Introduction
Acute myeloid meukemia (AML) is an aggressive,
heterogeneous malignancy defined by the clonal expansion of
immature myeloid blasts. Despite therapeutic advances, relapse
remains a huge challenge due to a reservoir of therapy-resistant
leukemic stem cells (LSCs) (1-3).
The bone marrow (BM) microenvironment, or niche, plays a crucial
role in nurturing these LSCs and promoting chemoresistance
(4).
Purinergic signaling, mediated by extracellular
nucleotides such as adenosine triphosphate (ATP) and their
receptors (P2X and P2Y families), is increasingly recognized as a
key regulator within the BM niche. Among these receptors, P2X7
receptor (P2X7R) is particularly notable due to its high
sensitivity to elevated extracellular adenosine triphosphate
(eATP), a common feature of tumor microenvironments (TME). As a
ligand-gated ion channel, its activation induces rapid
Na+ and Ca2+ influx and K+ efflux,
which promote cancer cell proliferation and migration (5,6).
Beyond the formation of non-selective ion channel pores, P2X7R can
form macropores in the cell membrane, particularly under sustained
high eATP concentrations. The formation of these macropores permits
the passage of large molecules and ultimately results in cell death
(6). The dual nature of P2X7R,
facilitating both pro-tumorigenic responses and cell death, is
often critically dysregulated in cancers.
P2X7R is highly expressed on hematopoietic cells,
where it functions as a key sensor of eATP, a ubiquitous 'danger
signal' released during cellular stress, injury, or death. The role
of P2X7R in hematopoiesis and immune regulation is multifaceted and
context-dependent. In hematopoietic stem and progenitor cells
(HSPCs), P2X7R signaling influences survival, proliferation, and
differentiation (7). In mature
immune cells, the effects of P2X7R activation are diverse. P2X7R
activation modulates macrophage polarization, enhances inflammatory
responses, and promotes antigen presentation (8,9).
In dendritic cells (DCs), it promotes cell maturation,
proinflammatory cytokine release, and antigen presentation
(10). In addition, it can
mediate cluster of differentiation (CD)8+ T-cell
activation and apoptosis (11).
In AML, the concentration of eATP is significantly
higher in the endosteal niche than in the vascular niche. Moreover,
LSCs preferentially localize to the endosteal region, a
distribution characteristic that provides crucial support for
maintaining the functional activity of LSCs (12). P2X7R is frequently overexpressed
on LSCs and AML blasts (13),
which constitute the self-renewing population responsible for
disease initiation, relapse, and chemoresistance. Within the unique
BM niche of AML, elevated eATP provides a persistent activation
signal for P2X7R. This signaling contributes to the maintenance of
LSCs, and supports the survival and proliferation of AML blasts
(5,12). Furthermore, by regulating the
metabolic status of LSCs, the P2X7R signaling pathway modulates
their homing and self-renewal, thereby driving disease progression
(12).
Given the central role in AML pathophysiology, P2X7R
has emerged as a promising biomarker for prognosis and a potential
therapeutic target. Small-molecule antagonists that block the
ATP-binding site to inhibit downstream signaling, and anti-P2X7R
antibodies that specifically eliminate P2X7R-expressing leukemia
cells, have been designed and are under investigation (7,13,14). The other promising avenue may
involve combining P2X7R inhibitors with conventional chemotherapy
(15). However, P2X7R targeted
therapy is still in the early research stage. It is speculated that
the main reason is the failure to achieve ideal therapeutic effects
and ensure the in vivo safety, indicating that further
exploration and improvement are still needed.
While recent reviews have discussed eATP and P2X7R
signaling across various cancers, the present review systematically
and comprehensively summarizes the roles of eATP and P2X7R in AML,
including the context-dependent duality of P2X7R in hematopoietic
cells, LSCs, and AML blasts, and the potential for targeting P2X7R
in AML therapy.
Structure and function of P2X7R
Structure of P2X7R
Human P2X7R is encoded by the P2X7R gene
located at chromosome 12q24.31. It typically assembles as a
homotrimer of three identical subunits, although heterotrimeric
assemblies also exist (16-20). Each subunit comprises ~595 amino
acids, contains a short intracellular N-terminus (26 amino acids),
two transmembrane helices (~24 amino acids each), a large
extracellular ligand-binding loop (282 amino acids), and a
characteristically long intracellular C-terminal tail. P2X7R
possesses the longest C-terminal intracellular domain (239 amino
acids) among all the P2X family members and mediates a bifunctional
response on stimulation (21,22). The 'dolphin anatomy' is commonly
adopted to identify specific regions in the tertiary structure of
P2X subunits and to describe P2X7R architecture. Each subunit
resembles the shape of a dolphin, with the transmembrane helices
corresponding to the flukes and the extracellular region forming
the body (Fig. 1) (21).
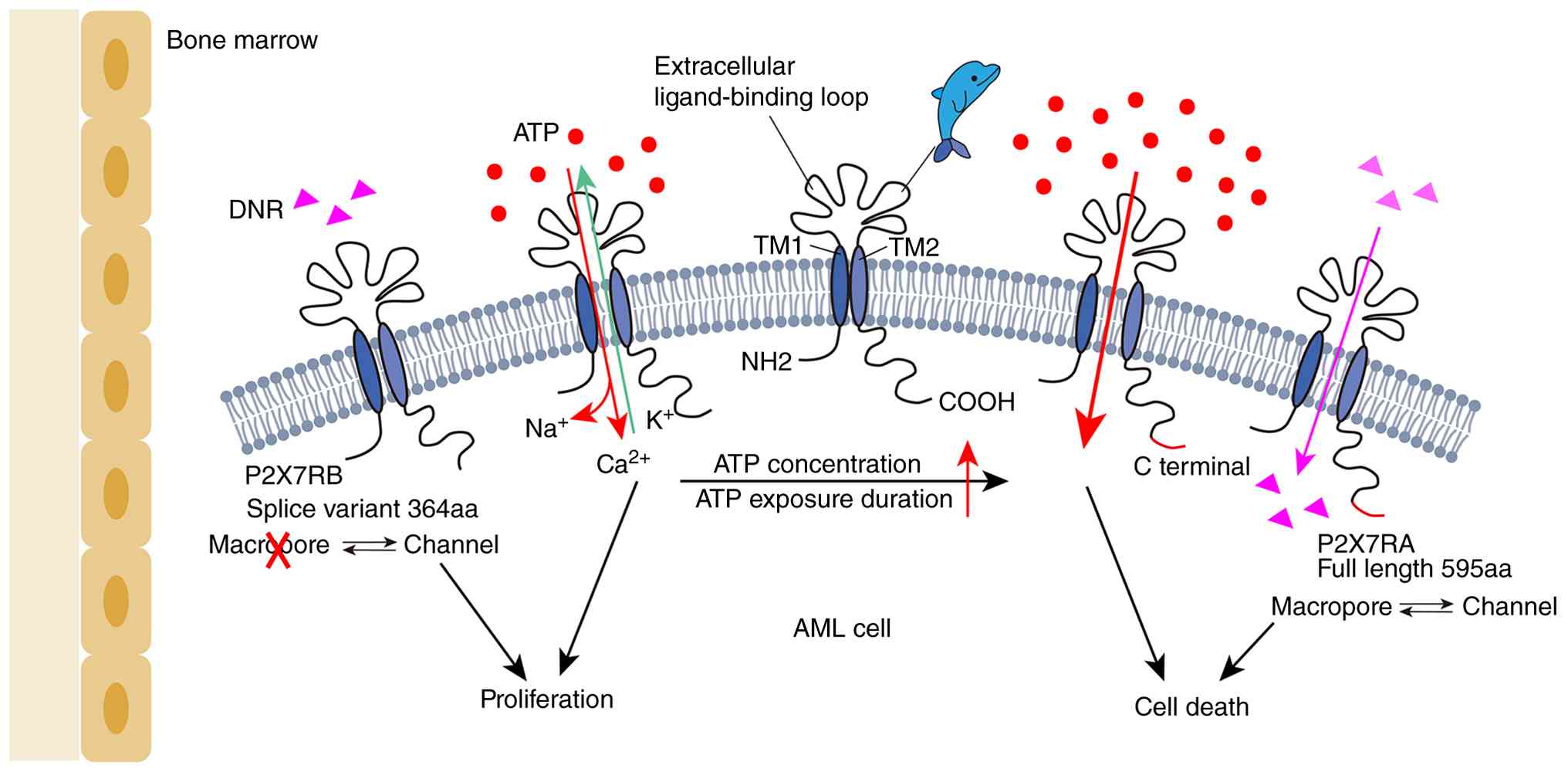 | Figure 1Structure and function of P2X7R.
