Introduction
Human papillomaviruses (HPVs) pertain to the
Papillomaviridae family, a highly diverse group of viruses
including >100 different types, and infect the epithelial cells
of several vertebrate species (1).
Condyloma acuminatum (CA) and verruca vulgaris (VV) are possibly
the most common epidermal infection caused by HPVs. CA, defined as
genital warts, in ~90% of cases is caused by the low risk HPV types
6 or 11, while the high risk HPV 16 or 18 mainly account for the
rest. CA is regarded as a benign tumor and may be contagious during
sexual activity. VV, which is caused mainly by HPV type 2, causes
recognizable warts that are more frequently observed in children,
occurring on the fingers and hands. VV is also benign and does not
involve internal organs. The genesis and development mechanism of
the warts caused by HPV infection is complex, and avoiding the host
immune response to HPVs has been considered a key factor in
triggering their development (2).
Furthermore, local inflammatory responses may promote neoplasia
progression through a number of different mechanisms (3,4).
HMGB1 is a fundamental nuclear DNA binding protein
(5) that is secreted into the
extracellular fluid to act as a proinflammatory cytokine. It binds
to TLR4 to activate nuclear factor-κB (NF-κB) (6,7),
which is involved in tumor progression (8). It has been recognized that HMGB1 has
important roles in inflammation and cancer (9,10).
Inflammation has been found to be intensified along with the
increase in epidermal tumor malignancy, and the TLR4 signaling
pathway has been considered to be a significant mediator of
inflammation in a number of malignant tumors (11).
Until now, the role of HMGB1, TLR4 and p65 in warts
caused by HPVs has not been elucidated. In the present study, their
involvement in warts caused by HPV was investigated, primarily by
examining the relation between HMGB1-TLR4 pathway-associated
inflammation and the development of CA and VV.
Materials and methods
Samples
The specimens were obtained from 30 cases of CA and
19 cases of VV, which were diagnosed based on clinically apparent
warts and histopathological features. The surrounding tissue of the
warts in 20 participants was used as normal contrasts. The
participants with immunological deficiency diseases were excluded.
A therapeutic washout period of three weeks for the warts was
implemented prior to specimen collection.
The study was performed in accordance with the
Declaration of Helsinki of 1964 and the subsequent amendments. The
Ethics Board of Tongji Medical College (Wuhan, China) and the
Ethics Board of Sun Yat-Sen Medical College (Zhuhai, China)
approved the experimental procedures. All of the subjects provided
written informed consent prior to participating in the study.
Antibodies and reagents
The antibodies and reagents used in the study
included: Anti-HMGB1 (2600-1; Epitomics, Burlingame, CA, USA),
anti-TLR4 (ab22048; Abcam, Cambridge, UK), anti-NF-κB p65 (SC-7151;
Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) and REALTMEn
Vision Detection kit (Dako, Carpinteria, CA, USA).
Immunohistochemistry
Immunohistochemistry EnVision was used to detect the
expression of HMGB1, TLR4 and NF-κB p65 in wart and normal skin
(NS) specimens. The 69 tissue specimens were routinely fixed in
formalin and embedded in paraffin. Sections of 4 μm thickness were
cut from paraffin-embedded tissue blocks and mounted on silanized
slides. Following dewaxing and rehydration, the sections were
antigen retrieved with ethylenediamine tetraacetic acid or citric
acid, incubated with 3% H2O2 for 10 min and
blocked with 5% bovine serum albumin for 20 min. The specimens were
then incubated with the primary antibodies (anti-HMGB1 1:800,
anti-TLR4 1:200 and anti-NF-κB p65 1:200) for 24 h at 4°C. Then,
the secondary antibodies were added (ChemMate TMEnVision +/
HRP-goat anti-mouse/rabbit secondary antibody, K5007; Dako,
Glostrup, Denmark) and the specimens were incubated for 45 min,
followed by the addition of 50–100 μl diaminobenzidine.
Dehydration, transparence and mounting of specimens for examination
were conducted using routine procedures. The specimens were
photographed with a Nikon Eclipse Ti-SR microscope equipped with a
Nikon DS-U3 digital camera (Nikon Incorporation, Tokyo, Japan). The
negative controls were obtained by omitting the primary
antibodies.
Two blinded pathologists conducted semi-quantitative
immunohistochemical grading of the nuclei, cytoplasm, cell
membranes, whole cells or intercellular spaces. The average scores
were used for analysis. The scoring system was as follows: 0, no
staining; 1, light brown yellow staining; 2, brown staining; and 3,
dark brown staining. Ten fields of view were counted on each slide
at a magnification of ×400. The average positive expression on each
slide was scored as: 1, <25%; 2, 25–49%; 3, 50–74%; and 4, ≥75%.
The product of the percentage of positive expression and the degree
of staining scores for each slide provided a final score, which was
used to indicate the following: 0–1 point, negative (−); 2–3
points, weakly positive (+); 4–6, moderately positive (++); and
>6, strongly positive (+++).
