Introduction
The emerging discipline of gaseous biology in
traditional Chinese medicine, has attracted significant attention
(1). Various gaseous signaling
molecules, including hydrogen sulfide (H2S) (2), carbon monoxide (3) and nitric oxide (NO) (4) are involved in regulating homeostasis
during acupuncture, in which NO as a messenger has been
well-documented under pathological and physiological conditions
(5).
NO, also termed endothelium-derived relaxing factor
(6), is one of the few gaseous
signaling molecules known. NO is biosynthesized from L-arginine,
oxygen and nicotinamide adenine dinucleotide phosphate by various
nitric oxide synthase (NOS) enzymes in vivo (7), including neuronal NO synthase (nNOS),
endothelial NO synthase and inducible NO synthase. NO can regulate
various biological processes in vertebrates, including the
regulation of blood flow (8),
blood flow metabolism coupling (9), neurotransmission (10), memory formation (11) and the prevention of apoptosis in
neurons (12). In particular, NO
is involved in regulating hypoxic-ischemic brain damage (HIBD)
(13). Excessive levels of NO can
cause reperfusion injury by reacting with superoxide to produce the
oxidant peroxynitrite (14),
indicating that downregulating the content of NO in cortical cells
may facilitate reperfusion injury recovery. Our previous study
(15) demonstrated that HIBD
upregulates the content of NO in rat cortical cells and that
electroacupuncture (EA) can protect this damage by downregulating
the NO content in cortical cells. However, the underlying mechanism
remains to be elucidated. Therefore, the present study aimed to
investigate the potential neuroprotective mechanism of NO
downregulation by EA, including the NF-κB/nNOS pathway.
In addition, cystathionine-β-synthase (CBS) is a
multi-domain enzyme, located mainly in the brain and nervous system
(16,17). It is able to catalyze the
transsulfuration pathway to generate H2S. H2S
has various physiological effects, including cysteine
S-sulfhydration (18,19), preventing cytokine or
oxidant-induced oxidative damage (20), inhibiting the expression of
proinflammatory factors by downregulating the activation of NF-κB
(21) or upregulating the
expression of heme oxygenase 1 (22). Therefore, in the present study it
was hypothesized that the NF-κB/nNOS and H2S/CBS
pathways crosstalk in the HIBD model. Consequently, the CBS
activator, S-adenosyl-L-methionine (SAM) and the CBS inhibitor,
hydroxylamine (HA) were used on the basis of the HIBD model.
Materials and methods
Animals and construction of the HIBD
model
A total of 96 specific pathogen-free Sprague-Dawley
rats (1 week-old, 12.9–21.0 g) were purchased and raised in the
Laboratory Animal Center of the Academy of Military Medical
Sciences (Beijing, China). The animals were housed at a temperature
of 25±2°C with a 12 h light/dark cycle and were breast fed by their
mothers. Each cage contained eight baby rats and their mother. The
animals were randomly divided into eight groups (n=12): Sham, Sham
+ EA, HIBD, HIBD + EA, HIBD + SAM, HIBD + SAM + EA, HIBD + HA and
HIBD + HA + EA. The rats were sacrificed using diethyl ether and
the four limbs were fixed to enable incision along the neck
midline. The left carotid artery communis was exposed through
stripping of the thyroid, vein and nervous tissues, which was then
ligated using 5/0 surgical line and sutured. After 2 h, the rats
were placed in a low-oxygen tank to maintain an appropriate
environmental temperature under continuous hypoxia with 8% oxygen
and 92% nitrogen for 2 h. The Sham-operated groups were subjected
to surgery, which also involved the exposure of the left carotid
artery communis, however no ligation was performed. This experiment
was approved by the Ethics Committee of the Chinese People’s
Liberation Army General Hospital (Beijing, China).
Intervention experiment of EA
The rats in the EA group were acupunctured at the
BaiHui acupoint, which is the crossing point either side of the
skull and linkline of the two ears and DaZhui acupoint, which lies
below the detail of the cervical spine using EA (~0.25 mm in
diameter and 10 mm in length, frequency, 2/100 Hz; intensity, 3 mA)
for a 30 min period for 14 days. The limbs of the rats in the
control group were simultaneously fixed down, but EA was not
performed. The rats in the HIBD + HA and HIBD + HA + EA groups were
injected with 12.5 mg/kg/d HA (Sigma-Aldrich, St. Louis, MO, USA),
an inhibitor of CBS, via the peritoneal cavity 20 min prior to the
acupuncture procedure or fixation. Similarly, the rats in the HIBD
+ SAM and the HIBD + SAM + EA groups were injected with 50 mg/kg/d
SAM (Sigma-Aldrich), an activator of CBS, via the peritoneal cavity
20 min prior to EA or fixation. The control group was injected with
an equal volume of normal saline. Subsequently, six rats from each
group were sacrificed by cervical dislocation and the brain cortex
tissues were obtained to determine the NO content. Tissues from the
remaining six animals in each group were perfused with 4%
paraformaldehyde for slicing and Nissl and immunohistochemistry
(IHC) staining.
