1. Cartilage: A closer look, cartilage
degradation and the challenge of regeneration
Cartilage is a resilient, yet flexible connective
tissue that performs several essential physiological functions,
including the maintenance of structural integrity and preservation
of shape, the absorption of mechanical shock during movement, and
enabling smooth joint motion (1).
There are three types of cartilage: Elastic cartilage,
fibrocartilage and hyaline cartilage. Hyaline cartilage is the most
abundant, which provides smooth, low-friction surfaces that
facilitate joint movement. It is located at the ends of bones in
synovial joints, and in structures such as the nasal septum. A
specialized form of hyaline cartilage, known as articular
cartilage, covers the articulating surfaces of bones within
synovial joints and functions in close interaction with synovial
fluid, which provides lubrication and facilitates smooth joint
movement (2,3). Articular cartilage is composed of a
complex extracellular matrix (ECM) that is rich in type II collagen
fibrils embedded within a highly hydrated proteoglycan network,
predominantly consisting of aggrecan (4).
This unique structure confers exceptional
biomechanical properties to articular cartilage, including low
friction, high compressive strength and marked resilience (5). Its smooth, lubricated surface enables
near-frictionless joint articulation, while its ability to
withstand substantial mechanical loads protects the underlying
subchondral bone from mechanical damage (4). Articular cartilage contains
chondrocytes, the only resident cell type, which are embedded
within an ECM that they synthesize, remodel and maintain. These
cells are distributed throughout the cartilage matrix in a distinct
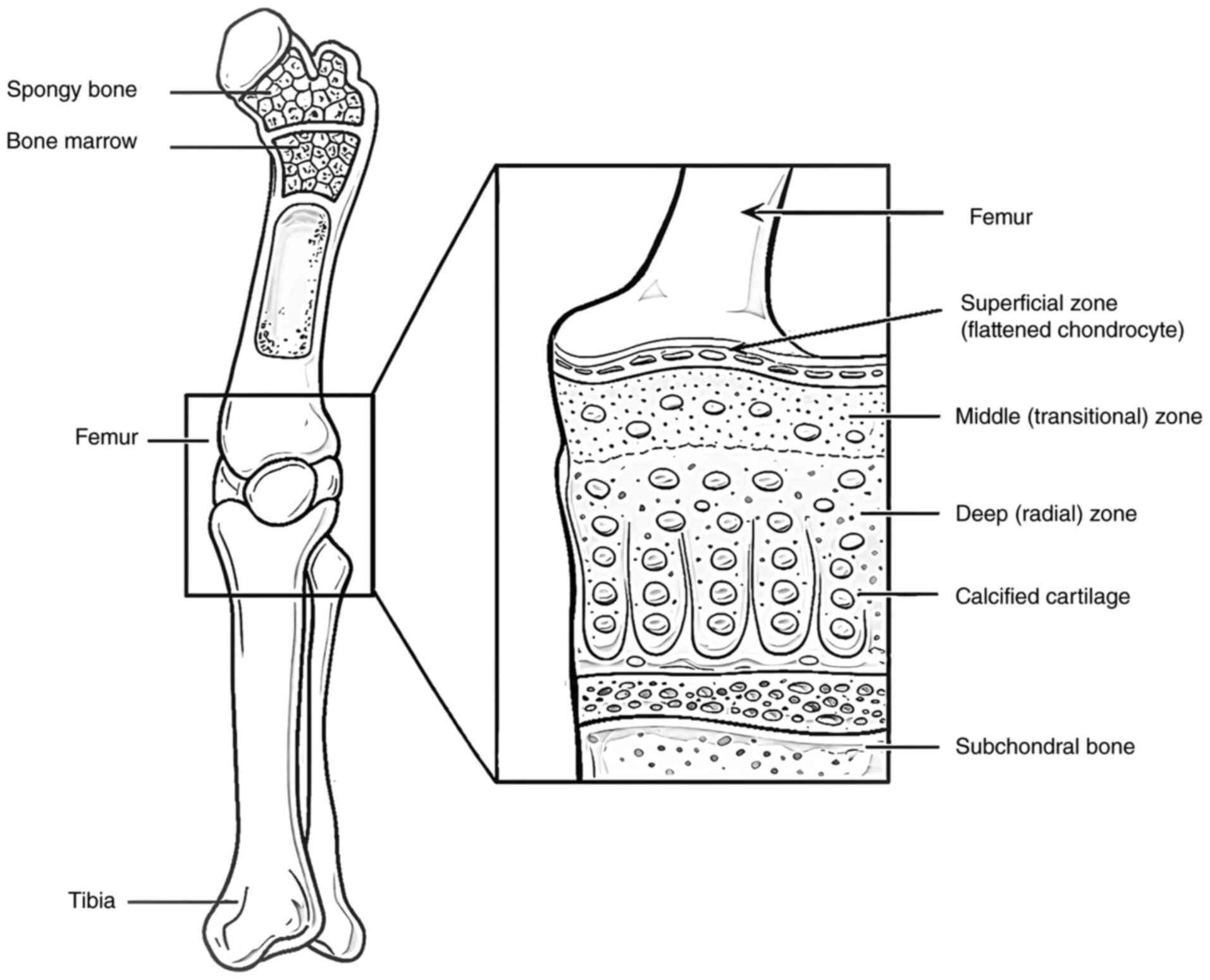
spatial organization, as illustrated in Fig. 1 (6-10).
Notably, articular cartilage is characterized as an avascular and
aneural tissue that significantly limits its intrinsic capacity for
repair following injury (3,11,12).
Consequently, damage to articular cartilage rarely heals
spontaneously and often leads to the progressive deterioration of
joint structure and function over time (6).
The limited regenerative capacity of articular
cartilage renders it particularly susceptible to injury and
degenerative joint diseases, such as osteoarthritis (OA).
Repetitive mechanical loading and age-related wear can initiate
cartilage degradation, leading to progressive structural and
functional deterioration. As degeneration advances, the protective
cartilage layer becomes progressively thinner, eventually exposing
the underlying subchondral bone to direct mechanical stress. This
process accelerates joint degeneration and ultimately contributes
to the development and progression of OA (12). OA, the most common form of arthritis,
is characterized by the progressive degeneration of articular
cartilage and alterations in subchondral bone, leading to chronic
pain, joint stiffness and reduced mobility.
Despite extensive research efforts, currently
available medical interventions have not reliably achieved the
complete regeneration of damaged cartilage nor provided consistent
long-term restoration of joint function. Consequently, considerable
attention has been directed towards the development of regenerative
therapeutic strategies. Emerging approaches, including stem cell
transplantation combined with biocompatible scaffolds and bioactive
growth factors, have been widely investigated in preclinical and
clinical settings, with promising results reported from in
vitro studies, animal models and early clinical applications
(13-18).
Conventional surgical techniques such as osteochondral plug
grafting and subchondral bone microfracture, which stimulate the
recruitment of endogenous bone marrow (BM)-derived cells have
demonstrated some effectiveness in treating small focal cartilage
defects. However, these approaches often fail to provide durable
repair in cases of extensive cartilage loss associated with
degenerative conditions such as OA (17,18).
Stem cell-based therapies provide significant promise by
introducing regenerative cells capable of promoting cartilage
repair, reducing inflammation, and alleviating pain, particularly
where conventional therapies are insufficient.
2. Stem cells and induced pluripotent stem
cells
Stem cells are characterized by their capacity for
self-renewal and multilineage differentiation (10). Based on their differentiation
potential, stem cells are classified as unipotent, multipotent and
pluripotent (19). Pluripotent stem
cells (PSCs), including embryonic stem cells (ESCs) and induced
PSCs (iPSCs) can differentiate into all somatic cell types
(20). ESCs are derived from the
inner cell mass of blastocysts, whereas iPSCs are generated through
the reprogramming of somatic cells, such as fibroblasts, via the
enforced expression of defined transcription factors. This landmark
discovery, recognized by the 2012 Nobel Prize in Physiology or
Medicine (https://www.nobelprize.org/prizes/medicine/2012/summary/),
revolutionized regenerative medicine. Cellular reprogramming
typically involves the transcription factors OCT4, SRY-related
HMG-box (SOX)2, KLF4 and c-Myc, which restore pluripotency by
resetting gene expression patterns. Although reprogramming
efficiency remains relatively low, successfully generated iPSCs
exhibit morphological, molecular and functional characteristics
comparable to those of ESCs. Their pluripotent status is commonly
confirmed through teratoma formation assays demonstrating
differentiation into tissues derived from all three germ layers
(21).
Human PSCs (hPSCs) represent a promising source of
chondroprogenitor cells owing to their unlimited proliferative
capacity and broad differentiation potential (22,23). In
addition to PSCs, mesenchymal stem cells (MSCs) have been
extensively investigated for cartilage repair. MSCs can be isolated
from multiple tissues, including BM, adipose tissue, synovium and
cartilage. They exhibit inherent chondrogenic potential with the
capacity for in vitro expansion (24). Of note, articular cartilage
progenitor cells (ACPCs) have been identified as a resident
progenitor cell population with potential relevance for cartilage
regeneration (25-27).
ACPCs predominantly localize in the superficial zone of articular
cartilage and exhibit key stem cell-like characteristics, including
clonogenicity, migratory capacity and chondrogenic differentiation
potential (25,27). Compared with fully differentiated
chondrocytes, ACPCs demonstrate enhanced proliferative capacity,
while maintaining a stable articular cartilage phenotype, rendering
them a promising cell source for regenerative strategies (26,27).
Recent research has highlighted their potential role in maintaining
cartilage homeostasis and contributing to tissue repair,
particularly during the early stages of OA (28). However, despite their therapeutic
promise, MSC-based cartilage repair approaches are frequently
complicated by phenotypic instability, including a tendency toward
hypertrophic differentiation and endochondral ossification
(29).
3. Chondrogenesis
Chondrogenesis is a tightly regulated developmental
process through which MSCs differentiate into chondrocytes, the
specialized cells responsible for producing and maintaining the ECM
of cartilage. This process involves sequential events, including
mesenchymal condensation, proliferation and the activation of
cartilage-specific gene expression programs (Fig. 2) (30). The transcription factor, SOX9, is
widely recognized as the master regulator of chondrogenesis
(31). During differentiation, MSCs
undergo characteristic morphological changes and initiate the
expression of key cartilage ECM components, including type II
collagen and aggrecan (4). Although
these molecules are not exclusively restricted to cartilage, they
represent the principal structural constituents responsible for the
unique mechanical and functional properties of cartilage tissue.
Chondrogenesis plays a fundamental role in embryonic skeletal
development and continues to be essential for the maintenance,
homeostasis, and repair of cartilage throughout life.
4. Proliferation and pre-hypertrophic
differentiation
BM-MSCs undergo staged differentiation toward mature
chondrocytes. The initial proliferative phase ensures sufficient
cell expansion (32). Subsequently,
cells enter a pre-hypertrophic stage marked by the upregulation in
the levels of chondrogenic markers, including aggrecan, Col2a1 and
SOX9, alongside the suppression of mesenchymal markers (33). ECM synthesis is initiated during this
phase.
5. Hypertrophic differentiation and
endochondral ossification
The differentiation of BM-MSCs into hypertrophic
chondrocytes represents the final stage of chondrogenic maturation.
During this phase, the cells undergo further phenotypic maturation
accompanied by distinct alterations in gene expression.
Hypertrophic chondrocytes are characterized by the markedly
elevated expression of type X collagen, alkaline phosphatase and
vascular endothelial growth factor (VEGF) (34). Type X collagen is widely recognized
as a hallmark marker of hypertrophic chondrocytes and contributes
to the structural organization of the developing cartilage matrix.
Increased alkaline phosphatase activity, an enzyme closely
associated with matrix mineralization, indicates the onset of the
transition toward ossification. In addition, VEGF promotes vascular
invasion, a critical event in the later stages of endochondral
ossification. This tightly regulated process involves the gradual
replacement of cartilage with bone and plays an essential role in
normal skeletal development. Under physiological conditions,
hypertrophic differentiation is carefully controlled to preserve
the stability and functional integrity of the cartilage matrix
(35). Consequently, a comprehensive
understanding of the molecular and cellular mechanisms governing
this differentiation process is crucial for the development of
effective therapeutic strategies for cartilage regeneration and
repair.
Early studies have demonstrated that hPSCs can
generate MSC-like cells through various approaches, including
co-culture with OP9 stromal cells (36), spontaneous differentiation under
standard culture conditions (37),
and direct mesengenesis under modified culture conditions (38,39).
