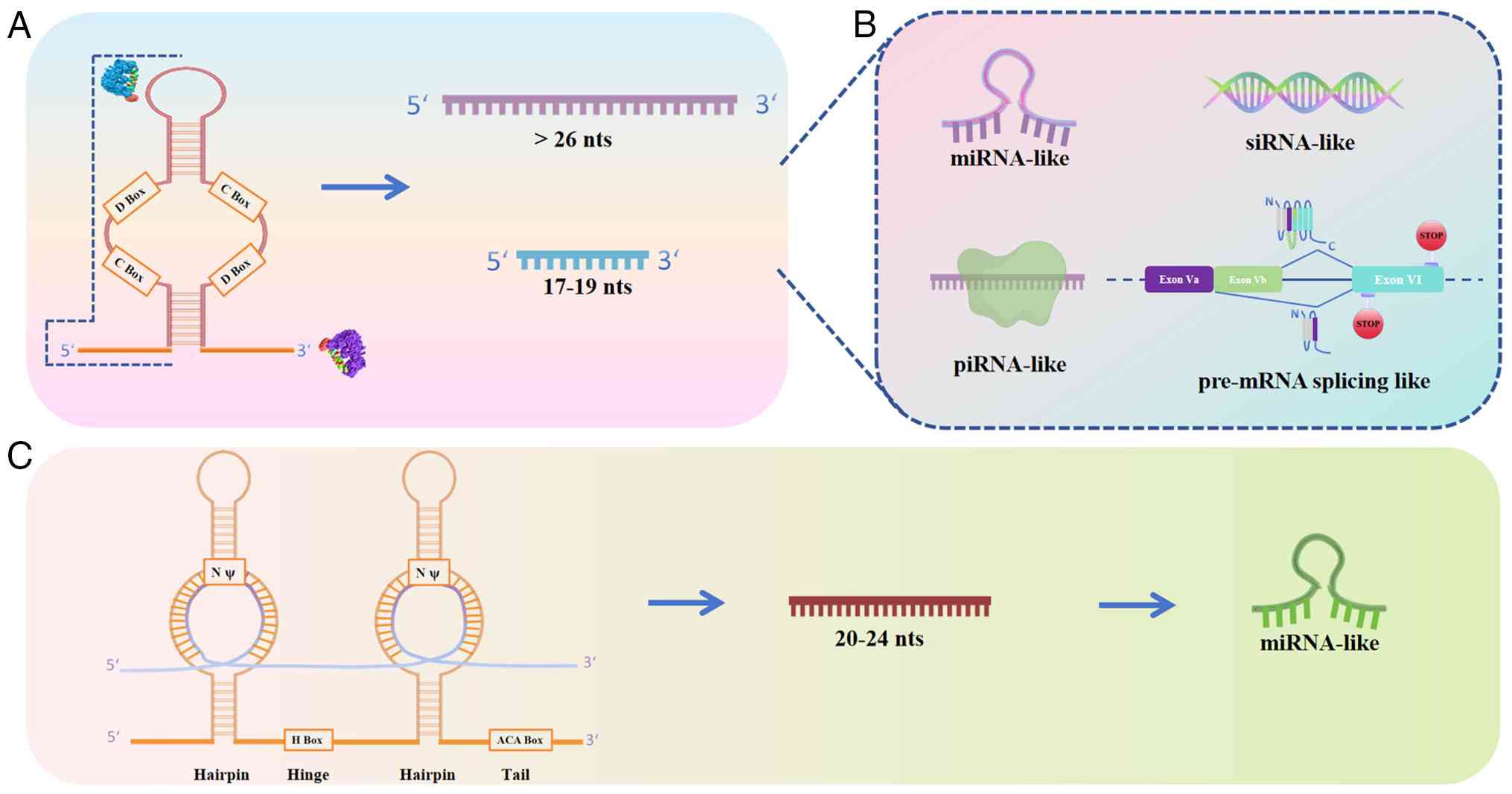 | Figure 2Biogenesis and function of sdRNAs.
(A) Processing pathways of C/D box snoRNA-derived fragments. C/D
box snoRNAs are processed via DGCR8-dependent or independent
pathways to yield longer (>26 nt) and shorter (17-19 nt)
fragments from the 5' end, which may adopt piRNA-like or miRNA-like
functions. (B) Functional outcomes of sdRNAs. miRNA-like sdRNAs
associate with AGO proteins and silence target mRNAs; piRNA-like
sdRNAs bind PIWI proteins and regulate gene expression
epigenetically; siRNA-like sdRNAs guide transcript cleavage through
near-perfect complementarity. In addition, both full-length snoRNAs
and specific sdRNAs have been implicated in the regulation of
pre-mRNA alternative splicing (such as SNORD115 and SNORD88C),
expanding their regulatory repertoire beyond RNA modification and
gene silencing. (C) Processing pathways of H/ACA box snoRNA-derived
fragments. H/ACA box snoRNAs are processed by DICER to generate
20-24 nt fragments predominantly from the 3' terminus, which can
function as miRNA-like molecules. Representative examples from each
category are indicated. snoRNA, small nucleolar RNA; sdRNA,
snoRNA-derived fragments; DGCR8, DiGeorge syndrome critical region
8; AGO, Argonaute; PIWI, P-element-induced wimpy testis; piRNA,
PIWI-interacting RNA; miRNA, microRNA; siRNA, small interfering
RNA; DICER, ribonuclease III Dicer; SNORD, C/D box snoRNA. |
miRNA-like functions
A particularly intriguing potential function of
sdRNAs lies in their capacity to serve as a novel source of
miRNA-like molecules. This hypothesis was initially investigated
through comprehensive analysis of small RNAs co-immunoprecipitated
with Agonaute RNA-induced silencing complex (RISC) catalytic
component (AGO)1 and AGO2 proteins in 293 cells. Notably, among the
AGO2-associated fragments, researchers identified a functionally
significant sdRNA derived from ACA45. Extensive functional
characterization, combining luciferase reporter assays with
sophisticated bioinformatic prediction tools, revealed that ACA45
exhibits canonical miRNA-like activity by specifically targeting
CDC2L6 mRNA. While ACA45 shares the requirement for DICER in its
maturation pathway with classical miRNAs, it displays a distinctive
processing mechanism that bypasses Drosha involvement (7). Evidence suggests that small RNAs
processed from snoRNAs can adopt microRNA (miRNA)-like functions,
though most associate with Argonaute proteins at levels
insufficient to elicit detectable silencing activity (56). Supporting this notion, Brameier
et al identified 11 distinct snoRNA-derived sequences
capable of integrating into the RISC and mediating effective gene
silencing. These 'sno-miRNAs' originate from SNORDs, including
snR39b, U3 (two loci), U78, HBII-336, HBII-429, HBII-142, U27,
U83a, U74, and U15a, and exhibit miRNA-like repression of
reporter-gene mRNAs (57).
piRNA-like functions
Emerging evidence has established snoRNAs as a
significant source of piRNAs, a distinct class of 26-31 nucleotide
ncRNAs that form functional complexes with PIWI proteins, a
specialized subgroup within the Argonaute protein family. These
snoRNA-derived piRNAs participate in crucial biological processes,
including transposon silencing and epigenetic regulation of gene
expression. In a pivotal study conducted in primary T lymphocytes,
researchers identified multiple piRNA candidates originating from
snoRNAs. Particularly noteworthy was the discovery of a piRNA
derived from SNORD63, which specifically interacts with an intronic
sequence of the IL-4 pre-mRNA. This interaction facilitates the
recruitment of the Trf4-Air2-Mtr4 complex, subsequently leading to
the targeted degradation of IL-4 mRNA by nuclear exosomes (58).
Further expanding our understanding of
snoRNA-derived piRNA functions, another investigation revealed that
a piRNA originating from SNORD75 exhibits dual binding capability
to PIWI-like protein 1 (PIWIL1) and PIWI-like protein 4 (PIWIL4)
proteins. This interaction triggers a significant epigenetic
transition, characterized by the replacement of repressive histone
marks (H3K27me3) with active marks (H3K4me3) at specific genomic
loci. This epigenetic reprogramming leads to transcriptional
activation of the TNF-related apoptosis-inducing ligand (TRAIL)
tumor suppressor gene, demonstrating the potential of
snoRNA-derived piRNAs in regulating critical cellular processes and
maintaining genomic stability (59).
siRNA-like function
Notably, a subset of sdRNAs exhibit near-perfect
complementarity to target mRNAs, enabling them to direct transcript
cleavage in a manner analogous to siRNAs.
Building on this concept, snoRNA modulator of gene
expression vectors have been developed as a versatile tool for
protein replacement in cultured cells. These vectors enable the
efficient substitution of endogenous cellular proteins with tagged
or mutated recombinant variants, offering a powerful approach for
functional studies (60).
Regulation of pre-mRNA splicing
SNORD115 has emerged as a key regulator in the
alternative splicing of HTR2C pre-mRNA, specifically facilitating
the inclusion of exon 5b to generate an extended variant. This
regulatory function is particularly noteworthy as it occurs
independently of the extensive RNA modifications typically required
for the synthesis of long mRNA isoforms (61). The splicing regulatory capacity of
SNORD115 extends beyond HTR2C, as it has been demonstrated to
influence alternative splicing events in other pre-mRNA targets
(62) Similarly, SNORD88C has
been identified as a modulator of FGFR3 splicing events. Both
SNORD115 and SNORD88C undergo specific processing to yield smaller
fragments, with several of these sdRNAs overlapping with regions
implicated in splicing regulation. However, the precise
contribution of these sdRNA fragments, as opposed to the
full-length snoRNAs, in splicing modulation remains to be fully
elucidated (48). The mechanistic
basis of sdRNA-mediated splicing regulation is not yet completely
understood, but emerging evidence suggests a potentially
significant role for interactions with a distinct group of
heterogeneous (hn) RNPs. These hnRNP interaction partners differ
from those associated with full-length snoRNAs and are themselves
known to participate in splicing regulation.
Biological roles of snoRNAs in cancer
Emerging research has revealed that abnormal
expression patterns and functional alterations of specific snoRNAs
are associated with various malignancies. Mechanistic
investigations have established that snoRNAs participate in
multiple oncogenic processes, influencing crucial aspects of cancer
biology such as tumor cell proliferation, metastatic potential,
apoptotic regulation, and therapeutic resistance (31). Furthermore, substantial clinical
evidence demonstrates a strong correlation between snoRNA
dysregulation and critical clinicopathological parameters,
including tumor staging, progression, and patient survival outcomes
(31,63).
However, while extensive research links abnormal
snoRNA expression to these malignancies (31,63), the evidential strength of these
associations varies significantly across the literature. To
systematically evaluate this landscape, as outlined in the
Introduction, current studies were stratified into three tiers: i)
Fundamental mechanistic investigations utilizing in vitro
and animal models; ii) clinical association studies relying on
retrospective analysis of tumor tissues; and iii) translational
research evaluating non-invasive biomarkers (liquid biopsies) and
therapeutic interventions. In the following sections, rather than
merely cataloging reported findings, the roles of snoRNAs across
specific cancer types are critically synthesized, explicitly
distinguishing between preliminary molecular observations,
established clinical correlations, and markers approaching
translational readiness. Summary of snoRNA functions and mechanisms
in various cancers is shown in Table
SI, and clinical implications of snoRNAs in cancers are shown
in Fig. 3.