Determination of the NO content in the
rat cortex cells
The brain cortex tissue (~50 mg) was homogenize in
nine volumes (w/v, ~0.5 ml) of 0.9% ice-cold sodium chloride. The
homogenate was centrifuged at 2,500 × g for 10 min and 200 μl
supernatant was obtained to measure the protein concentration. The
absolute absorbance value (A550 nm) was determined
according to the manufacturer’s instructions and the NO content was
calculated using the following formula: NO = (Asample −
Ablank)/(Astandard − Ablank) × X ×
Y, where X represents the standard sample concentration (μmol/l)
and Y represents the protein concentration (g/l). Column chart
analysis was performed using Origin 9.0 software (OriginLab Co.,
Northampton, MA, USA). Each experiment was repeated at least three
times.
Nissl staining
The slides (3–4 μm) were deparaffinized and
rehydrated, following which the frozen or vibratome sections were
mounted onto the slides and rehydrated. The sections were partially
over-stained with Nissl for ~5 min. The excess stain was removed
with tap water, followed by 100% ethanol for 1 min. The sections
were transferred to dimethylbenzene (Changhai Chemical Factory,
Beijing, China) for 1 h and differentiated with 95% ethanol. The
sections were then dehydrated and mounted with neutral balsam.
Images of the cortex were captured using a microscope connected to
a CCD camera (magnification, ×200; Olympus BX-41; Olypmus, Tokyo,
Japan). Each experiment was repeated at least three times.
IHC assay
The slides (3–4 μm) were deparaffinized, rehydrated,
post-fixed with 4% paraformaldehyde for 10 min and then washed
three times with 0.01 M phosphate-buffered saline (PBS). Endogenous
peroxidase was inactivated by incubating the sections in 3%
H2O2 for 30 min. The sections underwent
sequential incubations with 10% normal goat serum in 0.01 M PBS for
30 min at room temperature. The sections were incubated in rabbit
anti-nNOS (cat no. ZS-648; 1:100; Beijing Zhongshan, Golden Bridge
Biotechnology Co., Ltd, Beijing, China) and rabbit anti-NF-κB (cat
no. ab1650; 1:400; Abcam, Cambridge, UK) antibodies in PBS
containing 0.3% Triton X-100 overnight at 4°C. Following this, the
sections were washed three times with PBS for 5 min each and then
incubated in peroxidase-conjugated goat anti-rabbit IgG (1:200;
Zymed, San Fransisco, CA, USA) for 1 h at room temperature.
Subsequently, the sections were developed with diaminobenzidine
(Sigma, St. Louis, MO, USA) in 0.1 M tris-buffered saline
containing 0.001% H2O2 for 30–50 min.
Immunoreactions were observed under a microscope (Olympus, Tokyo,
Japan). For image analysis, the IHC sections were captured using a
microscope connected to a CCD camera (magnification, ×200). Images
of five specific areas in each region of the monitor were captured.
The quantity of immunopositive cells and total positive area in the
assigned subregions was measured using Image-Pro Plus 7.0 software
(Media Cybernetics, Inc., Rockville, MD, USA) and column chart
analysis was performed using Origin 9.0 software (OriginLab).
