Introduction
Gastric cancer (GC) is one of the most common
malignant tumors in the world with a high incidence (5.6% of all
cancers) (1,2). Genetic susceptibility is one of the
important risk factors for GC (3-5).
Germline mutations in the E-cadherin gene (CDH1) or microsatellite
instability in the DNA mismatch repair gene hMLH1 are associated
with familial GC (6,7). In addition to genetic factors, the
risk factors for GC are also associated with Helicobacter
pylori infection and unhealthy lifestyles, including alcohol
consumption, high-salt diet and smoking (2,8).
Due to the lack of clear clinical indications, most patients were
in the advanced stage of GC at diagnosis, accompanied by poor
prognosis (1,9). The innovation of molecular analysis
technology has improved the efficiency and accuracy of prediction
of diagnostic and targeted therapy markers (10). The diversification of treatment
methods such as chemotherapy, targeted therapy, and immunotherapy
and the innovation of molecular diagnostic technology have
prolonged the survival of patients, but the 5-year survival of
patients with GC is still less than 30% (9,11).
Therefore, it is essential to find effective early markers for the
diagnosis and treatment of GC.
T-box transcription factor 15 (TBX15) belongs to the
T-box gene family, encoding a phylogenetically conserved
transcription factor family. Members of this family have a highly
conserved characteristic sequence of 180 amino acid residues that
can bind to DNA, and their target genes contain one or more T
half-sites in the upstream promoter sequence (12,13). TBX15 has been found to play a role
in the regulation of various diseases, such as heart failure
(14), osteoporosis (15), obesity (16) and cancer development (17,18). In multiple tumor studies, TBX15
was associated with poor prognosis in patients with glioma
(18,19) and colorectal cancer (20). In addition, TBX15 enriched in
glioma was associated with immunosuppressive genes (19). It can be hypothesized that TBX15
may participate in the immune escape process of cancer cells. In a
GC study, TBX15 was implicated in GC cell proliferation and drug
antitumor pathways (21).
However, the regulatory pathways of TBX15 related to the prognosis
of patients with GC and their immune escape remain an unsolved
problem.
N6-methyladenosine (m6A) is a kind of RNA
modification, which widely exists in eukaryotic mRNA modification.
The m6A WERs ('writers', 'erasers' and 'readers') control the
dynamic modification process of m6A, which is mainly regulated by
proteins methyltransferase-like 3 (METTL3), methyltransferase 14
(METTL14), YTH-domain-containing proteins (YTHDFs), insulin-like
growth factor 2 mRNA-binding proteins (IGF2BPs) and demethylases
(FTO and ALKBH5) (22). m6A
methylation affects the physiological process of cells by
regulating the fate of downstream mRNAs. These include maintenance
of RNA stability (23-25), mRNA nuclear export (26,27) and microRNA processing (28). The effect on RNA stability is
mainly achieved by m6A 'reader' proteins that are connected to
downstream molecular pathways. In the cytoplasm, the YTHDF protein
family is able to preferentially recognize m6A-containing RNAs and
subsequently direct them to degradation pathways (29). A key member of the m6A
methyltransferase complex is METTL3, which acts as a reversible
epitranscriptomic regulator. During m6A modification, METTL3 acts
as an enzyme responsible for catalyzing m6A deposition and forms a
complex with several other proteins that act together to promote
m6A deposition on RNA (30,31). m6A modification plays a role in
the development of various types of cancer (32,33). In multiple cancer studies, METTL3
has been linked to the malignant advancement of tumors and poor
prognosis of patients, including colorectal cancer (34), cervical cancer (35), hepatocellular carcinoma (36) and GC (37). Its regulatory mechanisms include
glycolysis, self-renewal, tumorigenicity and metastasis of tumor
cells. However, the specific molecular mechanisms through which
METTL3 is involved in immune escape in GC cells remain unclear.
The present study aims to investigate the role of
TBX15 in mediating immune escape of GC cells, analyze and determine
the regulatory relationship between TBX15 mRNA and m6A
methylation modification, so as to enhance the understanding of
TBX15 in GC cell immune escape and provide new ideas for future
GC-targeted therapy.
Materials and methods
Materials
The human gastric mucosal epithelial cell (GES-1;
cat. no. AW-CNH199), human GC cell lines HGC27 (cat. no. AW-CNH127)
and MKN74 (cat. no. AW-CCH277), mouse forestomach carcinoma (MFC;
cat. no. AW-CCM404) and tohoku hospital pediatrics-1 (THP-1;
AW-CCH098) were purchased from Abiowell (https://www.abiowell.com/). All plasmids were
purchased from Honor Gene Co., Ltd. These included TBX15
silencing (si-TBX15; cat. no. HG-SH152380), TBX15
overexpression (oe-TBX15; cat. no. HG-HO152380), matrix
metalloproteinase 14 (MMP14) overexpression
(oe-MMP14; cat. no. HG-HO004995), METTL3 silencing
(si-METTL3#1-3; cat. no. HG-SH019852), MMP14-wt (cat.
no. HG-YO020961) and MMP14-mut (cat. no. HG-YO020961M). Male
C57BL/6 mice (6 weeks) were ordered from Hunan SJA Laboratory
Animal Co., Ltd. TRIzol (Thermo Fisher Scientific, Inc.), mRNA
reverse transcription kit (CWBIO), and UltraSYBR Mixture (CWBIO)
were used for reverse transcription-quantitative PCR (RT-qPCR)
analysis. 5-Ethynyl-2′-deoxyuridine (EdU) kit (cat. no. C10310) was
purchased from Guangzhou RiboBio Co., Ltd. Annexin V-APC assay kit
(Nanjing KeyGen Biotech Co., Ltd.) and TUNEL assay kit (cat. no.
40306ES50; Yeasen Biotechnology) (https://www.yeasen.com/) were used to detecT cell
death. The inducible nitric oxide synthase (iNOS; cat. no.
CSB-E08326m), IL-1β (cat. no. CSB-E08054m), TNF-α (cat. no.
CSB-E04741m), Arg-1 (cat. no. CSB-EL002005MO), IL-10 (cat. no.
CSB-E04594m) and CD206 (cat. no. CSB-EL014782MO) kits were
purchased from Wuhan Huamei Biotech Co., Ltd. The m6A assay kit
(cat. no. P-9005; EpiGentek) was used to analyze m6A detection.
MeRIP-qPCR kit (cat. no. 17-700) and Actinomycin D (cat. no. A4262)
were purchased from MilliporeSigma. A dual luciferase assay kit
(cat. no. E1910) was purchased from Promega Corporation. All
antibody information is shown in Table SI.
Data acquisition and bioinformatics
analysis
The RNA sequencing profiles and clinical data for 33
cancerous from the Cancer Genome Atlas (TCGA, https://portal.gdc.cancer.gov/) and
Genotype-Tissue Expression (GTEx) cohorts (GTEx Analysis Release
V8, https://gtexportal.org) were retrieved
from the University of California Santa Cruz Xena database
(https://xenabrowser.net/datapages/).
Univariate Cox regression (uniCox) and Kaplan-Meier analyses were
performed. The R packages 'survminer' and 'survival' visualized the
results, including overall survival (OS), disease-specific survival
(DSS), disease-free interval (DFI) and progression-free interval
(PFI) of patients with GC. JASPAR online software (https://jaspar.elixir.no/) was used to predict
potential downstream targets of transcription factors. SRAMP online
software (http://www.cuilab.cn/sramp) predicted
m6A modification sites on RNA. RM2Target (http://rm2target.canceromics.org/#/home) predicted
m6A-associated enzymes that bind to target genes.
Collection and processing of clinical
samples
GC tissue samples (n=10) and corresponding
paracancerous tissue samples were collected. All samples were
obtained from patients with GC in Zhuzhou Hospital Affiliated to
Xiangya School of Medicine, Central South University between May
2024 and May 2025. The procedures followed in the present study
were approved (approval no. ZZCHEC2022070-01) by the Ethics
Committee (Zhuzhou, China) and performed followed Declaration of
Helsinki Guideline. Written informed consent was obtained from each
enrolled subject. The baseline information statistics of patients
are shown in Table SII.
Cell culture and treatments
Cells were cultured in RPMI-1640 containing 10%
fetal bovine serum (both from Gibco; Thermo Fisher Scientific,
Inc.) and 1% penicillin/streptomycin. The cell incubator
environment was maintained at 37°C with a humidified atmosphere
containing 5% CO2.
To explore the effects of TBX15, MMP14 and METTL3 on
GC cells, plasmid intervention experiments of si-TBX15,
oe-TBX15, oe-MMP14 and si-METTL3#1-3 were
performed. The corresponding plasmids and negative control (si-NC
and oe-NC) were transfected into cells. si-TBX15#1-3
sequences are as follows: si-TBX15#1, 5′-GAC AAT AAA AGA TAC
AGA TAT-3′; si-TBX15#2, 5′-GGG GTG AAA ACG TTC AAC TTT-3′;
si-TBX15#3, 5′-GAC ATA TAC CCA AGA ACA AGA-3′.
si-METTL3#1-3 sequences are as follows: si-METTL3#1,
5′-GAG TTG ATT GAG GTA AAG CGA-3′; si-METTL3#2, 5′-TAG AGC
TAT TAA ATA CTA CAA-3′; si-METTL3#3, 5′-GAC GAA TTA TCA ATA
AAC ACA-3′. si-NC sequence is 5′-TGC GGG GCT AGG GTC CAA
CGG-3′.
A co-culture system of GC cell-TAM cells was
constructed. The THP-1 was resuspended in RPMI-1640 medium and
seeded in 6-well culture plates at a density of 7.5×105
cells/well. THP-1 cells were treated with phorbol 12-myristate
13-acetate (PMA; 320 nmol/l) for 6 h and then co-cultured with
tumor cells for 24 h. After washing the cells with PBS, they were
resuspended in serum-free RPMI-1640 medium. Macrophages were
harvested, and flow analysis was performed.
A co-culture system of MFC cell-CD8+ T
cells was established. Spleen-derived CD8+ T cells were
isolated using MojoSort™ Mouse CD8+ T cell Isolation kit
(BioLegend, Inc.). Isolated CD8+ T cells were activated
with Ultra-LEAF™ purified anti-mouse CD3/CD28 (BioLegend, Inc.) for
3 days following the manufacturer's protocol. IL-2 (10 ng/ml) was
added to RPMI-1640 medium for experiments. Transfected MFC cells
were co-incubated with activated CD8+ T cells for 48 h.
The ratio of tumor cells to CD8+ T cells was 1:1-1:20.
CD8+ T cells were collected and analyzed for granzyme B
(GZMB), perforin (PFP), CD100, IFN-γ and TNF-α expression.
Animals
A total of 48 C57BL/6 mice (male, 6 weeks old, ~20
g) were maintained under pathogen-free conditions for 1 week. Mice
were raised on a 12 h/12 h light/dark cycle (22~25°C) with ad
libitum access to water and food. All animal experiments were
conducted in accordance with animal care, animal welfare and
ethics. It has been approved by the Institutional Animal Care and
Use Committee (IACUC) of the Second Xiangya Hospital, Central South
University (approval no. 2022722).
Experiment 1: Mice were randomly divided into 3
groups (n=6): Control group, sh-NC group and sh-TBX15 group.
