Introduction
Prostaglandins (PGs) play significant roles as
autacoids of bone metabolism (1,2). It
is widely acknowledged that the metabolism of bone tissue requires
the concerted actions of bone formation and bone resorption, which
are regulated by osteoblasts and osteoclasts, respectively
(3). In bone metabolism, PGF2α is
known to act as a potent bone modulator via specific receptors (FP)
on osteoblasts (1,2). It has been reported that PGF2α
stimulates the proliferation of osteoblasts and inhibits their
differentiation (2). In our
previous studies (4,5), we revealed that PGF2α induces the
activation of protein kinase C via phosphoinositide-hydrolyzing
phospholipase C and phosphatidylcholine-hydrolyzing phospholipase D
in osteoblast-like MC3T3-E1 cells. However, PGF2α reportedly
stimulates the synthesis of vascular endothelial growth factor
(VEGF) and interleukin-6 (IL-6) in osteoblasts (2,6) and
these bioactive substances, released from osteoblasts, are known to
function as potent modulators in bone metabolism. However, the
exact mechanism behind the PGF2α-stimulated synthesis in
osteoblasts has yet to be clarified.
VEGF is a strong mitogen which has a high
specificity for vascular endothelial cells (7) and is synthesized and secreted by a
range of cell types. The protein increases capillary permeability
and stimulates endothelial cells to proliferate (7). During bone remodeling, capillary
endothelial cells provide microvasculature. The activities of
osteoblasts, osteoclasts and capillary endothelial cells are known
to be closely associated and to regulate bone metabolism (8). It is thought that these cell types
interact via humoral factors and cell-to-cell contact. A previous
study used a co-culture model of osteoblasts and endothelial cells
to reveal that VEGF is crucial for the differentiation of
osteoblasts (9). It is widely
acknowledged that VEGF is a significant regulator of the growth and
repair of bone. In our previous studies (6,10),
we revealed that in osteoblast-like MC3T3-E1 cells,
PGF2α-stimulated VEGF synthesis is positively regulated by p44/p42
mitogen-activated protein (MAP) kinase, p38 MAP kinase and
stress-activated protein kinase/c-Jun N-terminal kinase
(SAPK/JNK), which are members of the MAP kinase superfamily
(11). By contrast, IL-6 is a
multifunctional cytokine that has crucial effects on a wide range
of functions, including the promotion of B-cell differentiation and
T-cell activation and induction of acute phase proteins (12). In bone metabolism, IL-6 is
generally known to stimulate bone resorption and promote osteoclast
formation (13). It has been
reported that potent bone resorptive agents, including tumor
necrosis factor-α and IL-1, stimulate IL-6 synthesis in osteoblasts
(13). Accumulating evidence
suggests that IL-6 secreted from osteoblasts is crucial as a
downstream effector of bone resorptive agents in bone metabolism.
We previously demonstrated that PGF2α stimulates IL-6 synthesis via
the activation of protein kinase C (14).
Wnts are secreted glycoproteins that regulate early
embryonic development and cell proliferation, differentiation and
survival (15,16). β-catenin is one of the key
downstream effectors in the Wnt pathway. In the canonical
Wnt/β-catenin signaling pathway (15), Wnts bind to Frizzled receptors and
the low-density lipoprotein receptor-related protein 5 or 6
(LRP5/6) in vertebrates (16,17).
This complex leads to the phosphorylation and inactivation of
glycogen synthase kinase 3β (GSK3β), which destabilizes β-catenin
and suppresses the accumulation of β-catenin in the nucleus
(15). In bone tissue, it has been
shown that loss or gain of function mutations in LRP-5 are
associated with osteoporosis-pseudoglioma syndrome or a high bone
density syndrome, respectively, in humans (17). Moreover, the genetic deletion of
β-catenin from early osteoprogenitors results in a lack of mature
osteoblasts in the mouse embryo (17), whereas the forced activation of
β-catenin greatly enhances osteogenesis (18). Accumulating evidence concerning the
roles of molecules downstream of LRP5, including GSK3β and
β-catenin, indicates that the Wnt signaling pathway is pivotal in
bone metabolism, especially bone formation (17,19,20).
The exact roles of the Wnt/β-catenin signaling pathway in
osteoblasts have yet to be elucidated, although osteoblasts have
been reported to produce Wnt proteins (17). Among the Wnt-ligands, it has been
shown that Wnt3a mainly activates the canonical pathway in
osteoblasts (21). In the present
study, we investigated whether Wnt3a affects the PGF2α-stimulated
synthesis of VEGF and IL-6 in osteoblast-like MC3T3-E1 cells. We
showed that Wnt3a enhances PGF2α-stimulated VEGF synthesis via the
canonical Wnt/β-catenin pathway in these cells while Wnt3a had
little effect on the IL-6 synthesis.
Materials and methods
Materials
PGF2α, Wnt3a and the mouse VEGF and IL-6
enzyme-linked immunosorbent assay (ELISA) kits were purchased from
R&D Systems, Inc. (Minneapolis, MN, USA). SB216763 was obtained
from Calbiochem-Novabiochem Co. (La Jolla, CA, USA).
Phospho-specific p44/p42 MAP kinase antibodies, p44/p42 MAP kinase
antibodies, phospho-specific p38 MAP kinase antibodies, p38 MAP
kinase antibodies, phospho-specific SAPK/JNK antibodies and
SAPK/JNK antibodies were purchased from Cell Signalling Technology,
Inc. (Beverly, MA, USA). The ECL western blotting detection system
was purchased from GE Healthcare UK Ltd. (Buckinghamshire, UK).
