Introduction
Dental pulp tissues have the ability to repair and
regenerate following injury. In the presence of an infection within
the dentin-pulp complex, oral microorganisms and their components
diffuse into the pulp through the dentinal tubules. This type of
stimulation induces the mixed population of pulp cells (1,2),
which include stem cells and odontoblast progenitor cells, to
differentiate and form a dentin-like mineralized matrix. This is
marked by an increase in the expression of hard-tissue-forming
proteins, including alkaline phosphatase (ALP), dentin
sialophosphoprotein (DSPP) and dentin matrix protein-1 (DMP-1)
(3,4).
Lipopolysaccharides (LPS) comprise a major molecular
component of the outer cell wall of Gram-negative bacteria and are
potent virulence factors that contribute to bacterially induced
pathology, including general sepsis (5), lung disease (6), periodontal disease (7) and pulpitis (8). In several experiments, LPS has been
used to form a model of inflammation and the protective
inflammatory reaction evoked by LPS is considered to induce signal
transduction through Toll-like receptor 4 (TLR4) (9,10).
In our previous study, the mRNA and protein expression of TLR4 was
observed in the cells of the odontoblast layer and pulp tissues
(11). TLRs, broadly distributed
pattern recognition receptors, are also involved in the
pathogenesis of chronic inflammatory diseases. LPS and
peptidoglycans, which are TLR4 and TLR agonists, have been observed
to induce the upregulation of osteogenesis-associated factors in
human aortic valve interstitial cells (hAVICs) (12,13).
To date, few studies have examined the effects of LPS or TLRs on
hDPC differentiation.
Furthermore, LPS is known to activate NF-κB,
predominantly through the TLR4 downstream pathway (14). Increased expression levels of
intercellular adhesion molecule-1 and vascular cell adhesion
molecule-1 are associated with the translocation of NF-κB from the
nucleus to the cytoplasm in hDPCs following incubation with LPS
(15).
hDPCs can promote reparative dentin formation by
odontoblastic differentiation, resulting in the production of
mineralized matrix (1,3). This property of differentiation by
the pulp cells is critical for its repair following injury or
inflammation and is similar to the processes of osteogenic
differentiation and bone formation. In addition to LPS and TLR4,
NF-κB may also contribute to osteogenic differentiation (12,13)
and the present study hypothesized that they may be involved in the
odontoblastic differentiation of hDPCs. In the present study, hDPCs
were treated with Escherichia coli LPS in growth medium and
odontogenic induction medium (OM), respectively. Subsequently, an
ALP enzymatic assay, reverse transcription quantitative polymerase
chain reaction (RT-qPCR), western blotting and alizarin red
staining were used to investigate and confirm the upregulation of
mineralization indicators in the hDPCs following stimulation. The
translocation of NF-κB in the cells was evaluated using
immunofluorescence microscopy.
Materials and methods
hDPC cultures
Extracted premolars or third molars were collected
from patients (13–25 years old) at the Department of Oral and
Maxillofacial Surgery, Guanghua School and Hospital of Stomatology,
Sun Yat-sen University (Guangzhou, China). Informed consent was
obtained from each patient and the study was performed under the
approval of the ethics committee of Guanghua School and Hospital of
Stomatology, Sun Yat-sen University (Guangzhou, China). The hDPCs
were isolated, as previously reported (16) and cultured in Dulbecco’s modified
Eagle’s medium (DMEM; Gibco-BRL, Grand Island, NY, USA)
supplemented with 100 U/ml penicillin, 100 μg/ml streptomycin
(Sigma-Aldrich, St Louis, MO, USA) and 10% fetal bovine serum (FBS;
Gibco-BRL). The cultures were maintained at 37°C in an incubator
(5% CO2/20% O2). The cells obtained between
passages three and four were used in the subsequent
experiments.
ALP
The hDPCs (5×105 cells per well) were
seeded into 12-well culture plates. The cells in the experimental
groups were treated with either 0.1, 1.0 and 10.0 μg/ml LPS (E.
coli 0111:B4; Sigma) in growth medium or odontogenic induction
medium (OM; 2%FBS, 10 mmol/l β-glycerophosphate, 50 μmol/l ascorbic
acid and 100 nmol/l dexamethasone in DMEM) for 1, 3, 5 and 7 days,
respectively, while those belonging to the control group were
incubated with growth medium and OM alone. The cultures were
maintained at 37°C in an incubator (5% CO2/20%
O2). Following treatment at each time point, the plates
were washed twice with phosphate-buffered-saline (PBS) and lysed
with 200 μl 1% Triton X-100. After 16 h, 30 μl aliquots of cell
lysate per well were subjected to ALP activity and protein content
measurement using an ALP kit (Nanjing Jiancheng Bioengineering
Institute, Nanjing, China) and a bicinchoninic acid assay kit
(Boshide Bioengineering Co., Ltd., Wuhan, China). All results were
normalized by total protein content.
