Transforming growth factor-β1 (TGF-β1), a
pleiotropic cytokine, has a dynamic biphasic effect on cancer
development and progression. The mechanism underlying its early
tumour suppression and late tumour promotion has become a key
scientific issue in the field of tumour therapy (1). Under physiological conditions,
TGF-β1 maintains epithelial homeostasis through the suppressor of
mother against decapentaplegic (SMAD)-dependent pathway. However,
during tumour progression, this pathway is often inactivated
because of SMAD mutations, receptor expression downregulation or
negative regulator overexpression, resulting in the loss of its
tumour-suppressive function (2,3).
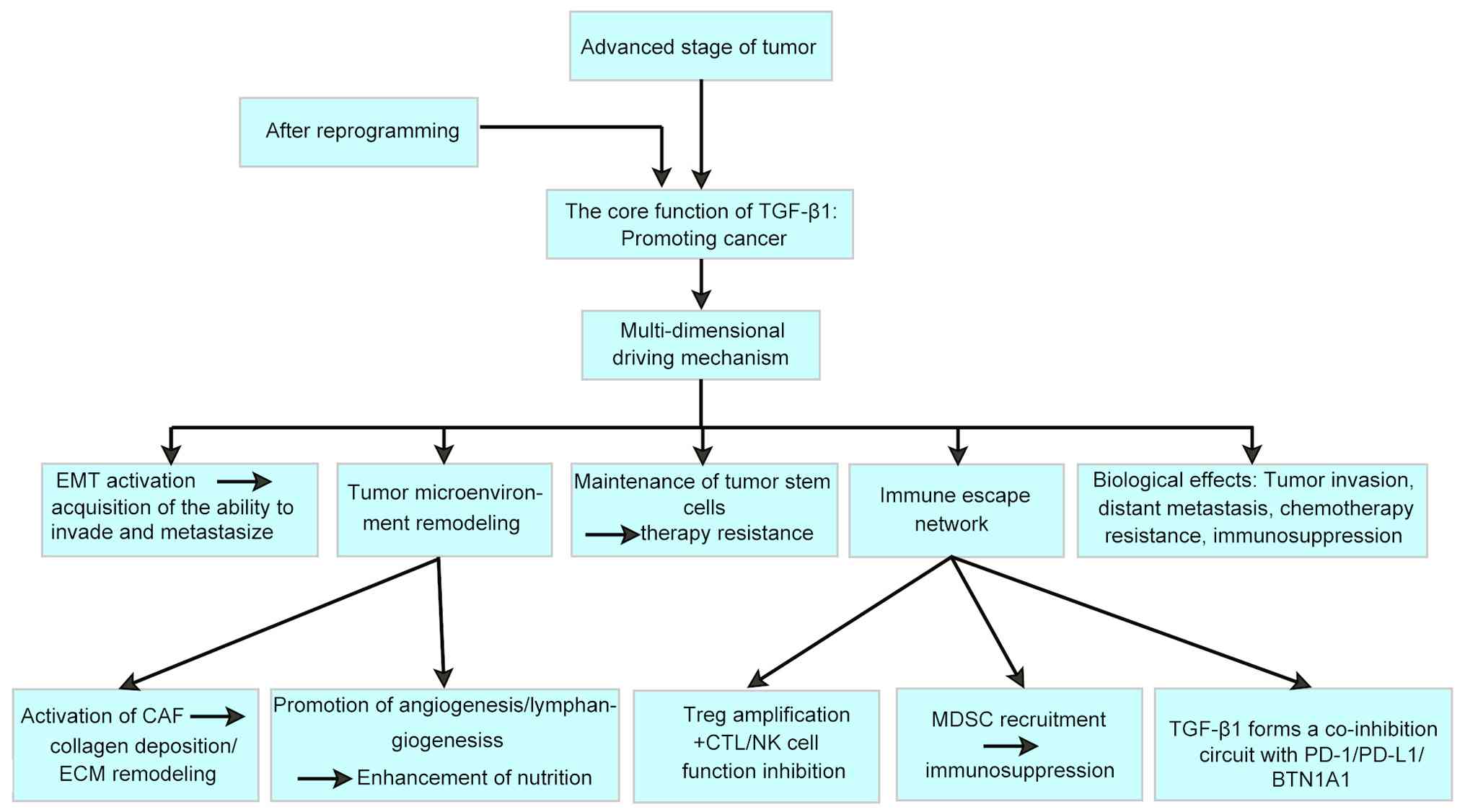
With tumour progression, TGF-β1 signalling induces tumour-promoting
reprogramming, which activates epithelial-mesenchymal transition
(EMT), non-SMAD pathways (such as phosphatidylinositol PI3K/AKT and
MAPK), and the tumour stem cell maintenance network, inhibiting the
binding ability of alkB homolog 5 to the forkhead box A1 (FOXA1)
coding sequence region and thereby driving metastatic phenotype
formation and chemotherapy resistance (2-4).
More notably, TGF-β1 signalling establishes a
'vicious cycle' that promotes metastasis by reshaping the
three-dimensional regulatory network of the tumour
microenvironment, including activating cancer-associated
fibroblasts (CAFs), promoting angiogenesis and constructing an
immunosuppressive microenvironment such as expanding T-regulatory
(Treg) cells and inhibiting cytotoxic T lymphocyte (CTL) function
(1,5,6).
Recent studies have particularly emphasised that TGF-β1 forms a
coinhibitory loop with immune checkpoints such as programmed cell
death protein 1 (PD-1)/programmed cell death ligand 1 (PD-L1),
providing a theoretical basis for combined blocking strategies
(6-8). Although TGF-β1-targeting therapeutic
strategies (such as antibodies and kinase inhibitors) face
challenges, including heterogeneous efficacy and toxicity in
clinical translation, it is anticipated that by accurately
distinguishing tumour development stages, analysing signal network
heterogeneity and developing combined immunotherapy regimens, the
current bottleneck can be overcome (5,9,10).
This review systematically summarises the multidimensional
mechanisms of action of TGF-β1 related to cancer progression,
aiming to provide a molecular basis for the development of adaptive
treatment strategies, which has important clinical significance for
improving the survival prognosis in patients with advanced
cancer.
As a pleiotropic cytokine, the physiological
functions of TGF-β1 include maintaining epithelial-mesenchymal
balance, cell cycle arrest, apoptosis induction and DNA damage
repair. In healthy tissues, TGF-β1 activates cyclin-dependent
kinase inhibitor 2B (p15INK4b, CDKN2B) and CDKN1A (p21CIP1)
expression through the SMAD2/3-dependent pathway (11), inducing G1-phase arrest and
inhibiting excessive cell proliferation. In addition, TGF-β1
maintains the epithelial phenotype and inhibits the EMT process by
regulating E-cadherin expression (12).
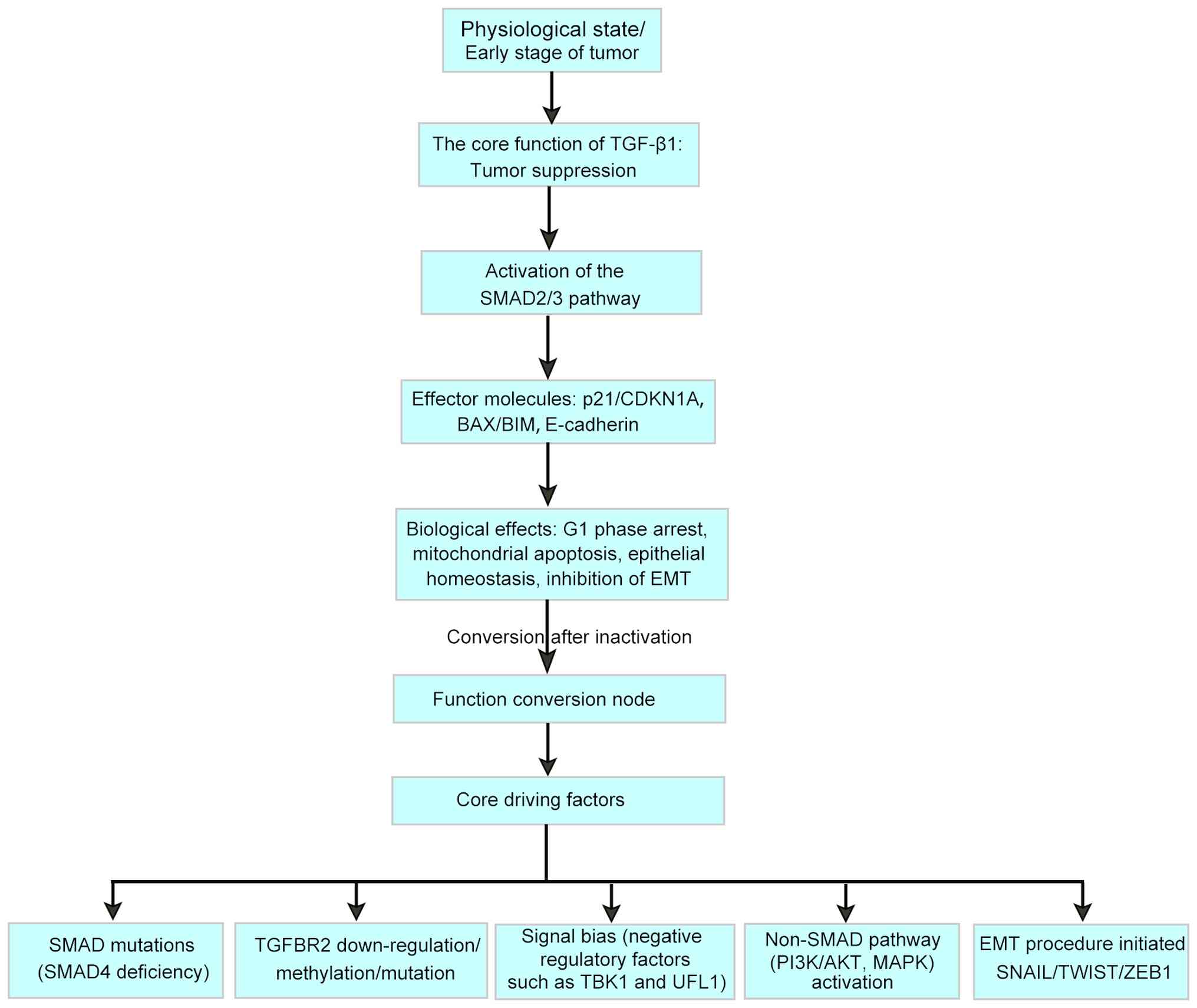
As tumours progress, inflammatory factors and gene
mutations in the tumour microenvironment can lead to a shift in
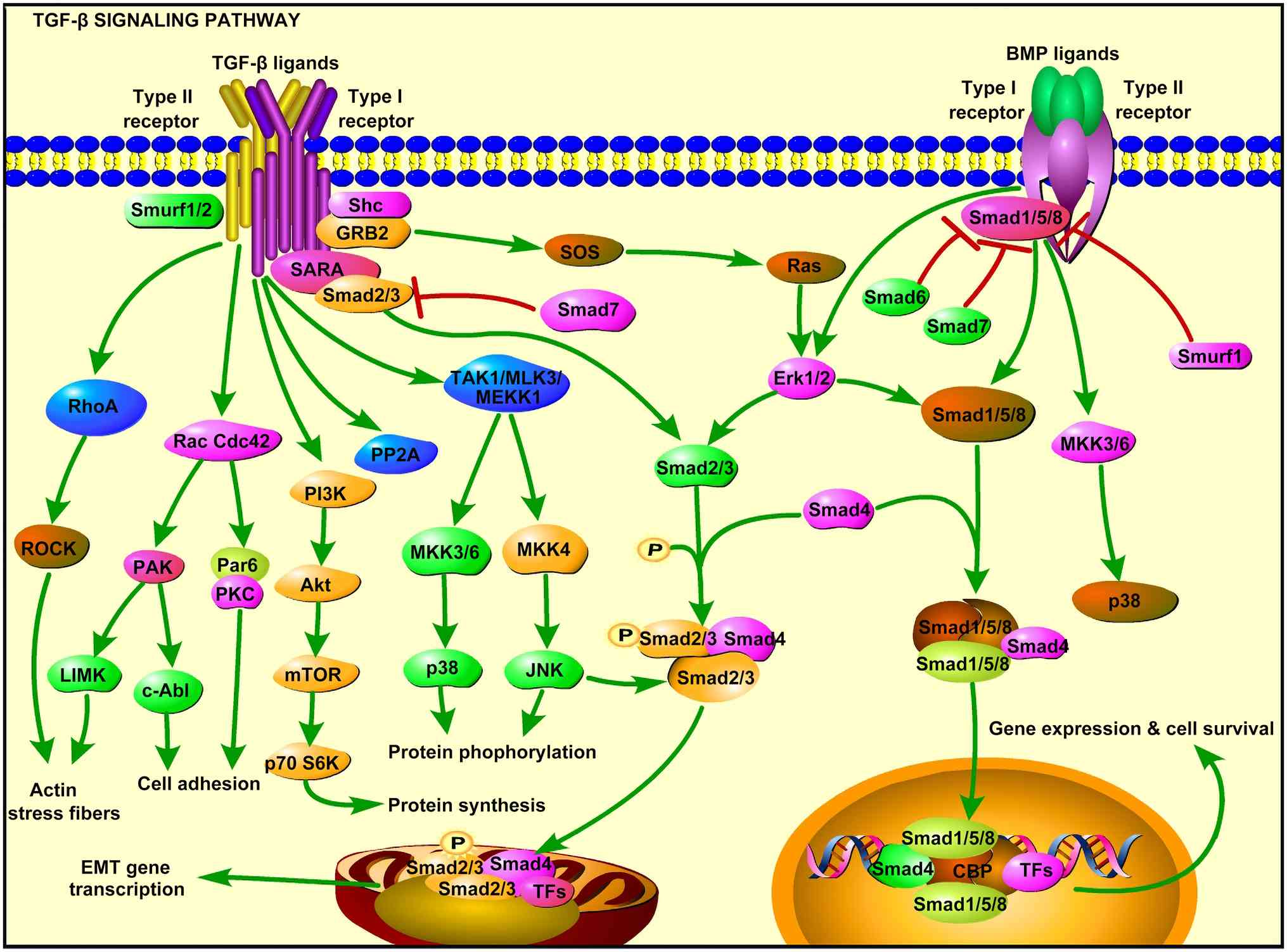
TGF-β1 signalling towards a tumour-promoting phenotype (15), as shown in Fig. 1 and Table I. In the early stages of tumour
progression, the inactivation of TGF-β1 signalling mainly occurs
through three mechanisms, including SMAD4 gene mutations or
deletions (16,17), TGF-β receptor 2 (TGFβR2) promoter
methylation (18-20) and signalling pathway bias.
These key bias nodes include: i) The Ras homolog
gene family member A/Rho-associated coiled-coil containing protein
kinase (RhoA/ROCK) pathway-it involves inhibiting cofilin through
LIM domain kinase 1 phosphorylation, promoting actin remodelling
and increasing the efficiency of pseudopodia formation by a factor
of two (21); ii) Notch
crosstalk-TGF-β1 upregulates and activates the Notch1 intracellular
domain, synergistically enhancing matrix metallopeptidase 9 (MMP9)
expression with Smad3 and increasing vascular density in breast
cancer models (22); and iii)
PD-L1 bidirectional regulation-PD-L1 can increase the transcription
and expression of TGF-β1 in tumour cells and knockdown of PD-L1
expression can significantly inhibit TGF-β1-induced EMT,
proliferation and migration (23).
With the accumulation of genomic instability and
microenvironmental alterations, tumour cells escape their
growth-inhibiting effects through acquired mutations (such as SMAD4
deletion) or epigenetic modifications (such as downregulation of
the expression of TGF-β receptor II) to promote invasion and
metastasis (24-27). The critical point of this
functional transition is usually associated with the activation of
the EMT program. TGF-β1 drives tumour cells to acquire migration
and invasion characteristics by regulating the expression of core
EMT-related transcription factors (such as Snail family
transcriptional repressor, Twist family bHLH transcription factor
and zinc finger E-box binding homeobox 1) (Table II). The EMT-promoting effect of
TGF-β1 has been confirmed in various cancer models, and its
mechanism involves FOXA1 and non-coding RNA regulation (2,28).
Changes in the expression of key EMT regulators are
significantly associated with clinical metastasis risk. In a