P2X7R presents a typical dolphin-like structural conformation
(21). Low concentrations of ATP
stimulation rapidly open P2X7R as an ion channel, while sustained
activation, particularly under high concentrations of eATP,
gradually opens macropores (24-26). In AML, the full-length of P2X7RA
mediates both ion channel activation and macropore formation, while
the P2X7RB variant only retains basic channel activity due to the
lack of the C-terminal domain. The macropores formed by P2X7RA
permit the passage of DNR, thereby promoting the chemosensitivity
of AML cells (37). P2X7R, P2X7
receptor; ATP, adenosine triphosphate; eATP, extracellular
adenosine triphosphate; AML, acute myeloid leukemia; DNR,
daunorubicin; TM2, second transmembrane helices. |
Dual functional characteristics of
P2X7R
Differences in ATP concentration and duration of
action can switch P2X7R between ion channel and large-pore
conformations, thereby mediating the dual biological effects of
cell survival or cell death. Receptor activation initiates when a
low concentration of ATP ligand (≥100 μM, sub-millimolar
range) binds to the orthosteric site of P2X7R (11) or non-nucleotide agonists engage
allosteric sites. Ligand binding induces conformational changes
within the extracellular domain that propagate to the transmembrane
helices (23). This rearrangement
specifically alters the position of the second transmembrane
helices (TM2), widening the central pore to permit rapid cation
flux (Na+ and Ca2+ influx, K+
efflux) within milliseconds, representing its canonical ion channel
function (Fig. 1) (24). Among these, the calcium channel
activity of P2X7R plays a central role in promoting proliferation
(6). Sustained activation,
particularly under high concentrations of eATP (≥0.3-0.5 mM)
(23), induces a conformational
change in P2X7R, and the ion channel gradually transforms into
large membrane pores (25,26).
This large conductance pore permits the passage of molecules up to
~900 Da (27,28), mediating cell death by disrupting
intracellular homeostasis and inducing necrotic cell lysis
(6). The extended C-terminal tail
is indispensable for this pore dilation (21,29), likely through interactions with
membrane lipids (such as phosphatidylinositol 4,5-bisphosphate
(PIP2)] and cytosolic proteins that relay conformational
signals. However, the precise mechanism of macropore formation
remains actively debated, potentially involving: i) Intrinsic
dilation of the P2X7R pore itself; ii) recruitment of an accessory
protein (such as pannexin-1 hemichannels) to form the conduit; or
iii) oligomerization of additional P2X7R subunits (30).
Differences in P2X7R variants also play an important
role in regulating its function. Human P2X7R exhibits significant
polymorphism, with at least nine splice variants identified. The
full-length variant (P2X7RA) requires its intact C-terminus for
macropore function, as truncation abolishes large solute uptake.
P2X7RB, a common splice variant lacking the final 249 C-terminal
amino acids, retains ion channel capability but cannot form
functional macropores independently (Fig. 1) (31-36). This is mainly manifested in AML as
distinct sensitivity to daunorubicin (DNR). AML blasts with high
P2X7RA expression are more prone to cell death upon exposure to DNR
due to its large pore formation. By contrast, cells highly
expressing P2X7RB can only form ion channels, leading to strong
cell viability and even resistance to the cytotoxic effect of DNR
(37). Notably, co-expression of
P2X7RB with P2X7RA generates heterotrimers that exhibit enhanced
ATP affinity, amplified macropore activity, and increased support
for cellular energy metabolism and proliferation compared with
P2X7RA homotrimers (23).
Another variant, nfP2X7R (potentially arising from
alternative splicing or mutations), primarily resides
intracellularly but translocates to the plasma membrane under high
eATP conditions. Similar to P2X7RB, nfP2X7R functions as a small
ion channel that promotes tumor survival and proliferation but
lacks ethidium-permeable macropore capability. Under stimulation by
high concentrations of eATP in the TME, nfP2X7R expression is
upregulated while the expression of functional P2X7R is
downregulated, thereby protecting tumor cells (38). These variant-specific functional
differences highlight the complex regulation of P2X7R in
physiological and disease conditions.
The human P2X7R gene also exhibits a high
degree of polymorphism. Notably, >13,000 single nucleotide
polymorphisms (SNPs) associated with P2X7R have been
identified in the human genome (39). These SNPs can affect receptor
function by causing amino acid substitutions or altering the
eATP-binding sites, ultimately leading to the loss of the original
channel function or large-pore function of P2X7R (40-42). Previous research on P2X7R
SNPs in leukemia has mainly focused on chronic lymphocytic leukemia
(CLL). Researchers analyzed the 1513 A>C polymorphism in
populations of different geographic origins, but no association
between this SNP and the progression of CLL was found (39).
Dynamic alterations in the TME, particularly eATP
fluctuations induced by chemotherapy, can directly regulate the
activation of P2X7R. In AML, certain chemotherapeutic agents, such
as DNR, can induce immunogenic cell death of cancer cells, thereby
triggering effective antitumor T-cell immune responses (43). The eATP then acts on P2X7R to
further activate downstream signaling pathways.
Expression and function of P2X7R in
hematopoietic cells in AML
P2X7R is widely expressed in human hematopoietic
cells, encompassing HSPCs as well as differentiated lineages such
as monocytes/macrophages, DCs, and lymphocytes (8,44).
Its function exhibits significanT cell specificity and context
dependence, especially in AML (Fig.
2).
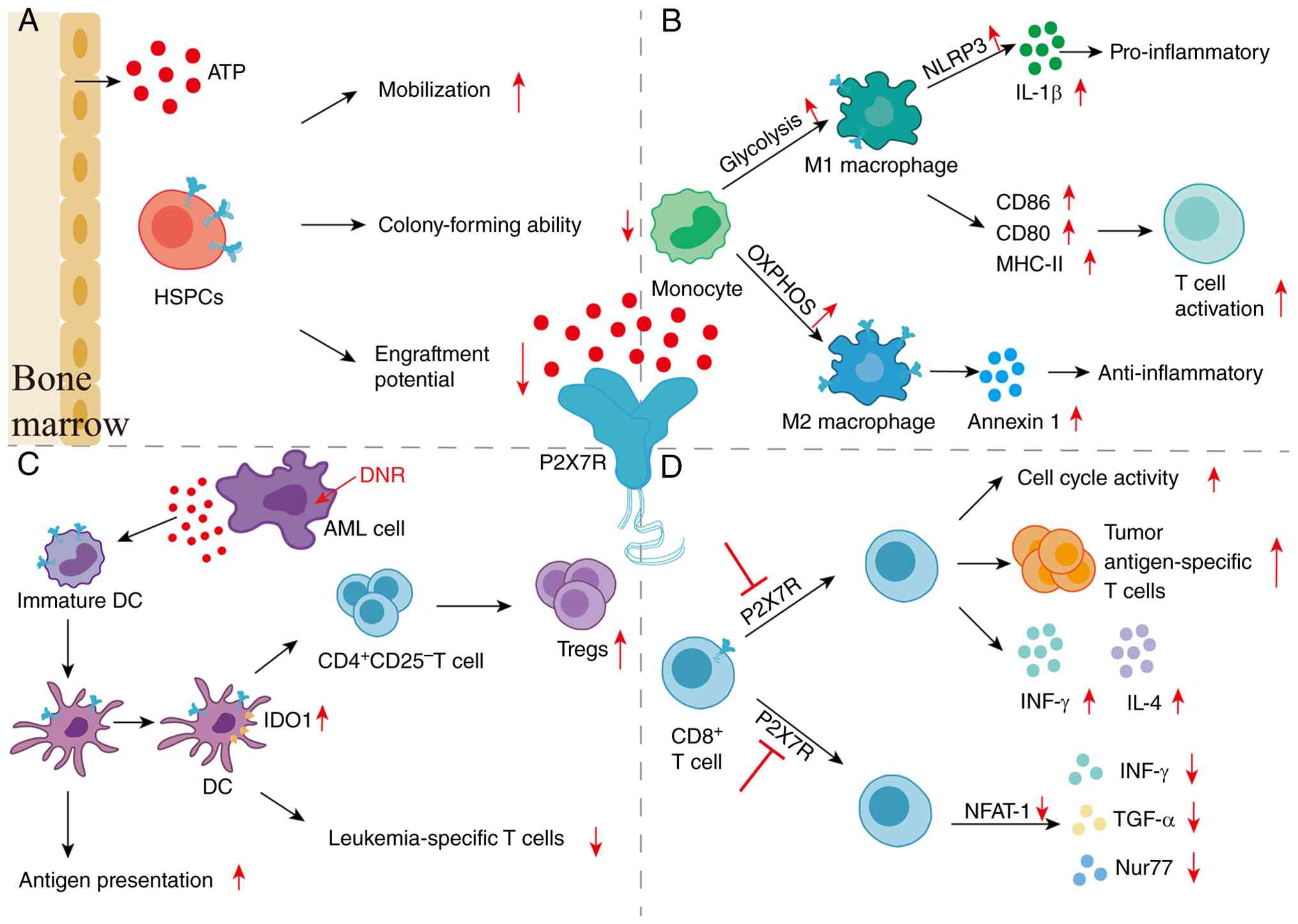 | Figure 2Effects of P2X7R on HSPCs,
macrophages, DCs, and CD8+ T cells in the BM niche of
AML. (A) ATP released into the BM niche of AML activates P2X7R on
HSPCs, promoting mobilization, inhibiting colony-forming ability
and engraftment potential (49).