Statistical analysis
All data are expressed as the mean ± standard
deviation. Statistical analysis was performed using SPSS software
(version 21.0; IBM, Armonk, NY, USA) and Student’s t-test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Downregulation of NO in the cortex of
HIBD rats by EA
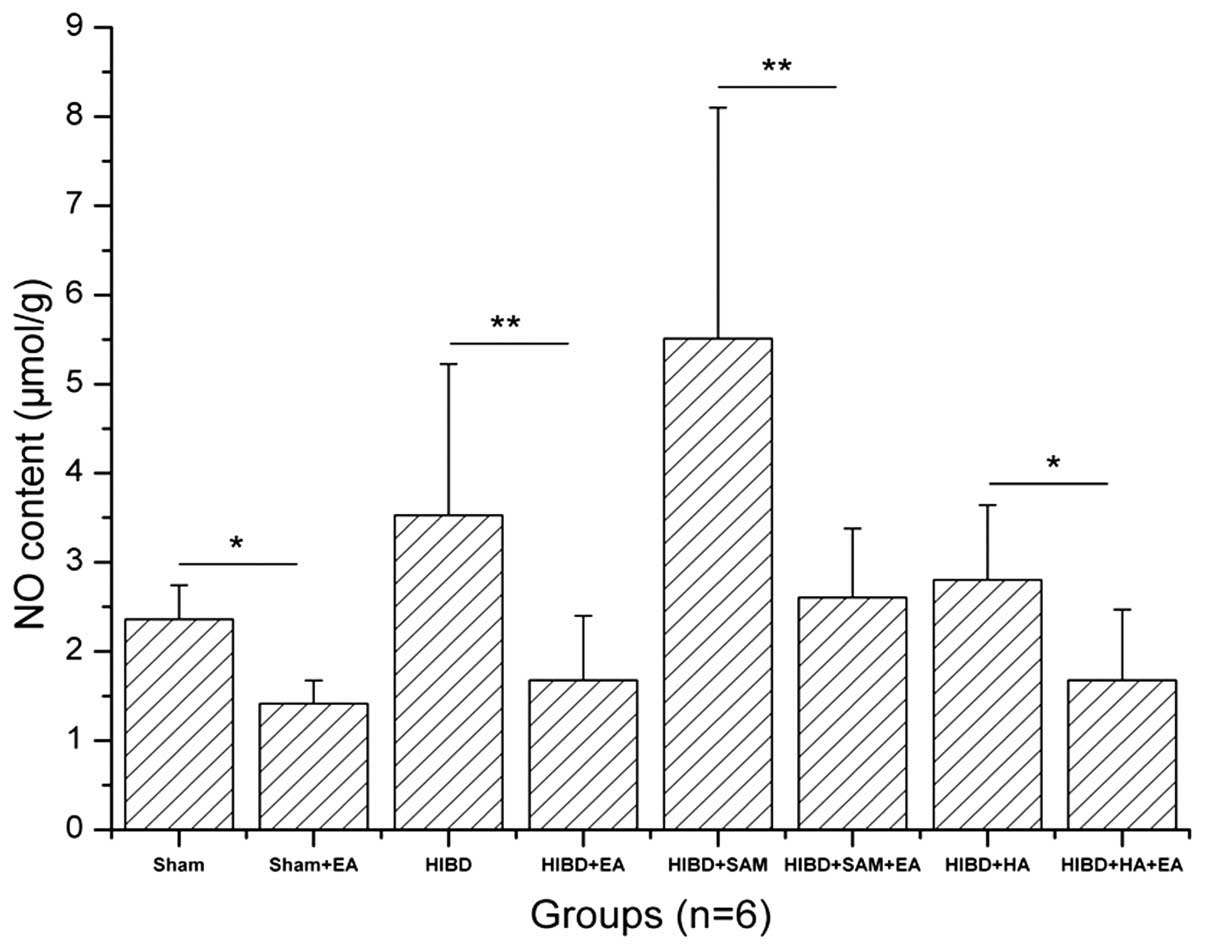
The NO content in the cortex of the Sham, Sham + EA,
HIBD, HIBD + EA, HIBD + SAM, HIBD + SAM + EA, HIBD + HA and HIBD +
HA + EA groups was 2.3614±0.3807, 1.4165±0.2592, 3.5269±1.6970,
1.6787±0.7213, 5.5101±2.5914, 2.6041±0.7773, 2.8041±0.8377 and
1.6784±0.7917, respectively (Table
I). HIBD significantly upregulated the NO content in the cortex
cells compared with the Sham group. In addition, treatment with SAM
further upregulated the NO content of the cortex cells in the HIBD
rats compared with the Sham group. However, treatment with HA
downregulated the NO content of the cortex cells compared with the
Sham group. Furthermore, EA treatment downregulated the NO content
in the Sham + EA, HIBD + EA, HIBD + SAM + EA and HIBD + HA + EA
groups compared with those of the control groups, including Sham
(*P<0.05), HIBD (**P<0.01), HIBD + SAM
(**P<0.01) and HIBD + HA (*P<0.05),
particularly in the HIBD and HIBD + SAM groups (Fig. 1).
 | Table INO content assay in rat cortex
cells. |
Table I
NO content assay in rat cortex
cells.