RT-qPCR
The cells from the experimental groups were treated
with 0.1 μg/ml LPS, in growth medium and OM for 3 days, while the
control groups were incubated with growth medium and OM alone. The
cultures were maintained at 37°C in an incubator (5%
CO2/20% O2). Following the treatment, total
RNA was extracted using TRIzol (Invitrogen Life Technologies,
Carlsbad, CA, USA). Quantification of the mRNA levels of DSPP,
DMP-1 and GAPDH were performed based on previously described
methods (17). Complementary DNA
was synthesized from 2 μg total RNA using Revert AidTM First Strand
cDNA Synthesis kit (Fermentas, Ottawa, Canada) according to the
manufacturer’s instructions. RT-qPCR was performed to detect genes
of DSPP, DMP-1 on the Chromo4 four-color Real-Time PCR system
(Bio-Rad Laboratories, Inc., Hercules, CA, USA) using SYBR Green
Realtime PCR Master mix (Toyobo, Osaka, Japan). The sequences of
the primers used are listed in Table
I. The conditions for RT-qPCR were as follows: 95°C for 10 min
for activation, followed by 40 cycles of denaturation at 95°C for
15 sec and primer extension at 60°C for 1 min. All reactions were
performed in triplicate for the independent experiments. The mean
cycle threshold (ΔCt) value of each target gene was normalized
against ΔCt value of house-keeping gene GAPDH and the relative
expression calculated using the following formula:
2−(normalized average ΔCts)x104. The
expression of the target gene was subsequently converted to the
fold-change of the control gene.
 | Table IPrimer sequences used in reverse
transcription quantitative polymerase chain reaction. |
Table I
Primer sequences used in reverse
transcription quantitative polymerase chain reaction.
| Gene | Primer sequence | Length (bp) |
|---|
| DSPP | Forward:
5′-GCCACTTTCAGTCTTCAAAGAGA-3′
Reverse: 5′-GCCCAAATGCAAAAATATGTAA-3′ | 130 |
| DMP1 | Forward:
5′-TGGGCATAGATTTCCTCTTTG-3′
Reverse: 5′-TGAGCAGGATGCTGATCTTC-3′ | 121 |
| GAPDH | Forward:
5′-AAGGTGAAGGTCGGAGTCAA-3′
Reverse: 5′-AATGAAGGGGTCATTGATGG-3′ | 108 |
Western blotting
The cells from the experimental groups were treated
with 0.1, 1.0 and 10.0 μg/ml LPS, in growth medium and in OM for 3
days, while the control group cells were incubated with growth
medium and OM alone. The cultures were maintained at 37°C in an
incubator (5% CO2/20% O2). Following
treatment, the hDPCs were harvested in lysis buffer containing 1%
Igepal CA-630 (Sigma-Aldrich), 0.5% sodium deoxycholate and 0.1%
sodium dodecylsulfate-polyacrylamide. Protease inhibitor cocktail
(Merck Millipore, Darmstadt, Germany), containing AEBSF,
hydrochloride, aprotinin, bovine lung, crystalline, E-64 protease
inhibitor, EDTA, disodium, leupeptin and hemisulfate, was added
prior to cell lysis. Western blotting was performed, as described
previously (11). The proteins
were detected using mouse polyclonal antibodies against human
dental sialoprotein (DSP; 1:1,000; Santa Cruz Biotechnology, Inc.,
Santa Cruz, CA, USA) and mouse monoclonal antibodies against human
GAPDH (1:3,000; EarthOx, San Francisco, CA, USA).
Immunofluorescence
The hDPCs (5.0×105 cells per well) were
seeded into 12-well culture plates and cultured overnight. The
experimental group was then treated with 0.1 μg/ml LPS,
with/without OM, for 16 h, while the control group was incubated
with growth medium and OM alone. The cultures were maintained at
37°C in an incubator (5% CO2/20% O2).
Following treatment, the cells were fixed with methanol for 15 min
and washed with PBS. The cells were then permeabilized with 0.1%
Triton X-100 in PBS for 10 min, incubated in 3% bovine serum
albumin in PBS for 30 min and then with the primary monoclonal
anti-NF-κB antibody (1:50; Cell Signaling Technology, Inc., Boston,
MA, USA) and primary monoclonal anti-c-Jun antibody (1:50;
Millipore, Temecula, CA, USA) simultaneously at 4°C overnight. The
cells were rinsed and then incubated with fluorescein
isothiocyanate-conjugated secondary antibody (1:200; Cell Signaling
Technology, Inc.). Detection was then performed using a
fluorescence microscope (Axio Observer Z1; Zeiss, Oberkochen,
Germany).
Alizarin red stain
The hDPCs (1.0×106 cells per well) were
seeded into 6-well culture plates and cultured overnight. The
experimental group was then treated with 0.1 μg/ml LPS in growth
medium and OM for 14 days and the control group was incubated with
growth medium and OM alone. Following treatment, at each time
point, the mineralized cells were washed with 1X PBS and fixed
using 95% ethanol at room temperature for 10 min. Calcium
accumulation was detected using 40 mM alizarin red stain solution
(Sigma-Aldrich). Following washing with distilled water, images of
the dried-culture dishes were captured under a fluorescence
microscope (Axio Observer Z1; Zeiss). The positively stained
nodules appeared orange/red.
