Introduction
Liver cancer is a malignant disease, which forms on
the surface or within the liver, including in blood vessels or the
bile duct (1–3). It is one of the most common types of
solid tumor worldwide and is the third leading cause of
cancer-associated mortality (4).
Primary liver cancer is the most common type of malignant tumor,
with poor prognosis and a high mortality rate (5,6). It
currently leads to 360,000 incident cases and the mortality of
350,000 individuals annually in China (7). The incidence and mortality rates of
hepatocellular carcinoma in China account for 40–45% and 18.8% of
the patients with liver cancer worldwide, respectively. Untreated
patients with advanced disease usually survive <6 months
(8) and, in 25 randomized clinical
trials, the 1 and 2 year survival rates of untreated patients were
between 10 and 72% and between 8 and 50%, respectively (8). Thus, liver cancer is one of the major
diseases affecting health.
Tumors occur in several diseases and result from
disordered cell cycle regulation or loss of control over cell
division. The unchecked cells may eventually form malignancies,
with the activation of oncogenes or inhibition of tumor suppressor
genes (9,10).
The cyclin kinase inhibitor p27kip1 is a
cell-cycle regulatory protein, which is one of the most important
regulators of the cell cycle (11). It interacts with cyclin-dependent
kinase (CDK)2 and CDK4 to inhibit cell cycle progression at the G1
phase and is also the checkpoint of the G1-S phase transition
(12,13). The degradation of
p27kip1 is dependent on the ubiquitin-proteasome
pathway. Thus is generally triggered by phosphorylation of
conserved threonine (Thr187) and subsequent ubiquitination by Skp,
Cullin and F-box (SCF) complexes, and is required for cell division
(14,15). The ubiquitin-proteasome system is
mediated by three enzymes, ubiquitin-activating enzyme,
ubiquitin-conjugating enzyme and E3 ubiquitin ligase. S-phase
kinase-associated protein 2 (Skp2) is one type of E3 ubiquitin
ligase for recruiting a specific substrate and determining their
specificity (16). Thus, Skp2 acts
as an adaptor of the SCF-Skp2 complex and recognizes phosphorylated
substrates, including p27kip1 and p21.
Skp2 acts as an oncogene and overexpression of Skp2
is frequently observed in several types of human tumor, including
lymphoma, prostate cancer, melanoma, nasopharyngeal carcinoma,
pancreatic cancer and breast carcinomas, which promotes the
progression and metastasis of human cancer (17–20).
By comparison, p27kip1, a tumor inhibitor protein,
usually inhibits the progression of human cancer. In the present
study, the role of the Skp2-p27 pathway in the progression of human
liver cancer was investigated and the underlying mechanism was
examined. Clarification of the process and progression of
carcinogenesis in liver tissues may provide novel targets for the
effective prevention of liver cancer.
Materials and methods
Liver cancer cell lines
The HepG2 and SSMC-7721 liver cancer cell lines
(American Type Culture Collection, Manassas, VA, USA) were
maintained in the laboratory at Jinan Central Hospital Affiliated
to Shandong University (Shandong, China) and were cultured in
Dulbecco’s modified Eagle’s medium (DMEM) with 10% fetal bovine
serum (HyClone, GE Healthcare Life Sciences, Little Chalfont, UK),
1% penicillin and 1% streptomycin (Gibco-BRL, Invitrogen Life
Technologies, Carlsbad, CA, USA).
Small interfering (si)RNA and
antibodies
The cells were planted in 48-well plates and
subsequent to a 24-h resting period, the Skp2 p45 siRNA and
scramble siRNA was transfected with Lipofectamine 2000 (Invitrogen
Life Technologies). Following 8 h of transfection, the DMEM medium
was replaced and the lysates were prepared subsequent to
transfection for 48 h. The Skp2 p45 siRNA was purchased from Santa
Cruz Biotechnology, Inc. (sc-36499; Dallas, TX, USA) and scramble
siRNA was synthesized by Shanghai Jima Company (Shanghai, China).
