Non-small cell lung cancer (NSCLC) is the primary
histological type of lung cancer, accounting for ~85% of all lung
cancer cases. Its subtypes include adenocarcinoma, squamous cell
carcinoma, and large-cell carcinoma (1-3).
NSCLC exhibits marked molecular heterogeneity and a complex tumor
microenvironment, both of which contribute to tumor stemness,
metastatic potential and therapeutic resistance (4,5).
Although significant progress has been made in surgery,
radiotherapy, chemotherapy, targeted therapy and immunotherapy, the
majority of patients are diagnosed at advanced stages, with the
5-year survival rate still <25%. Issues, such as drug resistance
and recurrence urgently require innovative therapies (6,7).
Neural stem cells (NSCs) are a type of progenitor cells with
self-renewal capacity and multi-lineage differentiation potential,
residing in the central nervous system of adult mammals (8). Under physiological conditions, NSCs
undergo asymmetric division to produce neurons and glial cells
(9), maintaining adult
neurogenesis, supporting learning, memory and nerve injury repair,
thus endowing the nervous system with high plasticity and
regenerative potential (10).
In recent years, the crosstalk between NSCs and
tumor cells has gradually emerged as a critical perspective for
unraveling the mechanisms underlying tumor initiation and
progression (11,12). Although the tumor microenvironment
(TME) is commonly recognized as a sophisticated ecosystem composed
of tumor cells, immune cells, fibroblasts and extracellular matrix,
the pivotal roles of neural components and stem-like properties of
tumor cells within this ecosystem have long been overlooked.
Accumulating evidence currently demonstrates that NSCLC cells
express a variety of NSC markers and activate relevant signaling
pathways to exhibit 'NSC-like behaviors (13). Moreover, neurogenic factors,
neurotransmitters and direct cell-cell contact can profoundly
modulate the proliferation, stemness maintenance, invasion and
metastasis capacities of tumor cells (14-16). This emerging research niche has
been conceptualized as the 'NSC-NSCLC axis'. Within this framework,
NSCLC cells augment cancer stemness and therapeutic resistance by
hijacking multiple core signaling cascades of neural stem cells;
nerve fibers residing in the TME remodel the immune
microenvironment and facilitate angiogenesis via the secretion of
neurotrophic factors. Intriguingly, tumors can reciprocally
regulate peripheral neurons and potential NSC-like cell
populations, thereby establishing a bidirectional regulatory loop.
Nevertheless, the precise molecular mechanisms and therapeutic
targets in this field remain incompletely characterized.
Therefore, analyzing the key molecular drivers of
the 'NSCs-NSCLC axis', the associated microenvironment cross-talk
networks and their functions in malignant progression will help
uncover novel biological targets and provide innovative insights
for the precise diagnosis and treatment of NSCLC. The present
review aimed to summarize the latest research advances in this
field and provide perspectives on potential therapeutic strategies
(Fig. 1).
NSCs are the main population of endogenous adult
stem cells in the central nervous system, primarily residing in the
subventricular zone and the subgranular zone of the hippocampal
dentate gyrus (17). Under
pathological conditions, NSCs can also be derived from reactive
astrocytes or neural crest-derived progenitor cells (18,19). Their core characteristics involve
the long-term maintenance of a dynamic balance between
'self-renewal' and 'multi-lineage differentiation' potential
(20,21). This balance is regulated by a set
of highly conserved molecular markers, such as Nestin, SOX2,
Musashi-1, CD133, along with core signaling pathways such as Notch,
Wnt/β-catenin, Shh and BMP (22,23). Notably, these molecules and
pathways are also highly active in cancer stem cells (CSCs). This
'molecular homology' is not coincidental, but rather reflects the
evolutionary co-option of developmental programs by tumor cells
(24). For example, the Notch
signaling pathway regulates cell fate determination in normal NSCs,
whereas in NSCLC CSCs, it is frequently constitutively activated,
thereby promoting stemness maintenance and chemotherapeutic
resistance (4). However, the
majority of existing studies remain largely descriptive of this
overlap (24,25). The crosstalk among distinct
signaling pathways within specific TME contexts, as well as their
spatiotemporal dynamics, remains poorly understood, representing a
critical area for future mechanistic investigation.
In the TME, NSCs can be actively recruited.
Chemokines and growth factors, including SDF-1α/CXCL12, SCF, IL-6
and TGF-β secreted by tumor cells and tumor-associated macrophages
bind to surface receptors, such as CXCR4 and c-Kit on NSCs,
subsequently activating the PI3K/Akt, MAPK and JAK/STAT3 signaling
pathways to drive the directional migration of NSCs toward tumor
lesions (26-28). In addition, the hypoxic
microenvironment upregulates VEGF expression via HIF-1α, which
cooperates with exosome-carried miR-21 to further facilitate the
homing and survival of NSCs (29). These mechanisms lay a cellular
foundation for the subsequent functional aberrance of NSCs within
the TME. After entering the TME, the intrinsic normal
differentiation program of NSCs is hijacked. Tumor cell-derived
Notch ligands (Jagged1/DLL4) persistently activate intracellular
Notch signaling in NSCs to repress cell differentiation (30); miR-21 and miR-210 encapsulated in
tumor-derived exosomes remodel the survival and metabolic
phenotypes of NSCs by targeting PTEN and inhibiting mitochondrial
metabolism, respectively (31);
furthermore, hypoxia upregulates the levels of pluripotency
factors, including Oct4 and Nanog in a HIF-1α-dependent manner,
arresting the differentiation of NSCs into mature neural cells
(32). Collectively, these
regulatory mechanisms endow NSCs with a pro-tumorigenic
phenotype
NSCLC CSCs achieve the functional hijacking of the
NSC programs by aberrantly activating core molecular signatures and
signaling networks that highly overlap with those of NSCs,
exhibiting significant 'NSC-like behavior (33). This 'co-option of homologous
programs' not only endows NSCLC CSCs with extreme stemness,
therapeutic resistance and metastatic potential, but also provides
a molecular basis for the tumor cells to efficiently recruit and
utilize authentic NSCs within the TME.
A summary of the functional divergence of certain
shared markers and their clinical significance is provided in
Table I (34-73). Taking SOX2 as an example, in
normal NSCs, it forms a positive feedback loop with other factors
(such as Nestin and TLX) to maintain stemness homeostasis (34). However, in NSCLC CSCs, SOX2 often
collaborates with ASCL1 to induce neuroendocrine differentiation
(35), and interacts with various
factors such as miRNAs, lncRNAs, STAT3 and Wnt/β-catenin to drive
tumor progression and drug resistance (36,37), becoming an independent predictor
of a poor prognosis (40).
Notably, discrepancies exist among studies regarding the reported
functions of these markers, which may arise from tumor
heterogeneity, differences in model systems (such as cell lines,
organoids and in vivo models),and the complexity of
microenvironmental signals. For example, CD133 expression is
associated with multiple pro-tumor characteristics: It can maintain
stemness by activating the Src/PI3K/mTOR pathway (49); under hypoxic conditions,
CD133+ cells enhance adaptability by stabilizing
HIF1α/2α (50). Furthermore, this
subpopulation exhibits a strong potential for initiating brain
metastasis (53,54). However, the expression and
function of CD133 are not fixed and may be dynamically regulated by
microenvironmental factors such as hypoxia and inflammation.
Therefore, future research should not be limited to descriptive
correlations of markers, but should also delve into their upstream
regulatory mechanisms and downstream effect networks in specific
clinical subtypes or microenvironmental contexts.
Notably, the hijacking of neural programs is not
confined to stemness and therapeutic resistance, but extends
directly to metastasis. Several of the shared markers and pathways
co-opted by NSCLC CSCs are mechanistically linked to invasive and
metastatic behavior. CD133+ subpopulations represent the
strongest initiators of brain metastasis (53,54), and Nestin serves as an independent
risk factor for brain and bone metastasis in lung adenocarcinoma by
forming complexes with vimentin/Snail/Slug to drive
epithelial-mesenchymal transition (58-60). Moreover, as detailed below,
functionally reprogrammed NSCs further promote metastasis by
secreting VEGF to induce angiogenesis and releasing MMP-2/9 to
degrade the extracellular matrix, thereby paving the way for the
invasion and metastatic dissemination of CSCs. This metastatic
dimension is further reinforced at the level of neural signaling
and of the brain-metastatic niche, as discussed in the
corresponding sections. In this sense, the same neural-program
hijacking that sustains stemness and resistance also drives the
metastatic potential of NSCLC.
After entering the TME, the normal differentiation
program of NSCs is hijacked, which in turn acts reciprocally on
NSCLC CSCs to form a malignant amplification loop.
Tumor-cell-secreted Notch ligands (Jagged1/DLL4) can persistently
activate intracellular Notch signaling in NSCs and inhibit their
differentiation (74); miR-21 and
miR-210 derived from tumor exosomes remodel the survival and
metabolic phenotypes of NSCs by targeting PTEN and suppressing
mitochondrial metabolism, respectively (31,75); the hypoxic microenvironment
upregulates pluripotency factors, such as Oct4 and Nanog via
HIF-1α, thereby blocking the differentiation of NSCs into mature
neural cells (76). Following
functional aberrance, these NSCs secrete neurotrophic factors,
including brain-derived neurotrophic factor (BDNF), nerve growth
factor (NGF) and glial cell line-derived neurotrophic factor (GDNF)
in a paracrine manner to activate PI3K/Akt,MAPK/ERK and other
signaling cascades in NSCLC CSCs, elevating the expression of
stemness markers and conferring chemoresistance (77,78). Furthermore, NSCs produce VEGF to
facilitate angiogenesis and release MMP-2/9 to degrade the
extracellular matrix, paving the way for the invasion and
metastasis of CSCs (79). A
subset of NSCs can further differentiate into neuron-like cells
that form synapse-like junctions with tumor cells to supply
sustained neurotrophic support (80). Collectively, the aforementioned
mechanisms constitute the molecular basis underlying the
bidirectional crosstalk between NSCs and NSCLC CSCs.