Statistical analysis
The SPSS 16.0 software package (SPSS, Inc, Chicago,
IL, USA) was used to perform statistical analyses. The ALP
activity, protein expression of DSP and gene expression of DSPP in
the different treatment groups were assessed using the Least
Significant Difference test. The gene expression of DMP-1 was
assessed using the Kruskal-Wallis and Bonferronni’s test due to
heterogeneity of variance. P<0.05 and P<0.01 were considered
to indicate statistically significant differences.
Results
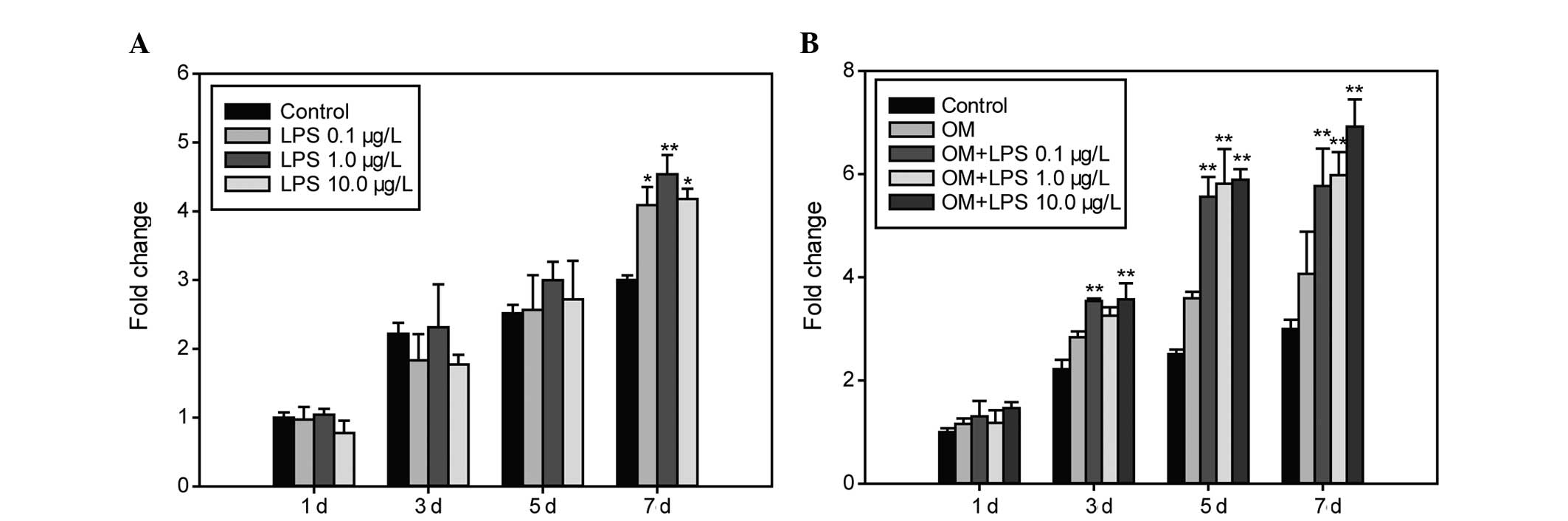
Effect of LPS on ALP activity
The activity of ALP was examined at different time
points using an ALP enzymatic assay and was found to be higher in
the hDPCs treated with LPS at all concentrations in the growth
medium on the 7th day (P<0.05; Fig.
1A), while no changes were observed on the 3rd and 5th days. In
OM, the ALP activity of cells exposed to three concentrations of
LPS increased at each time point (P<0.01; Fig. 1B) with the exception of the 1st day
and at the final concentration of 1.0 μg/l LPS on the 3rd day.
Exposure of the stimulated cells to 10.0 μg/l resulted in the
highest ALP activity on day 7.
