Introduction
Tumor cells with tumorigenic potential constitute
small populations, referred to as cancer-initiating cells (CICs),
which have the ability to self-renew and are multipotent. CICs are
found in a number of tumors (1–12). As
CICs efficiently eliminate antitumor chemicals, are resistant to
radiotherapy and degrade reactive oxygen species (ROS), they are
considered to cause cancer recurrence and/or metastasis (13–16).
Hodgkin's lymphoma (HL) is a monoclonal lymphoid
neoplasm diagnosed by the presence of multinucleated
(Reed-Sternberg; RS) cells admixed with singly nucleated Hodgkin
cells (tumor cells) and various inflammatory cells. Although the
cellular origin of HL remains controversial, recent molecular
biological studies have revealed that HL is principally a B-cell
neoplasm (17,18). The CICs of HL have not been
extensively investigated. Jones et al, in a study on HL
patients, described a small circulating clonotypic B-cell
population expressing high levels of aldehyde dehydrogenase and
CD27; these cells efficiently formed colonies in vitro
(19). Nakashima et al
investigated ‘side populations’ (SPs), which were negative for
CD133 and CD44, but exhibited the characteristics of CICs, in
several tumors. The SPs consisted of small mononuclear cells and
those of HL were resistant to chemotherapeutic agents (20). We recently demonstrated that
intracellular ROS levels were lower in a proportion of singly
nucleated HL cells compared with those in the RS cells of HL. The
ROS-low cells exhibited anti-apoptotic and tumorigenic potential,
similar to CICs, and abundantly expressed Forkhead box O3a
(FoxO3a), a transcription factor regulating the expression of genes
encoding ROS-degrading enzymes. These results suggested that these
FoxO3a-expressing cells may constitute the CICs of HL (3,21,22).
Inhibitor of DNA-binding (ID) proteins constitute a
family of highly conserved transcriptional regulators that play
pivotal roles during development and in the maintenance of adult
tissue homeostasis. The major biological effects of ID proteins are
inhibition of differentiation and maintenance of the self-renewal
ability and multipotency of stem cells (23). ID proteins are overexpressed in a
number of human cancers; deregulation of ID protein expression is
directly involved in cancer initiation, maintenance, progression
and development of drug resistance (characteristics of CICs). ID1
transcription is negatively regulated by FoxO3a in leukemic cells
(24). In nasopharyngeal carcinoma,
Epstein-Barr virus (EBV)-encoded latent membrane protein (LMP) 1
phosphorylates and inactivates FoxO3a, thereby upregulating ID1
(25–27). These results suggest that the
pathogenetic progression of certain tumors may be controlled by
these three factors, namely ID1, FoxO3a and LMP1. EBV has been
hypothesized to play a role in the pathogenesis of HL, and
EBV-positive HL tissue expresses LMP1. To the best of our
knowledge, no study has yet investigated the associations among
ID1, FoxO3a and LMP1 in HL. The aim of the present study was to
address this topic.
Materials and methods
Patients
This study received ethical approval from the
Institutional Review Board of Osaka University (12467–2). From 1999
to 2015, informed consent was obtained from 31 HL patients who were
included in this study. Formalin-fixed paraffin-embedded (FFPE)
diagnostic samples were stored in the dark room of the Department
of Pathology of Osaka University Hospital at room temperature, and
4-µm sections were obtained prior to staining with hematoxylin and
eosin, followed by routine immunohistochemical evaluation.
Immunohistochemical analysis of
FoxO3a, ID1 and LMP1 expression
Anti-FoxO3a (rabbit monoclonal, cat. no. 12,829,
Cell Signaling Technology Inc., Beverly, MA, USA), anti-ID1 (rabbit
monoclonal, ab134163, Abcam Ltd., Cambridge, UK) and anti-LMP1
(mouse monoclonal, cat. no. M0897, Dako A/S, Glostrup, Denmark)
antibodies were used for immunohistochemical analysis. Antigen
retrieval was performed with the aid of a Pascal pressurized
heating chamber (Dako). The sections were incubated with
anti-FoxO3a (100-fold dilution), anti-ID1 (200-fold dilution) and
anti-LMP1 (100-fold dilution) antibodies, and color was developed
with the aid of the ChemMate EnVision kit (Dako). Diaminobenzidine
(DAB) (Dako) served as the chromogen. The negative controls
underwent all the abovementioned steps, except for incubation with
the primary antibodies. The intensity of immunohistochemical
staining was categorized as none, weak to moderate, and strong.