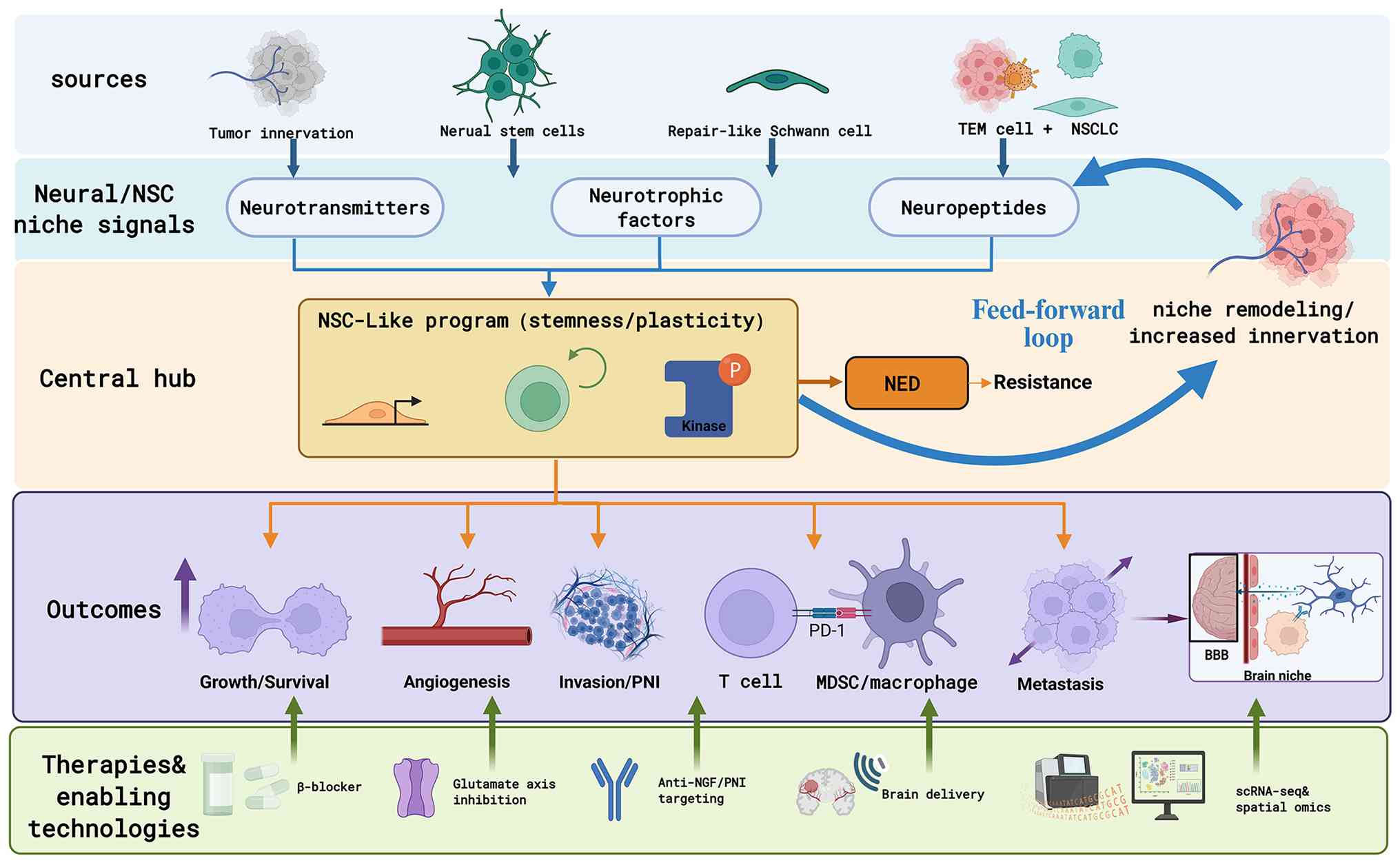
NSCLC is not a disease process driven solely by
genetic mutations. An increasing body of evidence suggests that,
under sustained neural input, tumor cells can undergo alterations
in receptor expression profiles and reprogram downstream signal
transduction, thereby converting external neural stimuli into a
broad spectrum of biological effects, including enhanced
proliferation and survival, invasion and metastasis, angiogenesis,
immune suppression and therapeutic resistance (81,82). These neural signals include
neurotransmitters, neurotrophic factors and neuropeptides. It
should be emphasized, however, that substantial heterogeneity
exists among studies with respect to experimental models, exposure
paradigms and endpoint definitions. As a result, the magnitude and
even the direction of the effects attributed to a given 'neural
signal' are not always consistent across different investigations.
Accordingly, this section is structured based on the relevance of
each signaling axis to NSCLC and the relative contribution of
direct versus indirect effects, thereby clarifying which findings
are approaching consensus and which should be interpreted as
preliminary or suggestive (Fig.
2).
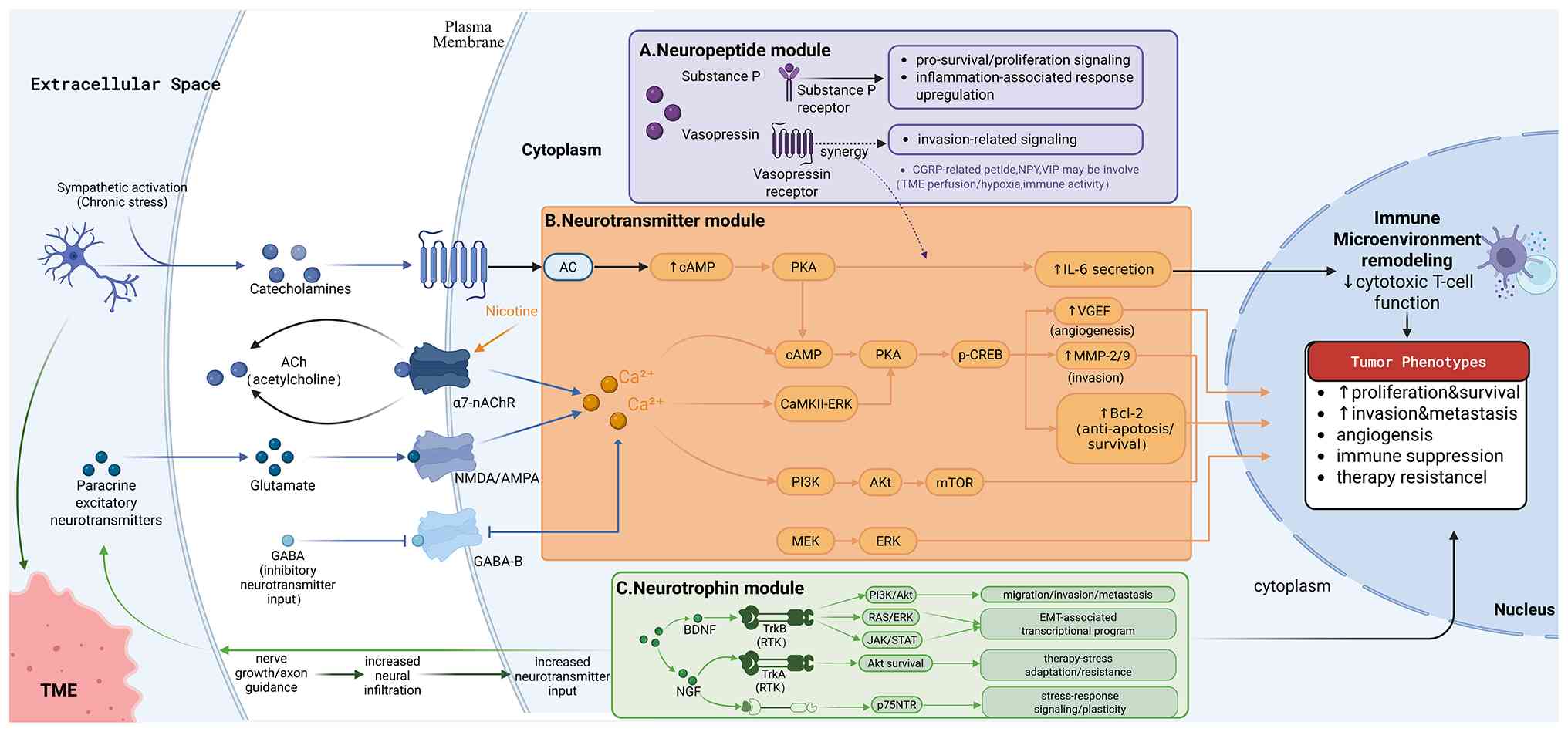
Among the currently available studies on NSCLC,
neurotransmitters exhibit the most direct associations (83-85). Evidence indicates that NSCLC cells
are not merely passively exposed to the surrounding neurochemical
milieu, but are frequently subjected to sustained and dynamic
neurotransmitter input (86). For
example, under chronic stress conditions, sympathetic excitation
increases the release of catecholamines, while excitatory
neurotransmitters, such as glutamate may also be present in the TME
via paracrine mechanisms (87).
Notably, NSCLC cells may even establish autocrine loops,
autonomously synthesizing acetylcholine (ACh), thereby conferring a
local 'autocrine growth factor-like' effect within the tumor niche
(88). Corresponding to these
diverse sources of neurotransmitters, NSCLC cells often upregulate
and exploit multiple classes of neurotransmitter receptors to
accomplish signal sensing and transduction. These include
β2-adrenergic receptor, nicotinic and muscarinic acetylcholine
receptors (particularly α7-nAChR), as well as NMDA/AMPA and GABA-B
receptors. The activation of these receptors converts external
neural stimuli into changes in second messengers, such as cAMP or
Ca2+, which are subsequently propagated through key
oncogenic pathways including PI3K/AKT/mTOR, MEK/ERK and CaMKII-ERK,
thereby driving malignant biological behaviors such as
proliferation and survival, angiogenesis, invasion and migration,
and immune suppression (Fig.
2B).
Catecholamine signaling provides a relatively
well-characterized example of neurotransmitter-mediated regulation
in NSCLC. Upon binding to β2-adrenergic receptors on NSCLC cells,
catecholamines stimulate adenylate cyclase, leading to elevated
intracellular cAMP levels and the activation of PKA. Activated PKA
phosphorylates CREB, inducing the upregulation of VEGF, MMP-2/9 and
Bcl-2, which respectively promote angiogenesis, enhance invasive
capacity and increase resistance to apoptosis (89). In parallel, this signaling axis
can stimulate the secretion of inflammatory cytokines, such as IL-6
(90), thereby remodeling the
immune microenvironment and attenuating cytotoxic T-cell activity,
resulting in a more pronounced immunosuppressive tumor phenotype.
By contrast, cholinergic signaling in NSCLC more commonly manifests
as a sustained driver of tumor growth and survival. Research
indicates show that NSCLC cells are capable of synthesizing and
releasing ACh. The immediate targets of ACh are cholinergic
receptors on the cell membrane, including both nicotinic and
muscarinic acetylcholine receptors, among which the α7-nAChR is
most frequently implicated in NSCLC research. Upon binding to
α7-nAChR, ACh triggers the earliest and most reproducible
event:receptor-mediated ion channel opening and Ca2+
influx (86). This
Ca2+ entry subsequently mobilizes multiple canonical
cancer-associated signaling modules. For example,
Ca2+-dependent nodes, such as CaMKII can participate in
signal amplification and couple with ERK activation, while the
PI3K/Akt/mTOR and MEK/ERK pathways are also commonly observed to be
upregulated downstream of α7-nAChR activation (91). In this context, nicotine, acting
as a high-affinity agonist of nAChRs, can markedly amplify
cholinergic signaling, thereby promoting aberrant clonal expansion
and enhancing phenotypes related to anti-apoptosis and resistance
to therapy (92).
Taken together, neurotransmitters represent the most
direct and environmentally sensitive class of neural inputs in
NSCLC. Through specific receptors, they translate host systemic
factors and local neural activity within the tumor into executable
intracellular events, such as cAMP or Ca2+, which are
then integrated through classical oncogenic modules, such as
PI3K/AKT/mTOR and MEK/ERK to remodel proliferation and survival,
invasion and migration, angiogenesis and the immune
microenvironment (93,94). Key questions for future
investigation include the following: First, the relative temporal
and spatial contributions of different neurotransmitter sources to
disease progression. Second, whether receptor expression profiles
are predictable and stratifiable in the context of tumor
heterogeneity. Third, whether neurotransmitter signaling represents
merely a correlate of treatment response or a causal driver of
therapeutic resistance in the setting of targeted therapy and
immunotherapy.
Neurotrophic factors constitute a central hub in
tumor-nerve interactions in NSCLC. Their sources are diverse,
including neurons, Schwann cells (SCs), tumor cells and stromal
cells (95). These factors
activate downstream signaling pathways through receptors, helping
tumor cells survive under stress conditions, such as detachment
from the matrix, hypoxia and treatment pressure. Studies indicate
that neuronal factors not only participate in regulating tumor
angiogenesis and microenvironmental remodeling (96), but also that NGF signaling is
closely related to the initiation and progression of
tumor-associated inflammation (97). Further exploration has revealed
the multi-layered mechanisms and translational prospects of the
'neuro-immune-tumor' axis in tumors such as gliomas (98). Notably, neurotrophic factors
inherently promote neuronal growth and axonal guidance (99), which may drive nerve fiber
infiltration and the increase of neuro-related structures within
tumors, thereby enhancing the input of neurotransmitters, such as
catecholamines and acetylcholine. This suggests that there may be a
positive feedback loop of mutual reinforcement between tumors and
neural components, where tumors are more likely to survive and
spread under the support of neurotrophic factors and neural
components, driven by neurotrophic factors, become further
enriched. The enhanced neural input continuously stimulates the
tumor and promotes its progression. In terms of the existing
evidence in NSCLC, BDNF and NGF are the most discussed neurotrophic
factors; however, their predominant roles are not identical. BDNF
is more commonly associated with migration, invasion, metastasis
and phenotype changes, translating external or tumor-derived
stimuli into stronger motility and dissemination capabilities. By
contrast, NGF is more often used to explain adaptive survival under
therapeutic stress, and its correspondence with the increase in
neural components within tumors is clearer, rendering it more
likely to be incorporated into the framework of tumor-neuron mutual
reinforcement. Collectively, the BDNF-TrkB and NGF-TrkA/p75NTR axes
constitute the core of neurotrophin signaling in NSCLC (Fig. 2C).