pancreatic cancer model, peptidyl-prolyl cis-trans isomerase
NIMA-interacting 1 (Pin1) protein promotes fibroblast growth factor
pathway activation by stabilising the conformation of TGFβR1,
leading to cytoskeleton remodelling and enhanced motility.
Knockdown of Pin1 expression weakens tumour cell migration
(29). Platelet-derived TGF-β1
induces autophagy in hepatocellular carcinoma (HCC) cells through
the AMP kinase/mammalian target of rapamycin (mTOR) axis,
increasing the number of autophagosomes in an HCC orthotopic model
and significantly increasing both invasion and metastasis rates
(30). This synergistic effect of
multiple pathways explains why inhibition of a single SMAD pathway
often fails to completely block the metastatic effect of
TGF-β1.
The metastatic mechanism of EMT involves multiple
biological processes. Studies have shown that TGF-β1 significantly
increases the proportion of tumour stem cells by regulating the
Wnt/β-catenin pathway (31). In
addition, TGF-β1 promotes collagen deposition and changes in amino
acid metabolism through prolyl 4-hydroxylase subunit α3-mediated
proline hydroxylation, increasing the cell migration speed by a
factor of two (32). TGF-β1 can
also remodel the microenvironment. TGF-β1 induces autophagy in CAFs
and decreases the microenvironment pH through upregulation of
monocarboxylate transporter 4 expression, thus promoting
angiogenesis (33).
TGF-β1 maintains tumour stem cell characteristics
through a dual mechanism. First, TGF-β1 directly activates SRY-box
transcription factor 4 (SOX4) expression, and SOX4 overexpression
in acute myeloid leukemia-resistant cell lines significantly
increases chemotherapeutic drug tolerance (34). Second, by upregulating TGF-β1
autocrine secretion through C-terminal tensin-like protein, a
positive feedback loop is formed in bladder cancer tissues, leading
to a significant increase in the proportion of paclitaxel-resistant
cells (35). A study on
pancreatic cancer has shown that deficient expression of trefoil
factor family 1 promotes EMT, significantly increasing the
proportion of CD133+ cells (a tumour stem cell marker) and
significantly decreasing the proportion of gemcitabine-sensitive
cells (36).
This reprogramming of the stem cell microenvironment
makes it difficult for traditional chemotherapy to eradicate tumour
cells, suggesting that combining TGF-β pathway inhibitors may
improve treatment efficacy. In patients with head and neck squamous
cell carcinoma, SMAD4-mutant cells respond strongly to
TGF-β1-induced stem cell marker expression (37), providing a molecular subtyping
basis for precision treatment.
CAFs are the main effector cells of TGF-β1-mediated
matrix remodelling. TGF-β1 drives CAF activation through
SMAD-dependent and SMAD-independent pathways, thereby stimulating
collagen crosslinking and fibronectin deposition to form a
metastatic, sclerotic matrix. In a gallbladder cancer model, TGF-β1
upregulates FOXA1 expression through m6A modification, increasing
the expression of α-smooth muscle actin (α-SMA) and fibronectin in
CAFs (2). In types such as colon
cancer, TGF-β1-induced CAFs can increase the collagen fibre
diameter through Janus kinase (JAK)/signal transducer and activator
of transcription 3 (STAT3) signalling, resulting in the formation
of a dense physical barrier. Treatment that neutralises TGF-β1 can
promote the recruitment of antigen-specific CD8+ T cells into
tumours (38,39). This fibrotic microenvironment
activates latent TGF-β1 through integrin αvβ6, forming a positive
feedback loop that maintains the concentration of active TGF-β1 at
a high level (40). In a liver
cancer model, the stabilising effect of TGFβ receptor-associated
protein 1 (TGFβRAP1) on TGFβR1 enhances the contractility of CAFs,
an effect that can be reversed by small interfering
(si)RNA-mediated knockout of TGFβRAP1 (3). During the transformation of ductal
carcinoma in situ to invasive carcinoma of the breast,
TGF-β1 induces the loss of cadherin in myoepithelial cells via EMT
and upregulates the expression of MMP-9 and IL-6 (41,42), significantly increasing the
disruption of basement membrane integrity.
Microenvironmental factors play important regulatory
roles in the remodelling of TGF-β1 signalling. Under hypoxic
conditions, the hypoxia-inducible factor 1α-microRNA (miR)145 axis
promotes tumour angiogenesis by inhibiting
TGFβ1-SMAD2/3-thrombospondin-1 signalling (43).
In mouse models, platelet-released TGF-β1 increases
microvascular density and enhances vascular permeability (44). TGF-β1-induced
endothelial-mesenchymal transition significantly enhances tumour
angiogenesis (45), and a breast
cancer mouse model showed increased microvascular density in the
TGF-β1-treated group, accompanied by activation of the Notch
pathway (22).
Lymphangiogenesis depends mainly on the upregulation
of vascular endothelial growth factor C (VEGF-C) expression. As
observed in a tube formation assay and a tumour xenograft mouse
model, TGF-β1 induced high expression of VEGF-C by activating the
Smad2/3 pathway, thereby significantly promoting lymphangiogenesis
in a model of gastric cancer (46). Notably, transglutaminase 1 has
been shown to participate in vascular basement membrane
remodelling, and its inhibitors can significantly reduce the
perivascular cell coverage of tumours, thereby enhancing the
penetration of chemotherapeutic drugs (47).
