1. Introduction
To date, studies have indicated that there is a
strong association between diabetes and heart failure. Diabetes
mellitus (DM) is one of the most common risk factors for the
development of non-ischemic heart failure, and its identification
results in the initiation of cardioprotective therapy (1). Therefore, cardiovascular complications
constitute the main cause of morbidity and mortality in diabetic
patients. In total, 80% of related deaths occur due to
cardiovascular diseases in these patients (2). These complications include
atherosclerosis, coronary artery disease, and myocardial infarction
(3). Diabetic cardiomyopathy (DCM),
is another cardiac disorder caused by diabetes (4).
DCM is defined as a heart disorder of a diabetic
patient in which abnormal myocardial structures, such as coronary
lesions, valvular heart disease and hypertension are absent. This
disorder is characterized by diastolic or systolic dysfunction,
combined with structural changes, such as fibrosis and hypertrophy
(5). The pathophysiology of DCM is
complex and involves various mechanisms, such as inflammation,
mitochondrial dysfunction, apoptosis, oxidative stress,
lipotoxicity, fibrosis and disruptions in insulin signaling
(6). Even though extensive research
has been conducted concerning diabetic cardiovascular disorders,
the efficacy of treatment remains a challenge (7,8).
Multiple contributing mechanisms are associated with
diabetic heart disease. It is well established that hyperglycemia
plays a critical role in diabetes-related heart failure (9,10).
Following a protein modification mechanism and the induction of
epigenetic alterations, as well as mitochondrial damage,
hyperglycemia results in myocardial dysfunction. Oxidative stress
is also associated with both non-cardiac and cardiac
diabetes-related complications. Oxidative stress usually provokes
impaired cardiomyocyte calcium handling and leads to reduced
cardiac contractility and relaxation. Inflammation is another
mechanism that leads to the progression of diabetes. Increased
inflammatory signaling often results in macrophage infiltration and
thus, in cardiac DM-related complications (9).
Moreover, autonomic dysfunction has an impact on
myocardial performance, and systemic and coronary vascular function
(11). Autonomic dysfunction is a
significant complication of DM that is firmly linked to a roughly
five-fold increased risk of cardiovascular mortality. Its
presentation ranges from resting tachycardia and a fixed heart rate
to the development of silent myocardial infarction (12).
Consequently, myocardial hypertrophy, fibrosis and
myocardial dysfunction lead to the progression of heart failure.
Cardiac complications, such as disrupted insulin signaling, the
renin-angiotensin-aldosterone system, and small and large-vessel
disease lead to alterations in myocardial energetics, cardiac
remodeling and impaired perfusion (11). Myocardial energy depletion in
individuals suffering from DM is multifactorial and is associated
with limitations in the uptake and utilization of substrates,
mitochondrial dysfunction and affected energy transfer from
