Introduction
At the 2006 FIFA World Cup in Germany, millions of
people followed live the head-butt given by the French superstar
Zinédine Zidane to the Italian defender Marco Materazzi.
Whether this aggressive behavior is the result of
innate factors or acquired elements is a matter for debate. Animal
studies have indicated a link between incidents of aggression and
the individual level of circulating testosterone. However, results
in relation to primates, particularly humans, are less clear cut
and are at best only suggestive of a positive association in
certain contexts (1).
It has been observed in male athletes that
testosterone levels before a competition exhibit an increase that
precedes the start of the competition and reflects the quality of
the performance. In fact, testosterone levels measured in athletes
with high-level performances are increased compared with those
detected in low-level athletes (2).
It must be emphasized, nevertheless, that several
studies in humans did not observe any correlation between
testosterone and aggression (3–5). The
existence of such a correlation could explain the so-called ‘roid
rage’, which is the extremely aggressive behavior associated with
the intake of large amounts of anabolic steroids (6,7).
However, even if there is an effect of testosterone on aggression
at extremely high doses, this does not necessarily suggest that
testosterone is effective at physiological concentrations.
It has been suggested that prenatal androgens affect
the developing brain by increasing its sensitivity to circulating
testosterone later in life (8,9).
These effects may include increased self-confidence (10), search persistence (11) and risk preferences (12–14),
as well as intensified vigilance and quickened reaction times
(15). A number of markers have
been proposed for evaluating the effects of prenatal androgens
(16), but the most suitable is
likely to be the second-to-fourth digit length ratio (2D:4D), with
a relatively longer fourth finger (i.e., lower 2D:4D ratio)
indicating higher fetal androgen levels (17). However, a previous study (18) concluded that, due to the
considerable within-group variability and between-group overlap,
digit ratio is not a good marker of individual differences in
prenatal androgen exposure. Recently, Manning (19) suggested that 2D:4D is determined
not by prenatal androgens alone but by the balance of prenatal
androgen to prenatal estrogen signaling in a narrow time window of
fetal digit development. For ease of measurement and
reproducibility, 2D:4D is used as a substitute measure for prenatal
androgen exposure. Support for this use originates from the fact
that digit growth and gonadal development are linked by the common
influence of Hox genes (17,20).
These findings suggest that sex steroids produced by the developing
gonads exert significant modulatory effects on digit growth
(21). Lower digit ratios have
also been shown to correlate with increased sensitivity of androgen
receptors (22) and human male
reproductive function is negatively correlated with 2D:4D ratio
(23). Moreover, 2D:4D ratios
appear to be predictive of success among high-frequency financial
traders (24), predicting entrance
to and success in Medical Schools of state-run Italian Universities
(25) and performance of
competitive sports, such as basketball (26), skiing (27) and soccer (28).
Professional soccer players have lower 2D:4D ratios
than controls. Soccer players in the 1st team squads have lower
2D:4D ratios than reserves or youth team players. Men who had
represented their country had lower ratios than those who had not,
and there was a significant (one-tailed) negative association
between 2D:4D ratios and the number of international appearances
after the effect of country was removed. These data suggest that
prenatal and adult testosterone promotes the development and
maintenance of traits that are useful in sports and athletics
disciplines and in male:male fighting (28).
The present study was performed to test the
hypothesis that not only high levels of testosterone in adulthood
but also higher prenatal testosterone exposure may affect the
aggressive behavior of professional soccer players. Specifically,
we predicted that players with a lower 2D:4D ratio, due to their
aggressive behavior, commit a high number of fouls during the
match, punished by the referee by a caution (yellow card) or
sending-off (red card).
To test our predictions, we recruited 18 male
professional players from a professional soccer team of the ‘Series
A’ Italian Football League. We used the numbers of yellow as well
as red cards obtained by players as the primary measure of their
aggressive behavior. We also measured the levels of salivary
testosterone (Sal/T) in players and their aggression using the
Picture Frustration test (PFT) by Rosenzweig (29).
Materials and methods
Participants
We recruited 18 male professional players from a
first level Italian Soccer Team (Calcio Catania S.p.A.) which
participated in the ‘Serie A’ championship 2010/2011 of the Italian
Football League. The players received an introductory note that
explained briefly that we were looking at the effects of prenatal
testosterone on the shape of their right hand, but no information
was provided about our hypothesis. Prior to providing a handprint,
all the subjects completed a short questionnaire pertaining to
their age, medical history and, in particular, whether they had
broken the index or ring finger on their right hand. The
participants also signed an informed consent form. Participants had
a mean height of 1.78 m (±0.06), a mean body mass of 76.9 kg
(±5.56) and a mean body mass index (BMI) of 24.2 (±0.53).