| Data Groups | NO content (mean ±
SD; n=6; μmol/g) |
|---|
| Sham | 2.3614±0.3807 |
| Sham + EA | 1.4165±0.2592a |
| HIBD | 3.5269±1.6970 |
| HIBD + EA | 1.6787±0.7213b |
| HIBD + SAM | 5.5101±2.5914 |
| HIBD + SAM + EA | 2.6041±0.7773b |
| HIBD + HA | 2.8041±0.8377 |
| HIBD + HA + EA | 1.6784±0.7917a |
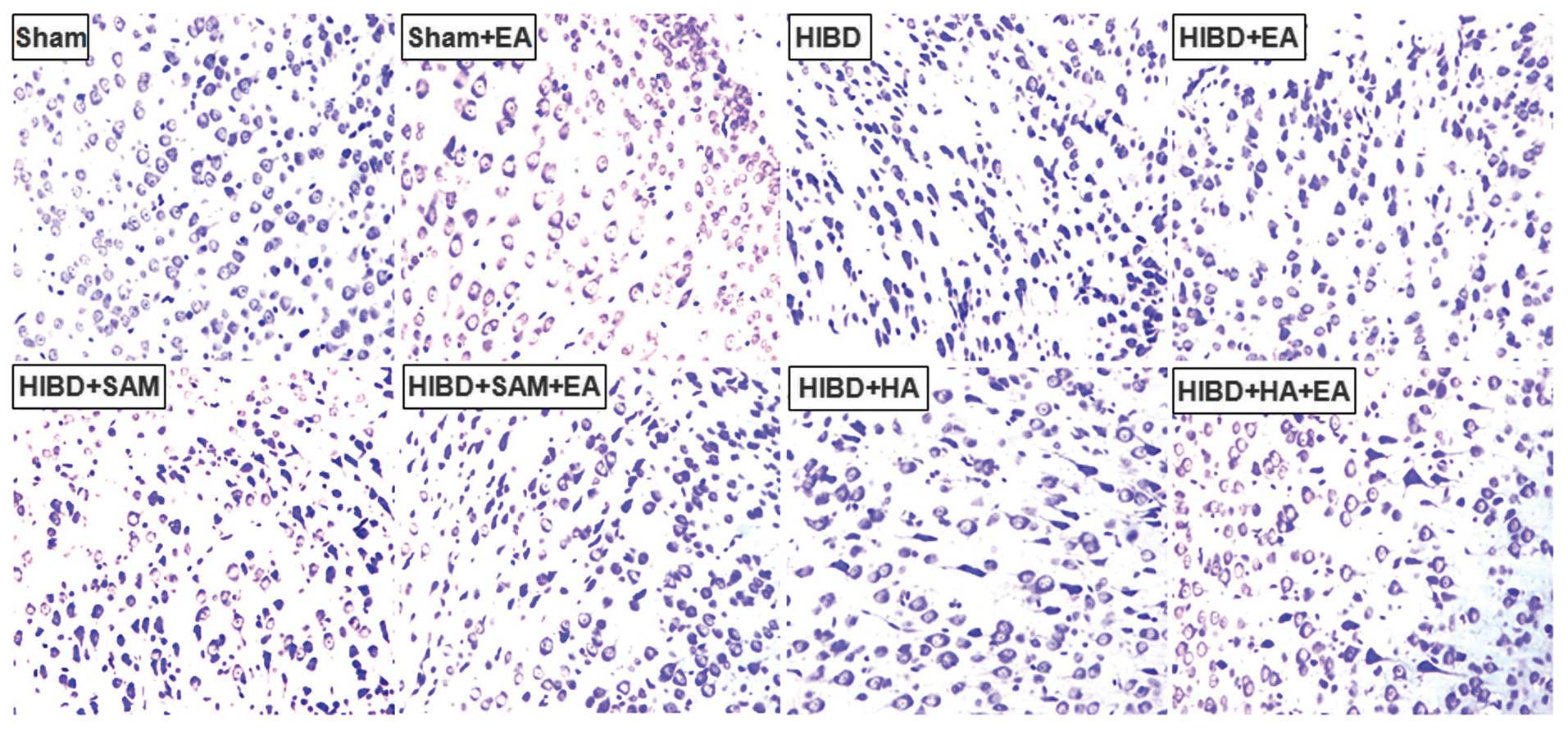
Alleviation of cell damage in the cortex
of the HIBD rats by EA
EA treatment alleviated the damage to the cortex in
the HIBD rats and decreased the number of positive cells in the
Sham + EA, HIBD + EA, HIBD + SAM + EA and HIBD + HA + EA groups
compared with those of the control groups, including the Sham,
HIBD, HIBD + SAM and HIBD + HA groups (Fig. 2). This result indicated that
hypoxia triggered severe damage to the cortex cells, which was
aggravated by the CBS activator SAM, but was alleviated by the CBS
inhibitor HA (Fig. 2).