Mice in the sh-NC and sh-TBX15 groups were injected with
1×106 MFC cells (100 µl, resuspended in PBS) that
had been transfected with sh-NC and sh-TBX15 plasmids. At
room temperature, 3 µg of sh-NC and sh-TBX15 plasmids were
transfected into MFC cells by using Lipofectamine™ 2000
(Invitrogen; Thermo Fisher Scientific, Inc.). At 48 h after
transfection, cells were collected immediately for injection. The
injection site was the right underarm. MFC cells injected in the
Control group were not treated. The body weight and tumor were
observed twice a week, and the tumor volume was measured with
vernier calipers. The tumor volume was calculated by the formula
V=(L × W2)/2, where V was the volume (mm3), L
was the maximum diameter (mm), and W was the minimum diameter (mm).
The experiment was finished 21 days after tumor seeding, and
animals were euthanized by intraperitoneal injection of 200 mg/kg
sodium pentobarbital solution. Samples were received and images
were captured after euthanasia. sh-TBX15 sequence is 5′-GAC
AAT AAA AGA TAC AGA TAT-3′ and sh-NC sequence is 5′-TGC GGG GCT AGG
GTC CAA CGG-3′.
Experiment 2: Mice were randomly divided into 5
groups (n=6): Control group, sh-NC group, sh-METTL3 group,
sh-METTL3 + oe-NC group, sh-METTL3 + oe-TBX15
group. Consistent with the aforementioned intervention procedure,
MFC cells subjected to sh-METTL3 and oe-TBX15 interventions
were injected into the right armpit of mice. After 21 days, mice
were euthanized by intraperitoneal injection of sodium
pentobarbital solution (200 mg/kg) and tumors were collected.
sh-METTL3 sequence is 5′-GAC GAA TTA TCA ATA AAC ACA-3′.
sh-NC sequence is 5′-TGC GGG GCT AGG GTC CAA CGG-3′.
RT-qPCR analysis
To detect gene expression in tissues and cells, qPCR
analysis was performed (38).
Total RNA was extracted from GC tissues, mouse tumor bodies, and
cells using TRIzol reagent, and the concentration was determined by
ultraviolet spectrophotometer. cDNA was synthesized using an mRNA
reverse transcription kit according to the manufacturer's
instructions. qPCR was applied to detect the mRNA levels using
UltraSYBR Mixture. PCR reaction conditions were 95°C for 10 min,
and 95°C for 15 sec, and 60°C for 30 sec, for 40 cycles. Melting
curve analysis was performed with a temperature gradient of
60-95°C. The primers were designed by primer5 (https://premierbiosoft.com/) and synthesized by the
TsingKe Biological Technology. The relative mRNA levels of the
target genes were determined by the 2−ΔΔCq method
(39) and normalized to β-actin
(as an endogenous control). All primer sequences are included in
Table SIII.
Western blotting
To determine the protein levels of TBX15, MMP14,
Granzyme B, Perforin, CD100, IFN-γ, TNF-α and METTL3, western
blotting was performed. Referring to a previous study (40), total protein was extracted from GC
tissues and cells using the RIPA lysis buffer (Beyotime Institute
of Biotechnology). A BCA assay kit was performed to detect protein
concentrations. Equal amounts of protein (15-20 µg) were
separated on a 10% SDS-PAGE gel and then transferred to a
nitrocellulose membrane. After 5% skim milk was used for incubation
and blocking for 90 min at room temperature, freshly prepared
specific primary antibodies were used to incubate the membranes at
4°C overnight. After incubation with the secondary antibody (90
min, room temperature) was completed, ECL chemiluminescence
solution was added to the membrane, followed by imaging using a gel
imaging system (ChemiScope6100, Clinx). β-actin was applied to
normalize the target protein levels. Relative expression level of
the target protein=gray value of target protein/gray value of
internal reference (β-actin).
Immunohistochemistry (IHC) analysis
After the mouse tumor and GC tissues were fixed in
paraffin for 24 h at room temperature, the samples were sectioned
(2 µm) (25). After
deparaffinization with xylene, it was then rehydrated with graded
ethanol series. The sections were immersed in 0.01 M sodium citrate
antigen repair solution (pH 6.0) and heated in a microwave oven for
thermal antigen repair. Incubation with 1% periodate for 15 min was
used to inactivate endogenous enzymes. After incubation with
specific primary antibodies (4°C overnight) anti-TBX15, anti-Ki67,
anti-PD-L1, anti-CD100, anti-IFN-γ and anti-TNF-α, sections were
subjected to secondary antibody incubation for 30 min at 37°C.
After DAB color development, hematoxylin counterstain was
performed, after which section images were randomly acquired. Data
are presented as the mean IOD and positive rate. Positive rate was
calculated as follows: (Number of positive cells/the number of
total cells) ×100%.
Cell proliferation assays
To detect the proliferative activity of GC cells,
Cell Counting Kit-8 (CCK-8) assay (Dojindo Laboratories, Inc.) and
5-Ethynyl-2′-deoxyuridine (EdU) analysis were performed. For CCK-8
assay, 10 µl CCK-8 reagent was added to prepared cells
(5×103 cells) and incubated for 4 h at 37°C, followed by
detection of absorbance values (OD values) in each well at 480
nm.
The EdU cell proliferation kit was performed. Cells
were seeded into a culture medium containing EdU reagent and
incubated overnight. Cells were fixed by incubation with 4%
paraformaldehyde for 30 min at room temperature. After incubation
with 100 µl of 1X Apollo® reaction solution for
staining, 1X Hoechst33342 reaction solution was added for staining
(30 min at room temperature). Images were captured using a
fluorescence microscope. Cell proliferation rate was calculated as
follows: (Number of Edu-positive cells/total number of cells)
×100%.
Flow cytometry (FCM)
To analyze the level of apoptosis, an Annexin V-APC
assay kit was performed (25).
Prepared cells were harvested. The Annexin V-APC and Propidium
Iodide reagents were added. Reaction was performed at room
temperature and in the dark for 10 min before analysis using flow
cytometry (CytoFLEX; Beckman Coulter, Inc.). Data were analyzed
with the use of CytExpert software (version 2.5; Beckman Coulter,
Inc.). Apoptotic rate (%) was calculated as follows: Early
apoptotic rate (%) + late apoptotic rate (%).
To analyze the polarization of M1 and M2
macrophages, CD86 and CD206 positive cells were identified using
flow cytometry. After digestion and washing of the cells, anti-CD86
or anti-CD206 antibodies were added. FCM was used for analysis
after 30 min of staining in the dark. Percentage of positive cells
(%) was calculated as follows: (Cells expressing a marker/targeT
cell population) ×100%.
Wound healing assay
GC cells were seeded in 6-well culture plates until
a monolayer was formed. A micropipette tip (200 µl) was used
to generate the wound. Sterile PBS was used to wash and remove
suspended cells. Image of the scratch was captured at 0 h. After
incubation in serum-free Dulbecco's Modified Eagle Medium (DMEM)
medium for 24 and 48 h, images were captured again for recording. A
total of 3 visual fields were taken at each time point.
Invasion assays
Cell invasion ability was determined by performing
Transwell assays (41) with
8-µm-pore membranes. Stably transfected GC cells
(2×106 cells/well) were seeded into the upper chamber,
and 10% fetal bovine serum culture medium was added to the lower
chamber. After culturing at 37°C for 48 h, cells on the upper
surface of the filter were removed by wiping with a cotton ball.
Next, the filter was fixed with 4% paraformaldehyde for 20 min and
stained with 0.1% crystal violet for 5 min at room temperature. A
total of three randomly selected fields were counted under a light
microscope.
TUNEL staining
To detect apoptosis in the tumor tissue, TUNEL
staining was performed using a TUNEL assay kit according to the
manufacturer's instructions. DAPI working solution (5 µg/ml)
was used to stain the nuclei. A fluorescence microscope was used to
view the images. A total of 6 fields of view were randomly selected
for assessment. Data are presented in the form of positive rates.
Positive rate was calculated as follows: (Number of positive
cells/the number of total cells) ×100%.
Measurement of iNOS, IL-1β, TNF-α, Arg-1,
IL-10 and CD206 levels
After total protein extraction from mouse tumor
bodies, the levels of iNOS, IL-1β, TNF-α, Arg-1, IL-10 and CD206
were measured by commercial kits according to the manufacturer's
instructions. An absorbance (OD) value at 450 nm was used to
quantify protein levels. Final sample concentration equals measured
concentration/protein concentration.
Dual-luciferase reporter assay
A dual luciferase assay kit was used to detect
targeting between TBX15 and MMP14 according to manufacturer's
instructions. The MMP14-wt, MMP14-mut and
TBX15 overexpression plasmid were transiently transfected
into cells using Lipofectamine™ 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). A reference reporter plasmid containing
Renilla luciferase (pRL-TK) was co-transfected into the
cells to regulate the transfection efficiency. After 48 h, cells
were harvested and lysed for 15 min. Cell lysates were obtained for
analysis of luciferase activity. The MMP14-mut sequence is:
5′-ACC ACA CT-3′.
ChIP-qPCR
ChIP-qPCR was performed to analyze the targeting
relationship between TBX15 and MMP14 promoters.
Immunoprecipitation was performed using anti-TBX15 and IgG. qPCR
was performed to quantify the immunoprecipitated DNA. The primers
used for ChIP-qPCR are shown in Table SI.
Detection of m6A methylation levels
The m6A assay kit was performed to detect the global
m6A levels. The cells and tissues were harvested and total RNA was
extracted. The working solution of anti-m6A was used to incubate
the samples for 30 min. The reaction was performed with the
developer solution at room temperature in the dark for 10 min.
After the terminator stop solution for 10 min, the m6A level was
quantified by observing the absorbance value (OD values) at 450 nm.
m6A (ng)=[Sample OD -NC OD)/Slope, m6A %=(m6A Amount (ng)]/S ×100%.
Slope is the slope of the standard curve, and S is the amount of
input sample RNA in ng (200 ng).
1MeRIP-qPCR
A MeRIP-qPCR kit was applied to detect the m6A
levels of the target genes (25).
Total RNA was extracted from GC cells. Samples were coupled to
protein A/G magnetic beads (40 µl) overnight in IP buffer
containing anti-m6A and anti-IgG. m6A-modified TBX15 was
enriched. qPCR was performed to detect TBX15 levels with m6A
modification.
RNA pull-down assays
METTL3 and TBX15 mRNA binding was identified.
The prepared cells were harvested and total protein was extracted.
The TBX15 mRNA probe, which was used as a bait factor, was
bound to streptavidin magnetic beads. Total protein was mixed with
magnetic beads after probe binding and incubated for 60 min. After
elution, proteins were collected and assayed for METTL3 protein
expression.
RNA stability assay
GC cells were cultured in a culture medium
containing Actinomycin D (5 µg/ml) for specific periods of
time (42). Cells were collected
for total RNA extraction. The stability of TBX15 mRNA was
analyzed by measuring the mRNA level of TBX15 by qPCR.
Statistical analysis
All experimental data were presented as the mean ±
standard deviation (SD). GraphPad Prism (version 9.0.0; Dotmatics)
was employed to perform data analysis. The Shapiro-Wilk test was
employed to analyze the normality of the data. The two-tailed
t-test (data comparison between two groups), one-way ANOVA, and
two-way ANOVA (data comparison between multiple groups) were
applied. Post hoc tests for multiple group comparisons were
performed using Tukey's multiple comparisons test. P<0.05 was
considered to indicate a statistically significant for comparison
between groups.