The number of played games, the number of min played
in each game, and the number of yellow and red cards (fouls) were
obtained for each player. In this way, the number of fouls/game (1
game = 90 min) was calculated.
The study was approved by the Ethics Committee of
the Medical School at the University of Catania.
Digit ratio measurement
The method used for measuring digit ratio was
described by our group in a previous study (25). Briefly, to determine the 2D:4D
ratio, we photocopied the players’ right hands and measured the
digit length from the metacarpo-phalangeal crease to the finger
tip. It has been observed that this crease appears around the 9th
week of gestation and is one of the primary creases of the hand
(30).
The 2D:4D ratio was determined from only the right
hand, as the right-hand digit ratios exhibit more robust gender
differences and appear to be more sensitive to prenatal androgens
(31,32).
To measure the 2D:4D ratio we used practical
recommendations suggested by Voracek et al(33) and those recently described by
Coates and Hebert (34). In soft
tissue, care must be taken to distinguish regular from irregular or
secondary creases. Irregular creases form later than regular
creases, after the 11th week of gestation when the fingers start to
bend, disrupting the dermal surface (30,35).
The handprints of the players were measured to determine 2D:4D
ratio by one of the authors (M.C.) using calipers accurate to 0.2
mm.
Hormone assessment
The method used for testosterone assessment was
described by our group in a previous study (25). Briefly, saliva samples (1 ml) were
collected at rest in sterile containers and stored at −80°C.
Sugar-free gum (Vivident Xylit) was used to increase saliva flow in
the participants (36). Since in
adult males, the excretion of testosterone in saliva appeared to
follow a circadian rhythm (37)
and to be a pulsatile secretion (38), four samples were collected, at
intervals of 30 min, between 9:00 and 1:00 a.m. Saliva was assayed
using diagnostic kits (Diagnostic Systems Laboratories Inc.,
Webster, TX, USA) and modified radioimmunoassay methods (39). The testosterone assay sensitivity
was 0.3 pg/ml, with intra-and inter-assay CVs of <9.2 and
<8.3%, respectively. Only the highest value of the four measures
obtained from each subject was used for the experiments.
Picture frustration test
The method used for PFT assessment was described by
our group in a previous study (25). Briefly, the Italian version of the
PFT (29,40) permits an evaluation of the
subject’s preferred way of handling aggressive impulses. The
subject is asked to react verbally to 24 drawings showing common
frustration situations by filling out an empty speech bubble for a
character experiencing the frustration. The answers are assigned to
3 directions of aggression: ‘extraggression’, ‘intraggression’ and
‘imaggression’. The types of aggression include attending to the
frustrating barrier (obstacle dominance), defending the
organization of personality (ego-defense) or finding solutions
(need-persistence), and some special indices. Descriptions of each
factor are shown in Table I [based
on (41)].
 | Table IConstructs of reaction to
frustration.a |
Table I
Constructs of reaction to
frustration.a
| Type of
aggression |
|---|
|
|
|---|
| Direction of
aggression | Obstacle-dominance
(O-D) | Ego-defence
(etho-defense) (E-D) | Need-persistence
(N-P) |
|---|
| Extragression
(E-A) | E1
(Extrapeditive): The presence of the frustrating obstacle is
(insistently) pointed out. | E
(Extrapunitive):
Blame, hostility are turned against some person or thing in the
environment.
E (a variant of E):
subject denies that he is responsible for some offense with which
he is charged. | e
(Extrapersistive):
A solution to the frustrating situation is expected of someone
else. |
| Intraggression
(I-A) | I1
(Intropeditive):
Obstacle is construed as non-frustrating even beneficial; subject
can also emphasize his embarrassment for causing another person’s
frustration | I
(Intropunitive):
Blame, censure are directed by the subject - upon
himself.