SB216763 was dissolved in dimethyl sulfoxide. Other materials and
chemicals were obtained from commercial sources. The maximum
concentration of dimethyl sulfoxide was 0.1%, which did not affect
the ELISA or western blot analysis.
Cell culture
The cloned osteoblast-like MC3T3-E1 cells, which are
derived from newborn mouse calvaria (22), were maintained as previously
described (23). Briefly, the
cells were cultured in α-minimum essential medium (α-MEM)
containing 10% fetal calf serum (FCS) at 37°C in a humidified
atmosphere of 5% CO2/95% air. The cells were seeded in
35-mm diameter dishes (5×104 cells/dish) or 90-mm
diameter dishes (20×104 cells/dish) in α-MEM containing
10% FCS. After 5 days, the medium was changed to α-MEM containing
0.3% FCS and the cells were incubated for 48 h. The cells were then
used for subsequent experiments.
Assay for VEGF and IL-6
The cultured cells were pretreated with various
doses of Wnt3a or SB216763 for 60 min and then stimulated with 10
mM PGF2α or vehicle in α-MEM containing 0.3% FCS for the indicated
periods. The conditioned medium was then collected and VEGF or IL-6
in the medium was measured using the mouse VEGF or IL-6 ELISA kit
according to the manufacturer’s instructions. The absorbance of the
ELISA samples was measured at 450 nm using EL 340 Bio Kinetic
Reader (Bio-Tek Instruments, Inc., Winooski, VT, USA).
Western blot analysis
Western blot analysis was performed as previously
described (24). In brief, the
cultured cells were pretreated with various doses of Wnt3a for 60
min and stimulated with 10 mM PGF2α or vehicle in α-MEM containing
0.3% FCS for the indicated periods. The cells were washed twice
with phosphate-buffered saline and then lysed, homogenized and
sonicated in a lysis buffer containing 62.5 mM Tris/HCl; pH 6.8, 3%
sodium dodecyl sulfate (SDS), 50 mM dithiothreitol and 10%
glycerol. SDS-polyacrylamide gel electrophoresis (PAGE) was
performed as described by Laemmli (25) using a 10% polyacrylamide gel. The
protein (10 mg) was fractionated and transferred onto an Immun-Blot
PVDF membrane (Bio-Rad, Hercules, CA, USA). The membranes were