Within BDNF-related mechanisms, binding of BDNF to
TrkB activates receptor tyrosine kinase signaling and elicits
multiple downstream pathways (100). Research has provided relatively
consistent evidence that the PI3K-AKT axis represents a major
component of this signaling, being closely associated with enhanced
cellular survival under adverse conditions (101). Additional studies suggest the
involvement of RAS-ERK-related pathways, which are linked to
migration- and invasion-associated transcriptional programs, as
well as cytoskeletal remodeling (102). In certain experimental models,
outputs related to JAK-STAT signaling have also been observed
(103). Taken together, the
functional outcomes of these signaling pathways place the BDNF-TrkB
axis predominantly within a pro-migratory, pro-invasive and
pro-metastatic framework, with established links to transcriptional
regulatory networks associated with epithelial-mesenchymal
transition. In this sense, the BDNF axis can be viewed as a
signaling conduit that translates extrinsic or tumor-derived cues
into enhanced cellular motility and dissemination capacity.
In NGF-related mechanisms, the binding of NGF to
TrkA similarly triggers receptor tyrosine kinase signaling, with
research indicating that one of its key outputs remains the
AKT-associated survival pathway, which can be linked to the
tolerance of tumor cells to chemotherapy and other stresses
(104). Unlike TrkA, p75NTR does
not rely on intrinsic kinase activity, but instead modulates
stress-responsive pathways through the formation of complexes with
distinct intracellular binding proteins, thereby contributing to
cellular state plasticity and stress tolerance (105). In NSCLC, a key implication of
the NGF axis is that it points in two directions simultaneously: On
the one hand, toward intrinsic tumor cell survival and therapeutic
adaptation, and on the other hand, toward the correspondence
between its neurotrophic growth-promoting effects and increased
intratumoral neural structures. From this perspective, NGF is
better viewed as mediating a bidirectional association: NGF-related
signaling enables tumor cells to better withstand therapeutic
pressure, while simultaneously facilitating the expansion of neural
components within the tumor. The resulting increase in neural
inputs subsequently provides sustained exogenous stimulation via
neurotransmitter release, thereby reinforcing and perpetuating this
process.
The role of neuropeptides in NSCLC resembles that of
neurotrophic factors, but is better conceptualized as a signal
amplifier that converts neural activity into tumor behavior. By
binding to receptors on tumor cells or microenvironmental
components, neuropeptides translate external neural inputs into
signaling responses within the tumor (106,107). Overall, neuropeptides are
typically invoked to explain three categories of phenomena. First,
they facilitate tumor cell survival and tolerance under hypoxic
conditions, and conditions of nutrient fluctuations or therapeutic
stress. Second, they alter cell adhesion and motility, thereby
promoting migration and invasion and associated with an increased
risk of metastasis. Third, they remodel the TME by influencing
vascular status, inflammatory responses and immune cell function,
ultimately affecting tumor growth rate and treatment
responsiveness. As these effects often occur in parallel,
neuropeptides function less as singular growth factors and more as
regulatory links between neural input and tumor progression
(108) (Fig. 2A). Precisely as their actions are
distributed and highly context-dependent, the critical challenge in
neuropeptide research is not the continued expansion of their
catalog, but the identification of signaling axes supported by
causal evidence and druggability, which can be prioritized for
translational development.
From the perspective of translational potential and
research depth, substance P is the most worthy of priority
discussion. Closely associated with sensory nerves, substance P can
be detected within the TME (109). After its receptor is activated,
intracellular pro-survival and pro-proliferative pathways are
enhanced, often accompanied by the upregulation of
inflammation-related responses (110). In some studies, blocking this
receptor has been associated with reduced cell growth or increased
therapy-induced cell death, thus being repeatedly discussed as a
potential intervention target (111). However, much of the existing
evidence remains confined to functional associations observed in
in vitro systems or specific models. The critical next step
is to determine whether this axis truly determines treatment
response differences in well-stratified NSCLC population, and
whether blocking it could generate reproducible benefits in the
context of immunotherapy or chemoradiotherapy, rather than merely a
'universal effect' of inhibiting proliferation.
Another commonly discussed axis involves neurotensin
and its receptor. In certain NSCLC specimens, receptor expression
is higher and associated with greater invasiveness (112). This pathway is considered to
activate growth- and migration-related signaling and may synergize
with common growth factor receptor pathways, thus rendering the
tumor more likely to maintain proliferation and acquire stronger
migratory and invasive abilities (113).Additionally, other neuropeptides,
such as calcitonin gene-related peptide, neuropeptide Y and
vasoactive intestinal peptide have also been suggested to be
involved in regulation, primarily from a microenvironment
perspective, such as influencing perfusion and hypoxia or
modulating immune activity (108). Overall, neuropeptide research
should shift from 'which neuropeptides are involved' to 'which axes
are most capable of determining clinically relevant phenotypes and
are most amenable to pharmacological targeting'. Within the current
evidence framework, the substance P receptor axis and the
neurotensin receptor axis have higher priority and are closer to a
feasible translational intervention path.
In NSCLC, the 'neural-stem cell crosstalk' is not
merely regulated via paracrine signaling mediated by
neurotransmitters, neurotrophic factors or neuropeptides; instead,
it is predominantly manifested as niche remodeling driven by nerve
fiber infiltration and glial cell recruitment (114). Neural signals deliver not only
transient stimulatory cues, but may also reshape the TME into a
neutralized microenvironment that favors the survival, migration
and therapeutic escape of CSCs by altering local tissue
architecture and cellular composition (115,116).
Notably, intratumoral nerve fibers are
heterogeneously distributed and preferentially enriched at invasive
fronts, perivascular regions and sites with prominent stromal
remodeling, implying their synergistic interplay with invasive
pathways (117). Accordingly,
perineural invasion (PNI) enables tumor cells to spread along nerve
tracts and gain survival privileges, which is associated with
elevated risks of local recurrence and distant metastasis.
Moreover, PNI provides pathological evidence for spatially focused
neural input: Nerve tracts function as anatomically structured
migratory conduits on the one hand, and confer localized survival
benefits via continuous, short-range signaling and nutritional
supply on the other. In other words, PNI should no longer be
regarded solely as a passive consequence of advanced tumor
progression; it serves as a pathologically accessible research
model to verify whether neural-associated signaling pathways
actively facilitate tumor invasion and metastasis.
Furthermore, the core feature of neural invasion/PNI
lies not in the simple upregulation of soluble transmitters, but in
the transformation of neural input from intermittent to persistent
stimulation. Direct juxtaposition between nerve fibers and tumor
cells or NSC-like cells establishes stable local concentration
gradients, which endows CSC subsets with high receptor expression
with persistent selective advantages and shifts intratumoral
heterogeneity toward the emergence of neural-dependent clones
(118).
In addition to neuronal axons, SCs represent another
key cellular component of the peripheral nervous system whose role
in the neural microenvironment of NSCLC may have been substantially
underestimated. Under physiological conditions, SCs exhibit a
pronounced injury-response and repair capacity: Following nerve
injury, they can dedifferentiate into a 'repair-like' state
characterized by enhanced migratory ability, the secretion of
multiple growth factors and chemokines, and active remodeling of
the extracellular matrix to support axonal regeneration (119,120). As the TME and chronic
inflammation share substantial signaling similarities with 'tissue
injury', it is plausible that tumors co-opt the SC repair program,
positioning SCs as a potential hub linking neural input to the
maintenance of tumor phenotypes, including stemness-associated
states (121,122).
It should be emphasized that, to date, the NSCLC
field lacks direct experimental studies centered on 'SC-tumor
interactions'. Accordingly, the following discussion is primarily
intended to propose transferable and testable mechanistic
frameworks based on evidence from other solid tumors, rather than
to draw definitive conclusions for NSCLC. Existing studies
nonetheless provide at least two lines of evidence that are highly
relevant to NSCLC (93,94). First, in melanoma models, tumors
have been shown to reprogram surrounding SCs into a repair-like
state and activate 12/15-LOX (Alox15) and COX2-associated
eicosanoid pathways, thereby increasing the production of
immunosuppressive lipid mediators, such as PGE2. These mediators
subsequently suppress T-cell activation via EP4 signaling on
T-cells. By analogy, if similar repair-like SCs are present in the
peritumoral regions of NSCLC, they may contribute to the formation
of an immunosuppressive niche through a '12/15-LOX/COX2-PGE2-EP4'
axis (93). Second, in pancreatic
ductal adenocarcinoma, cancer cells can induce the c-Jun-dependent
injury-repair-like reprogramming of non-myelinating SCs, leading to
the formation of dynamic 'tumor-activated Schwann cell tracks'
(TASTs) that provide both physical paths and mechanical support for
tumor cell migration and local invasion (94). Correspondingly, in NSCLC
subpopulations exhibiting neural infiltration or PNI phenotypes,
whether SCs display c-Jun activation and form TAST-like structures,
and whether such structures spatially colocalize with the invasive
front, represents a question of high priority for experimental
validation.
Beyond their roles in immunity and invasion, SCs are
also key sources of neurotrophic signals. SCs can produce
neurotrophic factors, such as NGF, BDNF and GDNF, which may enhance
tumor cell survival and stress tolerance on the one hand (123), while promoting further growth
and the branching of nerve fibers on the other hand (124). Together, these effects may
reinforce intratumoral neural input. Moreover, dedifferentiated
repair-like SCs can participate in matrix remodeling and
alterations in adhesion molecule networks, facilitating tumor cell
migration along neural or stromal tracks and thereby providing
structural conditions for PNI and the formation of invasive fronts
(125,126). When tumor cells are chronically
exposed to a combination of neurotrophic factors, inflammatory
mediators and hypoxia within SC-enriched regions, they are more
likely to sustain the functional activity of proteins, such as SOX2
and Nestin, and to acquire enhanced resistance under therapeutic
pressure (127,128).
Therefore, the significance of discussing SCs in
NSCLC extends beyond merely 'identifying an additional cell type'.
Rather, SCs provide a more explanatory and potentially actionable
upstream framework for understanding why neural-related signals can
form localized, high-intensity and sustained inputs within tumors,
and how such persistent inputs establish more coherent causal links
with key phenotypes, such as immunosuppression, invasion and
metastasis, and therapeutic resistance. On this basis, validation
studies focusing on SC spatial localization, repair-like markers
and key signaling axes may help advance 'neural-related phenomena'
from correlative observations toward mechanistically tractable
targets.
Across the NSCs-NSCLC axis, inflammation emerges as
an integral intermediary rather than an incidental feature. Neural
signaling and inflammatory responses are tightly intertwined at
multiple nodes discussed throughout the present review. NGF
signaling is closely associated with the initiation and progression
of tumor-associated inflammation (97); catecholamine/β2-adrenergic
signaling stimulates the secretion of inflammatory cytokines, such
as IL-6, thereby remodeling the immune microenvironment and
attenuating cytotoxic T-cell activity (90); the activation of the substance P
receptor is frequently accompanied by upregulation of
inflammation-related responses (110) and repair-like Schwann cells
generate immunosuppressive lipid mediators, such as PGE2 via the
12/15-LOX/COX2-EP4 axis (93).
Inflammatory mediators act as key intermediaries linking neural
input to immune remodeling, stemness maintenance, and therapeutic
resistance.