TGF-β1 promotes the expansion and functional
activation of Tregs through the SMAD2/3-dependent pathway. In a
multiple myeloma model, Treg-derived TGF-β1 inhibited cyclic
GMP-AMP synthase/stimulator of interferon genes signalling and
promoted major histocompatibility complex class I (MHC I) molecule
loss and PD-L1 expression upregulation (48). In a type 1 diabetes study,
TGF-β1-regulated T-cell immunoreceptor with Ig and ITIM domains+
Tregs significantly inhibited the cytotoxicity of CD226+CD8+ T
cells (49). In patients with
colorectal cancer, the expression level of TGFBR2 in
tumour-infiltrating Tregs was several times greater than that in
peripheral blood, and its inhibitory activity was positively
correlated with the concentration of TGF-β1 (50).
TGF-β1 can limit the ability of CD4+ (but not CD8+)
anti-TGF-β chimeric antigen receptor (CAR) T-cells to partially
inhibit tumour growth through the secretion of granzyme B and
interferon (IFN)-γ. CD4+ T28zT2 T cells treated with TGF-β1
survived continuously in peripheral blood and tumours, maintained
mitochondrial morphology and function, and did not induce in
vivo toxicity. These findings indicate that reprogramming the
TGF-β signalling pathway using T28zT2 (anti-TGF-β CAR T cells) in
CD4+ T cells is a promising strategy for eliminating solid tumours
(51). This bidirectional
regulation is achieved through a Smad3-dependent mechanism: TGF-β1
promotes Foxp3 expression by phosphorylating Smad3, while Smad3
knockout completely reverses the amplification effect of Tregs
(52).
MDSCs are considered major tumour promoters and
markers of cancer immune evasion mechanisms (53). TGF-β inhibits the activation and
antitumour function of T lymphocytes, dendritic cells and natural
killer (NK) cells while promoting the production of regulatory T
cells and MDSCs, thereby inhibiting the antitumour activity of T
cells (54). In patients with
renal cell carcinoma, cellular homolog of Sloan-Kettering viral
oncogene Ski (c-Ski) protein degradation is induced by TGF-β. The
downregulation of TGF-β expression remodels the immunosuppressive
tumour microenvironment by limiting tumour-infiltrating MDSCs,
while c-Ski protein degradation is also inhibited (55). The long noncoding RNA
Exo-AGAP2-AS1 from glioma cells regulates TGF-β1 expression in
MDSCs through binding to miR-486-3p; thus, Exo-AGAP2-AS1
upregulation promotes glioblastoma multiforme cell growth and
metastasis through MDSCs (56).
TGF-β1 activates the TGF-β1/2-Smad2/3 pathway by
upregulating the expression of FAT atypical cadherin 1 (FAT1),
promoting the recruitment of MDSCs to tumour foci and increasing
the proportion of MDSCs. In glioma tumours, FAT1 expression is
positively correlated with the expression of surrogate markers of
MDSCs, including PD-L1, PD-L2 and IL-10 (57), suggesting that FAT1 may play a
role in MDSC-mediated immunosuppression.
TGF-β1 exhibits dose-dependent inhibition of NK
cells. In patients with metastatic melanoma, the use of anti-TGF-β1
antibodies restored the NK cell killing efficiency to baseline
levels (58). These findings
suggest that the relationship between high levels of TGF-β1 and NK
cell inhibition in cancer patients is a major mechanism underlying
tumour immune escape. This inhibitory mechanism involves the
downregulation of the expression of the natural killer group 2D
(NKG2D) ligands MHC class I polypeptide-related sequence A/B and
PD-L1 via Smad4 by TGF-β1, thereby blocking NK cell activation
signals (59).
The inhibition of NK cell function involves multiple
mechanisms. For example, TGF-β1 induces receptor expression
downregulation. After NK cells from patients with gastric cancer
were treated with TGF-β1, the expression levels of NKp30, NKp46,
NKG2D and DNAX accessory molecule-1 were significantly reduced
(60). Second, during metabolic
reprogramming, TGF-β1 significantly reduced mitochondrial oxidative
phosphorylation efficiency through mTOR complex 1 inhibition
(61,62). In response to cytokine
dysregulation, IL-15-driven proliferation signalling is blocked by
TGF-β1, and the proportion of cells undergoing cell cycle arrest in
the G1 phase increases (63).
TGF-β1 forms a positive feedback loop with the
PD-1/PD-L1 pathway. In chronic inflammation models, TGF-β1
upregulates PD-1 expression in macrophages via synergistic
SMAD3/STAT3 signalling, and PD-1+ macrophages can secrete TGF-β1,
resulting in the formation of an autocrine loop (64). In triple-negative breast cancer
cells, TGF-βRII deficiency leads to a doubling of PD-L1 expression,
while the proportion of lymphocyte activation gene 3+ T cells
increases, resulting in resistance to immune checkpoint inhibitor
(ICI) therapy (18,65).
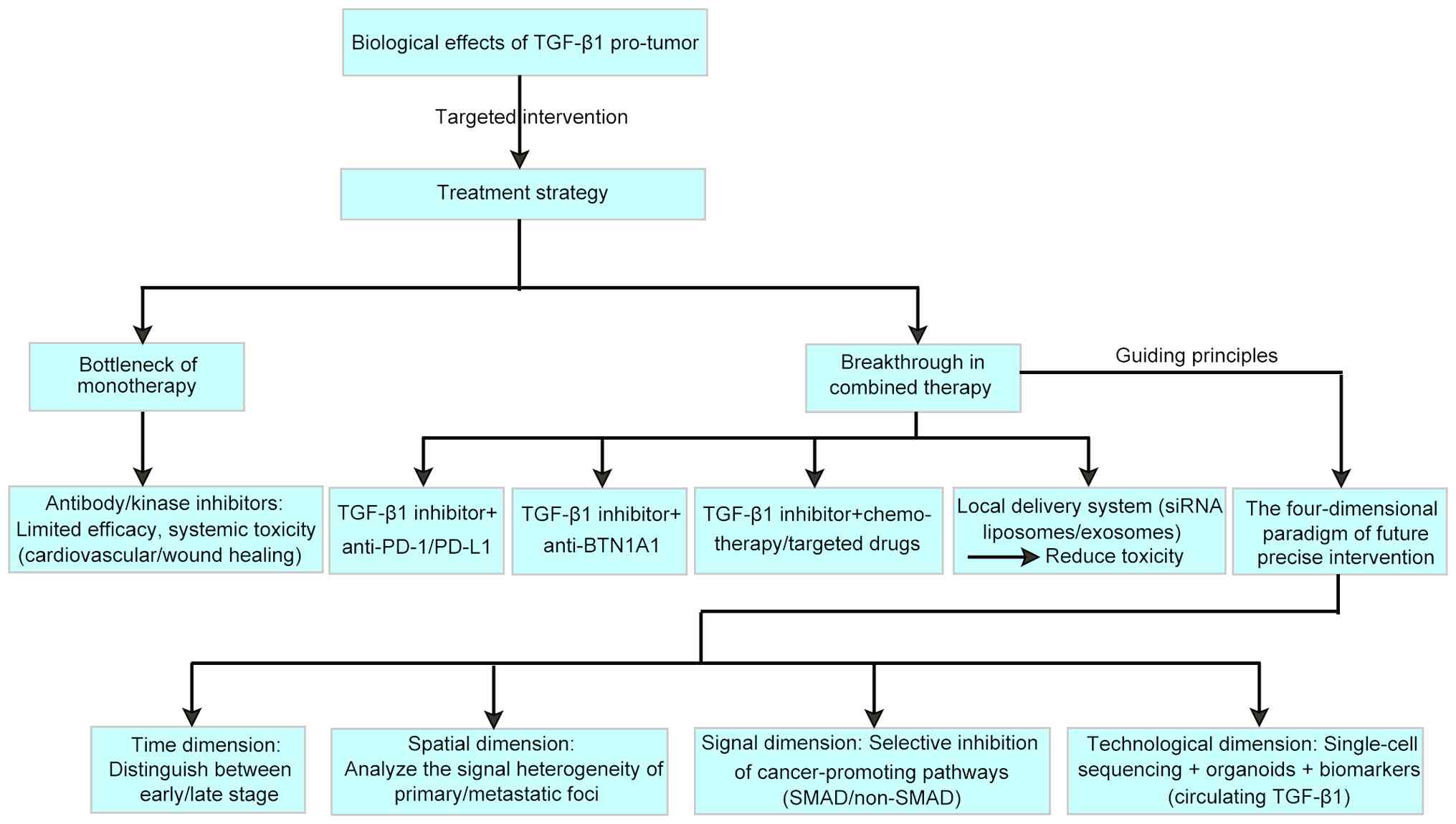
Selective TGF-β1 inhibitors, such as compound 67,
can block the EMT process and significantly reduce the number of
metastatic lesions in an A549 lung cancer model (66). Tumour-activating prodrugs
targeting TGFβR1, such as the LY2157299 precursor (galunisertib),
are designed to selectively release the active drug within the
tumour microenvironment, thereby minimising systemic exposure and
reducing off-target toxicity. LY2157299 has been shown to decrease
the incidence of cardiovascular events (67,68) by limiting systemic TGF-β pathway
inhibition, which can otherwise lead to adverse effects. This
targeted approach enhances the therapeutic window by balancing
efficacy and safety.
In combination immunotherapy, TGF-β1 antibodies and
PD-1 inhibitors can synergistically remodel the immune
microenvironment. In patients with advanced colorectal cancer, the
combination therapy group showed a significant increase in CD8+ T
cell infiltration and a significant improvement in the objective
response rate (69-71). These advances provide a precise
intervention tool for understanding the dual role of TGF-β1.
BTN1A1 is a novel immune checkpoint mutually
exclusive with PD-L1, and its inhibitory effect on T cell
activation has been demonstrated both in vitro and in
vivo (65). Increased
expression of BTN1A1, a member of the butyrophilin subfamily, has
been detected in BRAF-mutant tumours (72). Disruption of intact BTN1A1 protein
expression leads to a large accumulation of Xanthine dehydrogenase
in the cytoplasm, inducing the activation of acute-phase response
genes and phosphorylated leukemia inhibitory factor genes
downstream of STAT3 (73). Novel
strategies have shown that simultaneous blockade of TGF-β1 and
BTN1A1 can increase CD8+ T-cell infiltration in the tumour
microenvironment by a factor of two, which is superior to the
effect of monotherapy (65).
These findings provide new insight into overcoming
ICB resistance. In PD-1 resistance models, TREX1 inhibitors
combined with anti-TGF-β1 antibodies can increase tumour regression
rates (74,75).