Weak to moderate and strong were considered as positive signals.
When signals from FoxO3a and ID1 were detected in >1% of Hodgkin
and RS cells, that case was adjudged positive, as CICs constitute a
minority of tumor cells. In addition to tumor cells, some non-tumor
cells, such as inflammatory cells, macrophages and vascular
endothelial cells, were also positive for FoxO3a and ID1, which
were considered to be positive controls (3,28). When
LMP1 expression was detected in >20% of tumor cells, that case
was adjudged as positive (EBV-positive criterion) (29). All staining data were independently
evaluated by two pathologists (J. I. and E.M.) and the evaluation
results were matched.
In situ hybridization (ISH)
ISH using an EBV-encoded small RNA (EBER) probe was
performed to determine whether the EBV genome was present in the
FFPE sections; the EBER DAB application kit (Dako) was used.
Briefly, the sections were treated with proteinase K diluted 1:10
in TBS (50 mmol/l Tris-HCl buffered saline containing 150 mmol/l
NaCl; pH 7.6) and then hybridized with the EBER peptide nucleic
acid probe tagged with fluorescein (Dako) at 55°C for 90 min. After
blocking endogenous peroxidase activity, the sections were
incubated with rabbit anti-fluorescein isothiocyanate antibody
(dilution 1:50; rabbit polyclonal, cat. no. 71-1900, Invitrogen,
Carlsbad, CA, USA) at room temperature for 30 min, followed by
incubation with the ChemMate ENVISION/HRP polymer (Dako) at room
temperature for 30 min. DAB was used for color development. As
previously suggested, when signal from EBER was evident in >20%
of tumor cells, that case was adjudged as EBV-positive (30). All staining data were independently
evaluated by two pathologists (J.I. and E.M.).
Statistical analysis
Statistical analyses were performed with the aid of
JMP software (SAS Institute Inc., Cary, NC, USA). The Chi-square
and Fisher's exact probability tests were used to assess the
correlations among three groups, namely those expressing ID1,
FoxO3a and LMP1 among HL cells. A P-value of <0.05 was
considered to reflect statistical significance.
Results
Histological subtypes of HL
A total of 31 patients diagnosed with HL were
investigated following approval of the study protocol by the
Institutional Review Board of Osaka University Hospital. The
histological subtypes were classified as 15 mixed-cellularity
classical HL (MCCHL), 13 nodular sclerosis classical HL (NSCHL), 1
lymphocyte-rich classical HL (LRCHL) and 2 nodular
lymphocyte-predominant HL (NLPHL).
Immunohistochemistry for ID1 and
FoxO3a
To determine the association between ID1 and FoxO3a
expression, ID1 and FoxO3a were immunohistochemically detected.
Tumor cells stained for ID1 (strong cytoplasmic staining) and
FoxO3a (strong nuclear staining) (Fig.
1A-D). Of the 31 cases, 10 were ID1- and 20 FoxO3a-positive
(Table I). The ID1 expression level
was inversely correlated with that of FoxO3a (Table II; P=0.00035).