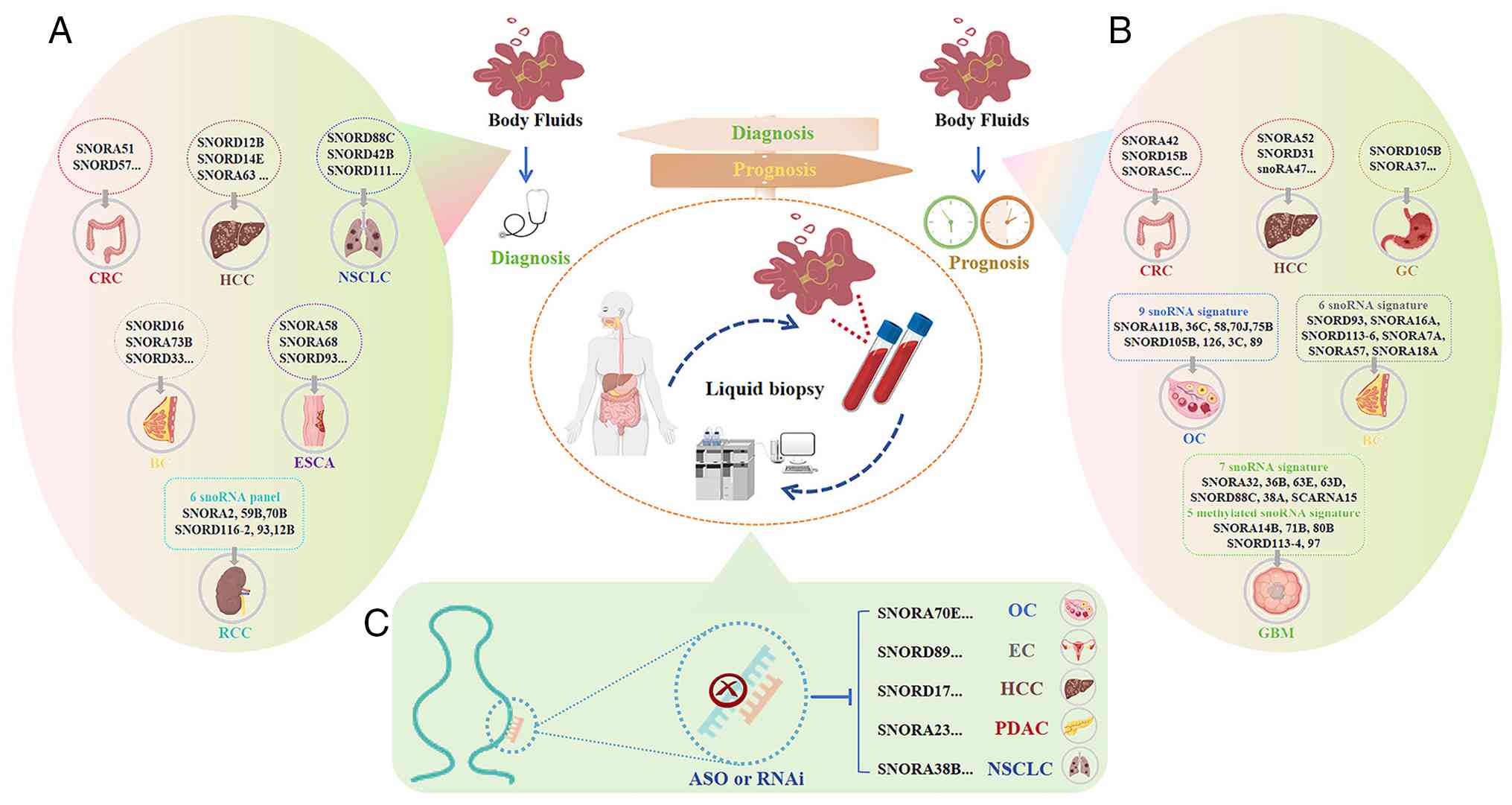 | Figure 3Representative examples highlighting
the cross-cancer clinical implications of snoRNAs. (A) snoRNAs as
diagnostic biomarkers: Emphasizing their cross-cancer applicability
in liquid biopsies for non-invasive detection (such as SNORA51 in
fecal samples for CRC, and SNORD33 in plasma for TNBC). (B) snoRNAs
as prognostic biomarkers: Highlighting their conserved roles in
driving shared oncogenic phenotypes, such as tumor proliferation,
cell-cycle dysregulation, and invasion/metastasis, where the
elevated expression of oncogenic snoRNAs (such as SNORA28 in CRC,
and SNORD17 in HCC) consistently correlates with advanced TNM
staging and poor overall survival across diverse malignancies. (C)
snoRNAs as therapeutic targets: Demonstrating the pan-cancer
potential of targeted interventions, where ASOs directed against
key oncogenic snoRNAs (such as SNORD17, and SNORA74A) effectively
suppress tumor growth and metastatic progression in preclinical
models. This figure presents selected representative examples
rather than an exhaustive overview to illustrate broader pan-cancer
themes. snoRNA, small nucleolar RNA; CRC, colorectal cancer; SNORD,
C/D box snoRNA; TNBC, triple-negative breast cancer; HCC,
hepatocellular carcinoma; ASO, antisense oligonucleotide; SNORA,
H/ACA box snoRNA; NSCLC, non-small cell lung cancer; BC, breast
cancer; ESCA, esophageal carcinoma; OC, ovarian cancer; EC,
endometrial cancer; PDAC, pancreatic ductal adenocarcinoma. |
Colorectal cancer (CRC)
Mechanistic drivers of
pathogenesis
Fundamental in vitro and in vivo
investigations have revealed extensive snoRNA involvement in CRC
progression. For instance, SNORA28 functioned as a molecular decoy
recruiting bromodomain-containing protein 4, leading to increased
H3K9 acetylation at the leukemia promoter, activating the
JAK1/STAT3 pathway, which promoted CRC cell proliferation and
conferred radioresistance (64).
SNORA56 played a critical role in the pseudouridylation of 28S rRNA
at the U1664 site, facilitating glutamate cysteine ligase catalytic
subunit translation to inhibit ferroptosis and sustain tumor
survival (65). SNORD11B
exemplifies a dual regulatory mechanism in CRC through
2'-O-methylation, which simultaneously promoted 18S rRNA maturation
at the G509 site to enhance ribosome biogenesis, and mediated the
degradation of MIRLET7A1HG (pri-let-7a) at the G225 site to
suppress let-7a-5p and upregulate oncogenic targets (66). Small Cajal body-specific RNA
(SCARNA)12 accelerated CRC progression by activating the PI3K/AKT
pathway (67). Another recent
study showed that SNORA21 enhanced CRC cell proliferation,
invasion, and migration through Hippo and Wnt signaling pathways,
two critical cascades involved in cancer development and metastasis
(68). SNORA24 was shown to
promote tumor growth by promoting G1/S phase transition
and modulated p53 protein stability via the ubiquitin-proteasome
pathway, suggesting its potential as a therapeutic target (69). SNORA42 has been implicated as a
putative oncogene in CRC, promoting multiple malignant phenotypes
including enhanced proliferation, migration, invasion, anoikis
resistance, and overall tumorigenic potential (70). SNORA71A exerted its oncogenic
effects through modulation of NF-κB and Toll-like receptor
pathways, thereby promoting CRC cell proliferation, migration, and
invasion (71).
SNORD12C and SNORD78 were demonstrated to catalyze
2'-O-methylation of 28S rRNA, stabilizing transcripts such as
eukaryotic factor 4A-III and laminim subunit γ2 to promote CRC
migration and survival (72).
SNORD1C enhanced stemness and metastasis via Wnt/β-catenin pathway.
Its suppression reduced β-catenin/transcription factor 7
expression, inhibiting tumorigenic properties (73). Conversely, SNORD44 and GAS5
exhibited tumor-suppressive properties by inhibiting growth and
inducing apoptosis (74), while
SNORD50A/B depletion amplified K-Ras activity and increased
tumorigenic risk (75).
Tissue-based clinical and prognostic
associations
These mechanistic findings are frequently supported
by retrospective analyses of patient tissues, highlighting strong
prognostic value. Clinically, high SNORA28 expression was shown to
be associated with poor prognosis (64). Elevated SNORD11B indicated
advanced TNM staging, lymph node metastasis, and a poor prognosis
(66). High expression of
SCARNA12 was linked to poor patient outcomes (67), and SNORA71A was correlated with
advanced TNM staging and metastasis (71). SNORA42 served as an independent
predictor of poor clinical outcomes (70).
Translational readiness and
non-invasive diagnostics
The most translationally mature evidence involves
liquid and fecal biopsies. SNORA51 in fecal samples exhibited
strong promise for CRC screening, particularly in fecal
immunochemical test-positive patients (76). SNORD15B and SNORA5C served as
independent prognostic markers, linked to lymphatic invasion and
colon polyp history (77). Serum
SNORD1C discriminated patients with CRC from healthy controls [area
under the curve (AUC)=0.748] and improved diagnostic accuracy with
carcinoembryonic antigen (CEA) (AUC=0.838) (78). Circulating SNORD57 has emerged as
a sensitive biomarker for early detection (79). Moreover, serum SNORA33 expression
was closely associated with vascular invasion and declined
following radical resection, demonstrating its diagnostic and
monitoring potential. Notably, serum SNORA33 represents a promising
non-invasive diagnostic biomarker for CRC, with its diagnostic
performance minimally affected by patient age (80).
In summary, while the mechanistic landscape of
snoRNAs in CRC, ranging from rRNA modification to epigenetic
regulation, is increasingly well-defined, their clinical
application remains in the exploratory phase. The dual role of
snoRNAs as both functional drivers and stable liquid biopsy
candidates highlights their diagnostic potential; however, bridging
the gap between these preclinical findings and translational
readiness will require rigorous independent validation in
large-scale longitudinal cohorts and a deeper understanding of
their roles in the complex tumor microenvironment (TME).
Hepatocellular carcinoma (HCC)
Mechanistic drivers of
pathogenesis
In HCC, snoRNAs function as versatile regulators
that drive malignancy by orchestrating distinct molecular axes
beyond their canonical roles. In terms of metabolic reprogramming,
transient receptor potential cation channel subfamily M member
8-mediated modulation of SNORA55 disrupted oxidative
phosphorylation by targeting adenosine triphosphate (ATP) synthase
subunits ATP5A1 and ATP5B, highlighting a direct link between
snoRNAs and mitochondrial bioenergetics (81). Simultaneously, a broad spectrum of
snoRNAs hijacks classical oncogenic signaling to promote invasion
and epithelial-mesenchymal transition (EMT). For instance, SNORD126
and SNORA11 (also known as ACA11) activated the PI3K/AKT/mTOR
cascade to drive robust cellular proliferation and migration
(82-84), whereas snoU2_19 and SNORD76
perturbed the Wnt/β-catenin pathway to accelerate cell cycle
progression and induce EMT (85,86).
Notably, snoRNAs target tumor suppressor
surveillance to evade apoptosis through multi-faceted inactivation
of the p53 pathway. SNORD17 was shown to exert a dual regulatory
influence, whereby its sequestration of nucleophosmin 1 (NPM1) and
MYB binding protein 1a (MYBBP1A) in the nucleolus suppressed
p53-mediated apoptosis. Conversely, its depletion triggered the
nucleoplasmic translocation of NPM1/MYBBP1A, which stabilized p53
via NPM1/mouse double minute 2 homolog (MDM2) interaction and
activated p53 through MYBBP1A/p300-mediated acetylation. Notably,
activated p53 reciprocally inhibited SNORD17 expression via
promoter acetylation, establishing a sophisticated self-regulatory
feedback loop (87). This
regulatory network was further diversified by SNORA18L5, which
promoted the MDM2-mediated degradation of p53 to inhibit apoptotic
signaling (88), and SNORA42,
which downregulated p53 to release cell cycle checkpoints and
suppress apoptosis (89).
Beyond the p53 axis, SNORD52, which is frequently
upregulated in HCC tissues, further drrove tumorigenesis by
elevating and stabilizing cyclin-dependent kinase 1 (CDK1) levels,
a process that was inversely correlated with up-frameshift
suppressor 1 homolog expression (90). Furthermore, specific snoRNAs are
pivotal for maintaining liver cancer stemness and translational
fidelity. For instance, SNORD88B anchored WRN to inactivate the
Hippo pathway and drive self-renewal, a capacity that was severely
impaired upon its deficiency (91). In terms of post-transcriptional
control, SNORD72 drove invasion and colony formation by
counteracting lncRNA-LALR1-mediated suppression to stabilize
inhibitor of DNA binding 2 mRNA (92). At the ribosomal level, snoRNAs
exert bidirectional control: SNORA24 guided the pseudouridylation
of 18S rRNA at positions 609 and 863, where loss of these
modifications altered translational efficiency to promote survival
in RAS-driven HCC (93); whereas
SNORA23 acted as a tumor suppressor by orchestrating 2'-O-ribose
methylation of 28S rRNA (cytidine4506) and inhibiting eukaryotic
translation initiation factor 4E-binding protein 1 phosphorylation
to restrict tumor growth (94).
Tissue-based clinical and prognostic
associations
A landmark analysis from The Cancer Genome Atlas
revealed 54 upregulated and 14 downregulated snoRNAs, establishing
distinct prognostic signatures (95). Elevated SNORA47 expression was
shown to be strongly correlated with intrahepatic metastasis,
lymphatic invasion, and advanced TNM stage, and exhibited
significantly shorter overall survival (OS) and higher recurrence
rates, underscoring its prognostic value (96). Similarly, SNORD78 was revealed to
be an adverse prognostic marker demonstrating a strong association
with increased tumor burden (multifocal lesions), advanced stage,
and distant metastasis, and was linked to poorer OS and
recurrence-free survival (97).
High SNORA51 was correlated with aggressive features including
portal vein tumor thrombus, vascular invasion, advanced TNM stage,
and shorter median survival (98). Conversely, downregulation of
tumor-suppressors carries severe prognostic implications: Reduced
SNORA52 predicted poor differentiation, capsular invasion, advanced
TNM stage, and worse disease-free survival (DFS) and OS (99); decreased SNORD31 was correlated
with larger tumor size, vascular invasion, and poor long-term
survival (100); and reduced
SNORD113-1 was strongly associated with worse survival outcomes
(101). Furthermore, diminished
SNORA71A was correlated with larger tumor size, multifocal lesions,
capsular invasion, poor differentiation, and advanced TNM stage,
predicting higher postoperative recurrence risk and reduced OS,
serving as an independent prognostic indicator (102). SNORA11, SNORD124, and SNORD46
also demonstrated potential diagnostic and prognostic utility
(103).
Translational readiness and
non-invasive diagnostics
SNORA47 and SNORD126 were shown to be highly
predictive in non-viral HCC, while SNORA80E and SNORA12 exhibited
favorable outcomes in combined analyses (104). In HBV-related HCC, signatures
(downregulated SNORD12B/SNORD14E; upregulated SNORA63) offered
diagnostic utility for early detection (105).
In summary, the role of snoRNAs in HCC spans a broad
spectrum of evidential strength. At the mechanistic level,
extensive in vitro and in vivo studies confirm that
snoRNAs are fundamental drivers of HCC pathogenesis, orchestrating
metabolic reprogramming, cell cycle dysregulation, and apoptosis
evasion. Clinically, large-scale retrospective tissue analyses
provide significant correlative evidence: Dysregulation of specific
snoRNAs, whether through upregulation of oncogenic drivers or the
suppression of tumor suppressors, acts as a powerful, independent
prognostic indicator for intrahepatic metastasis, high recurrence
risk, and poor OS. Most importantly, moving beyond retrospective
tissue associations, specific snoRNA signatures are rapidly
approaching translational readiness. Diagnostic panels,
particularly those tailored for the early screening of HBV-related
HCC, highlight the actionable, clinical utility of snoRNAs in
precision oncology.
Gastric cancer (GC)
Mechanistic drivers of
pathogenesis
In GC, in vitro evidence highlights complex
snoRNA-protein interactions. SNORA37 facilitated the cap
methyltransferase 1 (CMTR1)-ELAV-like RNA binding protein 1
(ELAVL1) complex to modulate CD44 alternative splicing, thereby
promoting tumor cell proliferation, invasive potential, and
metastatic dissemination (106).
SNORD105B interacted with aldolase A to modulate the c-Myc
signaling cascade, which ultimately enhanced the proliferative and
metastatic capacities of GC cells (107). Furthermore, SNORA21 promoted
gastric tumorigenesis by attenuating p53 activation through the
SNORA21-CHK1-PERP-MDM2 regulatory axis (108). Additionally, the coordinated
action of SNORA42, SNORA74A, and SNORD10 within
serine/arginine-rich protein-specific kinase 1-mediated signaling
networks directly facilitated tumor cell growth and GC development
(109).