Western blot analysis
Western immunoblot analysis was used to detect the
expression of NF-κB p65 in nuclei isolated from HPV-infected and
normal tissues. Dermal and subcutaneous tissues were removed from
the specimens and the epidermis was sliced into small sections to
prepare the epithelial nuclear proteins using a cytoplasmic/nuclear
protein extraction kit (Beyotime Co., Shanghai, China). Equal
quantities of cytoplasmic and nuclear extracts were subjected to
10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and
transferred to nitrocellulose membranes. The membranes were blocked
overnight at 4°C in phosphate-buffered saline and 0.1% Tween-20
buffer containing 5% non-fat dried milk proteins. The membranes
were then blotted for 2 h at room temperature with the primary
anti-NF-κB p65 antibody diluted at 1:500. The membrane-bound
protein-antibody complexes were labeled with horseradish
peroxidase-conjugated anti-IgG diluted at 1:3,000. Histone H3 was
used as an equal protein loading control. An enhanced
chemiluminescence system (Pierce Biotechnology, Inc., Rockford, IL,
USA) was used for detection.
Statistical analysis
Data are expressed as the mean ± standard error of
the mean (SEM). The between-group differences were analyzed by
one-way analysis of variance (ANOVA) followed by the Bonferroni
method for normally distributed datasets. The Kruskal-Wallis test
followed by the Nemenyi multiple comparisons test was used for
skewed datasets. Spearman’s correlation test was used for the
correlation analysis. P<0.05 and P<0.01 were considered to
indicate a statistically significant difference. Statistical
analysis was performed using R software, v3.0 (GNU Project, Boston,
MA, USA).
Results
Expression of HMGB1 in warts caused by
HPV
In VV, there was diffuse weakly to moderately
positive expression of HMGB1 in the epithelial nuclei and weak
expression in the cytoplasm. A weak expression level of
extracellular HMGB1 was also found in the intercellular spaces of
the epithelium. As for the associated inflammatory and vascular
endothelial cells, moderately positive HMGB1 expression was
observed in the nuclei and cytoplasm from VV samples (Fig. 1A).
 | Figure 1HMGB1 expression in NS and wart
samples determined by immunohistochemistry EnVision (magnification,
×400). (A to C) HMGB1 positive expression is located in the nuclei,
cytoplasm, cells and/or intercellular spaces following stimulation
of cell necrosis or inflammation. The red arrows represent HMGB1
expression in the epithelial intercellular spaces, the black arrows
represent HMGB1 expression in the epithelial cell nuclei, the blue
arrows represent HMGB1 expression in inflammatory cells, and the
green arrows represent HMGB1 expression in the vascular endothelial
cells. (D) HMGB1 expression in the epithelial intercellular spaces.
(E) HMGB1 expression in the epithelial cell nuclei. (F) HMGB1
expression in the inflammatory cells. The error bars represent the
standard error of the mean. *P<0.05 and
**P<0.01. HMGB1, high mobility group protein box 1;
CA, condyloma acuminatum; VV, verruca vulgaris; NS, normal
skin. |
In CA, there was diffuse moderately positive
expression of HMGB1 in the epithelial nuclei but minimal expression
of HMGB1 in the cytoplasm and weakly positive expression of
extracellular HMGB1 in the epithelial intercellular spaces. There
was moderately positive HMGB1 expression in the nuclei and
cytoplasm of the associated inflammatory cells and vascular
endothelial cells (Fig. 1B).
In the NS, HMGB1 exhibited moderately to strongly
positive diffuse expression in the squamous epithelial nuclei and
occasionally focal expression in the squamous epithelial cytoplasm.
Weak extracellular HMGB1 expression in the intercellular spaces of
the normal squamous epithelium was observed. There were a few
inflammatory cells in the NS, which exhibited a weak HMGB1
expression in the nuclei or cytoplasm, and there was moderately to
strongly positive HMGB1 expression in the nuclei and cytoplasm of
vascular endothelial cells (Fig.
1C).
ANOVA demonstrated that the expression of HMGB1 in
epithelial intercellular spaces of VV samples was significantly
higher than in the NS (P=0.0308), and that the expression of HMGB1
in CA was significantly higher than in the NS and VV samples
(P=0.0000 and P=0.0399, respectively; Fig. 1D). ANOVA also revealed that the
HMGB1 expression level in the epithelial nuclei of the cells in the
VV sample was significantly lower than in the NS (P=0.0194), and
there was no statistically significant difference between HMGB1 in
the epithelial nuclei of VV and CA samples (Fig. 1E). In addition, the expression of
HMGB1 in the associated inflammatory cells of VV and CA was
significantly higher than in the NS (P=0.0001 and P=0.0000,
respectively). No significant difference was identified between the
HMGB1 expression in the inflammatory cells in VV and CA samples
(Fig. 1F).