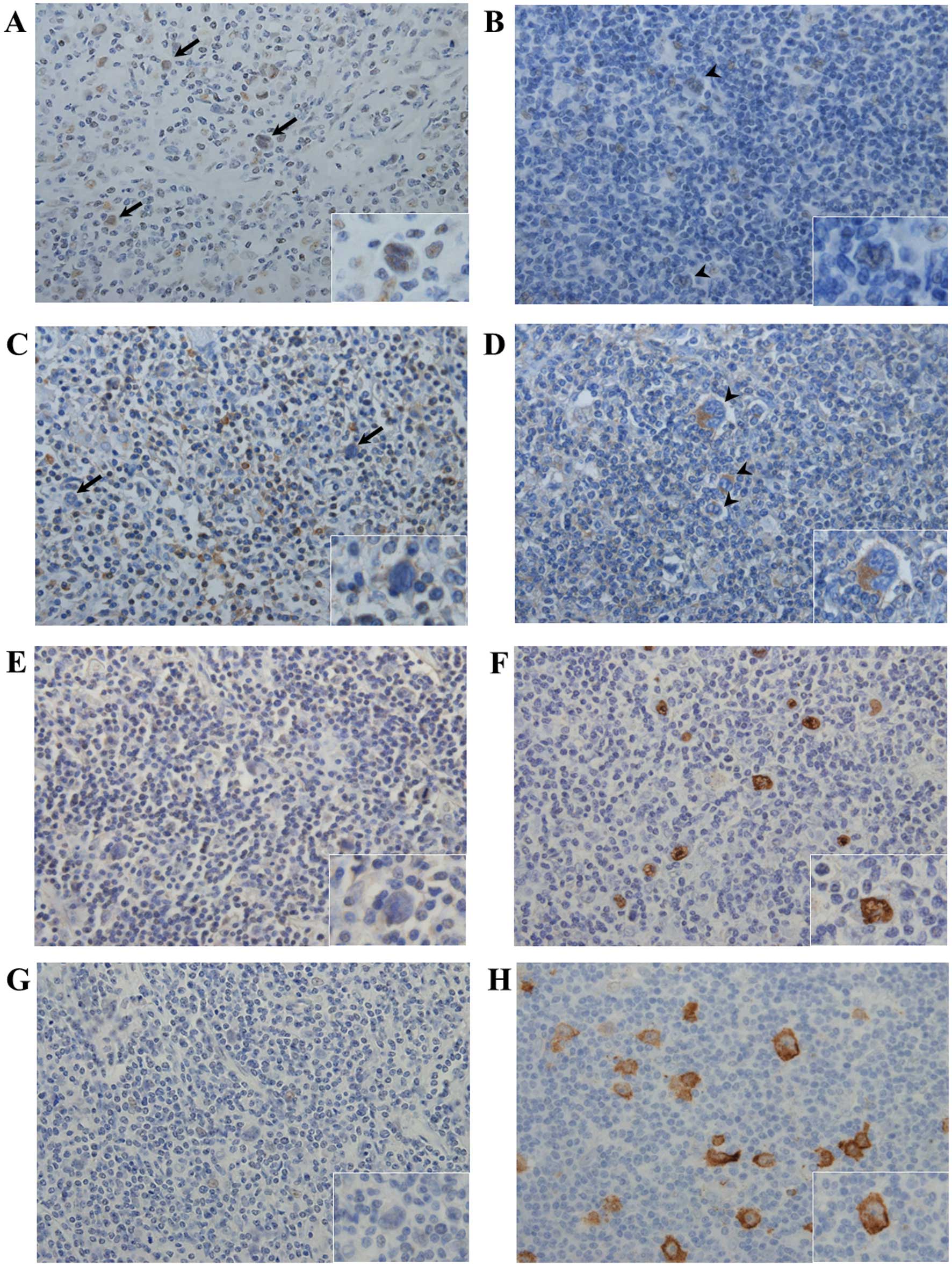 | Figure 1.FoxO3a, ID1 and LMP1 expression in HL.