Tissue-based clinical and prognostic
associations
These fundamental pathways manifest clinically as
poor prognostic markers. In GC patient cohorts, elevated tissue
expression of SNORA37 (alongside CMTR1, ELAVL1, and CD44) showed
significant association with reduced OS, poor therapeutic response,
and advanced disease progression (106). Similarly, elevated SNORD105B and
SNORA21 was correlated with increased tumor burden, poor
histological differentiation, and advanced TNM staging (107,108).
In GC, oncogenic snoRNAs drive progression via
complex RNA-protein interactions. Clinically, their tissue
upregulation strongly predicted advanced disease and poor survival.
However, translationally actionable tools such as non-invasive
liquid biopsies remain notably absent, representing a critical gap
for future research.
Non-small cell lung cancer (NSCLC)
Mechanistic drivers of
pathogenesis
Preclinical NSCLC models demonstrated that SNORD88C
mediates 2'-O-methylation of 28S rRNA to enhance the translation of
stearoyl-CoA desaturase-1, thereby promoting tumor cell
proliferation and metastatic dissemination (110). SNORA42 was shown to modulate
p53-dependent apoptotic mechanisms to drive tumorigenesis (111), while SNORA71A overcame
G0/G1 checkpoints via MEK/ERK pathway
activation, stimulating tumor cell growth, invasiveness, and
metastasis (112). Additionally,
NOP10 deletion suppressed cancer cell proliferation and reduced
migratory and invasive capacities through the dysregulation of
SNORA65, SNORA7A, and SNORA7B (113). Extensive research has elucidated
the critical role of SNORA38B in driving NSCLC progression through
dual mechanisms. Intracellularly, nuclear-localized SNORA38B
exerted its oncogenic effects by direct physical interaction with
the E2F transcription factor 1, which activates the GRB2-associated
binding protein 2/AKT/mTOR signaling cascade to promote tumor cell
proliferation and survival. Extracellularly, SNORA38B shaped an
immunosuppressive TME by inducing IL-10 secretion. This cytokine
shift recruited CD4+ FOXP3+ regulatory T
cells while simultaneously reducing the infiltration of
CD3+ CD8+ cytotoxic T lymphocytes,
effectively enabling the tumor to evade immune surveillance.
Translationally, targeting this axis using SNORA38B locked nucleic
acids (LNAs) successfully restored CD3+ CD8+
T-cell infiltration in the TME, thereby increasing the sensitivity
of NSCLC to immune checkpoint blockade (ICB) therapy (114).
Tissue-based clinical and prognostic
associations
Clinically, these molecular events are strictly
mirrored in retrospective tissue cohorts. Elevated tissue
expression of SNORD88C, SNORD42B, SNORD111, and SNORD83A was
consistently observed in patients with NSCLC relative to healthy
individuals (110,115,116). Notably, the significant
upregulation of SNORD60 in lung adenocarcinoma was directly
correlated with adverse clinicopathological features, including
lymph node metastasis and advanced TNM stage (117). Additionally, comprehensive
sequencing analyses highlighted SNORA21 as a valuable tissue-based
biomarker for predicting patient prognosis (118).
Translational readiness and
non-invasive diagnostics
NSCLC research features encouraging translational
readiness, particularly in non-invasive diagnostics and targeted
therapies. Circulating SNORD88C, SNORD42B, and SNORD111 showed
consistent elevation in plasma, enabling early-stage disease
detection (110,115). Circulating SNORD83A combined
with CEA demonstrated enhanced diagnostic capability (116), while circulating SNORD60 yielded
74.2% sensitivity and 75.3% specificity for early diagnosis
(117).
Taken together, oncogenic snoRNAs in NSCLC drive
proliferation, metastasis, and immune evasion. Clinically, their
elevation in tumor tissues strictly correlates with advanced TNM
stage, metastasis, and poor prognosis. Notably, circulating snoRNAs
in NSCLC are emerging as promising candidate non-invasive
biomarkers and potential therapeutic targets (such as LNA
therapeutics), although their transition to clinical practice
requires independent validation in large-scale cohorts.
Breast cancer (BC)
Mechanistic drivers of
pathogenesis
Mechanistic evidence in BC models reveals that
snoRNAs influence ribosomes, stemness, and immune evasion.
Knockdown studies confirmed that SNORA7B depletion enhanced
proliferative, invasive, and migratory capacities (119), while targeting SNORD50A/B
triggered cell cycle arrest and apoptosis via tripartite motif
containing (TRIM)21-guanosine monophosphate synthetase (120), and targeting U3/U8 suppressed
tumorigenic potential via impaired pre-rRNA processing (121). Conversely, SNORA71A acted as a
powerful driver of metastasis through a precise molecular cascade
by directly interacting with the G3BP stress granule assembly
factor 1 protein, which subsequently stabilized Rho associated
coiled-coil containing protein kinase 2 transcripts and modulated
the TGF-β signaling pathway. This orchestrated signaling ultimately
drove EMT, severely enhancing the proliferation, migration, and
invasion of BC cells (122).
SNORA68 was shown to bind U2 small nuclear RNA auxiliary factor 2
to upregulate nucleolar ribosomal protein L (RPL)23 and c-Myc,
promoting cancer stemness and tumorigenesis (123). SNORA51 maintained stem cell-like
properties via RPL3/NPM1/c-MYC (124). Furthermore, SNORA38 emerged as
an oncogenic driver closely linked to the expression of the
stemness marker octamer-binding transcription factor 4. SNORD67
directed U6 snRNA methylation to regulate lymph node metastasis
(125), and SNORA47 promoted
RPL11 transfer via early B-cell factor (EBF)3 to upregulate the
expression of the oncogene c-Myc (126). SNORD46 inhibition enhanced NK
cell-mediated antitumor immunity against triple-negative breast
cancer (TNBC) (43), while U50A
mediated everolimus resistance through mTOR downregulation
(127).
Tissue-based clinical and prognostic
associations
Multiple snoRNAs (SNORD15A, SNORD15B, SNORD22,
SNORD17, and SNORD87) are frequently overexpressed (128). SNORA7B functions as a potential
prognostic biomarker (119).
Mirroring its mechanistic role in EMT, SNORA71A exhibited marked
overexpression specifically in metastatic breast cancer tissues
compared with non-metastatic cases (122). High SNORA51 was associated with
poor prognosis (reduced OS and DFS) (124), while SNORA68 was strongly
correlated with Ki-67 index and TNM stage (123). SNORA47 was associated with high
TNM staging in Luminal A cohorts (126). SNORD90, SCARNA2, and SNORD78
effectively discriminated luminal subtypes; SNORD124 indicated
aggressive HER2+ variants; and a six-snoRNA signature
was shown to hold prognostic value for predicting locoregional
metastasis and clinical outcomes (129). SNORD16, SNORA73B, SCARNA4, and
SNORD49B were demonstrated to possess early-stage diagnostic
capability (130). Additionally,
elevated SNORA38 was strongly associated with advanced disease
characteristics, including increased tumor burden, nodal
involvement, and advanced TNM stage, as well as poor OS outcomes
(131).
Translational readiness and
non-invasive diagnostics
Moving beyond tissue profiling, snoRNAs are
demonstrating significant translational utility in liquid biopsies.
A panel of four snoRNAs (SNORD16, SNORA73B, SCARNA4, and SNORD49B)
has demonstrated significant capability as non-invasive biomarkers
for the early-stage detection of BC (AUC=0.7305) (130). Furthermore, circulating SNORD33
levels demonstrated a significant correlation with platinum-based
chemotherapy response in patients with metastatic TNBC, suggesting
strong utility as a predictive liquid biopsy biomarker for
chemotherapy efficacy (132).
Taken together, snoRNAs in BC regulate stemness,
metastasis, and immune evasion through intricate RNA-protein
networks. Clinically, specific snoRNA signatures possess potent
prognostic and molecular subtyping capabilities in tumor tissues.
Translationally, the identification of circulating snoRNAs as
non-invasive diagnostic panels and predictive markers for
chemotherapy response highlights their translational potential,
warranting further investigation in prospective clinical
trials.
Ovarian cancer (OC)
Mechanistic drivers of
pathogenesis
In vitro assays have demonstrated that
snoRNAs drive OC stemness and progression. SNORA70E was shown to
promote OC development by mediating RAP1B pseudouridylation and
PARP1 binding protein splicing (133). SNORA72 (134) and SNORD89 (135) upregulated stemness markers and
activated Notch receptor 1 (Notch1)/c-Myc, thereby enhancing
self-renewal, proliferation, and migratory potential. SNORD9
recruited methyltransferase like 3, to catalyze nuclear
transcription factor Y subunit α methylation (136). Conversely, SNORA81 knockdown
impaired 28S rRNA pseudouridylation, resulting in the suppression
of cancer cell proliferation and migration (137).
Tissue-based clinical and prognostic
associations
An H/ACA snoRNA signature (SNORA81, SNORA19,
SNORA56) was shown to distinguish high-grade serous ovarian
carcinoma from borderline tumors (137). Furthermore, a nine-snoRNA
prognostic signature was established as an independent predictor of
OC outcomes, serving as a highly promising prognostic biomarker
(138).
In OC, oncogenic snoRNAs predominantly drive cancer
stemness and migration by modulating key pathways such as
Notch1/c-Myc. Clinically, multi-snoRNA signatures in tissues act as
candidate diagnostic classifiers and independent prognostic
predictors. However, OC currently lacks mature translational
evidence for non-invasive liquid biopsies, indicating a critical
gap for future development.
Endometrial cancer (EC)
Mechanistic drivers of
pathogenesis
In EC, snoRNAs execute multifaceted oncogenic
functions, primarily through RNA modifications. Notably, SNORA73B
was shown to operate via a dual mechanism. First, it elevated the
pseudouridine modification of mindbomb E3 uniquitin-protein ligase
3 mRNA, increasing its stability and protein expression, which
subsequently reduced Jagged1 ubiquitination and led to the
sustained activation of the Notch signaling pathway. Additionally,
SNORA73B modulated the alternative splicing of regulator of
chromosome condensation 1 (RCC1), favoring the production of
oncogenic isoforms (RCC1-T2 and RCC1-T3). Together, these
mechanisms potently enhance tumor cell proliferation, migration,
and invasion while suppressing apoptosis (139). Furthermore, SNORD60 was shown to
exhibit carcinogenic properties by binding fibrillarin, a
2'-O-methyltransferase, to catalyze the 2'-O-methylation
modification of phosphatidylinositol-4,5-biphosphate 3-kinase
catalytic subunit α mRNA, thereby driving the PI3K/AKT/mTOR cascade
and tumor progression (140).
SNORD89 mediated 2'-O-methylation of Bim mRNA to disrupt the
Bcl-2/Bax apoptotic balance (141), SNORD104 modified PARP1 mRNA to
accelerate tumor growth (142),
and SNORD99 modified gasdermin D to inhibit pyroptosis (143). Additionally, SNORD15B enhanced
proliferation, invasion, and tumorigenicity through TRIM25
upregulation and cytoplasmic p53 accumulation (144).
Tissue-based clinical and prognostic
associations
Clinically, the dysregulation of these mechanisms
is mirrored in patient tissues. For instance, SNORD89 expression
was shown to be significantly elevated in endometrial tumors with
lymph node metastasis compared with non-metastatic cases,
highlighting its potential as a tissue-based indicator of
aggressive disease (141).
To synthesize these findings, evidence in EC relies
heavily on mechanistic models where snoRNAs drive invasion and
apoptosis evasion primarily via aberrant RNA modifications and
alternative splicing. Clinically, their upregulation in tumor
tissues is directly linked to adverse features such as metastasis.
Yet, unlike HCC or NSCLC, translational tools such as non-invasive
circulating biomarkers remain largely unexplored in EC,
representing a significant translational gap for future
studies.
Glioma
Mechanistic drivers of
pathogenesis
U3 snoRNA was revealed to drive aerobic glycolysis
and proliferation via the U3/ZBTB7A/HK2/LDHA axis (145) and recruited Ku-dependent
DNA-dependent protein kinase sububit catalytic subunit to
phosphorylate TRIM24, facilitating malignant transformation into
epithelioid glioblastoma (GBM)-like tumors (146). SNORD113-3 was shown to govern
GBM cell growth and glycolipid metabolism via the ADAR2/PHKA2/EBF1
axis (147), and SNORD88C
enhances proliferation, migration, and invasion (148).
Tissue-based clinical and rognostic
associations
Researchers have developed promising prognostic
tools. A 7-snoRNA prognostic signature (SNORA32, SNORA36B,
SCARNA15, SNORA63E, SNORA63D, SNORD88C and SNORD38A) and associated
nomogram predicted lower-grade glioma survival with high
sensitivity (148). Similarly, a
5-methylated-snoRNA (SNORA14B, SNORD113-4, SNORA71B, SNORA80B and
SNORD97) risk signature has been validated as an independent
prognostic indicator for glioma (149).
In glioma, snoRNAs uniquely drive malignant
transformation by reprogramming aerobic glycolysis and lipid
metabolism. Clinically, these metabolic drivers translate into
highly sensitive prognostic nomograms and risk signatures in tumor
cohorts. However, actionable liquid biopsies for non-invasive
monitoring are still needed to bridge the gap toward clinical
readiness.
Renal cell carcinoma (RCC)
Mechanistic drivers of
pathogenesis
In clear cell RCC (ccRCC), in vitro and
in vivo models have demonstrated that SNORA33 acts as a
potent oncogenic driver. Mechanistically, it promoted tumor cell
invasion and metastasis while simultaneously conferring resistance
to the targeted therapy sunitinib through the activation of the
JAK/STAT signaling pathway (150).
Tissue-based clinical and prognostic
associations
Clinically, the dysregulation of this signaling
pathway was strictly mirrored in patient tissue samples. Elevated
tissue expression of SNORA33 served as a significant and direct
indicator of unfavorable prognosis and reduced survival outcomes in
ccRCC cohorts.