Results
Prognosis-related TBX15 is highly
expressed in GC
Based on TCGA and GTEx pan-cancer data, TBX15
expression was analyzed. TBX15 was enriched in cancer
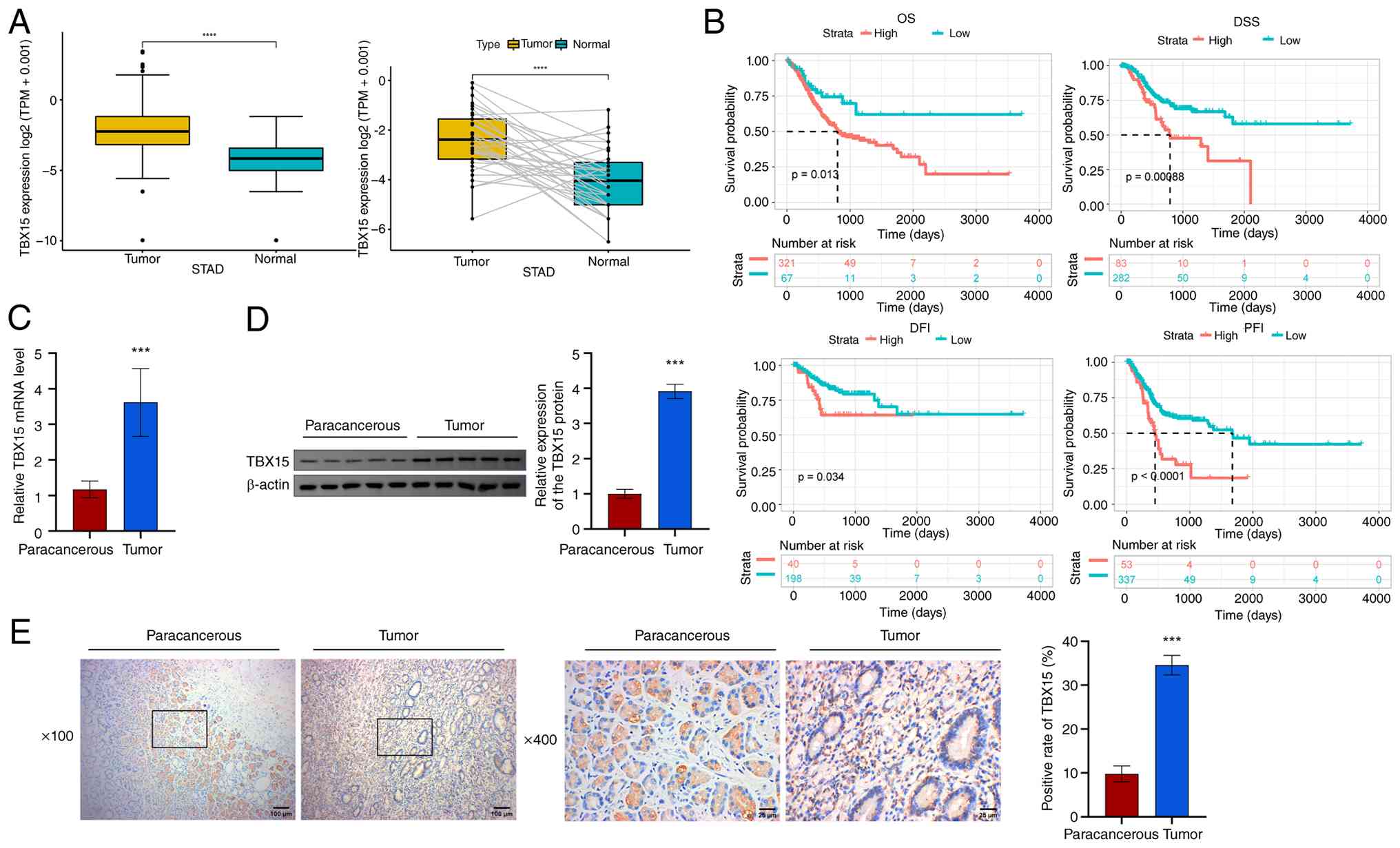
tissues in 21 tumors, including GC (TCGA-STAD) (Fig. S1A). In TCGA-STAD, TBX15
was significantly highly expressed in both unpaired and paired GC
samples (Fig. 1A). Pan-cancer
prognostic data for TBX15 is presented in Fig. S1B. TBX15 was a high-risk
factor for GC (TCGA-STAD) in OS, DSS and PFI analyses (OS, P=0.048;
DSS, P=0.007; PFI, P=0.003). Patients with GC with high
TBX15 predicted shorter survival time (OS, P=0.013; DSS,
P=0.00088; DFI, P=0.034; PFI, P<0.0001; Fig. 1B). Next, the TBX15 levels were
analyzed in the collected GC tissues [TBX15 mRNA: 3.62±0.95;
TBX15 (1G): 3.92±0.20; TBX15 (1H): 34.58±2.22] and their
paracancerous tissues [TBX15 mRNA: 1.18±0.23; TBX15 (1D):
1.00±0.13; TBX15 (1E): 9.80±1.80] (Fig. 1C-E). TBX15 was highly expressed in
GC. The aforementioned results proved that TBX15, which is
associated with poor prognosis in patients with GC, was enriched in
GC.
Knockdown of TBX15 inhibits the
proliferation, migration and invasion of GC cells
To further explore the involvement of TBX15 in GC
cells, the expression of TBX15 was identified in GC cells (HGC27
and MKN74). GES-1 (TBX15 mRNA: 1.00±0.05; TBX15: 0.09±0.01)
was used as control. TBX15 was highly expressed in HGC27
(TBX15 mRNA: 3.30±0.67; TBX15: 0.39±0.03) and MKN74
(TBX15 mRNA: 3.17±0.58; TBX15: 0.43±0.03) cells (Fig. 2A). The TBX15-silenced cells
were successfully constructed in HGC27 and MKN74 (Figs. 2B and S2A). Proliferation ability decreased
after silencing TBX15 (Fig. 2C
and D; Fig. S2B and C).
TBX15 silencing promoted GC cell apoptosis (Figs. 2E and S2D). The effect of TBX15
silencing intervention on GC cell migration was analyzed. At 24 and
48 h, the cell migration distance in the si-TBX15 group
(HGC27: 934.1±253.1; MKN74: 934.1±177.2) was lower than the si-NC
group (HGC27: 788.8±386.5; MKN74: 836.4±251.8) (Figs. 2F and S2E). Transwell was performed to analyze
the invasion ability of GC cells. TBX15 silencing inhibited
cell invasion (Figs. 2G and
S2F). These data proved that
knockdown of TBX15, which is highly expressed in HGC27 and
MKN74, inhibited the proliferation, migration and invasion of GC
cells.
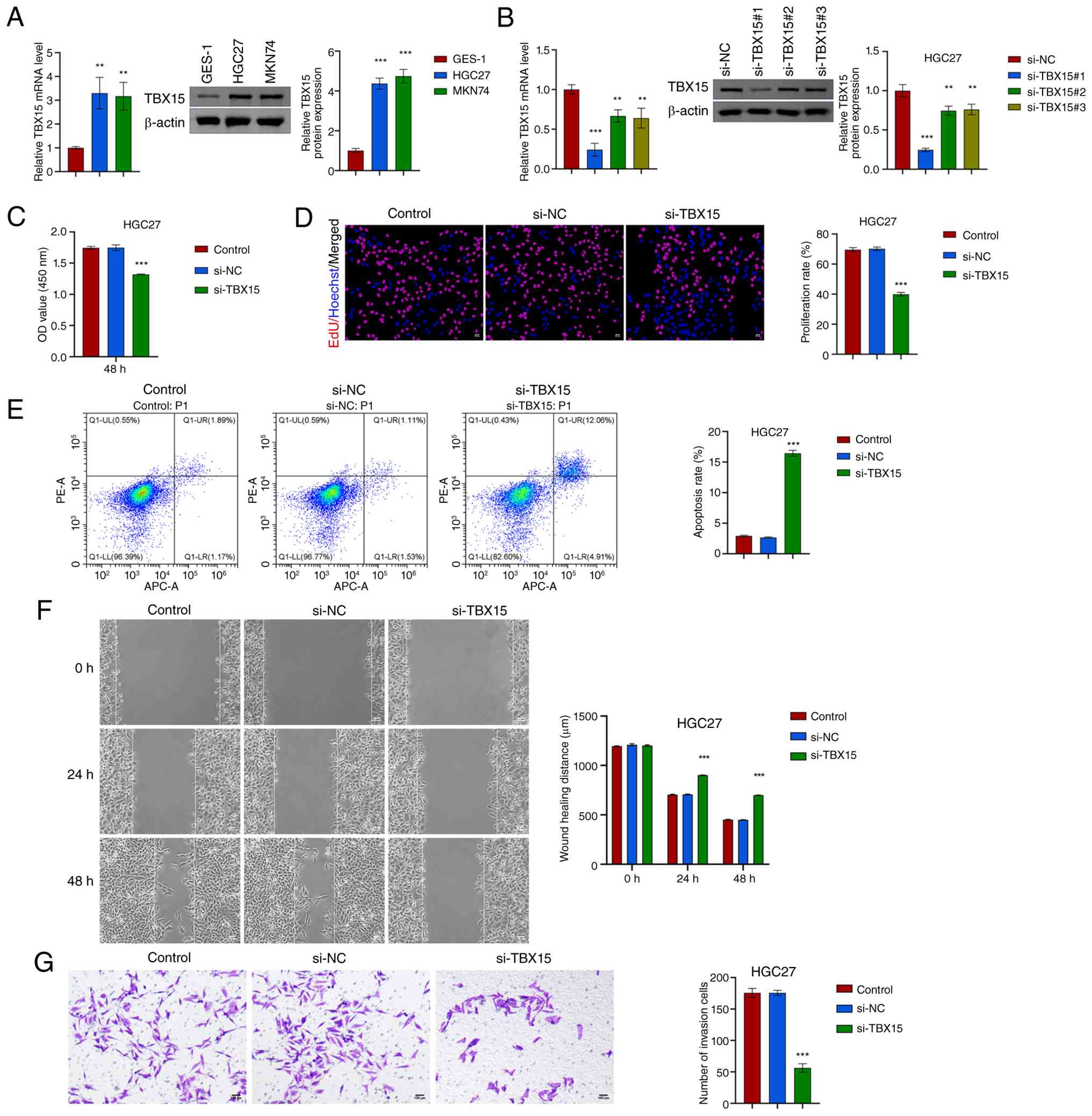 | Figure 2Silencing TBX15 inhibits the
proliferation, migration and invasion of HGC27 cells. (A) In GES-1
and GC cells (HGC27 and MKN74), TBX15 expression was verified.
**P<0.01 and ***P<0.001 vs. GES-1,
one-way ANOVA. (B) TBX15 mRNA and protein levels were
detected after silencing TBX15 in HGC27. (C) Cell Counting
Kit-8 assay. (D) EdU analysis. Scale bar, 50 µm. (E) Flow
cytometric analysis of GC cell apoptosis levels. (F) Wound healing
analysis. Scale bar, 100 µm. (G) Transwell assay was used to
analyze cell invasion ability. Scale bar, 100 µm.