Typical cases [case 4 (A, C, E and G) and case 1 (B, D, F and H)]
are shown. (A-D) Immunohistochemical detection of FoxO3a (A and B)
and ID1 (C and D). Positive FoxO3a staining was detected in a
limited number of tumor cells in case 4 (A) arrows and inset;
magnification, ×400. In case 1, tumor cells (B) arrowheads and
inset did not stain at all for FoxO3a, but certain non-tumor cells,
such as macrophages and vascular endothelial cells, stained
positive for FoxO3a (magnification, ×400). No ID1 signal was
detected from tumor cells in case 4 (C) arrows and inset;
magnification, ×400, whereas strong ID1 expression was evident in a
limited number of tumor cells in case 1 (D) arrowheads and inset;
magnification ×400. (E-H) EBER-1 in situ hybridization and
immunohistochemical staining for the detection of LMP1. No
EBER-1-positive/LMP1-expressing tumor cells were detected in case 4
(E and G), whereas tumor cells in case 1 stained positively for
both EBER-1 and LMP1 (F and H). FoxO3a, Forkhead box O3a; ID1,
inhibitor of DNA-binding protein 1; LMP1, latent membrane protein
1; EBER-1, Ebstein-Barr virus-encoded small RNA. |
 | Table I.Immunohistochemistry and ISH results
of 31 HL cases. |
Table I.
Immunohistochemistry and ISH results
of 31 HL cases.
|
| Histological
subtype | FoxO3a | ID1 | EBER-1 | LMP1 |
|---|
| Case 1 | MCCHL | − | + | + | + |
| Case 2 | MCCHL | − | + | + | + |
| Case 3 | MCCHL | − | + | + | + |
| Case 4 | MCCHL | + | − | − | − |
| Case 5 | MCCHL | + | − | − | − |
| Case 6 | MCCHL | + | − | − | − |
| Case 7 | MCCHL | + | − | + | + |
| Case 8 | MCCHL | + | − | + | + |
| Case 9 | MCCHL | + | − | + | + |
| Case 10 | MCCHL | + | + | − | − |
| Case 11 | MCCHL | − | + | + | + |
| Case 12 | MCCHL | − | + | + | + |
| Case 13 | MCCHL | − | − | + | + |
| Case 14 | MCCHL | − | − | + | + |
| Case 15 | MCCHL | − | − | − | − |
| Case 16 | NSCHL | + | − | − | − |
| Case 17 | NSCHL | + | − | − | − |
| Case 18 | NSCHL | + | − | − | − |
| Case 19 | NSCHL | + | − | − | − |
| Case 20 | NSCHL | + | − | − | − |
| Case 21 | NSCHL | + | − | − | − |
| Case 22 | NSCHL | + | − | − | − |
| Case 23 | NSCHL | + | − | − | − |
| Case 24 | NSCHL | + | − | − | − |
| Case 25 | NSCHL | + | − | − | − |
| Case 26 | NSCHL | + | − | − | − |
| Case 27 | NSCHL | + | + | − | − |
| Case 28 | NSCHL | − | + | + | + |
| Case 29 | LRCHL | − | + | + | + |
| Case 30 | NLPHL | − | + | − | − |
| Case 31 | NLPHL | + | − | + | + |
| Table II.Correlation between FoxO3a and ID1
expression in HL cases. |
Table II.
Correlation between FoxO3a and ID1
expression in HL cases.
|
| ID1 |
|
|---|
|
|
|
|
|---|
| Variable | (−) | (+) | P-value |
|---|
| FoxO3a |
|
|
|
| (−) | 3 | 8 | 0.00035 |
| (+) | 18 | 2 |
|
ISH for the detection of EBER-1 and
immunohistochemistry for LMP1
EBER-1 ISH was performed to determine whether the HL
cases were infected with EBV (Fig.
1E-H). Thirteen of the 31 cases were EBV-positive, and all were
also LMP1-positive, indicating that the EBV latent gene expression
pattern was of type II (Table I). By
histological subtype, 10 of 15 MCCHL (66.7%), 1 of 13 NSCHL (7.7%),
1 of 1 LRCHL (100%) and 1 of 2 NLPHL (50%) patients had EBER-1- and
LMP1-positive tumor cells. The EBV-positive rates were consistent
with those of previous studies, particularly in MCCHL and NSCHL
(29).
