Introduction
Pulmonary fibrosis is a progressive lung disease
characterized by critical injury of functional alveolar epithelial
cells and the excessive production of extracellular matrix proteins
(1–3). A number of novel therapeutic
approaches are emerging, following significant progress in the
understanding of the molecular events associated with fibrosis,
which involve cytokines (4–6),
growth factors (7) and biochemical
signaling pathways (6,8). However, there are currently no
effective treatments to reverse or delay the course of this disease
(9,10). The pathogenesis of pulmonary
fibrosis remains elusive; however, oxidative stress, hyperplasia,
denudation and apoptosis of type II alveolar epithelial cells (AEC
II) may be important early features in this condition (11,12).
There is increasing evidence that oxidative stress
is involved in the pathogenesis of pulmonary fibrosis. The
development of bleomycin (BLM)-induced pulmonary fibrosis is
dose-dependent and may occur due to an increase in the
concentration of reactive oxygen species (ROS) (13). Markers of oxidative stress have
been identified, demonstrating that aberrant antioxidant activity
may exacerbate pulmonary fibrosis in patients and animal models
(12,14). AEC II cells, which are known to be
vulnerable to oxidative stress, produce and secrete the surfactant
proteins (SPs), SP-A, SP-B, SP-C and SP-D (markers of alveolar
epithelial cell function) in order to maintain morphological
organization, biophysical functions, biochemical composition and
immune function in lung tissue (15).
Bone marrow mesenchymal stem cells (MSCs) are adult
stem cells that are present in the stroma of bone marrow and
certain other organs (16,17). Previous studies have shown that
MSCs have potential as regenerative therapeutics due to their
strong proliferative capacity and differentiation potential
(18,19). With the capacity for mesoderm-like
cell differentiation into, for example, adipocytes, osteoblasts and
chondrocytes, MSCs are able to repair damage in a number of tissues
(20). Stem cell therapy may prove
to be effective in treating lung disease. However, the mechanisms
underlying this effect have not yet been fully elucidated (21–23).
The present study hypothesized that systemically
transplanted exogenous MSCs migrate into damaged lung tissue and
develop functional characteristics of alveolar cells, thus slowing
the progression of pulmonary fibrosis. In addition, it was
postulated that this effect is associated with changes in oxidative
stress. To test these hypotheses, the effect of systemic MSC
transplantation on the lung tissue of a rat model of BLM-induced
pulmonary fibrosis was investigated at a number of time-points
(24). Reverse transcription
polymerase chain reaction (RT-PCR) and immunofluorescence staining
were used to detect male rat Y chromosome-specific sequences in the
recipient female rat lung tissue (25) and to assess the differentiation of
MSCs into alveolar cells. Experiments to assess superoxide
dismutase (SOD) activity and total antioxidative capability (T-AOC)
were also conducted.
Materials and methods
Animals
Pathogen-free Wistar rats were obtained from the
Animal Testing Center, The Second Affiliated Hospital of Harbin
Medical University, (Harbin, China). Their use was approved by the
Animal Care and Use Committee of Harbin Medical University. All
animals were kept in accordance with the National Institutes of
Health Guidelines on the Use of Laboratory Animals. Rats were fed
with a conventional laboratory diet, with ad libitum access to tap
water in an air-conditioned room at ~25°C.
Isolation, identification and labelling
of MSCs
MSCs from a four-week-old clean-grade male Wistar
rat were plated into tissue culture flasks. Adherent cells were
allowed to grow to ~75% confluency, trypsinized and reseeded at a
density of 105 cells/cm2 (26). This procedure was performed for
four passages. When the 5th generation of cells reached 50–60%
confluency, 5-bromo-2-deoxyuridine (BrdU; Sigma-Aldrich, St. Louis,
MO, USA) was added to obtain a final concentration of 10 μmol/l.
These cells were further incubated for 48 h (27). Following digestion and
centrifugation at 1,000 × g for 5 min, cells were washed in a flow
cytometry buffer (Becton Dickinson, San Jose, CA, USA) and
incubated for 30 min with fluorescein-5-isothiocyanate
(FITC)-conjugated anti-rat monoclonal antibodies, including
anti-CD44, anti-CD45 and anti-CD90 (1:100, BioLegend, Inc., San
Diego, CA, USA). Washed cells were pelleted and resuspended, and
fixed in 1% paraformaldehyde for 15 min. Non-specific fluorescence
was determined using equal aliquots of the cell preparation
incubated with anti-mouse monoclonal antibodies (1:4,000,
BioLegend, Inc.). Data were acquired and analyzed on FACSCalibur
with CellQuest software (Becton Dickinson) (28). Fluorescein isothiocyanate
(FITC)-conjugated anti-rat CD44, CD45 and CD90 antibodies were
obtained from BioLegend, Inc. (San Diego, CA, USA). A cell
suspension with a live cell density of 2.5×106 cells/ml
was prepared with serum-free Dulbecco’s modified Eagle’s medium
(DMEM) for transplantation (29).
Flasks, trypsin and DMEM were obtained from Gibco Life Technologies
(Carlsbad, CA, USA). Paraformaldehyde, chloral hydrate, NaCl,
H2O2, PBS, phenol, chloroform and agarose
were all obtained from Sigma-Aldrich (St. Louis, MO, USA).
Establishment of a rat pulmonary fibrosis
model and delivery of MSC transplantation
A total of 48 clean-grade female Wistar rats
(weight, 200–240 g) were randomly divided into four groups: BLM
control group; immediate MSC injection group (BLM with
administration of MSCs at day 0); 7-day MSC injection group (BLM
with administration of MSCs at day 7); and negative control group
(12). Rats in the BLM group were
given a slow intratracheal injection of BLM (Nippon Kayaku Co.,
Ltd., Tokyo, Japan) at a dose of 5.0 mg/kg following
intraperitoneal anesthesia with 10% chloral hydrate (0.03 ml/kg).
Following the injection, rats were held upright and rotated in
order to distribute the drug as evenly as possible. In the
immediate MSC injection group and the 7-day MSC injection group,
rats were injected with 1 ml BrdU-labeled MSCs from a male rat
(containing 2.5×106 cells) into the tail vein, at days 0
and 7 following BLM administration, respectively. The negative
control rats received an intratracheal injection of 0.9% NaCl. On
days 7, 14 and 28, four rats from each group were sacrificed with
10% chloral hydrate (0.3 ml/100 g) by intraperitoneal
injection.
Histopathological analysis
The lung tissue was fixed in 4% neutral-buffered
formaldehyde for paraffin embedding, cut into 4-μm sections.
Paraffin sections were de-waxed twice with dimethyl benzene at 37°C
for 15 min, then soaked in 95, 70 and 30% ethanol, and distilled
water for 2 min each, and washed twice in water (37°C) for 45 sec.