TGF-β1, a key regulator of the tumour
microenvironment, faces significant challenges related to targeted
therapy. Although systemic inhibition of TGF-β1 can enhance
antitumour immunity and limit the expansion, survival and function
of CD4+ T cells (51) (Table IV), dose adjustments are often
required because of cardiovascular toxicity and delayed wound
healing events (58,76). A recent study showed that
perfluorooctanoic acid induces pulmonary toxicity through the
TGF-β1/Smad pathway (77).
Therefore, an early phase Ib clinical trial of the use of
TGF-β-targeted drugs such as PF-03446962 in combination with
regorafenib for the treatment of metastatic colorectal cancer was
terminated because of serious adverse events (78) (Table IV). PF-03446962 is a monoclonal
antibody targeting activin receptor like kinase 1. However, a
recent study showed that the combination of PF-03446962 and
nivolumab has promising anti-cancer activity and safety in patients
with advanced HCC (79).
The clinical response rate to TGF-β1 monotherapy is
generally low, and its limitations also include the requirement
that the kinase inhibitor galunisertib achieves a 50% inhibitory
concentration of TGF-βRI in µmol (80).
However, the local delivery system of exosome-based
TGF-β1 siRNA significantly improved the degree of fibrosis in the
lung tissues of a fibrotic mouse model. siTGF-β1 effectively
delivers specific siRNAs to the lungs, leading to TGF-β1 mRNA
silencing and EMT pathway inhibition, thereby protecting lung
tissue from fibrotic damage (81). Recent research has indicated that
immunoliposomes can deliver siTGF-β1 to tumour cells (82), thereby improving a series of
effects, such as poor stability, a short half-life and high
toxicity of naked siRNA. In an A549 tumour-bearing nude mouse
model, transfection with siTGF-β1 significantly reduced tumour
growth and tumour volume (83).
These findings suggest that targeting siTGF-β1 with immunoliposomes
may be a novel strategy for cancer treatment.
In terms of selectively blocking downstream
pathways, the TAK1 inhibitor takinib, while preserving Smad2/3
signalling, specifically inhibited the noncanonical NF-κB pathway,
increasing the tumour growth inhibition rate in a melanoma model,
but wound healing impairment was not observed. Compared with
traditional inhibitors, this precise intervention significantly
improves the therapeutic index, particularly when combined with
ICIs (84-86). Consequently, future research is
expected to overcome the historical challenges associated with
TGF-β-targeted therapy.
The TGF-β1 and PD-1 pathways exhibit bidirectional
regulation. In colorectal and liver cancer models, TGF-β1 forms a
complex at the PD-1 promoter via SMAD3/STAT3 cosignalling,
significantly upregulating PD-1 expression in macrophages. PD-1+ T
cells, in turn, increase the secretion of active TGF-β1 (64,87), creating a positive feedback loop
of immunosuppression. Combined blockade can break this cycle. A
study showed that the combined blockade of TGF-β and PD-1 activates
GBM-infiltrating CD8+ T cells, which are characterised by
upregulated TGFβRI expression (88). This finding is consistent with
previous results from a phase 1b clinical trial in patients with
relapsed/refractory multiple myeloma (89).
Mechanistic studies have shown that combination
therapy enhances antitumour immunity through a triple effect
(90,91), significantly reducing the
proportion of Foxp3+ Tregs in tumours and decreasing
immunosuppressive capacity (92,93). This combined effect can also
remodel dendritic cell (DC) function, improve the
antigen-presenting capacity of DCs and promote the expression of
IFN-γ and granzyme b on CD8+ T cells and NK cells (94). Recent studies have shown that the
intrinsic tumour cell function of PD-L1 drives immunosuppression
and tumour progression through the PD-L1/JAK/STAT3/IL-6/MDSC axis
(95) and that inhibiting the
PD-L1 and TGF-β pathways has a synergistic effect on restoring CD4+
T-cell activity in vitro (96), thereby remodelling MDSC
polarisation.
The bidirectional regulatory nature of TGF-β1
signalling necessitates that intervention strategies must consider
disease stage specificity. In early-stage tumours, TGF-β1 maintains
genomic stability by activating p16INK4a (CDKN2A) and p21 wild-type
p53-activated fragment 1/CDK-interacting protein 1 (p21WAF1/CIP1)
(99,100), and targeted inhibition at this
stage may lead to cumulative DNA damage. A study using a
preclinical model has shown that in desmoglein-2 variant-induced
arrhythmic cardiomyopathy, overactivation of the activating
transcription factor 4/TGF-β1 pathway significantly promotes
collagen deposition only in the later stages of fibrosis, whereas
early inhibition of this pathway weakens the repair capacity of
cardiomyocytes (101).
A pancreatic ductal adenocarcinoma liver metastasis
model revealed that AVID200/BMS-986416-mediated depletion of TGF-β
can reduce the formation of metastatic lesions when it is applied
in the early stages of tumour development, but if treatment is
initiated in the late stages, it accelerates CAF-mediated immune
escape (reduced CD8+ T-cell infiltration) (103).
Single-cell transcriptome analysis revealed that
TGF-β1 is highly expressed in the tumour core (CD44+ stem cell-like
cells) in primary head and neck squamous cell carcinoma lesions
(103), whereas it is enriched
in the peripheral matrix (α-SMA + CAFs) in metastatic lesions
(104). Recent advances in
radiomics, such as the use of minimum redundancy maximum relevance
and recursive feature elimination algorithms, have enabled the
non-invasive prediction of TGF-β1 spatial distribution within
tumours. For instance, a CT radiomics model achieved an area under
the curve (AUC) of 0.849 in predicting TGF-β1 expression patterns
(105), highlighting its
potential as a tool for precision oncology. This approach leverages
quantitative imaging features to map TGF-β1 heterogeneity, which
may inform targeted therapeutic strategies. In a liver metastasis
model of colorectal cancer, Smad2/3 proteins are predominantly
phosphorylated in cells from primary lesions, whereas metastatic
lesions exhibit activation of the noncanonical
TGF-β/cysteine-and-glycine-rich protein 2 axis. This difference may
lead to a significant difference in the response rate to targeted
therapy (106,107).
Spatial heterogeneity is also reflected in the
immune microenvironment. TGF-β1 reduces CD8+ T-cell response
efficiency by inhibiting IL-4 secretion from follicular helper T
cells (108). Given the
significant differences in TGF-β1 activity across different regions
within the same tumour, this spatial heterogeneity directly leads
to stratification of immunotherapy response rates (109,110). In breast cancer tissues, regions
with high TGF-β1 activity have significantly lower CD8+ T-cell
infiltration density and significantly higher PD-1 expression
levels than regions with low TGF-β1 activity (111,112). By integrating single-cell
transcriptomics and spatial transcriptomics data, researchers have
mapped the quantitative correlation between TGF-β1 signalling
gradients and the degree of immunosuppression, providing new
evidence for precise intervention (113).
Researchers have developed spatial epigenomic
techniques for quantifying histone modifications, such as histone
H3 lysine 27 trimethylation (H3K27me3), H3K4me3 and H3K27ac, on the
basis of spatial transcriptomics (114). This remarkable approach can
reveal key spatial regulatory elements controlling identity (i.e.,
spatial enhancer prediction) and advance epigenetic research into
the spatial era.
Deterministic barcoding in tissue for spatial omics
sequencing supports the simultaneous recording of spatially
barcoded mRNA and analysis of proteins of interest (a set of 22
proteins) (115). The
development of spatial multiomics has also provided integrated,
spatial transcriptomics- and antibody-based proteomics analysis
without the need for complex infrastructure (116). This platform was tested in mouse
cerebral cortex samples and a significant correlation was observed
between specific mRNA and protein expression.
Patient-derived organoids (PDOs) combined with
single-cell multiomics can enable the dynamic optimisation of
treatment regimens (117). In a
study using a colorectal cancer model, gene or drug targeting of
TGF-βR1 enhanced NK cell-mediated PDO killing or activation
(118). AVID200 is a TGF-β
ligand-trapping agent and is reported to have a binding affinity
1,000 times greater than that of other trapping agents (102,119). Although studies on the effects
of AVID200 on solid tumours are limited, in a phase I study
(NCT03834662) of 19 patients with advanced or metastatic solid
tumours, AVID200 monotherapy was well tolerated and effectively
modulated TGF-β1/3 (120,121).
Compared with whole-cell RNA sequencing, single-cell
RNA sequencing can capture intratumoural heterogeneity.
Furthermore, cellular dynamic events can be studied using
pseudotime trajectory analysis of single-cell RNA sequencing data
(122). Currently, the Samsung
Medical Center colorectal cancer single-cell RNA sequencing dataset
can be used to identify important tumour epithelial cell subtypes
(122). Through pseudotime
trajectory analysis, researchers reported that TGFB1 and IL-1B may
be effective ligands and transcription factors that regulate
pseudotime-related gene expression. By constructing a LASSO Cox
regression for 20 pseudotime genes, the 3-year survival rate of
patients with colorectal cancer can be predicted with an AUC
>0.7 (122).
Current integrated technology pathways include
diagnostic, therapeutic and monitoring stages. For example,
epithelial cell adhesion molecule-based immunoisolation can be used
to capture circulating tumour cells (CTCs) that undergo EMT after
TGF-β1-induced stimulation (123,124). Spatial transcriptomics of a
gastric cancer brain metastasis model revealed atypical vascular
strategies supporting immune signatures. Tumours with high EMT and
transcriptomic gene expression signatures consisting of catenin β1,
secreted protein acidic and cysteine rich, ventral intermediate,
SMAD3, SMAD4, TGFB1, TGFB2 and TGFB3 are more prone to blood vessel
co-option than angiogenesis (125).
The role of DNA methylation driver genes (MDGs) in
pancreatic cancer has been assessed by integrating epigenomic,
transcriptomic and clinicopathological data. High signature
risk-scores (126), a
quantitative metrics derived from mathematical or bioinformatics
models based on MDG signature, were found to be associated with
poor histological grade and advanced TNM stage. The negative
regulatory genes of the cancer-immunity cycle demonstrated that the
immunosuppressive factors TGF-β1 and CD274 (PD-L1) were positively
correlated with risk scores. These factors are negatively
correlated with CD8+ lymphocytes but positively correlated with
MDSCs (126).
TGF-β1 exhibits a dual role in cancer progression,
acting as both a tumour suppressor and promoter through
stage-dependent mechanisms. Its signalling pathways drive tumour
invasion, metastasis and therapy resistance via SMAD and non-SMAD
pathways, EMT and immune evasion. Current TGF-β1-targeted therapies
face challenges, but combination immunotherapy holds promise.
Future precision strategies must integrate temporal, spatial,
cellular and technological dimensions to selectively modulate
TGF-β1's complex functions. Advancements in single-cell sequencing
and organoid models will be critical for guiding personalised
therapeutic interventions.
Not applicable.