Correlation between the expression of
LMP1 and ID1, and that of LMP1 and FoxO3a
To compare the expression of LMP1 with that of ID1
and FoxO3a, our data were subjected to Chi-square testing. The
expressions of LMP1 and ID1 were significantly associated (Table III; P=0.029), but LMP1 and FoxO3a
were not (Table III;
P=0.00085).
| Table III.Correlation among LMP1, ID1 and
FoxO3a expression in HL cases. |
Table III.
Correlation among LMP1, ID1 and
FoxO3a expression in HL cases.
|
| ID1 |
| FoxO3a |
|
|---|
|
|
|
|
|
|
|---|
| Variable | (−) | (+) | P-value | (−) | (+) | P-value |
|---|
| LMP1 |
|
|
(−) | 15 | 3 | 0.029 | 2 | 16 | 0.00085 |
|
(+) | 6 | 7 |
| 9 | 4 |
|
Discussion
The ID protein family contains four members; the ID
proteins maintain the self-renewal potential and multipotency of
stem cells. Overexpression of ID proteins has been reported in a
number of tumors, and such deregulation is associated with tumor
initiation and drug resistance (23,31). ID1
expression was found to be an independent (negative) prognostic
marker for nasopharyngeal carcinoma patients (32). In HL, ID2 interacts with the
retinoblastoma tumor suppressor protein to repress B-cell-specific
gene expression via inactivation of E2A (33,34). To
the best of our knowledge, no study to date has investigated the
role played by ID1 in HL. We herein addressed this issue, and found
that one-third of HL patients (10 of 31) expressed ID1. No
association was found between ID1 expression and histological HL
subtype (P=0.21, data not shown). ID1 and FoxO3a expressions were
inversely correlated (P=0.00035). Birkenkamp et al reported
that, in leukemia patients, ID1 transcription was negatively
regulated by FoxO3a (24). This may
also be the case in HL patients.
CIC detection is crucial for the development of
novel, targeted cancer therapies. CICs may eliminate ROS (14). In previous studies, we observed that
FoxO3a, a transcription factor upregulating genes encoding
ROS-degrading enzymes, was abundantly expressed in the ROS-low
regions of spreads of singly nucleated cells that resembled Hodgkin
cells, but not in the RS cells of HL. Gold standard markers for HL
are lacking, in contrast to the CD133 marker of brain and colon
carcinomas and mantle cell lymphoma in animal models. Our results
suggest that the FoxO3a-expressing cells may be the CICs of HL
(3,5,8,20–22,35).
However, we found that several HL cases were negative for FoxO3a,
and that the majority of FoxO3a-negative cases were EBV-positive (9
of 11). In nasopharyngeal carcinoma patients, EBV-encoded LMP1
inactivates FoxO3a (25–27). Similar to nasopharyngeal carcinoma,
FoxO3a appeared to be degraded by LMP1 in HL tissue. The HL cell
lines used in previous studies were EBV-negative, and populations
of FoxO3a-positive cells were evident (21,22).
Therefore, FoxO3a may be a marker of the CICs of EBV-negative HL.
The cells that eliminate ROS among the cells of EBV-positive HL
remain to be identified. EBV-positive HL tissue expresses ID1 at
high levels. Upon EBV infection, EBV-encoded LMP1 induces
phosphorylation and inactivation of FoxO3a, which is associated
with upregulation of ID1 (25–27). The
ID proteins control various genes involved in tumor initiation and
progression. For example, combined expression of ID1 and ID3
increased self-renewal and promoted tumor initiation by colon CICs
via downregulation of p21 (31).
Therefore, ID1 may be a useful marker of the CICs of EBV-positive
HL.
Recent studies have demonstrated that aberrant
levels of ID proteins are associated with the upregulation of
pro-survival and anti-apoptotic factors, including nuclear
factor-κB (NF-κB), B-cell lymphoma-2 and phosphoinositide
3-kinase-AKT, in several tumors (23,32,36).
Upon constitutive activation, NF-κB promotes proliferation and
abrogates apoptosis of the Hodgkin and RS cells of HL (37,38).