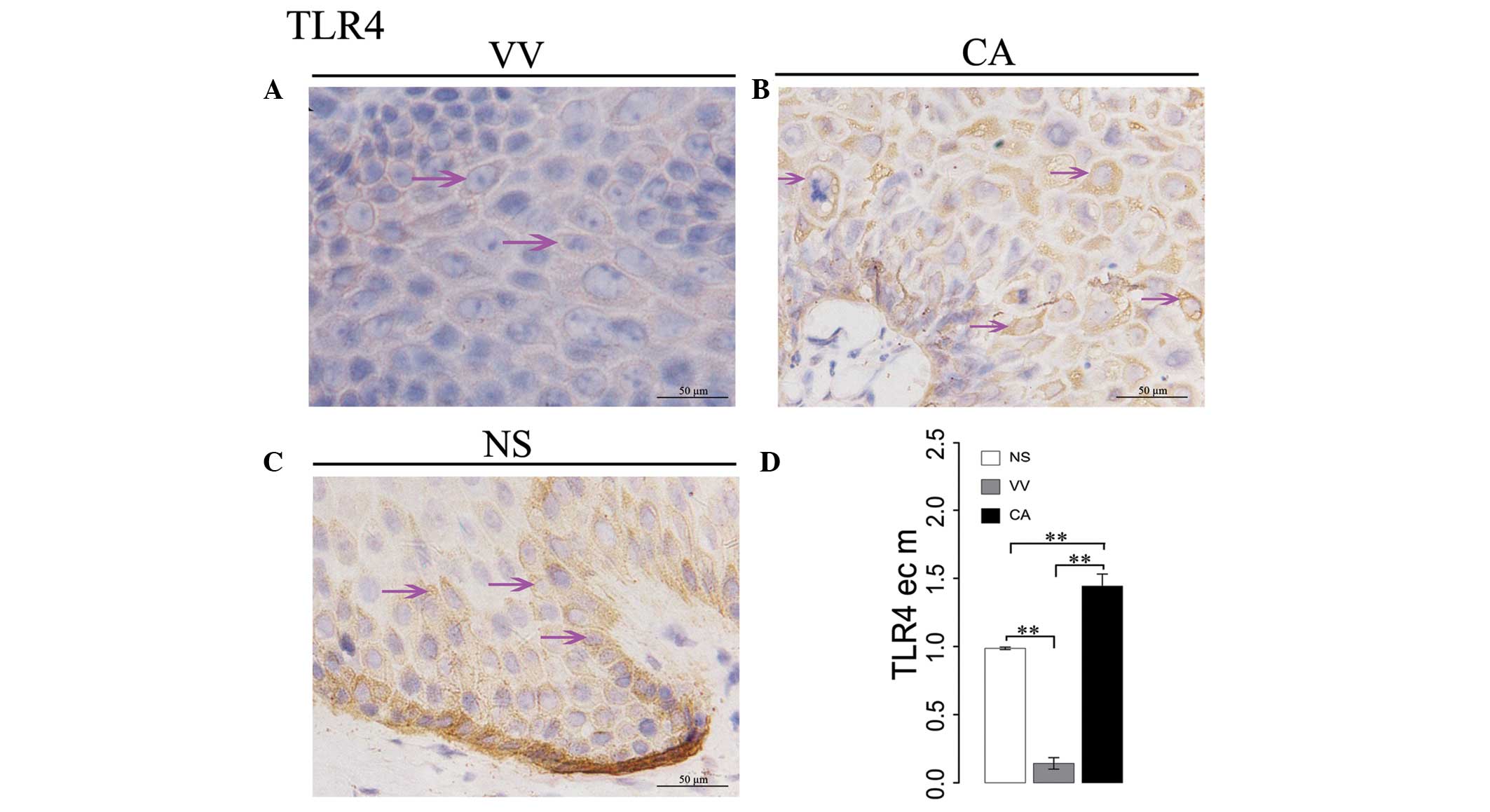
Expression of TLR4 in wart samples
In VV samples, only weak TLR4 expression was
observed on the surface of epithelial cell membranes (Fig. 2A). In CA samples, there was a weak
to moderately positive TLR4 expression on the surface of epithelial
membranes (Fig. 2B). In the NS,
weakly positive TLR4 expression was observed on the surface of the
epithelial cell membranes (Fig.
2C). ANOVA demonstrated that the expression of TLR4 on
epithelial cell membranes in CA samples was significantly higher
than in the NS (P=0.0010), and TLR4 on epithelial membranes in VV
samples was significantly lower than in the NS (P=0.0001; Fig. 2D).