Each section was stained with hematoxylin and eosin (H&E) and
Masson trichrome (Jiancheng Co., Ltd., Nanjing, China). The
remaining reagents were obtained from Beyotime Institute of
Biotechnology (Haimen, China). Standard scoring using the Szapiel
score was implemented for the semiquantitative assessment of the
severity of alveolitis and pulmonary fibrosis as described
previously (30). All histological
specimens were coded and interpreted in a blinded fashion. Three
sections were selected, and ten visual fields from each section
were observed. Images were evaluated by two assessors, and were
captured by an Optical Microscope (Olympus CX41, Olympus, Tokyo,
Japan).
Determination of hydroxyproline
content
The collagen content in the lung tissue was measured
using the hydroxyproline (Hyp) assay as described previously
(31). Hyp content was measured
using a Hyp detection kit (Jiancheng Biotechnology Institute,
Nanjing, China) according to the manufacturer’s instructions.
Immunofluorescence staining
Following sacrificing the rats, part of the right
lower lobes and part of the left lungs were removed, embedded in
polyvinyl alcohol and stored in liquid nitrogen. Serial sections of
~6 μm were prepared using a cryostat. The frozen sections were
treated with 3% H2O2 in order to block
endogenous peroxidase. Following DNA denaturation, 0.1% trypsin
pre-treatment was added. Normal goat serum (1:20; Sigma-Aldrich)
was used for blocking for 20 min. Subsequently, mouse monoclonal
anti-BrdU primary antibody (BU33, 1:100, Sigma-Aldrich) with or
without rabbit anti-SP-C polyclonal antibody (FL-197, 1:200, Santa
Cruz Biotechnology, Inc., Dallas, TX, USA) was added, tissue
sections were incubated overnight in a 4°C humidity chamber and
washed once with phosphate-buffered saline (PBS). Non-specific
serum, in place of the primary antibody, was used as the negative
control. An FITC-labeled goat anti-mouse secondary antibody (1:100,
Santa Cruz Biotechnology, Inc.) and a tetramethylrhodamine-labeled
goat anti-rabbit immunoglobulin G secondary antibody (1:100, Santa
Cruz Biotechnology, Inc.) were added. Tissue sections were then
incubated at 37°C in the dark for 1 h and washed once with PBS.
DAPI (Beyotime Institute of Biotechnology), 1:1,000 was added in
order to restain the nucleus and slides were then mounted. The
staining results were observed and radiographed using fluorescence
microscopy (Nikon MF30 LED, Nikon Corporation, Tokyo, Japan). Four
to six fields of view for each specimen were randomly selected and
observed under the microscope. The procedures for SP-C
immunofluorescence staining of fresh bone marrow aspirates and
cultured MSCs of the fifth generation were identical to those
described for assessment of the lung tissue, with the exception of
the use of 4% paraformaldehyde as the fixative solution.
PCR detection of sex-determining region Y
(SRY) gene
Data from a previous study (32) was used to identify and synthesize
the rat SRY gene product. The following primers were used: Forward,
5′-TTTAGTGTTCAGCCTACAGCC-3′ and reverse,
5′-GCACTTTAACCCTCGATGAGG-3′ for SRY, and forward,
5′-CACGATGGAGGGGCCGGACTCATC-3′ and reverse,
5′-TAAAGACCTCTATGCCAACACAGT-3′ for β-actin. Primers were
synthesized by Chaoshi Biotech Co. (Shanghai, China).” The lengths
of the amplified fragments for SRY and β-actin were 322 and 240 bp,
respectively. DNA from the lung tissue of female rats was extracted
using phenol and chloroform. PCR conditions were as follows:
Initial denaturation at 94°C for 5 min, denaturation at 94°C for 30
sec, annealing at 52°C for 30 sec, extension at 72°C for 30 sec for
30 cycles and full extension at 72°C for 5 min. Gel electrophoresis
in 2% agarose was conducted in order to analyze the PCR products.
An image scanner was used to radiograph and analyze the average
grayscale of target bands (TannonGIS gel imaging and analysis
system; Biotanon Biotech Co., Shanghai, China). The results were
compared with those of the female negative control and male
positive control groups.
RT-PCR analysis of SP-C and aquaporin 5
(AQP-5) mRNA expression
RNA from 100 mg lung tissue, (1–2)×107 fresh bone marrow
aspirates and 5×106 fifth generation MSCs was extracted
using a TRIzol® total RNA extraction kit (Invitrogen
Life Technologies, Carlsbad, CA, USA). Primers were synthesized by
Shanghai Chaoshi Biotech Co. (Shanghai, China). The following
primers were used: Forward, 5′-GTCCTTGTCGTCGTGGTGAT-3′ and reverse,
5′-AGGTAGCGATGGTGTCTGTGT-3′ for SP-C, and forward,
5′-AACCCAGCCCGATCTTTC-3′ and reverse, 5′-GGAAGAGCAGGTAGAAATAGAGG-3′
for AQP-5. The lengths of the amplified fragments for SP-C and
AQP-5 were 154 bp and 424 bp, respectively. RT-PCR was conducted
using a one-step reaction system kit (Takara Biotechnology Co.,
Ltd., Dalian, China), which consisted of complementary DNA (cDNA)
synthesis and pre-denaturation using the following conditions:
Reverse-transcription at 42°C for 30 min and 99°C for 5 min for one
cycle, PCR amplification with denaturation at 94°C for 30 sec,
annealing at 52°C for 30 sec, extension at 72°C for 30 sec for 30
cycles and final extension at 72°C for 5 min. Gel electrophoresis
and semi-quantitative mRNA detection were conducted using 2%
agarose gel electrophoresis of 10 μl of each of the amplified SP-C,
AQP-5 and β-actin products, followed by radiography using an image
scanner (WD-9413A, Liuyi Instrument Factory, Beijing, China).
Measurement of SOD activity and
T-AOC
SOD and T-AOC kits were purchased from Nanjing
Jiancheng Bioengineering Institute (Ninjing, China). BLM-injected
rats were treated with immediate injection of MSCs or with no
treatment and lung tissues were collected and homogenized. The lung
homogenates were centrifuged for 30 min at 3,000 × g at 4°C. The
content of protein in the supernatant was determined using a
Bradford protein assay kit (Beyotime Institute of Biotechnology).
The procedures used to quantify SOD activity and T-AOC were
conducted according to the manufacturer’s instructions on a plate
reader (Tecan Infinite 200, Eastwin Life Science, Beijing, China).
The absorbance change at 520 and 550 nm was monitored (33,34).
Data are expressed as the mean absorbance normalized to the
percentage of the vehicle control.