Therefore, high-level ID1 expression in cases of EBV-positive HL
may play an important role in the pathogenesis of the disease, via
the action of NF-κB. However, the correlation between NF-κB
activation and EBV-infected conditions was controversial, and
further studies are required (39).
In conclusion, the ID1 and FoxO3a expression levels
in clinical samples from HL patients were found to be inversely
correlated. LMP1 (EBV)-positive cases were usually FoxO3a-negative
and ID1-positive. Our previous suggestion that FoxO3a may be a
marker of CICs may not be applicable in cases of EBV-positive HL.
Further studies are required to elucidate whether ID1-positive
cells exhibit CIC-like characteristics.
Acknowledgements
The authors would like to thank Mr. Masaharu Kohara,
Ms. Etsuko Maeno and Ms. Takako Sawamura for their valuable
technical assistance. This study was supported by grants from the
Ministry of Education, Culture, Sports, Science and Technology,
Japan (nos. 25108507, 25460435, 15H00894 and T264604700).
References
|
1
|
Al-Hajj M, Wicha MS, Benito-Hernandez A,
Morrison SJ and Clarke MF: Prospective identification of
tumorigenic breast cancer cells. Proc Natl Acad Sci USA.
100:3983–3988. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bonnet D and Dick JE: Human acute myeloid
leukemia is organized as a hierarchy that originates from a
primitive hematopoietic cell. Nat Med. 3:730–737. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ikeda J, Tian T, Wang Y, Hori Y, Honma K,
Wada N and Morii E: Expression of FoxO3a in clinical cases of
malignant lymphoma. Pathol Res Pract. 209:716–720. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Matsui W, Wang Q, Barber JP, Brennan S,
Smith BD, Borrello I, McNiece I, Lin L, Ambinder RF, Peacock C, et
al: Clonogenic multiple myeloma progenitors, stem cell properties,
and drug resistance. Cancer Res. 68:190–197. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
O'Brien CA, Pollett A, Gallinger S and
Dick JE: A human colon cancer cell capable of initiating tumor
growth in immunodeficient mice. Nature. 445:106–110. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rahadiani N, Ikeda J, Mamat S, Matsuzaki
S, Ueda Y, Umehara R, Tian T, Wang Y, Enomoto T, Kimura T, et al:
Expression of aldehyde dehydrogenase 1 (ALDH1) in endometrioid
adenocarcinoma and its clinical implications. Cancer Sci.
102:903–908. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer, and cancer stem cells. Nature.
414:105–111. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ricci-Vitiani L, Lombardi DG, Pilozzi E,
Biffoni M, Todaro M, Peschle C and De Maria R: Identification and
expansion of human colon-cancer-initiating cells. Nature.
445:111–115. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Singh SK, Hawkins C, Clarke ID, Squire JA,
Bayani J, Hide T, Henkelman RM, Cusimano MD and Dirks PB:
Identification of human brain tumour initiating cells. Nature.