An improved understanding of the NSCs-NSCLC axis is
reshaping contemporary paradigms of cancer therapy. Accumulating
evidence suggests that neural signals not only directly promote
tumor growth, but may also function as an upstream regulatory layer
driving multiple malignant phenotypes, including immunosuppression,
the maintenance of stemness, metastatic colonization and the
evolution of therapeutic resistance. For example, Cao (129) proposed the concept of 'neural
stemness' as a basal state underlying cellular tumorigenesis and
differentiation potential, providing a theoretical framework for
tumor stemness maintenance. Cerv antes-Villagrana et al
(130) demonstrated that
tumor-induced neurogenesis and immune evasion may represent
actionable targets for innovative anti-cancer therapies. As regards
metastatic colonization, Ouyang et al (131) revealed that the neuropeptide
precursor, neural growth factor-inducible gene (VGF), can remodel
the microenvironment through paracrine loops to promote liver
metastasis of uveal melanoma, implying that analogous mechanisms
may also operate in NSCLC. By contrast, Rotow and Bivona (132) focused directly on NSCLC and
comprehensively reviewed multiple resistance mechanisms, including
those mediated by neural signaling. Collectively, these findings
indicate that interventions targeting neural inputs possess
multifaceted clinical value. Such strategies may not only directly
suppress tumor cell proliferation and invasion, but may also
enhance the efficacy of immune checkpoint inhibitors (ICIs) by
relieving neural-associated immunosuppression, while additionally
contributing to cancer pain control and an improved quality of
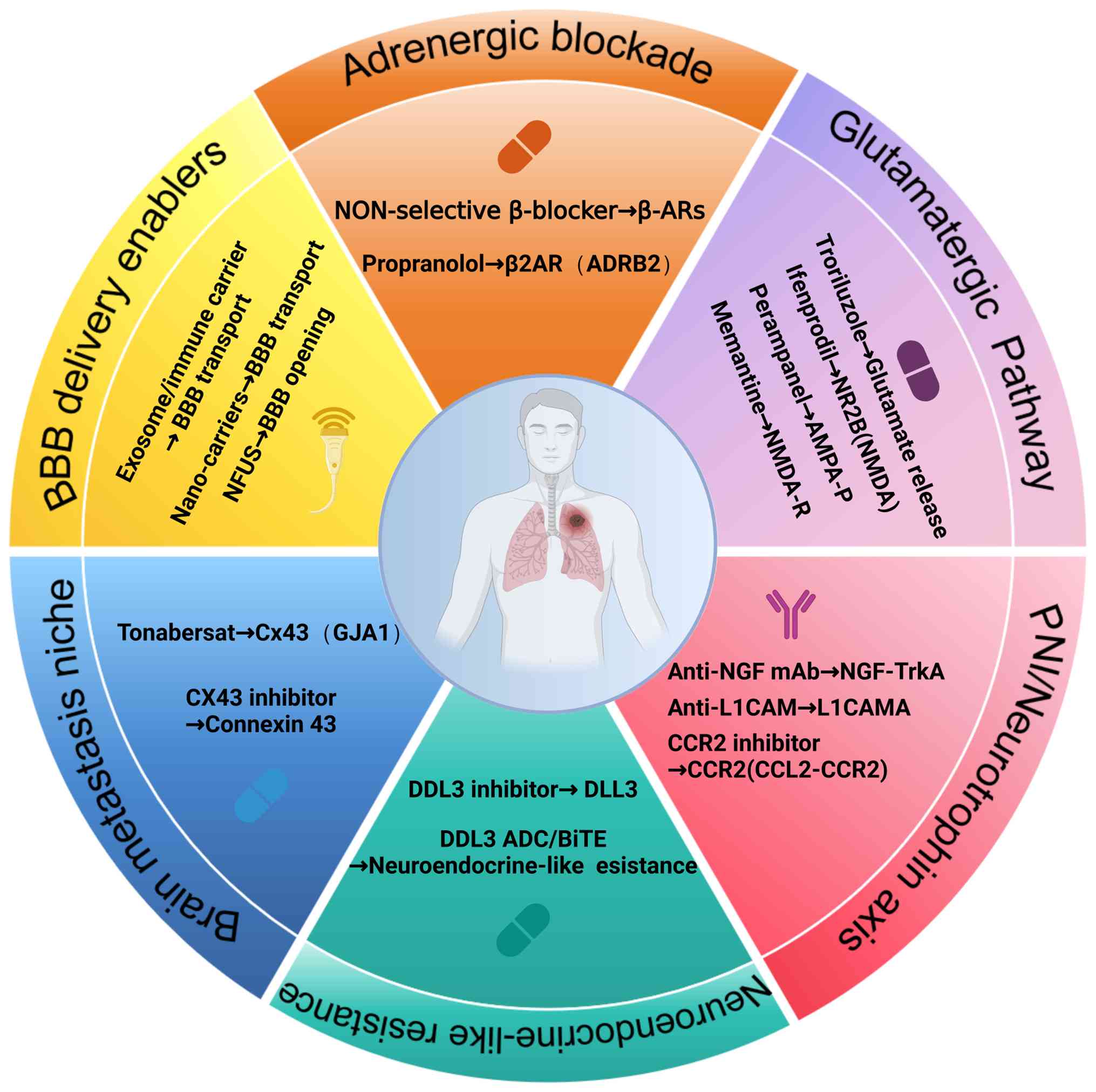
life. Current translational efforts mainly advance along five
directions: Receptor antagonism, intervention against PNI,
targeting neuroendocrine programs, neural adaptation in brain
metastases, and strategies for delivery and biomarker development
(Fig. 3).
Based on the growing understanding of
neurotransmitter signaling in NSCLC, clinical studies have begun to
explore therapeutic strategies targeting these systems. Clinical
evidence to date is mainly derived from retrospective and
observational studies. Among these, studies examining the impact of
non-selective β-adrenergic receptor blockers (β-blockers) on
patient survival have yielded notable progress. For example, a
scoping review published in 2024 summarized current evidence and
reported that the addition of β-blockers to standard lung cancer
therapies was associated with an improved overall survival (OS) and
recurrence-free survival, and a reduced incidence of metastasis
(133). However, findings in this field are not entirely consistent,
and more detailed subgroup analyses reveal a more complex image. A
large meta-analysis published in 2021, encompassing >30,000
patients, demonstrated that β-blocker use was not universally
associated with a significant improvement in OS among patients with
lung cancer (134).
Nevertheless, survival benefits were observed in specific
subpopulations, such as patients with stage III disease or those
who did not undergo surgical resection. Notably, the analysis
highlighted that the use of non-selective β-blockers was associated
with a worse OS (hazard ratio, 1.14), suggesting that distinct
β-blocker subtypes may exert divergent biological effects (134).
In contrast to these clinical observations,
preclinical and mechanistic studies provide the biological
rationale for this strategy. Mechanistic analyses have further
indicated that blockade of β-adrenergic signaling can reduce the
recruitment of myeloid-derived suppressor cells, downregulate PD-L1
expression and enhance CD8+ T-cell cytotoxic activity,
thereby providing a clear biological rationale for its combination
with ICIs (135). Accordingly,
prospective clinical trials can be designed to evaluate the
efficacy and safety of propranolol in combination with
immunotherapy in NSCLC and other solid tumors, with the aim of
enhancing immunotherapeutic responses and delaying the emergence of
resistance by alleviating stress-induced immunosuppression
(136).
Interventions targeting glutamatergic signaling in
NSCLC are currently achieved mainly through the use of pathway
inhibitors. At the preclinical level, NMDA receptor antagonists,
such as memantine and ifenprodil, have been shown in preclinical
models to suppress downstream signaling and reduce NSCLC cell
proliferation and migration (137). AMPA receptor antagonists,
including perampanel, are considered to disrupt excitatory
transmission between neurons and tumor cells, an effect that
appears particularly pronounced in the context of brain metastases
(138).
In terms of clinical translation, glutamate-release
inhibitors such as troriluzole can reduce glutamate levels within
the TME. The early clinical exploration of troriluzole in
combination with nivolumab has demonstrated preliminary
feasibility, providing a foundation for further optimization in
immunotherapy combinations and patient selection (139). Overall, the translational focus
of glutamatergic pathway-targeted strategies lies in defining
receptor expression profiles and identifying suitable patient
populations, particularly those with brain metastases or tumors
enriched in neural components, who are most likely to benefit from
interrupting excitatory neural inputs.
Evidence for these strategies currently stems
largely from preclinical and other tumor-type models. NGF and other
neurotrophic factors also represent key interventional entry
points. It has been demonstrated that the use of NGF-neutralizing
antibodies for analgesia can concomitantly attenuate tumor
dependence on neurotrophic support and growth signals, thereby
limiting PNI (140,141). In addition, interventions
targeting tumor migration along nerves may focus on disrupting cell
adhesion and recruitment networks. For example, interference with
L1CAM or the CCL2-CCR2 axis can weaken the cooperative interactions
between tumor cells, SCs and immune cells (142,143). The clinical value of such
strategies is more likely to manifest as a reduction in local
recurrence and nerve-related complications, while complementing
post-operative adjuvant therapy, radiotherapy, or other localized
treatments.
Neural-associated signaling can also contribute to
the evolution of therapeutic resistance through phenotypic
reprogramming, with neuroendocrine differentiation and small
cell-like transformation being the most representative examples.
Such transitions can be viewed as adaptive responses in which
tumors activate neural-related transcriptional programs under
therapeutic pressure to acquire increased invasiveness and
treatment resistance (144). As
DLL3 is highly expressed in small-cell lung cancer and
neuroendocrine phenotypes, but exhibits a relatively low expression
in normal tissues, it has emerged as an attractive
immunotherapeutic target (145).
Related strategies recognize and eliminate resistant clones based
on neural-like surface antigens: Clinically, the DLL3-targeting
bispecific T-cell engager (BiTE), tarlatamab, has entered phase I
evaluation in previously treated patients (146), whereas DLL3-targeting
antibody-drug conjugates currently remain at the preclinical stage
(147). A critical challenge in
this direction lies in early identification and dynamic monitoring:
Integrating histological and transcriptional features with liquid
biopsy to establish early warning and stratification systems before
and after phenotypic transformation, thereby enabling the
deployment of targeted strategies within the most appropriate
therapeutic window.
Brain metastasis is a major cause of mortality in
NSCLC. Its therapeutic intractability is attributable not only to
the blood-brain barrier (BBB), which restricts the penetration of
numerous pharmacologic agents, but also to the adaptive support
provided by immune networks formed by neural and glial cells within
the brain (148). At the
mechanistic level, preclinical studies have shown that brain
metastases can engage in metabolic exchange and signaling crosstalk
with astrocytes through Cx43-mediated gap junctions, thereby
establishing a tumor-favoring protective microenvironment and
enhancing resistance to chemotherapy (149). Consequently, Cx43 channel
blockers, such as tonabersat, represent a potential strategy to
disrupt astrocyte-mediated support (150,151).
In parallel, translational efforts are addressing
BBB-related delivery constraints, with encouraging progress in
receptor-mediated BBB nanocarriers, immune cell- or exosome-based
'Trojan horse' delivery and focused ultrasound (FUS)-mediated BBB
opening (152-155). Notably, these technologies are
not independent of neuro-oncological biology. If drugs targeting
neural-dependent pathways, such as inhibitors of the glutamatergic
signaling or glial interaction axes, can achieve effective
concentrations brain metastatic lesions (156-158), their efficacy signals may emerge
more clearly in brain metastases than in primary tumors, thereby in
turn accelerating drug development and biomarker maturation in this
field.
Single-cell sequencing and spatial omics
technologies are driving the field from mechanistic narratives
toward testable and stratifiable frameworks (159). At the single-cell level, these
approaches enable the identification of tumor cell subpopulations
with neural-like or stemness-associated features, as well as key
cellular states such as repair-like SCs, thereby illuminating
potential drug targets and trajectories of resistance evolution
(160). In parallel, spatial
transcriptomics preserves tissue architecture, while elucidating
spatial associations among nerve fibers, immune cells and tumor
cells, rendering phenomena such as the co-localization of increased
neural density and immune exclusion quantitatively assessable
(161,162). Based on these spatial and
single-cell insights, neural signal blockade may not only suppress
tumor growth, but may also enhance immunotherapeutic responses by
alleviating local immunosuppression, providing a more robust
rational for combining local neural modulation with systemic
therapies.