(B) P2X7R activation drives monocyte differentiation into M1
macrophages by enhancing glycolysis, while promoting
differentiation into immunosuppressive M2 macrophages through
enhancement of OXPHOS (53). (C)
DNR induces apoptosis of AML cells and triggers substantial ATP
release, thereby facilitating DC maturation and enhancing antigen
presentation. Elevated eATP upregulates IDO-1 expression in DCs,
which promotes the differentiation of
CD4+CD25− T cells into Tregs, and restrains
the immune response of leukemia-specific T cells (43). (D) In CD8+ T cells,
P2X7R inhibition enhances T-cell function by promoting cell cycle
activity, tumor antigen-specific T cells, and secretion of IFN-γ
and IL-4 (64-66). On the other hand, P2X7R inhibition
inhibits T-cell activity via the downregulation of NFAT-1, which in
turn reduces the secretion of IFN-γ, TGF-α and Nur77 (66). P2X7R, P2X7 receptor; HSPCs,
hematopoietic stem and progenitor cells; DCs, dendritic cells; BM,
bone marrow; AML, acute myeloid leukemia; ATP, adenosine
triphosphate; OXPHOS, oxidative phosphorylation; DNR, daunorubicin;
eATP, extracellular adenosine triphosphate; IDO-1, indoleamine
2,3-dioxygenase 1; Tregs, regulatory T cells; IFN-γ, interferon-γ;
IL-4, interleukin 4; NFAT-1, nuclear factor of activated T cells 1;
TGF-α, transforming growth factor-α. |
P2X7R and HSPCs in AML
HSPCs in AML display multiple abnormalities,
including suppression of normal hematopoiesis, disrupted phenotypic
differentiation, epigenetic and metabolic reprogramming, and drug
resistance, giving rise to a unique subset of HSPC-like LSCs
(45,46). Additionally, HSPCs in AML show
enhanced mitochondrial function, imbalanced fatty acid oxidation
and glycolysis, and reduced ROS levels. Aberrant activation of
stemness-related pathways in HSPCs mediates chemotherapy resistance
and relapse of AML (47).
P2X7R plays an important role in regulating the
functions of HSPCs (Fig. 2A).
P2X7R facilitates the mobilization of HSPCs. Upon P2X7R activation
by ATP, the NLR family pyrin domain-containing 3 (NLRP3)
inflammasome is assembled and activated, leading to the release of
pro-inflammatory cytokines such as interleukin (IL)-1β and IL-18,
which in turn trigger complement system activation, thereby
promoting the egress and mobilization of BM-derived stem and
progenitor cells into the peripheral blood. In
P2x7r−/− mice, granulocyte colony-stimulating
factor (CSF)-triggered mobilization of HSPCs is notably impaired
(48). Overexpression of P2X7R
also impaired the colony-forming ability of HSPCs in vitro.
In addition, HSPCs with P2X7R overexpression exhibited a
significant reduction in their engraftment potential (49). P2X7R also plays an important role
in HSPC response to acute genotoxic stress. Under such stressful
conditions, P2X7R is a direct transcriptional target of p53 in
HSPCs. The expression of P2X7R in HSPCs is upregulated through a
p53-dependent mechanism after whole-body irradiation. P2x7r
deficiency was shown not only to significantly prolong the survival
of mice with irradiation-induced hematopoietic failure but also to
enhance regenerative capacity (50). These findings suggest that
abnormal enhanced P2X7R signaling on HSPCs in AML may persistently
suppress normal hematopoiesis and exacerbate BM failure. Under
genotoxic stress such as chemotherapy, P2X7R was shown to further
impair HSPC regenerative potential and worsen therapy-related
myelosuppression and hematopoietic reconstitution defects.
P2X7R and macrophages in AML
The expression level of P2X7R in human monocytes and
monocyte-derived macrophages is significantly higher than in other
immune cells. P2X7R is the most abundant purinergic receptor in
macrophages, with an expression rate of ~90%. Furthermore, P2X7R
expression on monocytes is 4-5 times higher than on B cells, T
cells, and NKT cells, and its intracellular expression level of
P2X7R is much higher than that on the cell surface (51).
P2X7R was found to be involved in macrophage
function and polarization (Fig.
2B). Immediate stimulation with ATP or the P2X7R agonist
benzoylbenzoyl-ATP reduced C-C motif chemokine ligand 18 release in
a dose-dependent manner, whereas delayed stimulation had no such
effect (52). P2X7R has also been
shown to modulate macrophage polarization by influencing both
oxidative phosphorylation and glycolysis. Enhanced oxidative
metabolism was demonstrated to support M2 differentiation, while
increased glycolytic activity was reported to promote M1
polarization (53). In M1
macrophages, P2X7R was revealed to promote pro-inflammatory
responses (9,54-56), which were shown to exhibit
antitumor effects by mediating the activation of the NLRP3
inflammasome, releasing IL-1β, and inducing pyroptosis (57). In M2 macrophages, P2X7R was
observed to stimulate the secretion of anti-inflammatory mediators
such as Annexin 1 (9).
Macrophages in AML were shown to be reprogrammed
into an M2-like pro-leukemic phenotype through the
CSF-1/IL-10/STAT3 axis, accompanied by increased expression of
CD163 and CD206, which was observed to be correlated with poor
prognosis and chemotherapy resistance (58). M2-like alternatively activated
macrophages were shown to be markedly enriched in the BM of
patients with AML, and their high infiltration was associated with
enhanced stemness of LSCs and chemoresistance. Experiments have
validated that M2 macrophages directly promote leukemia progression
in vitro and in vivo (59). Furthermore, M2 macrophages have
been shown to sustain LSC survival via IL-10/TGF-β secretion and
modulation of angiogenesis (60).
The role of P2X7R in M2 macrophage function was demonstrated to
further contribute to leukemia development by enhancing the
immunosuppressive effects of M2-like macrophages within the BM
microenvironment in AML.
In the AML BM niche, the high expression of P2X7R on
monocytes and macrophages, together with its role in modulating M2
polarization, may promote the reprogramming of tumor-associated
macrophages toward a pro-leukemic M2-like phenotype. By enhancing
the immunosuppressive functions of M2-like macrophages, P2X7R has
been shown to contribute to the survival of LSCs, chemoresistance,
and leukemia progression, making it a potential immunotherapeutic
target within the AML BM microenvironment.
P2X7R and DCs in AML
Extracellular ATP was shown to promote DC maturation
by activating P2X7R and induce the release of pro-inflammatory
cytokines such as IFN-γ and IL-12 via the NF-κB pathway (61). In addition, P2X7R was demonstrated
to regulate the secretion of IL-12, IL-6 and IL-23 in DCs through
the NLRP3 inflammasome (62) and
to upregulate the expression of CD80 and CD86 (11). In addition, ATP released from
immunogenic cell death can also activate P2X7R (63), further strengthening antigen
presentation and ultimately enhancing T-cell activation.
However, the expression of P2X7R on DCs exhibits the
opposite effect in AML. ATP released from leukemic cells induced by
chemotherapy was shown to trigger potent anti-leukemia immune
responses by activating P2X7R on the surface of DCs, driving DCs to
fully mature and acquire efficient antigen-presenting capacity.
Subsequently, ATP was demonstrated to upregulate indoleamine
2,3-dioxygenase 1 (IDO1) expression in DCs through a
P2X7R-dependent pathway. IDO1 was revealed to mediate the
conversion of naive CD4+CD25− T cells into
functional regulatory T cells (Tregs). In addition,
leukemia-derived DCs were shown to suppress the generation of
leukemia-specific T cells via IDO1 (Fig. 2C). Such DCs can effectively induce
the generation of Tregs, which in turn further inhibit specific
T-cell immune responses against leukemia (43).
P2X7R activation on DCs by eATP released from
leukemic cells exerts dual effects in the AML BM niche. While it
promotes DC maturation and antigen presentation, it also induces a
P2X7R-dependent upregulation of IDO1, leading to the generation of
Tregs that suppress anti-leukemia immune responses. This
P2X7R-IDO1-Treg axis in AML-associated DCs may therefore represent
a critical immunosuppressive mechanism within the BM
microenvironment, contributing to immune evasion and limiting the
efficacy of chemotherapy-induced anti-leukemia immunity.
P2X7R and CD8+ T cells in
AML
P2X7R also exhibits dual functions in
CD8+ T cells (Fig.
2D). Knockout or inhibition of P2X7R in CD8+ T cells
was shown to enhance their antitumor activity. For instance,
P2x7r knockout in effector memory T cells increased cell
cycle activity upon stimulation (64,65). Adoptive transfer of
P2x7r−/− CD8+ T cells promoted T-cell
infiltration into tumors and elevated the proportion of tumor
antigen-specific T cells, thereby significantly suppressing tumor
growth (64). Inhibition of P2X7R
in γδ T cells not only markedly reduced apoptosis and improved
survival of unconventional T cells in vitro, but also
increased the number of cells secreting IFN-γ and IL-4 (66). Additionally, activation of P2X7R
induced T-cell senescence and limited its antitumor function
(64). On the other hand,
P2x7r deficiency impaired the antitumor capacity of T cells.
In P2x7r−/− CD8+ T cells, the
formation rate of initial calcium microdomains and the nuclear
translocation of nuclear factor of activated T cells 1 (NFAT-1)
were significantly reduced, a key transcription factor in T-cell
activation. These changes further led to downregulated expression
of INF-γ, TNF-α and nuclear receptor Nur77, ultimately resulting in
impaired cell proliferation (67).
T cells in AML are characterized by exhaustion,
aberrant differentiation, functional suppression and elevated Treg
proportions. Specifically, CD8+ T cells in AML highly
express exhaustion markers including PD-1, TIM-3, LAG-3 and CD244,
with decreased secretion of cytotoxic molecules and IFN-γ, leading
to impaired proliferation and cytolytic capacity (68). In addition, the CD8+
T-cell subsets from newly diagnosed and relapsed patients with AML
exhibit strong heterogeneity (69). At the initial diagnosis, the
proportion of terminally differentiated effector T cells
(CCR7−CD45RA+,
CD27−CD45RA+) is markedly elevated, while the
percentages of naive T cells (CCR7+CD45RA+)
and naive-like T cells (CD27+CD45RA+) are
decreased. Terminal differentiation as well as excessive clonal
expansion are commonly observed in relapsed cases. Notably,
persistently elevated Treg levels during disease remission also
indicate a significantly higher long-term risk of leukemia relapse
(70). However, the expression
pattern and functional mechanisms of P2X7R in CD8+ T
cell within AML still require further investigations.