mitochondria to myofibrils. These metabolic alterations combined
with affected myocardial perfusion, may result in the reduced
ability of the heart to cope with acute increases in workload
(13).
As a result, myocardial dysfunction, cardiac
stiffness and myocardial fibrosis occur. Finally, cytosolic calcium
trafficking and gene expression cause excitation-contraction
decoupling, and lead to cardiomyocyte contraction and relaxation
(11).
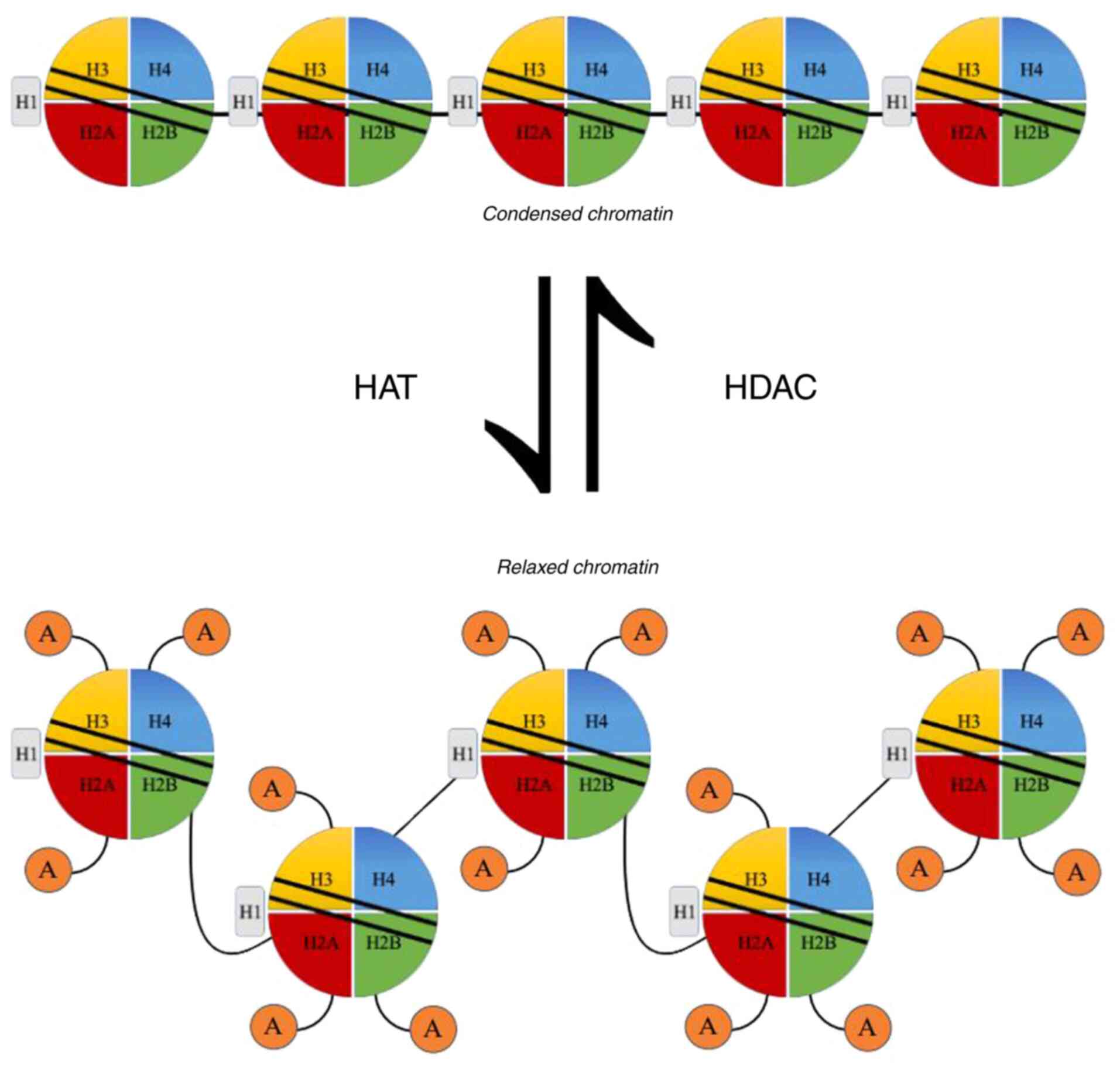
Histone deacetylases (HDACs) are enzymes that exert
their activities through the alteration of chromatin regulation.
The balance between HDACs and histone acetyltransferases (HATs)
controls that regulation. As their name suggests, they remove
acetyl-groups from histones. They actually regulate the interaction
between the negatively charged DNA and positively charged histones.
Thus, they serve as a repressor of transcription (14) (Fig.
1). The 18 HDACs are divided into four classes as follows: The
class I Rpd3-like proteins (HDAC1, HDAC2, HDAC3 and HDAC8); the
class II Hda1-like proteins (HDAC4, HDAC5, HDAC6, HDAC7, HDAC9 and
HDAC10); the class III Sir2-like proteins [sirtuin (SIRT)1, SIRT2,
SIRT3, SIRT4, SIRT5, SIRT6 and SIRT7]; and the class IV protein
(HDAC11) (15).
The HDACs of classes I, II and IV exhibit a
zinc-dependent activity. On the contrary, class III HDACs or SIRTs
(SIRT1-7) require nicotinamide adenine dinucleotide (NAD) for
catalysis (Table I) (14). Their crucial roles in epigenetics and
gene expression have rendered them potential targets in various
diseases, including both metabolic and cardiovascular entities
(16).
 | Table IHDAC classification. |
Table I
HDAC classification.
| Class I | Class II | Class III | Class IV |
|---|
|
Zn+-dependent | NAD-dependent |
Zn+-dependent |
|---|
| HDAC1 | IIA | SIRT1 | HDAC11 |
| HDAC2 |
HDAC4 | SIRT2 | |
| HDAC3 |
HDAC5 | SIRT3 | |
| HDAC8 |
HDAC7 | SIRT4 | |
| |
HDAC9 | SIRT5 | |
| | | SIRT6 | |
| | | SIRT7 | |
| | IIB | | |
| |
HDAC6 | | |
| |
HDAC10 | | |
HDAC inhibitors (HDACIs) are already used as
anticancer agents. Of note, four of them have already been approved
by the US Food and Drug Administration (FDA) against hematological
malignancies (14). Additionally,
their anticancer activity has been reported extensively in the
literature, including colorectal, hepatocellular, pancreatic,
breast, thyroid, lung, endometrial, and other cancers such as uveal
and cutaneous melanoma (17-27).
Furthermore, their role appear to be crucial in fighting other
diseases, including viral infections and both inflammatory and
neurological disorders (28,29). Hyperglycemia is a crucial factor that
contributes to the development of oxidative stress, mitochondrial
injury, reactive oxygen species (ROS) and myocardial inflammation
(30,31). Thus, HDACIs appear to be promising
therapeutic agents against cardiovascular diseases, including
DCM.
The present review article reports pre-clinical
trials identified in which HDACIs were tested in animal models of
DCM. The roles and mechanisms of action of HDACIs in metabolic and
cardiovascular diseases were also discussed.