These strategies yielded mesenchymal-like cells that often required
subsequent expansion in conventional MSC culture media to acquire
tri-lineage differentiation potential characteristic of MSCs
(40,41). Alternative methods, such as treatment
with the small molecule inhibitor, SB431542, have also been used to
produce a mesenchymal cell population. However, these cells
frequently exhibited heterogeneous developmental origins and
similarly required expansion in MSC-supportive media to attain
MSC-like properties (42). Although
these approaches have shown promise, the in vitro
chondrogenic capacity of PSC-derived MSCs has generally been
inferior to that of adult MSCs, particularly when transforming
growth factor-β (TGF-β) alone was used to induce chondrogenesis
(43). Comparative analyses of human
BM-MSCs and iPSC-derived MSCs from the same donor have revealed
significant functional differences, with iPSC-derived MSCs
demonstrating reduced chondrogenic potential (44,45).
6. MSC therapy for cartilage
regeneration
Recent advances in regenerative medicine have
increasingly focused on cell- and tissue-engineering strategies
that utilize articular chondrocytes or chondrogenic adult stem
cells (ASCs), such as MSCs derived from BM or adipose tissue
(46,47). In established clinical approaches,
selected cells are implanted into the defect site using a
periosteal patch in autologous chondrocyte implantation (ACI) or a
collagen membrane in matrix-induced ACI, to improve therapeutic
outcomes (18). Cell-based
therapies, specific stem cell populations such as BM-MSCs, which
possess the capacity to differentiate into chondrocytes, the
principal cell type responsible for cartilage matrix production,
are first isolated and selected. These cells are subsequently
expanded in vitro under growth factor-supplemented
conditions to generate sufficient numbers for therapeutic use.
Ex vivo expansion is often required to achieve an adequate
yield for effective treatment. However, prolonged culture and
expansion can alter cellular phenotype and functional properties,
thereby diminishing their suitability for cartilage repair
(48). Notably, in vivo
studies have demonstrated that uncultured or minimally expanded
articular chondrocytes are more capable of generating stable,
articular-like permanent cartilage compared with extensively
cultured cells (49,50). This observation underscores the
importance of preserving the native phenotype and functional
characteristics of therapeutic cells during preparation for
clinical use.
7. Use of iPSCs for cartilage
regeneration
The use of ESCs and ASCs, although promising for
cartilage regeneration, is associated with several critical
limitations. ESCs are derived from the inner cell mass of embryos,
and their isolation requires the destruction of the embryo, raising
substantial ethical concerns (51).
Moreover, ESC-based therapies carry a risk of teratoma formation as
residual undifferentiated cells may give rise to tumors following
transplantation (52). ASCs, while
more readily accessible from sources such as BM and adipose tissue,
possess limited proliferative capacity and may undergo phenotypic
drift during ex vivo expansion, potentially compromising
their chondrogenic potential (47).
Furthermore, achieving the consistent and reproducible
differentiation of both ESCs and ASCs into functional chondrocytes
remains challenging, as it requires precise control of culture
conditions and often involves complex combinations of growth
factors and biomaterials.
iPSCs provide several distinct advantages for
cartilage regeneration. Unlike ESCs, iPSCs can be generated from
the somatic cells of patients themselves, such as dermal
fibroblasts, eliminating ethical concerns related to embryo
destruction (1). This
patient-specific approach also reduces the risk of immune
rejection, enhancing the feasibility of clinical translation
(53). Additionally, iPSCs exhibit
broad differentiation potential and can be directed toward
chondrogenic lineages, including the generation of functional
chondrocytes (54). Their plasticity
enables the development of personalized cartilage grafts for
transplantation, representing a promising strategy for treating
cartilage defects and degenerative diseases, such as OA.
Despite these advantages however, several challenges
need to be addressed before the widespread clinical application of
iPSC-based therapies. A crucial unresolved question is whether
disease-associated abnormalities present in patients with
inflammatory joint disorders, such as OA or rheumatoid arthritis,
may be retained in iPSCs and their chondrocyte derivatives
(55). Addressing this issue will
require comprehensive comparative analyses of patient-derived
cells, together with the development of optimized culture and
differentiation protocols to ensure therapeutic safety and
efficacy.
For the successful clinical application of PSCs in
joint cartilage regeneration, several key objectives need to be
achieved. These include the efficient generation of
lineage-specific chondroprogenitors from hPSCs, the expansion of
these progenitors to clinically relevant numbers, without
compromising their chondrogenic capacity and their differentiation,
either in vitro or in vivo, into stable articular or
meniscal (permanent) chondrocytes rather than transient growth
plate chondrocytes (56). As
illustrated in Fig. 3, iPSCs are
derived from somatic cells such as fibroblasts and subsequently
differentiated into MSC-like intermediates prior to cartilage
formation. Chondrocyte differentiation is governed by complex and
tightly interconnected signaling pathways, as depicted in Figs. 3 and 4
(31,32,35,57-65).
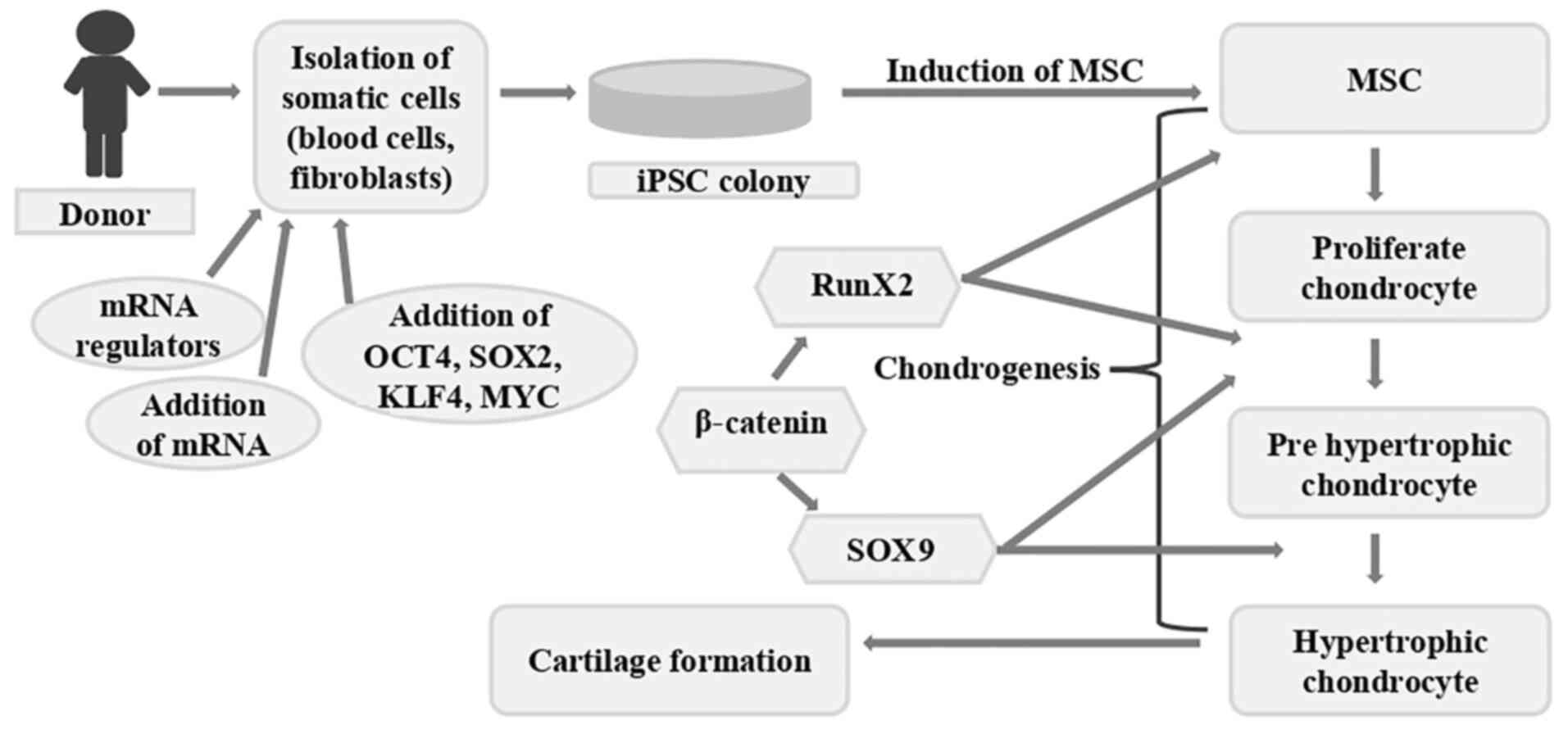 | Figure 3Diagrammatic illustration of
cartilage regeneration using iPSCs. By introducing particular
transcription factors, somatic cells can be transformed into iPSCs.
These iPSCs have the potential to develop into cartilage-producing
chondrocytes. The procedure for creating cartilage tissue from
cells iPSCs is shown in this schematic. Certain transcription
factors, including OCT4, SOX2, KLF4 and Myc, are introduced into
the somatic cells of an individual (fibroblasts, for example) to
reprogram them into iPSCs. These iPSCs can differentiate into a
variety of cell types, including chondrocytes, which are the cells
that form cartilage. A series of signaling events and the
activation of important transcription factors coordinate the
differentiation process. An early transcription factor called Runx2
is essential for starting chondrogenesis. Subsequently, genes
involved in the synthesis of cartilage matrix, including collagen
type II and aggrecan, are expressed in response to SOX9, another
essential transcription factor. Following a maturation process, the
differentiated chondrocytes go from proliferative to
pre-hypertrophic and ultimately hypertrophic. The information
presented in the image was obtained from previous studies (31,32,35,57-65).
iPSCs, induced pluripotent stem cells. |
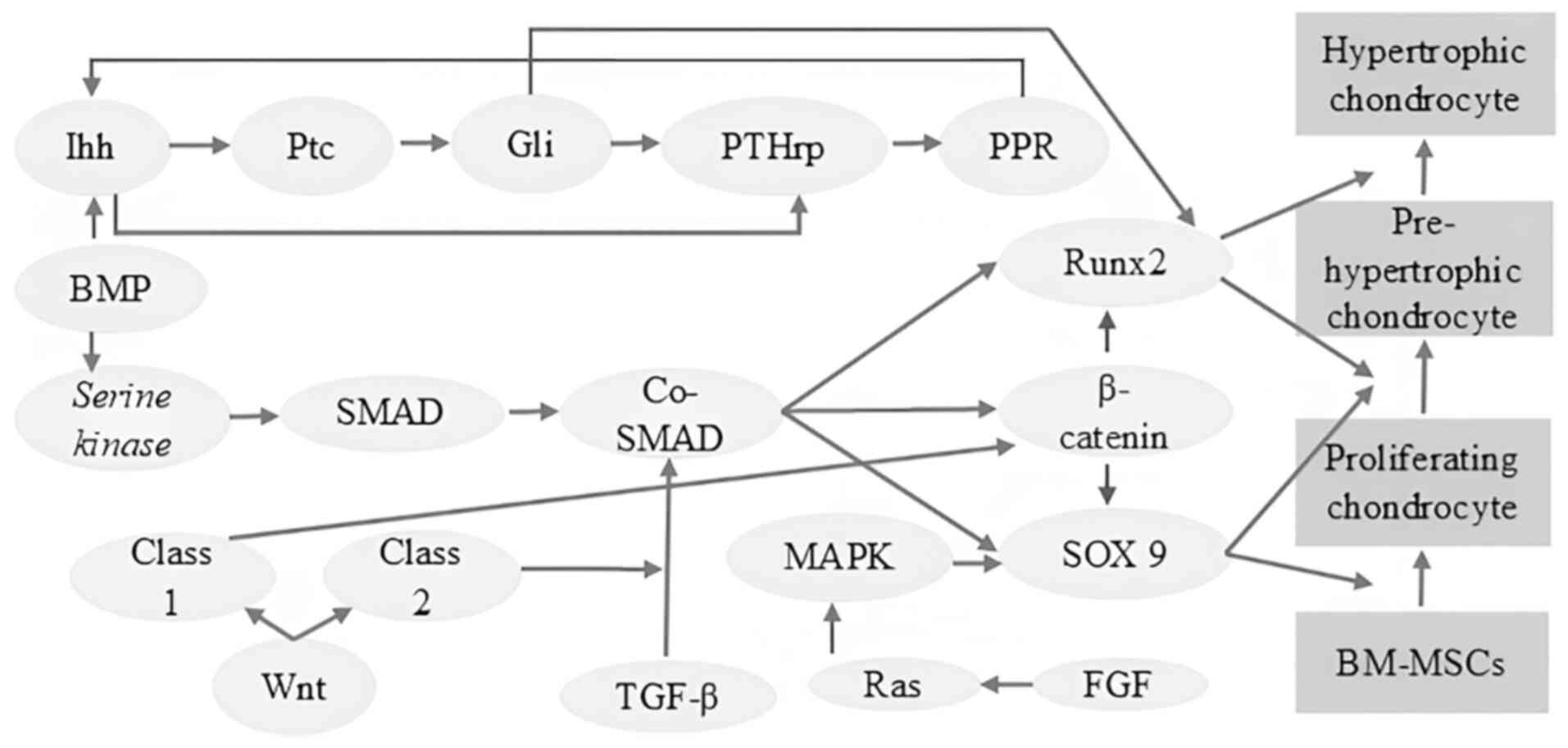 | Figure 4Schematic illustration of the complex
interplay of signaling pathways for chondrocyte differentiation.