Translational readiness and
non-invasive diagnostics
RCC research demonstrates notable translational
progress, particularly in the realm of liquid biopsies. SNORD15A,
SNORD35B, and SNORD60 exhibited significant diagnostic performance
when isolated from urine sediments for early-stage RCC (151). Furthermore, circulating SNORD63
(urine) and SNORD96A (plasma) demonstrated high diagnostic accuracy
(152). Most notably, a 6-snoRNA
diagnostic panel (SNORA2, SNORD116-2, SNORA59B, SNORD93, SNORD12B,
SNORA70B) isolated from urinary extracellular vesicles (EVs)
outperformed standard clinical parameters; when combined with the
Fuhrman grading system, its diagnostic AUC reached an impressive
0.800 (153).
Taken together, while mechanistic evidence in RCC
highlights the role of snoRNAs in driving invasion and targeted
therapy resistance, this field is uniquely characterized by its
advanced translational readiness. Highly accurate, non-invasive
snoRNA panels derived from urine and plasma are already
demonstrating the potential to outperform standard clinical
parameters for early cancer detection.
Other cancers
Mechanistic drivers of
pathogenesis
The oncogenic mechanisms of snoRNAs span diverse
tumor types, fundamentally driving progression and therapy
resistance. In cervical cancer, SNORD6 scaffolded the E6-E6AP-p53
degradation complex to dysregulate apoptotic pathways (154). In bladder cancer, SCARNA12
interacted with H2A histone family member Z to enhance
proliferative and metastatic capacities and to inhibit cell cycle
arrest and apoptosis (155).
SNORD35A modulated proliferation, invasion, and EMT in pancreatic
cancer via the hepatocyte growth factor/c-Met pathway (156). In neuroblastoma, the
SNHG25/SNORA50C axis stabilized histone deacetylase 1 to promote
tumor growth and migration (157). Furthermore, in esophageal
squamous cell carcinoma (ESCC), snoRNAs actively mediated treatment
resistance. Specifically, SNORA58 conferred radioresistance by
attenuating ionizing radiation (IR)-induced ferroptosis via the
SNORA58/CTCF/JNK1 regulatory axis (158). Concurrently, SNORA80B
established a feed-forward oncogenic loop by stabilizing
cholesterol metabolism transcripts through an m6A-YTH
domain containing 1-dependent mechanism, promoting the accumulation
of cholesterol/dihydrotestosterone (DHT) and lipid droplet
formation. In turn, DHT-activated androgen receptor (AR) further
upregulated SNORA80B expression, driving profound chemotherapy
resistance (159).
Tissue-based clinical and prognostic
associations
Clinically, the mechanistic dysregulation of these
snoRNAs is frequently observed in patient tissue cohorts. Across
these diverse malignancies, the elevated tissue expression of these
specific oncogenic snoRNAs consistently correlates with adverse
clinicopathological features, including advanced disease staging,
increased metastatic burden, and poor OS outcomes (154-159).
Translational readiness and
non-invasive diagnostics
Beyond tissue-based associations, emerging evidence
highlights the translational potential of snoRNAs in these cancers.
Most notably in esophageal cancer, the identification of a specific
snoRNA signature (SNORA58, SNORA68, and SNORD93) within
tumor-educated platelets (TEPs) demonstrated high capability for
early disease detection, representing a promising non-invasive
liquid biopsy application (160). Therapeutically, targeted
interventions are showing immense promise: Targeting the JNK
signaling pathway represents a viable strategy to improve
radiotherapy outcomes in patients with ESCC and elevated SNORA58
expression (158), while the
pharmacological inhibition of SNORA80B using clinofibrate exhibited
potent synergy with cisplatin to effectively overcome chemotherapy
resistance in ESCC models (159).
To synthesize these findings, snoRNAs consistently
act as fundamental drivers of tumor growth, metastasis, and therapy
resistance across a wide spectrum of malignancies. While their
prognostic value is firmly established in tumor tissues, innovative
translational applications, ranging from TEP profiling for early
diagnosis to targeted radiosensitization and chemosensitization
strategies, underscore the rapidly expanding clinical utility of
snoRNAs in pan-cancer settings.
sdRNAs in cancer
Given the growing recognition of sdRNAs in cancer
pathogenesis, it is essential to critically evaluate their
functional roles and clinical potential. Similar to full-length
snoRNAs, the current understanding of sdRNAs is synthesized across
three distinct tiers of evidential strength.
Mechanistic drivers of pathogenesis
Fundamental in vitro and in vivo
models have elucidated the capacity of sdRNAs to actively drive or
suppress tumor progression through complex molecular networks. In
androgen-resistant prostate cancer (AR-PCa), specific fragments
(sdRNA-D19b and sdRNA-A24) were shown to promote tumor
aggressiveness by suppressing the expression of tumor suppressor
genes CD44 and CDK12, thereby enhancing cellular proliferation,
metastatic potential, and chemoresistance (161). Similarly, Patterson et al
(162) revealed that SNORD93
undergoes processing into a functionally active, miRNA-like
fragment (sdRNA93). This specific sdRNA was shown to regulate
sarcosine metabolism by modulating pipecolic acid and sarcosine
oxidase expression, directly driving cancer cell invasiveness
(162). Conversely, certain
sdRNAs exert potent tumor-suppressive effects. In breast cancer
models, a specific sno-derived piRNA from the GAS5 locus (pi-sno75)
actively upregulated the TRAIL pathway to induce apoptosis
(59).
Tissue-based clinical and prognostic
associations
Clinically, the profound dysregulation of these
molecules is consistently reflected in patient tissue cohorts. In
pancreatic ductal adenocarcinoma (PDAC), comprehensive RNA
sequencing has identified severe sdRNA dysregulation, with specific
fragments such as hsa-sno-HBII-85-29 showing pronounced
downregulation and hsa-sno-HBII-296B exhibiting high
upregulation in tumor tissues (163). More broadly, by integrating
small RNA sequencing data from 10,262 patient samples across 32
cancer types, Chow et al (164) constructed a comprehensive
pan-cancer sdRNAome landscape. This landmark analysis revealed that
sdRNA expression signatures are highly cancer-specific and possess
the potential to independently stratify patient survival outcomes
(164).
Translational readiness and biomarker
potential
Beyond tissue associations, sdRNAs are emerging as
highly translatable clinical tools, particularly in the realm of
tumor immunology. A substantial cohort of sdRNAs demonstrates
notable correlations with critical features of the tumor-immune
microenvironment, including the expression of immunosuppressive
markers, levels of CD8+ T-cell infiltration, cytolytic
T-cell activity, and tumor vascularization patterns (164). These specific immune-associated
sdRNA signatures hold immense translational potential as predictive
biomarkers for stratifying patient responses to immunotherapy
(164). Furthermore, the
identification of functionally active sdRNAs driving
chemoresistance in advanced malignancies such as AR-PCa highlights
the need for their further investigation as novel therapeutic
targets (161).
To synthesize these findings, sdRNAs represent a
previously underappreciated yet functionally prevalent class of
ncRNAs. From orchestrating metabolic and epigenetic shifts to
serving as candidate prognostic indicators and
immune-microenvironment classifiers, sdRNAs demonstrate substantial
clinical relevance. Transitioning these molecules from mechanistic
discovery toward targeted clinical applications represents a
critical and highly promising frontier in precision oncology.
Extracellular snoRNAs for cancer diagnosis
and prognosis
Beyond their intracellular functions, snoRNAs are
consistently detectable in diverse body fluids, where they exhibit
remarkable stability due to two primary protective mechanisms,
including encapsulation within EVs or association with high-density
lipoprotein complexes (165,166). This stable circulation positions
them as highly promising non-invasive biomarkers for early cancer
detection and dynamic monitoring. A comprehensive summary of
snoRNAs with diagnostic, prognostic, or therapeutic significance is
provided in Table SII, while
their broader clinical implications are illustrated in Fig. 3.
Diagnostic performance in liquid
biopsies
Recent clinical investigations highlight the
diagnostic potential of circulating snoRNAs, particularly when
multiplexed with conventional markers. For instance, in NSCLC,
integrating diminished serum exosomal SNORD116 and SNORA21 levels
with conventional biomarkers (cytokeratin 19 fragment 21-1 and CEA)
surged the diagnostic accuracy to an AUC of 0.917, effectively
distinguishing metastatic from non-metastatic disease (167). Similarly, in ccRCC, a panel of
four EV-associated snoRNAs (SNORD99, SNORD22, SNORD26, and
SNORA50C) isolated from non-invasive urine samples has emerged as a
candidate diagnostic tool (153). Furthermore, a discovery-stage
clinical study in pancreatic cancer identified a multi-marker panel
(WASP family member 2, ADP-ribosylation factor 6, SNORA74A, and
SNORA25) that exhibited promising diagnostic value (AUC >0.9),
surpassing the standard carbohydrate antigen 19-9 biomarker in
accurately detecting early-stage (0-IIA) pancreatic cancer
(168).
Technical challenges and emerging
solutions
Despite their clinical promise, significant
technical hurdles hinder the accurate identification and
quantification of circulating snoRNAs. Key challenges include RNA
isolation biases stemming from their small size (60-300 nt), the
lack of polyadenylation tails that complicates standard library
preparation, and highly stable secondary structures that impair
reverse transcription efficiency. Furthermore, the rigorous
clinical translation of circulating snoRNAs is heavily dependent on
overcoming critical methodological bottlenecks. These include
standardizing specimen types (such as serum vs. plasma vs. EVs),
optimizing pre-analytical handling (such as centrifugation
protocols and minimizing freeze-thaw cycles), establishing
consensus strategies for data normalization (given the lack of
universal endogenous controls in biofluids), and selecting
appropriate detection platforms (reverse transcription-quantitative
PCR vs. small RNA-sequencing) to ensure cross-study
reproducibility. However, recent advances in small RNA-sequencing
technologies, including specialized adapter designs for
non-polyadenylated RNAs, improved reverse transcriptases for
structured RNAs, and snoRNA-specific bioinformatic pipelines, have
markedly enhanced analytical sensitivity (169,170). These methodological refinements
are actively accelerating the exploration of circulating snoRNAs as
candidate liquid biopsy biomarkers, although standardizing these
protocols for definitive clinical application requires further
extensive investigation.
SnoRNA-targeting tools
Target discovery and therapeutic
rationale
While the precise molecular mechanisms of snoRNAs
in tumorigenesis remain incompletely characterized, growing
evidence suggests these ncRNAs are potential candidates for cancer
therapy. The recent development of snoRNA-enriched
kethoxal-assisted RNA-RNA sequencing (snoKARR-seq) technology
represents a paradigm shift, enabling the transcriptome-wide
mapping of snoRNA-mRNA interactions and identifying over 1,000
novel snoRNA-mRNA pairs. These discoveries challenge traditional
paradigms by revealing modification-independent regulatory roles,
thereby exponentially expanding the potential therapeutic target
space for rational drug design (41). Furthermore, proof-of-concept
research, such as the engineered expression of SNORA73 fused to a
secreted green fluorescent protein reporter to modulate protein
secretion pathways, suggest that artificial snoRNAs could
potentially compensate for deficient secretory proteins in
malignancies (41).
NATs in preclinical models
The past decade has witnessed notable progress in
translating ncRNA targets into precision medicine (171). NATs, primarily antisense
oligonucleotides (ASOs), ASO gapmers, and LNA oligonucleotides,
have emerged as the most promising modalities for snoRNA-directed
interventions. Preclinical success using ASOs is well-documented
across multiple malignancies. For instance, ASO-mediated silencing
significantly suppressed tumor growth and tumorigenesis in HCC
(SNORD17, SNORD88B, SNORA74A) (87,91,172), PDAC (SNORA23) (173), OC (SNORA70E, SNORD9) (133,136), and EC (SNORD89, SNORD104,
SNORD99) (141-143). Beyond monotherapy, NATs show
profound synergistic potential and ASO-mediated inhibition of
SNORA13 was revealed to synergize with 5-fluorouracil in CRC
(174), while LNAs targeting
SNORA38B significantly enhanced CD8+ T-cell
infiltration, sensitizing NSCLC to ICB therapy (114).
Delivery hurdles and the frontier of
small-molecule drugs (SMDs)
Although intrathecal administration of ASOs has
demonstrated clinical safety in human trials (175), broad oncological application
faces significant barriers. To date, no ASO or LNA drugs have
received FDA approval specifically for cancer treatment, primarily
due to unresolved challenges regarding tumor-specific delivery and
immune modulation. Emerging non-viral delivery strategies, such as
engineered liposomes and hybrid carriers, show promise in
addressing these limitations.
Consequently, SMDs represent a highly attractive
parallel frontier. Compared with NATs, SMDs offer superior
solubility, enhanced metabolic stability, lower production costs,
and easier cellular penetration (176). Crucially, snoRNAs present
multiple druggable structural features, including complex
secondary/tertiary structures, conserved functional motifs, and
distinct protein interaction interfaces within snoRNP complexes.
While dedicated snoRNA-focused high-throughput screening campaigns
are still in their infancy, the demonstrated success of
miRNA-targeted screening platforms and in silico models
provides a strong and viable precedent for the development of
snoRNA-targeted SMDs (177).
Future challenges in clinical
translation
Despite these promising preclinical results, the
clinical translation of snoRNA-targeted therapeutics faces several
hurdles that must be addressed: i) Efficient delivery to
extrahepatic tumor tissues; ii) mitigating off-target effects and
innate immune sensor activation; iii) overcoming the nuclear
localization barrier of numerous snoRNAs; and iv) defining the
stoichiometric requirements for functionally disrupting snoRNP
complexes. Overcoming these barriers will require interdisciplinary
innovations combining RNA chemistry, nanomedicine, and tumor
biology.