blocked with 5% fat-free dry milk in Tris-buffered saline-Tween
(TBS-T; 20 mM Tris/HCl, pH 7.6, 137 mM NaCl, 0.1% Tween-20) for 2 h
prior to incubation with the primary antibodies. The rabbit
polyclonal phospho-specific p44/p42 MAP kinase antibodies, p44/p42
MAP kinase antibodies, phospho-specific SAPK/JNK antibodies,
SAPK/JNK antibodies, phospho-specific p38 MAP kinase antibodies or
p38 MAP kinase antibodies were used as primary antibodies.
Peroxidase-labeled antibodies raised in goat against rabbit IgG
were used as secondary antibodies. The primary and secondary
antibodies were diluted at 1:1,000 with 5% fat-free dry milk in
TBS-T. Peroxidase activity on the membrane was visualized on X-ray
film by means of the ECL western blotting detection system.
Statistical analysis
The data were analyzed by ANOVA followed by the
Bonferroni method for multiple comparisons between pairs and
P<0.05 was considered to indicate a statistically significant
result. Data were presented as the mean ± SEM of triplicate
determinations. Each experiment was repeated three times with
similar results.
Results
Effect of Wnt3a on the PGF2α-stimulated
VEGF release in osteoblast-like MC3T3-E1 cells
Since we previously showed that PGF2α significantly
stimulates VEGF synthesis in osteoblast-like MC3T3-E1 cells
(6), we first examined the effect
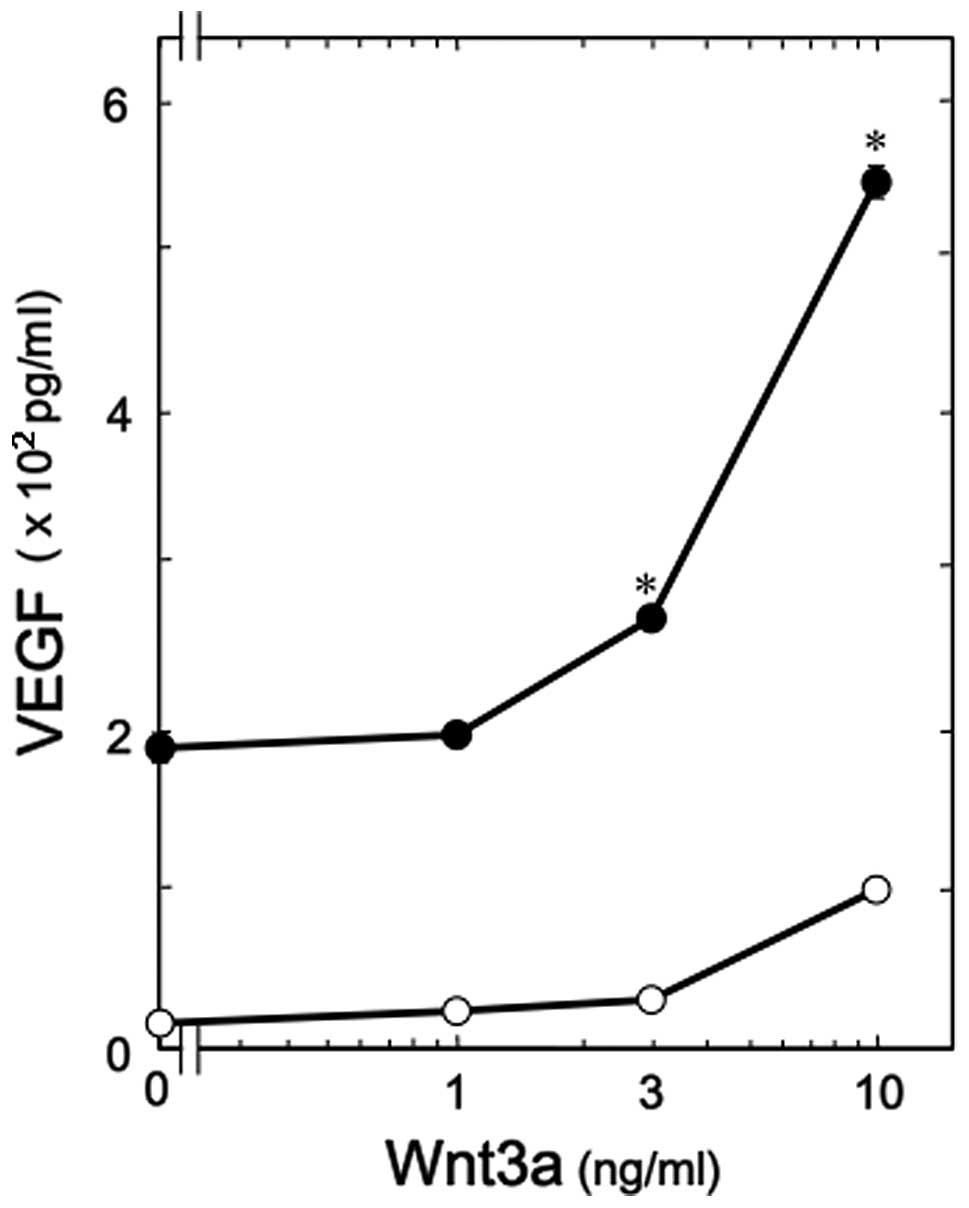
of Wnt3a on VEGF release induced by PGF2α in these cells. Wnt3a
significantly enhanced the PGF2α-induced VEGF release (Fig. 1). Amplification by Wnt3a was
dose-dependent at doses between 1 and 10 ng/ml. A dose of 10 ng/ml
Wnt3a resulted in a ~160% increase in the PGF2α effect.
Effect of Wnt3a on the PGF2α-stimulated
IL-6 release in osteoblast-like MC3T3-E1 cells
Previously, we demonstrated that PGF2α stimulates
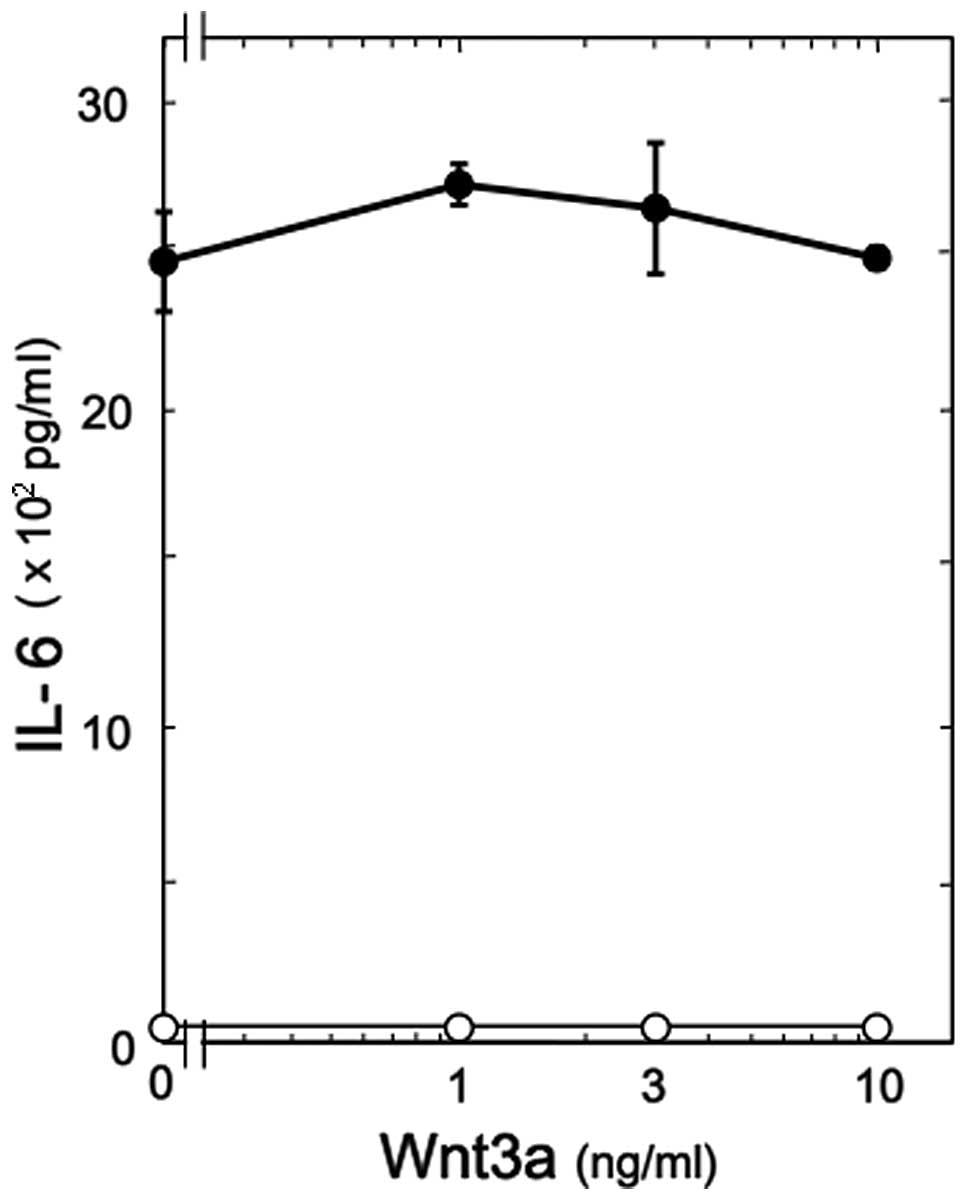
IL-6 synthesis in osteoblast-like MC3T3-E1 cells (14). In the present study, we examined
the effect of Wnt3a on IL-6 release induced by PGF2α in these
cells. However, Wnt3a, which alone had little effect on the IL-6
levels, did not affect the PGF2α-induced IL-6 release (Fig. 2).