The complex network of signaling pathways that control the
differentiation of BM-MSCs into chondrocytes is simplified in the
image. There are several phases in this developmental process, such
as pre-hypertrophy, hypertrophy and proliferation. These phases are
carefully coordinated by a wide range of signaling pathways,
including the Wnt/β-catenin pathway, which stimulates chondrocyte
proliferation and differentiation; the TGF-β/SMAD pathway, which is
essential for chondrogenesis as it controls the production of ECM
and chondrocyte maturation; the BMP/SMAD pathway, which stimulates
chondrocyte differentiation and controls matrix synthesis; the Ihh
signaling pathway and PTHrP signaling pathway, which controls
chondrocyte proliferation, hypertrophy and endochondral bone
formation; the FGF signaling pathway, which stimulates both
chondrocyte proliferation and differentiation; and the MAPK
signaling pathway, which is involved in multiple chondrogenesis,
including proliferation, differentiation and matrix synthesis. It
is important to understand that these signaling pathways show
complex interactions, with feedback loops and crosstalk having a
significant impact on chondrocyte development. Developing
successful strategies for cartilage tissue engineering and repair
requires a thorough understanding of these signaling mechanisms
(35,61,64,65).
BM-MSCs, bone marrow-derived mesenchymal stem cells; TGF-β,
transforming growth factor-β; Ihh, Indian hedgehog; PTHrP,
parathyroid hormone-related peptide; FGF, fibroblast growth
factor. |
The advantages and limitations of commonly used cell
sources for cartilage bioprinting are summarized in Table I.
 | Table IComparison of cell sources used in
cartilage bioprinting. |
Table I
Comparison of cell sources used in
cartilage bioprinting.
| Cell type | Advantages | Disadvantages | (Refs.) |
|---|
| Mesenchymal stem
cell (MSC) | Multipotent cells
capable of chondrogenic differentiation; widely available;
expandable in vitro | Risk of
hypertrophic differentiation; donor variability | (24,35,47,48) |
| Induced pluripotent
stem cell (iPSC) | Unlimited
proliferation; patient-specific; high differentiation
potential | Tumorigenic risk;
complex differentiation protocols | (3,53,55,59) |
| Articular
chondrocyte | Native
cartilage-producing cells produce collagen II and
proteoglycans | Limited
availability; dedifferentiation during expansion | (2,4,49,50) |
| Articular cartilage
progenitor cell (ACPC) | High proliferative
capacity; strong chondrogenic potential | Limited
availability; still under investigation | (18,56) |
8. Signaling pathways for chondrocyte
differentiation
SOX9 signaling, Wnt signaling, bone morphogenetic
protein (BMP) signaling, Indian Hedgehog (Ihh) signaling,
parathyroid hormone-related peptide (PTHrP) signaling, TGF-β
signaling, fibroblast growth factor (FGF) signaling and Runx Family
transcription factors are involved in chondrocyte differentiation
(34,57,60,61). In
addition, adenosine signaling, oxygen tension and reactive oxygen
species are involved in chondrocyte development (60).
SOX9, a member of the SRY-related HMG-box (SOX)
family of transcription factors, is a master regulator of
chondrogenesis, the process of cartilage formation. It plays a
pivotal role in orchestrating the differentiation of MSCs into
chondrocytes, the specialized cells that produce the ECM of
cartilage. SOX9 functions as an early marker of chondrogenic
commitment, initiating a cascade of events that leads to the
expression of genes essential for cartilage development, such as
collagen type II, collagen type IX and aggrecan. Furthermore, SOX9
plays a dual role in chondrocyte maturation, promoting early
chondrocyte hypertrophy while simultaneously inhibiting excessive
hypertrophy, a crucial step in endochondral bone formation. This
intricate balance is achieved through complex interactions with
other transcription factors, including SOX5 and SOX6, and by
modulating various signaling pathways, such as Wnt, TGF-β and BMP.
Numerous skeletal disorders have been linked to the dysregulation
of SOX9 expression or activity, underscoring the vital role of the
protein in preserving skeletal health and cartilage homeostasis
(30,31,57,60).
Wnt signaling influences skeletal development
through multiple mechanisms. β-catenin serves as the central
mediator of canonical Wnt signaling, which inhibits chondrocyte
differentiation while promoting osteoblast differentiation. By
contrast, non-canonical Wnt signaling pathways, involving factors
such as Wnt5a and Wnt11, regulate chondrocyte proliferation and
columnar organization and can also promote chondrocyte
differentiation. Wnt signaling further contributes to the spatial
orientation of chondrocytes within the growth plate, thereby
supporting proper skeletal patterning. The coordinated interplay
between canonical and non-canonical Wnt pathways maintains the
balance between chondrogenesis and osteogenesis, ensuring normal
skeletal development and homeostasis (30,60).
Ihh is a key regulatory molecule in endochondral
ossification. It controls chondrocyte proliferation and maturation
through both PTHrP-dependent and PTHrP-independent mechanisms. Ihh
stimulates PTHrP expression, which in turn delays chondrocyte
hypertrophy. Independent of PTHrP, Ihh directly promotes
chondrocyte proliferation through the activation of GLI family
transcription factors. The overexpression of PTHrP results in
delayed chondrocyte differentiation, whereas the deletion of PTHrP
leads to premature chondrocyte maturation at inappropriate
anatomical locations. Through these coordinated mechanisms, Ihh
ensures proper chondrocyte proliferation, differentiation and
subsequent bone formation during endochondral ossification
(60).
FGFs are essential signaling molecules in skeletal
development (61,62). FGF receptors (FGFR1, FGFR2 and FGFR3)
are expressed at various stages of chondrocyte development and
regulate both proliferation and hypertrophy (62-64).
Among the FGF ligands, FGF9 and FGF18 play particularly critical
roles in chondrogenesis. FGF9 promotes early chondrocyte
proliferation and hypertrophy, and also contributes to angiogenesis
(62,64). In later developmental stages, FGF18
exhibits a dual function, initially stimulating proliferation and
hypertrophy, followed by suppression of proliferation and delayed
hypertrophy (60).
BMPs are critical regulators of chondrogenesis. BMP
signaling is mediated through the receptors BMPR1 and BMPR2, the
latter possessing serine/threonine kinase activity. The activation
of BMP receptors induces the phosphorylation of SMAD transcription
factors, which then form complexes with co-SMADs and translocate to
the nucleus to regulate the expression of target genes, including
Runx2, SOX9 and β-catenin. β-catenin can, in turn, modulate the
expression of SOX9 and Runx2, both of which are essential for
chondrogenesis. In addition to SOX9, SOX5 and SOX6 are also
required for proper chondrocyte differentiation. SOX9 is
particularly critical during early chondrogenesis, as it promotes
the expression of genes involved in cartilage matrix synthesis. The
importance of BMP signaling in endochondral ossification is
highlighted by evidence indicating that the inhibition of BMP
signaling or genetic deletion of BMP receptors results in impaired
chondrocyte formation and maturation (60).
Runx2 and Runx3, members of the Runx transcription
factor family, are essential regulators of chondrocyte hypertrophy.
Runx2 plays a central role by directly inducing Ihh expression,
interacting with BMP-regulated SMADs, and activating hypertrophic
marker genes. Runx3 deficiency results in a relatively mild
phenotype, whereas Runx2 deficiency markedly impairs chondrocyte
hypertrophy. However, the simultaneous absence of both Runx2 and
Runx3 completely abolishes chondrocyte maturation, demonstrating
their complementary and indispensable roles in this developmental
process. Runx2 is particularly involved in the later stages of
chondrogenesis and contributes to hypertrophic differentiation
(34,60).
In contrast to adult BM-MSCs, which require TGF-β
for maintenance in defined culture conditions (65), PSC-derived chondrogenic cells, such
as those derived from neural crest-like progenitors, may exhibit
distinct growth regulatory mechanisms. When expanded in the
presence of SB431542, these cells demonstrated sustained
chondrogenic activity and reduced dependence on TGF-β signaling
(66). This finding suggests that
TGF-β may function as an intrinsic regulatory ‘brake’ on the
proliferation of certain PSC-derived embryonic cells, similar to
its established role in controlling endothelial cell growth
(67) and hematopoietic progenitor
proliferation (68). Notably,
although both PSC-derived ectomesenchymal cells and BM-MSCs have
been proposed to share neural crest origins (69-71),
their differential responses to TGF-β signaling indicate
fundamental differences in their growth characteristics and
differentiation potential.
9. Influence of 2D and 3D cell culture
systems on chondrogenesis
Cell culture conditions play a critical role in
regulating cellular phenotype, differentiation potential and ECM
production during cartilage tissue engineering. Traditional
two-dimensional (2D) monolayer culture systems are widely used for
cell expansion due to their simplicity and ease of manipulation.
However, these systems often fail to preserve the native
chondrocytic phenotype. Chondrocytes expanded in 2D culture
frequently undergo dedifferentiation, which is characterized by the
acquisition of a fibroblast-like morphology and reduced expression
of cartilage-specific markers such as type II collagen and
aggrecan, accompanied by increased expression of fibrocartilaginous
markers, including type I collagen (72,73).
Similarly, prolonged expansion of MSCs under 2D culture conditions
may alter their gene expression profiles and diminish their
subsequent chondrogenic differentiation potential.
In contrast, two-dimensional (3D) culture systems
including pellet cultures, hydrogels and scaffold-based matrices
more closely mimic the native cartilage microenvironment by
facilitating spatial cell-cell and cell-matrix interactions. These
interactions are essential for mesenchymal condensation and the
activation of chondrogenic transcriptional programs regulated by
factors, such as SOX9. Cells cultured in 3D environments typically
adopt a rounded morphology similar to native chondrocytes and
exhibit enhanced expression of cartilage-specific genes, including
COL2A1, aggrecan and SOX9, as well as increased deposition of ECM
components, such as glycosaminoglycan (GAG) and type II collagen
(74,75). Moreover, 3D culture conditions
promote more physiologically relevant biochemical and biomechanical
cues, enabling the development of tissue constructs with structural
and functional properties closer to native cartilage. For these
reasons, 3D culture platforms are widely regarded as more suitable
for cartilage regeneration studies. They are often integrated with
emerging technologies such as 3D bioprinting to generate biomimetic
cartilage constructs.
10. 3D bioprinting
Bioprinting is an emerging technology that utilizes
bioinks, composed of biomaterials and living cells to fabricate 3D
tissue constructs for regenerative medicine. These bioinks should
possess appropriate mechanical and rheological properties to ensure
printing accuracy, structural stability and the functional
performance of the resulting tissues (71). Biofabrication is a broader and
rapidly evolving field that focuses on the generation of
biologically functional constructs with hierarchical organization
(76,77). It encompasses a wide range of
technologies used to create biological substitutes, including
conventional tissue engineering strategies, such as scaffold
fabrication and cell seeding. Within this framework, bioprinting
represents a specialized subset of biofabrication that specifically
refers to the layer-by-layer fabrication of 3D tissue and organ
constructs using cell-laden bioinks.
3D bioprinting, a key modality within
biofabrication, enables the production of constructs with precise
spatial resolution, defined geometry and tunable mechanical
properties. This technique allows for the accurate placement of
biomaterials, cells and biological cues in a controlled,
layer-by-layer manner, rendering it a promising strategy for the
development of personalized regenerative implants (78). Articular cartilage, a thin and
avascular connective tissue, represents a relatively suitable
target for bioprinted regenerative therapies compared with highly
vascularized organs. Due to its low cell density and the lack of
vascular supply, cartilage has limited intrinsic regenerative
capacity (79). Consequently,
untreated articular cartilage injuries often progress to OA
(80).
Bioprinting technologies rely on computer-aided
design models to define the architecture of printed constructs.
Medical imaging modalities, including computed tomography (CT),
magnetic resonance imaging (MRI) and X-ray imaging, provide
detailed anatomical information that can be converted into
high-resolution 2D image slices and subsequently reconstructed into
3D digital models suitable for additive manufacturing and
stereolithography-based applications. Such 3D-printed anatomical
models are widely used in surgical planning and medical education,
particularly for complex anatomical structures. Overall, the major
advantages of bioprinting include high reproducibility, precise
control over construct architecture, and the potential for scalable
and high-throughput production. These features position bioprinting
as a promising platform in tissue engineering and regenerative
medicine for generating functional tissue substitutes. Bioprinting
technologies are generally classified into four principal
categories: Extrusion-based bioprinting, inkjet bioprinting,
laser-assisted bioprinting and stereolithography-based bioprinting
(76,81-85).