432:396–401. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sunayama J, Sato A, Matsuda K, Tachibana
K, Watanabe E, Seino S, Suzuki K, Narita Y, Shibui S, Sakurada K,
et al: FoxO3a functions as a key integrator of cellular signals
that control glioblastoma stem-like cell differentiation and
tumorigenicity. Stem Cells. 29:1327–1337. 2011.PubMed/NCBI
|
|
11
|
Touil Y, Zuliani T, Wolowczuk I, Kuranda
K, Prochazkova J, Andrieux J, Le Roy H, Mortier L, Vandomme J, Jouy
N, et al: The PI3K/AKT signaling pathway controls the quiescence of
the low-Rhodamine123-retention cell compartment enriched for
melanoma stem cell activity. Stem Cells. 31:641–651. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yin S, Li J, Hu C, Chen X, Yao M, Yan M,
Jiang G, Ge C, Xie H, Wan D, et al: CD133 positive hepatocellular
carcinoma cells possess high capacity for tumorigenicity. Int J
Cancer. 120:1444–1450. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Coffer PJ and Burgering BM: Stressed
marrow: FoxOs stem tumour growth. Nat Cell Biol. 9:251–253. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Diehn M, Cho RW, Lobo NA, Kalisky T, Dorie
MJ, Kulp AN, Qian D, Lam JS, Ailles LE, Wong M, et al: Association
of reactive oxygen species levels and radioresistance in cancer
stem cells. Nature. 458:780–783. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Santamaría CM, Chillón MC, García-Sanz R,
Pérez C, Caballero MD, Ramos F, de Coca AG, Alonso JM, Giraldo P,
Bernal T, et al: High FoxO3a expression is associated with a poorer
prognosis in AML with normal cytogenetics. Leuk Res. 33:1706–1709.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tothova Z, Kollipara R, Huntly BJ, Lee BH,
Castrillon DH, Cullen DE, McDowell EP, Lazo-Kallanian S, Williams
IR, Sears C, et al: FoxOs are critical mediators of hematopoietic
stem cell resistance to physiologic oxidative stress. Cell.
128:325–339. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kanzler H, Küppers R, Hansmann ML and
Rajewsky K: Hodgkin and Reed-Sternberg cells in Hodgkin's disease
represent the outgrowth of a dominant tumor clone derived from
(crippled) germinal center B cells. J Exp Med. 184:1495–1505. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Marofioti T, Hummel M, Foss HD, Laumen H,
Korbjuhn P, Anagnostopoulos I, Lammert H, Demel G, Theil J, Wirth T
and Stein H: Hodgkin and reed-sternberg cells represent an
expansion of single clone originating from a germinal center B-cell
with functional immunoglobulin gene rearrangements but defective
immunoglobulin transcription. Blood. 95:1443–1450. 2000.PubMed/NCBI
|
|
19
|
Jones RJ, Gocke CD, Kasamon YL, Miller CB,
Perkins B, Barber JP, Vala MS, Gerber JM, Gellert LL, Siedner M, et
al: Circulating clonotypic B cells in classic Hodgkin lymphoma.
Blood. 113:5920–5926. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nakashima M, Ishii Y, Watanabe M, Togano
T, Umezawa K, Higashihara M, Watanabe T and Horie R: The side
population, as a precursor of Hodgkin and Reed-Sternberg cells and
a target for nuclear factor-κB inhibitors in Hodgkin's lymphoma.
Cancer Sci. 101:2490–2496. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ikeda J, Mamat S, Tian T, Wang Y, Luo W,
Rahadiani N, Aozasa K and Morii E: Reactive oxygen species and
aldehyde dehydrogenase activity in Hodgkin lymphoma cells. Lab
Invest. 92:606–614. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ikeda J, Mamat S, Tian T, Wang Y,
Rahadiani N, Aozasa K and Morii E: Tumorigenic potential of
mononucleated small cells of Hodgkin lymphoma cell lines. Am J
Pathol. 177:3081–3088. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lasorella A, Benezra R and Iavarone A: The
ID proteins: Master regulators of cancer stem cells and tumour
aggressiveness. Nat Rev Cancer. 14:77–91. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Birkenkamp KU, Essafi A, van der Vos KE,
da Costa M, Hui RC, Holstege F, Koenderman L, Lam EW and Coffer PJ:
FoxO3a induces differentiation of Bcr-Abl-transformed cells through
transcriptional down-regulation of Id1. J Biol Chem. 282:2211–2220.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hau PM, Tsang CM, Yip YL, Huen MS and Tsao
SW: Id1 interacts and stabilizes the Epstein-Barr virus latent
membrane protein 1 (LMP1) in nasopharyngeal epithelial cells. PLoS
One. 6:e211762011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li HM, Zhuang ZH, Wang Q, Pang JC, Wang
XH, Wong HL, Feng HC, Jin DY, Ling MT, Wong YC, et al: Epstein-Barr
virus latent membrane protein 1 (LMP1) upregulates Id1 expression
in nasopharyngeal epithelial cells. Oncogene. 23:4488–4494. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lo AK, Dawson CW, Lo KW, Yu Y and Young
LS: Upregulation of Id1 by Epstein-Barr virus-encoded LMP1 confers
resistance to TGFbeta-mediated growth inhibition. Mol Cancer.