I (a variant of I):
subject admits his guilt but denies any essential fault by
referring to unavoidable circumstance. | i
(Intropersistive):
Amends are offered by the subject, usually from a sense of guilt,
to solve the problem. |
| Imaggression
(M-A) | M1
(Impeditive):
Obstacle of frustration is minimized almost to the point of denying
its existence. | M
(Impunitive):
Blame for the frustration is evaded altogether, the situation is
regarded as unavoidable; the frustrating individual is
absolved. | m
(Impersistive):
Hope is expressed that time or normally expected circumstances will
bring about a solution of the problem; patience and conformity are
characteristic. |
Statistical analysis
Data were reported as the means ± SD. Data were
collected and averaged, and then compared using unpaired Student’s
t-test or one-way repeated measures ANOVA (Friedman test) followed
by Dunn’s multiple comparison test. Correlation analysis was
carried out by using one-tailed Pearson’s correlation analysis;
significance was set at P<0.05. All analyses were performed
using Systat Software Package version 11 (Systat Inc., Evanston,
IL, USA). Statistical analysis was carried out according to
guidelines for reporting statistics in journals published by the
American Physiological Society (42).
Results
In the present study, we first correlated the
different aspects of aggression assessed by PFT with digit ratio.
As shown in Table II, the only
significant detected correlation was with
extraggression-extrapersistive (P<0.05). The other
extraggression scores, extrapeditive and extrapunitive, on the PFT
test had no significant correlation with the 2D:4D ratio.
Additionally, the intraggression and imaggression scores on the PFT
test showed no significant correlation.
| Table IICorrelation between RPF scores and
2D:4D ratio. |
Table II
Correlation between RPF scores and
2D:4D ratio.
| RPF test | P-value |
|---|
| Intraggression | Intropeditive | NS |
| Intropunitive | NS |
|
Intropersistive | NS |
| Imaggression | Impeditive | NS |
| Impunitive | NS |
| Impersistive | NS |
| Extraggression | Extrapeditive | NS |
| Extrapunitive | NS |
|
Extrapersistive |
<0.05 |