Effects of SB216763 on the
PGF2α-stimulated release of VEGF or IL-6 in MC3T3-E1 cells
In the canonical Wnt/β-catenin pathway, Wnt
stimulation inhibits the GSK3β activity that otherwise
phosphorylates β-catenin and induces its degradation (15,16).
It has been reported that β-catenin accumulation is stimulated by
an inhibitor of GSK3β as well as Wnt in pre-osteoblast CIMC-4 cells
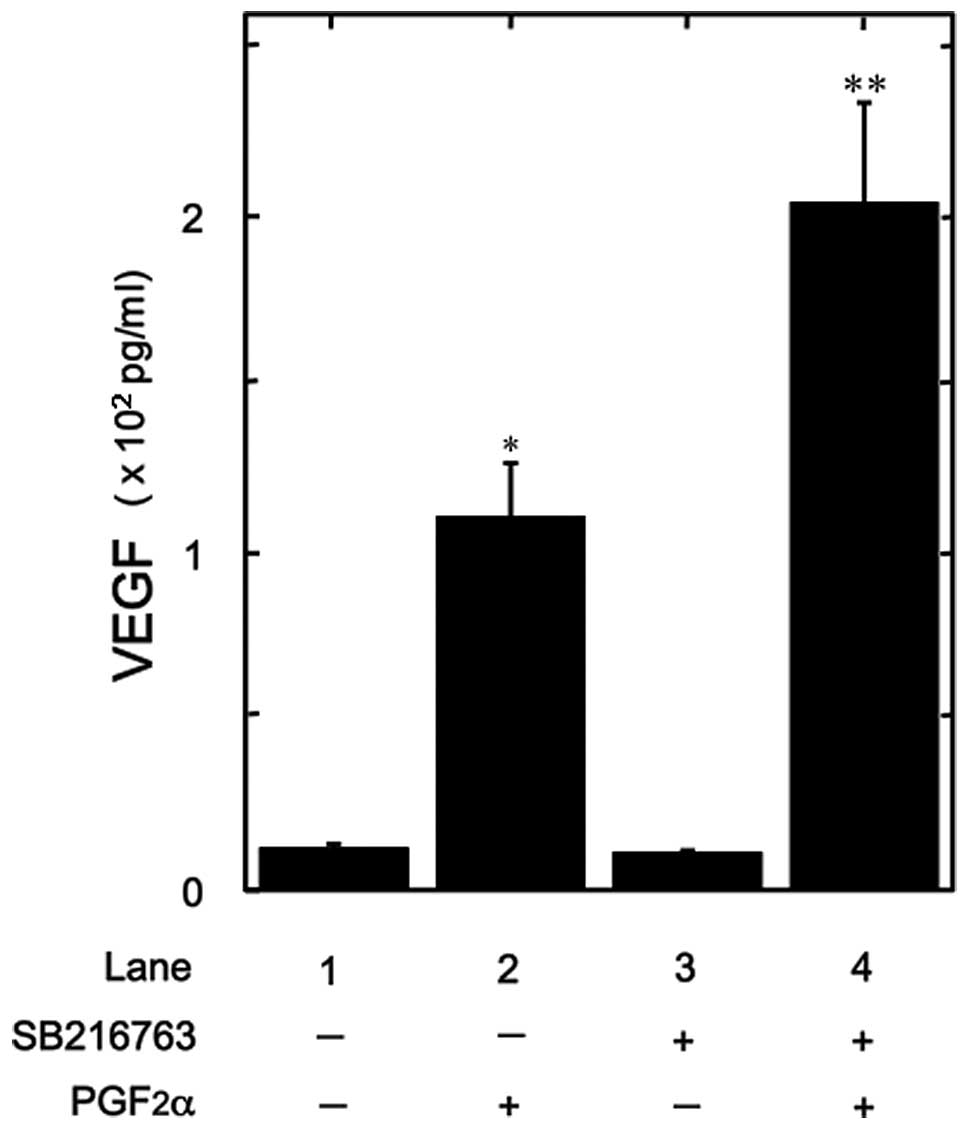
(26). Therefore, we examined the
effect of SB216763, an inhibitor of GSK3β (27), on the PGF2α-induced VEGF release.