Statistical analysis
Experimental data are expressed as the mean ±
standard error of the mean. SPSS version 17.0 (SPSS, Inc., Chicago,
IL, USA) was used to calculate analysis of variance in order to
compare the data between groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
Identification and labelling of MSCs
In the present study, the labeling rate of MSCs
accounted 98% after MSCs were incubated with BrdU (10 μmol/) for 48
h. Furthermore, the analysis of the cell surface phenotype
indicated that the MSC population was positive for CD44 and CD90
but negative for CD45, which was consistent with the International
Society for Cellular Therapy position statement for the minimum
criteria for an MSC (28).
MSCs reduce alveolitis and pulmonary
fibrosis in the injured lung
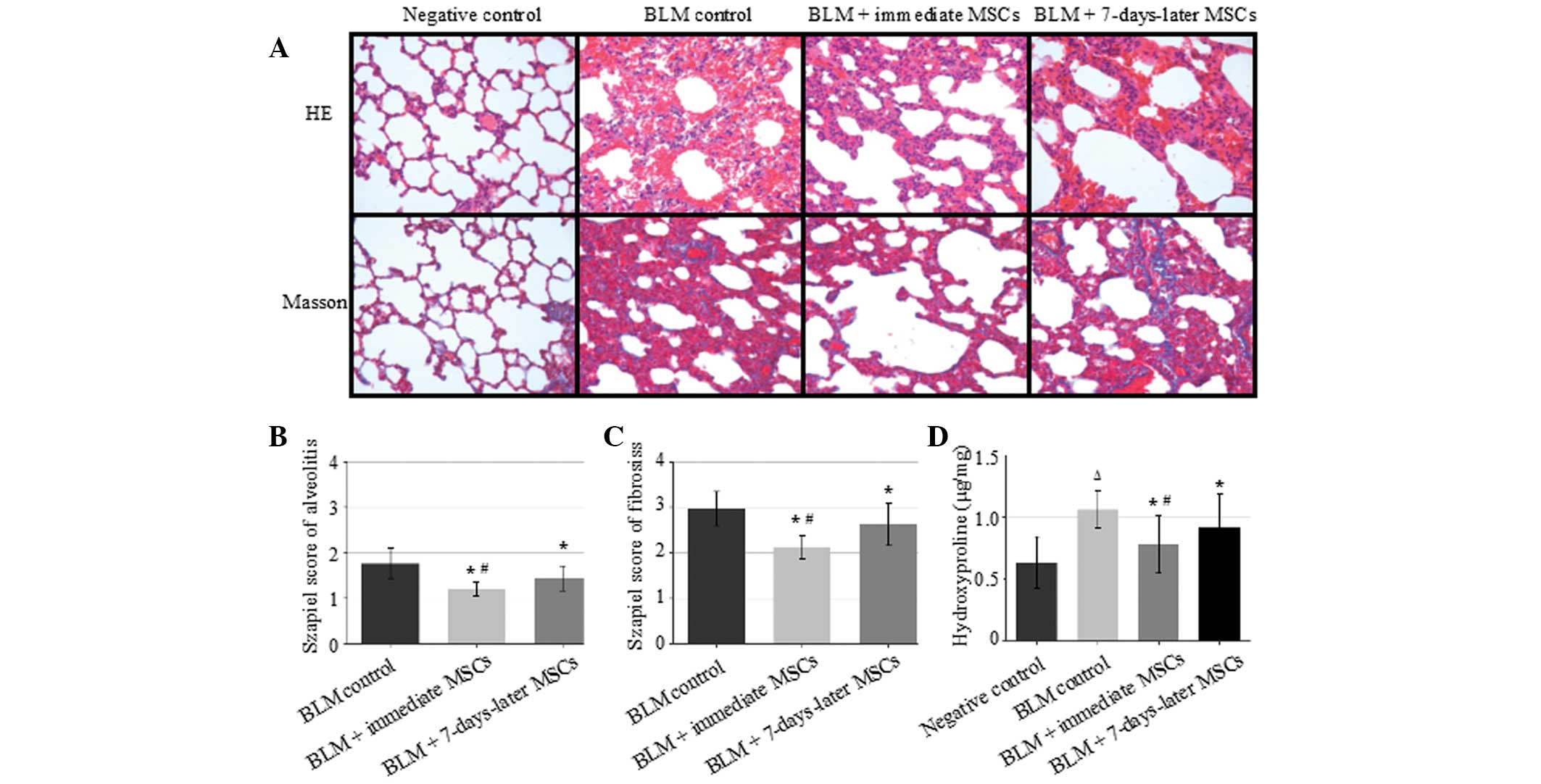
Rat lung sections from each group on day 28 are
shown in Fig. 1A. There was no
obvious lesion in the lungs of rats from the negative control
group. In the BLM control group, H&E staining showed that the
pulmonary alveolus cavity was markedly decreased in size, the
alveolar wall was thickened, and the mesenchyme was expanded, with
increased numbers of fibroblasts and macrophages compared with the
negative control group. Masson trichrome staining showed that
collagen was increased in the lung interstitium, as well as around
blood vessels and bronchi, when compared with the negative control
group. In the immediate MSCs group, the majority of alveoli were
intact. The thickness of the alveolar wall, the number of
fibroblasts and the quantity of collagen in the lung interstitium
were significantly reduced compared with the those in the BLM
control group. The Szapiel score for alveolitis and pulmonary
fibrosis on day 28 following BLM administration were significantly
reduced by transplantation of MSCs at day 0 and day 7, compared
with those in the BLM control group (P<0.05). Administration of
MSCs at day 0 further decreased the Szapiel score of alveolitis and
pulmonary fibrosis compared with treatment at day 7 (P<0.05;
Fig. 1B and C). Furthermore,
according to the Hyp content on day 28, there was a significant
difference in the lung collagen content between the immediate MSCs
group and the BLM control group (P<0.05; Fig. 1D).
Transplantation of the exogenous
MSCs
To detect whether BrdU-labelled exogenous MSCs
migrated to damaged lung tissue, stand-alone BrdU staining was
used. BrdU-labelled MSCs were identified in the frozen sections of
lung tissue from rats sacrificed on days 7, 14 and 28. These cells
showed green fluorescence. MSCs were predominantly distributed as
single cells. Clusters of two to three cells were also present. The
shapes of the MSCs were predominantly oval or cubic, and a number
were spindle-shaped at one end. The nuclei were round or oval in
shape and were relatively large. Morphologically, the MSC
distribution was consistent with the usual location and size of AEC
II. At the earlier time-points, MSCs were primarily distributed
around the interstitial lung and pulmonary blood vessels. However,
at day 28, MSCs were aggregated and were principally distributed in
the alveolar walls, with a number of MSCs distributed in the
collapsed alveolar spaces, in which the interstitial changes were
greatest. In the 7-day MSCs injection group, a small number of
BrdU-positive MSCs were observed. However, compared with the
immediate MSCs injection group, the number was significantly
smaller. No BrdU-positive cells were found in the negative control
group (Fig. 2A and B).