Table III shows
data obtained for each soccer player, i.e., their playing position
(goalkeeper, defender, midfielder or attacker), their 2D:4D ratio
and Sal/T (pg/ml), their score in PFT test
(extraggression-extrapersistive) and the number of fouls obtained
per game.
| Table IIIData obtained for the 18 professional
soccer players. |
Table III
Data obtained for the 18 professional
soccer players.
| Players | Role | 2D:4D | Sal/T (pg/ml) | PFT (E-A) e | Fouls/game |
|---|
| 1 | G | 0.99 | 18 | 0.36 | 0.14 |
| 2 | D | 0.94 | 24 | 0.42 | 0.22 |
| 3 | D | 0.98 | 17 | 0.42 | 0.15 |
| 4 | M | 0.98 | 22 | 0.46 | 0.16 |
| 5 | M | 0.96 | 26 | 0.58 | 0.18 |
| 6 | D | 0.94 | 32 | 0.63 | 0.18 |
| 7 | D | 0.96 | 20 | 0.38 | 0.17 |
| 8 | D | 0.98 | 17 | 0.33 | 0.12 |
| 9 | M | 0.97 | 19 | 0.33 | 0.13 |
| 10 | G | 0.96 | 27 | 0.54 | 0.14 |
| 11 | A | 0.95 | 26 | 0.54 | 0.17 |
| 12 | G | 0.95 | 26 | 0.33 | 0.16 |
| 13 | A | 0.97 | 18 | 0.29 | 0.13 |
| 14 | D | 0.97 | 20 | 0.29 | 0.17 |
| 15 | D | 0.97 | 19 | 0.12 | 0.14 |
| 16 | D | 0.95 | 22 | 0.46 | 0.17 |
| 17 | M | 0.94 | 25 | 0.50 | 0.20 |
| 18 | A | 0.96 | 20 | 0.58 | 0.16 |
| Mean | | 0.96 | 22.11 | 0.42 | 0.16 |
| SD | | 0.02 | 4.19 | 0.13 | 0.03 |
Firstly, we analyzed the correlation between 2D:4D
ratio and the number of fouls obtained by the player per game
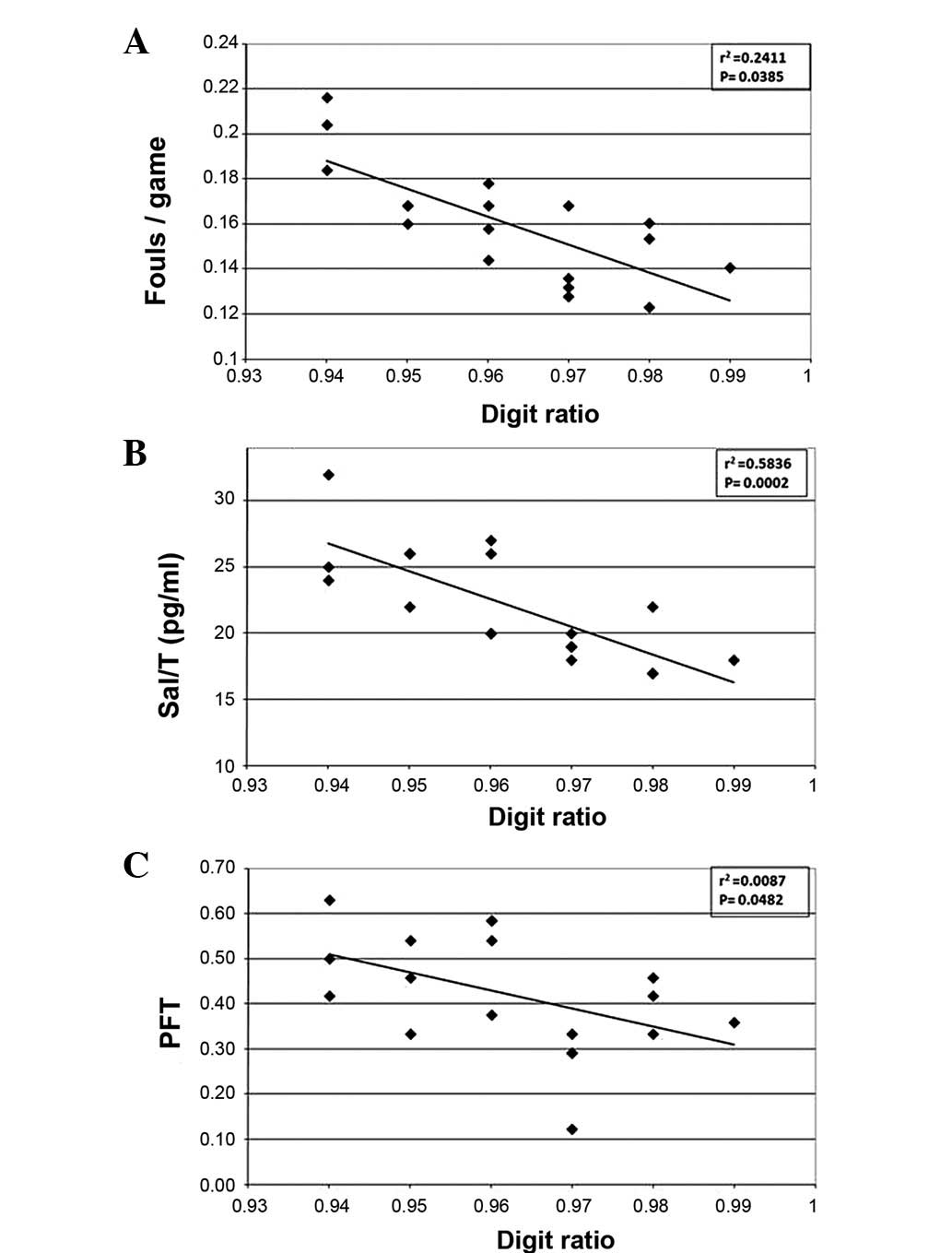
(Fig. 1A). As expected, players
with a lower 2D:4D ratio had a higher number of fouls per game. In
fact, the results showed that the lower a player’s 2D:4D ratio, the
higher his fouls/game ratio (P=0.0385). Moreover, we studied
possible correlations between the 2D:4D ratio of the players and
their salivary testosterone concentration (Sal/T). Mean Sal/T of
our players was 22.11 pg/ml (±4.19), indicating a significant
negative correlation between the two variables (P=0.0002) (Fig. 1B). When the 2D:4D ratio of our
players was plotted against their extrapersistive aggressiveness,
one of the extraggression characteristics from the PFT (Fig. 1C), we observed a significant
negative correlation between the two variables (P=0.0482). No
correlation was observed between 2D:4D ratio and the role of the
player in the team.