P2X7R plays a complex and context-dependent role in
CD8+ T-cell function in the AML BM niche. Although P2X7R
activation can promote T-cell senescence and limit antitumor
immunity, its deficiency may also impair T-cell activation and
proliferation. Given that CD8+ T cells in AML exhibit
exhaustion, terminal differentiation, and functional suppression,
the effect of P2X7R signaling in this setting remains to be
determined. Understanding how P2X7R modulates the balance between
T-cell activation and exhaustion within the AML BM microenvironment
could suggest strategies to restore effective antitumor immunity
while avoiding the pro-leukemic immunosuppressive effects.
Expression and function of P2X7R in LSCs and
AML blasts
Expression of P2X7R in patients with
AML
Compared with normal HSPCs, expression of P2X7R was
shown to be significantly elevated in AML, particularly in LSCs.
Overexpression of P2X7R was also observed in established human AML
cell lines and blasT cells derived from patients with AML (12,71,72). In patients with AML, the
expression levels of P2X7R were revealed to differ among various
subtypes, with higher levels in M4, M5, and M6 groups, but not in
M1 or M2 groups (13,72). Furthermore, P2X7R expression was
demonstrated to be associated with the malignant progression and
development of AML. In children with acute leukemia, P2X1R, P2X4R,
P2X5R, and P2X7R were simultaneously highly expressed, among which
the expression of P2X7R was reported to be even higher in relapsed
patients. However, the expression of P2X7R was shown to be
significantly decreased after chemotherapy upon reaching the
complete remission (CR) stage (73). These findings indicate the
heterogenous expression pattern of P2X7R in patients with AML.
The expression of P2X7R isoforms also varies among
patients with AML. It has been reported that P2X7RA and P2X7RB are
markedly upregulated in de novo patients with AML. However,
patients with relapsed or refractory AML were shown to exhibit
different expression patterns from de novo patients. P2X7RA
mRNA was demonstrated to be significantly decreased and P2X7RB mRNA
was reported to be substantially increased in patients with
relapsed AML, and the reason was revealed to be
chemotherapy-related positive selection of P2X7RB. In patients in
remission, both P2X7RA and P2X7RB expression were significantly
decreased compared with de novo patients with AML. These
findings suggest that high P2X7RB expression may confer
chemotherapy resistance (37). It
was verified by the formation of large pores by P2X7RA under high
ATP conditions, while P2X7RB was unable to form cytotoxic pores
in vivo (39).
P2X7R and LSCs/AML blasts
The expression of P2X7R in LSCs serves as a crucial
regulator in sustaining their functions, including proliferation,
survival and homing. High levels of eATP in AML BM microenvironment
and ATP/P2X7R-mediated signal were shown to be important for the
leukemogenic activities of LSCs (71). ATP/P2X7R signaling was revealed to
be essential for the homing of LSCs to their supportive BM niches
and for maintaining their self-renewal capacity, both of which
contribute to leukemogenesis. Moreover, ATP/P2X7R signaling was
demonstrated to directly stimulate the proliferation and enhance
the survival of LSCs (12).
Although P2X7R is expressed in both LSCs and normal
HSPCs, the biological roles in these two stem cell populations are
significantly different. Multiple subsets of LSCs express P2X7R,
including CD34−CD38−,
CD34+CD38−, CD34+CD38+,
and CD34−CD38+ subsets. Expression of P2X7R
in HSPCs was shown to be significantly lower than that in LSCs.
Studies have shown that high concentrations of eATP can induce LSC
apoptosis, while having little effect on the survival rate of
normal CD34+ HSPCs. P2X7RB isoform expressed on normal
HSPCs was also reported to lack the biological function of inducing
apoptosis (12,49,74). The differences of P2X7R may be
attributed to variations in the expression level or isoforms of
P2X7R between LSCs and HSPCs.
The expression of P2X7R on AML cells is also
involved in cell proliferation. P2X7R can maintain the activity and
quantity of mitochondria in leukemia cells by sensing high levels
of eATP, thereby promoting the proliferation of these cells.
Inhibition of P2X7R in AML cells or blockade of ATP release can
reduce leukemic cell proliferation (75).
Upstream and downstream signaling
pathways of P2X7R in AML
In the AML BM niche, multiple mechanisms have been
shown to drive the release of ATP and thereby activate P2X7R, which
in turn promotes AML progression (Fig. 3A). Osteoblasts in AML were
revealed to exhibit upregulated expression of pannexin 1 and
connexin-43, which facilitates ATP release (12). DNR treatment was demonstrated to
promote the release of ATP from dying leukemic cells, which
subsequently activates P2X7R (43). Extracellular nucleotide levels
were shown to be tightly regulated by extracellular nucleotidases
such as CD39 and CD73 (11,76), which were reported to hydrolyze
eATP into immunosuppressive adenosine (76), thereby limiting the activation of
P2X7R.
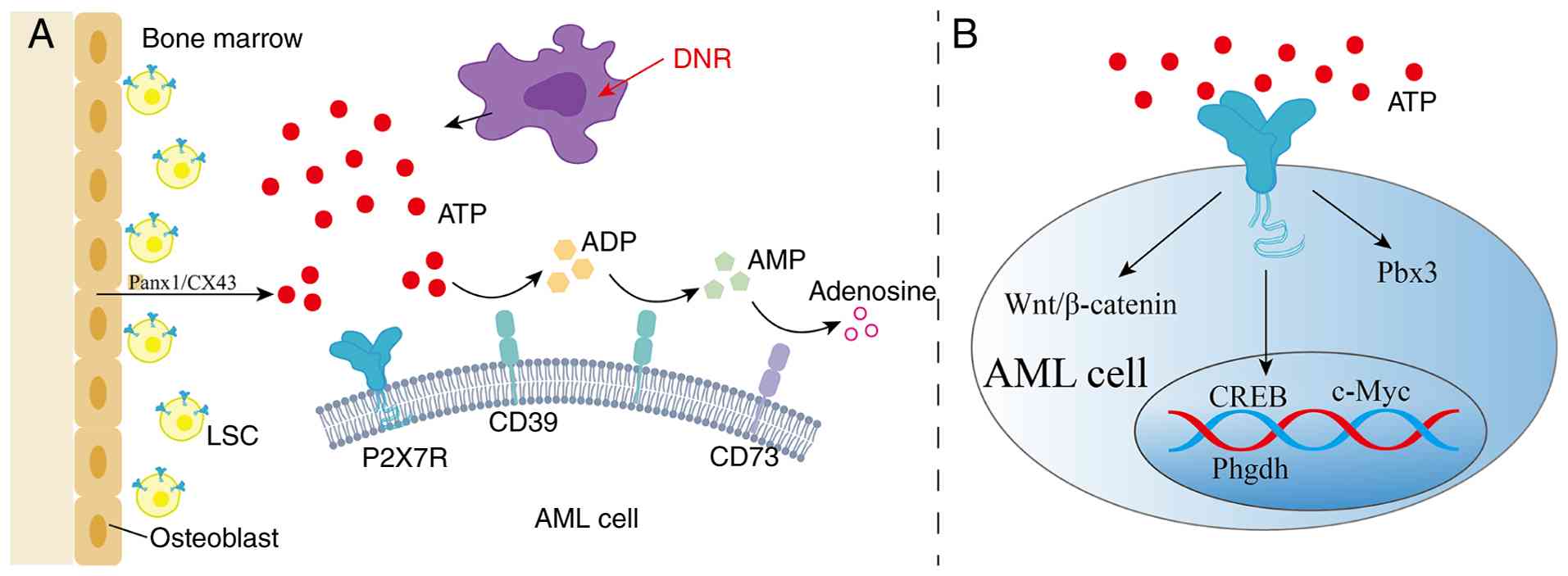 | Figure 3Upstream and downstream signaling
pathways of P2X7R in AML. (A) ATP released into the BM niche from
osteoblasts and dying AML cells can be sequentially hydrolyzed into
ADP, AMP and adenosine by CD39 and CD73 (43). (B) P2X7R promotes AML progression
by activating multiple downstream pathways, including the
CREB/Phgdh/serine metabolic axis (12), Pbx3 (13), Wnt/β-catenin (77) and c-Myc (37). P2X7R, P2X7 receptor; AML, acute
myeloid leukemia; ATP, adenosine triphosphate; BM, bone marrow;
ADP, adenosine diphosphate; AMP, adenosine monophosphate; CREB,
cAMP response element-binding protein; Phgdh, phosphoglycerate
dehydrogenase; Pbx3, PBX homeobox 3; Panx1, pannexin 1; CX43,
connexin-43; LSC, leukemia stem cell; DNR, daunorubicin; CREB, cAMP
response element-binding protein. |
An ATP-rich BM microenvironment in AML provides
sufficient conditions for robust activation of P2X7R, which then
triggers a series of downstream signaling cascades, including the
cAMP response element-binding protein (CREB)/phosphoglycerate
dehydrogenase (Phgdh)/serine metabolic axis (12), PBX homeobox 3 (Pbx3) (13), Wnt/β-catenin (77), and c-Myc (37) (Fig.