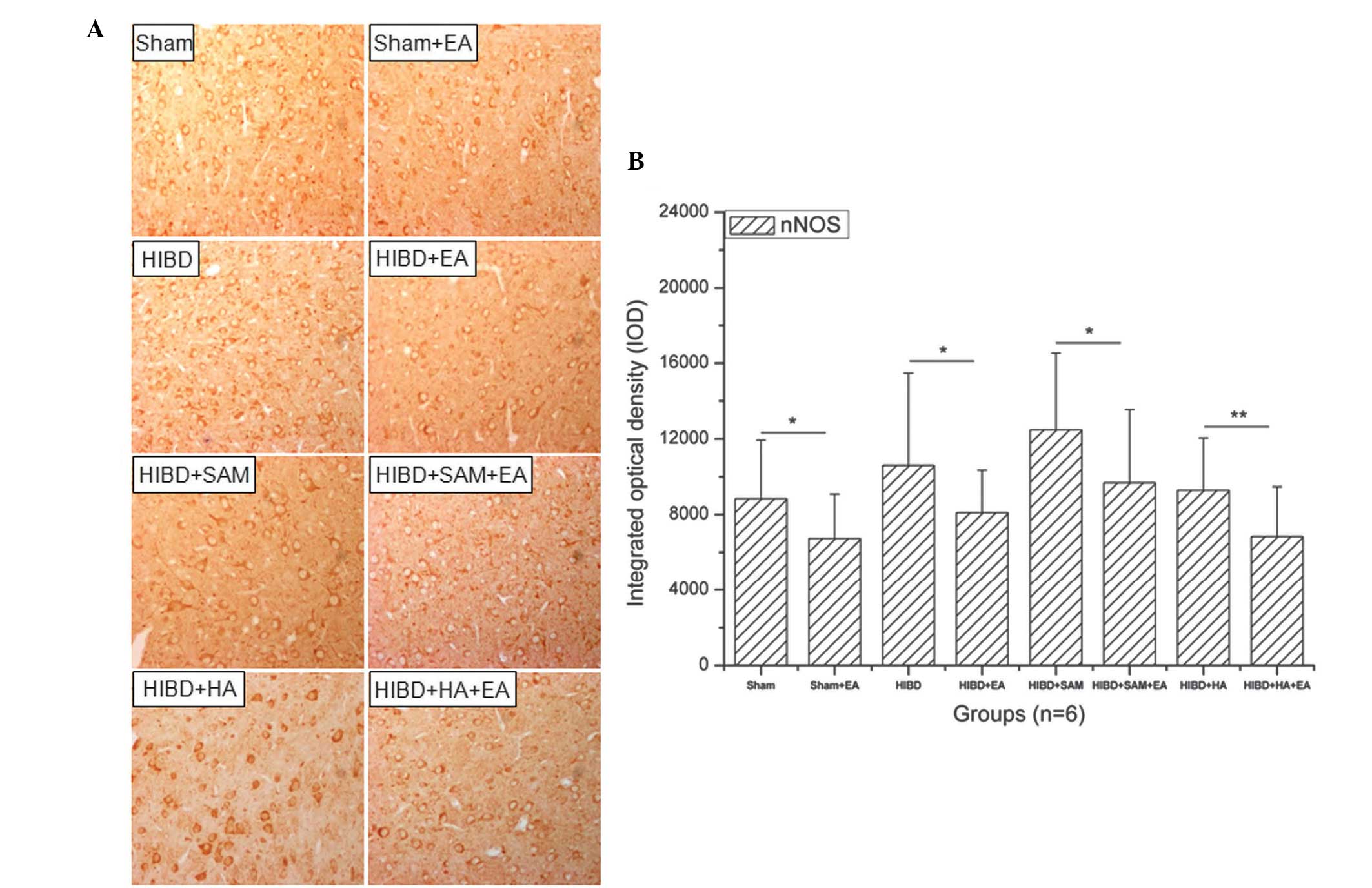
Downregulation of the expression of nNOS
in the cortex cells of HIBD rats by EA
HIBD upregulated the expression of nNOS in the
cortex cells compared with the Sham group and treatment with SAM
significantly upregulated the expression of nNOS in the cortex
cells of the HIBD rats compared with the Sham group. However,
treatment with HA downregulated the expression of nNOS in the
cortex cells compared with the Sham group. In addition, treatment
with EA downregulated the expression of nNOS in the cortex cells of
the HIBD rats compared with that of the control groups (Fig. 3A). The expression of nNOS was
significantly downregulated following treatment with EA (Fig. 3B). A significant difference in the
expression of nNOS was identified between the Sham + EA and the
Sham groups (Fig. 3B;
*P<0.05). Similarly, a significant difference in the
expression of nNOS was also identified between the other EA and the
control groups, including HIBD (Fig.
3B; *P<0.05), HIBD + SAM (Fig. 3B; *P<0.05) and HIBD +
HA (Fig. 3B;
*P<0.01).