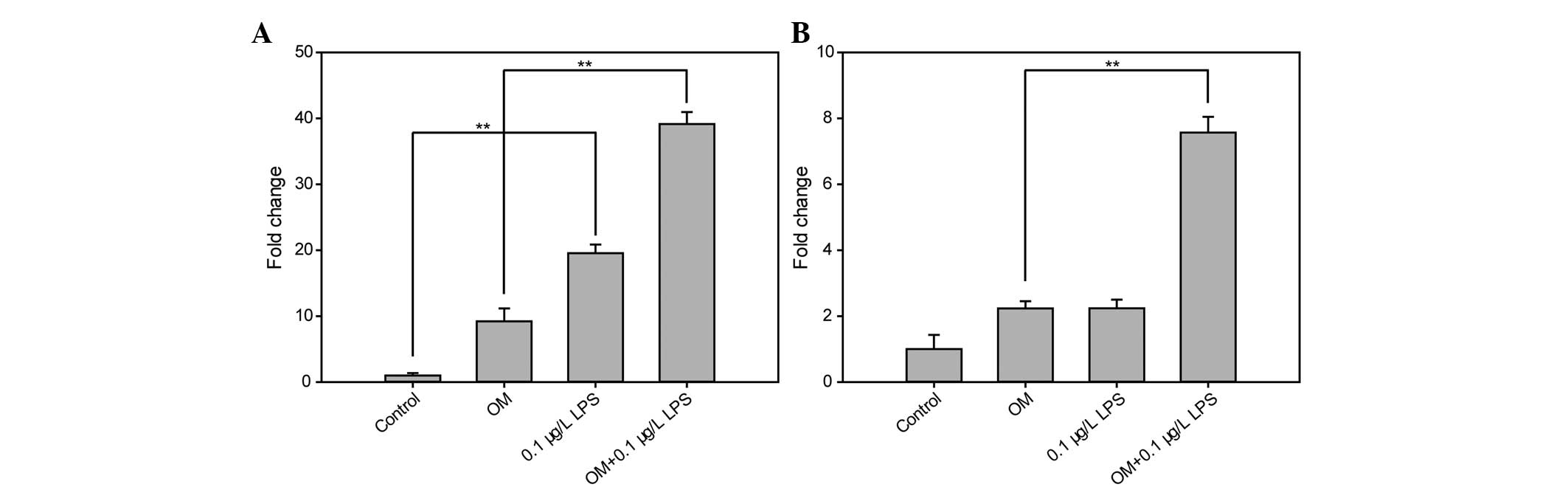
Effect of LPS on the gene expression
levels of DSPP and DMP-1
The gene expression levels of DSPP and DMP-1 were
examined using RT-qPCR on the 3rd day of treatment with 0.1 μg/ml
LPS in the growth medium and OM. The expression levels of DSPP
increased significantly in the hDPCs treated with LPS compared with
the control group in growth medium (P<0.05) and in OM
(P<0.01). The expression levels of DMP-1 were significantly
higher in the hDPCs treated with LPS compared with OM alone
(P<0.05; Fig. 2).
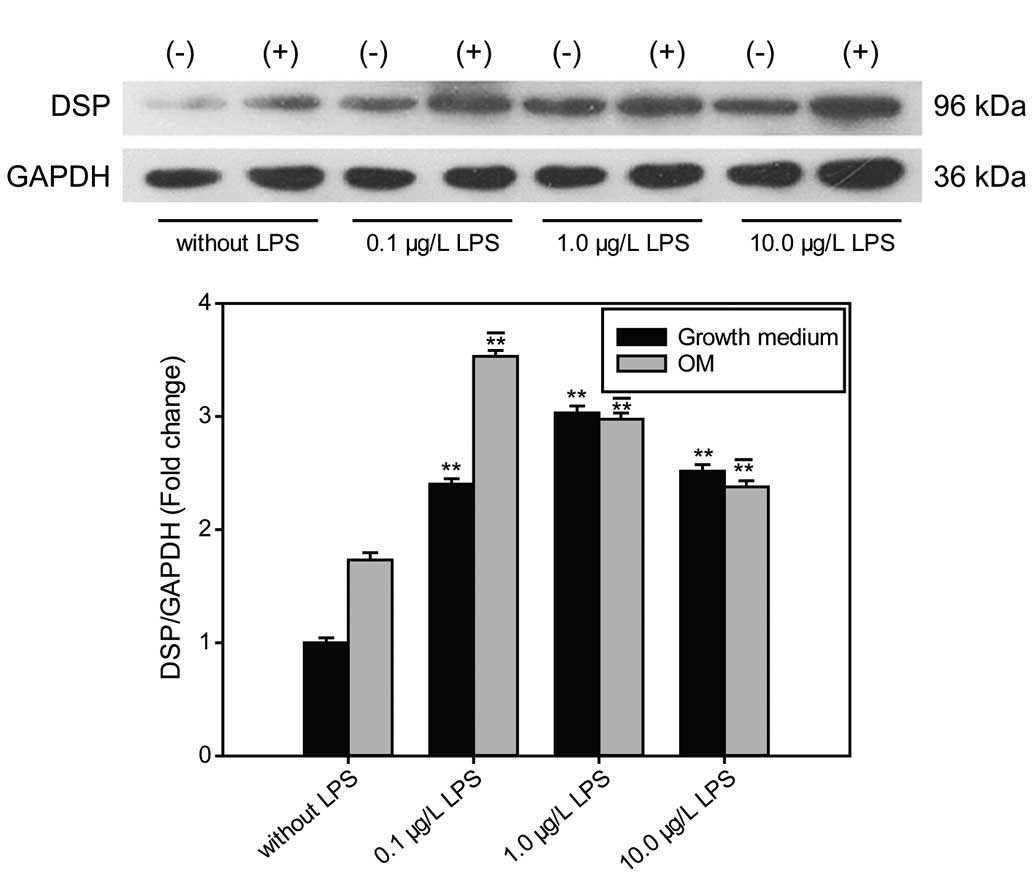
Effect of LPS on the expression of
DSP
After treatment with LPS for 3 days, the protein
levels of DSP were increased in the hDPCs at concentrations of 0.1,
1.0 and 10.0 μg/ml compared with the control group in growth medium
and OM (P<0.01). The highest expression level of DSP was
observed at a concentration of 0.1 μg/ml LPS in OM (P<0.01;
Fig. 3).