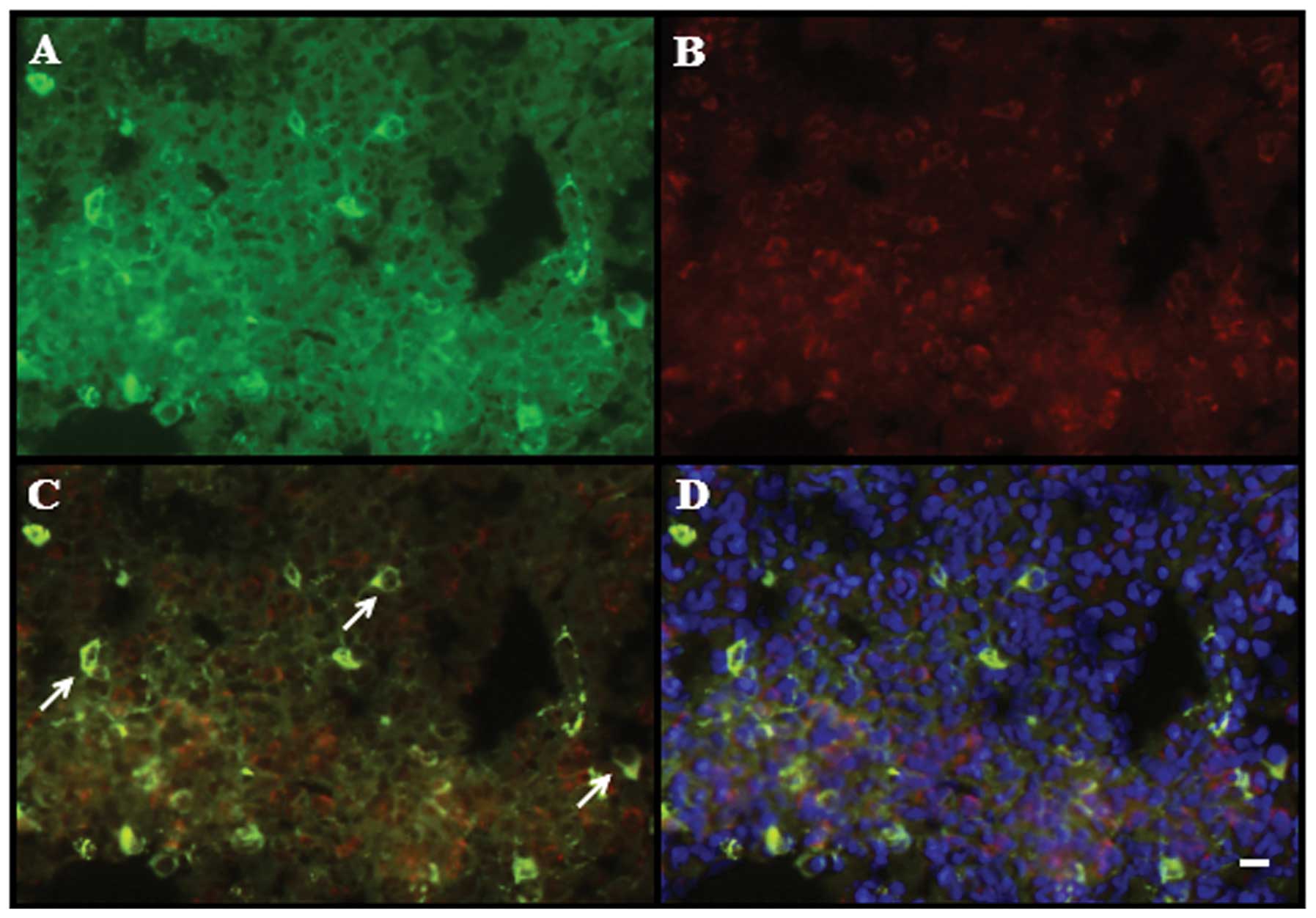
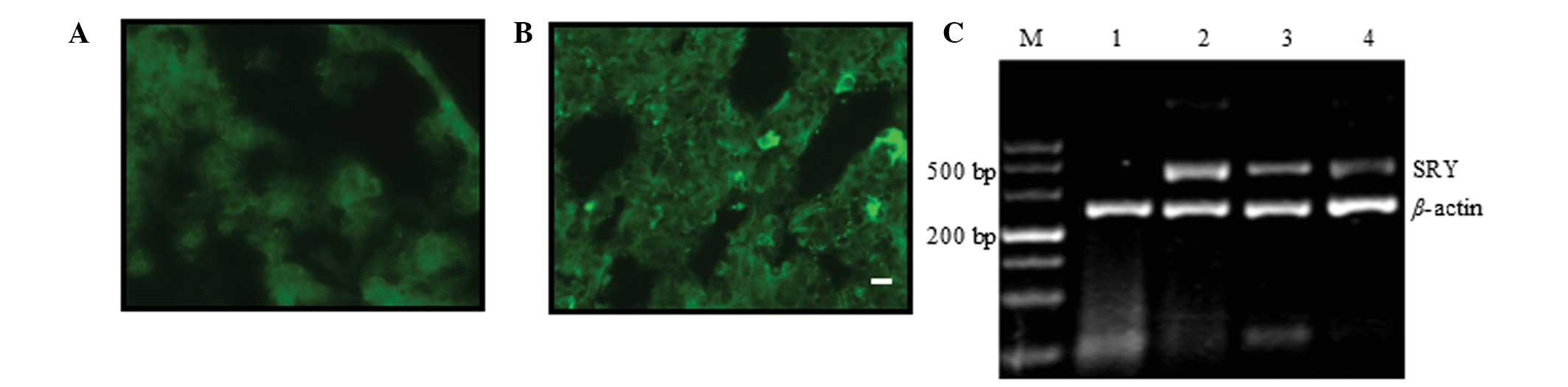 | Figure 2MSCs from male rats were transplanted
into BLM-induced female rat models of pulmonary fibrosis. (A and B)
Donor 5-bromo-2-deoxyuridine-labelled MSCs (green) were observed in
recipient lung tissue 28 days following injection of MSCs. (A)
Negative control group and (B) immediate MSCs injection group.