2. Role of HDACIs in experimental models of
DCM
In 2015, Chen et al (31) examined the role of sodium butyrate,
which is a specific HDACI, in preventing myocardial dysfunction in
diabetic mice. They established a model of diabetes using mice by
injecting the mice intraperitoneally with streptozocin. The next
step was the administration of sodium butyrate to certain subgroups
of these mice. The echocardiography findings revealed that cardiac
dysfunction was attenuated in he diabetic mice receiving sodium
butyrate. Additionally, the attenuation of cardiac remodeling and
interstitial fibrosis were noted. These findings were associated
with an increase in cardiac angiogenesis, and a decrease in both
apoptosis and active caspase 3. Glucose metabolism was improved
through the upregulation of glucose transporter (GLUT)-1 and -4.
Finally, sodium butyrate exerted an antioxidant effect on the
myocardium by elevating superoxide dismutase levels (31).
Lee et al (32,33)
conducted research on diabetic rats who received low doses of
streptozocin. MPT0E014, which is a pan-HDACI, was administered to a
subgroup of diabetic rats. The left-ventricular end-diastolic
diameter was smaller in the diabetic rats receiving the HDACI
compared to the diabetic rats not receiving the treatment. The
reduction in blood sugar and triglyceride levels revealed the
anti-hyperglycemic and hypolipidemic effects of MPT0E014. At the
cellular level, GLUT-4 expression was augmented through HDACI
administration via an insulin-independent pathway. The cleavage of
poly(ADP-ribose) polymerase 1 (PARP-1) was decreased, inhibiting
apoptosis. The levels of pro-inflammatory cytokines, such as tumor
necrosis factor α (TNF-α) or interleukin (IL)-6 were reduced
following treatment with MPT0E014(32). Finally, the procedure of autophagy,
which is necessary for the development and homeostasis of cells,
was accelerated following the administration of this HDACI
(33).
Wu et al (34)
investigated whether the suppression of HDACs in diabetic rat
hearts could protect the rats from myocardial ischemia/reperfusion
(MI/R) injury mediated via the PI3K/Akt pathway (34). This pathway results in the activation
of anti-apoptotic proteins that inhibit mitochondrial dysfunction
during MI/R injury (34,35). In their experiment, diabetic rats
underwent 45 min of ischemia followed by 3 h of reperfusion. They
used trichostatin A (TSA), a selective I and II HDACI, both in
vitro and in vivo experiments. Their study was the first
to demonstrate that MI/R injury and diabetes increased HDAC
activity in rat hearts. However, TSA protected the function and
integrity of mitochondria both in vivo and in vitro
through the expression of p-Akt via the increased phosphorylation
of p-Foxo3a and Foxo3a in the cytoplasm of myocardial cells
(34).
Leng et al (36) used the same methods and experimental
model as those in the study by Wu et al (34); however, they concentrated on the
inhibition of HDAC6 activity using tubastatin A (Tub A), a highly
selective HDAC6 inhibitor. Their research demonstrated that both
in vivo and in vitro, Tub A exhibited its
cardioprotective actions in diabetic rat hearts by modulating
peroxiredoxin 1 (Prdx1) acetylation and attenuating ROS generation
and subsequent cellular injury (36).
Another group of researchers examined the
association between the inhibition of HDAC3 and DCM. RGFP966, a
selective HDAC3 inhibitor, and valproic acid were administered to
mice with type 1 diabetes (37).
Treatment with RGFP966 prevented cardiac hypertrophy through
various mechanisms. Firstly, a reduction in heart hypertrophy and
fibrosis markers was noted. Histologically, cardiomyocyte size,
fibrosis and the accumulation of collagen were suppressed. Insulin
resistance, oxidative stress and inflammatory processes were all
diminished. TNF-α, plasminogen activator inhibitor-1 and ROS
production were downregulated, whereas the expression of GLUT-4 was
upregulated. The dual specificity phosphatase 5 (DUSP5), which is
an extracellular signal-regulated kinase 1 and 2 (ERK1/2) nuclear
phosphatase, inhibits phosphorylated extracellular signal-regulated
kinases ERK1/2, which accelerates cardiac hypertrophy (37). The administration of RGFP966 exerted
its cardioprotective effects through an increase in the levels of
DUSP5. Last but not least, they demonstrated that these effects
lasted for a period of time, since the protective activity of
RGFP966 remained for 3 months after the end of its administration
(37).
Zhang et al (7) performed a study in which they
administered sodium butyrate to mice with type 2 diabetes.
Treatment with sodium butyrate reduced obesity,
hypercholesterolemia, and glucose intolerance. In addition, the
wall thickness and the internal dimension of the left ventricle
were smaller in the diabetic group treated with sodium butyrate,
compared with the untreated diabetic group. Both the size of the
cardiomyocytes and collagen presence were limited in the mice
receiving the HDACI. The production of superoxide and active
caspase-3, which is an apoptotic agent, were significantly reduced.