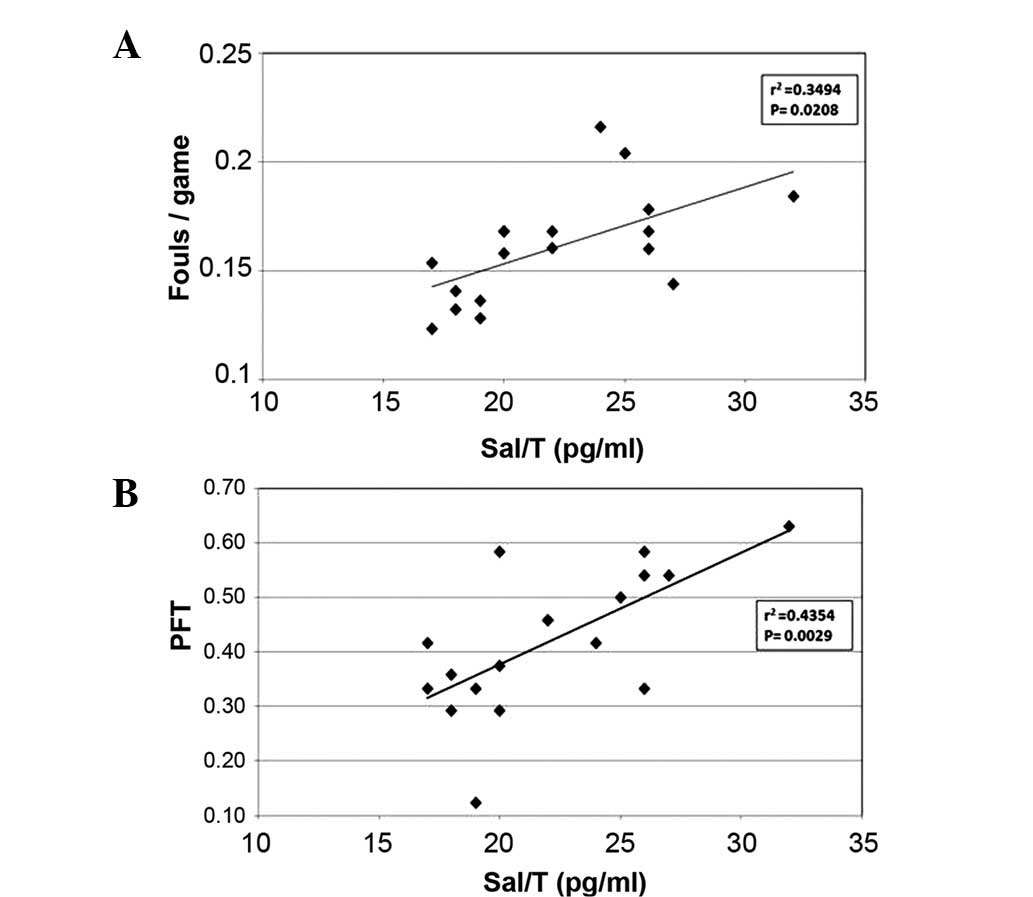
We also attempted to correlate the Sal/T values with
fouls/game score and the extrapersistive aggressiveness scores on
the PFT test. For the professional players included in this study,
a significant positive correlation was observed between Sal/T and
fouls/game score (P<0.01) and PFT (P<0.05) (Fig. 2). No significant correlation was
detected between Sal/T and the playing position in our sample of
soccer players.
Discussion
By summarizing our results, we found a significant
negative correlation between 2D:4D ratios in our sample of
professional soccer players and their salivary testosterone levels,
their extrapersistive aggressiveness (i.e., a mindset for which the
solution to the frustrating situation is expected of someone else)
and their number of fouls per game. No correlation was observed
between 2D:4D ratio and the playing positions. Moreover, in the
same players we detected a significant correlation between Sal/T
and both the number of fouls per game and aggressiveness, but not
with playing position.
Our results, beyond confirming the correlation
between testosterone levels and 2D:4D ratios, provide support for
the hypothesis that 2D:4D ratios predict a higher aggressive
behavior and, therefore, a higher risk of fouls during the
game.
Many investigators focus on the brain to explain
aggression. Electrical stimulation of the hypothalamus causes
aggressive behavior (43) and
receptors that modulate aggression levels have been identified in
the hypothalamus (44). These
brain areas have direct connections with both the brainstem nuclei
controlling vegetative functions, and with structures such as the
amygdala and prefrontal cortex.
Stimulation of the amygdala results in augmented
aggressive behavior in hamsters (45,46),
while in rhesus monkeys, neonatal lesions in the amygdala or
hippocampus result in reduced expression of social dominance,
related to the regulation of aggression and fear (47). In many mammals, the circuitry
within the amygdala appears to be involved in the control of
aggression. However, the role of the amygdala is less clear in
primates and appears to depend more on situational context, with
lesions leading to increases in either social affiliatory or
aggressive responses.