Discussion
The rapid advancement of high-throughput
transcriptomic and specialized RNA-interaction mapping technologies
has fundamentally reshaped our understanding of snoRNA and sdRNA
biology. Moving beyond their canonical roles in rRNA modification,
these molecules are now recognized as integral components of a
complex regulatory network, with their dysregulation being a
hallmark of numerous malignancies. The present review synthesized
evidence that snoRNAs and sdRNAs contribute to tumorigenesis
through a markedly diverse repertoire of mechanisms-from modulating
mRNA splicing, stability, and translation to directly interfacing
with signaling proteins and epigenetic regulators. Their presence
in EVs and notable stability in circulation further underscore
their potential as liquid biopsy biomarkers.
Beyond their cell-autonomous functions, emerging
evidence suggests that snoRNAs may play critical roles in the TME.
For instance, EV-associated snoRNAs have been detected in various
biofluids and may mediate intercellular communication between
cancer cells and stromal or immune cells. Additionally, SNORD46 has
been shown to modulate NK cell function in obesity-associated
cancer (43), hinting at broader
immunomodulatory roles. Future studies should explore whether
snoRNAs are actively transferred between cells in the TME and
whether they contribute to immune evasion or therapy
resistance.
However, despite this considerable progress, the
field stands at a critical juncture, facing three intertwined gaps
that must be bridged to translate mechanistic insights into
clinical utility.
The first and most fundamental gap is the
functional annotation deficit. While catalogs of dysregulated
snoRNAs and sdRNAs in cancer are rapidly expanding, the precise
molecular functions for the vast majority remain enigmatic, with
many still classified as 'orphans' (1). The full oncogenic or
tumor-suppressive potential of these RNAs cannot be harnessed
without a systems-level understanding of their interaction
networks, target spectra, and context-dependent roles. Closing this
gap requires a concerted shift from descriptive profiling to
mechanistic deconvolution. Emerging technologies such as
snoKARR-seq, which maps transcriptome-wide snoRNA-mRNA
interactions, alongside in situ RNA modification capture
techniques, are pivotal. Future research must prioritize the
systematic identification of bona fide targets (both RNA and
protein), the functional consequences of their binding, and the
integration of this data across different cancer types and states
(such as treatment-naïve vs. resistant).
The second gap is the methodological and
translational gap. The journey from identifying a promising
cancer-associated snoRNA to a validated clinical biomarker or
target is fraught with technical and biological challenges. Current
detection methods struggle with the unique biochemistry of snoRNAs,
including their small size, lack of poly-A tails, and stable
secondary structures, leading to biases in isolation, sequencing,
and quantification. Consequently, there is an urgent need for
standardized, sensitive, and reproducible protocols for
snoRNA/sdRNA detection from both tissues and liquid biopsies.
Furthermore, the heavy reliance of the field on synthetic,
unmodified RNAs for functional studies is a major limitation, as
endogenous snoRNAs are extensively post-transcriptionally modified.
Synthetic analogs may not recapitulate these native properties,
potentially leading to misleading conclusions. Developing
strategies to isolate and study endogenous snoRNPs in their native
state, and to chemically synthesize or engineer RNAs that
faithfully mimic their modified counterparts, is essential for
accurate functional validation and therapeutic design. A major
specific limitation in the field is the current lack of
standardized criteria for defining and annotating sdRNAs. Unlike
miRNAs, which benefit from well-established biogenesis hallmarks
and centralized annotation databases (such as miRBase), sdRNAs
exhibit significant heterogeneity in length, processing boundaries,
and functional modalities. Consequently, different research groups
often employ disparate bioinformatics pipelines, read-length
thresholds, and alignment strategies to identify these fragments
from small RNA sequencing data. This lack of methodological
consensus leads to substantial variability and discordance across
studies, severely complicating cross-study comparisons and
meta-analyses. Establishing a unified nomenclature and rigorous,
standardized annotation criteria, integrating precise cleavage
signatures with structural and functional validation, is an urgent
prerequisite to reliably distinguish bona fide regulatory
sdRNAs from stochastic snoRNA degradation products and to ensure
reproducibility in future clinical investigations (48,56).
The third gap is the therapeutic development gap.
While proof-of-concept studies using ASOs or LNAs against specific
oncogenic snoRNAs show impressive preclinical efficacy, significant
hurdles remain for clinical adoption (171). The core challenges of in
vivo delivery, achieving sufficient tumor-specific uptake while
minimizing off-target effects and immunostimulation, are amplified
for nuclear-localized targets. Moreover, the stoichiometric nature
of snoRNP complexes raises critical questions about whether partial
inhibition of a snoRNA is sufficient to disrupt an oncogenic
program, or if near-complete knockdown is required. Overcoming this
barrier will likely require exploring combination therapies
(pairing snoRNA inhibition with chemotherapy, targeted therapy, or
immunotherapy) and advancing the development of small-molecule
inhibitors that disrupt snoRNA-protein interactions or functions
(171,176,177).
Furthermore, the specificity of snoRNA-mRNA
interactions identified by crosslinking-based methods remains a
concern, as these techniques may capture transient or
non-functional associations. Future advances in high-resolution
structure determination and functional screening will be essential
to resolve these uncertainties.
Addressing these outlined gaps demands a
multidisciplinary approach, combining cutting-edge RNA biology,
sophisticated bioengineering, and comprehensive clinical research.
By elucidating function, refining analytical tools, and innovating
delivery platforms, the potential of these RNAs as precise
biomarkers and novel therapeutic levers can be fully realized,
potentially adding a powerful new dimension to precision
oncology.
Conclusion
SnoRNAs and their derived sdRNAs have firmly
emerged as critical, multidimensional regulators of cancer biology,
with diverse functions ranging from guiding RNA modifications to
modulating signaling pathways and immune responses. Their
dysregulation across multiple cancer types, coupled with their
stability in biofluids, positions them as promising non-invasive
biomarkers for early detection, prognosis, and treatment
monitoring. Furthermore, the development of ASO- and LNA-based
therapeutics targeting oncogenic snoRNAs has shown preclinical
efficacy, paving the way for clinical translation. Future research
should focus on: i) Elucidating the precise molecular mechanisms of
orphan snoRNAs using advanced technologies such as snoKARR-seq and
cryogenic electron microscopy; ii) validating circulating snoRNA
signatures in large, prospective cohorts; iii) exploring
combination therapies that integrate snoRNA targeting with ICB or
conventional chemotherapy; and iv) developing targeted delivery
systems to enhance tumor-specific uptake while minimizing
off-target effects. Addressing these challenges will be essential
for realizing the full potential of snoRNA-based strategies in
precision oncology.
Supplementary Data
Availability of data and materials
Not applicable.
Authors' contributions
JM and ZP designed the scope and and structure of
the review, performed structured literature searches, prepared the
figures, and wrote major sections of the manuscript. FG critically
synthesized and interpreted findings. HL and ZP revised major
sections of the manuscript. ZP provided conceptual guidance and
supervised the overall project. JM, HL and ZP secured the funding.
All authors read and approved the final manuscript. Data
authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
Not applicable.
Funding
The present review was supported by The Key Research and
Development Project of Zhenjiang (grant no. SH2022044), the General
Project of Xuzhou Basic Research Plan in 2023 (grant no. KC23070),
the Key Research and Development Program of Xuzhou City (grant no.
KC22155), and the Innovation and Entrepreneurship Project of Xuzhou
Medical University Science Park (grant no. CXCYZX202410).
References
|
1
|
Lestrade L and Weber MJ: snoRNA-LBME-db, a
comprehensive database of human H/ACA and C/D box snoRNAs. Nucleic
Acids Res. 34(Database Issue): D158–D162. 2006. View Article : Google Scholar :
|
|
2
|
Yoshihama M, Nakao A and Kenmochi N:
snOPY: A small nucleolar RNA orthological gene database. BMC Res
Notes. 6:4262013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Huang C, Shi J, Guo Y, Huang W, Huang S,
Ming S, Wu X, Zhang R, Ding J, Zhao W, et al: A snoRNA modulates
mRNA 3' end processing and regulates the expression of a subset of
mRNAs. Nucleic Acids Res. 45:8647–8660. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Falaleeva M, Pages A, Matuszek Z, Hidmi S,
Agranat-Tamir L, Korotkov K, Nevo Y, Eyras E, Sperling R and Stamm
S: Dual function of C/D box small nucleolar RNAs in rRNA
modification and alternative pre-mRNA splicing. Proc Natl Acad Sci
USA. 113:E1625–E1634. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gong J, Li Y, Liu CJ, Xiang Y, Li C, Ye Y,
Zhang Z, Hawke DH, Park PK, Diao L, et al: A Pan-cancer analysis of
the expression and clinical relevance of small nucleolar RNAs in
human cancer. Cell Rep. 21:1968–1981. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kawaji H, Nakamura M, Takahashi Y,
Sandelin A, Katayama S, Fukuda S, Daub CO, Kai C, Kawai J, Yasuda
J, et al: Hidden layers of human small RNAs. BMC Genomics.
9:1572008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ender C, Krek A, Friedlander MR,
Beitzinger M, Weinmann L, Chen W, Pfeffer S, Rajewsky N and Meister
G: A human snoRNA with microRNA-like functions. Mol Cell.
32:519–528. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dieci G, Preti M and Montanini B:
Eukaryotic snoRNAs: A paradigm for gene expression flexibility.
Genomics. 94:83–88. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Brown JWS, Marshall DF and Echeverria M:
Intronic noncoding RNAs and splicing. Trends Plant Sci. 13:335–342.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kufel J and Grzechnik P: Small nucleolar
RNAs tell a different tale. Trends Genet. 35:104–117. 2019.
View Article : Google Scholar
|
|
11
|
Zimta AA, Tigu AB, Braicu C, Stefan C,
Ionescu C and Berindan-Neagoe I: An emerging class of long
non-coding RNA with oncogenic role arises from the snoRNA host
genes. Front Oncol. 10:3892020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Matera AG, Terns RM and Terns MP:
Non-coding RNAs: Lessons from the small nuclear and small nucleolar
RNAs. Nat Rev Mol Cell Biol. 8:209–220. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dupuis-Sandoval F, Poirier M and Scott MS:
The emerging landscape of small nucleolar RNAs in cell biology.
Wiley Interdiscip Rev RNA. 6:381–397. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Warf MB and Berglund JA: MBNL binds
similar RNA structures in the CUG repeats of myotonic dystrophy and
its pre-mRNA substrate cardiac troponin T. RNA. 13:2238–2251. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Leader DJ, Clark GP, Watters J, Beven AF,
Shaw PJ and Brown JW: Clusters of multiple different small
nucleolar RNA genes in plants are expressed as and processed from
polycistronic pre-snoRNAs. EMBO J. 16:5742–5751. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Busch H, Reddy R, Rothblum L and Choi YC:
SnRNAs, SnRNPs, and RNA processing. Annu Rev Biochem. 51:617–654.
1982. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jack K, Bellodi C, Landry DM, Niederer RO,
Meskauskas A, Musalgaonkar S, Kopmar N, Krasnykh O, Dean AM,
Thompson SR, et al: rRNA pseudouridylation defects affect ribosomal
ligand binding and translational fidelity from yeast to human
cells. Mol Cell. 44:660–666. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bachellerie JP and Cavaillé J: Guiding
ribose methylation of rRNA. Trends Biochem Sci. 22:257–261. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liang J, Wen J, Huang Z, Chen XP, Zhang BX
and Chu L: Small nucleolar RNAs: Insight into their function in
cancer. Front Oncol. 9:5872019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Reichow SL, Hamma T, Ferré-D'Amaré AR and
Varani G: The structure and function of small nucleolar
ribonucleoproteins. Nucleic Acids Res. 35:1452–1464. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bachellerie JP, Cavaillé J and Huttenhofer
A: The expanding snoRNA world. Biochimie. 84:775–790. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Watkins NJ, Dickmanns A and Lührmann R:
Conserved stem II of the box C/D motif is essential for nucleolar
localization and is required, along with the 15.5K protein, for the
hierarchical assembly of the box C/D snoRNP. Mol Cell Biol.
22:8342–8352. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kiss-László Z, Henry Y, Bachellerie JP,
Caizergues-Ferrer M and Kiss T: Site-specific ribose methylation of
preribosomal RNA: A novel function for small nucleolar RNAs. Cell.
85:1077–1088. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tyc K and Steitz JA: U3, U8 and U13
comprise a new class of mammalian snRNPs localized in the cell
nucleolus. EMBO J. 8:3113–3119. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Girard JP, Lehtonen H, Caizergues-Ferrer
M, Amalric F, Tollervey D and Lapeyre B: GAR1 is an essential small
nucleolar RNP protein required for pre-rRNA processing in yeast.
EMBO J. 11:673–682. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ganot P, Caizergues-Ferrer M and Kiss T:
The family of box ACA small nucleolar RNAs is defined by an
evolutionarily conserved secondary structure and ubiquitous
sequence elements essential for RNA accumulation. Genes Dev.
11:941–956. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Watkins NJ, Gottschalk A, Neubauer G,
Kastner B, Fabrizio P, Mann M and Lührmann R: Cbf5p, a potential
pseudouridine synthase, and Nhp2p, a putative RNA-binding protein,
are present together with Gar1p in all H BOX/ACA-motif snoRNPs and
constitute a common bipartite structure. RNA. 4:1549–1568. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Massenet S, Bertrand E and Verheggen C:
Assembly and trafficking of box C/D and H/ACA snoRNPs. RNA Biol.
14:680–692. 2017. View Article : Google Scholar :
|
|
29
|
Meier UT: RNA modification in Cajal
bodies. RNA Biol. 14:693–700. 2017. View Article : Google Scholar :
|
|
30
|
Cheng Y, Wang S, Zhang H, Lee JS, Ni C,
Guo J, Chen E, Wang S, Acharya A, Chang TC, et al: A non-canonical
role for a small nucleolar RNA in ribosome biogenesis and
senescence. Cell. 187:4770–4789.e23. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Huo M, Rai SK, Nakatsu K, Deng Y and
Jijiwa M: Subverting the canon: Novel cancer-promoting functions
and mechanisms for snoRNAs. Int J Mol Sci. 25:29232024. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu B: Mapping snoRNA targets
transcriptome-wide with snoKARR-seq. ACS Chem Biol. 20:242–244.