Angiogenesis and capillary formation were enhanced using sodium
butyrate. Finally, sodium butyrate can activate signaling pathways,
such as mitogen-activated protein kinase 3
(MKK3)/p38/regulated-activated kinase (PRAK) and Akt-1, in order to
regulate proper cardiac function (7).
Bocchi et al (38) reported the action of suberoylanilide
hydroxamic acid (SAHA), a pan-HDACI, in the cardiomyocytes of
diabetic rats. Ventricular cardiomyocytes were extracted in order
to measure their contractility and calcium dynamics. These cells
were either treated with SAHA for 90 min or left untreated. The
group treated with SAHA exhibited higher contractility and calcium
dynamics. This was a result of the upregulation in ryanodine
receptors and a decrease in intracellular ROS levels (38).
Table II summarizes
the findings of all the aforementioned studies reporting the
potential role of HDACIs against DCM in experimental models
(5,31-34,36-38).
| Table IIPreclinical studies of histone
deacetylase inhibitors in diabetic cardiomyopathy. |
Table II
Preclinical studies of histone
deacetylase inhibitors in diabetic cardiomyopathy.
| Authors, year | Specimen | HDACI | Outcome | (Refs.) |
|---|
| Chen et al,
2015 | • Control mice | Sodium butyrate
(inhibitor of HDAC class I and IIA) | Reduction of
apoptosis, oxidative stress, cardiac remodeling, fibrosis.
Upregulation of GLUT-1 and -4, and angiogenesis. | (31) |
| • Control receiving
sodium butyrate mice |
| • Mice with
streptozocin-induced diabetes |
| • Mice with
streptozocin-induced diabetes receiving sodium butyrate |
| Zhang et al,
2017 | • Mice fed a
high-fat diet | Sodium
butyrate | Reduction of
obesity, hypercholesterolemia, hyperglycemia, apoptosis, oxidative
stress, left ventricular wall thickness and internal diameter.
Increase in angiogenesis. | (7) |
| • Mice fed a
high-fat diet and treated with sodium butyrate |
| • Mice fed a
standard chow diet |
| • Mice fed a
standard chow diet and treated with sodium butyrate |
| Wu et al,
2017 | • Control rats | Trichostatin A | Protected against
myocardial ischemia/reperfusion injury and hypoxia/reoxygenation
injury under diabetic. conditions | (34) |
| • Control rats
subjected to myocardial ischemia/reperfusion injury |
| • Diabetic
rats |
| • Diabetic rats
subjected to myocardial ischemia/reperfusion injury |
| Xu et al,
2017 | • Mice with type 1
diabetes | RGFP966 (Selective
HDAC-3 inhibitor), VPA (pan-HDACI) | Reduction of
cardiac hypertrophy, cardiomyocyte size, fibrosis, insulin
resistance, inflammation and oxidative stress. Increase in levels
of DUSP5. | (37) |
| • Mice with type 1
diabetes treated with VPA |
| • Mice with type 1
diabetes treated with RGFP966 |
| • Control |
| • Control and
VPA |
| • Control and
RGFP966 |
| Lee et al,
2016 and 2018 | • Control | MPT0E014
(pan-HDACI) | Reduction of
glucose, triglycerides, inflammatory cytokines and apoptosis.
Up-regulation of autophagy, GLUT-4 and homeostasis. | (32,33) |
| • Rats fed a
high-fat diet with streptozocin-induced T2D and treated with
MPT0E014 |
| • Rats fed a
high-fat diet with streptozocin-induced T2D |
| Leng et al,
2018 | • Control rats | Tubastatin A | Protected against
myocardial ischemia/reperfusion injury under diabetic
conditions | (36) |
| • Control rats with
ischemia/reperfusion injury |
| • Diabetic
rats |
| • Diabetic rats
with ischemia/reperfusion injury |
| Bocchi et
al, 2019 | • Control | SAHA
(pan-HDACI) | Increase in
contractility and calcium dynamics. Reduction in intracellular
oxidative stress. | (38) |
| • Mice with
streptozocin-induced type 2 diabetes |
| • Mice with
streptozocin-induced type 2 diabetes and treated with SAHA |
3. Mechanisms of prevention of cardiac
hypertrophy via the inhibition of HDACs
HDACIs are divided according to their actions on
HDACs. They are divided into hydroxamic acids, short-chain fatty
acids, cyclic peptides and benzamides. They are mostly studied and
used against malignancies (8).
However, extensive research into the possible therapeutic effects
of HDACIs on DCM has been performed (5,31-34,36-38).
HDAsC contribute to the development of DCM through various
metabolic and hypertrophic pathways (39).