***P<0.001 vs. si-NC, one-way ANOVA and two-way
ANOVA. TBX15, T-box transcription factor 15; GC, gastric cancer;
si-, small interfering; NC, negative control. |
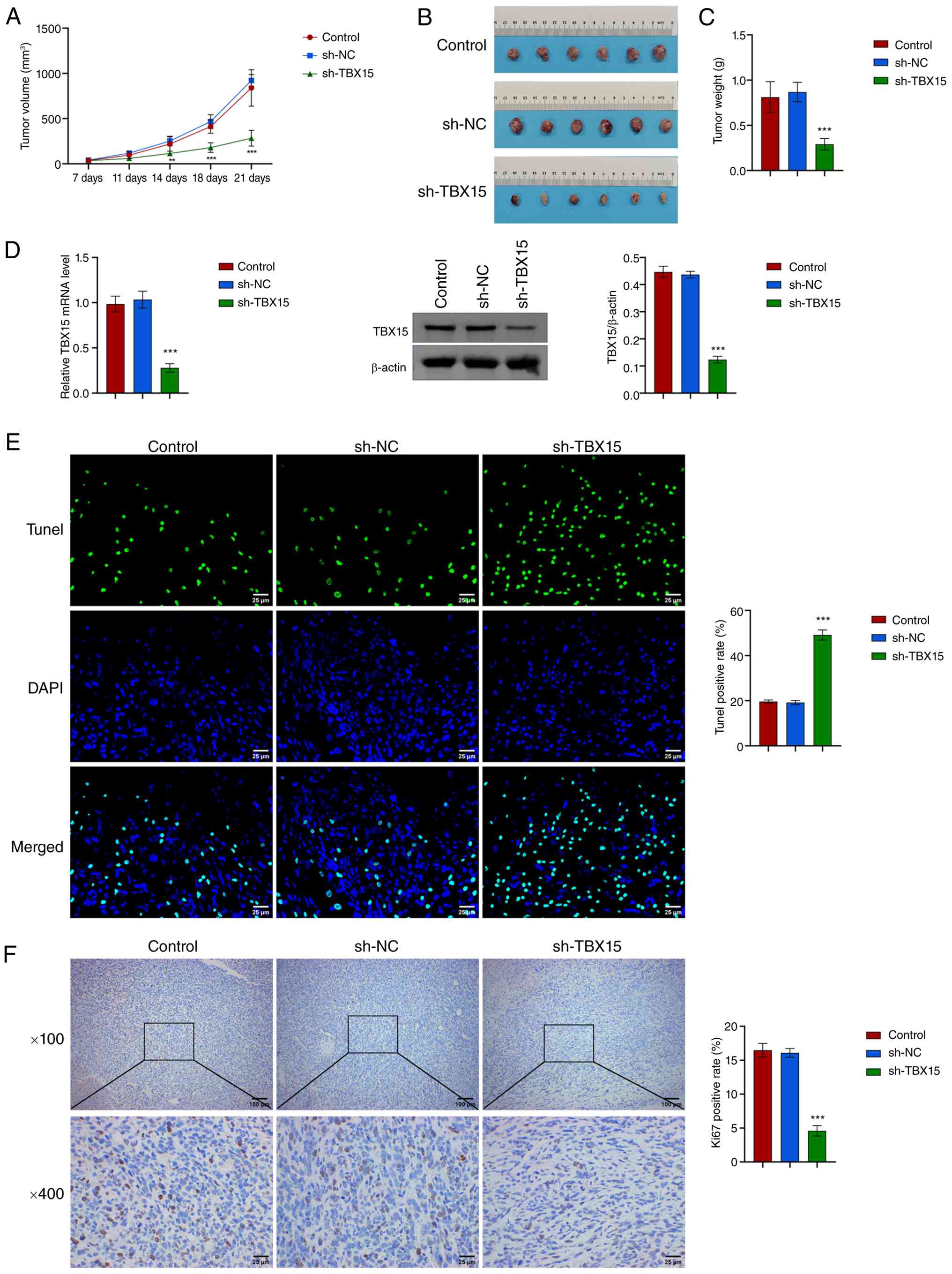
Knockdown of TBX15 inhibits the
tumorigenic ability of MFC cells in mice
Next, the effect of TBX15 on GC development was
analyzed by tumor burden. MFC cells with or without sh-TBX15
intervention were subcutaneously injected into mice to induce tumor
formation. The tumor volume (maximum diameter: 14.78 mm, maximum
volume: 1181.46 mm3) and weight decreased after
TBX15 silencing (Fig.
3A-C). TBX15 silencing reduced tumor burden in mice. The
apoptotic rate increased, and Ki67 levels decreased (Fig. 3D-F). These evidence demonstrated
that TBX15 silencing could inhibit GC development in
vivo.
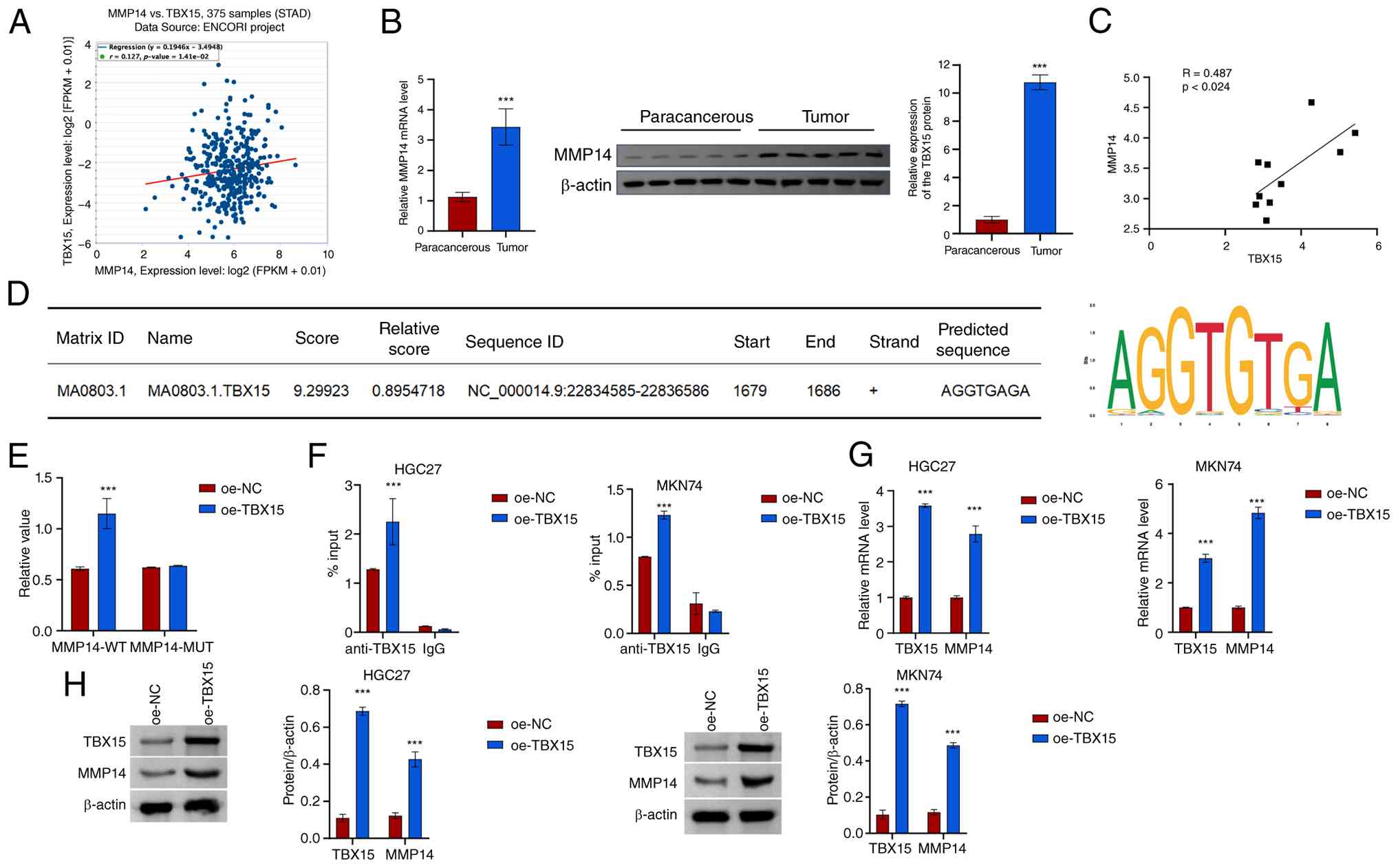
TBX15 regulates MMP14 transcription by
binding to the MMP14 promoter
MMP14 is a membrane-anchored matrix
metalloproteinase. Based on the patient data in TCGA-STAD, the
expression of TBX15 and MMP14 was predicted to be
positively correlated (Fig. 4A).
MMP14 mRNA and protein levels increased in GC tissues, and
TBX15 was positively correlated with MMP14 (Fig. 4B and C). JASPAR online prediction
found that the transcription factor TBX15 might target the
MMP14 promoter (Fig. 4D).
A dual luciferase reporter gene was used to verify the activity of
the MMP14 promoter. Overexpression of TBX15 increased
MMP14 promoter activity (Fig.
4E). ChIP-PCR assay showed that TBX15 interacted with the
MMP14 promoter (Fig. 4F).
Overexpression of TBX15 was found to promote MMP14
mRNA and protein expression (Fig. 4G
and H). These results suggested that the transcription factor
TBX15 may target the MMP14 promoter to regulate its
transcriptional activity.
Knockdown of TBX15 inhibits the immune
evasion of GC cells
The relationship between TBX15 and immune cells was
further explored. In the GC microenvironment, macrophages were
enriched, while CD8+ T cell infiltration level was
significantly reduced (43).
Macrophages play a crucial role in the innate immune system. Under
different microenvironmental stimuli, macrophages can transform
into two different subtypes (M1 and M2), which have completely
different molecular phenotypes and functional characteristics. TAMs
are macrophages that differentiate under the influence of various
factors in the tumor microenvironment (TME) and have the
characteristics and functions of M2-type macrophages. To analyze
the role of TBX15 and MMP14 in GC immune escape, a co-culture
system of GC cells and macrophages was constructed. After
intervention with TBX15 silencing and MMP14
overexpression, GC cells were then co-cultured with macrophages
(Fig. 5A; Fig. S3A and B). TBX15 silencing
promoted the levels of M1-type TAM cells and decreased the M2-type
levels (Fig. 5B and C; Fig. S3C and D). The gating strategy of
Fig. S4A and B was used to gate
the cell subsets. M1 macrophage markers (iNOS, IL-1β
and TNF-α) were increased after TBX15 silencing, and
the opposite was observed for M2 macrophage markers (Arg-1,
IL-10 and CD206) (Fig.
5D and E; Fig. S3E and F).
These results suggested that TBX15 silencing promotes TAMs
toward an M1 proinflammatory phenotype. Overexpression of
MMP14 decreased the levels of iNOS, IL-1β and
TNF-α and increased Arg-1, IL-10 and
CD206. This indicated that overexpression of MMP14
reduced the regulatory effect of TBX15 silencing on
macrophage polarization.