2025. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sharma S, Langhendries JL, Watzinger P,
Kötter P, Entian KD and Lafontaine DLJ: Yeast Kre33 and human NAT10
are conserved 18S rRNA cytosine acetyltransferases that modify
tRNAs assisted by the adaptor Tan1/THUMPD1. Nucleic Acids Res.
43:2242–2258. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Song Z, Bae B, Schnabl S, Yuan F, De Zoysa
T, Akinyi MV, Le Roux CA, Choquet K, Whipple AJ and Van Nostrand
EL: Mapping snoRNA-target RNA interactions in an RNA-binding
protein-dependent manner with chimeric eCLIP. Genome Biol.
26:392025. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nostramo RT and Hopper AK: Beyond rRNA and
snRNA: tRNA as a 2'-O-methylation target for nucleolar and Cajal
body box C/D RNPs. Genes Dev. 33:739–740. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang M, Li K, Bai J, Van Damme R, Zhang
W, Alba M, Stiles BL, Chen JF and Lu Z: A snoRNA-tRNA modification
network governs codon-biased cellular states. Proc Natl Acad Sci
USA. 120:e23121261202023. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chauhan W, Sudharshan S, Kafle S and
Zennadi R: SnoRNAs: Exploring their implication in human diseases.
Int J Mol Sci. 25:72022024. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kudla G, Granneman S, Hahn D, Beggs JD and
Tollervey D: Cross-linking, ligation, and sequencing of hybrids
reveals RNA-RNA interactions in yeast. Proc Natl Acad Sci USA.
108:10010–10015. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Nguyen TC, Cao X, Yu P, Xiao S, Lu J,
Biase FH, Sridhar B, Huang N, Zhang K and Zhong S: Mapping RNA-RNA
interactome and RNA structure in vivo by MARIO. Nat Commun.
7:120232016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sharma E, Sterne-Weiler T, O'Hanlon D and
Blencowe BJ: Global mapping of human RNA-RNA interactions. Mol
Cell. 62:618–626. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Liu B, Wu T, Miao BA, Ji F, Liu S, Wang P,
Zhao Y, Zhong Y, Sundaram A, Zeng TB, et al: snoRNA-facilitated
protein secretion revealed by transcriptome-wide snoRNA target
identification. Cell. 188:465–483.e22. 2025. View Article : Google Scholar
|
|
42
|
Xu L, Zhao XH, Zhang YY, Zhang MY, Zhang
LY, Ye KH, Teng L, Han MM, Yue YM, Yang J, et al: SNORD80-guided
2'-O-methylation stabilizes the lncRNA GAS5 to regulate cellular
stress responses. Proc Natl Acad Sci USA. 122:e24189961222025.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhang Y, Zhao Z, Huang LA, Liu Y, Yao J,
Sun C, Li Y, Zhang Z, Ye Y, Yuan F, et al: Molecular mechanisms of
snoRNA-IL-15 crosstalk in adipocyte lipolysis and NK cell
rejuvenation. Cell Metab. 35:1457–1473.e13. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Leroy E, Challal D, Pelletier S, Goncalves
C, Menant A, Marchand V, Jaszczyszyn Y, van Dijk E, Naquin D,
Andreani J, et al: A bifunctional snoRNA with separable activities
in guiding rRNA 2'-O-methylation and scaffolding gametogenesis
effectors. Nat Commun. 16:32502025. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Han C, Sun LY, Luo XQ, Pan Q, Sun YM, Zeng
ZC, Chen TQ, Huang W, Fang K, Wang WT and Chen YQ:
Chromatin-associated orphan snoRNA regulates DNA damage-mediated
differentiation via a non-canonical complex. Cell Rep.
38:1104212022. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Raabe CA, Voss R, Kummerfeld DM, Brosius
J, Galiveti CR, Wolters A, Seggewiss J, Huge A, Skryabin BV and
Rozhdestvensky TS: Ectopic expression of Snord115 in choroid plexus
interferes with editing but not splicing of 5-Ht2c receptor
pre-mRNA in mice. Sci Rep. 9:43002019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Coulson RL, Powell WT, Yasui DH, Dileep G,
Resnick J and LaSalle JM: Prader-Willi locus Snord116 RNA
processing requires an active endogenous allele and neuron-specific
splicing by Rbfox3/NeuN. Hum Mol Genet. 27:4051–4060.
2018.PubMed/NCBI
|
|
48
|
Scott MS, Ono M, Yamada K, Endo A, Barton
GJ and Lamond AI: Human box C/D snoRNA processing conservation
across multiple cell types. Nucleic Acids Res. 40:3676–3688. 2012.
View Article : Google Scholar :
|
|
49
|
Michel CI, Holley CL, Scruggs BS, Sidhu R,
Brookheart RT, Listenberger LL, Behlke MA, Ory DS and Schaffer JE:
Small nucleolar RNAs U32a, U33, and U35a are critical mediators of
metabolic stress. Cell Metab. 14:33–44. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Macias S, Cordiner RA, Gautier P, Plass M
and Cáceres JF: DGCR8 acts as an adaptor for the exosome complex to
degrade double-stranded structured RNAs. Mol Cell. 60:873–885.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Macias S, Plass M, Stajuda A, Michlewski
G, Eyras E and Cáceres JF: DGCR8 HITS-CLIP reveals novel functions
for the microprocessor. Nat Struct Mol Biol. 19:760–766. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Taft RJ, Glazov EA, Lassmann T,
Hayashizaki Y, Carninci P and Mattick JS: Small RNAs derived from
snoRNAs. RNA. 15:1233–1240. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Bratkovič T, Božič J and Rogelj B:
Functional diversity of small nucleolar RNAs. Nucleic Acids Res.
48:1627–1651. 2020. View Article : Google Scholar
|
|
54
|
Ono M, Scott MS, Yamada K, Avolio F,
Barton GJ and Lamond AI: Identification of human miRNA precursors
that resemble box C/D snoRNAs. Nucleic Acids Res. 39:3879–3891.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Scott MS, Avolio F, Ono M, Lamond AI and
Barton GJ: Human miRNA precursors with box H/ACA snoRNA features.
PLoS Comput Biol. 5:e10005072009. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Thomson DW, Pillman KA, Anderson ML,
Lawrence DM, Toubia J, Goodall GJ and Bracken CP: Assessing the
gene regulatory properties of Argonaute-bound small RNAs of diverse
genomic origin. Nucleic Acids Res. 43:470–481. 2015. View Article : Google Scholar :
|
|
57
|
Brameier M, Herwig A, Reinhardt R, Walter
L and Gruber J: Human box C/D snoRNAs with miRNA like functions:
Expanding the range of regulatory RNAs. Nucleic Acids Res.
39:675–686. 2011. View Article : Google Scholar :
|
|
58
|
Zhong F, Zhou N, Wu K, Guo Y, Tan W, Zhang
H, Zhang X, Geng G, Pan T, Luo H, et al: A SnoRNA-derived piRNA
interacts with human interleukin-4 pre-mRNA and induces its decay
in nuclear exosomes. Nucleic Acids Res. 43:10474–10491.
2015.PubMed/NCBI
|
|
59
|
He X, Chen X, Zhang X, Duan X, Pan T, Hu
Q, Zhang Y, Zhong F, Liu J, Zhang H, et al: An Lnc RNA
(GAS5)/SnoRNA-derived piRNA induces activation of TRAIL gene by
site-specifically recruiting MLL/COMPASS-like complexes. Nucleic
Acids Res. 43:3712–3725. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Ono M, Yamada K, Avolio F, Scott MS, van
Koningsbruggen S, Barton GJ and Lamond AI: Analysis of human small
nucleolar RNAs (snoRNA) and the development of snoRNA modulator of
gene expression vectors. Mol Biol Cell. 21:1569–1584. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kishore S and Stamm S: The snoRNA HBII-52
regulates alternative splicing of the serotonin receptor 2C.
Science. 311:230–232. 2006. View Article : Google Scholar
|
|
62
|
Bratkovič T, Modic M, Camargo Ortega G,
Drukker M and Rogelj B: Neuronal differentiation induces SNORD115
expression and is accompanied by post-transcriptional changes of
serotonin receptor 2c mRNA. Sci Rep. 8:51012018. View Article : Google Scholar
|
|
63
|
Zhang X, Wang C, Xia S, Xiao F, Peng J,
Gao Y, Yu F, Wang C and Chen X: The emerging role of snoRNAs in
human disease. Genes Dis. 10:2064–2081. 2022. View Article : Google Scholar
|
|
64
|
Liu X, Zhang H, Fan Y, Cai D, Lei R, Wang
Q, Li Y, Shen L, Gu Y, Zhang Q, et al: SNORA28 promotes
proliferation and radioresistance in colorectal cancer cells
through the STAT3 pathway by increasing H3K9 acetylation in the
LIFR promoter. Adv Sci (Weinh). 11:e24053322024. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Xu C, Bian Z, Wang X, Niu N, Liu L, Xiao
Y, Zhu J, Huang N, Zhang Y, Chen Y, et al: SNORA56-mediated
pseudouridylation of 28 S rRNA inhibits ferroptosis and promotes
colorectal cancer proliferation by enhancing GCLC translation. J
Exp Clin Cancer Res. 42:3312023. View Article : Google Scholar
|
|
66
|
Bian Z, Xu C, Xie Y, Wang X, Chen Y, Mao
S, Wu Q, Zhu J, Huang N, Zhang Y, et al: SNORD11B-mediated
2'-O-methylation of primary let-7a in colorectal carcinogenesis.
Oncogene. 42:3035–3046. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Zhang H, Liu X, Zhang W, Deng J, Lin C, Qi
Z, Li Y, Gu Y, Wang Q, Shen L and Wang Z: Oncogene SCARNA12 as a
potential diagnostic biomarker for colorectal cancer. Mol Biomed.
4:372023. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Yoshida K, Toden S, Weng W, Shigeyasu K,
Miyoshi J, Turner J, Nagasaka T, Ma Y, Takayama T, Fujiwara T and
Goel A: SNORA21-an oncogenic small nucleolar RNA, with a prognostic
biomarker potential in human colorectal cancer. EBioMedicine.
22:68–77. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Shen L, Lin C, Lu W, He J, Wang Q, Huang
Y, Zheng X and Wang Z: Involvement of the oncogenic small nucleolar
RNA SNORA24 in regulation of p53 stability in colorectal cancer.
Cell Biol Toxicol. 39:1377–1394. 2023. View Article : Google Scholar
|
|
70
|
Okugawa Y, Toiyama Y, Toden S, Mitoma H,
Nagasaka T, Tanaka K, Inoue Y, Kusunoki M, Boland CR and Goel A:
Clinical significance of SNORA42 as an oncogene and A prognostic
biomarker in colorectal cancer. Gut. 66:107–117. 2017. View Article : Google Scholar
|
|
71
|
Zhang Z, Tao Y, Hua Q, Cai J, Ye X and Li
H: SNORA71A promotes colorectal cancer cell proliferation,
migration, and invasion. Biomed Res Int. 2020:82845762020.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Wu H, Qin W, Lu S, Wang X, Zhang J, Sun T,
Hu X, Li Y, Chen Q, Wang Y, et al: Long noncoding RNA ZFAS1
promoting small nucleolar RNA-mediated 2'-O-methylation via NOP58
recruitment in colorectal cancer. Mol Cancer. 19:952020. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Liu Y, Zhao C, Wang G, Chen J, Ju S, Huang
J and Wang X: SNORD1C maintains stemness and 5-FU resistance by
activation of Wnt signaling pathway in colorectal cancer. Cell
Death Discov. 8:2002022. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Yuan S, Wu Y, Wang Y, Chen J and Chu L: An
oncolytic adenovirus expressing SNORD44 and GAS5 exhibits antitumor
effect in colorectal cancer cells. Hum Gene Ther. 28:690–700. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Siprashvili Z, Webster DE, Johnston D,
Shenoy RM, Ungewickell AJ, Bhaduri A, Flockhart R, Zarnegar BJ, Che
Y, Meschi F, et al: The noncoding RNAs SNORD50A and SNORD50B bind
K-Ras and are recurrently deleted in human cancer. Nat Genet.
48:53–58. 2016. View Article : Google Scholar
|
|
76
|
Gómez-Matas J, Duran-Sanchon S, Lozano JJ,
Ferrero G, Tarallo S, Pardini B, Naccarati A, Castells A and
Gironella M: SnoRNA profiling in colorectal cancer and assessment
of non-invasive biomarker capacity by ddPCR in fecal samples.
iScience. 27:1092832024. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Shen L, Lu W, Huang Y, He J, Wang Q, Zheng
X and Wang Z: SNORD15B and SNORA5C: Novel diagnostic and prognostic
biomarkers for colorectal cancer. Biomed Res Int. 2022:82608002022.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Liu Y, Zhao C, Sun J, Wang G, Ju S, Qian C
and Wang X: Overexpression of small nucleolar RNA SNORD1C is
associated with unfavorable outcome in colorectal cancer.