The inhibition of HDACs prevents cardiac hypertrophy
a number of mechanisms. As described above, active caspase-3 is a
protein which triggers apoptosis. The anti-apoptotic effects of
HDACIs are demonstrated through the reduction of apoptosis
(31). The inflammatory process is
reduced through the downregulation of pro-inflammatory cytokines,
such as TNF-α, IL-1, IL-6 and IFN-γ (40). Angiogenesis is accomplished through
the increased production of vascular endothelial growth factor
(VEGF) (41). The reduction of ROS
generation diminishes oxidative stress (37) and can be accomplished by the
modulation of acetylation of Prdx1(36). Autophagy is an adaptive mechanism,
whose absence induces intracellular death (42). As Lee et al (32,33)
demonstrated, HDACIs can increase autophagy and protect the
myocardium. Wu et al (34)
demonstrated that the protection of the function of mitochondria
via the regulation of P13K/Akt expression decreased cell apoptosis
and protected the diabetic heart. The transcription of GLUT-4 is
regulated by class II HDAC in skeletal muscles (43,44). The
upregulation of GLUT by HDACIs leads to the entrance of glucose
into cells and to a reduction in both insulin resistance and
hyperglycemia (32,45). Fibrosis is reduced by inhibiting
TGF-β and the angiotensin type 2 receptor (46). The mechanisms of action of HDACIσ
against DCM are reported in Table
III and are illustrated in Fig.
2.
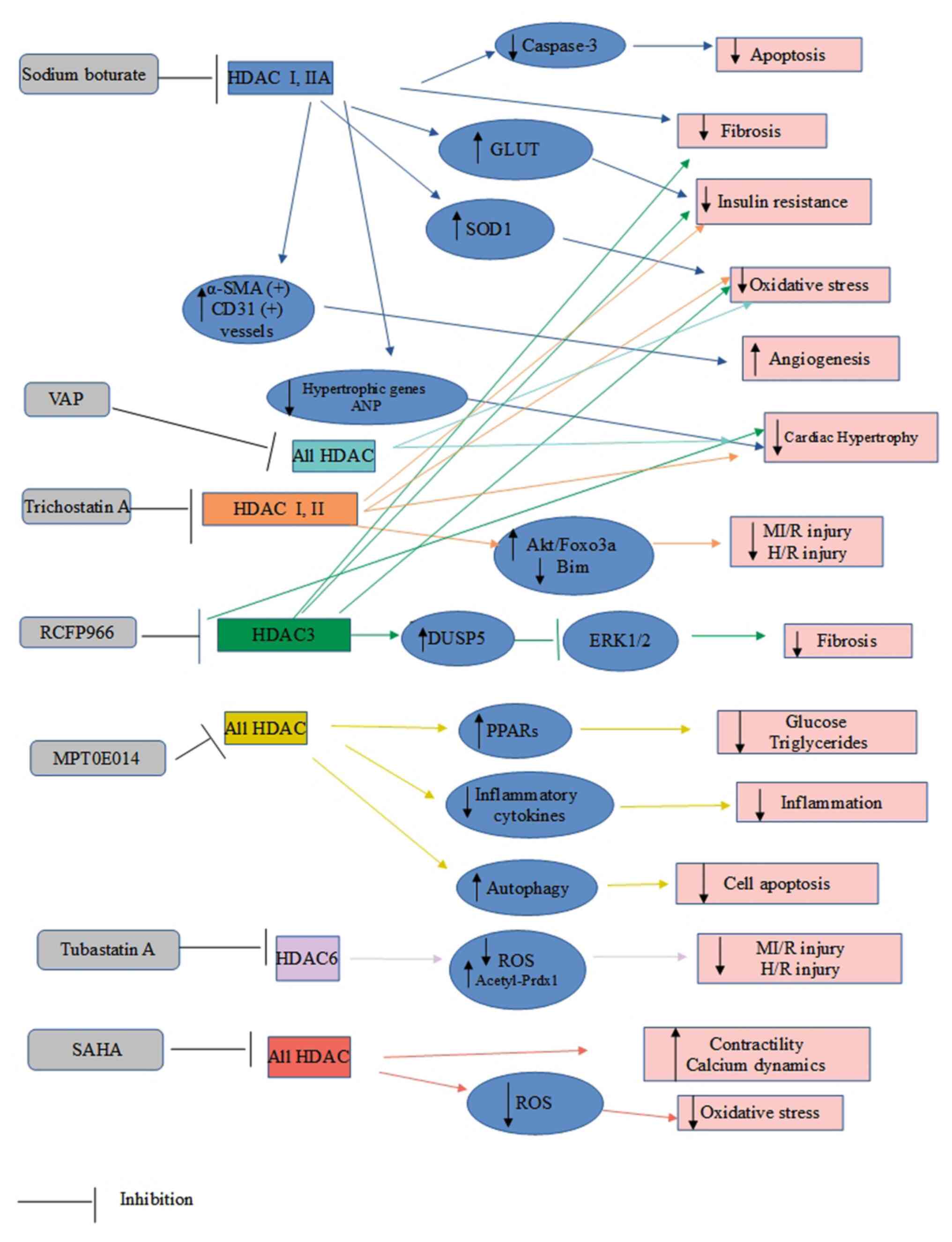 | Figure 2Mechanisms of action of HDAC
inhibitors against diabetic cardiomyopathy. ANP, atrial natriuretic
peptide; α-SMA, α smooth muscle actin; DUSP5, dual specificity
phosphatase 5; GLUT, glucose transporter; HDAC, histone
deacetylase; H/R, hypoxia/reoxygenation; MI/R, myocardial
ischemia/reperfusion; PPARs, peroxisome proliferator activated
receptors; PRDX1, peroxiredoxin 1; ROS, reactive oxygen species;
SAHA, suberoylanilide hydroxamic acid; SOD1, superoxide dismutase
1; T2D, type 2 diabetes; VPA, valproic acid. |
| Table IIIPreventive mechanisms of action of
HDACIs against DCM. |
Table III
Preventive mechanisms of action of
HDACIs against DCM.