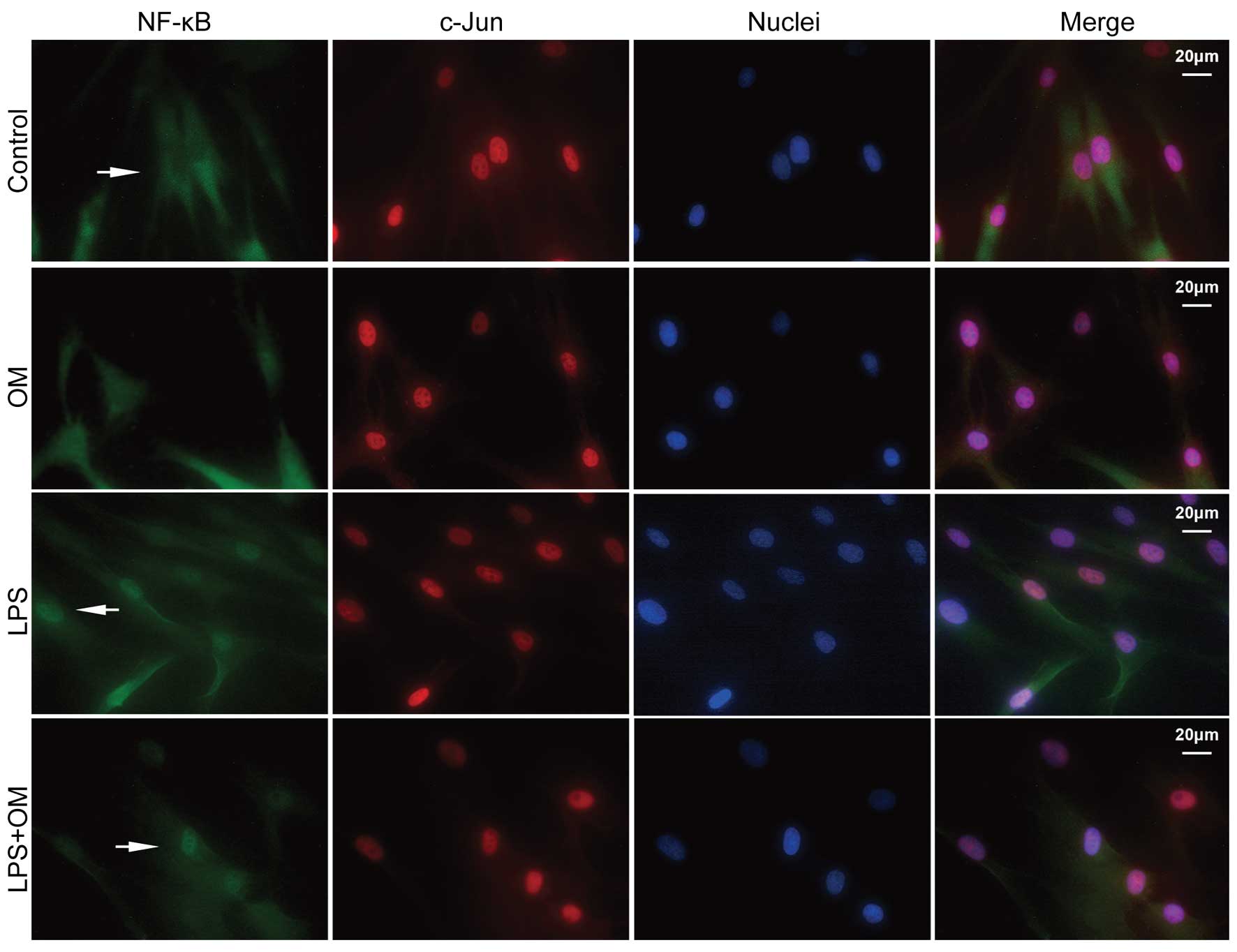
LPS induces the nuclear translocation of
NF-κB, but not c-Jun, in the hDPCs
The nuclear and cytosolic expression levels of NF-κB
and c-Jun were detected using immunofluorescence after 16 h
treatment with LPS (0.1 μg/ml) with or without OM. NF-κB was
localized predominantly in the cytoplasm of the cells treated with
2% FBS in DMEM alone and in the nuclei of the cells in the
experimental groups treated with LPS wit or without OM (Fig. 4). However, no change was noted in
the expression of c-Jun in the cells in either the control or
experimental group.
Effect of LPS on the mineralized nodule
formation of hDPCs
To determine whether E. coli LPS stimulation
increased cellular calcification, the cells were treated in OM with
or without 0.1 μg/ml LPS for 14 days (Fig. 5). The results revealed that LPS
promoted calcified nodule formation when the hDPCs were stimulated.
As shown in Fig. 5, nodules of
increased size and number were formed in the hDPCs exposed to OM
and LPS compared with those stimulated by OM alone.