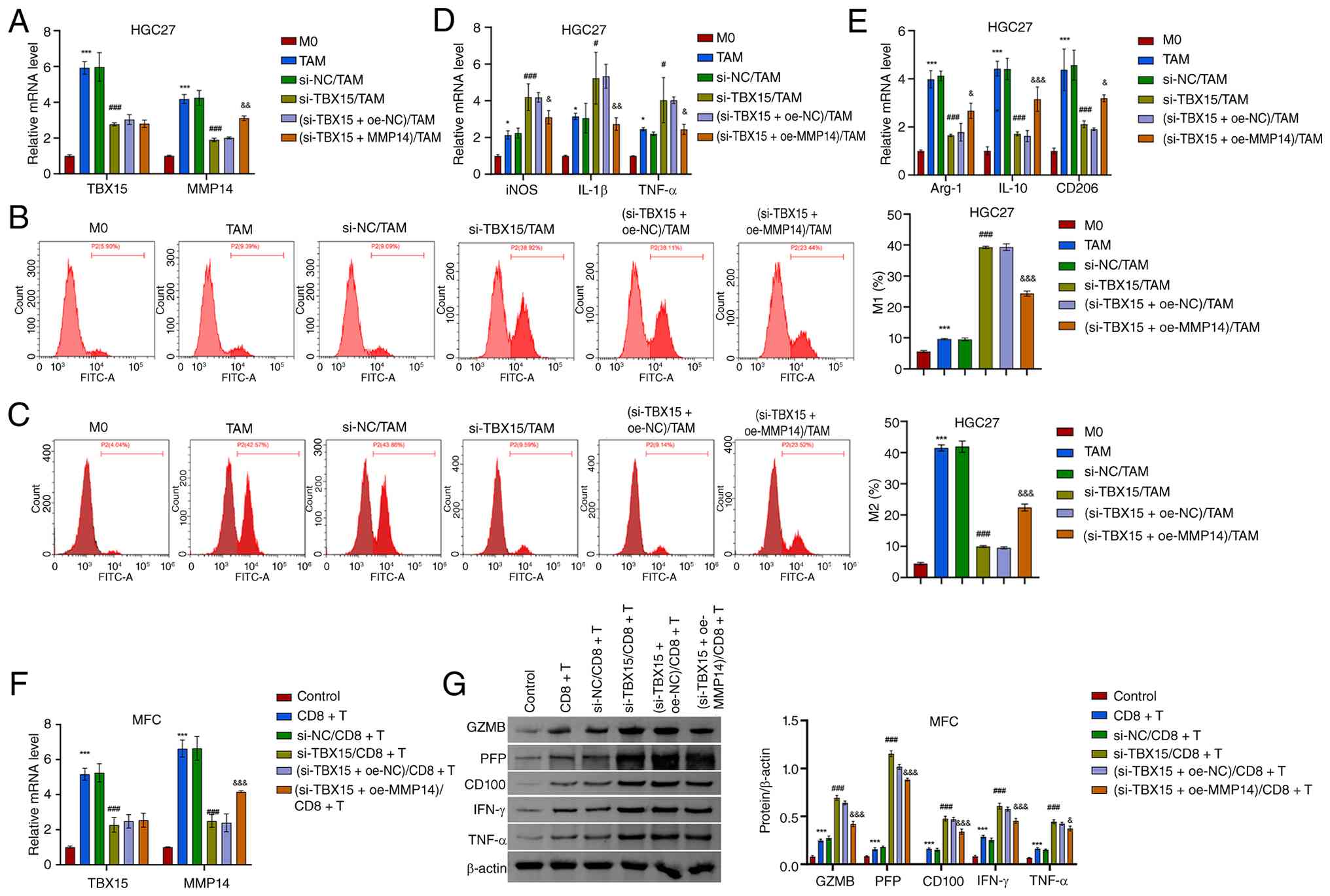 | Figure 5TBX15 and MMP14 participate in
macrophage polarization and CD8+ T cell processes. (A)
TBX15 and MMP14 mRNA levels in HGC27 cells. (B and C)
Percentage of M1- and M2-type macrophages in the co-culture system
of GC cell-TAMs. (D and E) Expression of iNOS, IL-1β,
TNF-α, Arg-1, IL-10 and CD206.
*P<0.05 and ***P<0.001 vs. M0;
#P<0.05 and ###P<0.001 vs. si-NC/TAM;
&P<0.05, &&P<0.01 and
&&&P<0.001 vs. (si-TBX15 +
oe-NC)/TAM, one-way ANOVA and two-way ANOVA. (F) TBX15 and
MMP14 mRNA levels in MFC. (G) Expression of GZMB, PFP,
CD100, IFN-γ and TNF-α in the co-culture system of MFC
cell-CD8+ T cells. ***P<0.001 vs. control;
###P<0.001 vs. si-NC/CD8+ T;
&P<0.05 and &&&P<0.001
vs. (si-TBX15 + oe-NC)/CD8+ T, two-way ANOVA.
TBX15, T-box transcription factor 15; MMP14, matrix
metalloproteinase 14; TAMs, tumor-associated macrophages; iNOS,
inducible nitric oxide synthase; oe-, overexpression; NC, negative
control; si-, small interfering; MFC, mouse forestomach carcinoma;
GZMB, granzyme B; PFP, perforin. |
In addition, CD8+ T cell-associated
cytokines GZMB, PFP, CD100, IFN-γ and TNF-α were identified in the
co-culture system constructed by MFC cells and CD8+ T
cells. In the TBX15-silenced co-culture system, GZMB, PFP,
CD100, IFN-γ and TNF-α levels increased. Silencing TBX15
enhanced CD8+ T cell activation and antitumor immunity.
Overexpression of MMP14 reduced the levels of GZMB, PFP,
CD100, IFN-γ, and TNF-α (Fig. 5F and
G).
In summary, TBX15 participates in the immune escape
process of GC by promoting macrophage M2 polarization and
inhibiting CD8+ T cell activation.
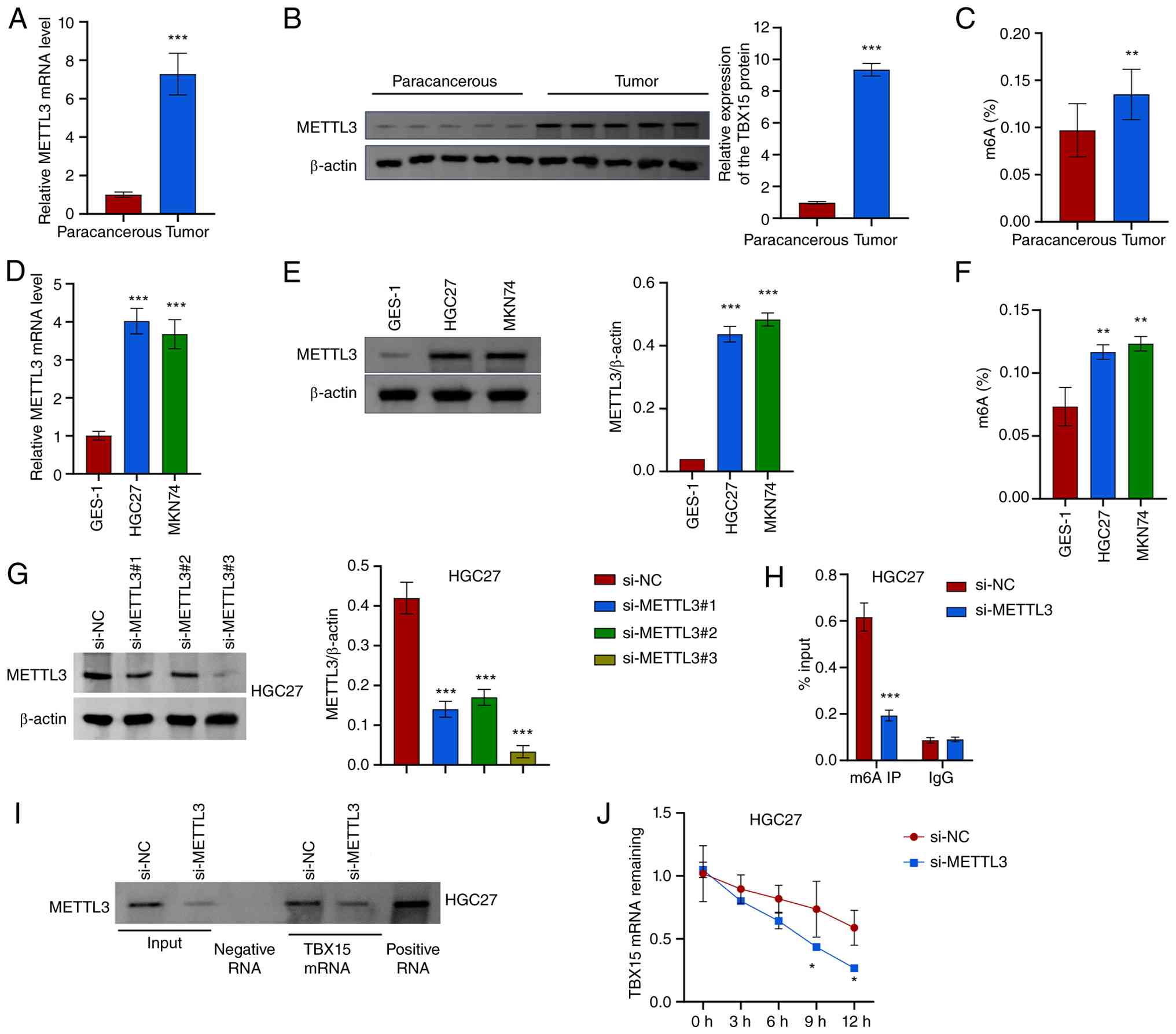
METTL3 mediates m6A modification of
TBX15
The SRAMP online website predicted that
TBX15 might undergo m6A modification, and RM2Target
predicted that TBX15 would bind to the m6A methyltransferase
METTL3 (Fig. S5A and B). In GC,
METTL3 expression and total m6A levels were increased (Fig. 6A-C). METTL3 expression and total
m6A level increased in HGC27 (METTL3 mRNA: 4.02±0.34;
METTL3: 0.44±0.03; m6A%: 0.12±0.01) and MKN74 (METTL3 mRNA:
3.68±0.38; METTL3: 0.48±0.02; m6A%: 0.12±0.01) compared with GES-1
(METTL3 mRNA: 1.00±0.11; METTL3: 0.04±0; m6A%: 0.07±0.02)
(Fig. 6D-F). To further explore
the regulatory relationship between METTL3 and TBX15,
METTL3 silenced cells were constructed (Figs. 6G and S5C). After silencing METTL3, the
m6A level of TBX15 was decreased (Figs. 6H and S5D). Results proved that METTL3 binds
to TBX15 mRNA (Figs. 6I
and S5E). METTL3
silencing reduced TBX15 mRNA stability (Figs. 6J and S5F). These results indicated that
silencing METTL3 inhibits the m6A level of TBX15 and reduces
TBX15 mRNA stability.
METTL3/TBX15/MMP14 signaling axis
regulates proliferation, migration ability and immune escape
process in GC
To explore whether METTL3 regulates GC cell
function and development through the TBX15/MMP14 signaling axis,
METTL3 silencing and TBX15 overexpression cells were
constructed. METTL3 silencing inhibited MMP14 expression,
and TBX15 overexpression attenuated the inhibitory effect of
METTL3 silencing on MMP14 expression (Figs. 7A and S6A). METTL3 silencing decreased
proliferation activity and increased apoptotic rate of GC cells
(Fig. 7B-D; Fig. S6B-D). Overexpression of
TBX15 increased proliferation and decreased apoptosis.
Overexpression of TBX15 reduced the inhibitory effect of
METTL3 silencing on migration (Figs. 7E and S6E). Under the intervention conditions
of METTL3 silencing and TBX15 overexpression, cell
invasion was altered, consistent with migration (Figs. 7F and S6F).