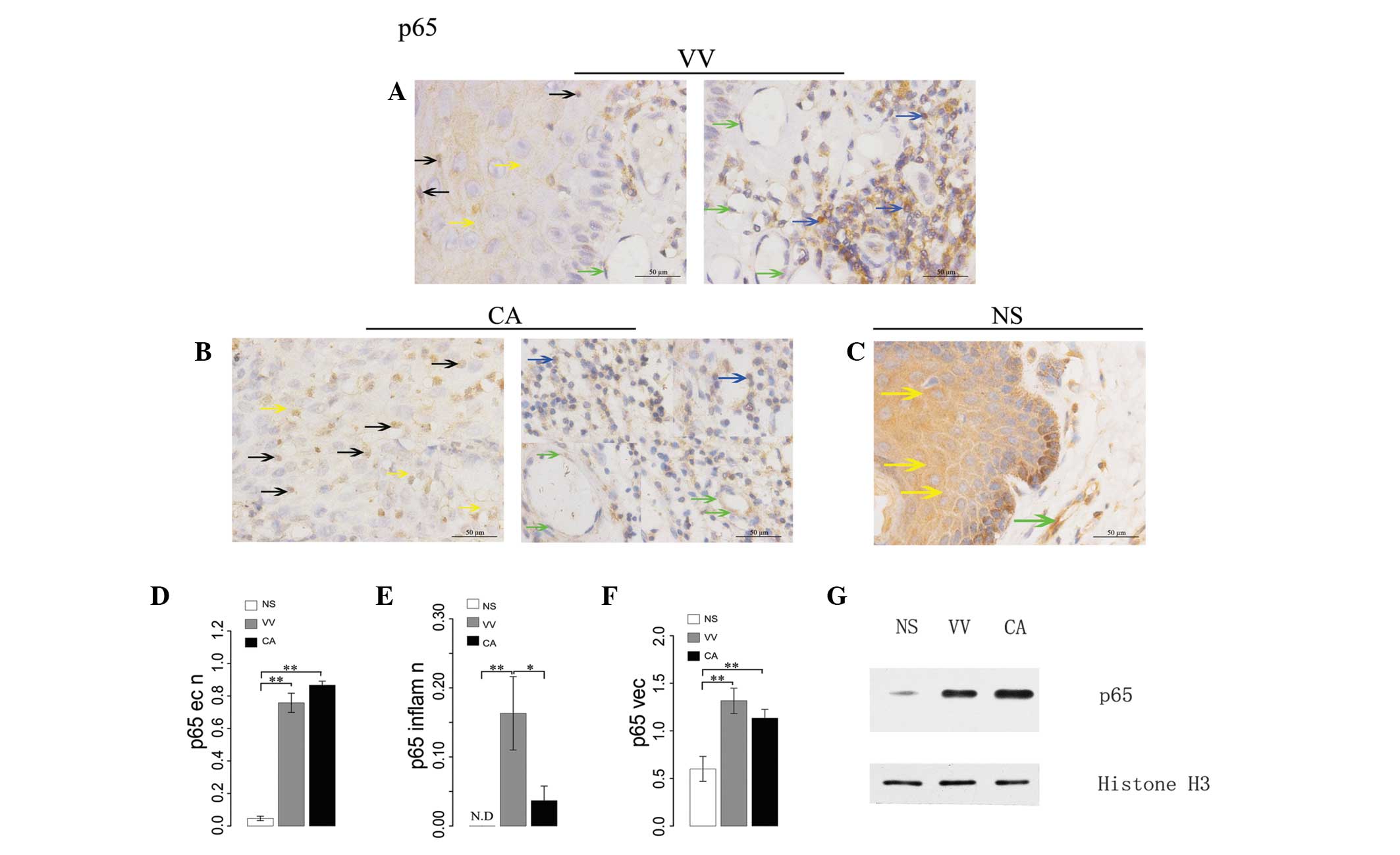
Expression of NF-κB p65 in warts
In VV samples, p65 exhibited weakly positive
expression in the epithelial nuclei and diffuse moderately positive
expression in the cytoplasm. In the associated inflammatory cells,
relatively high p65 expression was observed in the nuclei and a
weakly to moderately positive p65 expression in the cytoplasm was
noted. There was weakly to moderately positive p65 expression in
the associated vascular endothelial cells (Fig. 3A).
CA samples exhibited weakly positive p65 expression
in the nuclei and diffuse positive expression in the cytoplasm. In
the associated inflammatory cells, minimal p65 expression in the
nuclei and sporadic weakly positive expression in the cytoplasm was
identified. In addition, there was weakly positive p65 expression
in the associated vascular endothelial cells. (Fig. 3B).
In the NS, there was only minimal p65 expression in
the epithelial nuclei and focal positive expression in the
epithelial cytoplasm. Almost no p65 expression was observed in the
nuclei of associated inflammatory cells, and a degree of focal p65
expression was observed in the cytoplasm of the associated vascular
endothelial cells (Fig. 3C).