Each technique operates through distinct mechanisms and provides
specific advantages and limitations in terms of printing
resolution, cell viability and material compatibility.
Extrusion-based bioprinting is the most widely
applied technique in cartilage tissue engineering due to its
ability to process high-viscosity bioinks and cell-laden hydrogels.
In this approach, bioinks are extruded through a nozzle using
pneumatic, piston, or screw-driven pressure to generate continuous
filaments that are deposited layer by layer to form 3D structures.
This method supports a wide range of biomaterials and enables
printing at relatively high cell densities. However, shear stress
generated during extrusion may negatively affect cell viability,
and the printing resolution is typically limited by the nozzle
diameter (86).
Inkjet bioprinting deposits bioink droplets through
thermal or piezoelectric actuation in a non-contact manner. This
approach provides a high printing speed and good spatial
resolution; however, it is generally restricted to low-viscosity
bioinks and relatively low cell densities, which may limit its
ability to generate mechanically robust cartilage constructs.
Laser-assisted bioprinting utilizes laser pulses to create
localized pressure that propels droplets of bioink onto a receiving
substrate without the use of a nozzle. This nozzle-free technique
minimizes mechanical stress on cells and enables high spatial
precision with excellent cell viability. However, the high cost and
technical complexity of the system limit its widespread application
(87).
Stereolithography-based bioprinting relies on
light-induced photopolymerization to solidify photosensitive
bioinks layer by layer, enabling the fabrication of constructs with
high structural fidelity and resolution. Nevertheless, this method
is typically limited to photo-crosslinkable biomaterials and may
not be suitable for bioinks containing very high cell densities.
Among these techniques, extrusion-based bioprinting remains the
most commonly used approach in cartilage tissue engineering because
it allows the deposition of high-viscosity hydrogel bioinks
containing chondrocytes or mesenchymal stem cells, which are
essential for generating mechanically stable and biologically
functional cartilage constructs.
11. Bioink
3D bioprinting utilizes bioinks, which are printable
biomaterials designed for the fabrication of living tissues.
Although numerous biomaterials have been investigated for the
repair of diseased or damaged tissues, the majority remain
incompatible with current bioprinting technologies (88). Bioinks are broadly categorized into
two main types: Scaffold-based and scaffold-free systems.
Scaffold-free bioinks mimic aspects of embryonic development and
neo-tissue formation. This strategy relies on tissue spheroids,
cell pellets, or tissue strands to generate large-scale functional
constructs without the use of an exogenous scaffold (89). By contrast, scaffold-based bioinks
incorporate cells within supportive matrices such as hydrogels,
microcarriers, or decellularized ECM components. These matrices
provide structural support and promote cell proliferation,
differentiation and tissue maturation. Various bioprinting
techniques have been developed to optimize print fidelity and
spatial resolution when using these bioink systems (76).
Initially, additive manufacturing technologies were
developed for non-biological applications. Common materials used in
traditional 3D printing, including metals, ceramics and
thermoplastic polymers, are generally unsuitable for biological
applications as they often require high temperatures, organic
solvents, or harsh crosslinking conditions that are incompatible
with living cells. One of the greatest challenges in fabricating
functional tissues and organs using 3D printing lies in replicating
the mechanical, chemical and morphological properties of native
tissues. Bioinks play a critical role in overcoming these
challenges by protecting encapsulated cells from mechanical stress
during extrusion and from adverse environmental conditions during
the printing process (90).
An ideal bioink should possess several essential
characteristics, including printability, high mechanical integrity,
structural stability, resistance to dissolution in culture media, a
biodegradation rate compatible with tissue regeneration,
non-toxicity, non-immunogenicity, and the capacity to promote cell
adhesion and proliferation. Additionally, bioink materials should
be cost-effective and amenable to scalable manufacturing (91). Over the past decade, 3D printing
technologies have rapidly expanded and become a cornerstone of
tissue engineering research (92). A
wide range of materials including ceramics, polymers, elastomers,
hydrogels and lipids have been explored as bioinks for fabricating
3D constructs. Advanced bioinks can be classified into five main
categories: Multi-material bioinks, stimuli-responsive bioinks,
self-assembling bioinks, biomolecular bioinks and nanoengineered
bioinks, many of which exhibit shear-thinning properties that
facilitate extrusion-based printing (76).
In recent years, scaffold-based and scaffold-free
bioinks have emerged as the two principal strategies in
bioprinting. Scaffold-based systems consist of biomaterials
combined with cells that are co-deposited to form a construct, with
the biomaterial serving as structural support to promote cell
proliferation and differentiation (6,93). By
contrast, scaffold-free systems rely on cellular aggregates, such
as spheroids, tissue strands or cell pellets that secrete ECM-like
components to maintain structural integrity (94). Although scaffold-based approaches
remain the most widely used, both strategies present distinct
advantages and limitations.
The functional performance of bioprinted tissues
depends on the physicochemical and biological properties of the
bioink. For cartilage tissue engineering in particular, bioinks
should meet both material and biological requirements. Key
characteristics include appropriate printability, bioresorbability
and biodegradability (6). Additional
desirable features include high printing resolution,
cost-effectiveness, industrial scalability and rapid post-printing
maturation. In hydrogel-based systems, other key attributes include
reversible gelation (facilitating pre-culture prior to
implantation), rapid gelation kinetics and minimal volumetric
changes during crosslinking (95).
Successful 3D cell printing also requires bioinks
with optimal viscosity to ensure smooth extrusion, while
maintaining structural fidelity after deposition. Following
printing, the material must undergo rapid stabilization, typically
through solvent evaporation or polymer crosslinking to maintain the
intended 3D architecture. Notably, bioinks must remain
cytocompatible and non-toxic throughout both the printing and
crosslinking processes. Although certain robust materials require
high temperatures or toxic solvents for processing, such conditions
are generally incompatible with cell-laden bioprinting applications
(96).
In stem cell bioprinting, the preservation of cell
viability during printing and provision of a supportive
microenvironment for post-printing growth and differentiation are
essential. This requirement typically favors aqueous,
hydrogel-based bioinks. Both natural and synthetic polymers are
widely used. Natural ECM-derived polymers such as collagen, fibrin
and gelatin are particularly attractive due to their inherent
biocompatibility and ability to mimic the native cellular
microenvironment. Other natural polymers used as bioinks include
chitosan and alginate. Synthetic polymers, such as Pluronic F127,
polyethylene oxide and polyethylene glycol (PEG), provide tunable
mechanical and physicochemical properties. While natural polymers
provide excellent biocompatibility and cell-supportive properties,
they often exhibit limited mechanical strength and structural
stability. Therefore, they are frequently combined with synthetic
polymers to enhance versatility, stability and mechanical
performance in 3D bioprinting applications (86).
Bioinks designed for stem cell-based 3D bioprinting
need to achieve a balance between printability, biocompatibility,
and the capacity to support cell growth and function. Although
natural polymers, such as alginate, chitosan, agarose, hyaluronic
acid (HA) and fibrin provide strong biocompatibility, they may
require chemical or physical modification to improve mechanical
strength or printing resolution. For instance, alginate undergoes
rapid gelation in the presence of calcium ions, rendering it
suitable for high-throughput bioprinting (94,97-100).
Fibrin provides rapid crosslinking but has relatively low
mechanical strength and is often blended with other polymers to
enhance structural stability. Synthetic polymers, such as Pluronic
F127 and PEG provide adjustable properties and can be chemically
modified for specific applications. Ultimately, the choice and
composition of bioink materials should be tailored to the specific
cell type and target tissue being engineered (96,100).
Natural and synthetic hydrogels are widely used as
bioink materials in cartilage tissue engineering due to their
ability to mimic the hydrated ECM environment required for
chondrocyte survival and differentiation. Natural biomaterials,
such as alginate, gelatin, collagen, HA, agarose and chitosan are
particularly attractive as they exhibit excellent biocompatibility,
bioactivity and structural similarity to native cartilage ECM.
These materials often contain inherent cell-binding motifs that
support cell adhesion, proliferation and chondrogenic
differentiation. For example, alginate is widely used due to its
rapid ionic crosslinking and ability to maintain the rounded
morphology of chondrocytes, which is critical for preserving their
phenotype. Similarly, gelatin- and collagen-based hydrogels provide
cell-recognition sites that enhance cell attachment and ECM
deposition during cartilage formation (76,91).
However, natural hydrogels also present several
limitations, including relatively weak mechanical strength, rapid
degradation and batch-to-batch variability. These drawbacks can
compromise the structural integrity of printed constructs,
particularly when engineering load-bearing tissues such as
articular cartilage. In addition, although the majority of natural
polymers are biocompatible, certain materials may trigger mild
immune responses depending on their source and purification process
(101,102).
Synthetic polymers such as PEG, polycaprolactone
(PCL) and pluronic-based hydrogels have therefore been explored to
overcome these limitations. Synthetic bioinks provide highly
tunable mechanical properties, controlled degradation rates and
improved printability, rendering them suitable for fabricating
mechanically stable constructs. PEG-based hydrogels, for example,
allow for the precise modulation of stiffness and crosslinking
density, which can influence stem cell fate and chondrogenic
differentiation. Nevertheless, synthetic biomaterials generally
lack intrinsic bioactive signals required for cell adhesion and
matrix production, often necessitating modification with bioactive
peptides or blending with natural polymers to enhance cellular
responses (90,102). Hybrid bioinks that combine natural
and synthetic components such as gelatin methacrylate (GelMA),
alginate-gelatin blends and HA-PEG systems are therefore
increasingly investigated as promising candidates for cartilage
regeneration as they provide a balance between mechanical
stability, printability and biological functionality.
12. Lubrication
The mucinous glycoprotein lubricin, encoded by the
PRG4 gene is a principal boundary lubricant in diarthrodial joints.
Lubricin is an elongated molecule (~200 nm in length and a ~1-5 nm
in width) localized to the superficial zone of articular cartilage
and present in synovial fluid (103-105).
Its role in cartilage lubrication was first proposed by Radin et
al (106) in 1970, and the term
‘lubricin’ was coined in 1981 following its identification as a
mucin-like glycoprotein (107).
Data from the initial study by Radin et al (106) demonstrated that a lubricin solution
alone (without HA) reduced the coefficient of friction by ~2-fold
compared with saline. Subsequent studies consistently confirmed
that lubricin is a key mediator of boundary lubrication in synovial
joints (108-114).
Lubricin deficiency has been associated with severe joint
disorders, including camptodactyly-arthropathy-coxa
vara-pericarditis syndrome, which is characterized by accelerated
joint degeneration. In such cases, lubrication supplementation has
been shown to attenuate disease progression (112,115).
Cartilage degeneration typically begins at the
superficial zone and progresses to deeper layers, including the
subchondral bone, representing an early event in the pathogenesis
of OA (4,116). Articular cartilage regeneration,
particularly through human iPSC-derived cartilage, represents a
rational patient-specific strategy that may overcome several
limitations of conventional therapies (117). The development of 3D bioprinting
technologies has further accelerated progress in this field.
3D bioprinting involves the encapsulation of cells,
such as human iPSCs, human chondrocytes, or human MSCs within
hydrogel-based bioinks (e.g., gelatin and alginate), and in some
cases, nanomaterial-enhanced formulations (101,118).
Ex vivo research has demonstrated the promising potential of
3D bioprinting for generating articular cartilage constructs
(119), and various hydrogel
systems have been evaluated in vivo to improve mechanical
integrity and functional performance (120).
Targeting lubricin secretion has emerged as a major
strategy for restoring the functional lubrication properties of
regenerated cartilage, as native joint lubrication is highly
dependent on PRG4 expression and lubricin production. Recent 3D
bioprinting approaches have therefore focused on optimizing bioink
formulations and encapsulated cells to enhance lubricin secretion
(121,122). Lubricin functions as a protective
molecule within synovial fluid, and recombinant lubricin has been
shown to attenuate the onset of OA in experimental models (109,123).
An optimized bioink formulation consisting of 14% GelMA and 2%
oxidized methacrylated alginate has been identified as effective in
promoting lubricin secretion while maintaining structural stability
(122). This strategy may enhance
the long-term success of cartilage regeneration. Further
investigations are warranted to evaluate additional factors
influencing lubrication, including biomaterial stiffness and growth
factor incorporation. Moreover, iPSC-derived microtissues have been
generated using nanofibrillated cellulose and alginate-based
bioinks for in vivo mouse models, with modifications
designed to stimulate aggrecan production, a key GAG involved in
cartilage lubrication (124).
In addition to biological strategies, biomimetic
material design has provided alternative approaches to restoring
lubrication. A series of studies have described the synthesis of a
polymer that mimics the distinctive bottlebrush architecture of
lubricin, which is critical for its boundary lubrication function.