SB216763 markedly potentiated the VEGF release (Fig. 3). A dose of 10 mM of SB216763
resulted in a ~100% increase in the PGF2α-effect.
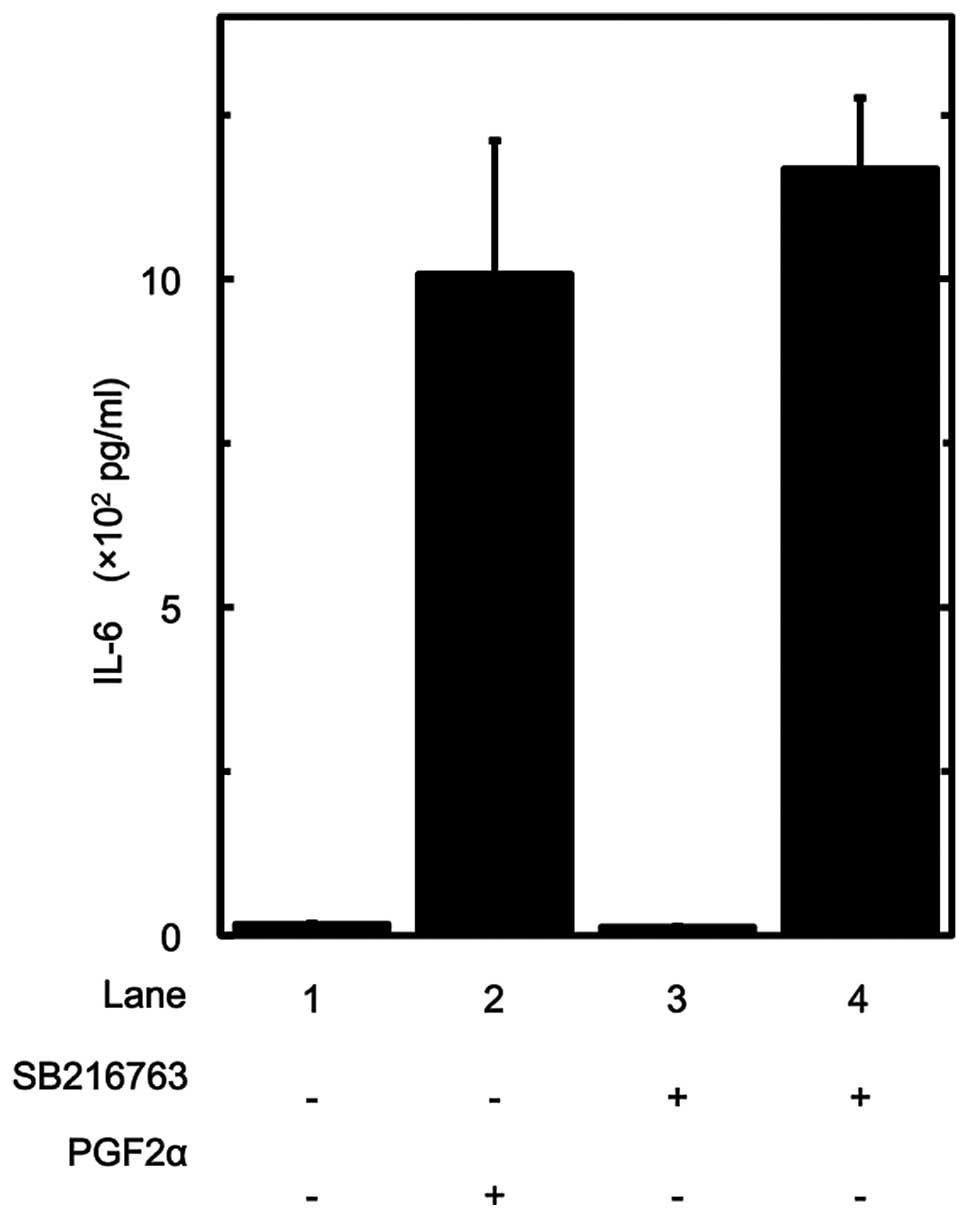
We also examined the effect of SB216763 on the
PGF2α-induced IL-6 release in MC3T3-E1 cells. However, SB216763 had
little effect on the PGF2α-induced IL-6 release (Fig. 4).