CH wrote and revised the manuscript, including
conception and design of the study and literature search/selection.
Data authentication is not applicable. The author has read and
approved the final manuscript.
Not applicable.
Not applicable.
The author declares that he has no competing
interests.
Not Applicable.
This work was supported by the Natural Science Foundation of
Shenzhen Municipality (grant no. JCYJ20240813115601003).
|
1
|
Chen Z, Ding C, Chen J, Zheng S and Li Q:
Pan-cancer analysis revealing the multidimensional expression and
prognostic and immunologic roles of TGFB1 in cancer. J Int Med Res.
52:30006052312213612024. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wu Z, Ke Q, Jiang L, Hong H, Pan W, Chen
W, Abudukeremu X, She F and Chen Y: TGF-β1 facilitates gallbladder
carcinoma metastasis by regulating FOXA1 translation efficiency
through m6A modification. Cell Death Dis. 15:4222024.
View Article : Google Scholar
|
|
3
|
Liu K, Tian F, Chen X, Liu B, Tian S, Hou
Y, Wang L, Han M, Peng S, Tan Y, et al: Stabilization of TGF-β
receptor 1 by a receptor-associated adaptor dictates feedback
activation of the TGF-β signaling pathway to maintain liver cancer
stemness and drug resistance. Adv Sci (Weinh). 11:e24023272024.
View Article : Google Scholar
|
|
4
|
Guan B, Liu Y, Xie B, Zhao S, Yalikun A,
Chen W, Zhou M, Gu Q and Yan D: Mitochondrial genome transfer
drives metabolic reprogramming in adjacent colonic epithelial cells
promoting TGFβ1-mediated tumor progression. Nat Commun.
15:36532024. View Article : Google Scholar
|
|
5
|
Tran TM, Chand Thakuri BK, Nurmukhambetova
S, Lee JJ, Hu P, Tran NQ, Steimle B, Dash P and Schneider D:
Armored TGFβRIIDN ROR1-CAR T cells reject solid tumors and resist
suppression by constitutively-expressed and treatment-induced
TGFβ1. J Immunother Cancer. 12:e0082612024. View Article : Google Scholar
|
|
6
|
Robbrecht D, Grob JJ, Bechter O, Simonelli
M, Doger B, Borbath I, Butler MO, Cheng T, Romano PM,
Pons-Tostivint E, et al: Biomarker and pharmacodynamic activity of
the transforming growth factor-beta (TGFβ) inhibitor SAR439459 as
monotherapy and in combination with cemiplimab in a phase I
clinical study in patients with advanced solid tumors. Clin Transl
Sci. 17:e137362024. View Article : Google Scholar
|
|
7
|
Barboy O, Bercovich A, Li H, Eyal-Lubling
Y, Yalin A, Shapir Itai Y, Abadie K, Zada M, David E,
Shlomi-Loubaton S, et al: Modeling T cell temporal response to
cancer immunotherapy rationalizes development of combinatorial
treatment protocols. Nat Cancer. 5:742–759. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu J, Wu M, Yang Y, Wang Z, He S, Tian X
and Wang H: γδ T cells and the PD-1/PD-L1 axis: A love-hate
relationship in the tumor microenvironment. J Transl Med.
22:5532024. View Article : Google Scholar
|
|
9
|
Longhi E, Carminati L, Carlessi E, Belotti
D and Taraboletti G: Thrombospondin-1 in drug activity and tumor
response to therapies. Semin Cell Dev Biol. 155:45–51. 2024.
View Article : Google Scholar
|
|
10
|
Guo Y, Wang Z, Zhou H, Pan H, Han W, Deng
Y, Li Q, Xue J, Ge X, Wang S, et al: First-in-human study of
GFH018, a small molecule inhibitor of transforming growth factor-β
receptor I inhibitor, in patients with advanced solid tumors. BMC
Cancer. 24:4442024. View Article : Google Scholar
|
|
11
|
Deb VK, Chauhan N and Jain U: Deciphering
TGF-β1's role in drug resistance and leveraging plant bioactives
for cancer therapy. Eur J Pharmacol. 988:1772182025. View Article : Google Scholar
|
|
12
|
Liu W, Lu D, Jia S, Yang Y, Meng F, Du Y,
Yang Y, Yuan L and Nan Y: Molecular mechanism of Gancao Xiexin
Decoction regulating EMT and suppressing hepatic metastasis of
gastric cancer via the TGF-β1/SMAD pathway. J Ethnopharmacol.
342:1194302025. View Article : Google Scholar
|
|
13
|
Wang Y, Lu H, Wang Z, Li Y and Chen X:
TGF-β1 promotes autophagy and inhibits apoptosis in breast cancer
by targeting TP63. Front Oncol. 12:8650672022. View Article : Google Scholar
|
|
14
|
Hinz B: More Velcro for the TGF-β1
straitjacket: A new antibody straps latent TGF-β1 to the matrix.
Sci Signal. 17:eado52792024. View Article : Google Scholar
|
|
15
|
Aashaq S, Batool A, Mir SA, Beigh MA,
Andrabi KI and Shah ZA: TGF-β signaling: A recap of
SMAD-independent and SMAD-dependent pathways. J Cell Physiol.
237:59–85. 2022. View Article : Google Scholar
|
|
16
|
Rossato VV, Silveira DA, Gupta S and
Mombach JCM: Towards the contribution of the p38MAPK pathway to the
dual role of TGFβ in cancer: A boolean model approach. Comput Biol
Med. 104:235–240. 2019. View Article : Google Scholar
|
|
17
|
Lin R, Li X, Wu S, Qian S, Hou H, Dong M,
Zhang X and Zhang M: Suppression of latent transforming growth
factor-β (TGF-β)-binding protein 1 (LTBP1) inhibits natural
killer/T cell lymphoma progression by inactivating the TGF-β/Smad
and p38MAPK pathways. Exp Cell Res. 407:1127902021.
View Article : Google Scholar
|
|
18
|
Zahraeifard S, Xiao Z, So JY, Ahad A,
Montoya S, Park WY, Sornapudi T, Andohkow T, Read A, Kedei N, et
al: Loss of tumor suppressors promotes inflammatory tumor
microenvironment and enhances LAG3+T cell mediated immune
suppression. Nat Commun. 15:58732024. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Miar P, Tabatabaiefar MA, Abdollahi Z,
Noruzi M, Kazemi M, Naimi A, Emami MH, Izadi S and Zeinalian M:
BAT25, ACVR2, and TGFBR2 mononucleotide STR markers: A triplex
panel for microsatellite instability testing in colorectal tumors.
Adv Biomed Res. 11:792022. View Article : Google Scholar :
|
|
20
|
Fricke F, Michalak M, Warnken U, Hausser
I, Schnölzer M, Kopitz J and Gebert J: SILAC-based quantification
of TGFBR2-regulated protein expression in extracellular vesicles of
microsatellite unstable colorectal cancers. Int J Mol Sci.
20:41622019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang H, Liu H, Cheng H, Xue X, Ge Y, Wang
X and Yuan J: Klotho stabilizes the podocyte actin cytoskeleton in
idiopathic membranous nephropathy through regulating the TRPC6/CatL
pathway. Am J Nephrol. 55:345–360. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li ZX, Chen JX, Zheng ZJ, Cai WJ, Yang XB,
Huang YY, Gong Y, Xu F, Chen YS and Lin L: TGF-β1 promotes human
breast cancer angiogenesis and malignant behavior by regulating
endothelial-mesenchymal transition. Front Oncol. 12:10511482022.
View Article : Google Scholar
|
|
23
|
Li Z, Wang F, Dang J, Cheng F and Zheng F:
Bidirectional regulation between tumor cell-intrinsic PD-L1 and
TGF-β1 in epithelial-to-mesenchymal transition in melanoma. Transl
Cancer Res. 11:3698–3710. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang J, Xiang H, Lu Y and Wu T: Role and
clinical significance of TGF-β1 and TGF-βR1 in malignant tumors
(Review). Int J Mol Med. 47:552021. View Article : Google Scholar
|
|
25
|
Lloyd EG, Jihad M, Manansala JS, Li W,
Cheng PSW, Mucciolo G, Zaccaria M, Pinto Teles S, Araos Henríquez
J, Harish S, et al: SMAD4 and KRAS status shapes cancer
cell-stromal cross-talk and therapeutic response in pancreatic
cancer. Cancer Res. 85:1368–1389. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sun M, Gu Y, Wang J, Zhang Z, Ling Z, Shao
F, Lin C, He H, Li R, Liu H and Xu J: Smad4 loss identifies
aggressive subtype with immunotherapy and anti-HER-2 treatment
resistance in gastric cancer. Br J Cancer. 132:1148–1157. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rizwan M, Mahjabeen I, Haris MS, Qayyum F
and Kayani MA: Deregulation of exosomal miR-17, miR-20a and TGFBR2
in head and neck cancer patients. Technol Cancer Res Treat.
24:153303382513233142025. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang X, Xu C, Liu C, Wu X, Chen X, Hou J
and Wang L: TGF-β1-Induced LINC01094 promotes
epithelial-mesenchymal transition in hepatocellular carcinoma
through the miR-122-5p/TGFBR2-SAMD2-SMAD3 axis. Funct Integr
Genomics. 24:1232024. View Article : Google Scholar
|
|
29
|
Bowman CL, Daniel CJ, Carlson EJ, Shah VM,
Farrell AS, Kresse KM, Wang X, Lindley KA, Kuhn MR,
MacPherson-Hawthorne K, et al: The prolyl isomerase PIN1 affects
fibroblast differentiation states and cross-talk in pancreatic
cancer. Cancer Res. 85:4899–4917. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lu M, Gong X, Zhang YM, Guo YW, Zhu Y,
Zeng XB, Gao JH, Liu LM, Shu D, Ma R, et al: Platelets promote
primary hepatocellular carcinoma metastasis through TGF-β1-mediated
cancer cell autophagy. Cancer Lett. 600:2171612024. View Article : Google Scholar
|
|
31
|
He H, Li Y, Wang Y and Li M: Geniposide
inhibits non-small cell lung cancer by regulating proliferation,
apoptosis, invasion, migration, epithelial-mesenchymal transition,
and cancer stem-like cell property Via Wnt/β-catenin pathway.
Biochem Genet. 64:707–726. 2026. View Article : Google Scholar
|
|
32
|
Nakasuka F, Tabata S, Sakamoto T, Hirayama
A, Ebi H, Yamada T, Umetsu K, Ohishi M, Ueno A, Goto H, et al:
TGF-β-dependent reprogramming of amino acid metabolism induces
epithelial-mesenchymal transition in non-small cell lung cancers.