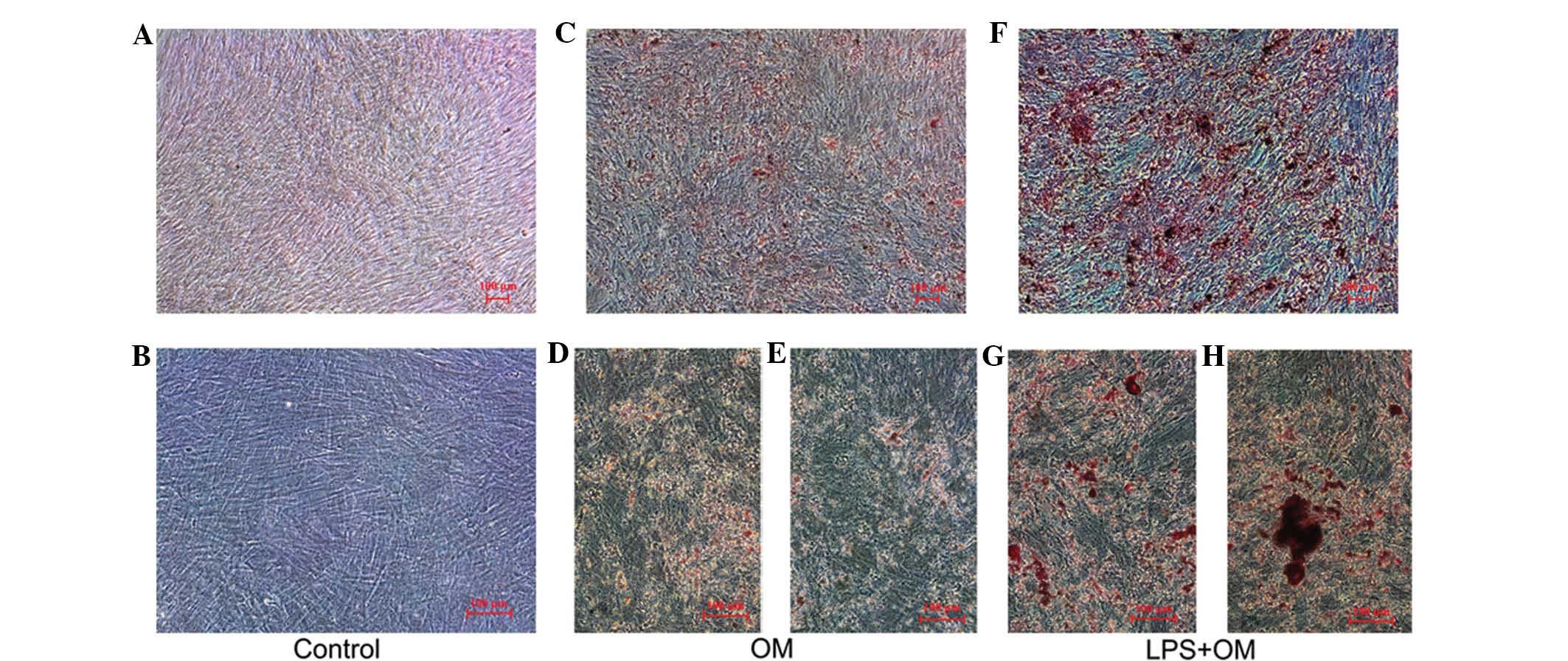 | Figure 5Effect of LPS on calcified nodule
formation of the hDPCs. The hDPCs were cultured in (A and B) growth
medium, (C, D and E) OM and (F, G and H) OM and LPS for 14 days,
respectively. (A, C and F, ×50; B, D and G, ×100). Alizarin red
staining revealed that the mineral nodule accumulation of the hDPCs
treated by OM and LPS was higher compared with the hDPCs treated
with OM only. LPS, lipopolysaccharide; hDPCs, human dental pulp
cells; OM, odontogenic induction medium. |
Discussion
Subsequent to injury or inflammation, reparative
dentin is formed by the odontoblastic differentiation of hDPCs
leading to increased production of mineralized matrix (1,3). As
bacteria are one of the most important causative agents of
pulpitis, LPS, which comprises a major molecular component of the
outer cell wall of Gram-negative bacteria, is often used to
establish a model of inflammation for the investigation of dental
pulp cells (8). The present study
used E. coli LPS, as it is a putative inflammation inducer,
evokes a protective inflammatory reaction and is hypothesized to
induce signal transduction through TLR4 (9,10).
The present study hypothesized that LPS and TLR4 may be involved in
the odontoblastic differentiation of hDPCs.
ALP, DSPP and DMP-1 are mineralization markers for
odontoblast and osteoblast-like differentiation of hDPCs (3,18).