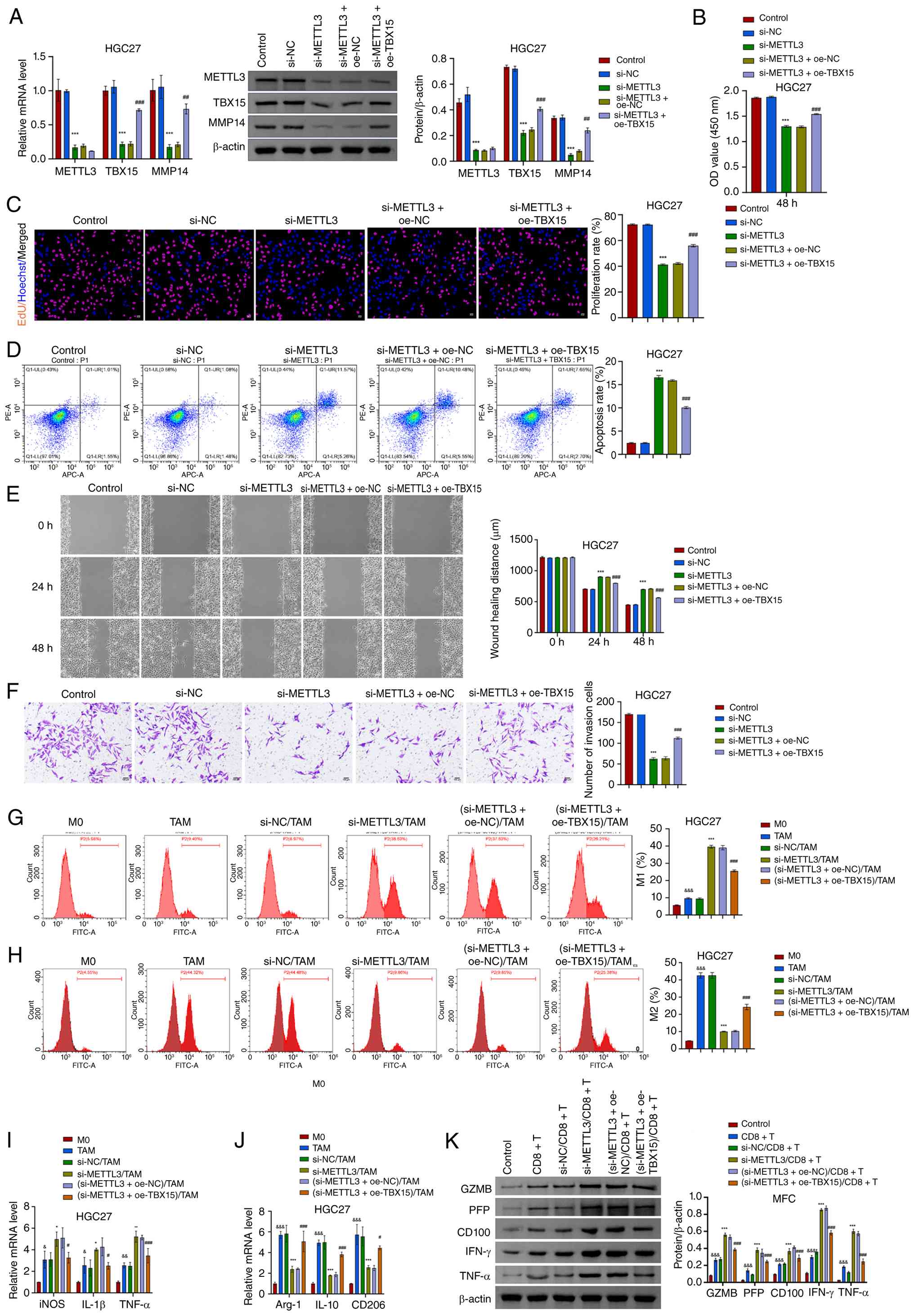 | Figure 7METTL3 regulates GC cell function and
development through the TBX15/MMP14 signaling axis. (A) The
expression of METTL3, TBX15 and MMP14 was identified after
METTL3 silencing and TBX15 overexpression. (B and C)
Analysis of GC cell proliferation activity. Scale bar, 50
µm. (D) Flow cytometric analysis of GC cell apoptosis. (E
and F) Migration and invasion assays. Scale bar, 100 µm.
***P<0.001 vs. si-NC; ##P<0.01 and
###P<0.001 vs. si-METTL3 + oe-NC, one-way
ANOVA and two-way ANOVA. (G and H) In the co-culture system of GC
cell-macrophages, the proportion of M1 and M2 macrophages was
analyzed after METTL3 silencing and TBX15
overexpression intervention. (I and J) Levels of iNOS,
IL-1β, TNF-α, Arg-1, IL-10 and
CD206. &P<0.05,
&&P<0.01 and
&&&P<0.001 vs. M0; *P<0.05,
**P<0.01 and ***P<0.001 vs si-NC/TAM;
#P<0.05 and ###P<0.001 vs.
(si-METTL3 + oe-NC)/TAM, one-way ANOVA and two-way ANOVA.
(K) Levels of CD8+ T cell-related cytokines (GZMB, PFP,
CD100, IFN-γ and TNF-α) expression after intervention of
METTL3 silencing and TBX15 overexpression.
&&&P<0.001 vs. control,
***P<0.001 vs. si-NC/CD8+ T and
###P<0.001 vs. (si-METTL3 +
oe-NC)/CD8+ T, two-way ANOVA. METTL3,
methyltransferase-like 3; GC, gastric cancer; TBX15, T-box
transcription factor 15; MMP14, matrix metalloproteinase 14; NC,
negative control; si-, small interfering; oe-, overexpression;
GZMB, granzyme B; PFP, perforin. |
Next, the role of METTL3/TBX15/MMP14 signaling axis
in GC cell immune escape was further analyzed. Silencing
METTL3 promoted the polarization of TAMs to M1 macrophages
in the co-culture system of GC cell-macrophages (Fig. 7G and H; Fig. S6G and H; Fig. S7A and B). Levels of iNOS,
IL-1β and TNF-α increased, and Arg-1,
IL-10 and CD206 decreased (Fig. 7I and J; Fig. S6I and J). Overexpression of
TBX15 reduced the effect of METTL3 silencing. In the
co-culture system of MFC cell-CD8+ T cells, the
expression of GZMB, PFP, CD100, IFN-γ and TNF-α increased after
METTL3 silencing. Overexpression of TBX15 inhibited
the regulation of METTL3 silencing on CD8+ T
cell-related cytokines GZMB, PFP, CD100, IFN-γ and TNF-α (Fig. 7K). METTL3 regulated proliferation,
migratory activity and immune escape process through TBX15/MMP14
signaling axis in GC.
METTL3 regulates the tumorigenic ability
of MFC cells in mice through TBX15/MMP14 axis
The effect of METTL3/TBX15/MMP14 signaling axis on
MFC cells' ability to induce tumorigenesis in mice was further
analyzed. MFC cells with METTL3 silencing and TBX15
overexpression were injected subcutaneously into mice. After 21
days, tumor volume and weight were reduced after METTL3
silencing. Overexpression of TBX15 attenuated the effect of
METTL3 silencing, and the tumor weight and volume (maximum
diameter: 14.99 mm, maximum volume: 1,385.54 mm3)
increased (Fig. 8A-C). Levels of
METTL3, TBX15 and MMP14 were reduced after METTL3 silencing.
Overexpression of TBX15 promoted TBX15 and MMP14 levels but
had no significant regulatory effect on METTL3 (Fig. 8D and E). In the tumor, the
apoptotic rate increased and the expression of Ki67 decreased after
METTL3 silencing. Overexpression of TBX15 decreased
the apoptotic rate and increased the expression of Ki67 (Fig. 8F and G). Similarly, the expression
of M1 macrophage markers (iNOS, IL-1β and TNF-α) was increased
after METTL3 silencing, and overexpression of TBX15
reduced the effect of METTL3 silencing. Levels of M2
macrophage markers (Arg-1, IL-10 and CD206) were opposite (Fig. 8H). Overexpression of TBX15
inhibited the regulation of METTL3 silencing on
CD8+ T cell-related cytokines GZMB, PFP, CD100, IFN-γ
and TNF-α (Figs. 8I and S8). The aforementioned results
suggested that silencing METTL3 inhibits tumor development,
via the TBX15/MMP14 signaling axis.
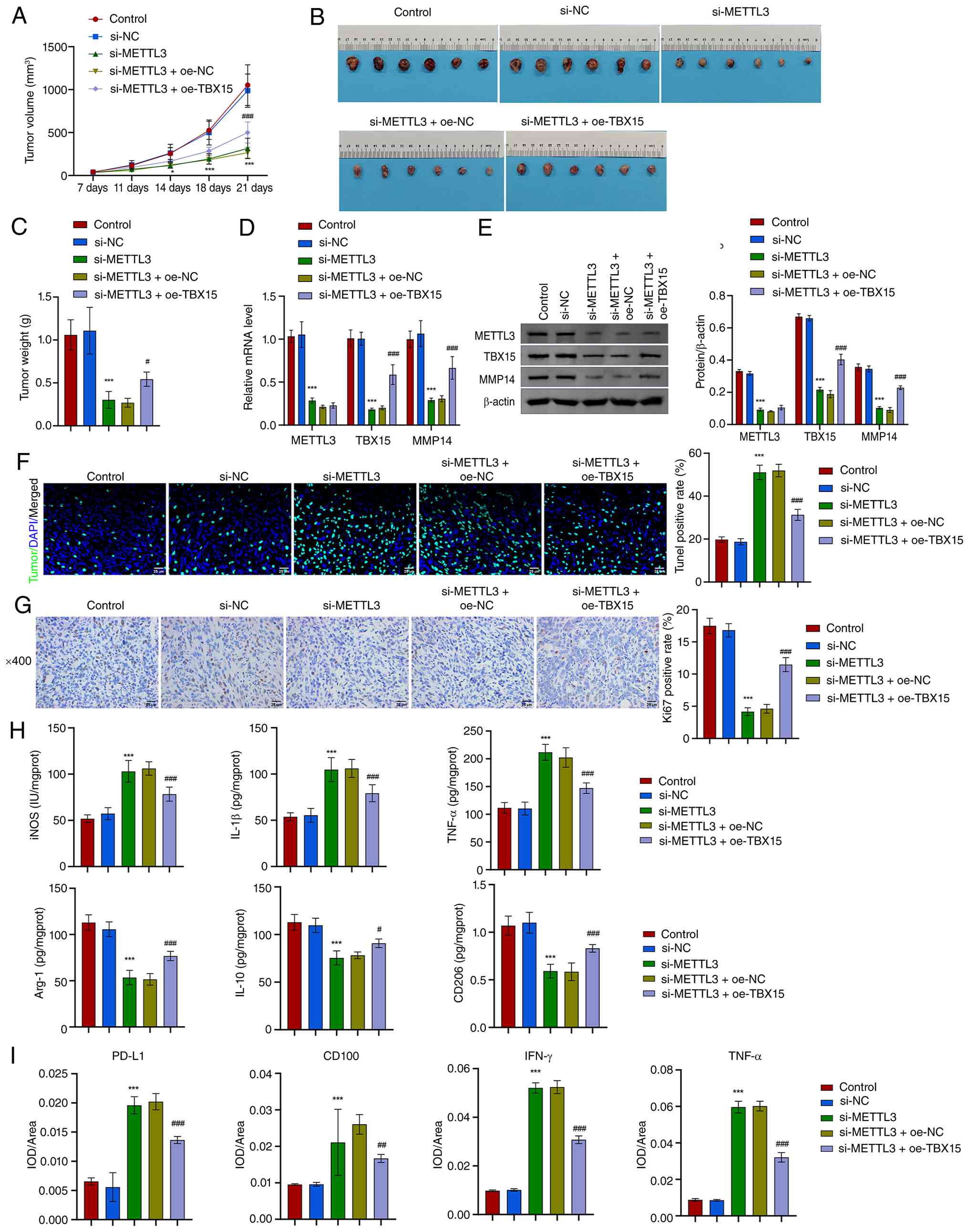 | Figure 8In vivo experimental
validation of the METTL3/TBX15/MMP14 axis. (A) Tumor growth curve.