Commun Biol. 4:7822021. View Article : Google Scholar
|
|
33
|
Jena BC, Das CK, Banerjee I, Bharadwaj D,
Majumder R, Das S, Biswas A, Kundu M, Roy PK, Kundu CN and Mandal
M: TGF-β1 induced autophagy in cancer associated fibroblasts during
hypoxia contributes EMT and glycolysis via MCT4 upregulation. Exp
Cell Res. 417:1131952022. View Article : Google Scholar
|
|
34
|
Liang X, Zhou J, Li C, Wang H, Wan Y, Ling
C, Pu L, Zhang W, Fan M, Hong J and Zhai Z: The roles and
mechanisms of TGFB1 in acute myeloid leukemia chemoresistance. Cell
Signal. 116:1110272024. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zou F, Zhang G, Mei G, Zhang H, Xie M and
Dan M: CTEN-induced TGF-β1 expression facilitates EMT and enhances
paclitaxel resistance in bladder cancer cells. Am J Transl Res.
16:3248–3258. 2024. View Article : Google Scholar
|
|
36
|
Yamaguchi J, Kokuryo T, Yokoyama Y, Oishi
S, Sunagawa M, Mizuno T, Onoe S, Watanabe N, Ogura A and Ebata T:
Trefoil factor 1 suppresses stemness and enhances chemosensitivity
of pancreatic cancer. Cancer Med. 13:e73952024. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bette M, Reinhardt L, Gansukh U,
Xiang-Tischhauser L, Meskeh H, Di Fazio P, Buchholz M, Stuck BA and
Mandic R: The role of TGF-β1 and mutant SMAD4 on
epithelial-mesenchymal transition features in head and neck
squamous cell carcinoma cell lines. Cancers (Basel). 16:31722024.
View Article : Google Scholar
|
|
38
|
Vienot A, Pallandre JR, Renaude E, Viot J,
Bouard A, Spehner L, Kroemer M, Abdeljaoued S, van der Woning B, de
Haard H, et al: Chemokine switch regulated by TGF-β1 in
cancer-associated fibroblast subsets determines the efficacy of
chemo-immunotherapy. Oncoimmunology. 11:21446692022. View Article : Google Scholar
|
|
39
|
Tan HX, Cao ZB, He TT, Huang T, Xiang CL
and Liu Y: TGFβ1 is essential for MSCs-CAFs differentiation and
promotes HCT116 cells migration and invasion via JAK/STAT3
signaling. Onco Targets Ther. 12:5323–5334. 2019. View Article : Google Scholar
|
|
40
|
Brown NF and Marshall JF:
Integrin-mediated TGFβ activation modulates the tumour
microenvironment. Cancers (Basel). 11:12212019. View Article : Google Scholar
|
|
41
|
Wang L, Xu C, Liu X, Yang Y, Cao L, Xiang
G, Liu F, Wang S, Liu J, Meng Q, et al: TGF-β1 stimulates
epithelial-mesenchymal transition and cancer-associated
myoepithelial cell during the progression from in situ to invasive
breast cancer. Cancer Cell Int. 19:3432019. View Article : Google Scholar
|
|
42
|
Effat H, El Houseini ME and Abohashem RS:
The combined impact of curcumin: Piperine and sorafenib on
microRNAs and different pathways in breast cancer cells. Indian J
Clin Biochem. 40:32–45. 2025. View Article : Google Scholar
|
|
43
|
Luo YW, Fang Y, Zeng HX, Ji YC, Wu MZ, Li
H, Chen JY, Zheng LM, Fang JH and Zhuang SM: HIF1α counteracts
TGFβ1-driven TSP1 expression in endothelial cells to stimulate
angiogenesis in the hypoxic tumor microenvironment. Cancer Res.
85:69–83. 2025. View Article : Google Scholar
|
|
44
|
Wang L, Wang H, Zhu M, Ni X, Sun L, Wang
W, Xie J, Li Y, Xu Y, Wang R, et al: Platelet-derived TGF-β1
induces functional reprogramming of myeloid-derived suppressor
cells in immune thrombocytopenia. Blood. 144:99–112. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Jiao Q, Xu X, Xu L, Wang Y, Pang S, Hao J,
Liu X, Zhao Y, Qi W, Qin L, et al: Knockdown of eIF3a alleviates
pulmonary arterial hypertension by inhibiting
endothelial-to-mesenchymal transition via TGFβ1/SMAD pathway. J
Transl Med. 23:5242025. View Article : Google Scholar
|
|
46
|
Pak KH, Park KC and Cheong JH: VEGF-C
induced by TGF-β1 signaling in gastric cancer enhances
tumor-induced lymphangiogenesis. BMC Cancer. 19:7992019. View Article : Google Scholar
|
|
47
|
Ebrahimi Samani S, Tatsukawa H, Hitomi K
and Kaartinen MT: Transglutaminase 1: Emerging functions beyond
skin. Int J Mol Sci. 25:103062024. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhang D, Zhan D, Zhang R, Sun Y, Duan C,
Yang J, Wei J, Li X, Lu Y and Lai X: Treg-derived TGF-β1 dampens
cGAS-STING signaling to downregulate the expression of class I MHC
complex in multiple myeloma. Sci Rep. 14:115932024. View Article : Google Scholar
|
|
49
|
Zhong T and Li X, Lei K, Tang R, Deng Q,
Love PE, Zhou Z, Zhao B and Li X: TGF-β-mediated crosstalk between
TIGIT+ Tregs and CD226+CD8+ T
cells in the progression and remission of type 1 diabetes. Nat
Commun. 15:88942024. View Article : Google Scholar
|
|
50
|
Wang J, Wang J, Gu Q, Yang Y, Ma Y and
Zhang Q: TGFβ1: An indicator for tumor immune microenvironment of
colon cancer from a comprehensive analysis of TCGA. Front Genet.
12:6120112021. View Article : Google Scholar
|
|
51
|
Zheng D, Qin L, Lv J, Che M, He B, Zheng
Y, Lin S, Qi Y, Li M, Tang Z, et al: CD4+ anti-TGF-β CAR
T cells and CD8+ conventional CAR T cells exhibit
synergistic antitumor effects. Cell Rep Med. 6:1020202025.
View Article : Google Scholar
|
|
52
|
Lodyga M and Hinz B: TGF-β1-A truly
transforming growth factor in fibrosis and immunity. Semin Cell Dev
Biol. 101:123–139. 2020. View Article : Google Scholar
|
|
53
|
Mojsilovic S, Mojsilovic SS, Bjelica S and
Santibanez JF: Transforming growth factor-beta1 and myeloid-derived
suppressor cells: A cancerous partnership. Dev Dyn. 251:105–124.
2022. View Article : Google Scholar
|
|
54
|
Villar VH, Subotički T, Đikić D,
Mitrović-Ajtić O, Simon F and Santibanez JF: Transforming growth
factor-β1 in cancer immunology: Opportunities for immunotherapy.
Adv Exp Med Biol. 1408:309–328. 2023. View Article : Google Scholar
|
|
55
|
Wang G, Zhou X, Guo Z, Huang N, Li J, Lv
Y, Han L, Zheng W, Xu D, Chai D, et al: The anti-fibrosis drug
Pirfenidone modifies the immunosuppressive tumor microenvironment
and prevents the progression of renal cell carcinoma by inhibiting
tumor autocrine TGF-β. Cancer Biol Ther. 23:150–162. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Tian Y, Gao X, Yang X, Chen S and Ren Y:
Glioma-derived exosome Lncrna Agap2-As1 promotes glioma
proliferation and metastasis by mediating Tgf-β1 secretion of
myeloid-derived suppressor cells. Heliyon. 10:e299492024.
View Article : Google Scholar
|
|
57
|
Irshad K, Srivastava C, Malik N, Arora M,
Gupta Y, Goswami S, Sarkar C, Suri V, Mahajan S, Gupta DK, et al:
Upregulation of atypical cadherin FAT1 promotes an
immunosuppressive tumor microenvironment via TGF-β. Front Immunol.
13:8138882022. View Article : Google Scholar
|
|
58
|
Mirjačić Martinović K, Vuletić A, Mališić
E, Srdić-Rajić T, Tišma Miletić N, Babović N and Jurišić V:
Increased circulating TGF-β1 is associated with impairment in NK
cell effector functions in metastatic melanoma patients. Growth
Factors. 40:231–239. 2022. View Article : Google Scholar
|
|
59
|
Leonteva A, Abdurakhmanova M, Bogachek M,
Belovezhets T, Yurina A, Troitskaya O, Kulemzin S, Richter V,
Kuligina E and Nushtaeva A: The activity of human NK cells towards
3D heterotypic cellular tumor model of breast cancer. Cells.
14:10392025. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Han B, Mao FY, Zhao YL, Lv YP, Teng YS,
Duan M, Chen W, Cheng P, Wang TT, Liang ZY, et al: Altered NKp30,
NKp46, NKG2D, and DNAM-1 expression on circulating NK cells is
associated with tumor progression in human gastric cancer. J
Immunol Res. 2018:62485902018. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Das R, Xu S, Nguyen TT, Quan X, Choi SK,
Kim SJ, Lee EY, Cha SK and Park KS: Transforming growth factor
β1-induced apoptosis in podocytes via the extracellular
signal-regulated kinase-mammalian target of rapamycin complex
1-NADPH oxidase 4 axis. J Biol Chem. 290:30830–30842. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wang D, Sun Z, Zhu X, Zheng X, Zhou Y, Lu
Y, Yan P, Wang H, Liu H, Jin J, et al: GARP-mediated active TGF-β1
induces bone marrow NK cell dysfunction in AML patients with early
relapse post-allo-HSCT. Blood. 140:2788–2804. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Zhang S, Huang H, Handley M, Griffin N,
Bai X and Shan F: A novel mechanism of lung cancer inhibition by
methionine enkephalin through remodeling the immune status of the
tumor microenvironment. Int Immunopharmacol. 99:1079992021.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Lei Z, Tang R, Wu Y, Mao C, Xue W, Shen J,
Yu J, Wang X, Qi X, Wei C, et al: TGF-β1 induces PD-1 expression in
macrophages through SMAD3/STAT3 cooperative signaling in chronic
inflammation. JCI Insight. 9:e1655442024. View Article : Google Scholar
|
|
65
|
Kim YS, Lee SH, Park AH, Wu C, Hong BK,
Jung H, Lin SH and Yoo SS: BTN1A1 is a novel immune checkpoint
mutually exclusive to PD-L1. J Immunother Cancer. 12:e0083032024.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Jeong JH, Jang HJ, Kwak S, Sung GJ, Park
SH, Song JH, Kim H, Na Y and Choi KC: Novel TGF-β1 inhibitor
antagonizes TGF-β1-induced epithelial-mesenchymal transition in
human A549 lung cancer cells. J Cell Biochem. 120:977–987. 2019.