This biomimetic polymer, synthesized from 2-hydroxyethyl acrylate
and 2-methacryloyloxyethyl phosphorylcholine, effectively replaced
natural lubricin when adsorbed onto damaged cartilage surfaces. The
system achieved an extremely low coefficient of friction,
comparable to that of native lubricin (125-127).
These findings highlight the potential of advanced material design
to generate high-performance, non-biological components for
integration into bioprinted constructs and for improving the
lubrication of damaged cartilage. Furthermore, in vitro
research has demonstrated that gelatin-based hydrogels
incorporating zwitterionic phosphocholine groups can enhance
surface organization and improve native lubrication properties
(128).
13. Comparison of 3D bioprinting with
current treatments for OA and stem cell therapy
Bioprinting, an advanced technology that integrates
biology with 3D printing, has transformed multiple medical fields,
including drug delivery, tissue engineering and regenerative
medicine. Bioprinted biomaterials provide a novel therapeutic
strategy for OA, in contrast to conventional treatments, such as
analgesic administration and joint replacement surgery, which are
often associated with limited long-term efficacy and potential
complications (129).
Several studies have demonstrated that stem cell
therapy can promote cartilage regeneration with partial success
(130). However, stem cell-based
approaches present several limitations, including limited control
over cell differentiation, poor cell survival and integration, and
variability in stem cell activity that complicates the prediction
of therapeutic outcomes and ethical concerns associated with the
use of ESCs. ASCs, due to their multipotent nature, possess
restricted differentiation capacity, whereas ESCs exhibit
pluripotency and can differentiate into a broad range of cell
types. Nevertheless, ethical concerns surrounding the use of ESCs
have limited their clinical application.
To address these challenges, innovative strategies,
such as the integration of 3D bioprinting with iPSCs have been
proposed. iPSCs are generated by reprogramming mature somatic
cells, such as dermal fibroblasts, into a pluripotent state. This
approach provides a virtually unlimited and patient-specific cell
source for regenerative applications, while avoiding the ethical
concerns associated with ESCs (35,87,131,132).
Importantly, previous studies focusing specifically
on cartilage tissue engineering have demonstrated improved outcomes
when using 3D bioprinted constructs compared with conventional
cell-based therapies, particularly in terms of enhanced cell
viability, spatial organization and extracellular matrix production
(133-137).
For example, bioprinted cartilage constructs containing MSCs and
hydrogel bioinks have been reported to achieve cell viability rates
typically exceeding 80-90% after printing, while maintaining high
chondrogenic differentiation capacity. In several in vitro
and animal studies, bioprinted cartilage constructs have shown
significant increases in GAG and collagen type II production
compared with traditional scaffold-based or cell injection
approaches (135,136,138).
Furthermore, preclinical studies using bioprinted MSC-laden
hydrogels have demonstrated cartilage defect filling and tissue
regeneration efficiencies of almost 70-80%, whereas conventional
microfracture or cell injection therapies often result in
fibrocartilage formation and regeneration rates <40-50%
(91,134,135).
These findings highlight the improved structural organization and
functional matrix production achieved through spatially controlled
cell deposition in bioprinted constructs.
Furthermore, this technology enables the fabrication
of complex 3D structures that closely replicate the architecture
and microenvironment of native tissues. The precise spatial
deposition of cells and biomaterials during bioprinting facilitates
the generation of constructs that mimic the native joint
microenvironment (102). By
reproducing the intricate structure of affected cartilage, 3D
bioprinting has the potential to promote tissue regeneration and
potentially slow disease progression.
One of the most critical advantages of bioprinting
is its capacity for personalized therapy (139). Bioprinted biomaterials can be
tailored to the specific anatomical and physiological
characteristics of each patient. By incorporating patient-specific
parameters, such as joint geometry, defect size and disease
severity, bioprinting enables the fabrication of customized
constructs designed to optimize clinical outcomes (140). In addition, high-resolution imaging
modalities, such as CT and MRI can be integrated into the
bioprinting workflow to generate constructs that accurately
replicate the geometry of cartilage defects and improve anatomical
integration with host tissue (141-143)
3D bioprinting is also highly automated and
scalable, enabling the relatively rapid fabrication of tissue
constructs. Unlike conventional stem cell therapy, which often
involves the transplantation of pre-differentiated chondrocytes,
bioprinting allows stem cells within the printed construct to
undergo proliferation and chondrogenic differentiation in
situ following implantation. Bioinks typically consist of
combinations of natural and synthetic polymers that provide a
supportive ECM enriched with nutrients and growth factors. This
microenvironment enhances cell viability and may promote more
sustained and long-term regeneration compared with traditional cell
injection approaches. Additionally, bioprinting allows for the
spatial incorporation of therapeutic agents within the bioink,
enabling the design of controlled and localized drug release
profiles. These release patterns can be tailored to target areas of
inflammation or cartilage degeneration according to
patient-specific needs. Bioinks can also be engineered to replicate
the mechanical properties of native cartilage, thereby providing
structural support while facilitating tissue regeneration (144).
Despite its significant promise, 3D bioprinting in
regenerative medicine continues to face important challenges. These
include the optimization of bioink formulations, achieving adequate
vascularization in larger constructs, addressing the technical
complexity of bioprinting processes, and overcoming high production
costs. Continued research and technological refinement are required
to translate this promising approach into widespread clinical
application.
14. Future directions
Future directions in cartilage bioprinting focus on
developing techniques capable of generating more functional,
durable and physiologically relevant tissue constructs. One
critical strategy involves the use of multi-material bioprinting
approaches to replicate the natural gradients present within
cartilage. Native cartilage exhibits region-specific variations in
mechanical properties, including stiffness and ECM composition. By
printing multiple biomaterials with distinct mechanical
characteristics, it is possible to recreate these gradients within
a single construct, thereby more closely mimicking the hierarchical
structure of native cartilage and improving functional performance
after implantation.
Cartilage is composed not only of chondrocytes, but
also of additional cell types that contribute to tissue development
and maintenance. Fibroblasts and stem cells, for example, play
vital roles in cartilage formation and repair. Accordingly,
co-culture strategies incorporating multiple cell types, such as
chondrocytes, fibroblasts and stem cells within bioprinted
constructs may enhance tissue complexity and more effectively
replicate the native cellular microenvironment. High-resolution
bioprinting further enables the fabrication of intricate and
spatially precise 3D architectures that resemble native cartilage.
Moreover, the integration of real-time monitoring and feedback
systems during printing may allow dynamic adjustments to process
parameters, thereby improving construct fidelity and biological
functionality.
To ensure reproducibility and biological
performance, the optimization of the printing process is essential.
High-precision fabrication of complex structures requires continued
advancements in nozzle design, printing speed, resolution, and
material deposition control (143).
While current research has primarily focused on printing small
tissue constructs, there is increasing interest in generating
entire complex organs such as the heart and brain. Although
progress has been achieved in disease modeling and partial tissue
repair in organs, including complex organs such as the heart,
liver, kidney, and neural tissues (135), fabrication of fully functional
complex organs remains a substantial challenge. In particular,
printing neural tissues such as the brain requires advanced
technologies capable of monitoring functional activity.
Advancements in printing technologies are also being
explored to improve structural accuracy and resolution. The
incorporation of advanced imaging modalities, including CT and MRI,
can facilitate the generation of detailed 3D models of affected
joints, thereby enhancing the anatomical precision of printed
constructs. A growing body of evidence supports the therapeutic
potential of bioprinting-enabled drug delivery systems for the
treatment of OA. Bioprinting allows localized and controlled drug
release within the affected joint, thereby improving targeting
accuracy and reducing systemic side effects. Notably, sustained
drug release profiles can be achieved, maintaining therapeutic
concentrations over extended periods and potentially providing
prolonged symptom relief and disease-modifying effects. In
addition, bioprinted constructs containing chondrocytes or MSCs
have demonstrated the ability to promote cartilage-like tissue
formation and support regenerative processes (145).
Preclinical studies using 3D bioprinting in rabbit
models of joint regeneration have demonstrated high post-printing
cell viability and functional tissue repair. For example, a GelMA
scaffold containing rabbit bone marrow MSCs showed ~81% viability
48 h after bioprinting and supported cartilage regeneration in a
New Zealand rabbit osteochondral defect model (146). Additionally, reviews of 3D
bioprinted cartilage point out that cell viability commonly reaches
~70-85% immediately after printing and can recover to high levels
approaching ~90% within days in similar hydrogel-based constructs
used for orthoregeneration (147,148).
Despite these encouraging results, comprehensive clinical trials
are required to establish the safety, efficacy and long-term
outcomes of bioprinted cartilage implants prior to routine clinical
applications. The incorporation of personalized medicine
strategies, including patient-specific design and fabrication, may
further enhance the success rates of cartilage repair. Furthermore,
long-term durability and performance of bioprinted cartilage
constructs depend on the integration of materials and structural
features that improve lubrication, a critical determinant of joint
function.
A major challenge in bioprinting remains the
development of optimal bioinks. Bioinks, composed of biomaterials,
cells, or their combinations, are essential for the fabrication of
structurally stable and biologically functional constructs. Despite
significant technological advancements, designing bioinks that
simultaneously satisfy mechanical, rheological and biological
requirements remains complex. Ongoing research is therefore
directed toward the development of novel bioink formulations and
the establishment of standardized evaluation methods. A notable
study introduced a method for assessing bioink shape fidelity by
analyzing filament fusion and collapse behavior in 3D-printed
structures, providing valuable insight into bioink suitability for
specific applications (149).
Of note, the translation of bioprinted cartilage
constructs from laboratory research to routine clinical practice
will depend on overcoming several technological and regulatory
barriers. Although promising results have been demonstrated in
vitro and in preclinical animal models, large-scale clinical
implementation requires rigorous evaluation of safety, long-term
functionality and reproducibility. Regulatory approval processes
for bioprinted tissues are particularly complex as these constructs
combine living cells, biomaterials and manufacturing technologies,
placing them at the intersection of medical devices, biologics and
advanced therapy medicinal products. Current regulatory frameworks
from agencies such as the US Food and Drug Administration (FDA) and
the European Medicines Agency (EMA) require extensive preclinical
validation, standardized manufacturing protocols and controlled
clinical trials before approval can be granted. Considering the
current pace of technological development and ongoing preclinical
studies, initial human trials of MSC-laden bioprinted cartilage
constructs may become feasible within the next ~5-10 years, while
broader clinical adoption is expected to take longer, depending on
progress in regulatory approval, manufacturing standardization, and
construct maturation (147,150,151).
Continued advancements in bioink development, printing resolution,
construct maturation, and regulatory standardization will therefore
play a crucial role in accelerating the translation of cartilage
bioprinting technologies into clinical therapies (134,135,152).
15. Conclusion
Bioprinting provides a powerful platform for the
fabrication of functional cartilage tissues through precise control
of cells and biomaterials. The use of iPSCs as a patient-specific
and readily available cell source further enhances the
translational potential of this approach. By rationally designing
bioinks with appropriate mechanical properties, incorporating
specific growth factors and using cues from the native ECM, it is
possible to effectively direct iPSC differentiation toward a
chondrogenic lineage. To achieve long-term functional outcomes,
strategies aimed at enhancing the lubrication properties of
regenerated cartilage are also essential. These may include the
incorporation of fluid-filled microchannels or HA within the
bioprinted construct to better replicate the native joint
environment. Although critical challenges remain, including the
optimization of vascularization and ensuring long-term structural
stability, continued research in this field holds substantial
promise for the development of personalized and effective
therapeutic strategies for cartilage regeneration.
Acknowledgements
The authors would like to thank Dr Sachinthani
Karunarathne, Department of Molecular Biology and Biotechnology,
University of Peradeniya, Peradeniya, Sri Lanka, for her valuable
feedback on the manuscript during its early development.
Funding
Funding: No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
GWNK was involved in the conceptualization of the
study, as well as in the study performing the literature search,
collecting and synthesizing relevant information from previous
studies, and in the writing of the manuscript (original draft
preparation). SR was involved in the reviewing and revising of the
manuscript, and supervised the study. Both authors have read and
approved the final manuscript. Data authentication is not
applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Use of artificial intelligence tools
During the preparation of this work, AI tools were
used solely to improve the readability and language of the
manuscript and for image upscaling, and subsequently, the authors
revised and edited the content as necessary, taking full
responsibility for the ultimate content of the present
manuscript.
References
|
1
|
Owaidah AY: Induced pluripotent stem cells
in cartilage tissue engineering: A literature review. Biosci Rep.
44(BSR20232102)2024.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Hunziker EB: Articular cartilage repair:
Basic science and clinical progress. A review of the current status
and prospects. Osteoarthritis Cartilage. 10:432–463.
2002.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Tsumaki N, Okada M and Yamashita A: iPSC
technologies and cartilage regeneration. Bone. 70:48–54.