3B). Activation of P2X7R was shown to facilitate
Ca2+ influx, which in turn promoted calcium-mediated
phosphorylation of CREB, upregulated Phgdh, and sustained serine
metabolism, ultimately enhancing the homing and self-renewal of
LSCs (12). In MLL-rearranged
AML, overexpression of P2X7R was demonstrated to upregulate Pbx3,
thereby promoting the proliferation of MLL-AF9-driven leukemic
cells and increasing the population of LSCs, ultimately exerting a
leukemogenic effect (13). A
previous study also revealed that the P2X7R-mediated Wnt/β-catenin
signaling pathway can promote the progression of AML (77). Combination treatment with DNR and
a P2X7R inhibitor prevented the downregulation of the
proto-oncogene c-Myc, which was not observed with DNR treatment
alone (37).
P2X7R as a prognosis marker and therapeutic
target in AML
Potential role of P2X7R as a biomarker
for AML prognosis
Elevated P2X7R expression has been found to be
linked to chemotherapy resistance and poor prognosis in AML
(78). The CR rate after one
induction therapy course was significantly lower in patients with
AML and high P2X7R than in those with low or negative P2X7R
(72). In AML mice with
MLL-rearrangements, peripheral blood leukemia cell counts rebound
more rapidly in the P2X7Rhigh group, and the reason was
that P2X7R accelerated the progression of AML by promoting cell
proliferation and increasing the number of LSCs (13). The overexpression of P2X7R also
enhanced the proliferative capacity of leukemic blasts, which play
a critical role in the recurrence and decreased survival rates of
AML (12,13,78). By contrast, when P2X7R was knocked
down, the survival of mice recipients transplanted with human AML
cell lines or primary leukemic cells was significantly prolonged
(12). Further studies have shown
that individuals with high P2X7RB frequently exhibit
chemoresistance and a higher risk of disease recurrence, suggesting
that P2X7RB may serve as a potential novel therapeutic target
(37). The key role of P2X7R in
the development of AML drug resistance and disease recurrence
provides the possibility for P2X7R to serve as a biomarker for AML
prognosis.
P2X7R blockade in AML therapy
Brilliant Blue G (BBG) and oxidized ATP are
first-generation P2X7R inhibitors that also exhibit inhibitory
activity against P2X1R and P2X4R (79). Their limited specificity has led
to the development of a second generation of P2X7R inhibitors,
which have optimized pharmacological properties, such as A438079,
AZ10606120, A740003, AZ11645373, JNJ-47965567 and ZINC 58368839
(80-83).
A740003 has been widely studied in AML and has
demonstrated anti-AML effects (13,78,84). A740003 is a highly selective P2X7R
antagonist that can block BzATP-induced intracellular calcium
concentration changes, P2X7R-mediated pore formation, and has
similar blocking activities in rats and humans (85,86). A740003 can inhibit the
proliferation of human and murine AML cells both in vitro
and in vivo, and impair the colony-forming ability of human
AML cells (12). Intratumoral
administration of A740003 reduced tumor formation in nude mouse
xenograft models (13). Previous
research also showed the anti-leukemia effects of A740003 in a
TIB-49 murine AML model (77).
Mechanistically, A740003 significantly reduced the levels of
phosphorylated CREB, CREB, and Phgdh proteins in wild-type AML
cells but not in P2x7r−/− cells. Treatment with
A740003 also significantly reduced CREB phosphorylation and Phgdh
levels in human AML samples (12).
4-Aminopyrazine (4-AP), one of the most commonly
used K+ channel inhibitors, can suppress the
proliferation of various types of cancer cells and induce cell
apoptosis, including AML cells (87,88). It has been proposed that 4-AP may
promote ATP release from apoptotic AML cells, and the released ATP
acts on P2X7R in an autocrine or paracrine manner, increasing the
intracellular Ca2+ concentration and thereby
participating in the regulation of the cell apoptosis (87).
P2X7R antagonists have also been shown to play a
critical role in graft-vs.-host disease (GVHD). In patients with
AML who had undergone allogeneic hematopoietic stem cell
transplantation, the combination of the P2X7R antagonist BBG with
cyclophosphamide not only significantly reduced the incidence of
hepatic GVHD but also did not impair graft-vs.-leukemia (GVL)
immune function (89).
Combination strategy targeting P2X7R and
chemotherapy
The association between P2X7R expression and
chemoresistance suggests that conventional chemotherapy may fail to
completely eradicate P2X7Rhigh LSCs (13), which provides a rationale for
combination therapy targeting P2X7R together with conventional
chemotherapy.
P2X7R antagonists combined with chemotherapeutic
agents have demonstrated synergistic antileukemic effects. P2X7R
antagonists and 6-mercaptopurine significantly enhanced the
antiproliferative response of leukemic cells in vitro
(75). Similarly, the combination
of P2X7R antagonist AZ10606120 with DNR effectively inhibited
leukemia growth by blocking the oncogenic c-Myc pathway (37). A more nuanced strategy involves
targeting specific P2X7R isoforms. The P2X7RA isoform was shown to
facilitate DNR uptake, increasing cellular sensitivity to
chemotherapy, whereas the P2X7RB isoform was demonstrated to be
highly expressed in relapsed AML cells. This differential
expression suggests a sequential combination strategy: Initial
administration of DNR to eliminate P2X7RA-expressing cells,
followed by the use of a P2X7RB-specific inhibitor to eradicate the
residual, resistant population (37,90).
Beyond antagonists, ATP itself can enhance
chemosensitivity in AML treatment. For instance, ATP administration
has been shown to increase the cytotoxicity of cytarabine against
AML cells (74). Furthermore, the
frequent co-expression of P2X7R and P2X4R offers another promising
strategy for combination therapy. Simultaneous targeting of these
receptors and their associated pathways holds promise for treating
a wider range of diseases driven by their dysfunction in the future
(91).
Research progress and limitations of
targeting P2X7R in AML
Considerable progress has been made in the
development of P2X7R inhibitors. Researchers constructed a 3D
pharmacophore model based on known antagonists (A740003, A804598,
and JNJ47965567) to screen for novel compounds targeting the
negative allosteric pocket of human P2X7R. This approach identified
three promising candidates (compounds 2, 2g, and 9), among which
the compound 2 family demonstrated significantly higher inhibitory
activity than the others (92).
Another design strategy stems from in-depth analysis of the P2X7R
structure. Using high-resolution cryo-electron microscopy to
resolve the full-length human P2X7R structure, researchers
successfully designed UB-MBX-46, a potent and selective antagonist
with a unique polycyclic scaffold. UB-MBX-46 not only exhibited
subnanomolar potency and near-irreversible binding capacity but
also had high selectivity for P2X7R (93). These advances provide critical
insights and a foundation for the development of inhibitors with
improved efficacy, specificity, and therapeutic potential. The
safety and efficacy of a variety of small-molecule inhibitors
targeting P2X7R have been preliminarily verified, such as A740003,
AZD9056 and A438079 (12,94-98).
Research has also expanded to anti-P2X7R antibodies
and nanobodies with superior specificity and efficacy (83,99). Specific antibodies targeting P2X7R
have been designed using gene fusion technology. Adeno-associated
viral vectors can also be used to express P2X7R-specific antibodies
with long-lasting biological effects (100). However, the specific
applications and detailed mechanisms of these antibodies in AML
treatment require further investigation.
The antibodies targeting nfP2X7R offer new
potential for cancer therapy. Antibodies against the E200 sequence
of P2X7R, such as BIL03s and BPM09, can specifically bind to
nfP2X7R without interacting with functional P2X7R (38). BIL010t, a first-in-class antibody
targeting nfP2X7R, has been tested in a phase I clinical trial for
basal cell carcinoma and has demonstrated a favorable safety
profile (14).
However, targeting P2X7R in AML therapy still has
multiple challenges. Although activation of P2X7R is generally
associated with poor prognosis in tumors, this receptor is neither
a simple oncogenic nor a tumor-suppressive molecule. A more
detailed understanding of its roles in distinct disease contexts
still needs to be explored. The expression profile of P2X7R in
distincT cell types within the BM microenvironment under
physiological conditions and in the pathological context of AML
still need to be systematically characterized (26). Another important aspect is to
characterize the functional diversity of P2X7R isoforms. P2X7R
antagonists/antibodies have also not reached approval for clinical
use due to poor pharmacokinetic properties, insufficient
selectivity, species differences, tissue distribution, and limited
clinical efficacy (92). Further
clinical trials are still required to validate the long-term safety
and efficacy of P2X7R-targeted approaches.
Conclusions
As an ATP-gated ion channel, P2X7R plays a critical
and dualistic role in AML pathogenesis and the immunosuppressive BM
niche. The formation of ion channels and large pores confers its
functional properties of promoting cell survival or inducing cell
death, which are significantly modulated by ATP concentration and
duration of action, P2X7R subtypes, and the TME. P2X7R is highly
expressed on LSCs, AML blasts, and multiple immune cells in AML.
Sustained activation by eATP in the BM niche promotes AML blast
survival, LSC maintenance, and suppresses anti-leukemic immunity
via downstream signaling pathways. The pathophysiological role of
P2X7R establishes it as a prognosis marker and potential
therapeutic target. Current strategies include small molecule
antagonists, specific antibodies, and combination with conventional
chemotherapy. Despite the rationale for targeting P2X7R in AML,
bridging the promising preclinical data to established clinical
therapy requires further extensive investigation.
Availability of data and materials
Not applicable.