ANOVA demonstrated that the expression of p65 in the
epithelial nuclei of cells from VV and CA samples was significantly
higher than that in the NS (P=0.0001 and P=0.0000, respectively),
and that the p65 level in CA was higher than in VV samples, however
this difference was not significant (Fig. 3D). By contrast, the expression of
p65 in the inflammatory cell nuclei of VV samples was significantly
higher than in the NS (P=0.0049) and than in the CA samples
(P=0.0412). There was no significant difference between p65
expression of inflammatory cell nuclei in CA and NS samples
(Fig. 3E). ANOVA also demonstrated
that the expression of p65 in vascular endothelial cells of NS was
significantly lower than in VV (P=0.0007) and CA (P=0.0069)
samples, and there was no significant difference in p65 expression
in the vascular endothelial cells between VV and CA (Fig. 3F).
Western immunoblot analysis identified different
levels of p65 expression in the epithelial nuclei of the NS and the
wart specimens. There was only a weak p65 expression in the
epithelial nuclei of the NS, whereas the epithelial nuclei of CA
and VV demonstrated strong expression. In addition, p65 epithelial
nuclear expression in CA was higher than that observed in VV
(Fig. 3G).
Correlation analysis
Spearman’s correlation analysis demonstrated that
the expression of p65 in the epithelial nuclei of the NS and the CA
and VV samples was negatively correlated with the HMGB1 level in
the epithelial cell nuclei (r=−0.1947, P>0.05; Fig. 4A and G), was positively correlated
with p65 in the inflammatory cell nuclei (r=0.0536, P>0.05;
Fig. 4B and 4G), was positively
correlated with TLR4 on the surface of the epithelial cell
membranes (r=0.0949, P>0.05; Fig.
4C and G), with p65 in the vascular endothelial cells
(r=0.4355, P=0.0002; Fig. 4D and
G), with HMGB1 in the inflammatory cells (r=0.4879, P=0.0000;
Fig. 4E and G) and with HMGB1 in
the epithelial intercellular spaces (r=0.5199, P=0.0000; Fig. 4F and G).
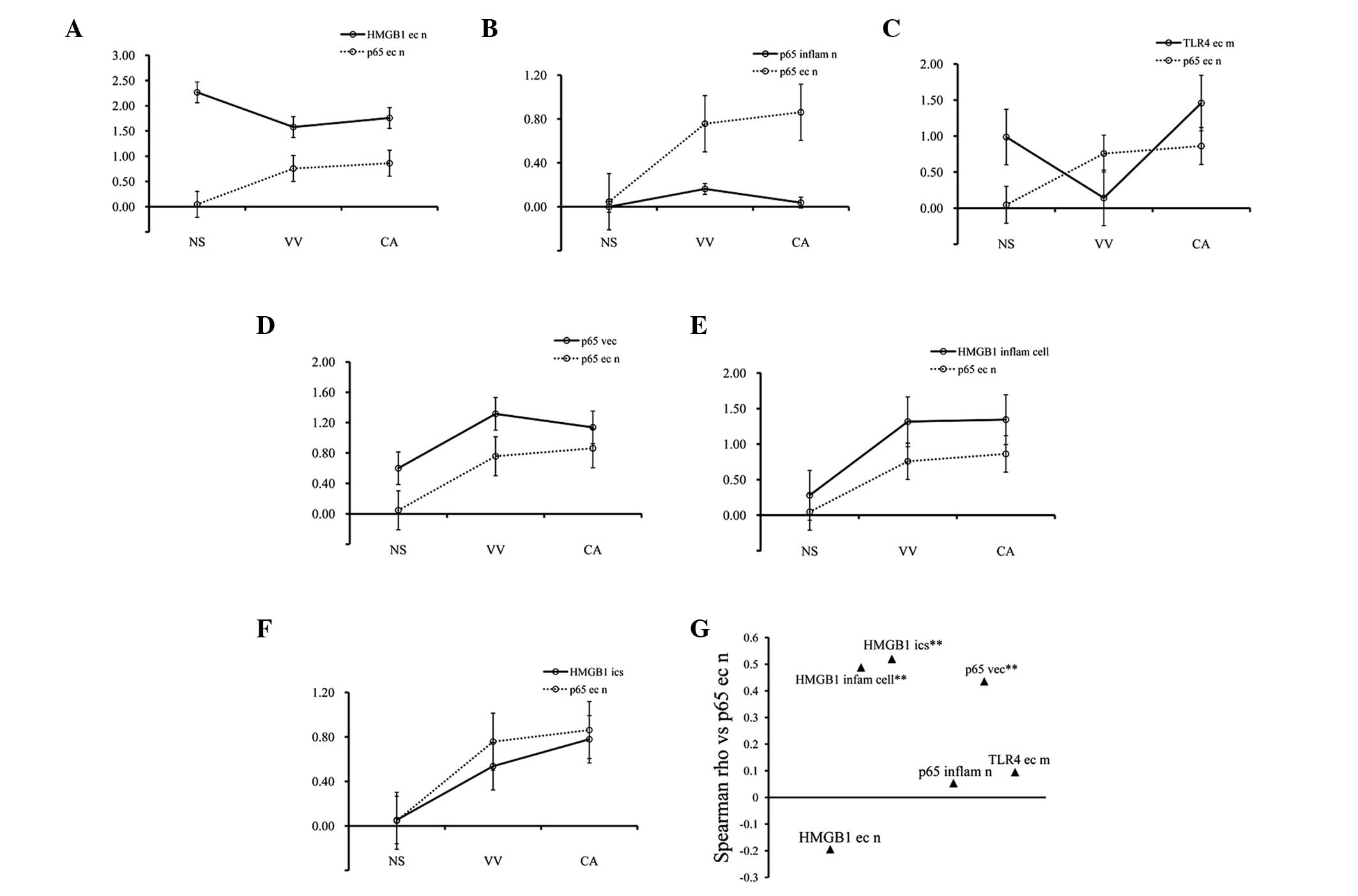 | Figure 4Correlation analysis of
immunohistochemistry. (A–F) The line-charts with the standard error
of the mean represent p65 expression in the epithelial cell nuclei
and other factors by Spearman’s correlation analysis. (G) The
Spearman’s correlation coefficients as compared with p65 in
epithelial cell nuclei. (A and G) p65 in epithelial cell nuclei vs.