In summary, translational pathways targeting the
NSCs-NSCLC axis are becoming increasingly defined, from rapid
clinical entry via β-blockers and glutamatergic pathway inhibitors
(163), to structural
interventions targeting neurotrophic signaling and perineural
invasion with added analgesic benefits (140), to the precision
immunotherapeutic targeting of neuroendocrine
differentiation-associated resistance branches (164), and further breakthroughs
achieved by integrating glial interaction targets and advanced
delivery technologies in the brain metastatic niche (80). Future research priorities may
focus on three major areas: First, precision stratification: Not
all NSCLCs are neural-dependent, underscoring the urgent need for
scalable biomarkers based on neural density, neurotransmitter
levels and receptor expression profiles to identify responsive
populations. Second, brain metastasis intervention: Dissection of
the 'neuron-astrocyte-tumor' triadic interaction network to develop
combination therapies capable of penetrating the BBB and disrupting
key interaction nodes. Third, multidisciplinary integration:
Advancing oncology-neuroscience convergence under the framework of
cancer neuroscience to open new therapeutic avenues for refractory
lung cancer.
The 'NSCs-NSCLC axis' framework proposed in the
present review provides an integrated analytical perspective for
understanding the interplay between neural stem cells and NSCLC. At
present, studies on the interaction between NSCs and NSCLC remain
limited, particularly as regards the regulatory mechanisms of the
NSC-NSCLC axis within the TME. Translating discoveries in this
field into clinical therapies presents both challenges and
opportunities. One major obstacle is the BBB, which severely
restricts the delivery of agents targeting key receptors, thereby
rendering the highly active NSC-tumor interactions within brain
metastatic lesions difficult to modulate effectively. Accordingly,
the development of novel drug-delivery systems capable of
traversing both the BBB and the blood-tumor barrier is of critical
importance. Moreover, the same molecular signal may exert opposing
effects depending on microenvironmental context or concentration,
rendering single-target interventions prone to compensatory
resistance or off-target toxicity. A more in-depth mechanistic
analysis may facilitate the design of more precise multi-target
combination strategies. Emerging technologies, including
single-cell sequencing and spatial omics, provide powerful tools to
overcome these limitations. These approaches enable the
characterization of NSC heterogeneity within the TME and the
delineation of their spatial distribution at the tumor-immune
interface, thereby identifying the specific NSC subpopulations that
truly drive therapeutic resistance and metastasis and furnishing a
solid basis for precision targeting. Looking ahead, the integration
of spatial multi-omics with artificial intelligence-based analysis
may enable the identification of patient-specific
'neural-immune-stem cell' molecular landscapes, allowing for
individualized stratification and precise modulation of the neural
microenvironment. Combined with the dynamic monitoring of
peripheral blood biomarkers, such approaches could support the
real-time adjustment of therapeutic regimens. Collectively, this
paradigm may shift lung cancer treatment from direct tumor
eradication toward reprogramming of the tumor microecosystem,
transforming neural stem cells from 'drivers' of tumor progression
into controllable therapeutic targets.
Not applicable.
YX, HD and FL contributed to the conception and
design of the study. YX, HD and PL were responsible for the writing
of the manuscript. PL and FL performed the analysis and
interpretation of the data from the literature presented in the
table. YX, HD and FL participated in manuscript revision. All
authors have reviewed and approved the final version of the
manuscript. FL served as the corresponding author, overseeing
correspondence and final manuscript approval. Data authentication
is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
The authors sincerely appreciate the assistance from
Hengyang Central Hospital, which was instrumental in facilitating
the research infrastructure, data collection and analysis. The
authors also extend our gratitude to the hospital administration
and relevant departments for their institutional support.
No funding was received.
|
1
|
Giaccone G and He Y: Current knowledge of
small cell lung cancer transformation from non-small cell lung
cancer. Semin Cancer Biol. 94:1–10. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jha SK, De Rubis G, Devkota SR, Zhang Y,
Adhikari R, Jha LA, Bhattacharya K, Mehndiratta S, Gupta G, Singh
SK, et al: Cellular senescence in lung cancer: Molecular mechanisms
and therapeutic interventions. Ageing Res Rev. 97:1023152024.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xie X, Li X, Tang W, Xie P and Tan X:
Primary tumor location in lung cancer: The evaluation and
administration. Chin Med J (Engl). 135:127–136. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
He T, Wang Y, Lv W, Wang Y, Li X, Zhang Q,
Shen HM and Hu J: FBP1 inhibits NSCLC stemness by promoting
ubiquitination of Notch1 intracellular domain and accelerating
degradation. Cell Mol Life Sci. 81:872024. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen M, Sun LX, Yu L, Liu J, Sun LC, Yang
ZH, Shu X and Ran YL: MYH9 is crucial for stem cell-like properties
in non-small cell lung cancer by activating mTOR signaling. Cell
Death Discov. 7:2822021. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Haratake N, Ozawa H, Morimoto Y, Yamashita
N, Daimon T, Bhattacharya A, Wang K, Nakashoji A, Isozaki H,
Shimokawa M, et al: MUC1-C is a common driver of acquired
osimertinib resistance in NSCLC. J Thorac Oncol. 19:434–450. 2024.
View Article : Google Scholar
|
|
7
|
Rotow JK, Lee JK, Madison RW, Oxnard GR,
Jänne PA and Schrock AB: Real-World genomic profile of EGFR
second-site mutations and other osimertinib resistance mechanisms
and clinical landscape of NSCLC post-osimertinib. J Thorac Oncol.
19:227–239. 2024. View Article : Google Scholar
|
|
8
|
Llorente V, Velarde P, Desco M and
Gómez-Gaviro MV: Current understanding of the neural stem cell
niches. Cells. 11:30022022. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bunel B, Leclercq R, Goïame R, Gautier A,
Morin X and Fischer E: Unequal mitochondrial segregation promotes
asymmetric fates during neurogenesis. Nat Commun. 16:110492025.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hussain G, Akram R, Anwar H, Sajid F, Iman
T, Han HS, Raza C and De Aguilar JG: Adult neurogenesis: A real
hope or a delusion. Neural Regen Res. 19:6–15. 2024. View Article : Google Scholar
|
|
11
|
Vanden Abeele F and Salzet M: The
neuro-immune oncology axis. Cancer Lett. 634:2180702025. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Qu J, Li P and Sun Z: Histone lactylation
regulates cancer progression by reshaping the tumor
microenvironment. Front Immunol. 14:12843442023. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hassan KA, Wang L, Korkaya H, Chen G,
Maillard I, Beer DG, Kalemkerian GP and Wicha MS: Notch pathway
activity identifies cells with cancer stem cell-like properties and
correlates with worse survival in lung adenocarcinoma. Clin Cancer
Res. 19:1972–1980. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Thaker PH, Han LY, Kamat AA, Arevalo JM,
Takahashi R, Lu C, Jennings NB, Armaiz-Pena G, Bankson JA, Ravoori
M, et al: Chronic stress promotes tumor growth and angiogenesis in
a mouse model of ovarian carcinoma. Nat Med. 12:939–944. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Venkataramani V, Tanev DI, Strahle C,
Studier-Fischer A, Fankhauser L, Kessler T, Körber C, Kardorff M,
Ratliff M, Xie R, et al: Glutamatergic synaptic input to glioma
cells drives brain tumour progression. Nature. 573:532–538. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li X, Ma G, Ma Q, Li W, Liu J, Han L, Duan
W, Xu Q, Liu H, Wang Z, et al: Neurotransmitter substance P
mediates pancreatic cancer perineural invasion via NK-1R in cancer
cells. Mol Cancer Res. 11:294–302. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Makrygianni EA and Chrousos GP: Neural
progenitor cells and the hypothalamus. Cells. 12:18222023.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wei J, Wang M, Li S, Han R, Xu W, Zhao A,
Yu Q, Li H, Li M and Chi G: Reprogramming of astrocytes and glioma
cells into neurons for central nervous system repair and
glioblastoma therapy. Biomed Pharmacother. 176:1168062024.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lin JD, Hammad R, Kumar P, Manzitti P, Wu
K, Alzubi J, Vlachos A, Cathomen T, Blesch A, Chu YH and Schachtrup
C: ID3 -depleted human induced pluripotent stem cell-derived neural
stem/progenitor cells promote neurorepair. Neural Regen Res.
21:3730–3740. 2026. View Article : Google Scholar
|
|
20
|
Yu J, Chen G, Zhu H, Zhong Y, Yang Z, Jian
Z and Xiong X: Metabolic and proteostatic differences in quiescent
and active neural stem cells. Neural Regen Res. 19:43–48. 2024.
View Article : Google Scholar
|
|
21
|
Foley T, Thetiot M and Bally-Cuif L:
Neural stem cell regulation in zebrafish. Annu Rev Genet.
58:249–272. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Reinhard J, Brösicke N, Theocharidis U and
Faissner A: The extracellular matrix niche microenvironment of
neural and cancer stem cells in the brain. Int J Biochem Cell Biol.
81(Pt A): 174–183. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Drysch A, Ahuja A, Prasad D, Jain R,
Romanos S, Alwakeal A and Ahuja C: Transcriptomic comparisons of
somatic and cancer stem cells. Biomedicines. 13:20392025.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ajani JA, Song S, Hochster HS and
Steinberg IB: Cancer stem cells: The promise and the potential.
Semin Oncol. 42(Suppl 1): S3–S17. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Takebe N, Miele L, Harris PJ, Jeong W,
Bando H, Kahn M, Yang SX and Ivy SP: Targeting Notch, Hedgehog, and
Wnt pathways in cancer stem cells: Clinical update. Nat Rev Clin
Oncol. 12:445–464. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ma S, Zhou J, Huang T, Zhang Z, Xing Q,
Zhou X, Zhang K, Yao M, Cheng T, Wang X, et al: Sodium
alginate/collagen/stromal cell-derived factor-1 neural scaffold
loaded with BMSCs promotes neurological function recovery after
traumatic brain injury. Acta Biomater. 131:185–197. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Molina-Peña R, Haji Mansor M, Najberg M,
Thomassin JM, Gueza B, Alvarez-Lorenzo C, Garcion E, Jérôme C and
Boury F: Nanoparticle-containing electrospun nanofibrous scaffolds
for sustained release of SDF-1α. Int J Pharm. 610:1212052021.
View Article : Google Scholar
|
|
28
|
Zhu M, Li S, Cao X, Rashid K and Liu T:
The STAT family: Key transcription factors mediating crosstalk
between cancer stem cells and tumor immune microenvironment. Semin
Cancer Biol. 88:18–31. 2023. View Article : Google Scholar
|
|
29
|
Man J, Yu X, Huang H, Zhou W, Xiang C,
Huang H, Miele L, Liu Z, Bebek G, Bao S and Yu JS: Hypoxic
induction of vasorin regulates Notch1 turnover to maintain glioma
stem-like cells. Cell Stem Cell. 22:104–118.e6. 2018. View Article : Google Scholar
|
|
30
|
You WK, Schuetz TJ and Lee SH: Targeting
the DLL/Notch signaling pathway in cancer: challenges and advances
in clinical development. Mol Cancer Ther. 22:3–11. 2023. View Article : Google Scholar :
|
|
31
|
Chawra HS, Agarwal M, Mishra A, Chandel
SS, Singh RP, Dubey G, Kukreti N and Singh M: MicroRNA-21's role in
PTEN suppression and PI3K/AKT activation: Implications for cancer
biology. Pathol Res Pract. 254:1550912024. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Carrica L, Li L, Newville J, Kenton J,
Gustus K, Brigman J and Cunningham LA: Genetic inactivation of
hypoxia inducible factor 1-alpha (HIF-1α) in adult hippocampal
progenitors impairs neurogenesis and pattern discrimination
learning. Neurobiol Learn Mem. 157:79–85. 2019. View Article : Google Scholar
|
|
33
|
Li J, Wang J, Xie D, Pei Q, Wan X, Xing HR
and Ye T: Characteristics of the PI3K/AKT and MAPK/ERK pathways
involved in the maintenance of self-renewal in lung cancer
stem-like cells. Int J Biol Sci. 17:1191–1202. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shimozaki K: Sox2 transcription network
acts as a molecular switch to regulate properties of neural stem
cells. World J Stem Cells. 6:485–490. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Borromeo MD, Savage TK, Kollipara RK, He
M, Augustyn A, Osborne JK, Girard L, Minna JD, Gazdar AF, Cobb MH
and Johnson JE: ASCL1 and NEUROD1 reveal heterogeneity in pulmonary
neuroendocrine tumors and regulate distinct genetic programs. Cell
Rep. 16:1259–1272. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mirzaei S, Paskeh M, Entezari M,
Mirmazloomi SR, Hassanpoor A, Aboutalebi M, Rezaei S, Hejazi ES,
Kakavand A, Heidari H, et al: SOX2 function in cancers: Association
with growth, invasion, stemness and therapy response. Biomed
Pharmacother. 156:1138602022. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liang Y, Wang N, Zhang Y, Jiang W, Fang C,
Feng Y, Ma H, Jiang F and Dong G: Self-restricted circular RNA
circSOX2 suppressed the malignant progression in SOX2-amplified
LUSC. Cell Death Dis. 13:8732022. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Schaefer T, Mittal N, Wang H, Ataman M,
Candido S, Lötscher J, Velychko S, Tintignac L, Bock T, Börsch A,
et al: Nuclear and cytosolic fractions of SOX2 synergize as
transcriptional and translational co-regulators of cell fate. Cell
Rep. 43:1148072024. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Park JH, Kim YH, Shim S, Shim S, Kim A,
Jang H, Lee SJ, Park S, Seo S, Jang WI, et al: Radiation-Activated
PI3K/AKT pathway promotes the induction of cancer stem-like cells
via the upregulation of SOX2 in colorectal cancer. Cells.