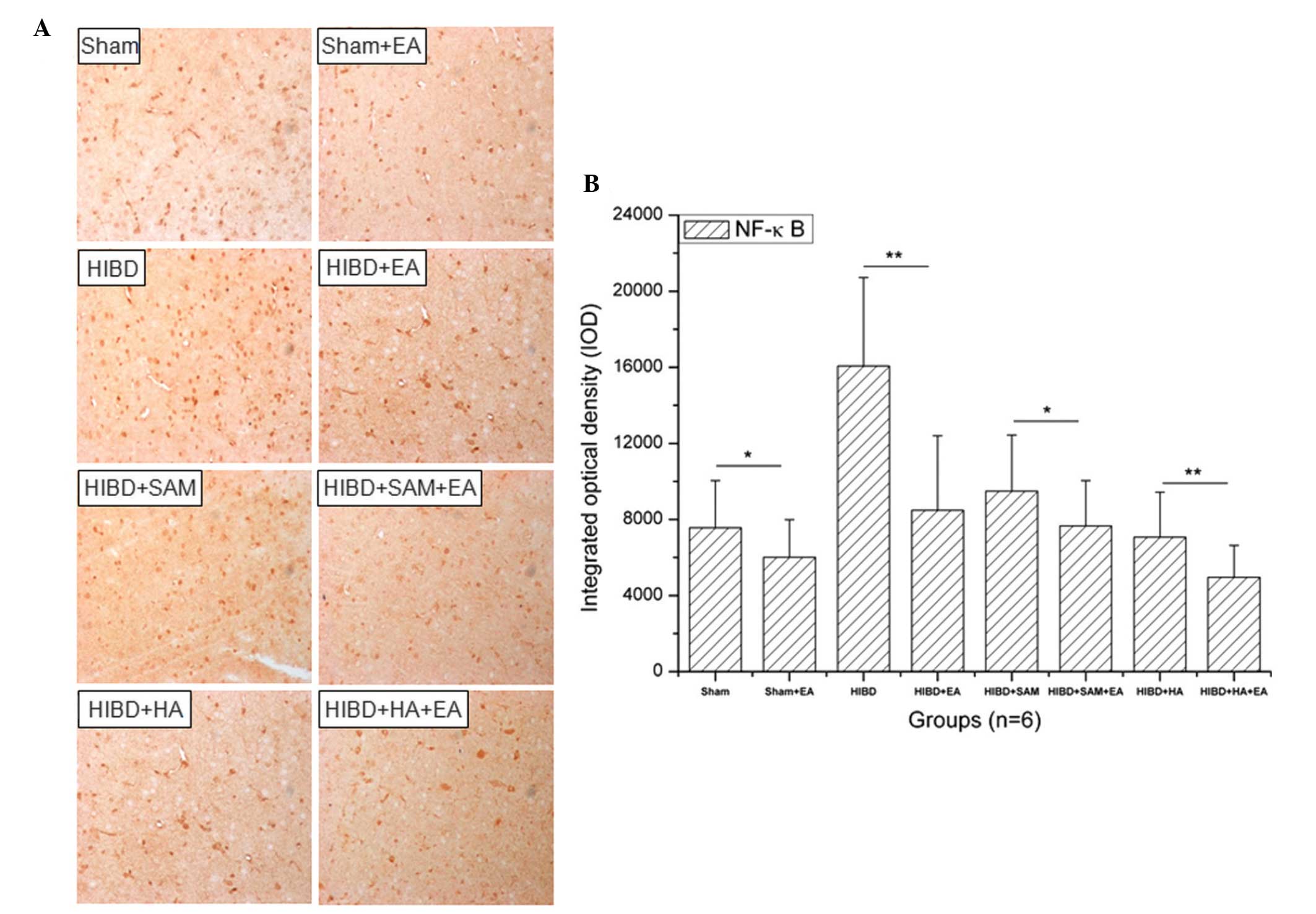
Downregulation in the expression of NF-κB
in the cortex cells of HIBD rats by EA
HIBD upregulated the expression of NF-κB in the
cortex cells compared with the Sham group and treatment with SAM
significantly upregulated the expression of NF-κB in the cortex
cells of HIBD rats compared with Sham treatment. However, treatment
with HA downregulated the expression of NF-κB in the cortex cells
of HIBD rats compared with the Sham group and treatment with EA
downregulated the expression of NF-κB in the cortex cells compared
with the control groups (Fig. 4A).
A significant difference in the expression of NF-κB was observed
between the Sham + EA and the Sham groups (Fig. 4B; **P<0.05).
Similarly, a significant difference was identified in the
expression of NF-κB between the other EA groups and the control
groups, including HIBD (Fig. 4B;
**P<0.01), HIBD + SAM (Fig. 4B; *P<0.05) and HIBD +
HA (Fig. 4B;
**P<0.01).
Discussion
The present study demonstrated that treatment with
EA can downregulate the NO content of cortical cells and alleviate
cortex cell damage in HIBD rats. The number of positive cells
significantly decreased following treatment with EA compared with
each control. In addition, treatment with EA downregulated the
expression of nNOS and NF-κB in the rat cortex cells, whereas
treatment with SAM significantly upregulated the expression of nNOS
and NF-κB in the rat cortex cells. Treatment with HA significantly
downregulated the expression of nNOS and NF-κB in the rat cortex
cells. These results suggested that the NF-κB/nNOS and
H2S/CBS pathways may crosstalk during the recovery of
HIBD-induced neuron damage.