HMGB1 in the epithelial nuclei (r=−0.1947, P>0.05). (B and G)
p65 in epithelial cell nuclei vs. p65 in the inflammatory nuclei
(r=0.0536, P>0.05). (C and G) p65 in epithelial cell nuclei vs.
TLR4 on the epithelial cell membranes (r=0.0949, P>0.05). (D and
G) p65 in epithelial cell nuclei vs. p65 in the vascular
endothelial cells (r=0.4355, **P<0.01). (E and G) p65
in epithelial cell nuclei vs. HMGB1 in the inflammatory cells
(r=0.4879, **P<0.01). (F and G) p65 in epithelial
cell nuclei vs. HMGB1 in the epithelial intercellular spaces
(r=0.5199, **P<0.01). HMGB1, high mobility group
protein box 1; TLR4, toll like receptor 4; CA, condyloma
acuminatum; VV, verruca vulgaris; NS, normal skin. |
Discussion
CA and VV are the most common epidermal infection
caused by HPVs. The majority of HPV infections are eliminated by an
effective immune response (12,13).
Failure to eradicate HPVs promotes wart development and is mainly
associated with the ability of HPV to avoid the immune defense
system of the body (2). This
process also involves a series of inflammatory responses, which
have been identified in recent years. Pro-inflammatory cytokines,
including several interleukins and tumor necrosis factor, are
important mediators of the development of skin and mucosal
inflammation (14,15). There is evidence that epidermal
cells are able to produce cytokines in response to diverse stimuli,
including viral infection (14).
However, data detailing the role of the HMGB1-TLR4
pathway-associated inflammation in the HPV-related epidermal warts
are lacking. The present study focused on the HMGB1 expression in
the epithelial intercellular spaces, TLR4 expression on the
epithelial cell membrane surface and p65 expression in the
epithelial nuclei in VV and CA.
HMGB1 is an essential component of mammalian
chromatin in the nuclei (16). It
is able to reach the intercellular spaces either through passive
release from necrotic cells (17),
or active secretion by macrophages and monocytes (18). Extracellular HMGB1 is a novel
pro-inflammatory cytokine, which acts by binding to TLR4, TLR2 or
receptors for advanced glycation end-products (19,20).
The immunohistochemistry results of the present study revealed that
HMGB1 expression was present in the epithelial intercellular spaces
in CA and VV samples as well as in the NS, and its expression level
increased significantly and progressively from the NS to VV to CA
(P<0.05). These findings imply that intracellular HMGB1 is
released outside of the HPV-infected cells to act as an
extracellular mediator of local inflammation. The difference in
extracellular HMGB1 expression between CA and VV, may be due to the
fact that the two tumors are caused by variant HPV types.
Expectedly, HMGB1 epithelial nuclear expression in the NS was
significantly higher than in VV (P<0.05), and there was no
significant difference identified in the HMGB1 epithelial nuclear
expression between VV and CA. This result is in accordance with our
recent study investigating the HMGB1 expression in a number of
malignant epidermal tumors (11).
This result also suggests that HMGB1 expression in the epithelial
nuclei may have a function in the stabilization of DNA and
chromosomes rather than acting as a factor that leads to wart
development. In addition, HMGB1 expression in the associated
inflammatory cells in VV and CA samples was higher than in the NS
(P<0.01), and the correlation analysis demonstrated that the
level of HMGB1 in inflammatory cells was positively correlated with
the level of p65 in the epithelial nuclei (r=0.4879, P<0.01).
These results suggest that HMGB1 may have a key role in
inflammation in the HPV-associated warts.