ALP activity is higher in odontoblast-like cells compared to
undifferentiated dental mesenchymal cells and is closely associated
with dentin formation (19). In
the present study, increased ALP activity was observed in the hDPCs
treated with LPS, which was more marked when the cells were
simultaneously stimulated with OM. The rise in ALP activity was
time-dependent, but not dose-dependent, indicating that treatments
with OM and LPS, with or without OM, may enhance odontoblastic
differentiation. DSP, which promotes predentin to transform into
mature dentin (20), is considered
to be a specific labeled protein for odontoblastic differentiation
(21). DSPP and DMP-1 are also
expressed by differentiating odontoblasts. Western blot analysis
revealed that the protein levels of DSP were similar to those of
ALP following treatment. Similarly, the mRNA levels of DSPP and
DMP-1 were reconfirmed by western blotting. Furthermore, larger and
increased numbers of nodules were formed in the hDPCs exposed to OM
and LPS compared with those stimulated by OM only. These
observations suggested that there may be an association between LPS
stimulation, with or without OM, and odontoblastic
differentiation.
The results of the present study are in agreement
with those of calcific aortic valve cells (hAVICs). Previous
studies on calcific aortic valve stenosis have revealed that LPS
stimulation of hAVICs induces inflammatory mediators and the gene
expression of osteogenic factors, similar to those induced by OM
(22,23). Osteogenic changes have been
associated with high expression levels of TLR4 (13). In addition, the combined effects of
LPS and OM have been demonstrated on the osteogenic differentiation
of hAVICs (12). As fibroblasts
form the major bulk of hDPCs, it they may have similarities to
hAVICs, which are myofibroblasts. It has been previously reported
that human dental pulp stem cells exposed to LPS, obtained from
Porphyromonas gingivalis, exhibit a reduction in the
expression levels of DSPP and osteocalcin and this effect can be
moderated by TLR2 inhibition (24). Based on this finding and the
results of the present study, it can be concluded that LPS from
different bacteria, including P. gingivalis and E.
coli, may not simulate the same receptor and may exert
different effects with different receptors.
NF-κB activation is known to contribute to cytokine
induction in response to bacterial products and is necessary for
innate immunity and inflammation (25,26).
It has been demonstrated that NF-κB is required for the LPS-induced
expression of interleukin-8 in the hDPSCs and NF-κB is important in
mediating the intracellular signaling generated by LPS, which
induces the expression of pro-inflammatory genes (27). LPS has also been demonstrated to
cause NF-κB intranuclear translocation and rapid phosphorylation in
hAVICs (12). In the present
study, the LPS-induced translocation of NF-κB was demonstrated
using immunofluorescence and was in accordance with the results of
western blot analysis reported in a previous study (15), indicating its role during the
LPS-induced odontoblastic differentiation of hDPCs. However, LPS
did not affect the translocation of c-Jun. The results suggested
that NF-κB was involved in the transcriptional activation of
various downstream genes and may have a similar role in the
odontoblastic differentiation of hDPCs, improved by OM and LPS with
or without OM.
In the present study, the expression of
mineralization markers for the odontoblast and osteoblast-like
differentiation of hDPCs stimulated by LPS, with or without OM,
increased, suggesting the involvement of TLR4 and NF-κB. To further
understand the roles of LPS, TLR4 and NF-κB in the odontoblastic
differentiation of hDPCs, further studies examining the involvement
of the TLR4 inhibitor, NF-κB inhibitor and the relevant signal
pathways involved are required.
In addition to inducing ALP activity, LPS was
observed to increase the gene expression levels of DSPP and DMP-1
and the protein expression of DSP in the hDPCs, particularly in
combination with OM. The nuclear translocation of NF-κB, induced by
LPS suggested that NF-κB is involved in the LPS-induced increase in
odontoblastic differentiation of hDPCs.
Acknowledgements
This study was supported by the Key Clinical Program
of the Ministry of Health, China (no. [2010] 439), the National
Nature Science Foundation of China (nos. 81100742 and 81200775) and
the Fundamental Research Funds for the Central Universities (no.
12ykpy65).
References
|
1
|
Gronthos S, Brahim J, Li W, et al: Stem
cell properties of human dental pulp stem cells. J Dent Res.
81:531–535. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Goldberg M, Farges JC, Lacerda-Pinheiro S,
et al: Inflammatory and immunological aspects of dental pulp
repair. Pharmacol Res. 58:137–147. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wei X, Ling J, Wu L, Liu L and Xiao Y:
Expression of mineralization markers in dental pulp cells. J Endod.
33:703–708. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wei X, Wu L, Ling J, et al: Differentially
expressed protein profile of human dental pulp cells in the early
process of odontoblast-like differentiation in vitro. J Endod.
34:1077–1084. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tobias PS, Mathison JC and Ulevitch RJ: A
family of lipopolysaccharide binding proteins involved in responses
to gram-negative sepsis. J Biol Chem. 263:13479–13481.
1988.PubMed/NCBI
|
|
6
|
Vernooy JH, Dentener MA, van Suylen RJ,
Buurman WA and Wouters EF: Long-term intratracheal
lipopolysaccharide exposure in mice results in chronic lung
inflammation and persistent pathology. Am J Respir Cell Mol Biol.