Rabbit polyclonal immunoglobulin G (IgG) Skp2 p45 antibody (200
μg/ml; sc-7164), mouse monoclonal IgG1 p27 antibody (200
μg/ml; sc-53906) and mouse monoclonal IgG1 β-actin
(AC-15) antibody (100 μg/ml; sc-69879), as well as horseradish
peroxidase-conjugated goat anti-mouse secondary antibody (sc-2005)
were all obtained from Santa Cruz Biotechnology, Inc.
Histology and immunohistochemistry
Liver tissue samples were obtained from Jinan
Central Hospital Affiliated to Shandong University. The subjects
included 50 cases of hepatocellular carcinoma and 40 paratumor
tissues with an average age of 58±13 years. The subjects were
recruited from March 2011 to December 2013. The specimens were
obtained during surgery. There were 52 male and 38 female patients
during the experiment. Written informed consent was also obtained
from the patients’ families. The study was approved by the ethics
committee of Jinan Central Hospital Affiliated to Shandong
University (Jinan, China). The liver tissues were fixed in neutral
buffered paraformaldehyde and processed for hematoxylin and eosin
(H&E) staining. Immunohistochemical staining was performed, as
previously described (21,22). Briefly, the paraffin-embedded
sections were dewaxed, rehydrated, inhibited and incubated
overnight with the primary antibodies (anti-p27 and anti-Skp2).
Subsequently, the samples were incubated with goat anti-mouse and
goat anti-rabbit secondary antibodies (Santa Cruz Biotechnology)
for 1 h. The slides were stained, dehydrated and cleared and the
stained slides were mounted in Permount and visualized using a
Nikon Eclipse Ti microscope (Nikon, Tokyo, Japan). Images were
captured using an FE330 digital camera (Olympus Corporation, Tokyo,
Japan), which was attached to the microscope.
Western blot analysis
The whole cell extracts were prepared and separated
by PAGE, as previously described (23–25)
using buffer and gel obtained from Shanghai Qcbio Science and
Technologies Co., Ltd. (Shanghai, China) and the machine
(Mini-Protean® Tetra Cell) from obtained from Bio-Rad
(Hercules, CA, USA). The proteins of the sample were separated
using gel electrophoresis and were subsequently transferred to a
polyvinylidine difluoride membrane (Millipore, Billerica, MA, USA).
Subsequently, blocking of non-specific binding was achieved by
placing the membrane in a solution of 5% bovine serum albumin.
Following blocking, a dilute solution of the appropriate primary
antibody was incubated with the membrane at 4°C overnight. The
antibodies used were as follows: Anti-p27, anti-Skp2, anti-β-actin
and horseradish peroxidase-conjugated goat anti-mouse secondary
antibody.
MTT assay
An MTT assay was performed, as described previously
(26–28). The cells (5×105) were
seeded into 48-well plates. Following culture for 24, 48 or 72 h,
the plates were read on a microplate reader (Varioskan Flash
Multimode Reader; Thermo Fisher Scientific, Waltham, MA, USA) at a
test wavelength of 490 nm and a reference wavelength of 570 nm.
Immunoprecipitation
Immunoprecipitation was performed, as previously
described (29–31). The cells were lysed and the sample
was passed over beads alone or bound to a relevant antibody to
absorb any proteins that non-specifically bind to the
immunoprecipitation components. For immunoprecipitation analysis,
the samples were immunoprecipitated with mouse monoclonal
hemagglutinin (HA)-tag antibodies (A012244-100; GenScript, Nanjing,
China). The immunocomplexes were separated by SDS-PAGE and treated
with the secondary antibody. The complexes of Skp2 or ubiquitin
with p27kip1 were determined by immunoprecipitating the
mouse monoclonal anti-Flag antibody (AF519; Beyotime Institue of
Biotechnology, Haimen, China) and normalized to the quantity of
antibodies. To assess the specificity of the primary antibodies,
negative controls were used, in which the antibodies used to
immunoprecipitate were incubated for 2 h at room temperature with
the respective immunogen peptide.