10:1352021. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ding LN, Yu YY, Ma CJ, Lei CJ and Zhang
HB: SOX2-associated signaling pathways regulate biological
phenotypes of cancers. Biomed Pharmacother. 160:1143362023.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
de Wet L, Williams A, Gillard M, Kregel S,
Lamperis S, Gutgesell LC, Vellky JE, Brown R, Conger K, Paner GP,
et al: SOX2 mediates metabolic reprogramming of prostate cancer
cells. Oncogene. 41:1190–1202. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen Y, Chen Y, Li Q, Liu H, Han J, Zhang
H, Cheng L and Lin G: Short C-terminal Musashi-1 proteins regulate
pluripotency states in embryonic stem cells. Cell Rep.
42:1133082023. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Dolcemascolo R, Heras-Hernández M, Goiriz
L, Montagud-Martínez R, Requena-Menéndez A, Ruiz R, Pérez-Ràfols A,
Higuera-Rodríguez RA, Pérez-Ropero G, Vranken WF, et al:
Repurposing the mammalian RNA-binding protein Musashi-1 as an
allosteric translation repressor in bacteria. Elife.
12:RP917772024. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sicking M, Falke I, Löblein MT, Eich HT,
Götte M, Greve B and Troschel FM: The Musashi RNA-binding proteins
in female cancers: Insights on molecular mechanisms and therapeutic
relevance. Biomark Res. 11:762023. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ren G, Yang EJ, Tao S, Mou PK, Pu Y, Chen
LJ and Shim JS: MDM2 inhibition is synthetic lethal with PTEN loss
in colorectal cancer cells via the p53-dependent mechanism. Int J
Biol Sci. 19:3544–3557. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liang W, Chen Y, Liu H, Zhao H, Luo T,
Tang H, Zhou X, Jiang E, Shao Z, Liu K and Shang Z: Cancer cells
corrupt normal epithelial cells through miR-let-7c-rich small
extracellular vesicle-mediated downregulation of p53/PTEN. Int J
Oral Sci. 14:362022. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Deng C, Yuan F, Li C, Tong X, Qin Y, Wen
H, Qin T, Liu Y, Wu T, Duan C, et al: Exosomes from
CD133(+)CD271(+) human urine-derived stem cells combined with a
novel photosensitive hydrogel promote repair after spinal cord
injury. J Transl Med. 24:22025. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Izumi H, Kaneko Y and Nakagawara A:
Molecular regulation of autophagy and asymmetric cell division by
cancer stem cell marker CD133. Cells. 12:8192023. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wu YY, Wu HC, Wu JE, Huang KY, Yang SC,
Chen SX, Tsao CJ, Hsu KF, Chen YL and Hong TM: The dual PI3K/mTOR
inhibitor BEZ235 restricts the growth of lung cancer tumors
regardless of EGFR status, as a potent accompanist in combined
therapeutic regimens. J Exp Clin Cancer Res. 38:2822019. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Moreno Roig E, Groot AJ, Yaromina A,
Hendrickx TC, Barbeau LMO, Giuranno L, Dams G, Ient J, Olivo
Pimentel V, van Gisbergen MW, et al: HIF-1α and HIF-2α differently
regulate the radiation sensitivity of NSCLC cells. Cells. 8:452019.
View Article : Google Scholar
|
|
51
|
Zhang GN, Zhang YK, Wang YJ, Gupta P,
Ashby CR Jr, Alqahtani S, Deng T, Bates SE, Kaddoumi A, Wurpel JND,
et al: Epidermal growth factor receptor (EGFR) inhibitor PD153035
reverses ABCG2-mediated multidrug resistance in non-small cell lung
cancer: In vitro and in vivo. Cancer Lett. 424:19–29. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Sun H, Yao N, Cheng S, Li L, Liu S, Yang
Z, Shang G, Zhang D and Yao Z: Cancer stem-like cells directly
participate in vasculogenic mimicry channels in triple-negative
breast cancer. Cancer Biol Med. 16:299–311. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Tournier N, Goutal S, Auvity S, Traxl A,
Mairinger S, Wanek T, Helal OB, Buvat I, Soussan M, Caillé F and
Langer O: Strategies to Inhibit ABCB1- and ABCG2-Mediated efflux
transport of erlotinib at the blood-brain barrier: A PET study on
nonhuman primates. J Nucl Med. 58:117–122. 2017. View Article : Google Scholar
|
|
54
|
Dong XD, Zhang M, Teng QX, Lei ZN, Cai CY,
Wang JQ, Wu ZX, Yang Y, Chen X, Guo H and Chen ZS: Mobocertinib
antagonizes multidrug resistance in ABCB1- and ABCG2-overexpressing
cancer cells: In vitro and in vivo studies. Cancer Lett.
607:2173092024. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Nouri P, Zimmer A, Brüggemann S, Friedrich
R, Kühn R and Prakash N: Generation of a NES-mScarlet red
fluorescent reporter human iPSC line for live cell imaging and flow
cytometric analysis and sorting using CRISPR-Cas9-Mediated gene
editing. Cells. 11:2682022. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Lubanska D, Qemo I, Stringer KF, Mahendran
HP, Fifield BA, Cieslukowski A, Alrashed S, El-Abed Y, Boujeke E,
Rodzinka A, et al: Atypical cell cycle regulation over neural stem
cell expansion. Stem Cell Reports. 21:1027522026. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Matsubara S, Matsuda T and Nakashima K:
Regulation of adult mammalian neural stem cells and neurogenesis by
cell extrinsic and intrinsic factors. Cells. 10:11452021.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Wang R, Khan S, Liao G, Wu Y and Tang DD:
Nestin modulates airway smooth muscle cell migration by affecting
spatial rearrangement of vimentin network and focal adhesion
assembly. Cells. 11:30472022. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Liu X, Feng Q, Zhang Y, Zheng P and Cui N:
Absence of EpCAM in cervical cancer cells is involved in
sluginduced epithelial-mesenchymal transition. Cancer Cell Int.
21:1632021. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Jaramillo-Rangel G, Chávez-Briones MD,
Ancer-Arellano A and Ortega-Martínez M: Nestin-Expressing cells in
the lung: The bad and the good parts. Cells. 10:34132021.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Swartchick CB, Dirak M, Wenger LCF, Tapia
Hernandez R, Lee MC and Chan J: Activity-Based bioluminescent
logic-gate probe reveals crosstalk between the inflammatory tumor
microenvironment and ALDH1A1 in cancer cells. JACS Au. 5:320–331.
2024. View Article : Google Scholar
|
|
62
|
Xin Q, Ji Q, Zhang Y, Ma W, Tian B, Liu Y,
Chen Y, Wang F, Zhang R, Wang X and Yuan J: Aberrant ROS served as
an acquired vulnerability of cisplatin-resistant lung cancer. Oxid
Med Cell Longev. 2022:11129872022. View Article : Google Scholar :
|
|
63
|
Pandya P, Al-Qasrawi DS, Klinge S and
Justilien V: Extracellular vesicles in non-small cell lung cancer
stemness and clinical applications. Front Immunol. 15:13693562024.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
van Kerkhof P, Kralj T, Spanevello F, van
Bloois L, Jordens I, van der Vaart J, Jamieson C, Merenda A,
Mastrobattista E and Maurice MM: RSPO3 Furin domain-conjugated
liposomes for selective drug delivery to LGR5-high cells. J Control
Release. 356:72–83. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Wang W, Lokman NA, Barry SC, Oehler MK and
Ricciardelli C: LGR5: An emerging therapeutic target for cancer
metastasis and chemotherapy resistance. Cancer Metastasis Rev.
44:232025. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Luo X, Wang J, Chen Y, Zhou X, Shao Z, Liu
K and Shang Z: Melatonin inhibits the stemness of head and neck
squamous cell carcinoma by modulating HA synthesis via the
FOSL1/HAS3 axis. J Pineal Res. 76:e129402024. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Espejo-Román JM, Rubio-Ruiz B,
Chayah-Ghaddab M, Vega-Gutierrez C, García-García G,
Muguruza-Montero A, Domene C, Sánchez-Martín RM, Cruz-López O and
Conejo-García A: N-aryltetrahydroisoquinoline derivatives as
HA-CD44 interaction inhibitors: Design, synthesis, computational
studies, and antitumor effect. Eur J Med Chem. 258:1155702023.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Jeibouei S, Monfared AK, Hojat A, Aref AR,
Shams F, Dolati M, Moradi A, Hosseini M, Javadi SM, Ajoudanian M,
et al: Human-derived Tumor-On-Chip model to study the heterogeneity
of breast cancer tissue. Biomater Adv. 162:2139152024. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Tsai T, Wu S, Lai Y, Wang H, Hou P, Huang
Y, Chen HH and Su W: CD44-hyaluronan mediating endocytosis of
iron-platinum alloy nanoparticles induces ferroptotic cell death in
mesenchymal-state lung cancer cells with tyrosine kinase inhibitor
resistance. Acta Biomater. 186:396–410. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Huang Q, Xu X, Halbiyat Z, Wei X, Wang L,
Ren J, Xu K, Huang T and Shuai Q: Collagen/fibronectin-based lung
carcinoma culture platform: development and characterization of a
new tumor model for vascular mimicry study. J Mater Chem B.
13:7449–7462. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Tan B, Zhang J, Wang W, Ma H and Yang Y:
Tumor-suppressive E3 ubiquitin ligase CHIP inhibits the PBK/ERK
axis to repress stem cell properties and radioresistance in
non-small cell lung cancer. Apoptosis. 28:397–413. 2023. View Article : Google Scholar
|
|
72
|
Vainorius G, Novatchkova M, Michlits G,
Baar JC, Raupach C, Lee J, Yelagandula R, Wernig M and Elling U:
Ascl1 and Ngn2 convert mouse embryonic stem cells to neurons via
functionally distinct paths. Nat Commun. 14:53412023. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Olsen RR, Ireland AS, Kastner DW, Groves
SM, Spainhower KB, Pozo K, Kelenis DP, Whitney CP, Guthrie MR, Wait
SJ, et al: ASCL1 represses a SOX9(+) neural crest stem-like state
in small cell lung cancer. Genes Dev. 35:847–869. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Androutsellis-Theotokis A, Leker RR,
Soldner F, Hoeppner DJ, Ravin R, Poser SW, Rueger MA, Bae SK,
Kittappa R and McKay RD: Notch signalling regulates stem cell
numbers in vitro and in vivo. Nature. 442:823–826. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Li F, Zhang J, Liao R, Duan Y, Tao L, Xu Y
and Chen A: Mesenchymal stem cellderived extracellular vesicles
prevent neural stem cell hypoxia injury via promoting miR2103p
expression. Mol Med Rep. 22:3813–3821. 2020.PubMed/NCBI
|
|
76
|
Mathieu J, Zhang Z, Zhou W, Wang AJ,
Heddleston JM, Pinna CM, Hubaud A, Stadler B, Choi M, Bar M, et al:
HIF induces human embryonic stem cell markers in cancer cells.