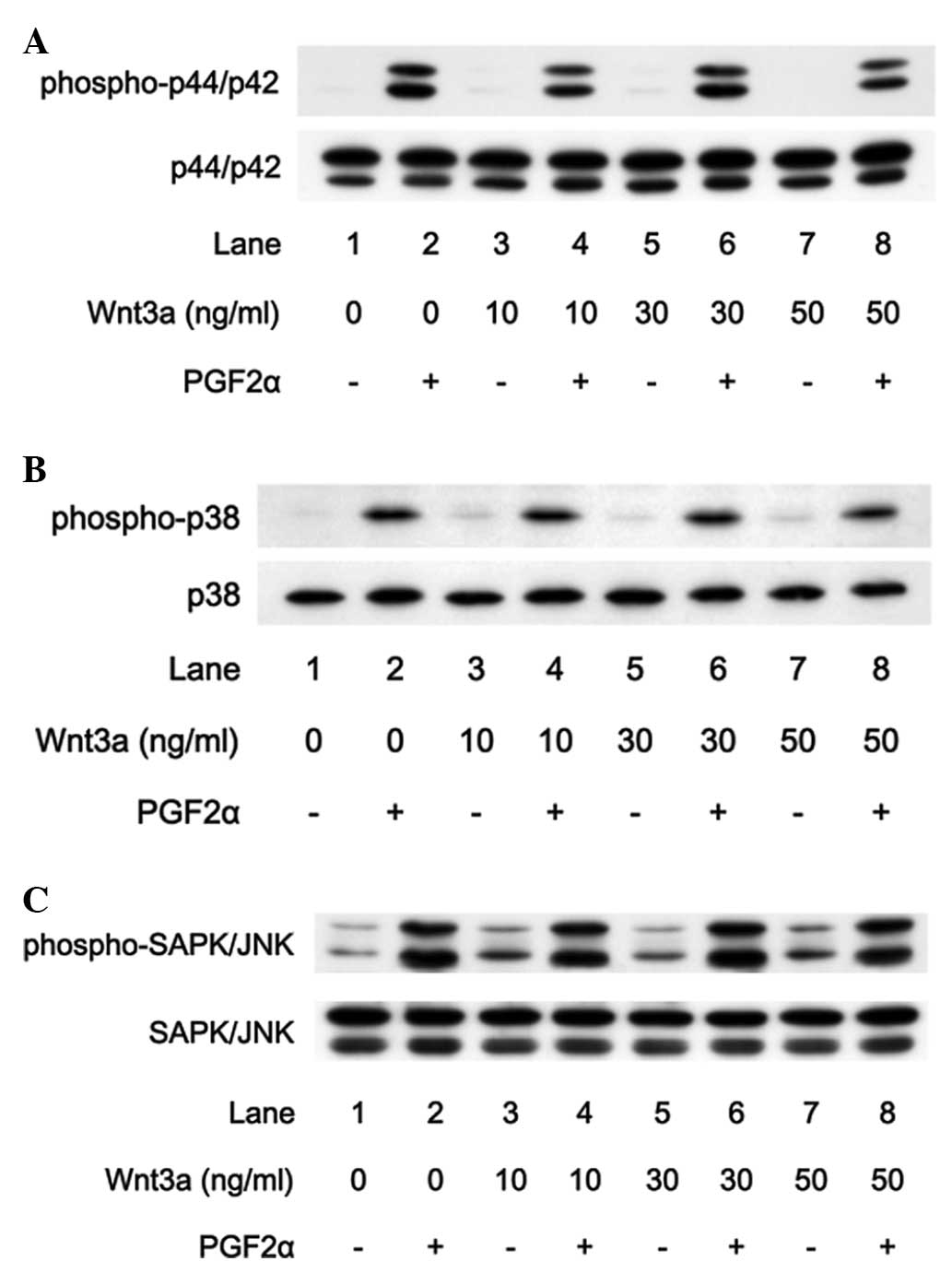
Effects of Wnt3a on the phosphorylation
of p44/p42 MAP kinase, SAPK/JNK and p38 MAP kinase induced by PGF2α
in MC3T3-E1 cells
In a previous study, we showed that p44/p42 MAP
kinase, p38 MAP kinase and SAPK/JNK are involved in the
PGF2α-stimulated VEGF synthesis in osteoblast-like MC3T3-E1 cells
(6,10). Therefore, to investigate whether
the amplifying effect of Wnt3a on the VEGF synthesis is dependent
on the MAP kinase pathway in MC3T3-E1 cells, we examined the
effects of Wnt3a on the PGF2α-induced phosphorylation of p44/p42
MAP kinase, p38 MAP kinase and SAPK/JNK. However, Wnt3a hardly
affected the PGF2α-stimulated phosphorylation of p44/p42 MAP kinase
at doses between 10 and 50 ng/ml (Fig.
5A). Furthermore, Wnt3a had little effect on the
phosphorylation of p38 MAP kinase (Fig. 5B) or SAPK/JNK (Fig. 5C) by PGF2α at doses between 10 and
50 ng/ml.
Discussion
In the present study, we demonstrated that Wnt3a
significantly enhanced VEGF synthesis stimulated by PGF2α in
osteoblast-like MC3T3-E1 cells. However, the PGF2α-stimulated IL-6
synthesis was not affected by Wnt3a in these cells. The activation
of the canonical pathway, which is well known as a major signaling
pathway involving Wnt, causes the accumulation of β-catenin in the
nucleus through the inactivation of GSK3β, which otherwise induces
the degradation of β-catenin (15,28).
It has been reported that Wnt3a increases β-catenin levels in
osteoblasts (28). We confirmed
that Wnt3a upregulated the protein levels of β-catenin in a
time-dependent manner in MC3T3-E1 cells. We also showed that
SB216763, a specific GSK3β inhibitor (27), markedly enhanced the VEGF synthesis
stimulated by PGF2α as well as Wnt3a in MC3T3-E1 cells. In
addition, SB216763 had little effect on the PGF2α-stimulated IL-6
synthesis, similar to Wnt3a. Therefore, our findings suggest that
Wnt3a upregulates PGF2α-stimulated VEGF synthesis via the canonical
Wnt/β-catenin pathway in osteoblast-like MC3T3-E1 cells.