The prefrontal area of the cerebral cortex is
involved in aggression, along with many other functions, including
inhibition of emotions. Reduced activity of the prefrontal cortex,
in particular its medial and orbitofrontal portions, has been
associated with violent/antisocial aggression (48).
In a previous study, a significant correlation
between 2D:4D ratios and salivary testosterone in adult men was
noted (49). However, in the same
study, no significant correlation was observed between digit ratio
and reactive aggression. The discrepancy may be due to the
different methods used for measuring aggression: in the present
study we used a validated and widely used test (PFT), whereas
Benderlioglu and Nelson (49)
utilized an adaptation of the Kulik and Brown (50) method, which is less widespread and
not yet validated.
The correlation between baseline testosterone
concentrations and aggressiveness, detected in the present study,
has been previously observed (25,51–53),
although other investigators have failed to replicate this finding
(54,55). These contradictory data for
aggression may be due, in part, to the use of self-report measures
as opposed to the direct measurement of aggressive behavior
(56). Additionally, circadian
rhythm (37), pulsatile secretion
(38) or dynamic fluctuations
(57) in testosterone
concentrations may be more related to aggressive behavior than mean
daily testosterone concentrations. To minimize these possible
errors, for the evaluation of aggression, we directly measured
aggression of our players by using one of the most commonly used
tests (PFT) for this purpose, while testosterone collection was
carried out between 9:00 and 12:00 a.m., and 4 samples were
collected, at intervals of 30 min. Only the highest value of the
four measures obtained from each subject was used for the present
experiments.
In conclusion, this study appears to show that, in
professional soccer players, aggressive behavior with the
consequent increased risk of fouls during the game, is more likely
in individuals with high testosterone levels, not only in
adulthood, but also during their intrauterine life.
References
|
1
|
van Bokhoven I, van Goozen SH, van
Engeland H, Schaal B, Arseneault L, Séguin JR, Assaad JM, Nagin DS,
Vitaro F and Tremblay RE: Salivary testosterone and aggression,
delinquency, and social dominance in a population-based
longitudinal study of adolescent males. Horm Behav. 50:118–125.
2006.PubMed/NCBI
|
|
2
|
Mazur A and Booth A: Testosterone and
dominance in men. Behav Brain Sci. 21:353–363. 1998. View Article : Google Scholar
|
|
3
|
Albert DJ, Walsh ML and Jonik RH:
Aggression in humans: what is its biological foundation? Neurosci
Biobehav Rev. 17:405–425. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Coccaro EF, Beresford B, Minar P, Kaskow J
and Geracioti T: CSF testosterone: relationship to aggression,
impulsivity, and venturesomeness in adult males with personality
disorder. J Psychiatr Res. 41:488–492. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Constantino JN, Grosz D, Saenger P,
Chandler DW, Nandi R and Earls FJ: Testosterone and aggression in
children. J Am Acad Child Adolesc Psychiatry. 32:1217–1222. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pibiri F, Nelson M, Carboni G and Pinna G:
Neurosteroids regulate mouse aggression induced by anabolic
androgenic steroids. Neuroreport. 17:1537–1541. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Choi PY, Cowan D and Parrott AC: High-dose
anabolic steroids in strength athletes: effects upon hostility and
aggression. Hum Psychopharm Clin. 5:349–356. 2004. View Article : Google Scholar
|
|
8
|
Breedlove SM and Hampson E: Sexual
differentiation of the brain and behaviour. Behavioral
Endocrinology. Becker J, Breedlove SM, Crews D and McCarthy MM: 2nd
edition. MIT Press; Cambridge, MA: pp. 75–114. 2002
|
|
9
|
Tobet S and Baum M: Role for prenatal
estrogen in the development of masculine sexual behavior in the
male ferret. Horm Behav. 21:419–429. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Boissy A and Bouissou M: Effects of
androgen treatment on behavioural and physiological responses of
heifers to fear-eliciting situations. Horm Behav. 28:66–83. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Andrew RJ and Rogers LJ: Testosterone,
search behaviour and persistence. Nature. 237:343–346. 1972.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Apicella CL, Dreber A, Campbell B, Gray
PB, Hoffman M and Little AC: Testosterone and financial risk
preferences. Evol Hum Behav. 29:384–390. 2008. View Article : Google Scholar
|
|
13
|
Booth A, Johnson D and Granger D:
Testosterone and men’s health. J Behav Med. 22:1–19. 1999.
|
|
14
|
van Honk J, Schutter DJ, Hermans EJ,
Putman P, Tuiten A and Koppeschaar H: Testosterone shifts the
balance between sensitivity for punishment and reward in healthy
young women. Psychoneuroendocrinology. 29:937–943. 2004.PubMed/NCBI
|
|
15
|
Salminen E, Portin R, Koskinen A, Helenius
H and Nurmi M: Associations between serum testosterone fall and
cognitive function in prostate cancer patients. Clin Cancer Res.