Scale bars, 10 μm; original magnification, ×400. (C) Polymerase
chain reaction analysis of the SRY gene in rat lung tissue. Lane M,
DL500 DNA marker; lane 1, control female rats; lane 2, control male
rats; lane 3, immediate MSCs injection group; and lane 4, 7-day
MSCs injection group. MSCs, mesenchymal stem cells; BLM, bleomycin;
SRY, sex-determining region Y gene. |
To further verify that the transplanted MSCs had
migrated into the lung tissue, PCR was used to detect the presence
of the male SRY gene on the Y chromosome in lung tissue in which
positive stand-alone BrdU staining was observed. The length of the
SRY gene fragment used for detection was 322 bp. The length of the
β-actin fragment, used as an internal control, was 240 bp.
Following PCR amplification of the DNA that was extracted from rat
lung tissue, electrophoresis demonstrated only a single 240 bp
internal control band in the female negative control rats, but an
additional 322 bp gene band in the male positive control rats. The
322 bp bands were also detected in the immediate MSCs injection
group and in the 7-day MSCs injection group, which each exhibited
identical electrophoresis patterns to those of the male positive
control male rats. However, the levels of SRY gene expression, as
measured by the average greyscale of the target bands, was
significantly lower in the 7-day MSC injection group (0.04) than in
the immediate MSC injection group (0.23; P<0.05; Fig. 2C).
Dual immunofluorescence staining of lung
tissue
To determine whether MSCs that had migrated into the
lung tissue differentiated into AEC II and exhibited AEC II
function, lung tissue was treated with dual immunofluorescence
staining for SP-C and BrdU. As shown in Fig. 3, green fluorescence staining
indicated BrdU-positive cells, red fluorescence in the cytoplasm
indicated SP-C-positive cells and yellow fluorescence indicated
BrdU- and SP-C-positive cells. Dual BrdU- and SP-C-positive cells
were observed in frozen sections of lung tissue from the immediate
MSC injection group on days 7, 14 and 28. The dual positive cells
were predominantly distributed in the alveolar walls and a number
were observed in the collapsed alveolar spaces, in which the
interstitial changes were greatest. In addition, all the
BrdU-positive cells in the lung tissue that were morphologically
similar to AEC II, but different from AEC I and fibroblast cells,
were able to differentiate into AEC II. A small number of dual
positive cells were also be found in the rat lung tissue from the
7-day MSC injection group.
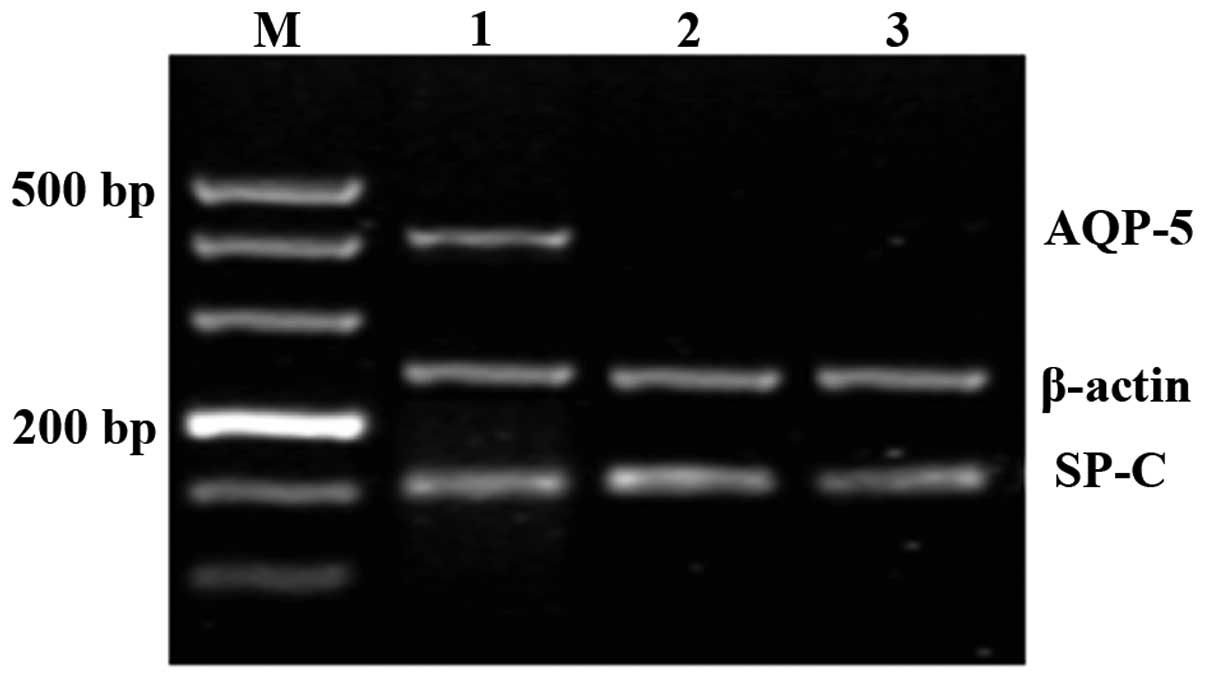
SP-C and AQP-5 mRNA expression in
different tissues and cells
It was shown that exogenous MSCs differentiated into
AEC II in the lung tissue of recipients. In order to investigate at
what point this differentiation occurred, the mRNA levels of SP-C,
a specific marker for AEC II, and AQP-5, a specific marker for AEC
I, were examined. As shown in Fig.
4, mRNA from each marker was expressed in normal lung tissue.
However, in fresh bone marrow, aspirates and the fifth generation
of cultured MSCs, only the 154 bp SP-C mRNA was expressed, as
detected by RT-PCR. In order to further validate the SP-C
expression in bone marrow stem cells, SP-C immunofluorescence
staining of fresh bone marrow aspirates and the fifth generation of
cultured MSCs was performed, and no positive cells were observed in
these samples.