Authors' contributions
LF conceived the study. YL wrote the original
draft. HX, HM, TS and ZL wrote, reviewed and edited the manuscript.
LF and NW designed the scope and structure of the review. LF
supervised the study and was responsible for funding acquisition,
LF. All authors read and approved the final manuscript. Data
authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
Not applicable.
Funding
The research was funded by the Natural Science Foundation of
Shandong Province (grant no. ZR2023MH233 and no. ZR2020MH122).
References
|
1
|
Bonnet D and Dick JE: Human acute myeloid
leukemia is organized as a hierarchy that originates from a
primitive hematopoietic cell. Nat Med. 3:730–737. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Döhner H, Weisdorf DJ and Bloomfield CD:
Acute myeloid leukemia. N Engl J Med. 373:1136–1152. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Marchand T and Pinho S: Leukemic stem
cells: From leukemic niche biology to treatment opportunities.
Front Immunol. 12:7751282021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Méndez-Ferrer S, Bonnet D, Steensma DP,
Hasserjian RP, Ghobrial IM, Gribben JG, Andreeff M and Krause DS:
Bone marrow niches in haematological malignancies. Nat Rev Cancer.
20:285–298. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Di Virgilio F, Sarti AC, Falzoni S, De
Marchi E and Adinolfi E: Extracellular ATP and P2 purinergic
signalling in the tumour microenvironment. Nat Rev Cancer.
18:601–618. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lara R, Adinolfi E, Harwood CA, Philpott
M, Barden JA, Di Virgilio F and McNulty S: P2X7 in cancer: From
molecular mechanisms to therapeutics. Front Pharmacol. 11:7932020.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
He X, Zhang Y, Xu Y, Xie L, Yu Z and Zheng
J: Function of the P2X7 receptor in hematopoiesis and
leukemogenesis. Exp Hematol. 104:40–47. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Acuña-Castillo C, Escobar A, García-Gómez
M, Bachelet VC, Huidobro-Toro JP, Sauma D and Barrera-Avalos C:
P2X7 receptor in dendritic cells and macrophages: Implications in
antigen presentation and T lymphocyte activation. Int J Mol Sci.
25:24952024. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
de Torre-Minguela C, Barberà-Cremades M,
Gómez AI, Martín-Sánchez F and Pelegrín P: Macrophage activation
and polarization modify P2X7 receptor secretome influencing the
inflammatory process. Sci Rep. 6:225862016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang Y, Story ME, Hao X, Sumpter TL and
Mathers AR: P2X7 receptor expression and signaling on dendritic
cells and CD4+ T cells is not required but can enhance Th17
differentiation. FronT cell Dev Biol. 10:6876592022. View Article : Google Scholar
|
|
11
|
Rivas-Yáñez E, Barrera-Avalos C,
Parra-Tello B, Briceño P, Rosemblatt MV, Saavedra-Almarza J,
Rosemblatt M, Acuña-Castillo C, Bono MR and Sauma D: P2X7 receptor
at the crossroads of T cell fate. Int J Mol Sci. 21:49372020.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
He X, Wan J, Yang X, Zhang X, Huang D, Li
X, Zou Y, Chen C, Yu Z, Xie L, et al: Bone marrow niche ATP levels
determine leukemia-initiating cell activity via P2X7 in leukemic
models. J Clin Invest. 131:e1402422021. View Article : Google Scholar
|
|
13
|
Feng W, Yang X, Wang L, Wang R, Yang F,
Wang H, Liu X, Ren Q, Zhang Y, Zhu X and Zheng G: P2X7 promotes the
progression of MLL-AF9 induced acute myeloid leukemia by
upregulation of Pbx3. Haematologica. 106:1278–1289. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gilbert SM, Gidley Baird A, Glazer S,
Barden JA, Glazer A, Teh LC and King J: A phase I clinical trial
demonstrates that nfP2X7-targeted antibodies provide a novel, safe
and tolerable topical therapy for basal cell carcinoma. Br J
Dermatol. 177:117–124. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Roger S and Pelegrin P: P2X7 receptor
antagonism in the treatment of cancers. Expert Opin Investig Drugs.
20:875–880. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Buell GN, Talabot F, Gos A, Lorenz J, Lai
E, Morris MA and Antonarakis SE: Gene structure and chromosomal
localization of the human P2X7 receptor. Recept Channels.
5:347–354. 1998.PubMed/NCBI
|
|
17
|
Jiang LH, Caseley EA, Muench SP and Roger
S: Structural basis for the functional properties of the P2X7
receptor for extracellular ATP. Purinergic Signal. 17:331–344.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Saul A, Hausmann R, Kless A and Nicke A:
Heteromeric assembly of P2X subunits. FronT cell Neurosci.
7:2502013. View Article : Google Scholar
|
|
19
|
Boumechache M, Masin M, Edwardson JM,
Górecki DC and Murrell-Lagnado R: Analysis of assembly and
trafficking of native P2X4 and P2X7 receptor complexes in rodent
immune cells. J Biol Chem. 284:13446–13454. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sheng D and Hattori M: Recent progress in
the structural biology of P2X receptors. Proteins. 90:1779–1785.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hattori M and Gouaux E: Molecular
mechanism of ATP binding and ion channel activation in P2X
receptors. Nature. 485:207–212. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jiang LH, Baldwin JM, Roger S and Baldwin
SA: Insights into the molecular mechanisms underlying mammalian
P2X7 receptor functions and contributions in diseases, revealed by
structural modeling and single nucleotide polymorphisms. Front
Pharmacol. 4:552013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Di Virgilio F, Schmalzing G and Markwardt
F: The elusive P2X7 macropore. Trends Cell Biol. 28:392–404. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cevoli F, Arnould B, Peralta FA and
Grutter T: Untangling macropore formation and current facilitation
in P2X7. Int J Mol Sci. 24:108962023. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang WJ, Hu CG, Zhu ZM and Luo HL: Effect
of P2X7 receptor on tumorigenesis and its pharmacological
properties. Biomed Pharmacother. 125:1098442020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang GP, Liao JX, Liu YY, Zhu FQ, Huang
HJ and Zhang WJ: Ion channel P2X7 receptor in the progression of
cancer. Front Oncol. 13:12977752023. View Article : Google Scholar
|
|
27
|
Di Virgilio F, Vultaggio-Poma V and Sarti
AC: P2X receptors in cancer growth and progression. Biochem
Pharmacol. 187:1143502021. View Article : Google Scholar
|
|
28
|
Kopp R, Krautloher A, Ramírez-Fernández A
and Nicke A: P2X7 interactions and signaling-making head or tail of
it. Front Mol Neurosci. 12:1832019. View Article : Google Scholar
|
|
29
|
Harkat M, Peverini L, Cerdan AH, Dunning
K, Beudez J, Martz A, Calimet N, Specht A, Cecchini M, Chataigneau
T and Grutter T: On the permeation of large organic cations through
the pore of ATP-gated P2X receptors. Proc Natl Acad Sci USA.
114:E3786–E3795. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Di Virgilio F, Giuliani AL, Vultaggio-Poma
V, Falzoni S and Sarti AC: Non-nucleotide agonists triggering P2X7
receptor activation and pore formation. Front Pharmacol. 9:392018.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Martínez-Cuesta MÁ, Blanch-Ruiz MA,
Ortega-Luna R, Sánchez-López A and Álvarez Á: Structural and
functional basis for understanding the biological significance of
P2X7 receptor. Int J Mol Sci. 21:84542020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ziberi S, Zuccarini M, Carluccio M,
Giuliani P, Ricci-Vitiani L, Pallini R, Caciagli F, Di Iorio P and
Ciccarelli R: Upregulation of Epithelial-to-Mesenchymal transition
markers and P2X7 receptors is associated to increased invasiveness
caused by P2X7 receptor stimulation in human glioblastoma stem
cells. Cells. 9:852019. View Article : Google Scholar
|
|
33
|
Ulrich H, Ratajczak MZ, Schneider G,
Adinolfi E, Orioli E, Ferrazoli EG, Glaser T, Corrêa-Velloso J,
Martins PCM, Coutinho F, et al: Kinin and purine signaling
contributes to neuroblastoma metastasis. Front Pharmacol.
9:5002018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Giuliani AL, Colognesi D, Ricco T, Roncato
C, Capece M, Amoroso F, Wang QG, De Marchi E, Gartland A, Di
Virgilio F and Adinolfi E: Trophic activity of human P2X7 receptor
isoforms a and B in osteosarcoma. PLoS One. 9:e1072242014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Benzaquen J, Heeke S, Janho dit Hreich S,
Douguet L, Marquette CH, Hofman P and Vouret-Craviari V:
Alternative splicing of P2RX7 pre-messenger RNA in health and
diseases: Myth or reality? Biomed J. 42:141–154. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
De Salis SKF, Li L, Chen Z, Lam KW,
Skarratt KK, Balle T and Fuller SJ: Alternatively spliced isoforms
of the P2X7 receptor: Structure, function and disease associations.
Int J Mol Sci. 23:81742022. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Pegoraro A, Orioli E, De Marchi E,
Salvestrini V, Milani A, Di Virgilio F, Curti A and Adinolfi E:
Differential sensitivity of acute myeloid leukemia cells to
daunorubicin depends on P2X7A versus P2X7B receptor expression.