2015.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Sophia Fox AJ, Bedi A and Rodeo SA: The
basic science of articular cartilage: Structure, composition, and
function. Sports Health. 1:461–468. 2009.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Massari L, Benazzo F, Falez F, Perugia D,
Pietrogrande L, Setti S, Osti R, Vaienti E, Ruosi C and Cadossi R:
Biophysical stimulation of bone and cartilage: State of the art and
future perspectives. Int Orthop. 43:539–551. 2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Roseti L, Cavallo C, Desando G, Parisi V,
Petretta M, Bartolotti I and Grigolo B: Three-dimensional
bioprinting of cartilage by the use of stem cells: A strategy to
improve regeneration. Materials (Basel). 11(1749)2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Jochimsen KN, Kim JS, Jayabalan P,
Lawrence C, Lewis CL, Prather H and Bostrom MP: Arthritis
foundation/HSS workshop on hip osteoarthritis, Part 3:
Rehabilitation and exercise. HSS J. 19:447–452. 2023.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Bilotsky Y and Gasik M: Modelling of
poro-visco-elastic biological systems. J Phys Conf Ser.
633(012134)2015.
|
|
9
|
Khalid Al-Hayali N, Salman Chiad J, Nacy S
and Hussein O: A review of passive and quasi-passive lower limb
exoskeletons for gait rehabilitation. J Mech Eng Res Dev. 44:73–86.
2021.
|
|
10
|
Weissman IL: Stem cells: Units of
development, units of regeneration, and units in evolution. Cell.
100:157–168. 2000.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Grässel S and Muschter D: Recent advances
in the treatment of osteoarthritis. F1000Res 9: F1000 Faculty
Rev-325, 2020.
|
|
12
|
Yu Y: Articular cartilage tissue
engineering using chondrogenic progenitor cell homing and 3D
bioprinting [dissertation]. University of Iowa, 2015.
|
|
13
|
Rahmadian R, Adly M, Dilogo IH, Revilla G
and Ariliusra Z: Single intra-articular injection of human synovial
membrane MSC from grade IV knee osteoarthritis patient improve
cartilage repair in OA rat model. J Orthop Surg Res.
19(710)2024.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Lee CH, Cook JL, Mendelson A, Moioli EK,
Yao H and Mao JJ: Regeneration of the articular surface of the
rabbit synovial joint by cell homing: A proof of concept study.
Lancet. 376:440–448. 2010.PubMed/NCBI View Article : Google Scholar
|
|
15
|
De Vries-van Melle ML, Tihaya MS, Kops N,
Koevoet WJ, Murphy JM, Verhaar JA, Alini M, Eglin D and van Osch
GJ: Chondrogenic differentiation of human bone marrow-derived
mesenchymal stem cells in a simulated osteochondral environment is
hydrogel dependent. Eur Cell Mater. 27:112–123. 2014.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Wakitani S, Imoto K, Yamamoto T, Saito M,
Murata N and Yoneda M: Human autologous culture expanded bone
marrow mesenchymal cell transplantation for repair of cartilage
defects in osteoarthritic knees. Osteoarthritis Cartilage.
10:199–206. 2002.PubMed/NCBI View Article : Google Scholar
|
|
17
|
De Bari C and Roelofs AJ: Stem cell-based
therapeutic strategies for cartilage defects and osteoarthritis.
Curr Opin Pharmacol. 40:74–80. 2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Makris EA, Gomoll AH, Malizos KN, Hu JC
and Athanasiou KA: Repair and tissue engineering techniques for
articular cartilage. Nat Rev Rheumatol. 11:21–34. 2015.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Foster CS, Dodson A, Karavana V, Smith PH
and Ke Y: An introduction to stem cells. J Pathol. 197:419–423.
2002.
|
|
20
|
Thomson JA, Itskovitz-Eldor J, Shapiro SS,
Waknitz MA, Swiergiel JJ, Marshall VS and Jones JM: Embryonic stem
cell lines derived from human blastocysts. Science. 282:1145–1147.
1998.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Takahashi K and Yamanaka S: Induction of
pluripotent stem cells from mouse embryonic and adult fibroblast
cultures by defined factors. Cell. 126:663–676. 2006.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Gadue P, Huber TL, Nostro MC, Kattman S
and Keller GM: Germ layer induction from embryonic stem cells. Exp
Hematol. 33:955–964. 2005.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Nishikawa S, Goldstein RA and Nierras CR:
The promise of human induced pluripotent stem cells for research
and therapy. Nat Rev Mol Cell Biol. 9:725–729. 2008.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Punwar S and Khan WS: Mesenchymal stem
cells and articular cartilage repair: Clinical studies and future
direction. Open Orthop J. 5:327–333. 2011.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Dowthwaite GP, Bishop JC, Redman SN, Khan
IM, Rooney P, Evans DJ, Haughton L, Bayram Z, Boyer S, Thomson B,
et al: The surface of articular cartilage contains a progenitor
cell population. J Cell Sci. 117:889–897. 2004.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Alsalameh S, Amin R, Gemba T and Lotz M:
Identification of mesenchymal progenitor cells in normal and
osteoarthritic human articular cartilage. Arthritis Rheum.
50:1522–1532. 2004.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Williams R, Khan IM, Richardson K, Nelson
L, McCarthy HE, Analbelsi T, Singhrao SK, Dowthwaite GP, Jones RE,
Baird DM, et al: Identification and clonal characterisation of a
progenitor cell sub-population in normal human articular cartilage.
PLoS One. 5(e13246)2010.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Rikkers M, Korpershoek JV, Levato R, Malda
J and Vonk LA: The clinical potential of articular
cartilage-derived progenitor cells: A systematic review. NPJ Regen
Med. 7(2)2022.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Sekiya I, Larson BL, Smith JR, Pochampally
R, Cui JG and Prockop DJ: Expansion of human adult stem cells from
bone marrow stroma: Conditions that maximize the yields of early
progenitors and evaluate their quality. Stem Cells. 20:530–541.
2002.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Chen H, Tan XN, Hu S, Liu RQ, Peng LH, Li
YM and Wu P: Molecular mechanisms of chondrocyte proliferation and
differentiation. Front Cell Dev Biol. 9(664168)2021.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Tian B, Zhang L, Zheng J and Kang X: The
role of NF-κB-SOX9 signalling pathway in osteoarthritis. Heliyon.
10(e37191)2024.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Denker AE, Haas AR, Nicoll SB and Tuan RS:
Chondrogenic differentiation of murine C3H10T1/2 multipotential
mesenchymal cells: I. Stimulation by bone morphogenetic protein-2
in high-density micromass cultures. Differentiation. 64:67–76.
1999.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Klampfleuthner FAM, Lotz B, Renkawitz T,
Richter W and Diederichs S: Stage-dependent activity and
pro-chondrogenic function of PI3K/AKT during cartilage neogenesis
from mesenchymal stromal cells. Cells. 11(2965)2022.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Studer D, Millan C, Öztürk E,
Maniura-Weber K and Zenobi-Wong M: Molecular and biophysical
mechanisms regulating hypertrophic differentiation in chondrocytes
and mesenchymal stem cells. Eur Cell Mater. 24:118–135.
2012.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Magne D, Vinatier C, Julien M, Weiss P and
Guicheux J: Mesenchymal stem cell therapy to rebuild cartilage.
Trends Mol Med. 11:519–526. 2005.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Barberi T, Willis LM, Socci ND and Studer
L: Derivation of multipotent mesenchymal precursors from human
embryonic stem cells. PLoS Med. 2(e161)2005.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Olivier EN, Rybicki AC and Bouhassira EE:
Differentiation of human embryonic stem cells into bipotent
mesenchymal stem cells. Stem Cells. 24:1914–1922. 2006.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Trivedi P and Hematti P: Derivation and
immunological characterization of mesenchymal stromal cells from
human embryonic stem cells. Exp Hematol. 36:350–359.
2008.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Gruenloh W, Kambal A, Sondergaard C, McGee
J, Nacey C, Kalomoiris S, Pepper K, Olson S, Fierro F and Nolta JA:
Characterization and in vivo testing of mesenchymal stem cells
derived from human embryonic stem cells. Tissue Eng Part A.
17:1517–1525. 2011.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Lian Q, Lye E, Suan Yeo K, Khia Way Tan E,
Salto-Tellez M, Liu TM, Palanisamy N, El Oakley RM, Lee EH, Lim B
and Lim SK: Derivation of clinically compliant MSCs from CD105+,
CD24-differentiated human ESCs. Stem Cells. 25:425–436.
2007.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Lian I, Kim J, Okazawa H, Zhao J, Zhao B,
Yu J, Chinnaiyan A, Israel MA, Goldstein LS, Abujarour R, et al:
The role of YAP transcription coactivator in regulating stem cell
self-renewal and differentiation. Genes Dev. 24:1106–1118.
2010.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Chen YS, Pelekanos RA, Ellis RL, Horne R,
Wolvetang EJ and Fisk NM: Small molecule mesengenic induction of
human induced pluripotent stem cells to generate mesenchymal
stem/stromal cells. Stem Cells Transl Med. 1:83–95. 2012.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Nakagawa T, Lee SY and Reddi AH: Induction
of chondrogenesis from human embryonic stem cells without embryoid
body formation by bone morphogenetic protein 7 and transforming
growth factor beta1. Arthritis Rheum. 60:3686–3692. 2009.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Diederichs S and Tuan RS: Functional
comparison of human-induced pluripotent stem cell-derived
mesenchymal cells and bone marrow-derived mesenchymal stromal cells
from the same donor. Stem Cells Dev. 23:1594–1610. 2014.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Xu M, Shaw G, Murphy M and Barry F:
Induced pluripotent stem cell-derived mesenchymal stromal cells are
functionally and genetically different from bone marrow-derived
mesenchymal stromal cells. Stem Cells. 37:754–765. 2019.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Caplan AI: Mesenchymal stem cells. J
Orthop Res. 9:641–650. 1991.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Pittenger MF, Mackay AM, Beck SC, Jaiswal
RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S and
Marshak DR: Multilineage potential of adult human mesenchymal stem
cells. Science. 284:143–147. 1999.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Bonab MM, Alimoghaddam K, Talebian F,
Ghaffari SH, Ghavamzadeh A and Nikbin B: Aging of mesenchymal stem
cell in vitro. BMC Cell Biol. 7(14)2006.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Dell'Accio F, De Bari C and Luyten FP:
Molecular markers predictive of the capacity of expanded human
articular chondrocytes to form stable cartilage in vivo. Arthritis
Rheum. 44:1608–1619. 2001.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Dell'Accio F, Vanlauwe J, Bellemans J,
Neys J, De Bari C and Luyten FP: Expanded phenotypically stable
chondrocytes persist in the repair tissue and contribute to
cartilage matrix formation and structural integration in a goat
model of autologous chondrocyte implantation. J Orthop Res.
21:123–131. 2003.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Robertson JA: Embryo culture and the
‘culture of life’: Constitutional issues in the embryonic stem cell
debate. U Chi Legal Forum. 1(40)2006.
|
|
52
|
Lo B and Parham L: Ethical issues in stem
cell research. Endocr Rev. 30:204–213. 2009.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Medvedev SP, Shevchenko AI and Zakian SM:
Induced pluripotent stem cells: Problems and advantages when
applying them in regenerative medicine. Acta Naturae. 2:18–28.
2010.PubMed/NCBI
|
|
54
|
Pezawas T, Diedrich A, Robertson D, Winker
R, Richter B, Wang L and Schmidinger H: Risk of arrhythmic death in
ischemic heart disease: A prospective, controlled, observer-blind
risk stratification over 10 years. Eur J Clin Invest. 47:231–240.
2017.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Eremeev A, Pikina A, Ruchko Y and
Bogomazova A: Clinical potential of cellular material sources in
the generation of iPSC-based products for the regeneration of
articular cartilage. Int J Mol Sci. 24(14408)2023.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Nakayama N, Pothiawala A, Lee JY, Matthias
N, Umeda K, Ang BK, Huard J, Huang Y and Sun D: Human pluripotent
stem cell-derived chondroprogenitors for cartilage tissue
engineering. Cell Mol Life Sci. 77:2543–2563. 2020.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Almalki SG and Agrawal DK: Key
transcription factors in the differentiation of mesenchymal stem
cells. Differentiation. 92:41–51. 2016.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Fakhry M: Molecular mechanisms of
mesenchymal stem cell differentiation towards osteoblasts. World J
Stem Cells. 5(136)2013.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Lach M, Trzeciak T, Richter M, Pawlicz J
and Suchorska WM: Directed differentiation of induced pluripotent
stem cells into chondrogenic lineages for articular cartilage
treatment. J Tissue Eng. 5(2041731414552701)2014.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Li J and Dong S: The signaling pathways
involved in chondrocyte differentiation and hypertrophic
differentiation. Stem Cells Int. 2016(2470351)2016.PubMed/NCBI View Article : Google Scholar
|
|
61
|
West-Livingston LN, Park J, Lee SJ, Atala
A and Yoo JJ: The role of the microenvironment in controlling the
fate of bioprinted stem cells. Chem Rev. 120:11056–11092.