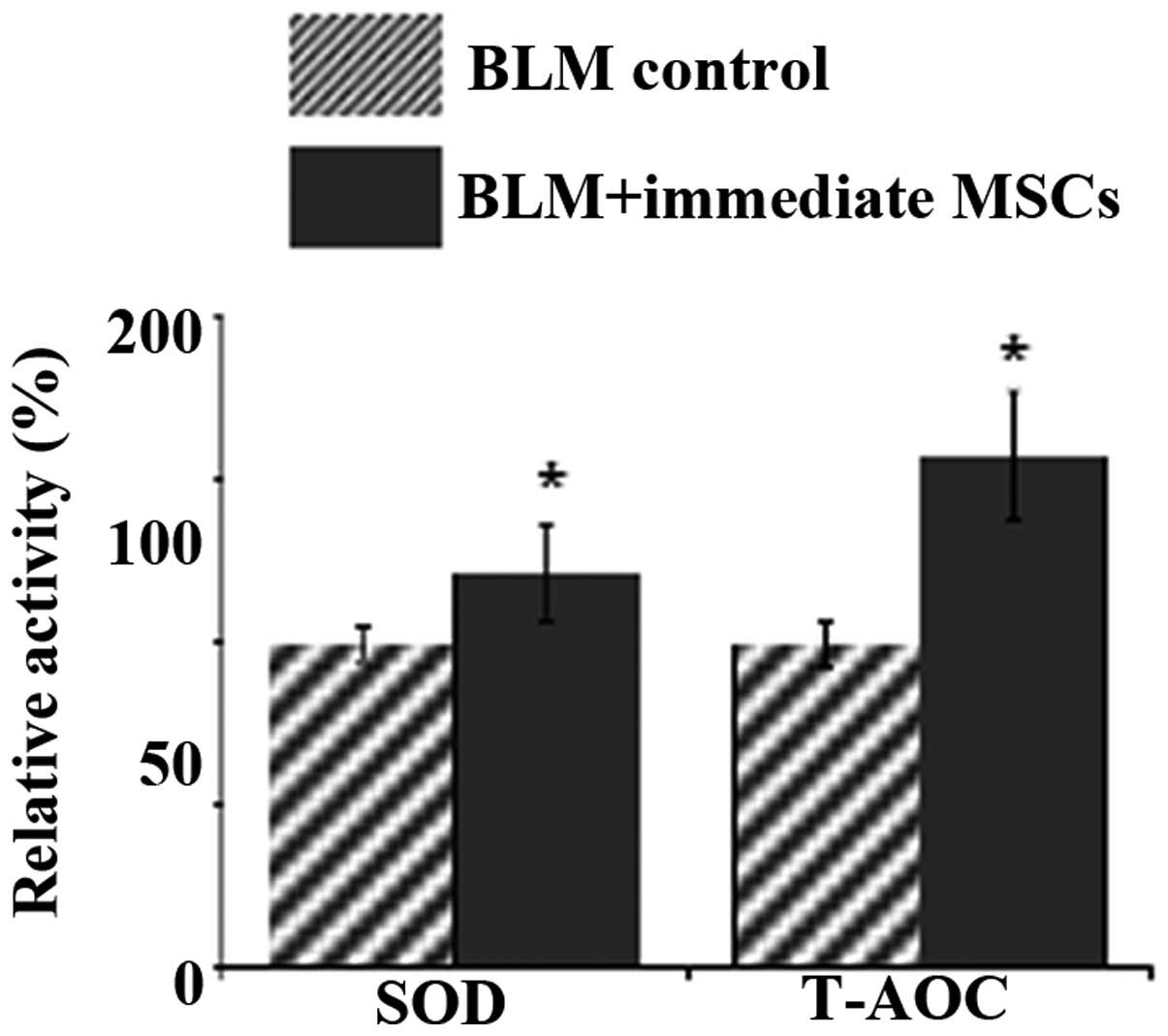
Oxidative stress detection
It was hypothesized that the alleviating effect of
pulmonary fibrosis following MSC injection may be partly due to the
alleviation of the oxidative damage by AEC II, which MSCs
differentiate into. SOD is an important component of the
antioxidant enzymatic defense system and converts the superoxide
radical to H2O2. T-AOC reflects the overall
cellular endogenous antioxidative capability, including enzymatic
as well as non-enzymatic antioxidants (35). SOD activity and T-AOC in the lung
tissue from the immediate MSC injection group were significantly
increased compared to those in the BLM control group (P<0.01,
Fig. 5). Thus, AEC II generation
as a result of injection of MSCs, significantly attenuated the
oxidative stress in pulmonary fibrosis.
Discussion
In the present study, bone marrow-derived MSCs were
shown to differentiate into AEC II when transplanted into rats
following BLM administration. In addition, their behavior was
altered by the microenvironment of injury.
BLM is one of the most extensively studied and
reproducible agents used to induce lung fibrosis in rats. When BLM
is given into the airway, it causes lung epithelial injury,
followed by an inflammatory response, which is followed by the
development of lung fibrosis that eventually resolves (36). Purified bone marrow cells that
proliferated in vitro exhibited morphological
characteristics of MSCs, as indicated by a
CD45−CD44+CD90+ cell surface
phenotype. In the present study, when transplanted into Wistar rats
with BLM-induced lung injury, MSCs were shown to promote the repair
process following lung injury and fibrosis. The lung injury Szapiel
score for inflammation and fibrosis on day 28 following BLM
administration was significantly reduced by the transplantation of
MSCs. Moreover, according to the Hyp content on day 28, there was a
significant difference in the collagen content of the lungs of rats
treated with MSCs compared with that in the BLM control group.
A characteristic function of mature AEC II is the
synthesis and secretion of pulmonary SPs, including SP-A, SP-B,
SP-C and SP-D. Notably, SP-C is exclusively expressed in AEC II and
therefore may be used for the identification of these cells
(37,38). AQP-5 is a specific marker for AEC
I, although in the present study, dual AQP-5 and BrdU
immunofluorescence staining was not performed as it was possible to
differentiate AEC I from AEC II (39). AEC II have been postulated to act
as stem cells in lung tissue (40), giving rise to AEC I in response to
injury, and are known targets of apoptotic signals induced by lung
lesions (39,41,42).
By replacing AEC II, MSCs may limit the effects of lung injury, and
their differentiation into AEC II may partially restore the stem
cell pool.
The results of the present study demonstrated that
when MSCs were administered immediately following the establishment
of the BLM-induced animal model of fibrosis, exogenous MSCs were
detected in lung tissues from rats killed on days 7, 14 and 28.
Furthermore, the number of MSCs in the lung tissue increased over
time. The distinct cellular localization, morphology and molecular
features of the engrafted cells indicated that they expressed the
type II pneumocyte phenotype, and that these cells had the SP-C
secreting function characteristic of AEC II. However, when MSCs
were administered at 7 days following the establishment of the
animal model, only a small number of dual positive cells were
detected in the lung tissue from rats killed on day 28. The PCR
results confirmed that male MSCs migrated into the lung tissue of
female recipients in varying quantities in the immediate and 7-day
MSC groups.
Under normal conditions, MSCs exist in G0
or G1 phase of the cell cycle and are thus in a
relatively static state. However, upon induction by certain
factors, MSCs may be stimulated to differentiate and migrate to
damaged tissues. Ortiz et al (22) showed that BLM-induced lung injury
promoted the migration of transplanted MSCs into the lungs as well
as the differentiation of MSCs into AEC II, resulting in repair of
lung damage and a reduction in pulmonary fibrosis. However, a study
by Kotton et al (21)
produced different results. Bone marrow stem cells that had
attached to petri dishes were cultured for one week and then
transplanted into rats with BLM-induced lung injury. These cells
were observed to migrate to the lungs and differentiate into AEC I.