Cell Death Dis. 11:8762020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gilbert S, Oliphant CJ, Hassan S, Peille
AL, Bronsert P, Falzoni S, Di Virgilio F, McNulty S and Lara R: ATP
in the tumour microenvironment drives expression of
nfP2X7, a key mediator of cancer cell survival.
Oncogene. 38:194–208. 2019. View Article : Google Scholar :
|
|
39
|
Pegoraro A, De Marchi E and Adinolfi E:
P2X7 variants in oncogenesis. Cells. 10:1892021. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Magni L, Yu H, Christensen NM, Poulsen MH,
Frueh A, Deshar G, Johansen AZ, Johansen JS, Pless SA, Jørgensen NR
and Novak I: Human P2X7 receptor variants Gly150Arg and Arg276His
polymorphisms have differential effects on risk association and
cellular functions in pancreatic cancer. Cancer Cell Int.
24:1482024. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shemon AN, Sluyter R, Fernando SL, Clarke
AL, Dao-Ung LP, Skarratt KK, Saunders BM, Tan KS, Gu BJ, Fuller SJ,
et al: A Thr357 to Ser polymorphism in homozygous and compound
heterozygous subjects causes absent or reduced P2X7 function and
impairs ATP-induced mycobacterial killing by macrophages. J Biol
Chem. 281:2079–2086. 2006. View Article : Google Scholar
|
|
42
|
Schäfer W, Stähler T, Pinto Espinoza C,
Danquah W, Knop JH, Rissiek B, Haag F and Koch-Nolte F: Origin,
distribution, and function of three frequent coding polymorphisms
in the gene for the human P2X7 ion channel. Front Pharmacol.
13:10331352022. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lecciso M, Ocadlikova D, Sangaletti S,
Trabanelli S, De Marchi E, Orioli E, Pegoraro A, Portararo P,
Jandus C, Bontadini A, et al: ATP release from chemotherapy-treated
dying leukemia cells elicits an immune suppressive effect by
increasing regulatory T cells and tolerogenic dendritic cells.
Front Immunol. 8:19182017. View Article : Google Scholar
|
|
44
|
Thomas LM and Salter RD: Activation of
macrophages by P2X7-induced microvesicles from myeloid cells is
mediated by phospholipids and is partially dependent on TLR4. J
Immunol. 185:3740–3749. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chen DW, Fan JM, Schrey JM, Mitchell DV,
Jung SK, Hurwitz SN, Perez EB, Muraro MJ, Carroll M, Taylor DM and
Kurre P: Inflammatory recruitment of healthy hematopoietic stem and
progenitor cells in the acute myeloid leukemia niche. Leukemia.
38:741–750. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Matthes T: Phenotypic analysis of
hematopoietic stem and progenitor cell populations in acute myeloid
leukemia based on spectral flow cytometry, a 20-color panel, and
unsupervised learning algorithms. Int J Mol Sci. 25:28472024.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bao J, Freund O, Sund L and Du W: Inside
the battle against acute myeloid leukemia: Biology, breakthroughs,
and hope. Cells. 15:3382026. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lenkiewicz AM, Adamiak M, Thapa A, Bujko
K, Pedziwiatr D, Abdel-Latif AK, Kucia M, Ratajczak J and Ratajczak
MZ: The Nlrp3 inflammasome orchestrates mobilization of bone
marrow-residing stem cells into peripheral blood. Stem Cell Rev
Rep. 15:391–403. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Feng W, Yang F, Wang R, Yang X, Wang L,
Chen C, Liao J, Lin Y, Ren Q and Zheng G: High level P2X7-mediated
signaling impairs function of hematopoietic stem/progenitor cells.
Stem Cell Rev Rep. 12:305–314. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Tung LT, Wang H, Belle JI, Petrov JC,
Langlais D and Nijnik A: p53-dependent induction of P2X7 on
hematopoietic stem and progenitor cells regulates hematopoietic
response to genotoxic stress. Cell Death Dis. 12:9232021.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Gu BJ, Zhang WY, Bendall LJ, Chessell IP,
Buell GN and Wiley JS: Expression of P2X(7) purinoceptors on human
lymphocytes and monocytes: Evidence for nonfunctional P2X(7)
receptors. Am J Physiol Cell Physiol. 279:C1189–C1197. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Scherr BF, Reiner MF, Baumann F, Höhne K,
Müller T, Ayata K, Müller-Quernheim J, Idzko M and Zissel G:
Prevention of M2 polarization and temporal limitation of
differentiation in monocytes by extracellular ATP. BMC Immunol.
24:112023. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Virgilio FD, Ben DD, Sarti AC, Giuliani AL
and Falzoni S: The P2X7 receptor in infection and inflammation.
Immunity. 47:15–31. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Pelegrin P and Surprenant A: Dynamics of
macrophage polarization reveal new mechanism to inhibit IL-1beta
release through pyrophosphates. EMBO J. 28:2114–2127. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Lopez-Castejón G, Baroja-Mazo A and
Pelegrín P: Novel macrophage polarization model: From gene
expression to identification of new anti-inflammatory molecules.
Cell Mol Life Sci CMLS. 68:3095–3107. 2011. View Article : Google Scholar
|
|
56
|
Chen SY, Feng WL, Yang X, Liao JF, Wang
LN, Lin YM, Ren Q and Zheng GG: Expression of P2X family receptors
in peritoneal macrophages of mouse with acute T lymphoblastic
leukemia. Zhongguo Shi Yan Xue Ye Xue Za Zhi. 22:623–628. 2014.In
Chinese. PubMed/NCBI
|
|
57
|
Pelegrin P: P2X7 receptor and the NLRP3
inflammasome: Partners in crime. Biochem Pharmacol. 187:1143852021.
View Article : Google Scholar
|
|
58
|
Miari KE, Guzman ML, Wheadon H and
Williams MTS: Macrophages in acute myeloid leukaemia: Significant
players in therapy resistance and patient outcomes. FronT cell Dev
Biol. 9:6928002021. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Weinhäuser I, Pereira-Martins DA, Almeida
LY, Hilberink JR, Silveira DRA, Quek L, Ortiz C, Araujo CL, Bianco
TM, Lucena-Araujo A, et al: M2 macrophages drive leukemic
transformation by imposing resistance to phagocytosis and improving
mitochondrial metabolism. Sci Adv. 9:eadf85222023. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Chen ZG, Yang H, Yang C, Xie YT, Li CM,
Xiao T, Wu JH, Gao MY, Wang CC, Zhao YL, et al: Macrophage
polarization in hematologic cancers: Mechanisms and therapeutic
strategies. Blood Res. 61:82026. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Yu Y, Feng S, Wei S, Zhong Y, Yi G, Chen
H, Liang L, Chen H and Lu X: Extracellular ATP activates
P2X7R-NF-κB (p65) pathway to promote the maturation of bone
marrow-derived dendritic cells of mice. Cytokine. 119:175–181.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Li R, Wang J, Li R, Zhu F, Xu W, Zha G, He
G, Cao H, Wang Y and Yang J: ATP/P2X7-NLRP3 axis of dendritic cells
participates in the regulation of airway inflammation and
hyper-responsiveness in asthma by mediating HMGB1 expression and
secretion. Exp Cell Res. 366:1–15. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Barrera-Avalos C, Briceño P, Valdés D,
Imarai M, Leiva-Salcedo E, Rojo LE, Milla LA, Huidobro-Toro JP,
Robles-Planells C, Escobar A, et al: P2X7 receptor is essential for
cross-dressing of bone marrow-derived dendritic cells. iScience.
24:1035202021. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Romagnani A, Rottoli E, Mazza EMC,
Rezzonico-Jost T, De Ponte Conti B, Proietti M, Perotti M,
Civanelli E, Perruzza L, Catapano AL, et al: P2X7 receptor activity
limits accumulation of T cells within tumors. Cancer Res.
80:3906–3919. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Grassi F: The P2X7 receptor as regulator
of T cell development and function. Front Immunol. 11:11792020.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Xu C, Obers A, Qin M, Brandli A, Wong J,
Huang X, Clatch A, Fayed A, Starkey G, D'Costa R, et al: Selective
regulation of IFN-γ and IL-4 co-producing unconventional T cells by
purinergic signaling. J Exp Med. 221:e202403542024. View Article : Google Scholar
|
|
67
|
Brock VJ, Lory NC, Möckl F, Birus M,
Stähler T, Woelk LM, Jaeckstein M, Heeren J, Koch-Nolte F, Rissiek
B, et al: Time-resolved role of P2X4 and P2X7 during CD8+ T cell
activation. Front Immunol. 15:12581192024. View Article : Google Scholar
|
|
68
|
Knaus HA, Berglund S, Hackl H, Blackford
AL, Zeidner JF, Montiel-Esparza R, Mukhopadhyay R, Vanura K, Blazar
BR, Karp JE, et al: Signatures of CD8+ T cell dysfunction in AML
patients and their reversibility with response to chemotherapy. JCI
Insight. 3:e1209742018. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Desai PN, Wang B, Fonseca A, Borges P,
Jelloul FZ, Reville PK, Lee E, Ly C, Basi A, Root J, et al:
Single-cell profiling of CD8+ T cells in acute myeloid leukemia
reveals a continuous spectrum of differentiation and clonal
hyperexpansion. Cancer Immunol Res. CIR-22-09612023.PubMed/NCBI
|
|
70
|
Li Z, Philip M and Ferrell PB: Alterations
of T-cell-mediated immunity in acute myeloid leukemia. Oncogene.