| Reduction | Increase |
|---|
| ROS | Autophagy |
| Inflammatory
cytokines (IL-1, IL-6, TNF-α, IFN-γ) | GLUT-1 and -4 |
| Apoptosis
(caspase-3) | Angiogenesis |
| Fibrosis | Contractility |
| Left ventricular
thickness and internal diameter Insulin resistance | Calcium
dynamics |
4. Conclusion and future perspectives
Conventionally, HDACIs are considered as anticancer
drugs. However, they appear to be useful in the treatment of other
non-malignant diseases. The present review article reported
numerous preclinical trials, which examined the association between
HDACs, HDACIs and DCM. The use of HDACIs has yielded encouraging
results against disease progression. Research has markedly enhance
current knowledge on HDACs and their roles in metabolic and
cardiovascular dysfunction. Additionally, HDACs are considered as
possible therapeutic targets against heart failure in general. Due
to the novelty of this approach, further research into the
mechanisms of action and biochemical pathways of HDACs and their
inhibitors is warranted prior to any clinical applications.
Extending current knowledge regarding the therapeutic potential of
HDACIs against these diseases will pave the way for the discovery
of novel targeted epigenetic drugs. Thus, millions of individuals
will be beneficially affected worldwide in the future.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
AG, CD, NG and VVK conceptualized the study. VEG,
DAS, PP, ED, AP, AS, KT, GM, Gky, GKo and DD analyzed the data from
the literature to be included in the review, and wrote and prepared
the draft of the manuscript. GKo and DD provided critical
revisions. ED, AS and AP prepared the figure and the tables. All
authors contributed to manuscript revision and have read and
approved the final version of the manuscript. Data authentication
is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wang Y, Yang H, Huynh Q, Nolan M, Negishi
K and Marwick TH: Diagnosis of nonischemic stage B heart failure in
type 2 diabetes mellitus. JACC Cardiovasc Imaging. 11:1390–1400.
2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Hölscher ME, Bode C and Bugger H: Diabetic
cardiomyopathy: Does the type of diabetes matter? Int J Mol Sci.
17(2136)2016.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Yilmaz S, Canpolat U, Aydogdu S and Abboud
HE: Diabetic cardiomyopathy; Summary of 41 years. Korean Circ J.
45:266–272. 2015.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Avogaro A, de Kreutzenberg SV, Negut C,
Tiengo A and Scognamiglio R: Diabetic cardiomyopathy: A metabolic
perspective. Am J Cardiol. 93:13A–16A. 2004.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Jia G, Hill MA and Sowers JR: Diabetic
cardiomyopathy: An update of mechanisms contributing to this
clinical entity. Circ Res. 122:624–638. 2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Dinarello CA: Anti-inflammatory agents:
Present and future. Cell. 140:935–950. 2010.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Zhang L, Du J, Yano N, Wang H, Zhao YT,
Dubielecka PM, Zhuang S, Chin YE, Qin G and Zhao TC: Sodium
butyrate protects-against high fat diet-induced cardiac dysfunction
and metabolic disorders in type II diabetic mice. J Cell Biochem.
118:2395–2408. 2017.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Jia G, Whaley-Connell A and Sowers JR:
Diabetic cardiomyopathy: A hyperglycaemia- and
insulin-resistance-induced heart disease. Diabetologia. 61:21–28.
2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Dhingra R and Vasan RS: Diabetes and the
risk of heart failure. Heart Fail Clin. 8:125–133. 2012.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Rosano GM, Vitale C and Seferovic P: Heart
failure in patients with diabetes mellitus. Card Fail Rev. 3:52–55.
2017.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Marwick TH, Ritchi R, Shaw JE and Kaye D:
Implications of underlying mechanisms for the recognition and
management of diabetic cardiomyopathy. J Am Coll Cardiol.