(B) Images of the tumor. (C) Tumor weight. (D and E) METTL3, TBX15
and MMP14 levels in tumors. (F) TUNEL analysis. Scale bar, 25
µm. (G) Ki67 expression via immunohistochemical analysis.
Scale bar, 25 µm. (H) Levels of iNOS, IL-1β, TNF-α, Arg-1,
IL-10 and CD206. (I) PD-L1, CD100, IFN-γ and TNF-α expression via
immunohistochemical analysis. *P<0.05 and
***P<0.001 vs. sh-NC; #P<0.05,
##P<0.01 and ###P<0.001 vs.
sh-METTL3 + oe-NC, one-way ANOVA and two-way ANOVA. METTL3,
methyltransferase-like 3; TBX15, T-box transcription factor 15;
MMP14, matrix metalloproteinase 14; iNOS, inducible nitric oxide
synthase; PD-L1, programmed death-ligand 1; IFN, interferon; NC,
negative control; oe-, overexpression; sh-, short hairpin. |
Discussion
The experimental design of the present study is
shown in Fig. S5. It is
suggested that high expression of TBX15 in GC was linked to shorter
survival of patients. Suppression of TBX15 led to elevated
levels of apoptosis and hindered the proliferation and invasion of
GC cells. Consistent with a previous study, TBX15 silencing
stimulated increased apoptosis in cancer cells (44), and high levels of TBX15
predicted poor prognosis in patients with cancer (19). This evidence suggested that
TBX15 is regulated by m6A methylation modification and
interacts with the m6A methyltransferase METTL3. In a GC study,
METTL3 was associated with tumor malignant progression and poor
prognosis of patients (37).
Silencing METTL3 reduced TBX15 mRNA stability.
Overexpression of TBX15 reduced the inhibitory effect of
METTL3 silencing on GC cell proliferation and invasion to a
certain extent. Meanwhile, TBX15 could regulate the transcription
of MMP14 by targeting its promoter. In GC, MMP14 is a target
enzyme used to detect GC peritoneal metastasis (45), and its high expression predicts
poor prognosis of patients with GC (46). As one of the membrane-type matrix
metalloproteinases, MMP14 plays an important role in tumor
development and metastasis by promoting the decomposition of cell
adhesion molecules, destroying the basement membrane and destroying
the extracellular matrix (47,48). Therefore, it is reasonable to
hypothesize that TBX15 regulated by m6A methylation
modification may have the involvement of MMP14 in controlling the
migration and invasion process of GC cells.
In addition, overexpression of TBX15 reduced
glycolysis, glucose uptake and lactate production in GC cells
(21). The 'glycolytic switch',
also known as the 'Warburg effect', can lead to the accumulation of
lactate and lactate in the tumor environment, thereby promoting
tumor immune escape (49).
Overexpression of MMP14 reversed the inhibitory effect of
TBX15 silencing on M2 macrophage polarization and the
promoting effect on M1 macrophages in the GC cell-TAM cell
co-culture system. M2 macrophages are important tumor supporting
cells in tumor development and promote the immune escape of tumor
cells by regulating the immune checkpoints (50,51). MMP14 was involved in the
polarization of M2 macrophages (52). Blockade of MMP14 increases iNOS
and polarizes macrophages to an antitumor phenotype (53). This further confirms the findings
that TBX15, an intracellular transcription factor, regulates
macrophage to tumor-promoting phenotypic polarization by targeting
MMP14, thereby promoting cancer development. On the other
hand, CD8+ T cells can participate in antitumor immunity
by secreting inflammatory cytokines (IFN-γ and TNF-α) and cytotoxic
factors (GZMB and PFP) (54). In
the MFC cell-CD8+ T cell co-culture system,
overexpression of MMP14 inhibited the expression of
antitumor related cytokines of CD8+ T cells, including
GZMB, PFP, IFN-γ and TNF-α. As effective effector cells in the TME,
CD8+ T cells can induce tumor cell apoptosis by
secreting cytotoxic factors (GZMB and PFP). In the tumor immune
microenvironment, cells with high levels of MMP14 can transport
tsRNA through exosomes and inhibit the activity of CD8+
T cells (55). MMP14 promotes the
shedding of membrane-bound CD100 (mCD100) from the membrane of
CD8+ T cells, thereby promoting the elevation of
CD8+ T cell response (56). Thus, the TBX15/MMP14 signaling
axis promotes tumor development and cancer cell immune escape in GC
cells, possibly by promoting M2 macrophage polarization and
inhibiting CD8+ T cell antitumor activity. In the
future, the regulatory mechanism of MMP14 in immune escape of GC
cells will be further refined.
It should be pointed out that the present study did
not investigate whether m6A readers are involved in TBX15 mRNA
stability or translational regulation. Considering that METTL3
mainly plays the function of m6A 'writer', the transmission of its
regulatory effect is often dependent on the downstream mediation of
the m6A readers. Therefore, the current study has not fully
revealed the regulatory pathway of 'METTL3-m6A-Reader-TBX15', which
is the main limitation and the direction of future research needs
to be focused on improving. In future studies, it is planned to
collect clinical samples and analyze the correlation between m6A
readers and TBX15 based on clinical samples and further explore its
internal regulatory pathways by RIP-qPCR and co-expression analysis
of m6A readers. The authors acknowledge that although the clinical
sample (n=10) provided preliminary evidence, it was insufficient to
ensure statistical generalizability, and potential selection bias
(for example from a single institution) cannot be ruled out.
Validation in a larger multicenter cohort is needed. Limited to a
single subcutaneous model may not fully recapitulate the
heterogeneous human GC immune microenvironment, particularly immune
cell composition and cytokine signaling. This is also one of the
limitations of the present study. It would be helpful to construct
an orthotopic tumor model to verify the role of target genes in
tumor development in numerous ways. Alternatively, it is noted that
the immortalized cell lines lack a patient-specific genetic
background. Future studies using primary cells are needed. Other
potential biases, including the lack of clinical follow-up and
survival data, are a limitation of the present study. Continuing to
collect patient follow-up and survival information is the authors'
next arrangement.
The results of the present study demonstrated that
the m6A modification of TBX15 is regulated by the m6A
methyltransferase METTL3. TBX15 regulates GC cell proliferation,
migration and invasion through MMP14. The METTL3/TBX15/MMP14
signaling axis promotes the polarization of macrophages to M2 and
inhibits the antitumor activity of CD8+ T cells. The
results of the present study suggested that TBX15 is involved in
the process of proliferation, migration and invasion of GC cells.
Combined with the results of GC cell-TAM cells and MFC
cell-CD8+ T cell co-culture systems, it is suggested
that TBX15 is connected with the polarization of macrophages in the
TME and the antitumor activity of CD8+ T cells. This
suggested that TBX15 may be a promising target for alleviating
immunosuppression.
Supplementary Data
Availability of data and materials
The data generated in the present study are
included in the figures and/or tables of this article. The data
generated in the present study may be requested from the
corresponding author.
Authors' contributions
WQ and HH conceptualized the study, acquired
funding and resources, and wrote, reviewed and edited the
manuscript. HH, RH, ML, XL, QA, MC, QW, WC and WQ curated data. HH
conducted rmal analysis, developed methodology, performed data
validation and visualization, and wrote the original draft. HH, RH,
ML, XL and MC conducted investigation. WQ conducted project
administration and supervised the study. All authors read and
approved the final version of the manuscript. WQ and HS confirm the
authenticity of all the raw data.
Ethics approval and consent to
participate
The procedures followed in the present study were
approved (approval no. ZZCHEC2022070-01) by the Ethics Committee of
Zhuzhou Central Hospital (Zhuzhou Hospital Affiliated to Xiangya
School of Medicine, Central South University; Zhuzhou, China).
Written informed consent was obtained from each enrolled subject.
All animal experiments were performed in accordance with the
relevant guidelines and regulations and were approved by Animal
Ethical and Welfare Committee of The Second Xiangya Hospital
(approval no. 2022722; Zhuzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
The present study was supported by the Natural Science
Foundation of Hunan (grant nos. 2021JJ70076, 2023JJ50217,
2025JJ70002 and 2022JJ50104).
References
|
1
|
Qiu H, Cao S and Xu R: Cancer incidence,
mortality, and burden in China: A time-trend analysis and
comparison with the United States and United Kingdom based on the
global epidemiological data released in 2020. Cancer Commun (Lond).
41:1037–1048. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tan P and Yeoh KG: Genetics and molecular
pathogenesis of gastric adenocarcinoma. Gastroenterology.
149:1153–1162.e3. 2015. View Article : Google Scholar
|
|
3
|
Zhang M, Zhong A, Liu H, Zhao L, Wang Y,
Lu Z, Zhang L, Pan X, Liang Z, Gao L, et al: EZH2 loss promotes
gastric squamous cell carcinoma. Nat Commun. 16:60322025.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tao B, Wang Z, Wang X, Song A, Liu J, Wang
J, Zhang Q, Chen Z, Wang Z, Xu W, et al: An inherited
predisposition allele promotes gastric cancer via enhancing
deubiquitination-mediated activation of epithelial-to-mesenchymal
transition signaling. J Clin Invest. 135:e1796172025. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cancer Genome Atlas Research Network:
Comprehensive molecular characterization of gastric adenocarcinoma.
Nature. 513:202–209. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Keller G, Grimm V, Vogelsang H, Bischoff
P, Mueller J, Siewert JR and Höfler H: Analysis for microsatellite
instability and mutations of the DNA mismatch repair gene hMLH1 in
familial gastric cancer. Int J Cancer. 68:571–576. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Richards FM, McKee SA, Rajpar MH, Cole TR,
Evans DG, Jankowski JA, McKeown C, Sanders DS and Maher ER:
Germline E-cadherin gene (CDH1) mutations predispose to familial
gastric cancer and colorectal cancer. Hum Mol Genet. 8:607–610.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lu L, Mullins CS, Schafmayer C, Zeißig S
and Linnebacher M: A global assessment of recent trends in
gastrointestinal cancer and lifestyle-associated risk factors.
Cancer Commun (Lond). 41:1137–1151. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ajani JA, Lee J, Sano T, Janjigian YY, Fan
D and Song S: Gastric adenocarcinoma. Nat Rev Dis Primers.
3:170362017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Matsuoka T and Yashiro M: Bioinformatics
analysis and validation of potential markers associated with
prediction and prognosis of gastric cancer. Int J Mol Sci.
25:58802024. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Guan WL, He Y and Xu RH: Gastric cancer
treatment: Recent progress and future perspectives. J Hematol
Oncol. 16:572023. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Farin HF, Bussen M, Schmidt MK, Singh MK,
Schuster-Gossler K and Kispert A: Transcriptional repression by the
T-box proteins Tbx18 and Tbx15 depends on Groucho corepressors. J
Biol Chem. 282:25748–25759. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kispert A and Herrmann BG: The Brachyury
gene encodes a novel DNA binding protein. Embo J. 12:3211–3220.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ahmed A, Syed JN, Chi L, Wang Y,
Perez-Romero C, Lee D, Kocaqi E, Caballero A, Yang J,
Escalante-Covarrubias Q, et al: KDM8 epigenetically controls
cardiac metabolism to prevent initiation of dilated cardiomyopathy.
Nat Cardiovasc Res. 2:174–191. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ye W, Wang Y, Hou S, Mei B, Liu X, Huang
H, Zhou Q, Niu Y, Chen Y, Zhang M and Huang Q: USF3 modulates
osteoporosis risk by targeting WNT16, RANKL, RUNX2, and two GWAS
lead SNPs rs2908007 and rs4531631. Hum Mutat. 42:37–49. 2021.