The morphological characteristics and molecular biological
phenotypes of endogenous AEC I were also detected in fresh bone
marrow aspirates and bone marrow cells that had been cultured for
one week. Furthermore, cultured bone marrow-derived cells exhibited
features of lung cell differentiation prior to engraftment.
Therefore, results regarding the conditions of MSC migration and
differentiation may markedly vary by experimental design. The
present study showed that the morphological characteristics and the
molecular biological phenotypes of endogenous AEC II were
detectable in all BrdU-labelled MSCs following migration into
recipient lung tissue. In fresh bone marrow aspirates and fifth
generation of cultured MSCs, RT-PCR detected mRNA expression of the
AEC II-specific marker SP-C, but not of the AEC I specific marker
AQP-5. SP-C immunofluorescence staining did not detect
SP-C-positive cells. The experimental design was considerably
different from that of Ortiz et al (22), but nevertheless yielded identical
results. By contrast, the experimental protocol was similar to that
used by Kotton et al but yielded contradictory results.
There are a number of possible reasons for these discrepancies.
Kotton et al used bone marrow cells for transplantation.
These had not been subcultured and purified, and were therefore not
MSCs. It is currently understood that bone marrow cells contain
three cell groups: Hematopoietic stem cells, MSCs and endothelial
precursor cells (43). The
morphological characteristics and molecular biological phenotypes
of AEC I that were detected by Kotton et al were likely to
have been produced by the endothelial precursor cells. In addition,
AEC II are functionally equivalent to the stem cells of the lungs,
in that they can reproduce to produce AEC II progeny or rapidly
differentiate into AEC I. The AEC I detected by Kotton et al
are likely to originate from differentiated AEC II.
Through differentiation into AEC II, MSCs may
partially restore the lung stem cell pool. The proliferation of
lung stem cells may also explain how the low number of MSCs that
migrated to the lungs had such a marked effect on the health status
of rats with BLM-induced lung damage. The results of the present
study suggested that exogenous MSCs migrated into injured lungs and
differentiated into AEC II. This differentiation potential was
present prior to transplantation, but the cells required a local
microenvironment in order to complete differentiation and to
develop the morphological characteristics and function of AEC II.
The MSC transplantation rate was higher when MSCs were injected in
the early stages of lung injury. This increased efficacy may be
associated with lung injury, and the chemokines produced by this
injury, which in turn promote MSC migration into the lungs and the
subsequent directional differentiation (44). MSC differentiation is key to the
therapeutic role of these cells in pulmonary fibrosis.
Oxidative stress has been implicated as a possible
molecular mechanism underlying fibrosis in a variety of organs,
including the lung. Accumulating evidence also indicates that BLM
induces lung injury as a result of its ability to generate ROS,
including superoxide and hydroxy radicals (45). Accordingly, in the present study,
the levels of markers of oxidative stress were investigated. It was
shown that the antioxidant enzyme activities of SOD and T-AOC were
significantly decreased in the immediate MSC injection group
compared with those in the BLM control group. These results
supported the hypothesis that the AEC II generation following
injection of MSCs may responsible for the alleviation of pulmonary
fibrosis, at least in part due blocking oxidative damage.
In the present study, MSCs were shown to
differentiate into AEC II, suggesting that differentiation may be
the primary mechanism through which MSCs exert a therapeutic
effect. MSCs possess this differentiation potential at the bone
marrow stage, but are only able to differentiate within a
microenvironment of injury. The stem cells repair damage caused by
BLM, at least in part, by blocking oxidative stress. Further
studies are required to investigate alternative and additional
mechanisms. In conclusion, MSC migration and differentiation may
provide an effective cell therapy for pulmonary fibrosis.
Acknowledgements
This study was supported by the Educational
Commission of Heilongjiang Province of China (grant no.
12541517).
References
|
1
|
Fernandez IE and Eickelberg O: New
cellular and molecular mechanisms of lung injury and fibrosis in
idiopathic pulmonary fibrosis. Lancet. 380:680–688. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Crestani B, Marchand-Adam S, Fabre A,
Dehoux M and Soler P: Mechanisms in pulmonary fibrosis. Rev Prat.
57:2222–2226. 2007.(In French).
|
|
3
|
Nuovo GJ, Hagood JS, Magro CM, et al: The
distribution of immunomodulatory cells in the lungs of patients
with idiopathic pulmonary fibrosis. Mod Pathol. 25:416–433. 2012.
View Article : Google Scholar :
|
|
4
|
Starosta V, Ratjen F, Rietschel E, Paul K
and Griese M: Anti-inflammatory cytokines in cystic fibrosis lung
disease. Eur Respir J. 28:581–587. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Simonian PL, Roark CL, Born WK, O’Brien RL
and Fontenot AP: Gammadelta T cells and Th17 cytokines in
hypersensitivity pneumonitis and lung fibrosis. Transl Res.
154:222–227. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tan HL, Regamey N, Brown S, et al: The
Th17 pathway in cystic fibrosis lung disease. Am J Respir Crit Care
Med. 184:252–258. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Okada M, Suzuki K, Matsumoto M, et al:
Effects of angiotensin on the expression of fibrosis-associated
cytokines, growth factors, and matrix proteins in human lung
fibroblasts. J Clin Pharm Ther. 34:288–299. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ruan W and Ying K: Abnormal expression of
IGF-binding proteins, an initiating event in idiopathic pulmonary
fibrosis? Pathol Res Pract. 206:537–543. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Datta A, Scotton CJ and Chambers RC: Novel
therapeutic approaches for pulmonary fibrosis. Br J Pharmacol.
163:141–172. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Maher TM: Idiopathic pulmonary fibrosis:
pathobiology of novel approaches to treatment. Clin Chest Med.
33:69–83. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sisson TH, Mendez M, Choi K, et al:
Targeted injury of type II alveolar epithelial cells induces
pulmonary fibrosis. Am J Respir Crit Care Med. 181:254–263. 2010.
View Article : Google Scholar :
|
|
12
|
Kliment CR and Oury TD: Oxidative stress,
extracellular matrix targets, and idiopathic pulmonary fibrosis.
Free Radic Biol Med. 49:707–717. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
El-Khouly D, El-Bakly WM, Awad AS,
El-Mesallamy HO and El-Demerdash E: Thymoquinone blocks lung injury
and fibrosis by attenuating bleomycin-induced oxidative stress and
activation of nuclear factor Kappa-B in rats. Toxicology.
302:106–113. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cheresh P, Kim SJ, Tulasiram S and Kamp
DW: Oxidative stress and pulmonary fibrosis. Biochim Biophys Acta.