10:7575–7582. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cohen-Bendahan CC, van de Beek C and
Berenbaum SA: Prenatal sex hormone effects on child and adult
sex-typed behavior: methods and findings. Neurosci Biobehav Rev.
29:353–384. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Manning JT, Scutt D, Wilson J and
Lewis-Jones DI: The 2nd to 4th digit length: a predictor of sperm
numbers and concentrations of testosterone, luteinizing hormone and
oestrogen. Hum Reprod. 13:3000–3004. 1998. View Article : Google Scholar
|
|
18
|
Berenbaum SA, Bryk KK, Nowak N, Quigley CA
and Moffat S: Fingers as a marker of prenatal androgen exposure.
Endocrinology. 150:5119–5124. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Manning JT: Resolving the role of prenatal
sex steroids in the development of digit ratio. Proc Natl Acad Sci
USA. 108:16143–16144. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Paul SN, Kato BS, Cherkas LF, Andrew T and
Spector TD: Heritability of the second to fourth digit ratio
(2d:4d): a twin study. Twin Res Hum Genet. 9:215–219. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
McIntyre M: The use of digit ratios as
markers for perinatal androgen action. Reprod Biol Endocrinol.
4:10–19. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Manning JT, Bundred PE, Newton DJ and
Flanagan BF: The 2nd to 4th digit ratio and variation in the
androgen receptor gene. Evol Hum Behav. 24:399–405. 2003.
View Article : Google Scholar
|
|
23
|
Auger J and Eustache F: Second to fourth
digit ratios, male genital development and reproductive health: a
clinical study among fertile men and testis cancer patients. Int J
Androl. 34:e49–e58. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Coates JM, Gurnell M and Rustichini A:
Second-to-fourth digit ratio predicts success among high-frequency
financial traders. Proc Natl Acad Sci USA. 106:623–628. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Coco M and Perciavalle V, Maci T,
Nicoletti F, Di Corrado D and Perciavalle V: The second-to-fourth
digit ratio correlates with the rate of academic performance in
medical school students. Mol Med Rep. 4:471–476. 2011.PubMed/NCBI
|
|
26
|
Tester N and Campbell A: Sporting
achievement: what is the contribution of digit ratio? J Pers.
75:663–677. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Manning JT: The ratio of 2nd to 4th digit
length and performance in skiing. J Sports Med Phys Fitness.
42:446–450. 2002.PubMed/NCBI
|
|
28
|
Manning JT and Taylor RP: Second to fourth
digit ratio and male ability in sport: implications for sexual
selection in humans. Evol Hum Behav. 22:61–69. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rosenzweig S: PFS picture-frustration
study: manuale integrato delle tre forme per adulti, bambini e
adolescenti. Organizzazioni Speciali; Firenze: pp. 2111992
|
|
30
|
Kimura S, Schaumann BA, Plato CC and
Kitagawa T: Embryological development and prevalence of digital
flexion creases. Anat Rec. 226:249–257. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Manning JT, Churchill AJ and Peters M: The
effects of sex, ethnicity, and sexual orientation on self-measured
digit ratio (2D:4D). Arch Sex Behav. 36:223–233. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Williams TJ, Pepitone ME, Christensen SE,
Cooke BM, Huberman AD, Breedlove NJ, Breedlove TJ, Jordan CL and
Breedlove SM: Finger-length ratios and sexual orientation. Nature.
404:455–456. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Voracek M, Bagdonas A and Dressler SG:
Digit ratio (2D:4D) in Lithuania once and now: testing for sex
differences, relations with eye and hair color, and a possible
secular change. Coll Antropol. 31:863–868. 2007.PubMed/NCBI
|
|
34
|
Coates JM and Hebert J: Endogenous
steroids and financial risk taking on a London trading floor. Proc
Natl Acad Sci USA. 105:6167–6172. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ashbaugh D: The identification process.
Quantitative-Qualitative Friction Ridge Analysis: An Introduction
to Basic and Advanced Ridgeology. CRC Press; Boca Raton, FL: pp.