Bioengineered. 12:8943–8952. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Tosar JP, Garcia-Silva MR and Cayota A:
Circulating SNORD57 rather than piR-54265 is a promising biomarker
for colorectal cancer: Common pitfalls in the study of somatic
piRNAs in cancer. RNA. 27:403–410. 2021. View Article : Google Scholar
|
|
80
|
Yuan Z, Chen Y, Liu S, Shen L and Wang X:
Evaluation of serum SNORA33 as a novel biomarker in colorectal
cancer screening. Biochem Biophys Res Commun. 791:1529112025.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Fu J, Liu G, Zhang X, Lei X, Liu Q, Qian
K, Tong Q, Qin W, Li Z, Cao Z, et al: TRPM8 promotes hepatocellular
carcinoma progression by inducing SNORA55 mediated
nuclear-mitochondrial communication. Cancer Gene Ther. 30:738–751.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Fang X, Yang D, Luo H, Wu S, Dong W, Xiao
J, Yuan S, Ni A, Zhang KJ, Liu XY and Chu L: SNORD126 promotes HCC
and CRC cell growth by activating the PI3K-AKT pathway through
FGFR2. J Mol Cell Biol. 9:243–255. 2017.
|
|
83
|
Xu W, Wu Y, Fang X, Zhang Y, Cai N, Wen J,
Liao J, Zhang B, Chen X and Chu L: SnoRD126 promotes the
proliferation of hepatocellular carcinoma cells through
transcriptional regulation of FGFR2 activation in combination with
hnRNPK. Aging (Albany NY). 13:13300–13317. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Wu L, Zheng J, Chen P, Liu Q and Yuan Y:
Small nucleolar RNA ACA11 promotes proliferation, migration and
invasion in hepatocellular carcinoma by targeting the PI3K/AKT
signaling pathway. Biomed Pharmacother. 90:705–712. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Wang H, Ma P, Liu P, Chen B and Liu Z:
Small nucleolar RNA U2_19 promotes hepatocellular carcinoma
progression by regulating Wnt/β-catenin signaling. Biochem Biophys
Res Commun. 500:351–356. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Wu L, Chang L, Wang H, Ma W, Peng Q and
Yuan Y: Clinical significance of C/D box small nucleolar RNA U76 as
an oncogene and a prognostic biomarker in hepatocellular carcinoma.
Clin Res Hepatol Gastroenterol. 42:82–91. 2018. View Article : Google Scholar
|
|
87
|
Liang J, Li G, Liao J, Huang Z, Wen J,
Wang Y, Chen Z, Cai G, Xu W, Ding Z, et al: Non-coding small
nucleolar RNA SNORD17 promotes the progression of hepatocellular
carcinoma through a positive feedback loop upon p53 inactivation.
Cell Death Differ. 29:988–1003. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Cao P, Yang A, Wang R, Xia X, Zhai Y, Li
Y, Yang F, Cui Y, Xie W, Liu Y, et al: Germline duplication of
SNORA18L5 increases risk for HBV-related hepatocellular carcinoma
by altering localization of ribosomal proteins and decreasing
levels of p53. Gastroenterology. 155:542–556. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Wang G, Li J, Yao Y, Liu Y, Xia P, Zhang
H, Yin M, Qin Z, Ma W and Yuan Y: Small nucleolar RNA 42 promotes
the growth of hepatocellular carcinoma through the p53 signaling
pathway. Cell Death Discov. 7:3472021. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Li C, Wu L, Liu P, Li K, Zhang Z, He Y,
Liu Q, Jiang P, Yang Z, Liu Z, et al: The C/D box small nucleolar
RNA SNORD52 regulated by Upf1 facilitates hepatocarcinogenesis by
stabilizing CDK1. Theranostics. 10:9348–9363. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Gu Y, Yi Z, Zhou Z, Wang J, Li S, Zhu P,
Liu N, Xu Y, He L, Wang Y and Fan Z: SNORD88B-mediated WRN
nucleolar trafficking drives self-renewal in liver cancer
initiating cells and hepatocarcinogenesis. Nat Commun. 15:67302024.
View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Mao LH, Chen SY, Li XQ, Xu F, Lei J, Wang
QL, Luo LY, Cao HY, Ge X, Ran T, et al: LncRNA-LALR1 upregulates
small nucleolar RNA SNORD72 to promote growth and invasion of
hepatocellular carcinoma. Aging (Albany NY). 12:4527–4546. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
McMahon M, Contreras A, Holm M, Uechi T,
Forester CM, Pang X, Jackson C, Calvert ME, Chen B, Quigley DA, et
al: A single H/ACA small nucleolar RNA mediates tumor suppression
downstream of oncogenic RAS. Elife. 8:e488472019. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Liu Z, Pang Y, Jia Y, Qin Q, Wang R, Li W,
Jing J, Liu H and Liu S: SNORA23 inhibits HCC tumorigenesis by
impairing the 2'-O-ribose methylation level of 28S rRNA. Cancer
Biol Med. 19:104–119. 2021.PubMed/NCBI
|
|
95
|
Yang H, Lin P, Wu HY, Li HY, He Y, Dang YW
and Chen G: Genomic analysis of small nucleolar RNAs identifies
distinct molecular and prognostic signature in hepatocellular
carcinoma. Oncol Rep. 40:3346–3358. 2018.PubMed/NCBI
|
|
96
|
Li G, He Y, Liu X, Zheng Z, Zhang M, Qin F
and Lan X: Small nucleolar RNA 47 promotes tumorigenesis by
regulating EMT markers in hepatocellular carcinoma. Minerva Med.
108:396–404. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Ma P, Wang H, Han L, Jing W, Zhou X and
Liu Z: Up-regulation of small nucleolar RNA 78 is correlated with
aggressive phenotype and poor prognosis of hepatocellular
carcinoma. Tumour Biol. 37:15753–15761. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Yu L, Zhang M, Ma Z and Wu S: Expression
of small nucleolar RNA SNORA51 and its clinical significance in
hepatocellular carcinoma. Oncol Lett. 27:552023. View Article : Google Scholar
|
|
99
|
Ding Y, Sun Z, Zhang S, Li Y, Han X, Xu Q,
Zhou L, Xu H, Bai Y, Xu C, et al: Downregulation of snoRNA SNORA52
and its clinical significance in hepatocellular carcinoma. Biomed
Res Int. 2021:70206372021. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Ding Y, Sun Z, Zhang S, Xu Q, Zhou L, Zhou
D, Li Y, Han X, Xu H, Bai Y, et al: Revealing the clinical
significance and prognostic value of small nucleolar RNA SNORD31 in
hepatocellular carcinoma. Biosci Rep. 40:BSR202014792020.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Xu G, Yang F, Ding CL, Zhao LJ, Ren H,
Zhao P, Wang W and Qi ZT: Small nucleolar RNA 113-1 suppresses
tumorigenesis in hepatocellular carcinoma. Mol Cancer. 13:2162014.
View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Ding Y, Sun Z, Zhang S, Zhou L, Xu Q, Zhou
D, Li Y, Han X, Xu H, Bai Y, et al: Identification of snoRNA
SNORA71A as a novel biomarker in prognosis of hepatocellular
carcinoma. Dis Markers. 2020:88799442020. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Xie Q, Zhang D, Ye H, Wu Z, Sun Y and Shen
H: Identification of key snoRNAs serves as biomarkers for
hepatocellular carcinoma by bioinformatics methods. Medicine
(Baltimore). 101:e308132022. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Mallela VR, Kasi PB, Shetti D, Trailin A,
Cervenkova L, Palek R, Daum O, Liska V, Hemminki K and
Ambrozkiewicz F: Small nucleolar RNA expression profiles: A
potential prognostic biomarker for non-viral hepatocellular
carcinoma. Noncoding RNA Res. 9:1133–1139. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Zhao X, Chen G, Wu Y, Li X, Zhang Z, Xie L
and Song X and Song X: TEP SNORD12B, SNORA63, and SNORD14E as novel
biomarkers for hepatitis B virus-related hepatocellular carcinoma
(HBV-related HCC). Cancer Cell Int. 24:32024. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Bao B, Tian M, Wang X, Yang C, Qu J, Zhou
S, Cheng Y, Tong Q and Zheng L: SNORA37/CMTR1/ELAVL1 feedback loop
drives gastric cancer progression via facilitating CD44 alternative
splicing. J Exp Clin Cancer Res. 44:152025. View Article : Google Scholar
|
|
107
|
Zhang C, Zhao LM, Wu H, Tian G, Dai SL,
Zhao RY and Shan BE: C/D-box Snord105b promotes tumorigenesis in
gastric cancer via ALDOA/C-Myc pathway. Cell Physiol Biochem.
45:2471–2482. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Wu F, Wu W, Ma X, Xu H and Zhang D: The
snoRNA SNORA21 promotes gastric tumorigenesis by attenuating P53
activity through CHK1 phosphorylation inhibition and PERP-dependent
feedback loops. Cell Signal. 139:1123262026. View Article : Google Scholar
|
|
109
|
Li Y, Yu S, Wang X, Ye X, He B, Quan M and
Gao Y: SRPK1 facilitates tumor cell growth via modulating the small
nucleolar RNA expression in gastric cancer. J Cell Physiol.
234:13582–13591. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Wang K, Wang S, Zhang Y, Xie L and Song X
and Song X: SNORD88C guided 2'-O-methylation of 28S rRNA regulates
SCD1 translation to inhibit autophagy and promote growth and
metastasis in non-small cell lung cancer. Cell Death Differ.
30:341–355. 2023. View Article : Google Scholar
|
|
111
|
Mei YP, Liao JP, Shen J, Yu L, Liu BL, Liu
L, Li RY, Ji L, Dorsey SG, Jiang ZR, et al: Small nucleolar RNA 42
acts as an oncogene in lung tumorigenesis. Oncogene. 31:2794–2804.
2012. View Article : Google Scholar
|
|
112
|
Tang G, Zeng Z, Sun W, Li S, You C, Tang
F, Peng S, Ma S, Luo Y, Xu J, et al: Small nucleolar RNA 71A
promotes lung cancer cell proliferation, migration and invasion via
MAPK/ERK pathway. J Cancer. 10:2261–2275. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Cui C, Liu Y, Gerloff D, Rohde C, Pauli C,
Köhn M, Misiak D, Oellerich T, Schwartz S, Schmidt LH, et al: NOP10
predicts lung cancer prognosis and its associated small nucleolar
RNAs drive proliferation and migration. Oncogene. 40:909–921. 2021.
View Article : Google Scholar :
|
|
114
|
Zhuo Y, Li S, Hu W, Zhang Y, Shi Y, Zhang
F, Zhang J, Wang J, Liao M, Chen J, et al: Targeting SNORA38B
attenuates tumorigenesis and sensitizes immune checkpoint blockade
in non-small cell lung cancer by remodeling the tumor
microenvironment via regulation of GAB2/AKT/mTOR signaling pathway.
J Immunother Cancer. 10:e0041132022. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Wang K and Song X, Wang S, Li X, Zhang Z,
Xie L and Song X: Plasma SNORD42B and SNORD111 as potential
biomarkers for early diagnosis of non-small cell lung cancer. J
Clin Lab Anal. 36:e247402022. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Wang K and Song X, Li X, Zhang Z, Xie L
and Song X: Plasma SNORD83A as a potential biomarker for early
diagnosis of non-small-cell lung cancer. Future Oncol. 18:821–832.
2022. View Article : Google Scholar
|
|
117
|
Zhou H, Yao Y, Li Y, Guo N, Zhang H, Wang
Z, Chen Y and Dai G: Identification of small nucleolar RNA SNORD60
as a potential biomarker and its clinical significance in lung
adenocarcinoma. Biomed Res Int. 2022:55011712022. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Gao L, Ma J, Mannoor K, Guarnera MA,
Shetty A, Zhan M, Xing L, Stass SA and Jiang F: Genome-wide small
nucleolar RNA expression analysis of lung cancer by next-generation
deep sequencing. Int J Cancer. 136:E623–E629. 2015. View Article : Google Scholar
|
|
119
|
Sun Y, Chen E, Li Y, Ye D, Cai Y, Wang Q,
Li Q and Zhang X: H/ACA box small nucleolar RNA 7B acts as an
oncogene and a potential prognostic biomarker in breast cancer.
Cancer Cell Int. 19:1252019. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Su X, Feng C, Wang S, Shi L, Gu Q, Zhang
H, Lan X, Zhao Y, Qiang W, Ji M and Hou P: The noncoding RNAs
SNORD50A and SNORD50B-mediated TRIM21-GMPS interaction promotes the
growth of p53 wild-type breast cancers by degrading p53. Cell Death
Differ. 28:2450–2464. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Langhendries JL, Nicolas E, Doumont G,
Goldman S and Lafontaine DLJ: The human box C/D snoRNAs U3 and U8
are required for pre-rRNA processing and tumorigenesis. Oncotarget.
7:59519–59534. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Hu T, Lu C, Xia Y, Wu L, Song J, Chen C
and Wang Q: Small nucleolar RNA SNORA71A promotes
epithelial-mesenchymal transition by maintaining ROCK2 mRNA
stability in breast cancer. Mol Oncol. 16:1947–1965. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Zhang W, Song X, Jin Z, Zhang Y, Li S, Jin
F and Zheng A: U2AF2-SNORA68 promotes triple-negative breast cancer
stemness through the translocation of RPL23 from nucleoplasm to
nucleolus and c-Myc expression. Breast Cancer Res. 26:602024.
View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Li S, Jin Z, Song X, Ma J, Peng Z, Yu H,
Song J, Zhang Y, Sun X, He M, et al: The small nucleolar RNA
SNORA51 enhances breast cancer stem cell-like properties via the
RPL3/NPM1/c-MYC pathway. Mol Carcinog. 63:1117–1132. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Chao YL, Zhou KI, Forbes KK, Porrello A,
Gentile GM, Zhu Y, Chack AC, John Mary DJS, Liu H, Cockman E, et
al: Snord67 promotes breast cancer metastasis by guiding U6
modification and modulating the splicing landscape. Nat Commun.
16:41182025. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Han Q, Zhou Y, Dong Z, Wang W, Wang M,
Pang M, Song X, Chen B and Zheng A: SNORA47 affects stemness and
chemotherapy sensitivity via EBF3/RPL11/c-Myc axis in luminal A
breast cancer. Mol Med. 31:1502025. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Li JN, Loh ZJ, Chen HW, Lee IY, Tsai JH
and Chen PS: SnoRNA U50A mediates everolimus resistance in breast
cancer through mTOR downregulation. Transl Oncol. 48:1020622024.
View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Su H, Xu T, Ganapathy S, Shadfan M, Long
M, Huang TH, Thompson I and Yuan ZM: Elevated snoRNA biogenesis is
essential in breast cancer. Oncogene. 33:1348–1358. 2014.