Extracellular HMGB-1 has been recognized to be
involved in the activation of NF-κB through its receptor TLR4
(7). The interference with the
expression of TLRs may effect the activation of antigen presenting
cells and phagocytes to enhance innate and adaptive responses
against pathogens, and therefore induce tumorigenesis and
development (21,22). From the present results that
demonstrated the signifcantly lower expression of TLR4 on the
surface of epithelial membranes in VV compared to normal skin
(P<0.01), it is deduced that TLR4 signaling may not be involved
in inducing inflammation in VV. Instead, the impairment of TLR4 may
be a factor for the development of VV. The present study also
demonstrated that the TLR4 expression on the surface of the
epithelial membrane in CA samples was significantly higher than
that of the NS (P<0.01), and that the extracellular HMGB1
expression in CA was significantly higher than in the NS
(P<0.01). These findings indicate that extracellular HMGB1 may
act as a pro-inflammatory factor to interact with TLR4, and that
the HMGB1-TLR4 signaling pathway-associated inflammation may have a
significant role in CA.
The NF-κB family mainly includes RELA (p65), NF-κB1
(p50; p105), NF-κB2 (p52; p100), c-REL and RELB (23,24).
NF-κB is sequestered in the cytoplasm inactively and bound by the
inhibitor proteins (24).
Activated by various stimuli, including different pro-inflammatory
cytokines, NF-κB is transported to the nuclei, where it binds with
sequences of various genes to induce the expression of inflammatory
factors, promote cell proliferation and prevent apoptosis, and may
therefore have a role in tumorigenesis and tumor development
(25,26). A previous study demonstrated that
NF-κB expression and activation are abnormal in human epidermal
tumors of seborrheic keratosis, basal cell carcinoma and squamous
cell carcinoma (11). The present
study revealed that there was weak positive expression of p65 in
the epithelial nuclei of VV and CA samples, while in the NS there
was only minimal p65 expression in the epithelial cell nuclei.
ANOVA demonstrated that the p65 expression in the epithelial nuclei
of VV and CA samples was significantly higher than in the NS
(P<0.01). These findings indicate that p65 is transported to the
nuclei due to NF-κB activation, and thus induces the expression of
inflammatory factors, promoting cell proliferation and preventing
apoptosis in the warts. Western blot analysis also demonstrated a
significant p65 expression in the epithelial nuclei of CA and VV
samples, whereas only a weak p65 expression in the NS was
identified. NF-κB is sequestered and bound by the IkB family of
inhibitor proteins in the cytoplasm in its inactive form. Various
stimuli, including proinflammatory cytokines and viral replication
are able to activate NF-κB by triggering the phosphorylation of
IkB, and induce its subsequent molecular translocation to the
nucleus. As expected, the immunohistochemistry and western blotting
results proved that the level of p65 expression in the epithelial
nuclei, which is indicative of NF-κB activation and inflammation
responses, was increased and significantly higher in VV and CA than
in the NS (P<0.01), suggesting that inflammation may be
intensified in the HPV-infected epithelial cells, which may lead to
the development of warts. The expression of p65 in the vascular
endothelial cells in the VV and CA samples was significantly higher
than in the NS (P<0.01). In addition, the p65 level in the
vascular endothelial cells was positively correlated with the p65
level in the epithelial nuclei of the tissues (r=0.4355,
P<0.01), indicating that NF-κB may activate the transcription
and translation of genes to promote angiogenesis in the warts.
Furthermore, the expression of p65 in the associated
inflammatory cell nuclei in the VV samples was higher than the
other tissues (P<0.05). Together with the results of the HMGB1
inflammatory cell expression, it is deduced that the inflammation
in VV may be mediated by inflammatory cells, and that the
inflammation of CA may be induced by infected epithelial cells
rather than inflammatory cells. Finally, correlation analysis
demonstrated that the level of HMGB1 in epithelial intercellular
spaces was positively correlated with the level of p65 in the
epithelial nuclei (r=0.5199, P<0.01), further indicating that
HMGB1 may be a mediator in inducing the local inflammation in the
warts.
In conclusion, the present study demonstrated that
inflammation was intensified in VV and CA samples. The HMGB1 and
TLR4 signaling pathway-associated inflammation may have a central
role in CA. Furthermore, HMGB1 may be one of mediators resulting in
the progression of inflammation in VV, while the TLR4 signaling
pathway does not appear to have a role in inducing inflammation in
VV. Therefore, anti-HMGB1 therapy may be useful for the treatment
of the warts caused by HPVs prior to their development into
malignant lesions.
Acknowledgements
This study was supported by the Natural Science
Foundation of China (grant no. 81373152).