FACS analysis
Cell apoptosis rate was determined by Annexin
V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) (Hangzhou
Lianke Biology Technology Limited Company, Hangzhou, China)
staining. The one-step staining procedure was selected, which takes
only 10 min to perform. Detection was analyzed by flow cytometry
(FC 500; Beckman Coulter, Brea, CA, USA), which differentiates
between apoptosis and necrosis following Annexin V-FITC and PI
staining.
Statistical analysis
Student’s t-test was used to evaluate statistical
significance. Data were analyzed using SPSS 19.0 (IBM SPSS, Armonk,
NY, USA) Data are expressed as the mean ± standard deviation.
P<0.05 were considered to indicate a statistically significant
difference.
Results
Positive Skp2 staining is observed in the
majority of liver cancer tissues
In the present study, 90 registered patients were
obtained from the database of Jinan Central Hospital Affiliated to
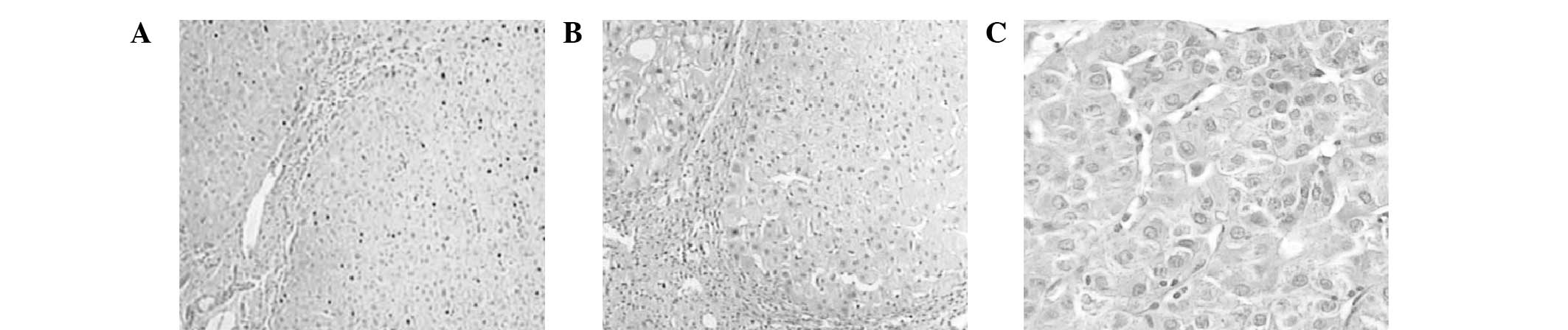
Shandong University. As shown in Fig.
1, H&E staining of the samples of paratumor liver tissue
and hepatocellular carcinoma tissue was compared with normal liver
tissues. It is reported that the Skp2-p27kip1 signaling
pathway is closely associated with cancer (32,33).
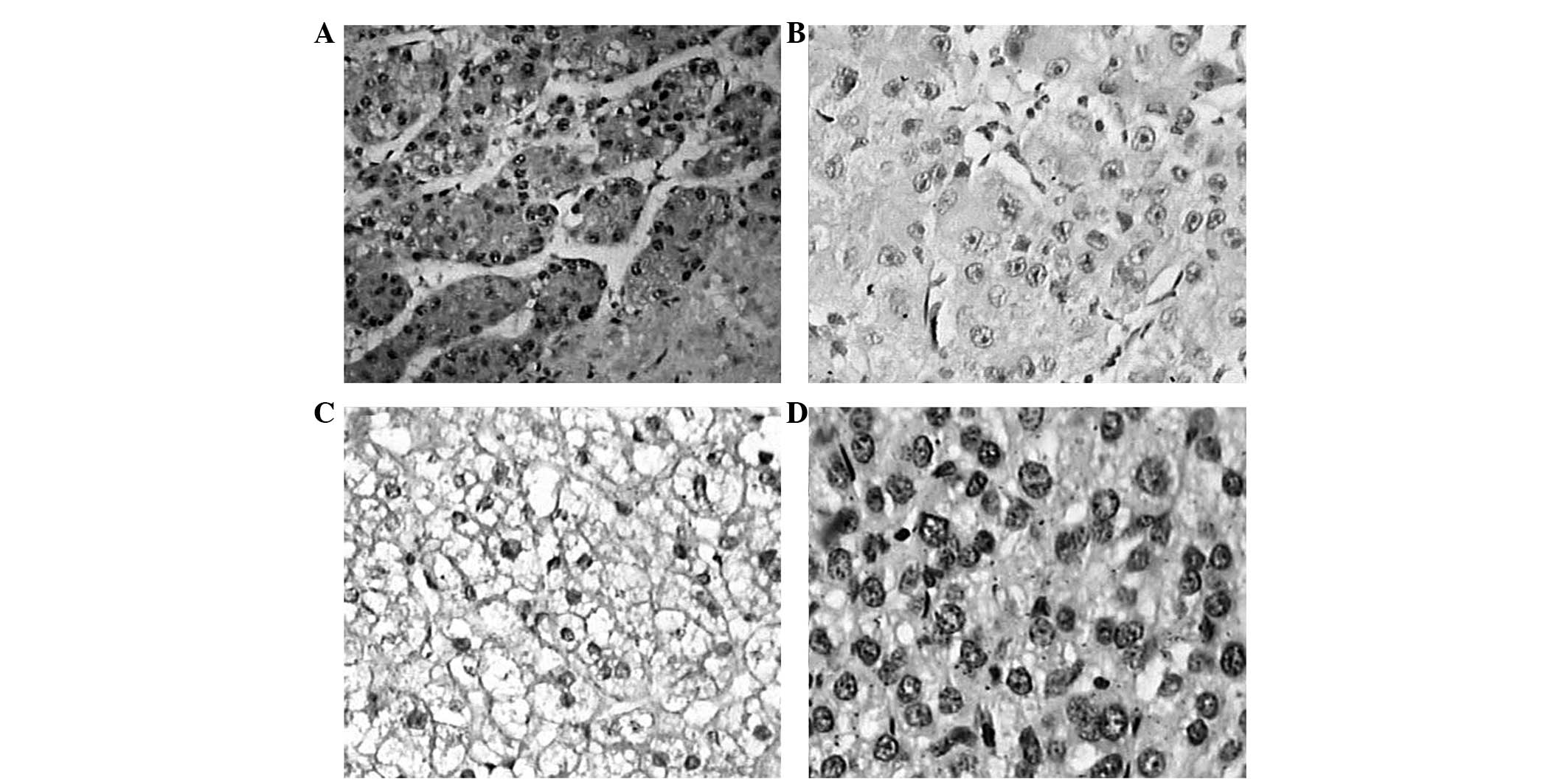
In the present study, the expression levels of Skp2 and
p27kip1 were detected by immunohistochemical analyses in
the paratumor and hepatocellular carcinoma tissues. The results