71:339–351. 2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Serhiyenko VA and Serhiyenko AA: Cardiac
autonomic neuropathy: Risk factors, diagnosis and treatment. World
J Diabetes. 9:1–24. 2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Levelt E, Rodgers CT, Clarke WT, Mahmod M,
Ariga R, Francis JM, Liu A, Wijesurendra RS, Dass S, Sabharwal N,
et al: Cardiac energetics, oxygenation, and perfusion during
increased workload in patients with type 2 diabetes mellitus. Eur
Heart J. 37:3461–3469. 2016.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Garmpi A, Garmpis N, Damaskos C, Valsami
S, Spartalis E, Lavaris A, Patelis N, Margonis GA, Apostolou KG,
Spartalis M, et al: Histone deacetylase inhibitors as a new
anticancer option: How far can we go with expectations? J BUON.
23:846–861. 2018.PubMed/NCBI
|
|
15
|
Seto E and Yoshida M: Erasers of histone
acetylation: The histone deacetylase enzymes. Cold Spring Harb
Perspect Biol. 6(a018713)2014.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Zwergel C, Stazi G, Valente S and Mai A:
Histone deacetylase inhibitors: Updated studies in various
epigenetic-related diseases. J Clin Epigenet. 2(1)2016.
|
|
17
|
Garmpis N, Damaskos C, Garmpi A, Nonni A,
Georgakopoulou VE, Antoniou E, Schizas D, Sarantis P, Patsouras A,
Syllaios A, et al: Histone deacetylases and their inhibitors in
colorectal cancer therapy: Current evidence and future
considerations. Curr Med Chem. 29:2979–2994. 2022.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Garmpis N, Damaskos C, Garmpi A,
Georgakopoulou VE, Sarantis P, Antoniou EA, Karamouzis MV, Nonni A,
Schizas D, Diamantis E, et al: Histone deacetylase inhibitors in
the treatment of hepatocellular carcinoma: Current evidence and
future opportunities. J Pers Med. 11(223)2021.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Damaskos C, Garmpis N, Karatzas T,
Nikolidakis L, Kostakis ID, Garmpi A, Karamaroudis S, Boutsikos G,
Damaskou Z, Kostakis A and Kouraklis G: Histone deacetylase (HDAC)
inhibitors: Current evidence for therapeutic activities in
pancreatic cancer. Anticancer Res. 35:3129–3135. 2015.PubMed/NCBI
|
|
20
|
Damaskos C, Garmpis N, Valsami S, Kontos
M, Spartalis E, Kalampokas T, Kalampokas E, Athanasiou A, Moris D,
Daskalopoulou A, et al: Histone deacetylase inhibitors: An
attractive therapeutic strategy against breast cancer. Anticancer
Res. 37:35–46. 2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Garmpis N, Damaskos C, Garmpi A,
Kalampokas E, Kalampokas T, Spartalis E, Daskalopoulou A, Valsami
S, Kontos M, Nonni A, et al: Histone deacetylases as new
therapeutic targets in triple-negative breast cancer: Progress and
promises. Cancer Genomics Proteomics. 14:299–313. 2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Damaskos C, Garmpis N, Valsami S,
Spartalis E, Antoniou EA, Tomos P, Karamaroudis S, Zoumpou T,
Pergialiotis V, Stergios K, et al: Histone deacetylase inhibitors:
A novel therapeutic weapon against medullary thyroid cancer?
Anticancer Res. 36:5019–5024. 2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Spartalis E, Athanasiadis DI, Chrysikos D,
Spartalis M, Boutzios G, Schizas D, Garmpis N, Damaskos C, Paschou
SA, Ioannidis A, et al: Histone deacetylase inhibitors and
anaplastic thyroid carcinoma. Anticancer Res. 39:1119–1127.
2019.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Damaskos C, Tomos I, Garmpis N,
Karakatsani A, Dimitroulis D, Garmpi A, Spartalis E, Kampolis CF,
Tsagkari E, Loukeri AA, et al: Histone deacetylase inhibitors as a
novel targeted therapy against non-small cell lung cancer: Where
are we now and what should we expect? Anticancer Res. 38:37–43.
2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Garmpis N, Damaskos C, Garmpi A, Spartalis
E, Kalampokas E, Kalampokas T, Margonis GA, Schizas D, Andreatos N,
Angelou A, et al: Targeting histone deacetylases in endometrial
cancer: A paradigm-shifting therapeutic strategy? Eur Rev Med
Pharmacol Sci. 22:950–960. 2018.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Moschos MM, Dettoraki M, Androudi S,
Kalogeropoulos D, Lavaris A, Garmpis N, Damaskos C, Garmpi A and
Tsatsos M: The role of histone deacetylase inhibitors in uveal
melanoma: Current evidence. Anticancer Res. 38:3817–3824.
2018.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Garmpis N, Damaskos C, Garmpi A,
Dimitroulis D, Spartalis E, Margonis GA, Schizas D, Deskou I, Doula
C, Magkouti E, et al: Targeting histone deacetylases in malignant
melanoma: A future therapeutic agent or just great expectations?