2020.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Jacob AL, Smith C, Partanen J and Ornitz
DM: Fibroblast growth factor receptor 1 signaling in the
osteo-chondrogenic cell lineage regulates sequential steps of
osteoblast maturation. Dev Biol. 296:315–328. 2006.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Ornitz DM and Marie PJ: FGF signaling
pathways in endochondral and intramembranous bone development and
human genetic disease. Genes Dev. 16:1446–1465. 2002.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Minina E, Kreschel C, Naski MC, Ornitz DM
and Vortkamp A: Interaction of FGF, Ihh/Pthlh, and BMP signaling
integrates chondrocyte proliferation and hypertrophic
differentiation. Dev Cell. 3:439–450. 2002.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Ng F, Boucher S, Koh S, Sastry KS, Chase
L, Lakshmipathy U, Choong C, Yang Z, Vemuri MC, Rao MS and Tanavde
V: PDGF, TGF-beta, and FGF signaling is important for
differentiation and growth of mesenchymal stem cells (MSCs):
Transcriptional profiling can identify markers and signaling
pathways important in differentiation of MSCs into adipogenic,
chondrogenic, and osteogenic lineages. Blood. 112:295–307.
2008.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Umeda K, Oda H, Yan Q, Matthias N, Zhao J,
Davis BR and Nakayama N: Long-term expandable SOX9+ chondrogenic
ectomesenchymal cells from human pluripotent stem cells. Stem Cell
Reports. 4:712–726. 2015.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Watabe T, Nishihara A, Mishima K,
Yamashita J, Shimizu K, Miyazawa K, Nishikawa S and Miyazono K:
TGF-beta receptor kinase inhibitor enhances growth and integrity of
embryonic stem cell-derived endothelial cells. J Cell Biol.
163:1303–1311. 2003.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Batard P, Monier MN, Fortunel N, Ducos K,
Sansilvestri-Morel P, Phan T, Hatzfeld A and Hatzfeld JA:
TGF-(beta)1 maintains hematopoietic immaturity by a reversible
negative control of cell cycle and induces CD34 antigen
up-modulation. J Cell Sci. 113:383–390. 2000.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Takashima Y, Era T, Nakao K, Kondo S,
Kasuga M, Smith AG and Nishikawa SI: Neuroepithelial cells supply
an initial transient wave of MSC differentiation. Cell.
129:1377–1388. 2007.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Morikawa S, Mabuchi Y, Kubota Y, Nagai Y,
Niibe K, Hiratsu E, Suzuki S, Miyauchi-Hara C, Nagoshi N, Sunabori
T, et al: Prospective identification, isolation, and systemic
transplantation of multipotent mesenchymal stem cells in murine
bone marrow. J Exp Med. 206:2483–2496. 2009.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Nagoshi N and Okano H: Characterisation of
neural crest-derived stem cells in different tissues. In: Neural
crest stem cells: Breakthroughs and applications. World Scientific
Publishing Co., pp87-107, 2012.
|
|
72
|
Benya PD and Shaffer JD: Dedifferentiated
chondrocytes reexpress the differentiated collagen phenotype when
cultured in agarose gels. Cell. 30:215–224. 1982.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Darling EM and Athanasiou KA: Rapid
phenotypic changes in passaged articular chondrocyte
subpopulations. J Orthop Res. 23:425–432. 2005.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Johnstone B, Hering TM, Caplan AI,
Goldberg VM and Yoo JU: In vitro chondrogenesis of bone
marrow-derived mesenchymal progenitor cells. Exp Cell Res.
238:265–272. 1998.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Caron MMJ, Emans PJ, Coolsen MME, Voss L,
Surtel DA, Cremers A, van Rhijn LW and Welting TJ:
Redifferentiation of dedifferentiated human articular chondrocytes:
Comparison of 2D and 3D cultures. Osteoarthritis Cartilage.
20:117–126. 2012.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Mobaraki M, Ghaffari M, Yazdanpanah A, Luo
Y and Mills DK: Bioinks and bioprinting: A focused review.
Bioprinting. 18(e00080)2020.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Bajaj P, Schweller RM, Khademhosseini A,
West JL and Bashir R: 3D biofabrication strategies for tissue
engineering and regenerative medicine. Annu Rev Biomed Eng.
16:247–276. 2014.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Groll J, Boland T, Blunk T, Burdick JA,
Cho DW, Dalton PD, Derby B, Forgacs G, Li Q, Mironov VA, et al:
Biofabrication: Reappraising the definition of an evolving field.
Biofabrication. 8(013001)2016.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Almarza AJ and Athanasiou KA: Design
characteristics for the tissue engineering of cartilaginous
tissues. Ann Biomed Eng. 32:2–17. 2004.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Prakash D and Learmonth D: Natural
progression of Osteo-chondral defect in the femoral condyle. Knee.
9:7–10. 2002.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Li W, Miao W, Liu Y, Wang T, Zhang Y, Wang
W, Lu D, Zhou X, Jiao X, Jia X and Lin Y: Bioprinted constructs
that mimic the ossification center microenvironment for targeted
innervation in bone regeneration. Adv Funct Mater.
32(2109871)2022.
|
|
82
|
Zhao T, Liu Y, Wu Y, Zhao M and Zhao Y:
Controllable and biocompatible 3D bioprinting technology for
microorganisms: Fundamental, environmental applications and
challenges. Biotechnol Adv. 69(108243)2023.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Ma Y, Deng B, He R and Huang P:
Advancements of 3D bioprinting in regenerative medicine: Exploring
cell sources for organ fabrication. Heliyon.
10(e24593)2024.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Zhang H, Wang M, Wu R, Guo J, Sun A, Li Z,
Ye R, Xu G and Cheng Y: From materials to clinical use: Advances in
3D-printed scaffolds for cartilage tissue engineering. Phys Chem
Chem Phys. 25:24244–24263. 2023.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Zhang H, Zhou Z, Zhang F and Wan C:
Hydrogel-based 3D bioprinting technology for articular cartilage
regenerative engineering. Gels. 10(430)2024.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Irvine SA and Venkatraman SS: Bioprinting
and differentiation of stem cells. Molecules.
21(1188)2016.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Huang G, Zhao Y, Chen D, Wei L, Hu Z, Li
J, Zhou X, Yang B and Chen Z: Applications, advancements, and
challenges of 3D bioprinting in organ transplantation. Biomater
Sci. 12:1425–1448. 2024.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Gudapati H, Dey M and Ozbolat I: A
comprehensive review on droplet-based bioprinting: Past, present
and future. Biomaterials. 102:20–42. 2016.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Cui X, Boland T, D'Lima DD and Lotz MK:
Thermal inkjet printing in tissue engineering and regenerative
medicine. Recent Pat Drug Deliv Formul. 6:149–155. 2012.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Rutz AL, Lewis PL and Shah RN: Toward
next-generation bioinks: Tuning material properties pre- and
post-printing to optimize cell viability. MRS Bull. 42:563–570.
2017.
|
|
91
|
Hospodiuk M, Dey M, Sosnoski D and Ozbolat
IT: The bioink: A comprehensive review on bioprintable materials.
Biotechnol Adv. 35:217–239. 2017.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Guvendiren M, Molde J, Soares RMD and Kohn
J: Designing biomaterials for 3D printing. ACS Biomater Sci Eng.
2:1679–1693. 2016.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Roseti L, Parisi V, Petretta M, Cavallo C,
Desando G, Bartolotti I and Grigolo B: Scaffolds for bone tissue
engineering: State of the art and new perspectives. Mater Sci Eng C
Mater Biol Appl. 78:1246–1262. 2017.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Gruene M, Deiwick A, Koch L, Schlie S,
Unger C, Hofmann N, Bernemann I, Glasmacher B and Chichkov B: Laser
printing of stem cells for biofabrication of scaffold-free
autologous grafts. Tissue Eng Part C Methods. 17:79–87.
2010.PubMed/NCBI View Article : Google Scholar
|
|
95
|
You F, Wu X, Kelly M and Chen X:
Bioprinting and in vitro characterization of alginate
dialdehyde-gelatin hydrogel bio-ink. Bio-des Manuf. 3:48–59.
2020.
|
|
96
|
Skardal A and Atala A: Biomaterials for
integration with 3-D bioprinting. Ann Biomed Eng. 43:730–746.
2015.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Fedorovich NE, Schuurman W, Wijnberg HM,
Prins HJ, Van Weeren PR, Malda J, Alblas J and Dhert WJ:
Biofabrication of osteochondral tissue equivalents by printing
topologically defined, cell-laden hydrogel scaffolds. Tissue Eng
Part C Methods. 18:33–44. 2012.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Schütz K, Placht AM, Paul B, Brüggemeier
S, Gelinsky M and Lode A: Three-dimensional plotting of a
cell-laden alginate/methylcellulose blend: Towards biofabrication
of tissue engineering constructs with clinically relevant
dimensions. J Tissue Eng Regen Med. 11:1574–1587. 2017.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Wüst S, Müller R and Hofmann S: Controlled
positioning of cells in Biomaterials-approaches towards 3D tissue
printing. J Funct Biomater. 2:119–154. 2011.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Carrow JK, Kerativitayanan P, Jaiswal MK,
Lokhande G and Gaharwar AK: Polymers for bioprinting. In:
Essentials of 3D biofabrication and translation. Academic Press,
pp229-248, 2015.
|
|
101
|
Nguyen D, Hägg DA, Forsman A, Ekholm J,
Nimkingratana P, Brantsing C, Kalogeropoulos T, Zaunz S, Concaro S,
Brittberg M, et al: Cartilage tissue engineering by the 3D
Bioprinting of iPSCs in a nanocellulose/alginate bioink. Sci Rep.
7(658)2017.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Liang Q, Ma Y, Yao X and Wei W: Advanced
3D-printing bioinks for articular cartilage repair. Int J
Bioprinting. 8(511)2022.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Andrades JA, Motaung SC, Jiménez-Palomo P,
Claros S, López-Puerta JM, Becerra J, Schmid TM and Reddi AH:
Induction of superficial zone protein (SZP)/lubricin/PRG 4 in
muscle-derived mesenchymal stem/progenitor cells by transforming
growth factor-β1 and bone morphogenetic protein-7. Arthritis Res
Ther. 14(R72)2012.PubMed/NCBI View
Article : Google Scholar
|
|
104
|
Flannery CR, Hughes CE, Schumacher BL,
Tudor D, Aydelotte MB, Kuettner KE and Caterson B: Articular
cartilage superficial zone protein (SZP) is homologous to
megakaryocyte stimulating factor precursor and is a multifunctional
proteoglycan with potential growth-promoting, cytoprotective, and
lubricating properties in cartilage metabolism. Biochem Biophys Res
Commun. 254:535–541. 1999.PubMed/NCBI View Article : Google Scholar
|
|
105
|
Jay GD, Tantravahi U, Britt DE, Barrach HJ
and Cha CJ: Homology of lubricin and superficial zone protein
(SZP): Products of megakaryocyte stimulating factor (MSF) gene
expression by human synovial fibroblasts and articular chondrocytes
localized to chromosome 1 q25. J Orthop Res. 19:588–593.
2001.PubMed/NCBI View Article : Google Scholar
|
|
106
|
Radin EL, Swann DA and Weisser PA:
Separation of a hyaluronate-free lubricating fraction from synovial
fluid. Nature. 228:377–378. 1970.PubMed/NCBI View Article : Google Scholar
|
|
107
|
Swann DA, Slayter HS and Silver FH: The
molecular structure of lubricating glycoprotein-I, the boundary
lubricant for articular cartilage. J Biol Chem. 256:5921–5925.
1981.PubMed/NCBI
|
|
108
|
Chen SS, Falcovitz YH, Schneiderman R,
Maroudas A and Sah RL: Depth-dependent compressive properties of
normal aged human femoral head articular cartilage: Relationship to
fixed charge density. Osteoarthritis Cartilage. 9:561–569.
2001.PubMed/NCBI View Article : Google Scholar
|
|
109
|
Flannery CR, Zollner R, Corcoran C, Jones
AR, Root A, Rivera-Bermúdez MA, Blanchet T, Gleghorn JP, Bonassar
LJ, Bendele AM, et al: Prevention of cartilage degeneration in a
rat model of osteoarthritis by intra-articular treatment with
recombinant lubricin. Arthritis Rheum. 60:840–847. 2009.PubMed/NCBI View Article : Google Scholar
|
|
110
|
Gleghorn JP, Jones ARC, Flannery CR and
Bonassar LJ: Boundary mode lubrication of articular cartilage by
recombinant human lubricin. J Orthop Res. 27:771–777.