There are two important mechanisms underlying the
regulation of the biological function of NO, namely, S-nitrosation
of thiols (23) and nitrosylation
of transition metal ions (24).
S-nitrosation transfers thiol groups from the cysteine residues of
proteins to form S-nitrosothiols and nitrosylation is able to
transfer NO to a transition metal ion. Under physiological
conditions, NO acts as a tonic inhibitory modulator to regulate
carotid body chemosensory discharge by indirectly modifying
vascular tone and oxygen delivery and/or directly modulating the
excitability of the glomus cells and petrosal neurons (25). In addition, NO has a dual
dose-dependent effect on carotid body chemosensory discharge
(26).
Acupuncture is used as a curative tool in
traditional Chinese medicine and is significantly neuroprotective
in organisms through gaseous signaling molecules (27). Acupuncture increases the local
generation of NO (4) and increases
its content on the surface of the skin at acupoints (28). An increased level of NO has a
curative effect on brain damage in rats (29), including transient middle cerebral
artery occlusion and HIBD. Acupuncture is also able to cause a
decrease in the nNOS/NO system to recover neuronal function,
however, the underlying mechanism remains to be elucidated
(15). In the present study,
treatment with EA downregulated the NO content of the cortical
cells in the Sham + EA group as well as following HIBD and
treatment with SAM and HA. Treatment with EA also had a significant
curative effect on HIBD-induced rat brain damage and downregulated
the expression of nNOS and NF-κB in the rat brain. These results
suggested that EA may cure brain damage by downregulating the
NF-κB/nNOS pathway and that this process is associated with the
H2S/CBS pathway. However, the present study did not
investigate the detailed regulatory association between the
NF-κB/nNOS and H2S/CBS pathways. Future studies are
required to examine this regulatory effect.
In conclusion, the present study revealed that EA
can alleviate HIBD in rats by downregulating the NO content of
cortex cells. These results provide a significant reference for the
prevention and treatment of HIBD using the EA technique and also
describe a novel protective mechanism.
Acknowledgements
This study was sponsored and supported by the
International Science and Technology Cooperation Foundation of the
Ministry of Science and Technology of China (grant no.
2008DFA31850), the International Cooperation of Science and
Technique Foundation of Beijing (grant no. 2007G05) and the Beijing
Chinese Medicine Projects (grant no. JJ2005-17).
References
|
1
|
Kajimura M, Nakanishi T, Takenouchi T, et
al: Gas biology: Tiny molecules controlling metabolic systems.
Respir Physiol Neurobiol. 184:139–148. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu Y, Zou LP, Du JB and Wong V:
Electro-acupuncture protects against hypoxic-ischemic brain-damaged
immature rat via hydrogen sulfide as a possible mediator. Neurosci
Lett. 485:74–78. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
He J: Analysis on therapeutic effect of
acupuncture combined with hyperbaric oxygenation on delayed
encephalopathy in the patient of acute carbon monoxide poisoning.
Zhongguo Zhen Jiu. 28:30–32. 2008.(In Chinese). PubMed/NCBI
|
|
4
|
Ha Y, Kim M, Nah J, Suh M and Lee Y:
Measurements of location-dependent nitric oxide levels on skin
surface in relation to acupuncture point. Evid Based Complement
Alternat Med. 2012:7814602012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ma SX, Li XY, Sakurai T and Pandjaitan M:
Evidence of enhanced non-enzymatic generation of nitric oxide on
the skin surface of acupuncture points: An innovative approach in
humans. Nitric Oxide. 17:60–68. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ignarro LJ, Buga GM, Wood KS, Byrns RE and
Chaudhuri G: Endothelium-derived relaxing factor produced and
released from artery and vein is nitric oxide. Proc Natl Acad Sci
USA. 84:9265–9269. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Palmer RM, Ashton DS and Moncada S:
Vascular endothelial cells synthesize nitric oxide from l-arginine.
Nature. 333:664–666. 1988. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schmidl D, Boltz A, Kaya S, et al: Role of
nitric oxide in optic nerve head blood flow regulation during
isometric exercise in healthy humans. Invest Ophthalmol Vis Sci.