26:152–159. 2002. View Article : Google Scholar
|
|
7
|
Hanazawa S, Nakada K, Ohmori Y, Miyoshi T,
Amano S and Kitano S: Functional role of interleukin 1 in
periodontal disease: induction of interleukin 1 production by
Bacteroides gingivalis lipopolysaccharide in peritoneal macrophages
from C3H/HeN and C3H/HeJ mice. Infect Immun. 50:262–270.
1985.PubMed/NCBI
|
|
8
|
Vianna ME, Horz HP, Conrads G, Zaia AA,
Souza-Filho FJ and Gomes BP: Effect of root canal procedures on
endotoxins and endodontic pathogens. Oral Microbiol Immunol.
22:411–418. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hoshino K, Takeuchi O, Kawai T, et al:
Cutting edge: Toll-like receptor 4 (TLR4)-deficient mice are
hyporesponsive to lipopolysaccharide: evidence for TLR4 as the Lps
gene product. J Immunol. 162:3749–3752. 1999.PubMed/NCBI
|
|
10
|
Yang X, Coriolan D, Murthy V, Schultz K,
Golenbock DT and Beasley D: Proinflammatory phenotype of vascular
smooth muscle cells: role of efficient Toll-like receptor 4
signaling. Am J Physiol Heart Circ Physiol. 289:H1069–H1076. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jiang HW, Zhang W, Ren BP, Zeng JF and
Ling JQ: Expression of toll like receptor 4 in normal human
odontoblasts and dental pulp tissue. J Endod. 32:747–751. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Meng X, Ao L, Song Y, et al: Expression of
functional Toll-like receptors 2 and 4 in human aortic valve
interstitial cells: potential roles in aortic valve inflammation
and stenosis. Am J Physiol Cell Physiol. 294:C29–C35. 2008.
View Article : Google Scholar
|
|
13
|
Yang X, Fullerton DA, Su X, Ao L and Meng
X: Pro-osteogenic phenotype of human aortic valve interstitial
cells is associated with higher levels of Toll-like receptors 2 and
4 and enhanced expression of bone morphogenetic protein 2. J Am
Coll Cardiol. 53:491–500. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
O’Neill LA and Bowie AG: The family of
five: TIR-domain-containing adaptors in Toll-like receptor
signalling. Nat Rev Immunol. 7:353–364. 2007. View Article : Google Scholar
|
|
15
|
Lee JC, Yu MK, Lee R, et al: Terrein
reduces pulpal inflammation in human dental pulp cells. J Endod.
34:433–437. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang J, Wei X, Ling J, Huang Y and Gong Q:
Side population increase after simulated transient ischemia in
human dental pulp cell. J Endod. 36:453–458. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jiang HW, Ling JQ and Gong QM: The
expression of stromal cell-derived factor 1 (SDF-1) in inflamed
human dental pulp. J Endod. 34:1351–1354. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yamada Y, Fujimoto A, Ito A, et al:
Cluster analysis and gene expression profiles: a cDNA microarray
system-based comparison between human dental pulp stem cells
(hDPCs) and human mesenchymal stem cells (hMSCs) for tissue
engineering cell therapy. Biomaterials. 27:3766–3781. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shiba H, Mouri Y, Komatsuzawa H, et al:
Enhancement of alkaline phosphatase synthesis in pulp cells
co-cultured with epithelial cells derived from lower rabbit
incisors. Cell Biol Int. 27:815–823. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Paine ML, Luo W, Wang HJ, et al: Dentin
sialoprotein and dentin phosphoprotein overexpression during
amelogenesis. J Biol Chem. 280:31991–31998. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Begue-Kirn C, Krebsbach PH, Bartlett JD
and Butler WT: Dentin sialoprotein, dentin phosphoprotein,
enamelysin and ameloblastin: tooth-specific molecules that are
distinctively expressed during murine dental differentiation. Eur J
Oral Sci. 106:963–970. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Babu AN, Meng X, Zou N, et al:
Lipopolysaccharide stimulation of human aortic valve interstitial
cells activates inflammation and osteogenesis. Ann Thorac Surg.
86:71–76. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lopez J, Fernandez-Pisonero I, Duenas AI,
et al: Viral and bacterial patterns induce TLR-mediated sustained
inflammation and calcification in aortic valve interstitial cells.
Int J Cardiol. 158:18–25. 2012. View Article : Google Scholar
|
|
24
|
Yamagishi VT, Torneck CD, Friedman S,
Huang GT and Glogauer M: Blockade of TLR2 inhibits Porphyromonas
gingivalis suppression of mineralized matrix formation by human
dental pulp stem cells. J Endod. 37:812–818. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Janssens S and Beyaert R: A universal role
for MyD88 in TLR/IL-1R-mediated signaling. Trends Biochem Sci.
27:474–482. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chang J, Zhang C, Tani-Ishii N, Shi S and
Wang CY: NF-κB activation in human dental pulp stem cells by TNF
and LPS. J Dent Res. 84:994–998. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
He W, Qu T, Yu Q, et al: LPS induces IL-8
expression through TLR4, MyD88, NF-kappaB and MAPK pathways in
human dental pulp stem cells. Int Endod J. 46:128–136. 2013.
View Article : Google Scholar
|