revealed that the positive brown staining for Skp2 was
predominantly observed in the nuclei with occasional cytoplasmic
expression (Fig. 2). Compared with
the immnunostaining of the paratumor tissues, nuclear and
cytoplasmic localization of Skp2 was observed in the majority of
the examined samples in the patients with hepatocellular carcinoma
(Table I) with 70.0% of the
malignant tumors demonstrating varying levels of positive
immunostaining. A significant difference was observed between the
two groups (P<0.01).
 | Table IExpression of Skp2 in hepatocellular
carcinorma and paratumor tissues. |
Table I
Expression of Skp2 in hepatocellular
carcinorma and paratumor tissues.
| Tissue | Sample no. | Skp2 | Positive rate
(%) |
|---|
|
|---|
| − | + |
|---|
| Hepatocellular
carcinoma | 50 | 15 | 35 | 70.0a |
| Paratumor | 40 | 34 | 6 | 15.0 |
Positive Skp2 staining is limited in
hepatocellular carcinoma
The present study then detected the expression and
localization of p27kip1 in the hepatocellular carcinoma
and paratumor tissues. As shown in Fig. 2, the results of the immunostaining
demonstrated that p27kip1 exhibited a primarily nuclear
pattern of expression. As shown in Table II, 84.0% (42/50) of the malignant
hepatocellular carcinoma tissues demonstrated negative
immunostaining; however, negative immunostaining was observed in
only 10% (4/40) of the paratumor tissues, with a significant
difference observed between the two groups (P<0.01).