2009.PubMed/NCBI View Article : Google Scholar
|
|
111
|
Jay GD, Torres JR, Rhee DK, Helminen HJ,
Hytinnen MM, Cha CJ, Elsaid K, Kim KS, Cui Y and Warman ML:
Association between friction and wear in diarthrodial joints
lacking lubricin. Arthritis Rheum. 56:3662–3669. 2007.PubMed/NCBI View Article : Google Scholar
|
|
112
|
Rhee DK, Marcelino J, Baker M, Gong Y,
Smits P, Lefebvre V, Jay GD, Stewart M, Wang H, Warman ML and
Carpten JD: The secreted glycoprotein lubricin protects cartilage
surfaces and inhibits synovial cell overgrowth. J Clin Invest.
115:622–631. 2005.PubMed/NCBI View Article : Google Scholar
|
|
113
|
Waller KA, Zhang LX, Elsaid KA, Fleming
BC, Warman ML and Jay GD: Role of lubricin and boundary lubrication
in the prevention of chondrocyte apoptosis. Proc Natl Acad Sci USA.
110:5852–5857. 2013.PubMed/NCBI View Article : Google Scholar
|
|
114
|
Zappone B, Ruths M, Greene GW, Jay GD and
Israelachvili JN: Adsorption, lubrication, and wear of lubricin on
model surfaces: Polymer brush-like behavior of a glycoprotein.
Biophys J. 92:1693–1708. 2007.PubMed/NCBI View Article : Google Scholar
|
|
115
|
Bahabri SA, Suwairi WM, Laxer RM,
Polinkovsky A, Dalaan AA and Warman ML: The
camptodactyly-arthropathy-coxa vara-pericarditis syndrome: Clinical
features and genetic mapping to human chromosome 1. Arthritis
Rheum. 41:730–735. 1998.PubMed/NCBI View Article : Google Scholar
|
|
116
|
Klein TJ, Rizzi SC, Reichert JC, Georgi N,
Malda J, Schuurman W, Crawford RW and Hutmacher DW: Strategies for
zonal cartilage repair using hydrogels. Macromol Biosci.
9:1049–1058. 2009.PubMed/NCBI View Article : Google Scholar
|
|
117
|
Kamaraj A, Kyriacou H, Seah KM and Khan
WS: Use of human induced pluripotent stem cells for cartilage
regeneration in vitro and within chondral defect models of knee
joint cartilage in vivo: A Preferred Reporting Items for Systematic
Reviews and Meta-Analyses systematic literature review.
Cytotherapy. 23:647–661. 2021.PubMed/NCBI View Article : Google Scholar
|
|
118
|
Jeon O, Lee YB, Lee SJ, Guliyeva N, Lee J
and Alsberg E: Stem cell-laden hydrogel bioink for generation of
high-resolution and fidelity engineered tissues with complex
geometries. Bioact Mater. 15:185–193. 2021.PubMed/NCBI View Article : Google Scholar
|
|
119
|
Gatenholm B, Lindahl C, Brittberg M and
Simonsson S: Collagen 2A type B induction after 3D bioprinting
chondrocytes in situ into osteoarthritic chondral tibial lesion.
Cartilage. 13 (2_Suppl):1755S–1769S. 2020.PubMed/NCBI View Article : Google Scholar
|
|
120
|
Möller T, Amoroso M, Hägg D, Brantsing C,
Rotter N, Apelgren P, Lindahl A, Kölby L and Gatenholm P: In vivo
chondrogenesis in 3D bioprinted human cell-laden hydrogel
constructs. Plast Reconstr Surg Glob Open. 5(e1227)2017.PubMed/NCBI View Article : Google Scholar
|
|
121
|
Lee Y, Choi J and Hwang NS: Regulation of
lubricin for functional cartilage tissue regeneration: A review.
Biomater Res. 22(9)2018.PubMed/NCBI View Article : Google Scholar
|
|
122
|
Martyniak K, Kennedy S, Karimzadeh M, Cruz
MA, Jeon O, Alsberg E and Kean TJ: Optimizing bioink composition
for human chondrocyte expression of lubricin. Bioengineering.
10(997)2023.PubMed/NCBI View Article : Google Scholar
|
|
123
|
Cui Z, Xu C, Li X, Song J and Yu B:
Treatment with recombinant lubricin attenuates osteoarthritis by
positive feedback loop between articular cartilage and subchondral
bone in ovariectomized rats. Bone. 74:37–47. 2015.PubMed/NCBI View Article : Google Scholar
|
|
124
|
Pettersson I and Olsson Widjaja A: 3D
bioprinting meniscus for future tissue replacement in
osteoarthritis affected knee joints [master's thesis]. Chalmers
tekniska högskola, 2020.
|
|
125
|
Fu PJ, Zheng SY, Luo Y, Ren ZQ, Li ZH,
Wang YP and Lu BB: Prg4 and osteoarthritis: Functions, regulatory
factors, and treatment strategies. Biomedicines.
13(693)2025.PubMed/NCBI View Article : Google Scholar
|
|
126
|
Ignatyeva N, Gavrilov N, Timashev PS and
Medvedeva EV: Prg4-expressing chondroprogenitor cells in the
superficial zone of articular cartilage. Int J Mol Sci.
25(5605)2024.PubMed/NCBI View Article : Google Scholar
|
|
127
|
Xiong D, Gong C, Shi Z and Cui L: Boundary
film-liquid mixed lubrication of biomimetic lubricants on cartilage
interfaces. Friction. 13(9441029)2025.
|
|
128
|
Zhang K, Yang J, Sun Y, Wang Y, Liang J,
Luo J, Cui W, Deng L, Xu X, Wang B and Zhang H: Gelatin-based
composite hydrogels with biomimetic lubrication and sustained drug
release. Friction. 10:232–246. 2022.
|
|
129
|
Armiento AR, Stoddart MJ, Alini M and
Eglin D: Biomaterials for articular cartilage tissue engineering:
Learning from biology. Acta Biomater. 65:1–20. 2018.PubMed/NCBI View Article : Google Scholar
|
|
130
|
Filardo G, Perdisa F, Roffi A, Marcacci M
and Kon E: Stem cells in articular cartilage regeneration. J Orthop
Surg Res. 11(53)2016.PubMed/NCBI View Article : Google Scholar
|
|
131
|
Nakamura A, Murata D, Fujimoto R, Tamaki
S, Nagata S, Ikeya M, Toguchida J and Nakayama K: Bio-3D printing
iPSC-derived human chondrocytes for articular cartilage
regeneration. Biofabrication. 13(045009)2021.PubMed/NCBI View Article : Google Scholar
|
|
132
|
Ogorevc J, Orehek S and Dovč P: Cellular
reprogramming in farm animals: An overview of ipsc generation in
the mammalian farm animal species. J Anim Sci Biotechnol.
7(57)2016.PubMed/NCBI View Article : Google Scholar
|
|
133
|
Huang J, Xiong J, Wang D, Zhang J, Yang L,
Sun S and Liang Y: 3D bioprinting of hydrogels for cartilage tissue
engineering. Gels. 7(144)2021.PubMed/NCBI View Article : Google Scholar
|
|
134
|
Daly AC, Critchley SE, Rencsok EM and
Kelly DJ: A comparison of different bioinks for 3D bioprinting of
fibrocartilage and hyaline cartilage. Biofabrication.
8(045002)2016.PubMed/NCBI View Article : Google Scholar
|
|
135
|
Mouser VHM, Levato R, Bonassar LJ, D'Lima
DD, Grande DA, Klein TJ, Saris DBF, Zenobi-Wong M, Gawlitta D and
Malda J: Three-dimensional bioprinting and its potential in the
field of articular cartilage regeneration. Cartilage. 8:327–340.
2017.PubMed/NCBI View Article : Google Scholar
|
|
136
|
Markstedt K, Mantas A, Tournier I,
Martínez Ávila H, Hägg D and Gatenholm P: 3D bioprinting human
chondrocytes with nanocellulose-alginate bioink for cartilage
tissue engineering applications. Biomacromolecules. 16:1489–1496.
2015.PubMed/NCBI View Article : Google Scholar
|
|
137
|
Zhou J, Li Q, Tian Z, Yao Q and Zhang M:
Recent advances in 3D bioprinted cartilage-mimicking constructs for
applications in tissue engineering. Mater Today Bio.
23(100870)2023.PubMed/NCBI View Article : Google Scholar
|
|
138
|
Sang S, Mao X, Cao Y, Liu Z, Shen Z, Li M,
Jia W, Guo Z, Wang Z and Xiang C: 3D bioprinting using
synovium-derived MSC-laden photo-cross-linked ECM bioink for
cartilage regeneration. ACS Appl Mater Interfaces. 15:8895–8913.
2023.PubMed/NCBI View Article : Google Scholar
|
|
139
|
Zelinka A, Roelofs AJ, Kandel RA and De
Bari C: Cellular therapy and tissue engineering for cartilage
repair. Osteoarthritis Cartilage. 30:1547–1560. 2022.PubMed/NCBI View Article : Google Scholar
|
|
140
|
Lafuente-Merchan M, Ruiz-Alonso S,
García-Villén F, Gallego I, Gálvez-Martín P, Saenz-del-Burgo L and
Pedraz JL: Progress in 3D bioprinting technology for osteochondral
regeneration. Pharmaceutics. 14(1578)2022.PubMed/NCBI View Article : Google Scholar
|
|
141
|
Martínez-Moreno D, Venegas-Bustos D, Rus
G, Gálvez-Martín P, Jiménez G and Marchal JA: Chondro-inductive
B-TPUE-based functionalized scaffolds for application in cartilage
tissue engineering. Adv Healthc Mater. 11(e2200251)2022.PubMed/NCBI View Article : Google Scholar
|
|
142
|
Gao G and Cui X: Three-dimensional
bioprinting in tissue engineering and regenerative medicine.
Biotechnol Lett. 38:203–211. 2016.PubMed/NCBI View Article : Google Scholar
|
|
143
|
Yang X, Liu P, Zhang Y, Lu J and Zhao H:
Bioprinting-enabled biomaterials: A cutting-edge strategy for
future osteoarthritis therapy. Int J Nanomedicine. 18:6213–6232.
2023.PubMed/NCBI View Article : Google Scholar
|
|
144
|
Echave MC, Hernáez-Moya R, Iturriaga L,
Pedraz JL, Lakshminarayanan R, Dolatshahi-Pirouz A, Taebnia N and
Orive G: Recent advances in gelatin-based therapeutics. Expert Opin
Biol Ther. 19:773–779. 2019.PubMed/NCBI View Article : Google Scholar
|
|
145
|
Liu Y, Peng L, Li L, Huang C, Shi K, Meng
X, Wang P, Wu M, Li L, Cao H, et al: 3D-bioprinted BMSC-laden
biomimetic multiphasic scaffolds for efficient repair of
osteochondral defects in an osteoarthritic rat model. Biomaterials.
279(121216)2021.PubMed/NCBI View Article : Google Scholar
|
|
146
|
Pei Z, Gao M, Xing J, Wang C, Zhao P,
Zhang H and Qu J: Experimental study on repair of cartilage defects
in the rabbits with GelMA-MSCs scaffold prepared by
three-dimensional bioprinting. Int J Bioprint.
9(662)2023.PubMed/NCBI View Article : Google Scholar
|
|
147
|
Murphy SV, De Coppi P and Atala A:
Opportunities and challenges of translational 3D bioprinting. Nat
Biomed Eng. 4:370–380. 2020.PubMed/NCBI View Article : Google Scholar
|
|
148
|
Kolesky DB, Truby RL, Gladman AS, Busbee
TA, Homan KA and Lewis JA: 3D bioprinting of vascularized,
heterogeneous cell-laden tissue constructs. Adv Mater.
26:3124–3130. 2014.PubMed/NCBI View Article : Google Scholar
|
|
149
|
Ribeiro A, Blokzijl MM, Levato R, Visser
CW, Castilho M, Hennink WE, Vermonden T and Malda J: Assessing
bioink shape fidelity to aid material development in 3D
bioprinting. Biofabrication. 10(014102)2017.PubMed/NCBI View Article : Google Scholar
|
|
150
|
Bliley JM, Shiwarski DJ and Feinberg AW:
3D-bioprinted human tissue and the path toward clinical
translation. Sci Transl Med. 14(eabo7047)2022.PubMed/NCBI View Article : Google Scholar
|
|
151
|
Boretti G, Amirfallah A, Edmunds KJ,
Hamzehpour H and Sigurjónsson ÓE: Advancing cartilage tissue
engineering: A review of 3D bioprinting approaches and bioink
properties. Tissue Eng Part B Rev. 31:357–373. 2025.PubMed/NCBI View Article : Google Scholar
|
|
152
|
Murphy SV and Atala A: 3D bioprinting of
tissues and organs. Nat Biotechnol. 32:773–785. 2014.PubMed/NCBI View Article : Google Scholar
|