Cancer Res. 71:4640–4652. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Venkatesh HS, Johung TB, Caretti V, Noll
A, Tang Y, Nagaraja S, Gibson EM, Mount CW, Polepalli J, Mitra SS,
et al: Neuronal activity promotes glioma growth through
neuroligin-3 secretion. Cell. 161:803–816. 2015. View Article : Google Scholar
|
|
78
|
Lagadec C, Meignan S, Adriaenssens E,
Foveau B, Vanhecke E, Romon R, Toillon RA, Oxombre B, Hondermarck H
and Le Bourhis X: TrkA overexpression enhances growth and
metastasis of breast cancer cells. Oncogene. 28:1960–1970. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Barkho BZ, Munoz AE, Li X, Li L,
Cunningham LA and Zhao X: Endogenous matrix metalloproteinase
(MMP)-3 and MMP-9 promote the differentiation and migration of
adult neural progenitor cells in response to chemokines. Stem
Cells. 26:3139–3149. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Zeng Q, Michael IP, Zhang P, Saghafinia S,
Knott G, Jiao W, McCabe BD, Galván JA, Robinson HPC, Zlobec I, et
al: Synaptic proximity enables NMDAR signalling to promote brain
metastasis. Nature. 573:526–531. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Jiang SH, Hu LP, Wang X, Li J and Zhang
ZG: Neurotransmitters: Emerging targets in cancer. Oncogene.
39:503–515. 2020. View Article : Google Scholar
|
|
82
|
Liu YZ, Liu WX and Deng WH: Advances in
the study of the relationship between neurotransmitters and gastric
cancer. World J Gastroenterol. 31:1137932025. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Mancusi R and Monje M: The neuroscience of
cancer. Nature. 618:467–479. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Schaal C and Chellappan SP:
Nicotine-mediated cell proliferation and tumor progression in
smoking-related cancers. Mol Cancer Res. 12:14–23. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Nilsson MB, Sun H, Diao L, Tong P, Liu D,
Li L, Fan Y, Poteete A, Lim SO, Howells K, et al: Stress hormones
promote EGFR inhibitor resistance in NSCLC: Implications for
combinations with beta-blockers. Sci Transl Med. 9:eaao43072017.
View Article : Google Scholar
|
|
86
|
Zhao Q, Gu X, Zhang C, Lu Q, Chen H and Xu
L: Blocking M2 muscarinic receptor signaling inhibits tumor growth
and reverses epithelial-mesenchymal transition (EMT) in non-small
cell lung cancer (NSCLC). Cancer Biol Ther. 16:634–643. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Chen W, Yu J, Ge S, Yuan T, Ding X, Yan B,
Ye L, Wang Y and Jia R: Neurotransmitters in tumors: Chemical
cross-talk shaping tumor progression. Biomark Res. 13:1252025.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Friedman JR, Richbart SD, Merritt JC,
Brown KC, Nolan NA, Akers AT, Lau JK, Robateau ZR, Miles SL and
Dasgupta P: Acetylcholine signaling system in progression of lung
cancers. Pharmacol Ther. 194:222–254. 2019. View Article : Google Scholar
|
|
89
|
Emery AC, Xu W, Eiden MV and Eiden LE:
Guanine nucleotide exchange factor Epac2-dependent activation of
the GTP-binding protein Rap2A mediates cAMP-dependent growth arrest
in neuroendocrine cells. J Biol Chem. 292:12220–12231. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Chio CC, Chang YH, Hsu YW, Chi KH and Lin
WW: PKA-dependent activation of PKC, p38 MAPK and IKK in
macrophage: Implication in the induction of inducible nitric oxide
synthase and interleukin-6 by dibutyryl cAMP. Cell Signal.
16:565–575. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Zhao Q, Yue J, Zhang C, Gu X, Chen H and
Xu L: Inactivation of M2 AChR/NF-κB signaling axis reverses
epithelial-mesenchymal transition (EMT) and suppresses migration
and invasion in non-small cell lung cancer (NSCLC). Oncotarget.
6:29335–29346. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Czyżykowski R, Połowinczak-Przybyłek J and
Potemski P: Nicotine-induced resistance of non-small cell lung
cancer to treatment-possible mechanisms. Postepy Hig Med Dosw
(Online). 70:186–193. 2016. View Article : Google Scholar
|
|
93
|
Kruglov O, Vats K, Soman V, Tyurin VA,
Tyurina YY, Wang J, Williams L, Zhang J, Donahue Carey C, Jaklitsch
E, et al: Melanoma-associated repair-like Schwann cells suppress
anti-tumor T-cells via 12/15-LOX/COX2-associated eicosanoid
production. Oncoimmunology. 12:21920982023. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Deborde S, Gusain L, Powers A, Marcadis A,
Yu Y, Chen CH, Frants A, Kao E, Tang LH, Vakiani E, et al:
Reprogrammed schwann cells organize into dynamic tracks that
promote pancreatic cancer invasion. Cancer Discov. 12:2454–2473.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Huang L, Qi G, Chen G, Duan J, Dai C, Lu Y
and Zhou Q: Tumor-associated Schwann cells as new therapeutic
target in non-neurological cancers. Cancer Lett. 624:2177482025.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Shalabi S, Belayachi A and Larrivée B:
Involvement of neuronal factors in tumor angiogenesis and the
shaping of the cancer microenvironment. Front Immunol.
15:12846292024. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Ferraguti G, Terracina S, Tarani L,
Fanfarillo F, Allushi S, Caronti B, Tirassa P, Polimeni A,
Lucarelli M, Cavalcanti L, et al: Nerve growth factor and the role
of inflammation in tumor development. Curr Issues Mol Biol.
46:965–989. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Xu L, Chen S, Fu Y, Zhou T, Yu J, Li J and
Chen W: Neuro-immune-tumor axis in gliomas: A review of mechanisms,
models, and translational opportunities. Front Immunol.
16:16823222025. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Eftimiadi G, Soligo M, Manni L, Di Giuda
D, Calcagni ML and Chiaretti A: Topical delivery of nerve growth
factor for treatment of ocular and brain disorders. Neural Regen
Res. 16:1740–1750. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Lee J, Jiffar T and Kupferman ME: A novel
role for BDNF-TrkB in the regulation of chemotherapy resistance in
head and neck squamous cell carcinoma. PLoS One. 7:e302462012.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Nakazawa T, Tamai M and Mori N:
Brain-derived neurotrophic factor prevents axotomized retinal
ganglion cell death through MAPK and PI3K signaling pathways.
Invest Ophthalmol Vis Sci. 43:3319–3326. 2002.PubMed/NCBI
|
|
102
|
Yuan Y, Ye HQ and Ren QC: Upregulation of
the BDNF/TrKB pathway promotes epithelial-mesenchymal transition,
as well as the migration and invasion of cervical cancer. Int J
Oncol. 52:461–472. 2018.PubMed/NCBI
|
|
103
|
Chen B, Liang Y, He Z, An Y, Zhao W and Wu
J: Autocrine activity of BDNF induced by the STAT3 signaling
pathway causes prolonged TrkB activation and promotes human
non-small-cell lung cancer proliferation. Sci Rep. 6:304042016.
View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Chakravarthy R, Mnich K and Gorman AM:
Nerve growth factor (NGF)-mediated regulation of p75(NTR)
expression contributes to chemotherapeutic resistance in triple
negative breast cancer cells. Biochem Biophys Res Commun.
478:1541–1547. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Teng HK, Teng KK, Lee R, Wright S, Tevar
S, Almeida RD, Kermani P, Torkin R, Chen ZY, Lee FS, et al: ProBDNF
induces neuronal apoptosis via activation of a receptor complex of
p75NTR and sortilin. J Neurosci. 25:5455–5463. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Coveñas R and Muñoz M: Involvement of the
substance p/neurokinin-1 receptor system in cancer. Cancers
(Basel). 14:35392022. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Sigorski D, Sejda A, Abualsaud N, Krawczyk
E, Izycka-Swieszewska E and Kitlinska J: Neuropeptide Y in
cancer-biological functions and potential clinical implications.
Cancer Metastasis Rev. 44:212025. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Dong Z, Wang Y and Jin W: The neuroscience
of cancer: Focus on neuropeptidergic systems. Acta Pharm Sin B.
15:2323–2350. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Mashaghi A, Marmalidou A, Tehrani M, Grace
PM, Pothoulakis C and Dana R: Neuropeptide substance P and the
immune response. Cell Mol Life Sci. 73:4249–4264. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Fiebich BL, Schleicher S, Butcher RD,
Craig A and Lieb K: The neuropeptide substance P activates p38
mitogen-activated protein kinase resulting in IL-6 expression
independently from NF-kappa B. J Immunol. 165:5606–5611. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Garnier A, Vykoukal J, Hubertus J, Alt E,
von Schweinitz D, Kappler R, Berger M and Ilmer M: Targeting the
neurokinin-1 receptor inhibits growth of human colon cancer cells.
Int J Oncol. 47:151–160. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Wu Z, Martinez-Fong D, Trédaniel J and
Forgez P: Neurotensin and its high affinity receptor 1 as a
potential pharmacological target in cancer therapy. Front
Endocrinol (Lausanne). 3:1842013. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Alifano M, Souazé F, Dupouy S,
Camilleri-Broët S, Younes M, Ahmed-Zaïd SM, Takahashi T,
Cancellieri A, Damiani S, Boaron M, et al: Neurotensin receptor 1
determines the outcome of non-small cell lung cancer. Clin Cancer
Res. 16:4401–4410. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Zheng Y, Li L, Shen Z, Wang L, Niu X, Wei
Y, Sun S and Zhao J: Mechanisms of neural infiltration-mediated
tumor metabolic reprogramming impacting immunotherapy efficacy in
non-small cell lung cancer. J Exp Clin Cancer Res. 43:2842024.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Winkler F, Venkatesh HS, Amit M, Batchelor
T, Demir IE, Deneen B, Gutmann DH, Hervey-Jumper S, Kuner T,
Mabbott D, et al: Cancer neuroscience: State of the field, emerging
directions. Cell. 186:1689–1707. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Sloan EK: Harnessing brain-body
communication to understand cancer. Genes Dev. 38:820–822. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Chen X, Geng Y, Wei G, He D, Lv J, Wen W,
Xiang F, Tao K and Wu C: Neural circuitries between the brain and
peripheral solid tumors. Cancer Res. 84:3509–3521. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Chen MM, Gao Q, Ning H, Chen K, Gao Y, Yu
M, Liu CQ, Zhou W, Pan J, Wei L, et al: Integrated single-cell and
spatial transcriptomics uncover distinct cellular subtypes involved
in neural invasion in pancreatic cancer. Cancer Cell.
43:1656–1676.e10. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Zhang L, Xie J, Dai W, Lu B and Yi S:
Schwann cells in regeneration and cancer. Front Pharmacol.
16:15065522025. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Bunimovich YL, Keskinov AA, Shurin GV and
Shurin MR: Schwann cells: A new player in the tumor
microenvironment. Cancer Immunol Immunother. 66:959–968. 2017.