It is generally known that members of the MAP kinase
superfamily, including p44/p42 MAP kinase, p38 MAP kinase and
SAPK/JNK, are central elements used by mammalian cells to transmit
various extracellular messages (11). With regard to VEGF synthesis in
osteoblasts, we have previously reported that the activation of
three major MAP kinases, p44/p42 MAP kinase, p38 MAP kinase and
SAPK/JNK, positively regulates PGF2α-stimulated VEGF synthesis in
osteoblast-like MC3T3-E1 cells (6,10).
Thus, we investigated the correlation between the Wnt pathway and
the MAP kinase pathway in the PGF2α-stimulated VEGF synthesis in
MC3T3-E1 cells. However, Wnt3a did not increase the PGF2α-induced
phosphorylation levels of p44/p42 MAP kinase. In addition, the
PGF2α-induced phosphorylation levels of p38 MAP kinase and SAPK/JNK
were not affected by Wnt3a. Based on these findings, it appears to
be unlikely that Wnt3a affects PGF2α-stimulated VEGF synthesis via
the modulation of p44/p42 MAP kinase, p38 MAP kinase or SAPK/JNK in
osteoblast-like MC3T3-E1 cells. Therefore, it is likely that Wnt3a
modulates PGF2α-induced VEGF synthesis at a point downstream of the
MAP kinases or that the effect of Wnt3a is exerted via the
modulation of other signaling pathways.
In bone tissue, the expansion of a capillary network
providing microvasculature is an essential process in bone
remodeling (9). Since VEGF is a
specific mitogen of vascular endothelial cells (7), VEGF released from osteoblasts acts as
a significant intercellular mediator between osteoblasts and
vascular endothelial cells. Moreover, VEGF is reportedly involved
in trabecular bone formation and the expansion of the hypertrophic
chondrocyte zone in the epiphyseal growth plate of mice (29), indicating the significance of VEGF
in bone metabolism. However, bone is capable of regenerating and
repairing itself after suffering from damage such as fractures. It
is known that the localized activation of signal cascades is
required for bone regeneration and fracture healing (30). Moreover, the Wnt/β-catenin
signaling pathway is known to play an anabolic role in bone
metabolism, leading to bone formation (17). The Wnt signaling pathway has been
reported to be activated and induce bone regeneration during bone
fracture repair, resulting in the increase of bone mass (30). In the present study, we
demonstrated that Wnt3a enhanced the PGF2α-stimulated VEGF
synthesis without affecting the synthesis of IL-6, a potent bone
resorptive agent (13), in
osteoblast-like MC3T3-E1 cells. Taking all our results into
account, it is probable that Wnt3a-enhanced VEGF synthesis in
osteoblasts is crucial in the physiological process of skeletal
maintenance, including bone remodeling and fracture healing, by
upregulating the proliferation of capillary endothelial cells.
These findings lead us to speculate that the effect of Wnt in bone
metabolism is mediated, at least in part, by VEGF release from
osteoblasts. Further investigation is necessary to clarify the
exact mechanism of the Wnt pathway in osteoblasts.
In conclusion, our results strongly suggest that
Wnt3a amplifies PGF2α-stimulated VEGF synthesis via the activation
of the canonical Wnt/β-catenin signaling pathway in
osteoblasts.
Acknowledgements
We are grateful to Yoko Kawamura and Emiko Fuseya
for their skillful technical assistance. This investigation was
supported in part by a Grant-in-Aid for Scientific Research
(19591042) from the Ministry of Education, Science, Sports and
Culture of Japan, the Foundation for Growth Science, the Research
Grants for Longevity Sciences (21A-1, 21A-4 and 21A-22) the
Ministry of Health, Labour and Welfare of Japan.
References
|
1
|
Pilbeam CC, Harrison JR and Raisz LG:
Prostaglandins and bone metabolism. Principles of Bone Biology.
Bilezikian JP, Raisz LG and Rodan GA: Academic press; San Diego:
pp. 715–728. 1996
|
|
2
|
Hikiji H, Takato T, Shimizu T and Ishii S:
The roles of prostanoids, leukotrienes, and platelet-activating
factor in bone metabolism and disease. Prog Lipid Res. 47:107–126.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Karsenty G and Wagner EF: Reaching a
genetic and molecular understanding of skeletal development. Dev
Cell. 2:389–406. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Miwa M, Tokuda H, Tsushita K, Kotoyori J,
Takahashi Y, Ozaki N, Kozawa O and Oiso Y: Involvement of
pertussis-toxin-sensitive GTP-binding protein in prostaglandin
F2α-induced phosphoinositide hydrolysis in osteoblast-like cells.
Biochem Biophys Res Commun. 171:1229–1235. 1990.
|
|
5
|
Kozawa O, Suzuki A, Kotoyori J, Tokuda H,
Watanabe Y, Ito Y and Oiso Y: Prostaglandin F2α-activates
phospholipase D independently from activation of protein kinase C
in osteoblast-like cells. J Cell Biochem. 55:373–379. 1994.
|
|
6
|
Tokuda H, Harada A, Hirade K, Matsuno H,
Ito H, Kato K, Oiso Y and Kozawa O: Incadronate amplifies
prostaglandin F2α-induced vascular endothelial growth factor in
osteoblasts: enhancement of MAP kinase activity. J Biol Chem.