Abbreviations:
|
CA
|
condyloma acuminatum
|
|
HMGB1
|
high mobility group protein box 1
|
|
HPV
|
human papillomavirus
|
|
NF-κB
|
nuclear factor-κB
|
|
TLR4
|
toll like receptor 4
|
|
VV
|
verruca vulgaris
|
References
|
1
|
Bernard HU, Burk RD, Chen Z, van Doorslaer
K, zur Hausen H and de Villiers EM: Classification of
papillomaviruses (PVs) based on 189 PV types and proposal of
taxonomic amendments. Virology. 401:70–79. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Stanley MA, Pett MR and Coleman N: HPV:
from infection to cancer. Biochem Soc Trans. 35:1456–1460. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Aggarwal BB, Shishodia S, Sandur SK,
Pandey MK and Sethi G: Inflammation and cancer: how hot is the
link? Biochem Pharmacol. 72:1605–1621. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Balkwill F and Coussens LM: Cancer: an
inflammatory link. Nature. 431:405–406. 2004. View Article : Google Scholar
|
|
5
|
Thomas JO and Travers AA: HMG1 and 2, and
related ‘architectural’ DNA-binding proteins. Trends Biochem Sci.
26:167–174. 2001.
|
|
6
|
Tadie J, Bae HB, Deshane J, et al:
Toll-like receptor 4 engagement inhibits adenosine
5′-monophosphate-activated protein kinase activation through a high
mobility group box 1 protein-dependent mechanism. Mol Med.
18:659–668. 2012.PubMed/NCBI
|
|
7
|
Dai S, Sodhi C, Cetin S, et al:
Extracellular high mobility group box-1 (HMGB1) inhibits enterocyte
migration via activation of Toll-like receptor-4 and increased
cell-matrix adhesiveness. J Biol Chem. 285:4995–5002. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Naugler WE and Karin M: NF-kappaB and
cancer-identifying targets and mechanisms. Curr Opin Genet Dev.
18:19–26. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fiuza C, Bustin M, Talwar S, et al:
Inflammation-promoting activity of HMGB1 on human microvascular
endothelial cells. Blood. 101:2652–2660. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Poser I, Golob M, Buettner R and
Bosserhoff AK: Upregulation of HMG1 leads to melanoma inhibitory
activity expression in malignant melanoma cells and contributes to
their malignancy phenotype. Mol Cell Biol. 23:2991–2998. 2003.
View Article : Google Scholar
|
|
11
|
Weng H, Deng Y, Xie Y, Liu H and Gong F:
Expression and significance of HMGB1, TLR4 and NF-κB p65 in human
epidermal tumors. BMC Cancer. 13:3112013.
|
|
12
|
Frazer IH: Interaction of human
papillomaviruses with the host immune system: a well evolved
relationship. Virology. 384:410–414. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Scott M, Nakagawa M and Moscicki AB:
Cell-mediated immune response to human papillomavirus infection.
Clin Diagn Lab Immunol. 8:209–220. 2001.PubMed/NCBI
|
|
14
|
Gröne A: Keratinocytes and cytokines. Vet
Immunol Immunopathol. 88:1–12. 2002.
|
|
15
|
Locksley RM, Killeen N and Lenardo MJ: The
TNF and TNF receptor superfamilies: integrating mammalian biology.
Cell. 104:487–501. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Vaccari T, Beltrame M, Ferrari S and
Bianchi ME: Hmg4, a new member of the Hmg1/2 gene family. Genomics.
49:247–252. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zeh HJ III and Lotze MT: Addicted to
death: invasive cancer and the immune response to unscheduled cell
death. J Immunother. 28:1–9. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ulloa L and Messmer D: High-mobility group
box 1 (HMGB1) protein: friend and foe. Cytokine Growth Factor Rev.
17:189–201. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim SW, Lim CM, Kim JB, et al:
Extracellular HMGB1 released by NMDA treatment confers neuronal
apoptosis via RAGE-p38 MAPK/ERK signaling pathway. Neurotox Res.
20:159–169. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nogueira-Machado JA, Volpe CM, Veloso CA
and Chaves MM: HMGB1, TLR and RAGE: a functional tripod that leads
to diabetic inflammation. Expert Opin Ther Targets. 15:1023–1035.
2011. View Article : Google Scholar
|
|
21
|
Kawai T and Akira S: The roles of TLRs,
RLRs and NLRs in pathogen recognition. Int Immunol. 21:317–337.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hasan UA, Bates E, Takeshita F, et al:
TLR9 expression and function is abolished by the cervical
cancer-associated human papillomavirus type 16. J Immunol.
178:3186–3197. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Verma IM, Stevenson JK, Schwarz EM, Van
Antwerp D and Miyamoto S: Rel/NF-kappa B/I kappa B family: intimate
tales of association and dissociation. Genes Dev. 9:2723–2735.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ghosh S, May MJ and Kopp EB: NF-kappa B
and Rel proteins: evolutionarily conserved mediators of immune
responses. Annu Rev Immunol. 16:225–260. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ghosh S and Hayden MS: New regulators of
NF-kappaB in inflammation. Nat Rev Immunol. 8:837–848. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Karin M and Greten FR: NF-kappaB: linking
inflammation and immunity to cancer development and progression.
Nat Rev Immunol. 5:749–759. 2005. View
Article : Google Scholar : PubMed/NCBI
|