View Article : Google Scholar :
|
|
121
|
Stratton JA, Shah PT, Kumar R, Stykel MG,
Shapira Y, Grochmal J, Guo GF, Biernaskie J and Midha R: The
immunomodulatory properties of adult skin-derived precursor Schwann
cells: Implications for peripheral nerve injury therapy. Eur J
Neurosci. 43:365–375. 2016. View Article : Google Scholar
|
|
122
|
Zhang Y, Yuan Y, Wang Y, Ye Z, Liu T, Lv G
and Chen G: The bridging role of schwann cells in the interaction
between tumors and the nervous system: A potential target for
cancer therapy. Mol Cancer Res. 23:494–502. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Glenn TD and Talbot WS: Signals regulating
myelination in peripheral nerves and the Schwann cell response to
injury. Curr Opin Neurobiol. 23:1041–1048. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Taveggia C: Schwann cells-axon interaction
in myelination. Curr Opin Neurobiol. 39:24–29. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Ebrahim NAA, Soliman SMA, Othman MO and
Tahoun NS: Molecular mechanisms and clinical significance of
perineural invasion in malignancies: The pivotal role of
tumor-associated Schwann cells in cancer progression and
metastasis. Med Oncol. 42:1712025. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Deborde S and Wong RJ: How Schwann cells
facilitate cancer progression in nerves. Cell Mol Life Sci.
74:4405–4420. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Abd GM, Laird MC, Ku JC and Li Y:
Hypoxia-induced cancer cell reprogramming: A review on how cancer
stem cells arise. Front Oncol. 13:12278842023. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Chen SH, Zhang BY, Zhou B, Zhu CZ, Sun LQ
and Feng YJ: Perineural invasion of cancer: A complex crosstalk
between cells and molecules in the perineural niche. Am J Cancer
Res. 9:1–21. 2019.PubMed/NCBI
|
|
129
|
Cao Y: Neural is fundamental: neural
stemness as the ground state of cell tumorigenicity and
differentiation potential. Stem Cell Rev Rep. 18:37–55. 2022.
View Article : Google Scholar
|
|
130
|
Cervantes-Villagrana RD, Albores-García D,
Cervantes-Villagrana AR and García-Acevez SJ: Tumor-induced
neurogenesis and immune evasion as targets of innovative
anti-cancer therapies. Signal Transduct Target Ther. 5:992020.
View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Ouyang S, Shi S, Ding W, Ge Y, Su Y, Mo J,
Peng K, Zhang Q, Liu G, Xiao W, et al: Neuropeptide precursor VGF
promotes liver metastatic colonization of Gαq mutant uveal melanoma
by facilitating tumor microenvironment via paracrine loops. Adv Sci
(Weinh). 11:e24079672024. View Article : Google Scholar
|
|
132
|
Rotow J and Bivona TG: Understanding and
targeting resistance mechanisms in NSCLC. Nat Rev Cancer.
17:637–658. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Garramona FT, Cunha TF, Vieira JS, Borges
G, Santos G, de Castro G, Ugrinowitsch C and Brum PC: Increased
sympathetic nervous system impairs prognosis in lung cancer
patients: A scoping review of clinical studies. Lung Cancer Manag.
12:LMT632024. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Lei Z, Yang W and Zuo Y: Beta-blocker and
survival in patients with lung cancer: A meta-analysis. PLoS One.
16:e02457732021. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Daher C, Vimeux L, Stoeva R, Peranzoni E,
Bismuth G, Wieduwild E, Lucas B, Donnadieu E, Bercovici N,
Trautmann A and Feuillet V: Blockade of β-Adrenergic receptors
improves CD8+ T-cell priming and cancer vaccine
efficacy. Cancer Immunol Res. 7:1849–1863. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Tokunaga K, Fujiwara Y, Omori R, Sato T,
Ahluwalia MS and Mukherjee S: The efficacy and safety of
beta-blockers and immune checkpoint inhibitors in patients with
cancer: A systematic review and meta-analysis. Target Oncol.
20:893–905. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Bian W, Chen Y, Ni Y, Lv B, Gong B, Zhu K,
Gao W, Zeng L, Lu W and Zhang B: Efficacy of GluN2B-Containing NMDA
receptor antagonist for antitumor and antidepressant therapy in
non-small cell lung cancer. Eur J Pharmacol. 980:1768602024.
View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Tabaee Damavandi P, Pasini F, Fanella G,
Cereda GS, Mainini G, DiFrancesco JC, Trinka E and Lattanzi S:
Perampanel in brain tumor-related epilepsy: A systematic review.
Brain Sci. 13:3262023. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Silk AW, Saraiya B, Groisberg R, Chan N,
Spencer K, Girda E, Shih W, Palmeri M, Saunders T, Berman RM, et
al: A phase Ib dose-escalation study of troriluzole (BHV-4157), an
oral glutamatergic signaling modulator, in combination with
nivolumab in patients with advanced solid tumors. Eur J Med Res.
27:1072022. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Jimenez-Andrade JM, Ghilardi JR,
Castañeda-Corral G, Kuskowski MA and Mantyh PW: Preventive or late
administration of anti-NGF therapy attenuates tumor-induced nerve
sprouting, neuroma formation, and cancer pain. Pain. 152:2564–2574.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Zhu Z, Friess H, diMola FF, Zimmermann A,
Graber HU, Korc M and Büchler MW: Nerve growth factor expression
correlates with perineural invasion and pain in human pancreatic
cancer. J Clin Oncol. 17:2419–2428. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Acevedo DS, Fang WB, Rao V, Penmetcha V,
Leyva H, Acosta G, Cote P, Brodine R, Swerdlow R, Tan L, et al:
Regulation of growth, invasion and metabolism of breast ductal
carcinoma through CCL2/CCR2 signaling interactions with MET
receptor tyrosine kinases. Neoplasia. 28:1007912022. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Xu M, Wang Y, Xia R, Wei Y and Wei X: Role
of the CCL2-CCR2 signalling axis in cancer: Mechanisms and
therapeutic targeting. Cell Prolif. 54:e131152021. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Jiang J, Han D, Wang J, Wen W, Zhang R and
Qin W: Neuroendocrine transdifferentiation in human cancer:
Molecular mechanisms and therapeutic targets. MedComm (2020).
5:e7612024. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Schmitt M, Bohnenberger H, Bartsch DK,
Wagner DC, Litmeyer AS, Grass A, Rinke A, Koch C, Kremer M, Evert
M, et al: DLL3 expression in neuroendocrine carcinomas and
neuroendocrine tumours: Insights from a multicentric cohort of 1294
pulmonary and extrapulmonary neuroendocrine neoplasms. Endocr
Pathol. 36:92025. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Minocha M, Thompson CG, Murphy A, Zhou Y,
Brandl C, Parkes A, Chen X, Yu B, Martinez P and Houk BE:
Pharmacokinetics of tarlatamab, a delta-like ligand-3 (DLL3)
targeted half-life extended bispecific T-cell engager
(BiTE®) immunotherapy in adult patients with previously
treated small-cell lung cancer: Results from DeLLphi-300, a phase I
multiple-dose-escalation study. Clin Pharmacokinet. 63:1757–1768.
2024. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Lin S, Zhang Y, Yao J, Yang J, Qiu Y, Zhu
Z and Hua H: DB-1314, a novel DLL3-targeting ADC with DNA
topoisomerase I inhibitor, exhibits promising safety profile and
therapeutic efficacy in preclinical small cell lung cancer models.
J Transl Med. 22:7662024. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Stelzer KJ: Epidemiology and prognosis of
brain metastases. Surg Neurol Int. 4(Suppl 4): S192–S202. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Osswald M, Jung E, Sahm F, Solecki G,
Venkataramani V, Blaes J, Weil S, Horstmann H, Wiestler B, Syed M,
et al: Brain tumour cells interconnect to a functional and
resistant network. Nature. 528:93–98. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Gao R, Zhou R, Chen X, Gao L, Du J, Chen
Z, Chang L, Song Y, Wu Y and Li H: Inhibition of astrocytic AT1R
ameliorates sleep deprivation induced Aβ deposition and glymphatic
dysfunction via the MAPK/Cx43 pathway. Sleep Med. 136:1068472025.
View Article : Google Scholar
|
|
151
|
Chen Q, Boire A, Jin X, Valiente M, Er EE,
Lopez-Soto A, Jacob L, Patwa R, Shah H, Xu K, et al:
Carcinoma-astrocyte gap junctions promote brain metastasis by cGAMP
transfer. Nature. 533:493–498. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Neves V, Aires-da-Silva F, Morais M, Gano
L, Ribeiro E, Pinto A, Aguiar S, Gaspar D, Fernandes C, Correia JDG
and Castanho MARB: Novel peptides derived from dengue virus capsid
protein translocate reversibly the blood-brain barrier through a
receptor-free mechanism. ACS Chem Biol. 12:1257–1268. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Patel M, Panja S, Zaman LA, Yeapuri P,
Bhattarai S, Gorantla S, Chang L, Heredia A, Walczak P, Hanson B,
et al: CCR5-ligand decorated rilpivirine lipid-based nanoparticles
for sustained antiretroviral responses. Nat Commun. 16:5132025.
View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Haroon J, Aboody K, Flores L, McDonald M,
Mahdavi K, Zielinski M, Jordan K, Rindner E, Surya J, Venkatraman
V, et al: Use of transcranial low-intensity focused ultrasound for
targeted delivery of stem cell-derived exosomes to the brain. Sci
Rep. 13:177072023. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Zhao P, Wu T, Tian Y, You J and Cui X:
Recent advances of focused ultrasound induced blood-brain barrier
opening for clinical applications of neurodegenerative diseases.
Adv Drug Deliv Rev. 209:1153232024. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Chen Y, Liu H, Zhang L, Zeng H, Jiang L,
Qin Q, Li D and Lu G: Glutamate molecular structure and protein
affect the inhibition of breast cancer cell metastasis:
Cell-derived exosomes inhibitory effects through the MAPK signaling
pathway. Int J Biol Macromol. 300:1402642025. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Sui XY, Cao SW, Song XQ, Liu XY, Chen C,
Yan Q, Wang ZQ, Zhang WJ, Ma LX, Jin X, et al: MORF4L2 induces
immunosuppressive microenvironment and immunotherapy resistance
through GRHL2/MORF4L2/H4K12Ac/CSF1 axis in triple-negative breast
cancer. Biomark Res. 13:62025. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Zhou C, Yang Y, Cui H, Li S, Wang Z, Chen
L, Feng M, Li D, Chen X, Hao B, et al: SEC61G facilitates brain
metastases via antagonizing PGAM1 ubiquitination and immune
microenvironment remodeling in non-small cell lung cancer. Int J
Biol Sci. 21:1436–1458. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Stuart T and Satija R: Integrative
single-cell analysis. Nat Rev Genet. 20:257–272. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Krishna S, Lowery FJ, Copeland AR,
Bahadiroglu E, Mukherjee R, Jia L, Anibal JT, Sachs A, Adebola SO,
Gurusamy D, et al: Stem-like CD8 T cells mediate response of
adoptive cell immunotherapy against human cancer. Science.
370:1328–1334. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Meng F, Sun Y, Li J and Xu L: Single-cell
and spatial transcriptome landscapes reveal the spatial immune
defense pattern shared by primary and brain metastatic lung
adenocarcinoma. J Thorac Dis. 17:9927–9942. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Wang Z, Wang Y, Chang M, Wang Y, Liu P, Wu
J, Wang G, Tang X, Hui X, Liu P, et al: Single-cell transcriptomic
analyses provide insights into the cellular origins and drivers of
brain metastasis from lung adenocarcinoma. Neuro Oncol.
25:1262–1274. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Yap A, Lopez-Olivo MA, Dubowitz J, Pratt
G, Hiller J, Gottumukkala V, Sloan E, Riedel B and Schier R: Effect
of beta-blockers on cancer recurrence and survival: A meta-analysis
of epidemiological and perioperative studies. Br J Anaesth.
121:45–57. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Beltran H, Rickman DS, Park K, Chae SS,
Sboner A, MacDonald TY, Wang Y, Sheikh KL, Terry S, Tagawa ST, et
al: Molecular characterization of neuroendocrine prostate cancer
and identification of new drug targets. Cancer Discov. 1:487–495.
2011. View Article : Google Scholar
|