View Article : Google Scholar
|
|
67
|
Zhang Y, Parrish KE, Tortolani DR, Poss
MA, Huang A, Wan H, Purandare AV, Donnell AF, Kempson J, Hou X, et
al: Long-acting tumor-activated prodrug of a TGFβR inhibitor. J Med
Chem. 64:15787–15798. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Liu T, Sun S, Huang Y, E Y, Li W, Xu F,
Liu Z, Luo X, Lu C and Yu C: The integration of single-cell and
metabolomics reveals the increase of oxidative phosphorylation
during the liver metastasis of colorectal cancer. Cancer Metab.
13:412025. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Mortezaee K and Majidpoor J: Transforming
growth factor-β signalling in tumour resistance to the anti-PD-(L)1
therapy: Updated. J Cell Mol Med. 27:311–321. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Lim YW, Coles GL, Sandhu SK, Johnson DS,
Adler AS and Stone EL: Single-cell transcriptomics reveals the
effect of PD-L1/TGF-β blockade on the tumor microenvironment. BMC
Biol. 19:1072021. View Article : Google Scholar
|
|
71
|
Yi M, Li T, Niu M, Wu Y, Zhao Z and Wu K:
TGF-β: A novel predictor and target for anti-PD-1/PD-L1 therapy.
Front Immunol. 13:10613942022. View Article : Google Scholar
|
|
72
|
Lund-Andersen C, Torgunrud A, Kanduri C,
Dagenborg VJ, Frøysnes IS, Larsen MM, Davidson B, Larsen SG and
Flatmark K: Novel drug resistance mechanisms and drug targets in
BRAF-mutated peritoneal metastasis from colorectal cancer. J Transl
Med. 22:6462024. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Jeong J, Kadegowda AKG, Meyer TJ, Jenkins
LM, Dinan JC, Wysolmerski JJ, Weigert R and Mather IH: The
butyrophilin 1a1 knockout mouse revisited: Ablation of Btn1a1 leads
to concurrent cell death and renewal in the mammary epithelium
during lactation. FASEB Bioadv. 3:971–997. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Toufektchan E, Dananberg A, Striepen J,
Hickling JH, Shim A, Chen Y, Nichols A, Duran Paez MA, Mohr L,
Bakhoum SF and Maciejowski J: Intratumoral TREX1 induction promotes
immune evasion by limiting type I IFN. Cancer Immunol Res.
12:673–686. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Hanks BA: Unlocking the therapeutic
potential of the cGAS-STING pathway through TREX1 targeting. Cancer
Res. 85:2778–2780. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
de Streel G and Lucas S: Targeting
immunosuppression by TGF-β1 for cancer immunotherapy. Biochem
Pharmacol. 192:1146972021. View Article : Google Scholar
|
|
77
|
Elsheikh AA, Shalaby AM, Alabiad MA,
Abd-Almotaleb NA and Khayal EES: Perfluorooctanoic acid induced
lung toxicity via TGF-β1/Smad pathway, crosstalk between airway
hyperresponsiveness and fibrosis: Withdrawal impact. Environ Sci
Pollut Res Int. 32:4989–5007. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Clarke JM, Blobe GC, Strickler JH, Uronis
HE, Zafar SY, Morse M, Dropkin E, Howard L, O'Neill M, Rushing CN,
et al: A phase Ib study of the combination regorafenib with
PF-03446962 in patients with refractory metastatic colorectal
cancer (REGAL-1 trial). Cancer Chemother Pharmacol. 84:909–917.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Hsu C, Chang YF, Yen CJ, Xu YW, Dong M and
Tong YZ: Combination of GT90001 and nivolumab in patients with
advanced hepatocellular carcinoma: A multicenter, single-arm, phase
1b/2 study. BMC Med. 21:3952023. View Article : Google Scholar :
|
|
80
|
Maher JM, Zhang R, Palanisamy G, Perkins
K, Liu L, Brassil P, McNamara A, Lo A, Hughes AD, Kanodia J, et al:
Lung-restricted ALK5 inhibition avoids systemic toxicities
associated with TGFβ pathway inhibition. Toxicol Appl Pharmacol.
438:1159052022. View Article : Google Scholar
|
|
81
|
Qiu C, Zhao Z, Xu C, Yuan R, Ha Y, Tu Q,
Zhang H, Mu Z, Xin Q, Tian Y, et al: Nebulized milk exosomes loaded
with siTGF-β1 ameliorate pulmonary fibrosis by inhibiting EMT
pathway and enhancing collagen permeability. J Nanobiotechnology.
22:4342024. View Article : Google Scholar
|
|
82
|
Wang X, Huang H, Xu W, Gong Y, Shi S, Wan
X and Li P: TGF-β1 and FOXM1 siRNA co-loaded nanoparticles by
disulfide crosslinked PEG-PDMAEMA for the treatment of
triple-negative breast cancer and its bone metastases in vitro.
Drug Dev Ind Pharm. 50:777–788. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Shi Y, Zhang H, Chen H, Guo J, Yuan R,
Tian Y, Xin Q, Mu Z, Tao Y, Chu Y, et al: Cetuximab-immunoliposomes
loaded with TGF-β1 siRNA for the targeting therapy of NSCLC:
Design, and in vitro and in vivo evaluation. Int J Mol Sci.
26:11962025. View Article : Google Scholar
|
|
84
|
Kim BG, Malek E, Choi SH, Ignatz-Hoover JJ
and Driscoll JJ: Novel therapies emerging in oncology to target the
TGF-β pathway. J Hematol Oncol. 14:552021. View Article : Google Scholar
|
|
85
|
Totzke J, Scarneo SA, Yang KW and Haystead
TAJ: TAK1: A potent tumour necrosis factor inhibitor for the
treatment of inflammatory diseases. Open Biol. 10:2000992020.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Canè S, Van Snick J, Uyttenhove C, Pilotte
L and Van den Eynde BJ: TGFβ1 neutralization displays therapeutic
efficacy through both an immunomodulatory and a non-immune
tumor-intrinsic mechanism. J Immunother Cancer. 9:e0017982021.
View Article : Google Scholar
|
|
87
|
Zhang B, Wang CM, Wu HX, Wang F, Chai YY,
Hu Y, Wang BJ, Yu Z, Xia RH, Xu RH and Cao XT: MFSD2A potentiates
gastric cancer response to anti-PD-1 immunotherapy by reprogramming
the tumor microenvironment to activate T cell response. Cancer
Commun (Lond). 43:1097–1116. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Kim AR, Jeon SH, Park J, Kim ES, Kwon M,
Yoo J, Kang SG, Park SH, Chang JH and Shin EC: Co-blockade of TGFβ
and PD-1 reinvigorates glioblastoma-infiltrating CD8+ T cells that
characteristically upregulate TGFβRI expression. Clin Cancer Res.
31:3306–3316. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Malek E, Rana PS, Swamydas M, Daunov M,
Miyagi M, Murphy E, Ignatz-Hoover JJ, Metheny L, Kim SJ and
Driscoll JJ: The TGFβ type I receptor kinase inhibitor vactosertib
in combination with pomalidomide in relapsed/refractory multiple
myeloma: A phase 1b trial. Nat Commun. 15:73882024. View Article : Google Scholar
|
|
90
|
Niu M, Yi M, Wu Y, Lyu L, He Q, Yang R,
Zeng L, Shi J, Zhang J, Zhou P, et al: Synergistic efficacy of
simultaneous anti-TGF-β/VEGF bispecific antibody and PD-1 blockade
in cancer therapy. J Hematol Oncol. 16:942023. View Article : Google Scholar
|
|
91
|
Yi M, Wu Y, Niu M, Zhu S, Zhang J, Yan Y,
Zhou P, Dai Z and Wu K: Anti-TGF-β/PD-L1 bispecific antibody
promotes T cell infiltration and exhibits enhanced antitumor
activity in triple-negative breast cancer. J Immunother Cancer.
10:e0055432022. View Article : Google Scholar
|
|
92
|
Zong Y, Deng K and Chong WP: Regulation of
Treg cells by cytokine signaling and co-stimulatory molecules.
Front Immunol. 15:13879752024. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Kujan O, Agag M, Smaga M, Vaishnaw Y,
Idrees M, Shearston K and Farah CS: PD-1/PD-L1, Treg-related
proteins, and tumour-infiltrating lymphocytes are associated with
the development of oral squamous cell carcinoma. Pathology.
54:409–416. 2022. View Article : Google Scholar
|
|
94
|
Lucarini V, Melaiu O, D'Amico S, Pastorino
F, Tempora P, Scarsella M, Pezzullo M, De Ninno A, D'Oria V, Cilli
M, et al: Combined mitoxantrone and anti-TGFβ treatment with PD-1
blockade enhances antitumor immunity by remodelling the tumor
immune landscape in neuroblastoma. J Exp Clin Cancer Res.
41:3262022. View Article : Google Scholar
|
|
95
|
Jeong H, Koh J, Kim S, Yim J, Song SG, Kim
H, Li Y, Lee SH, Chung YK, Kim H, et al: Cell-intrinsic PD-L1
signaling drives immunosuppression by myeloid-derived suppressor
cells through IL-6/Jak/Stat3 in PD-L1-high lung cancer. J
Immunother Cancer. 13:e0106122025. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Wang Z, Hu Y, Song J, Ma P and Xia H:
Polymorphonuclear myeloid-derived suppressor cells regulates immune
recovery during HIV infection through PD-L1 and TGF-β pathways.
Front Cell Infect Microbiol. 14:15164212024. View Article : Google Scholar
|
|
97
|
Huang H, Zhu X, Yu Y, Li Z, Yang Y, Xia L
and Lu S: EGFR mutations induce the suppression of CD8+
T cell and anti-PD-1 resistance via ERK1/2-p90RSK-TGF-β axis in
non-small cell lung cancer. J Transl Med. 22:6532024. View Article : Google Scholar
|
|
98
|
Quach HT, Hou Z, Bellis RY, Saini JK,
Amador-Molina A, Adusumilli PS and Xiong Y: Next-generation
immunotherapy for solid tumors: Combination immunotherapy with
crosstalk blockade of TGFβ and PD-1/PD-L1. Expert Opin Investig
Drugs. 31:1187–1202. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Hara D, Sasaki K, Doi S, Ike T, Maeda K,
Yoshida M, Takahashi A, Osaki Y, Ishiuchi N, Maeoka Y, et al:
Targeting MLL1/WDR5-mediated epigenetic regulation mitigates
peritoneal fibrosis by reducing p16INK4a. FASEB J.
39:e705432025. View Article : Google Scholar
|
|
100
|
Ji YF, Li T, Jiang F, Ni WK, Guan CQ, Liu
ZX, Lu CH, Ni RZ, Wu W and Xiao MB: Correlation between S100A11 and
the TGF-β1/SMAD4 pathway and its effects on the
proliferation and apoptosis of pancreatic cancer cell line PANC-1.