9:1552010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gupta GP, Perk J, Acharyya S, de Candia P,
Mittal V, Todorova-Manova K, Gerald WL, Brogi E, Benezra R and
Massagué J: ID genes mediate tumor reinitiation during breast
cancer lung metastasis. Proc Natl Acad Sci USA. 104:19506–19511.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wada N, Ikeda J, Hori Y, Fujita S, Ogawa
H, Soma T, Sugiyama H, Fukuhara S, Kanamaru A, Hino M, et al:
Epstein-barr virus in diffuse large B-cell lymphoma in
immunocompetent patients in Japan is as low as in Western
Countries. J Med Virol. 83:317–321. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hummel M, Anagnostopoulos I, Dallenbach F,
Korbjuhn P, Dimmler C and Stein H: EBV infection patterns in
Hodgkin's disease and normal lymphoid tissue: Expression and
cellular localization of EBV gene products. Br J Haematol.
82:689–694. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
O'Brien CA, Kreso A, Ryan P, Hermans KG,
Gibson L, Wang Y, Tsatsanis A, Gallinger S and Dick JE: ID1 and ID3
regulate the self-renewal capacity of human colon cancer-initiating
cells through p21. Cancer Cell. 21:777–792. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sun W, Guo MM, Han P, Lin JZ, Liang FY,
Tan GM, Li HB, Zeng M and Huang XM: Id-1 and the p65 subunit of
NF-κB promote migration of nasopharyngeal carcinoma cells and are
correlated with poor prognosis. Carcinogenesis. 33:810–817. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mathas S, Janz M, Hummel F, Hummel M,
Wollert-Wulf B, Lusatis S, Anagnostopoulos I, Lietz A, Sigvardsson
M, Jundt F, et al: Intrinsic inhibition of transcription factor E2A
by HLH proteins ABF-1 and Id2 mediates reprogramming of neoplastic
B cells in Hodgkin lymphoma. Nat Immunol. 7:207–215. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Renné C, Martin-Subero JI, Eickernjäger M,
Hansmann ML, Küppers R, Siebert R and Bräuninger A: Aberrant
expression of ID2, a suppressor of B-cell-specific gene expression,
in Hodgkin's lymphoma. Am J Pathol. 169:655–664. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Medina DJ, Abass-Shereef J, Walton K,
Goodell L, Aviv H, Strair RK and Budak-Alpdogan T: Cobblestone-area
forming cells derived from patients with mantle cell lymphoma are
enriched for CD133+ tumor-initiating cells. PLoS One.
9:e910422014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ling MT, Wang X, Ouyang XS, Xu K, Tsao SW
and Wong YC: Id-1 expression promotes cell survival through
activation of NF-kappaB signalling pathway in prostate cancer
cells. Oncogene. 22:4498–4508. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hinz M, Lemke P, Anagnostopoulos I, Hacker
C, Krappmann D, Mathas S, Dörken B, Zenke M, Stein H and
Scheidereit C: Nuclear factor kappaB-dependent gene expression
profiling of Hodgkin's disease tumor cells, pathogenetic
significance, and link to constitutive signal transducer and
activator of transcription 5a activity. J Exp Med. 196:605–617.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hinz M, Löser P, Mathas S, Krappmann D,
Dörken B and Scheidereit C: Constitutive NF-kappaB maintains high
expression of a characteristic gene network, including CD40, CD86,
and a set of antiapoptotic genes in Hodgkin/Reed-Sternberg cells.
Blood. 97:2798–2807. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Küppers R: The biology of Hodgkin's
lymphoma. Nat Rev Cancer. 9:15–27. 2009. View Article : Google Scholar : PubMed/NCBI
|