1832.1028–1040. 2012.
|
|
15
|
Lu TH, Chen CH, Lee MJ, et al:
Methylmercury chloride induces alveolar type II epithelial cell
damage through an oxidative stress-related mitochondrial cell death
pathway. Toxicol Lett. 194:70–78. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shi C: Recent progress toward
understanding the physiological function of bone marrow mesenchymal
stem cells. Immunology. 136:133–138. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Charbord P: Bone marrow mesenchymal stem
cells: historical overview and concepts. Hum Gene Ther.
21:1045–1056. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Daba MH, El-Tahir KE, Al-Arifi MN and
Gubara OA: Drug-induced pulmonary fibrosis. Saudi Med J.
25:700–706. 2004.PubMed/NCBI
|
|
19
|
Kan I, Melamed E and Offen D: Integral
therapeutic potential of bone marrow mesenchymal stem cells. Curr
Drug Targets. 6:31–41. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jiang Y, Jahagirdar BN, Reinhardt RL, et
al: Pluripotency of mesenchymal stem cells derived from adult
marrow. Nature. 418:41–49. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kotton DN, Ma BY, Cardoso WV, et al: Bone
marrow-derived cells as progenitors of lung alveolar epithelium.
Development. 128:5181–5188. 2001.PubMed/NCBI
|
|
22
|
Ortiz LA, Gambelli F, McBride C, et al:
Mesenchymal stem cell engraftment in lung is enhanced in response
to bleomycin exposure and ameliorates its fibrotic effects. Proc
Natl Acad Sci USA. 100:8407–8411. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mora AL and Rojas M: Aging and lung injury
repair: a role for bone marrow derived mesenchymal stem cells. J
Cell Biochem. 105:641–647. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li HP, Li X, He GJ, Yi XH and Kaplan AP:
The influence of dexamethasone on the proliferation and apoptosis
of pulmonary inflammatory cells in bleomycin-induced pulmonary
fibrosis in rats. Respirology. 9:25–32. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Padilla-Nash HM, Barenboim-Stapleton L,
Difilippantonio MJ and Ried T: Spectral karyotyping analysis of
human and mouse chromosomes. Nat Protoc. 1:3129–3142. 2006.
View Article : Google Scholar
|
|
26
|
Zhao F, Zhang YF, Liu YG, et al:
Therapeutic effects of bone marrow-derived mesenchymal stem cells
engraftment on bleomycin-induced lung injury in rats. Transplant
Proc. 40:1700–1705. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Feng SW, Yao XL, Li Z, et al: In vitro
bromodeoxyuridine labeling of rat bone marrow-derived mesenchymal
stem cells. Di Yi Jun Yi Da Xue Xue Bao. 25:184–186. 2005.(In
Chinese). PubMed/NCBI
|
|
28
|
Dominici M, Le Blanc K, Mueller I, et al:
Minimal criteria for defining multipotent mesenchymal stromal
cells. The International Society for Cellular Therapy position
statement. Cytotherapy. 8:315–317. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li X, Zhang Y and Qi G: Evaluation of
isolation methods and culture conditions for rat bone marrow
mesenchymal stem cells. Cytotechnology. 65:323–334. 2013.
View Article : Google Scholar :
|
|
30
|
Szapiel SV, Elson NA, Fulmer JD,
Hunninghake GW and Crystal RG: Bleomycin induced interstitial
pulmonary disease in the nude, athymic mouse. Am Rev Respir Dis.
120:893–899. 1979.PubMed/NCBI
|
|
31
|
Tanaka K, Ishihara T, Azuma A, et al:
Therapeutic effect of lecithinized superoxide dismutase on
bleomycin-induced pulmonary fibrosis. Am J Physiol Lung Cell Mol
Physiol. 298:L348–L360. 2010. View Article : Google Scholar
|
|
32
|
Narahara M, Yamada A, Hamada-Kanazawa M,
Kawai Y and Miyake M: cDNA cloning of the Sry-related gene Sox6
from rat with tissue-specific expression. Biol Pharm Bull.
25:705–709. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu J, Wang X, Wang F, Teng L and Cao J:
Attenuation effects of heparin-superoxide dismutase conjugate on
bleomycin-induced lung fibrosis in vivo and radiation-induced
inflammatory cytokine expression in vitro. Biomed Pharmacother.
63:484–491. 2009. View Article : Google Scholar
|
|
34
|
Wei X, Liu H, Sun X, et al: Hydroxysafflor
yellow A protects rat brains against ischemia-reperfusion injury by
antioxidant action. Neurosci Lett. 386:58–62. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Qi HP, Wei SQ, Gao XC, et al:
Ursodeoxycholic acid prevents selenite-induced oxidative stress and
alleviates cataract formation: In vitro and in vivo studies. Mol
Vis. 18:151–160. 2012.PubMed/NCBI
|
|
36
|
Bowden DH: Unraveling pulmonary fibrosis:
the bleomycin model. Lab Invest. 50:487–488. 1984.PubMed/NCBI
|
|
37
|
Wang D, Haviland DL, Burns AR, Zsigmond E
and Wetsel RA: A pure population of lung alveolar epithelial type
II cells derived from human embryonic stem cells. Proc Natl Acad
Sci USA. 104:4449–4454. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang D, Morales JE, Calame DG, Alcorn JL
and Wetsel RA: Transplantation of human embryonic stem cell-derived
alveolar epithelial type II cells abrogates acute lung injury in
mice. Mol Ther. 18:625–634. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kapanci Y, Weibel ER, Kaplan HP and
Robinson FR: Pathogenesis and reversibility of the pulmonary
lesions of oxygen toxicity in monkeys. II. Ultrastructural and
morphometric studies. Lab Invest. 20:101–118. 1969.PubMed/NCBI
|
|
40
|
Castranova V, Rabovsky J, Tucker JH and
Miles PR: The alveolar type II epithelial cell: a multifunctional
pneumocyte. Toxicol Appl Pharmacol. 93:472–483. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Fehrenbach H: Alveolar epithelial type II
cell: defender of the alveolus revisited. Respir Res. 2:33–46.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Emura M, Ochiai A, Singh G, et al: In
vitro reconstitution of human respiratory epithelium. In Vitro Cell
Dev Biol Anim. 33:602–605. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jiang Y, Jahagirdar BN, Reinhardt RL, et
al: Pluripoteney of mesenehymal stem cells derived from adult
marrow. Nature. 418:41–49. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ablin RJ: The need for personalized
therapy and companion diagnostics in prostate cancer. Biomark Med.
5:281–283. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Caspary WJ, Lanzo DA and Niziak C: Effect
of deoxyribonucleic acid on the production of reduced oxygen by
bleomycin and iron. Biochemistry. 21:334–338. 1982. View Article : Google Scholar : PubMed/NCBI
|