2341999
|
|
36
|
Dabbs JM Jr: Salivary testosterone
measurements: collecting, storing, and mailing saliva samples.
Physiol Behav. 49:815–817. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Landman AD, Sanford LM, Howland BE, Dawes
C and Pritchard ET: Testosterone in human saliva. Experientia.
32:940–941. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Keenan DM and Veldhuis JD: Disruption of
the hypothalamic luteinizing hormone pulsing mechanism in aging
men. Am J Physiol Regul Integr Comp Physiol. 281:R1917–R1924.
2001.PubMed/NCBI
|
|
39
|
Granger DA, Schwartz EB, Booth A and
Arentz M: Salivary testosterone determination in studies of child
health and development. Horm Behav. 35:18–27. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Rauchfleisch U: Handbuch zum Rosenzweig
Picture-Frustration Test (PFT). Huber; Bern: pp. 3991979
|
|
41
|
Tatsuki DH: If my complaints could
passions move: an interlanguage study of aggression. J Pragm.
32:1003–1017. 2000. View Article : Google Scholar
|
|
42
|
Curran-Everett D and Benos DJ: Guidelines
for reporting statistics in journals published by the American
Physiological Society. Am J Physiol Regul Integr Comp Physiol.
97:457–459. 2004.
|
|
43
|
Kruk MR, Van der Poel AM, Meelis W,
Hermans J, Mostert PG, Mos J and Lohman AH: Discriminant analysis
of the localization of aggression-inducing electrode placements in
the hypothalamus of male rats. Brain Res. 260:61–79. 1983.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ferris CF, Melloni RH Jr, Koppel G, Perry
KW, Fuller RW and Delville Y: Vasopressin/serotonin interactions in
the anterior hypothalamus control aggressive behavior in golden
hamsters. J Neurosci. 17:4331–4340. 1997.
|
|
45
|
Potegal M, Hebert M, DeCoster M and
Meyerhoff JL: Brief, high-frequency stimulation of the
corticomedial amygdala induces a delayed and prolonged increase of
aggressiveness in male Syrian golden hamsters. Behav Neurosci.
110:401–412. 1996. View Article : Google Scholar
|
|
46
|
Potegal M, Ferris CF, Hebert M, Meyerhoff
J and Skaredoff L: Attack priming in female Syrian golden hamsters
is associated with a c-fos-coupled process within the corticomedial
amygdala. Neuroscience. 75:869–880. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bauman MD, Toscano JE, Mason WA, Lavenex P
and Amaral DG: The expression of social dominance following
neonatal lesions of the amygdala or hippocampus in rhesus monkeys
(Macaca mulatta). Behav Neurosci. 120:749–760. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Paus T: Mapping brain development and
aggression. Can Child Adolesc Psychiatr Rev. 14:10–15.
2005.PubMed/NCBI
|
|
49
|
Benderlioglu Z and Nelson RJ: Digit length
ratios predict reactive aggression in women, but not in men. Horm
Behav. 46:558–564. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kulik JA and Brown R: Frustration,
attribution of blame, and aggression. J Exp Soc Psychol.
15:183–194. 1979. View Article : Google Scholar
|
|
51
|
Dabbs JM, Carr TS, Frady RL and Riad JK:
Testosterone, crime, and misbehavior among 692 male prison inmates.
Pers Indiv Differ. 18:627–633. 1995. View Article : Google Scholar
|
|
52
|
Archer J: Testosterone and human
aggression: an evaluation of the challenge hypothesis. Neurosci
Biobehav Rev. 30:319–345. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Sellers JG, Mehl MR and Josephs RA:
Hormones and personality: testosterone as a marker of individual
differences. J Res Pers. 41:126–138. 2007. View Article : Google Scholar
|
|
54
|
Josephs RA, Sellers JG, Newman ML and
Mehta PH: The mismatch effect: when testosterone and status are at
odds. J Pers Soc Psychol. 90:999–1013. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Stanton SJ and Schultheiss OC: Basal and
dynamic relationship between implicit power motivation and
estradiol in women. Horm Behav. 52:571–580. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Klinesmith J, Kasser T and McAndrew FT:
Guns, testosterone and aggression: an experimental test of a
mediational hypothesis. Psychol Sci. 17:568–571. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Hermans EJ, Ramsey NF and van Honk J:
Exogenous testosterone enhances responsiveness to social threat in
the neural circuitry of social aggression in humans. Biol
Psychiatry. 63:263–270. 2008. View Article : Google Scholar : PubMed/NCBI
|