View Article : Google Scholar
|
|
16
|
Pan DZ, Miao Z, Comenho C, Rajkumar S,
Koka A, Lee SHT, Alvarez M, Kaminska D, Ko A, Sinsheimer JS, et al:
Identification of TBX15 as an adipose master trans regulator of
abdominal obesity genes. Genome Med. 13:1232021. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li P, Li Y, Bai S, Zhang Y and Zhao L:
miR-4732-3p prevents lung cancer progression via inhibition of the
TBX15/TNFSF11 axis. Epigenomics. 15:195–207. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ge Y, Jia B, Zhang P, Chen B, Liu L, Shi
Y, Huang S, Liu X, Wang R, Xie Y, et al: TBX15 facilitates
malignant progression of glioma by transcriptional activation of
TXDNC5. iScience. 27:1089502024. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yan D, Yu Y, Ni Q, Meng Q, Wu H, Ding S,
Liu X, Tang C, Liu Q and Yang K: The overexpression and clinical
significance of TBX15 in human gliomas. Sci Rep. 13:97712023.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Golozar M, Motlagh AV, Mahdevar M, Peymani
M, InanlooRahatloo K and Ghaedi K: TBX15 and SDHB expression
changes in colorectal cancer serve as potential prognostic
biomarkers. Exp Mol Pathol. 136:1048902024. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gu P and Wu LN: Sulforaphane targets the
TBX15/KIF2C pathway to repress glycolysis and cell proliferation in
gastric carcinoma cells. Nutr Cancer. 75:1263–1270. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wei G: RNA m6A modification, signals for
degradation or stabilisation? Biochem Soc Trans. 52:707–717. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hu Y, Gong C, Li Z, Liu J, Chen Y, Huang
Y, Luo Q, Wang S, Hou Y, Yang S and Xiao Y: Demethylase ALKBH5
suppresses invasion of gastric cancer via PKMYT1 m6A modification.
Mol Cancer. 21:342022. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shu F, Liu H, Chen X, Liu Y, Zhou J, Tang
L, Cao W, Yang S, Long Y, Li R, et al: m6A modification promotes
EMT and metastasis of castration-resistant prostate cancer by
upregulating NFIB. Cancer Res. 84:1947–1962. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang J, Zhang J, Liu H, Meng L, Gao X,
Zhao Y, Wang C, Gao X, Fan A, Cao T, et al: N6-methyladenosine
reader hnRNPA2B1 recognizes and stabilizes NEAT1 to confer
chemoresistance in gastric cancer. Cancer Commun (Lond).
44:469–490. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jin T, Yang L, Chang C, Luo H, Wang R, Gan
Y, Sun Y, Guo Y, Tang R, Chen S, et al: HnRNPA2B1 ISGylation
regulates m6A-Tagged mRNA selective export via ALYREF/NXF1 complex
to foster breast cancer development. Adv Sci (Weinh).
11:e23076392024. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Qiao Y, Sun Q, Chen X, He L, Wang D, Su R,
Xue Y, Sun H and Wang H: Nuclear m6A reader YTHDC1 promotes muscle
stem cell activation/proliferation by regulating mRNA splicing and
nuclear export. Elife. 12:e827032023. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Alarcón CR, Goodarzi H, Lee H, Liu X,
Tavazoie S and Tavazoie SF: HNRNPA2B1 is a mediator of
m(6)A-dependent nuclear RNA processing events. Cell. 162:1299–1308.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang X, Lu Z, Gomez A, Hon GC, Yue Y, Han
D, Fu Y, Parisien M, Dai Q, Jia G, et al:
N6-methyladenosine-dependent regulation of messenger RNA stability.
Nature. 505:117–120. 2014. View Article : Google Scholar :
|
|
30
|
Bokar JA, Shambaugh ME, Polayes D, Matera
AG and Rottman FM: Purification and cDNA cloning of the
AdoMet-binding subunit of the human mRNA
(N6-adenosine)-methyltransferase. RNA. 3:1233–1247. 1997.PubMed/NCBI
|
|
31
|
Zaccara S, Ries RJ and Jaffrey SR:
Reading, writing and erasing mRNA methylation. Nat Rev Mol Cell
Biol. 20:608–624. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zeng Y, Luo Y, Zhao K, Liu SS, Wu KK, Wu
YY, Du KK, Pan W, Dai Y, Liu Y, et al: m6A-mediated induction of
7-dehydrocholesterol reductase stimulates cholesterol synthesis and
cAMP signaling to promote bladder cancer metastasis. Cancer Res.
84:3402–3418. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang J, Fan P, Shen P, Fan C, Zhao P, Shen
Y, Dong K, Ling R, Chen S and Zhang J: XBP1s activates
METTL3/METTL14 for ER-phagy and paclitaxel sensitivity regulation
in breast cancer. Cancer Lett. 596:2168462024. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li T, Hu PS, Zuo Z, Lin JF, Li X, Wu Q,
Chen ZH, Zeng ZL, Wang F, Zheng J, et al: METTL3 facilitates tumor
progression via an m(6)A-IGF2BP2-dependent mechanism in colorectal
carcinoma. Mol Cancer. 18:1122019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang Q, Guo X, Li L, Gao Z, Su X, Ji M and
Liu J: N(6)-methyladenosine METTL3 promotes cervical cancer
tumorigenesis and Warburg effect through YTHDF1/HK2 modification.
Cell Death Dis. 11:9112020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang J, Yu H, Dong W, Zhang C, Hu M, Ma W,
Jiang X, Li H, Yang P and Xiang D: N6-Methyladenosine-Mediated
Up-Regulation of FZD10 regulates liver cancer stem cells'
properties and lenvatinib resistance through WNT/β-catenin and
hippo signaling pathways. Gastroenterology. 164:990–1005. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wei X, Huo Y, Pi J, Gao Y, Rao S, He M,
Wei Q, Song P, Chen Y, Lu D, et al: METTL3 preferentially enhances
non-m(6)A translation of epigenetic factors and promotes
tumourigenesis. NaT cell Biol. 24:1278–1290. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Xu X and Li Y, Wu Y, Wang M, Lu Y, Fang Z,
Wang H and Li Y: Increased ATF2 expression predicts poor prognosis
and inhibits sorafenib-induced ferroptosis in gastric cancer. Redox
Biol. 59:1025642023. View Article : Google Scholar
|
|
39
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
40
|
He Z, Jiao H, An Q, Zhang X, Zengyangzong
D, Xu J, Liu H, Ma L and Zhao W: Discovery of novel
4-phenylquinazoline-based BRD4 inhibitors for cardiac fibrosis.
Acta Pharm Sin B. 12:291–307. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zheng Z, Lin F, Zhao B, Chen G, Wei G,
Chen X, Nie R, Zhang R, Zhao Z, Zhou Z, et al: ALKBH5 suppresses
gastric cancer tumorigenesis and metastasis by inhibiting the
translation of uncapped WRAP53 RNA isoforms in an m6A-dependent
manner. Mol Cancer. 24:192025. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wei J, Yin Y, Zhou J, Chen H, Peng J, Yang
J and Tang Y: METTL3 potentiates resistance to cisplatin through
m(6) A modification of TFAP2C in seminoma. J Cell Mol Med.
24:11366–11380. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Thorsson V, Gibbs DL, Brown SD, Wolf D,
Bortone DS, Yang TH, Porta-Pardo E, Gao GF, Plaisier CL, Eddy JA,
et al: The immune landscape of cancer. Immunity. 48:812–830.e814.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Arribas J, Giménez E, Marcos R and
Velázquez A: Novel antiapoptotic effect of TBX15: overexpression of
TBX15 reduces apoptosis in cancer cells. Apoptosis. 20:1338–1346.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ogawa S, Kubo H, Murayama Y, Kubota T,
Yubakami M, Matsumoto T, Ohashi T, Okamoto K, Kuriki Y, Hanaoka K,
et al: Matrix metalloprotease-14 is a target enzyme for detecting
peritoneal metastasis in gastric cancer. Photodiagnosis Photodyn
Ther. 35:1024202021. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kasurinen A, Gramolelli S, Hagström J,
Laitinen A, Kokkola A, Miki Y, Lehti K, Yashiro M, Ojala PM,
Böckelman C and Haglund C: High tissue MMP14 expression predicts
worse survival in gastric cancer, particularly with a low PROX1.
Cancer Med. 8:6995–7005. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Chen WC, Chang AC, Tsai HC, Liu PI, Huang
CL, Guo JH, Liu CL, Liu JF, Thuong LHH and Tang CH: Bone
sialoprotein promotes lung cancer osteolytic bone metastasis via
MMP14-dependent mechanisms. Biochem Pharmacol. 211:1155402023.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Chen M, Qu H, Liang X, Huang Y, Yang Z, Lu
P, Shi K, Chen P, Zhang Y, Zhou H, et al: Brachyury promotes
proliferation and migration of colorectal cancer cells by targeting
MMP14. Cancer Cell Int. 25:1322025. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Siska PJ, Singer K, Evert K, Renner K and
Kreutz M: The immunological Warburg effect: Can a
metabolic-tumor-stroma score (MeTS) guide cancer immunotherapy?
Immunol Rev. 295:187–202. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
DeNardo DG, Barreto JB, Andreu P, Vasquez
L, Tawfik D, Kolhatkar N and Coussens LM: CD4(+) T cells regulate
pulmonary metastasis of mammary carcinomas by enhancing protumor
properties of macrophages. Cancer Cell. 16:91–102. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yang H, Zhang Q, Xu M, Wang L, Chen X,
Feng Y, Li Y, Zhang X, Cui W and Jia X: CCL2-CCR2 axis recruits
tumor associated macrophages to induce immune evasion through PD-1
signaling in esophageal carcinogenesis. Mol Cancer. 19:412020.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Qian J, Li J, Ma H and Ji W: Exosomal
circ-ADRM1 promotes lung adenocarcinoma progression and induces
macrophage M2 polarization through regulating MMP14 mRNA and
protein. Anticancer Drugs. 34:333–343. 2023. View Article : Google Scholar
|
|
53
|
Ager EI, Kozin SV, Kirkpatrick ND, Seano
G, Kodack DP, Askoxylakis V, Huang Y, Goel S, Snuderl M, Muzikansky
A, et al: Blockade of MMP14 activity in murine breast carcinomas:
Implications for macrophages, vessels, and radiotherapy. J Natl
Cancer Inst. 107:djv0172015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Chen Q, Yin H, Jiang Z, He T, Xie Y, Mao
W, Han J, Liu S, Lou W, Wu W, et al: Poor clinical outcomes and
immunoevasive contexture in CD161+CD8+ T
cells barren human pancreatic cancer. J Immunother Cancer.
12:e0086942024. View Article : Google Scholar
|
|
55
|
Wang J, Zheng C, Lu J, Xu X, Xiang G, Li
J, Zhang J, Mu X and Lu Q: The mechanism of MMP14-positive
tumor-associated fibroblast subsets in inhibiting PD-1
immunotherapy for esophageal cancer through exosomal tsRNA-10522.
Funct Integr Genomics. 24:1862024. View Article : Google Scholar
|
|
56
|
Wang HM, Zhang XH, Ye LQ, Zhang K, Yang
NN, Geng S, Chen J, Zhao SX, Yang KL and Fan FF: Insufficient CD100
shedding contributes to suppression of CD8(+) T-cell activity in
non-small cell lung cancer. Immunology. 160:209–219. 2020.
View Article : Google Scholar : PubMed/NCBI
|