Anticancer Res. 37:5355–5362. 2017.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Gray SG: Epigenetic treatment of
neurological disease. Epigenomics. 3:431–450. 2011.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Savi M, Bocchi L, Sala R, Frati C,
Lagrasta C, Madeddu D, Falco A, Pollino S, Bresciani L, Miragoli M,
et al: Parenchymal and stromal cells contribute to pro-inflammatory
myocardial environment at early stages of diabetes: Protective role
of resveratrol. Nutrients. 8(729)2016.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Bugger H and Abel ED: Molecular mechanisms
of diabetic cardiomyopathy. Diabetologia. 57:660–671.
2014.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Chen Y, Du J, Zhao YT, Zhang L, Lv G,
Zhuang S, Qin G and Zhao TC: Histone deacetylase (HDAC) inhibition
improves myocardial function and prevents cardiac remodeling in
diabetic mice. Cardiovasc Diabetol. 14(99)2015.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Lee TI, Kao YH, Tsai WC, Chung CC, Chen YC
and Chen YJ: HDAC inhibition modulates cardiac PPARs and fatty acid
metabolism in diabetic cardiomyopathy. PPAR Res.
2016(5938740)2016.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Lee TI, Bai KJ, Chen YC, Lee TW, Chung CC,
Tsai WC, Tsao SY and Kao YH: Histone deacetylase inhibition of
cardiac autophagy in rats on a high-fat diet with low-dose
streptozotocin-induced type 2 diabetes mellitus. Mol Med Rep.
17:594–601. 2018.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Wu Y, Leng Y, Meng Q, Xue R, Zhao B, Zhan
L and Xia Z: Suppression of excessive histone deacetylases activity
in diabetic hearts attenuates myocardial ischemia/reperfusion
injury via mitochondria apoptosis pathway. J Diabetes Res.
2017(8208065)2017.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Ban K, Cooper AJ, Samuel S, Bhatti A,
Patel M, Izumo S, Penninger JM, Backx PH, Oudit GY and Tsushima RG:
Phosphatidylinositol 3-kinase gamma is a critical mediator of
myocardial ischemic and adenosine-mediated preconditioning. Circ
Res. 103:643–653. 2008.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Leng Y, Wu Y, Lei S, Zhou B, Qiu Z, Wang K
and Xia Z: Inhibition of HDAC6 activity alleviates myocardial
ischemia/reperfusion injury in diabetic rats: Potential role of
peroxiredoxin 1 acetylation and redox regulation. Oxid Med Cell
Longev. 2018(9494052)2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Xu Z, Tong Q, Zhang Z, Wang S, Zheng Y,
Liu Q, Qian LB, Chen SY, Sun J and Cai L: Inhibition of HDAC3
prevents diabetic cardiomyopathy in OVE26 mice via epigenetic
regulation of DUSP5-ERK1/2 pathway. Clin Sci (Lond). 131:1841–1857.
2017.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Bocchi L, Motta BM, Savi M, Vilella R,
Meraviglia V, Rizzi F, Galati S, Buschini A, Lazzaretti M,
Pramstaller PP, et al: The histone deacetylase inhibitor
suberoylanilide hydroxamic acid (SAHA) restores cardiomyocyte
contractility in a rat model of early diabetes. Int J Mol Sci.
20:1873–1885. 2019.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Bagshi RA and Weeks KL: Histone
deacetylases in cardiovascular and metabolic diseases. J Mol Cell
Cardiol. 130:151–159. 2019.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Larsen L, Tonnesen M, Ronn SG, Størling J,
Jørgensen S, Mascagni P, Dinarello CA, Billestrup N and
Mandrup-Poulsen T: Inhibition of histone deacetylases prevents
cytokine-induced toxicity in beta cells. Diabetologia. 50:779–789.
2007.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Bogaard HJ, Mizuno S, Hussaini AA, Toldo
S, Abbate A, Kraskauskas D, Kasper M, Natarajan R and Voelkel NF:
Suppression of histone deacetylases worsens right ventricular
dysfunction after pulmonary artery banding in rats. Am J Respir
Crit Cure Med. 183:1402–1410. 2011.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Hur KY, Jung HS and Lee MS: Role of
autophagy in β-cell function and mass. Diabetes Obes Metab. 2 (12
Suppl):S20–S26. 2010.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Weems J and Olson AL: Class II histone
deacetylases limit GLUT4 gene expression during adipocyte
differentiation. J Biol Chem. 286:460–468. 2011.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Thai MV, Guruswamy S, Cao KT, Pessin JE
and Olson AL: Myocyte enhancer factor 2 (MEF2)-binding site is
required for GLUT4 gene expression in transgenic mice. Regulation
of MEF2 DNA binding activity in insulin-deficient diabetes. J Biol
Chem. 273:14285–14292. 1998.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Shao D and Tian R: Glucose transporters in
cardiac metabolism and hypertrophy. Compr Physiol. 6:331–351.
2015.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Kao YH, Liou JP, Chung CC, Lien GS, Kuo
CC, Chen SA and Chen YJ: Histone deacetylase inhibition improved
cardiac functions with direct antifibrotic activity in heart
failure. Int J Cardiol. 168:4178–4183. 2013.PubMed/NCBI View Article : Google Scholar
|