View Article : Google Scholar
|
|
129
|
Escuin D, Bell O, García-Valdecasas B,
Clos M, Larrañaga I, López-Vilaró L, Mora J, Andrés M, Arqueros C
and Barnadas A: Small non-coding RNAs and their role in
locoregional metastasis and outcomes in early-stage breast cancer
patients. Int J Mol Sci. 25:39822024. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Li X, Zhao X, Xie L and Song X and Song X:
Identification of four snoRNAs (SNORD16, SNORA73B, SCARNA4, and
SNORD49B) as novel non-invasive biomarkers for diagnosis of breast
cancer. Cancer Cell Int. 24:552024. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Song J, Zheng A, Li S, Zhang W, Zhang M,
Li X, Jin F and Ji Z: Clinical significance and prognostic value of
small nucleolar RNA SNORA38 in breast cancer. Front Oncol.
12:9300242022. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Wang B, Zhao Y, Li Y, Xu Y, Chen Y, Jiang
Q, Yao D, Zhang L, Hu X, Fu C, et al: A plasma SNORD33 signature
predicts platinum benefit in metastatic triple-negative breast
cancer patients. Mol Cancer. 21:222022. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Chen S, Li QH, Chen X, Bao HJ, Wu W, Shen
F, Lu BF, Jiang RQ, Zong ZH and Zhao Y: SNORA70E promotes the
occurrence and development of ovarian cancer through
pseudouridylation modification of RAP1B and alternative splicing of
PARPBP. J Cell Mol Med. 26:5150–5164. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Zhang L, Ma R, Gao M, Zhao Y, Lv X, Zhu W,
Han L, Su P, Fan Y, Yan Y, et al: SNORA72 activates the
Notch1/c-Myc pathway to promote stemness transformation of ovarian
cancer cells. Front Cell Dev Biol. 8:5830872020. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Zhu W, Niu J, He M, Zhang L, Lv X, Liu F,
Jiang L, Zhang J, Yu Z, Zhao L, et al: SNORD89 promotes stemness
phenotype of ovarian cancer cells by regulating Notch1-c-Myc
pathway. J Transl Med. 17:2592019. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Chen S, Wen JT, Zhang S, Wang JL, Yuan J,
Bao HJ, Chen X and Zhao Y: SNORD9 promotes ovarian cancer
tumorigenesis via METTL3/IGF2BP2-mediated NFYA m6A modification and
is a potential target for antisense oligonucleotide therapy. Life
Sci. 368:1235272025. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Faucher-Giguère L, Roy A,
Deschamps-Francoeur G, Couture S, Nottingham RM, Lambowitz AM,
Scott MS and Abou Elela S: High-grade ovarian cancer associated
H/ACA snoRNAs promote cancer cell proliferation and survival. NAR
Cancer. 4:zcab0502022. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Zhu W, Zhang T, Luan S, Kong Q, Hu W, Zou
X, Zheng F and Han W: Identification of a novel nine-SnoRNA
signature with potential prognostic and therapeutic value in
ovarian cancer. Cancer Med. 11:2159–2170. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Chen X, Li QH, Xie BM, Ji YM, Han Y and
Zhao Y: SNORA73B promotes endometrial cancer progression through
targeting MIB1 and regulating host gene RCC1 alternative splicing.
J Cell Mol Med. 27:2890–2905. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Wu W, Chen X, Liu X, Bao HJ, Li QH, Xian
JY, Lu BF, Zhao Y and Chen S: SNORD60 promotes the tumorigenesis
and progression of endometrial cancer through binding PIK3CA and
regulating PI3K/AKT/mTOR signaling pathway. Mol Carcinog.
62:413–426. 2023. View Article : Google Scholar
|
|
141
|
Bao HJ, Chen X, Liu X, Wu W, Li QH, Xian
JY, Zhao Y and Chen S: Box C/D snoRNA SNORD89 influences the
occurrence and development of endometrial cancer through
2'-O-methylation modification of Bim. Cell Death Discov. 8:3092022.
View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Lu B, Chen X, Liu X, Chen J, Qin H, Chen S
and Zhao Y: C/D box small nucleolar RNA SNORD104 promotes
endometrial cancer by regulating the 2'-O-methylation of PARP1. J
Transl Med. 20:6182022. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Xian JY, Wu W, Chen X, Bao HJ, Zhang S,
Sheng XJ and Chen S: SNORD99 promotes endometrial cancer
development by inhibiting GSDMD-mediated pyroptosis through
2'-O-methylation modification. J Cell Mol Med. 28:e185002024.
View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Wen JT, Chen X, Liu X, Xie BM, Chen JW,
Qin HL and Zhao Y: Small nucleolar RNA and C/D Box 15B regulate the
TRIM25/P53 complex to promote the development of endometrial
cancer. J Oncol. 2022:77627082022. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Dong W, Liu Y, Wang P, Ruan X, Liu L, Xue
Y, Ma T, E T, Wang D, Yang C, et al: U3 snoRNA-mediated degradation
of ZBTB7A regulates aerobic glycolysis in isocitrate dehydrogenase
1 wild-type glioblastoma cells. CNS Neurosci Ther. 29:2811–2825.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Xu C, Chen G, Yu B, Sun B, Zhang Y, Zhang
M, Yang Y, Xiao Y, Cheng SY, Li Y and Feng H: TRIM24 cooperates
with ras mutation to drive glioma progression through snoRNA
recruitment of PHAX and DNA-PKcs. Adv Sci (Weinh). 11:e24000232024.
View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Cui Z, Liu X, E T, Lin H, Wang D, Liu Y,
Ruan X, Wang P, Liu L and Xue Y: Effect of SNORD113-3/ADAR2 on
glycolipid metabolism in glioblastoma via A-to-I editing of PHKA2.
Cell Mol Biol Lett. 30:52025. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Zhou Y, Yin W, Kuang Y, Wu Z, Huang H, Liu
W, Jiang X and Ren C: A prognostic signature based on snoRNA
predicts the overall survival of lower-grade glioma patients. Front
Immunol. 14:11383632023. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Yue X, Zheng Y, Li L, Yang Z, Chen Z, Wang
Y, Wang Z, Zhang D, Bian E and Zhao B: Integrative analysis of a
novel 5 methylated snoRNA genes prognostic signature in patients
with glioma. Epigenomics. 14:1089–1104. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Sun J, Zhao S, Ren X, Du Q, Chang Q, Guo
W, Qiu L, Yang L, Zhang N, Zhao Z, et al: SNORA33 promotes clear
cell renal cell carcinoma development and resistance to sunitinib
through triggering the JAK/STAT pathway. IUBMB Life. 77:e700582025.
View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Zhang Y, Shang X, Yu M, Bi Z, Wang K,
Zhang Q, Xie L and Song X and Song X: A three-snoRNA signature:
SNORD15A, SNORD35B and SNORD60 as novel biomarker for renal cell
carcinoma. Cancer Cell Int. 23:1362023. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Shang X and Song X, Wang K, Yu M, Ding S,
Dong X, Xie L and Song X: SNORD63 and SNORD96A as the non-invasive
diagnostic biomarkers for clear cell renal cell carcinoma. Cancer
Cell Int. 21:562021. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Grützmann K, Salomo K, Krüger A,
Lohse-Fischer A, Erdmann K, Seifert M, Baretton G, Aust D, William
D, Schröck E, et al: Identification of novel snoRNA-based
biomarkers for clear cell renal cell carcinoma from urine-derived
extracellular vesicles. Biol Direct. 19:382024. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Li Q, Xie B, Chen X, Lu B, Chen S, Sheng X
and Zhao Y: SNORD6 promotes cervical cancer progression by
accelerating E6-mediated p53 degradation. Cell Death Discov.
9:1922023. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Lu Q, Wang J, Tao Y, Zhong J, Zhang Z,
Feng C, Wang X, Li T, He R, Wang Q and Xie Y: Small cajal
body-specific RNA12 promotes carcinogenesis through modulating
extracellular matrix signaling in bladder cancer. Cancers (Basel).
16:4832024. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Chen F, Zheng Y, Zhou H and Li C: The
regulatory role of SNORD35A in pancreatic cancer involves the
HGF/C-Met pathway. Cancer Biother Radiopharm. 39:211–222. 2024.
|
|
157
|
Zeng H, Pan J, Hu C, Yang J, Li J, Tan T,
Zheng M, Shen Y, Yang T, Deng Y and Zou Y: SNHG25 facilitates
SNORA50C accumulation to stabilize HDAC1 in neuroblastoma cells.
Cell Death Dis. 13:5972022. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Zheng Y, Liu F, Huang Y, Feng Y, Yang X,
Wu J, Yang X, Lin X, Jiang L, Zeng T, et al: SNORA58 facilitates
radioresistance via suppressing JNK1-mediated ferroptosis in
esophageal squamous cell carcinoma. Adv Sci (Weinh). 12:e085152025.
View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Yuan H, Ge G, Liu L, Hu S, Tian M, Nie Y,
Zhao Z and Song Y: Clinofibrate disrupts the SNORA80B/YTHDC1-driven
M6A modification to suppress cholesterol metabolism and cisplatin
resistance in ESCC. Adv Sci (Weinh). 13:e095742026. View Article : Google Scholar :
|
|
160
|
Zhang Q, Bi Z and Song X, Zhang Y, Wang S,
Xie L and Song X: Tumor-educated platelet SNORA58, SNORA68 and
SNORD93 as novel diagnostic biomarkers for esophageal cancer.
Future Oncol. 19:651–661. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Coley AB, Stahly AN, Kasukurthi MV,
Barchie AA, Hutcheson SB, Houserova D, Huang Y, Watters BC, King
VM, Dean MA, et al: MicroRNA-like snoRNA-derived RNAs (sdRNAs)
promote castration-resistant prostate cancer. Cells. 11:13022022.
View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Patterson DG, Roberts JT, King VM,
Houserova D, Barnhill EC, Crucello A, Polska CJ, Brantley LW,
Kaufman GC, Nguyen M, et al: Human snoRNA-93 is processed into a
microRNA-like RNA that promotes breast cancer cell invasion. NPJ
Breast Cancer. 3:252017. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Müller S, Raulefs S, Bruns P, Afonso-Grunz
F, Plötner A, Thermann R, Jäger C, Schlitter AM, Kong B, Regel I,
et al: Next-generation sequencing reveals novel differentially
regulated mRNAs, lncRNAs, miRNAs, sdRNAs and a piRNA in pancreatic
cancer. Mol Cancer. 14:942015. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Chow RD and Chen S: Sno-derived RNAs are
prevalent molecular markers of cancer immunity. Oncogene.
37:6442–6462. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Srinivasan S, Yeri A, Cheah PS, Chung A,
Danielson K, De Hoff P, Filant J, Laurent CD, Laurent LD, Magee R,
et al: Small RNA sequencing across diverse biofluids identifies
optimal methods for exRNA isolation. Cell. 177:446–462.e16. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Vickers KC, Palmisano BT, Shoucri BM,
Shamburek RD and Remaley AT: MicroRNAs are transported in plasma
and delivered to recipient cells by high-density lipoproteins. Nat
Cell Biol. 13:423–433. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Li L, Zhang Z, Xu W, Wang J and Feng X:
The diagnostic value of serum exosomal SNORD116 and SNORA21 for
NSCLC patients. Clin Transl Oncol. 27:650–659. 2025. View Article : Google Scholar
|
|
168
|
Kitagawa T, Taniuchi K, Tsuboi M,
Sakaguchi M, Kohsaki T, Okabayashi T and Saibara T: Circulating
pancreatic cancer exosomal RNAs for detection of pancreatic cancer.
Mol Oncol. 13:212–227. 2019. View Article : Google Scholar :
|
|
169
|
Shi J, Zhou T and Chen Q: Exploring the
expanding universe of small RNAs. Nat Cell Biol. 24:415–423. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Watkins CP, Zhang W, Wylder AC, Katanski
CD and Pan T: A multiplex platform for small RNA sequencing
elucidates multifaceted tRNA stress response and translational
regulation. Nat Commun. 13:24912022. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Kulkarni JA, Witzigmann D, Thomson SB,
Chen S, Leavitt BR, Cullis PR and van der Meel R: The current
landscape of nucleic acid therapeutics. Nat Nanotechnol.
16:630–643. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Zhou Z, Gu Y, Yi Z, Wang J, Xiong Z, Guo
H, Du Y, Zhu X, He L, Ren W, et al: SNORA74A drives self-renewal of
liver cancer stem cells and hepatocarcinogenesis through activation
of Notch3 signaling. Adv Sci (Weinh). 12:e25040542025. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Cui L, Nakano K, Obchoei S, Setoguchi K,
Matsumoto M, Yamamoto T, Obika S, Shimada K and Hiraoka N: Small
nucleolar noncoding RNA SNORA23, up-regulated in human pancreatic
ductal adenocarcinoma, regulates expression of spectrin
repeat-containing nuclear envelope 2 to promote growth and
metastasis of xenograft tumors in mice. Gastroenterology.
153:292–306.e2. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Wang Y, An H, Zhang Y, Lyu QR and Zhang Z:
SNORA13 antisense oligonucleotides enhances the therapeutical
effects of 5-fluorouracil in colon adenocarcinoma. Front Pharmacol.
16:15646822025. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Miller TM, Pestronk A, David W, Rothstein
J, Simpson E, Appel SH, Andres PL, Mahoney K, Allred P, Alexander
K, et al: An antisense oligonucleotide against SOD1 delivered
intrathecally for patients with SOD1 familial amyotrophic lateral
sclerosis: A phase 1, randomised, first-in-man study. Lancet
Neurol. 12:435–442. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Sterling T and Irwin JJ: ZINC 15-ligand
discovery for everyone. J Chem Inf Model. 55:2324–2337. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Velagapudi SP, Costales MG, Vummidi BR,
Nakai Y, Angelbello AJ, Tran T, Haniff HS, Matsumoto Y, Wang ZF,
Chatterjee AK, et al: Approved anti-cancer drugs target oncogenic
non-coding RNAs. Cell Chem Biol. 25:1086–1094.e7. 2018. View Article : Google Scholar : PubMed/NCBI
|