54:1964–1970. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Guarini G, Ohanyan VA, Kmetz JG, et al:
Disruption of trpv1-mediated coupling of coronary blood flow to
cardiac metabolism in diabetic mice: Role of nitric oxide and bk
channels. Am J Physiol Heart Circ Physiol. 303:H216–H223. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Phattanarudee S, Towiwat P, Maher TJ and
Ally A: Effects of medullary administration of a nitric oxide
precursor on cardiovascular responses and neurotransmission during
static exercise following ischemic stroke. Can J Physiol Pharmacol.
91:510–520. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ito E, Matsuo R and Okada R: Involvement
of nitric oxide in memory formation in microbrains. Neurosci Lett.
541:1–3. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Martin LJ, Adams NA, Pan Y, Price A and
Wong M: The mitochondrial permeability transition pore regulates
nitric oxide-mediated apoptosis of neurons induced by target
deprivation. J Neurosci. 31:359–370. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tsuji M, Higuchi Y, Shiraishi K, Kume T,
Akaike A and Hattori H: Protective effect of aminoguanidine on
hypoxic-ischemic brain damage and temporal profile of brain nitric
oxide in neonatal rat. Pediatr Res. 47:79–83. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fabian RH, Perez-Polo JR and Kent TA:
Perivascular nitric oxide and superoxide in neonatal cerebral
hypoxia-ischemia. Am J Physiol Heart Circ Physiol. 295:H1809–H1814.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu Y, Zou LP and Du JB: Nitric
oxide-mediated neuronal functional recovery in hypoxic-ischemic
brain damaged rats subjected to electrical stimulation. Brain Res.
1383:324–328. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gong QH, Shi XR, Hong ZY, Pan LL, Liu XH
and Zhu YZ: A new hope for neurodegeneration: Possible role of
hydrogen sulfide. J Alzheimers Dis. 24:173–182. 2011.PubMed/NCBI
|
|
17
|
Abe K and Kimura H: The possible role of
hydrogen sulfide as an endogenous neuromodulator. J Neurosci.
16:1066–1071. 1996.PubMed/NCBI
|
|
18
|
Edwards G, Félétou M and Weston AH:
Hydrogen sulfide as an endothelium-derived hyperpolarizing factor
in rodent mesenteric arteries. Circ Res. 110:e13–e14. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mustafa AK, Sikka G, Gazi SK, et al:
Hydrogen sulfide as endothelium-derived hyperpolarizing factor
sulfhydrates potassium channels. Circ Res. 109:1259–1268. 2012.
View Article : Google Scholar
|
|
20
|
Liu C, Gu X and Zhu YZ: Synthesis and
biological evaluation of novel leonurine-sprc conjugate as
cardioprotective agents. Bioorg Med Chem Lett. 20:6942–6946. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hu LF, Wong PT, Moore PK and Bian JS:
Hydrogen sulfide attenuates lipopolysaccharide-induced inflammation
by inhibition of p38 mitogen-activated protein kinase in microglia.
J Neurochem. 100:1121–1128. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kim KM, Pae HO, Zhung M, et al:
Involvement of anti-inflammatory heme oxygenase-1 in the inhibitory
effect of curcumin on the expression of pro-inflammatory inducible
nitric oxide synthase in raw264.7 macrophages. Biomed Pharmacother.
62:630–636. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yanagimoto T, Toyota T, Matsuki N, Makino
Y, Uchiyama S and Ohwada T: Transnitrosation of thiols from
aliphatic n-nitrosamines: S-nitrosation and indirect generation of
nitric oxide. J Am Chem Soc. 129:736–737. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kovacs I and Lindermayr C: Nitric
oxide-based protein modification: Formation and site-specificity of
protein s-nitrosylation. Front Plant Sci. 4:1372013.PubMed/NCBI
|
|
25
|
Avolio A and Grassi G: Haemoglobin: Subtle
scavenger of nitric oxide and modulator of retinal blood flow. J
Hypertens. 31:661–662. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Badn W and Siesjö P: The dual role of
nitric oxide in glioma. Curr Pharm Des. 16:428–430. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang R, Huang ZN and Cheng JS:
Anticonvulsion effect of acupuncture might be related to the
decrease of neuronal and inducible nitric oxide synthases. Acupunct
Electrother Res. 25:137–143. 2000.
|
|
28
|
Jou NT and Ma SX: Responses of nitric
oxide-cgmp release in acupuncture point to electroacupuncture in
human skin in vivo using dermal microdialysis. Microcirculation.
16:434–443. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang XY, Li XJ, Li DL, Wang CR and Guo XP:
Progesterone exerts neuroprotective effect on hypoxic-ischemic
encephalopathy-induced brain damage via inhibition expression of
inducible nitric oxide synthase and nitric oxide production.
Zhongguo Ying Yong Sheng Li Xue Za Zhi. 28:253–254. 2622012.(In
Chinese).
|