Mol Cell Biochem. 450:53–64. 2019. View Article : Google Scholar
|
|
101
|
Zhang B, Wu Y, Zhou C, Xie J, Zhang Y,
Yang X, Xiao J, Wang DW, Shan C, Zhou X, et al: Hyperactivation of
ATF4/TGF-β1 signaling contributes to the progressive cardiac
fibrosis in Arrhythmogenic cardiomyopathy caused by DSG2 variant.
BMC Med. 22:3612024. View Article : Google Scholar
|
|
102
|
Chen SY, Kung HC, Espinoza B, Washington
I, Chen K, Wang J, Zlomke H, Loycano M, Wang R, Pickup M, et al:
Targeting heterogeneous tumor microenvironments in pancreatic
cancer mouse models of metastasis by TGF-β depletion. JCI Insight.
9:e1827662024. View Article : Google Scholar
|
|
103
|
Yokoyama S, Shigeishi H, Murodumi H,
Sakuma M, Kato H, Higashikawa K, Ohta K, Sugiyama M and Takechi M:
TGF-β1 induces amoeboid-to-mesenchymal transition of
CD44high oral squamous cell carcinoma cells via miR-422a
downregulation through ERK activation and cofilin-1
phosphorylation. J Oral Pathol Med. 50:155–164. 2021. View Article : Google Scholar
|
|
104
|
Yang W, Zhang S, Li T, Zhou Z and Pan J:
Single-cell analysis reveals that cancer-associated fibroblasts
stimulate oral squamous cell carcinoma invasion via the TGF-β/Smad
pathway. Acta Biochim Biophys Sin (Shanghai). 55:262–273. 2022.
View Article : Google Scholar
|
|
105
|
Qin K, Gong C, Cheng Y, Li L, Liu C, Yang
F, Rao J and Li Q: Radiomics-based model for prediction of TGF-β1
expression in head and neck squamous cell carcinoma. Am J Nucl Med
Mol Imaging. 14:239–252. 2024. View Article : Google Scholar
|
|
106
|
Li Y, Fujishita T, Mishiro-Sato E, Kojima
Y, Niu Y, Taketo MM, Urano Y, Sakai T, Enomoto A, Nishida Y and
Aoki M: TGF-β signaling promotes desmoid tumor formation via CSRP2
upregulation. Cancer Sci. 115:401–411. 2024. View Article : Google Scholar :
|
|
107
|
Fasano M, Pirozzi M, Miceli CC, Cocule M,
Caraglia M, Boccellino M, Vitale P, De Falco V, Farese S, Zotta A,
et al: TGF-β modulated pathways in colorectal cancer: New potential
therapeutic opportunities. Int J Mol Sci. 25:74002024. View Article : Google Scholar
|
|
108
|
Joachims ML, Leehan KM, Dozmorov MG,
Georgescu C, Pan Z, Lawrence C, Marlin MC, Macwana S, Rasmussen A,
Radfar L, et al: Sjögren's syndrome minor salivary gland
CD4+ memory T cells associate with glandular disease
features and have a germinal center T follicular helper
transcriptional profile. J Clin Med. 9:21642020. View Article : Google Scholar
|
|
109
|
Hajiran A, Chakiryan N, Aydin AM, Zemp L,
Nguyen J, Laborde JM, Chahoud J, Spiess PE, Zaman S, Falasiri S, et
al: Reconnaissance of tumor immune microenvironment spatial
heterogeneity in metastatic renal cell carcinoma and correlation
with immunotherapy response. Clin Exp Immunol. 204:96–106. 2021.
View Article : Google Scholar
|
|
110
|
John M, Helal M, Duell J, Mattavelli G,
Stanojkovska E, Afrin N, Leipold AM, Steinhardt MJ, Zhou X, Žihala
D, et al: Spatial transcriptomics reveals profound subclonal
heterogeneity and T-cell dysfunction in extramedullary myeloma.
Blood. 144:2121–2135. 2024. View Article : Google Scholar
|
|
111
|
Honda CK, Kurozumi S, Fujii T, Pourquier
D, Khellaf L, Boissiere F, Horiguchi J, Oyama T, Shirabe K, Colinge
J, et al: Cancer-associated fibroblast spatial heterogeneity and
EMILIN1 expression in the tumor microenvironment modulate TGF-β
activity and CD8+ T-cell infiltration in breast cancer.
Theranostics. 14:1873–1885. 2024. View Article : Google Scholar
|
|
112
|
Chatterjee S, Chatterjee A, Jana S, Dey S,
Roy H, Das MK, Alam J, Adhikary A, Chowdhury A, Biswas A, et al:
Transforming growth factor beta orchestrates PD-L1 enrichment in
tumor-derived exosomes and mediates CD8 T-cell dysfunction
regulating early phosphorylation of TCR signalome in breast cancer.
Carcinogenesis. 42:38–47. 2021. View Article : Google Scholar
|
|
113
|
Gray GK, Li CM, Rosenbluth JM, Selfors LM,
Girnius N, Lin JR, Schackmann RCJ, Goh WL, Moore K, Shapiro HK, et
al: A human breast atlas integrating single-cell proteomics and
transcriptomics. Dev Cell. 57:1400–1420.e7. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Wu Y, Cheng Y, Wang X, Fan J and Gao Q:
Spatial omics: Navigating to the golden era of cancer research.
Clin Transl Med. 12:e6962022. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Liu Y, Yang M, Deng Y, Su G, Enninful A,
Guo CC, Tebaldi T, Zhang D, Kim D, Bai Z, et al:
High-spatial-resolution multiomics sequencing via deterministic
barcoding in tissue. Cell. 183:1665–1681.e18. 2020. View Article : Google Scholar
|
|
116
|
Vickovic S, Lötstedt B, Klughammer J,
Mages S, Segerstolpe Å, Rozenblatt-Rosen O and Regev A: SM-Omics is
an automated platform for high-throughput spatial multi-omics. Nat
Commun. 13:7952022. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Palacios PA, Flores I, Cereceda L, Otero
FF, Müller M, Brebi P, Contreras HR and Carreño LJ: Patient-derived
organoid models for NKT cell-based cancer immunotherapy. Cancers
(Basel). 17:4062025. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
von Kries A, Garcés-Lázaro I, Balzasch BM,
Sticht C, Shaltiel IA, Boonekamp KE, Sams A, Triassi A, Hofman T,
Burgermeister E, et al: Colorectal cancer organoids drive hypoxia,
TGF-β, and patient-specific diversification of NK cell activation
programs. J Immunother Cancer. 13:e0129882025. View Article : Google Scholar
|
|
119
|
Varricchio L, Iancu-Rubin C, Upadhyaya B,
Zingariello M, Martelli F, Verachi P, Clementelli C, Denis JF,
Rahman AH, Tremblay G, et al: TGF-β1 protein trap AVID200
beneficially affects hematopoiesis and bone marrow fibrosis in
myelofibrosis. JCI Insight. 6:e1456512021. View Article : Google Scholar
|
|
120
|
National Library of Medicine: A trial of
AVID200, a transforming growth factor β (TGFβ) inhibitor, in
patients malignancies. https://clinicaltrials.gov/ct2/show/NCT03834662.
Accessed September 20, 2024
|
|
121
|
Yap TA, Lakhani NJ, Araujo DV, Ahnert JR,
Chandana SR, Sharma M, Denis JF, Gruosso T, Tremblay G, O'Connor M,
et al: AVID200, first-in-class TGF-beta 1 and 3 selective and
potent inhibitor: Safety and biomarker results of a phase I
monotherapy dose-escalation study in patients with advanced solid
tumors. J Clin Oncol. 38(Suppl 15): S35872020. View Article : Google Scholar
|
|
122
|
Bazyari MJ, Saadat Z, Firouzjaei AA and
Aghaee-Bakhtiari SH: Deciphering colorectal cancer progression
features and prognostic signature by single-cell RNA sequencing
pseudotime trajectory analysis. Biochem Biophys Rep.
35:1014912023.PubMed/NCBI
|
|
123
|
Myung JH, Cha A, Tam KA, Poellmann M,
Borgeat A, Sharifi R, Molokie RE, Votta-Velis G and Hong S:
Dendrimer-based platform for effective capture of tumor cells after
TGFβ1-induced epithelial-mesenchymal transition. Anal
Chem. 91:8374–8382. 2019. View Article : Google Scholar
|
|
124
|
Alonso-Alconada L, Muinelo-Romay L,
Madissoo K, Diaz-Lopez A, Krakstad C, Trovik J, Wik E, Hapangama D,
Coenegrachts L, Cano A, et al: Molecular profiling of circulating
tumor cells links plasticity to the metastatic process in
endometrial cancer. Mol Cancer. 13:2232014. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Liu K, Wang Y, Wang C, Guo C and Zhang D,
Zhong Y, Yin L, Lu Y, Liu F, Zhang Y and Zhang D: Spatial
transcriptomics of gastric cancer brain metastasis reveals atypical
vasculature strategies with supportive immune profiles.
Gastroenterol Rep (Oxf). 12:goae0672024. View Article : Google Scholar :
|
|
126
|
Xiao M, Liang X, Yan Z, Chen J, Zhu Y, Xie
Y, Li Y, Li X, Gao Q, Feng F, et al: A DNA-methylation-driven genes
based prognostic signature reveals immune microenvironment in
pancreatic cancer. Front Immunol. 13:8039622022. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Chen L, Qiu X, Dupre A, Pellon-Cardenas O,
Fan X, Xu X, Rout P, Walton KD, Burclaff J, Zhang R, et al: TGFB1
induces fetal reprogramming and enhances intestinal regeneration.
Cell Stem Cell. 30:1520–1537.e8. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Lengrand J, Pastushenko I, Vanuytven S,
Song Y, Venet D, Sarate RM, Bellina M, Moers V, Boinet A, Sifrim A,
et al: Pharmacological targeting of netrin-1 inhibits EMT in
cancer. Nature. 620:402–408. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Yi L, Jiang X, Zhou Z, Xiong W, Xue F, Liu
Y, Xu H, Fan B, Li Y and Shen J: A hybrid nanoadjuvant
simultaneously depresses PD-L1/TGF-β1 and activates cGAS-STING
pathway to overcome radio-immunotherapy resistance. Adv Mater.
36:e23043282024. View Article : Google Scholar
|
|
130
|
Sparano C, Solís-Sayago D, Zangger NS,
Rindlisbacher L, Van Hove H, Vermeer M, Westermann F, Mussak C,
Rallo E, Dergun S, et al: Autocrine TGF-β1 drives tissue-specific
differentiation and function of resident NK cells. J Exp Med.
222:e202409302025. View Article : Google Scholar
|
|
131
|
Hanusek K, Rybicka B, Popławski P,
Adamiok-Ostrowska A, Głuchowska K, Piekiełko-Witkowska A and
Bogusławska J: TGF-β1 affects the renal cancer miRNome and
regulates tumor cells proliferation. Int J Mol Med. 49:522022.
View Article : Google Scholar
|