278:18930–18937. 2003.
|
|
7
|
Ferrara N: Vascular endothelial growth
factor: basic science and clinical progress. Endoc Rev. 25:581–611.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Erlebacher A, Filvaroff EH, Gitelman SE
and Derynck R: Toward a molecular understanding of skeletal
development. Cell. 80:371–378. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Grellier M, Ferreira-Tojais N, Bourget C,
Bareille R, Guillemot F and Amédée J: Role of vascular endothelial
growth factor in the communication between human osteoprogenitors
and endothelial cells. J Cell Biochem. 106:390–398. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tokuda H, Takai S, Matsushima-Nishiwaki R,
Akamatsu S, Hanai Y, Hosoi T, Harada A, Ohta T and Kozawa O:
(-)-epigallocatechin gallate enhances prostaglandin F2α-induced
VEGF synthesis via up-regulating SAPK/JNK activation in
osteoblasts. J Cell Biochem. 100:1146–1153. 2007.PubMed/NCBI
|
|
11
|
Kyriakis JM and Avruch J: Mammalian
mitogen-activated protein kinase signal transduction pathways
activated by stress and inflammation. Physiol Rev. 81:807–869.
2001.PubMed/NCBI
|
|
12
|
Kishimoto T: Interleukin-6: from basic
science to medicine - 40 years in immunology. Annu Rev Immunol.
23:1–21. 2005.PubMed/NCBI
|
|
13
|
Kwan Tat S, Padrines M, Théoleyre S,
Heymann D and Fortun Y: IL-6, RANKL, TNF-alpha/IL-1: interrelations
in bone resorption pathophysiology. Cytokine Growth Factor Rev.
15:49–60. 2004.
|
|
14
|
Kozawa O, Suzuki A, Tokuda H and Uematsu
T: Prostaglandin F2α stimulates interleukin-6 via activation of PKC
in osteoblast-like cells. Am J Physiol. 272:E208–E211. 1997.
|
|
15
|
Moon RT, Bowerman B, Boutros M and
Perrimon N: The promise and perils of Wnt signalling through
β-catenin. Science. 296:1644–1646. 2002.PubMed/NCBI
|
|
16
|
van Amerongen R and Nusse R: Towards an
integrated view of Wnt signalling in development. Development.
136:3205–3214. 2009.PubMed/NCBI
|
|
17
|
Milat F and Ng KW: Is Wnt signalling the
final common pathway leading to bone formation? Mol Cell
Endocrinol. 310:52–62. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rodda SJ and McMahon AP: Distinct roles
for hedgehog and canonical Wnt signalling in specification,
differentiation and maintenance of osteoblast progenitors.
Development. 133:3231–3244. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hu H, Hilton MJ, Tu X, Yu K, Ornitz DM and
Long F: Sequential roles of Hedgehog and Wnt signalling in
osteoblast development. Development. 132:49–60. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Krishnan V, Bryant HU and Macdougald OA:
Regulation of bone mass by Wnt signalling. J Clin Invest.
116:1202–1209. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Almeida M, Han L, Bellido T, Manolagas SC
and Kousteni S: Wnt proteins prevent apoptosis of both uncommitted
osteoblast progenitors and differentiated osteoblasts by β-catenin-
dependent and -independent signalling cascades involving Src/ERK
and phosphatidylinositol 3-kinase/AKT. J Biol Chem.
280:41342–41351. 2005.PubMed/NCBI
|
|
22
|
Sudo H, Kodama H, Amagai Y, Yamamoto S and
Kasai S: In vitro differentiation and calcification in a new clonal
osteogenic cell line derived from newborn mouse calvaria. J Cell
Biol. 96:191–198. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kozawa O, Tokuda H, Miwa M, Kotoyori J and
Oiso Y: Cross-talk regulation between cyclic AMP production and
phosphoinositide hydrolysis induced by prostaglandin E2 in
osteoblast-like cells. Exp Cell Res. 198:130–134. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kato K, Ito H, Hasegawa K, Inaguma Y,
Kozawa O and Asano T: Modulation of the stress-induced synthesis of
hsp27 and αB-crystallin by cyclic AMP in C6 rat glioma cells. J
Neurochem. 66:946–950. 1996.
|
|
25
|
Laemmli UK: Cleavage of structural
proteins during the assembly of the head of bacteriophage T4.
Nature. 227:680–685. 1970. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Case N, Ma M, Sen B, Xie Z, Gross TS and
Rubin J: β-catenin levels influence rapid mechanical responses in
osteoblasts. J Biol Chem. 283:29196–29205. 2008.
|
|
27
|
Carmichael J, Sugars KL, Bao YP and
Rubinsztein DC: Glycogen synthase kinase-3β inhibitors prevent
cellular polyglutamine toxicity caused by the Huntington’s disease
mutation. J Biol Chem. 277:33791–33798. 2002.
|
|
28
|
Chung YS, Baylink DJ, Srivastava AK, Amaar
Y, Tapia B, Kasukawa Y and Mohan S: Effects of secreted
frizzled-related protein 3 on osteoblasts in vitro. J Bone Miner
Res. 19:1395–1402. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gerber HP, Vu TH, Ryan AM, Kowalski J,
Werb Z and Ferrara N: VEGF couples hypertrophic cartilage
remodeling, ossification and angiogenesis during endochondral bone
formation. Nature Med. 5:623–628. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Secreto FJ, Hoeppner LH and Westendorf JJ:
Wnt signalling during fracture repair. Curr Osteoporos Rep.
7:64–69. 2009. View Article : Google Scholar : PubMed/NCBI
|