39:3611–3619. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Pegoraro A and Adinolfi E: The ATP/P2X7
axis is a crucial regulator of leukemic initiating cells
proliferation and homing and an emerging therapeutic target in
acute myeloid leukemia. Purinergic Signal. 17:319–321. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Zhang XJ, Zheng GG, Ma XT, Yang YH, Li G,
Rao Q, Nie K and Wu KF: Expression of P2X7 in human hematopoietic
cell lines and leukemia patients. Leuk Res. 28:1313–1322. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Chong JH, Zheng GG, Zhu XF, Guo Y, Wang L,
Ma CH, Liu SY, Xu LL, Lin YM and Wu KF: Abnormal expression of P2X
family receptors in Chinese pediatric acute leukemias. Biochem
Biophys Res Commun. 391:498–504. 2010. View Article : Google Scholar
|
|
74
|
Salvestrini V, Orecchioni S, Talarico G,
Reggiani F, Mazzetti C, Bertolini F, Orioli E, Adinolfi E, Di
Virgilio F, Pezzi A, et al: Extracellular ATP induces apoptosis
through P2X7R activation in acute myeloid leukemia cells but not in
normal hematopoietic stem cells. Oncotarget. 8:5895–5908. 2017.
View Article : Google Scholar :
|
|
75
|
Ledderose C, Woehrle T, Ledderose S,
Strasser K, Seist R, Bao Y, Zhang J and Junger WG: Cutting off the
power: Inhibition of leukemia cell growth by pausing basal ATP
release and P2X receptor signaling? Purinergic Signal. 12:439–451.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Mucha PT, Brahmachari A, Frańczak MA,
Tomczyk M, Kutryb-Zając B, Koszałka P, Giovannetti E and Peters GJ:
The role of the ecto-nucleotidases CD73 and CD39 in chemo- and
immunotherapy. Cancers (Basel). 18:9572026. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Feng L, Zhang H, Mao C, De Andrade Mello
P, Stroopinsky D, Csizmadia E, Zhou J, Avigan D, Yu J, Gao W and
Robson SC: Cd39 and P2rx7-wnt signaling enhance blast pathogenicity
in an experimental model of acute myeloid leukemia. Haematologica.
110:212–217. 2024.PubMed/NCBI
|
|
78
|
De Marchi E, Orioli E, Pegoraro A,
Sangaletti S, Portararo P, Curti A, Colombo MP, Di Virgilio F and
Adinolfi E: The P2X7 receptor modulates immune cells infiltration,
ectonucleotidases expression and extracellular ATP levels in the
tumor microenvironment. Oncogene. 38:3636–3650. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Greve AS, Skals M, Fagerberg SK, Tonnus W,
Ellermann-Eriksen S, Evans RJ, Linkermann A and Praetorius HA:
P2X1, P2X4, and P2X7 receptor knock out mice expose differential
outcome of sepsis induced by α-haemolysin producing escherichia
coli. FronT cell Infect Microbiol. 7:1132017. View Article : Google Scholar
|
|
80
|
Bin Dayel A, Evans RJ and Schmid R:
Mapping the site of action of human P2X7 receptor antagonists
AZ11645373, brilliant blue G, KN-62, calmidazolium, and
ZINC58368839 to the intersubunit allosteric pocket. Mol Pharmacol.
96:355–363. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Filippin KJ, De Souza KFS, De Araujo
Júnior RT, Torquato HFV, Dias DA, Parisotto EB, Ferreira AT and
Paredes-Gamero EJ: Involvement of P2 receptors in hematopoiesis and
hematopoietic disorders, and as pharmacological targets. Purinergic
Signal. 16:1–15. 2020. View Article : Google Scholar :
|
|
82
|
Drill M, Jones NC, Hunn M, O'Brien TJ and
Monif M: Antagonism of the ATP-gated P2X7 receptor: A potential
therapeutic strategy for cancer. Purinergic Signal. 17:215–227.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Monaco S, Browne J, Wallace M, Angulo J
and Stokes L: On-cell saturation transfer difference NMR
spectroscopy on ion channels: Characterizing negative allosteric
modulator binding interactions of P2X7. J Am Chem Soc.
147:32400–32411. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
De Marchi E, Pegoraro A and Adinolfi E:
P2X7 receptor in hematological malignancies. FronT cell Dev Biol.
9:6456052021. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Honore P, Donnelly-Roberts D, Namovic MT,
Hsieh G, Zhu CZ, Mikusa JP, Hernandez G, Zhong C, Gauvin DM,
Chandran P, et al: A-740003 [N-(1-{[(cyanoimino)(5-quinolinylamino)
methyl]amino}-2,2-dimethylpropyl)-2-(3,4
-dimethoxyphenyl)acetamide], a novel and selective P2X7 receptor
antagonist, dose-dependently reduces neuropathic pain in the rat. J
Pharmacol Exp Ther. 319:1376–1385. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Anderson CM and Nedergaard M: Emerging
challenges of assigning P2X7 receptor function and immunoreactivity
in neurons. Trends Neurosci. 29:257–262. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Wang W, Xiao J, Adachi M, Liu Z and Zhou
J: 4-aminopyridine induces apoptosis of human acute myeloid
leukemia cells via increasing [Ca2+]i through P2X7
receptor pathway. Cell Physiol Biochem. 28:199–208. 2011.
View Article : Google Scholar
|
|
88
|
Rybalchenko V, Prevarskaya N, Van
Coppenolle F, Legrand G, Lemonnier L, Le Bourhis X and Skryma R:
Verapamil inhibits proliferation of LNCaP human prostate cancer
cells influencing K+ channel gating. Mol Pharmacol. 59:1376–1387.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Cuthbertson P, Button A, Sligar C, Elhage
A, Vine KL, Watson D and Sluyter R: Post-transplant
cyclophosphamide combined with brilliant blue G reduces
graft-versus-host disease without compromising
graft-versus-leukaemia immunity in humanised mice. Int J Mol Sci.
25:17752024. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Adinolfi E, Cirillo M, Woltersdorf R,
Falzoni S, Chiozzi P, Pellegatti P, Callegari MG, Sandonà D,
Markwardt F, Schmalzing G and Di Virgilio F: Trophic activity of a
naturally occurring truncated isoform of the P2X7 receptor. FASEB
J. 24:3393–3404. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Pérez-Flores G, Lévesque SA, Pacheco J,
Vaca L, Lacroix S, Pérez-Cornejo P and Arreola J: The P2X7/P2X4
interaction shapes the purinergic response in murine macrophages.
Biochem Biophys Res Commun. 467:484–490. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Zuanon M, Brancale A and Young MT:
Identification of new human P2X7 antagonists using ligand- and
structure-based virtual screening. J Chem Inf Model. 65:7143–7155.
2025. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Oken AC, Turcu AL, Tzortzini E, Georgiou
K, Nagel J, Westermann FG, Barniol-Xicota M, Seidler J, Kim GR, Lee
SD, et al: A polycyclic scaffold identified by structure-based drug
design effectively inhibits the human P2X7 receptor. Nat Commun.
16:82832025. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Zhang WJ, Luo HL, Liu JP, Xu YS, Wang WL
and Huang C: P2X7 receptor promotes the growth and metastasis of
gastric cancer by activating P13/AKT/GSK-3 beta signaling
(experimental research). Int J Surg Lond Engl. 111:3752–3766. 2025.
View Article : Google Scholar
|
|
95
|
Zhang WJ, Luo C, Huang C, Pu FQ, Zhu JF
and Zhu ZM: PI3K/akt/GSK-3β signal pathway is involved in P2X7
receptor-induced proliferation and EMT of colorectal cancer cells.
Eur J Pharmacol. 899:1740412021. View Article : Google Scholar
|
|
96
|
Zhang WJ, Hu CG, Luo HL and Zhu ZM:
Activation of P2×7 receptor promotes the invasion and migration of
colon cancer cells via the STAT3 signaling. FronT cell Dev Biol.
8:5865552020. View Article : Google Scholar
|
|
97
|
Keystone EC, Wang MM, Layton M, Hollis S
and McInnes IB; D1520C00001 Study Team: Clinical evaluation of the
efficacy of the P2X7 purinergic receptor antagonist AZD9056 on the
signs and symptoms of rheumatoid arthritis in patients with active
disease despite treatment with methotrexate or sulphasalazine. Ann
Rheum Dis. 71:1630–1635. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Eser A, Colombel J-F, Rutgeerts P,
Vermeire S, Vogelsang H, Braddock M, Persson T and Reinisch W:
Safety and efficacy of an oral inhibitor of the purinergic receptor
P2X7 in adult patients with moderately to severely active Crohn's
disease: A randomized placebo-controlled, double-blind, phase IIa
study. Inflamm Bowel Dis. 21:2247–2253. 2015.PubMed/NCBI
|
|
99
|
Danquah W, Meyer-Schwesinger C, Rissiek B,
Pinto C, Serracant-Prat A, Amadi M, Iacenda D, Knop JH, Hammel A,
Bergmann P, et al: Nanobodies that block gating of the P2X7 ion
channel ameliorate inflammation. Sci Transl Med. 8:366ra1622016.
View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Koch-Nolte F, Eichhoff A, Pinto-Espinoza
C, Schwarz N, Schäfer T, Menzel S, Haag F, Demeules M, Gondé H and
Adriouch S: Novel biologics targeting the P2X7 ion channel. Curr
Opin Pharmacol. 47:110–118. 2019. View Article : Google Scholar : PubMed/NCBI
|