| Table IIExpression of p27kip1 in
hepatocellular carcinorma and paratumor tissues. |
Table II
Expression of p27kip1 in
hepatocellular carcinorma and paratumor tissues.
| Tissue | Sample no. |
p27kip1 | Positive rate
(%) |
|---|
|
|---|
| − | + |
|---|
| Hepatocellular
carcinoma | 50 | 42 | 8 | 16.0a |
| Paratumor | 40 | 4 | 36 | 90.0 |
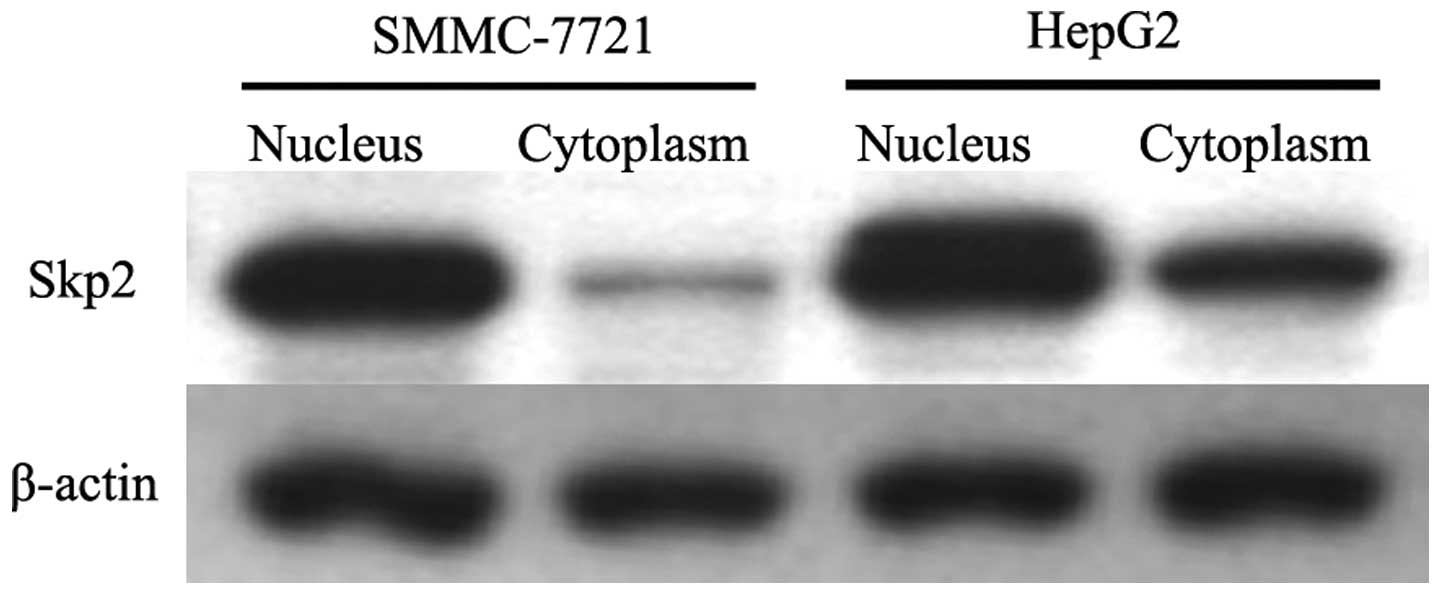
Localization of Skp2 detected by western
blot analysis
In order to confirm the localization of Skp2 in SSMC
7721 and HepG2 cells, cytoplasmic and nucleic proteins were
prepared and western blot analysis was performed. As shown in
Fig. 3, the results revealed that
the majority of Skp2 was located in the nuclei of the cells;
although there was low expression detected in the cytoplasm.
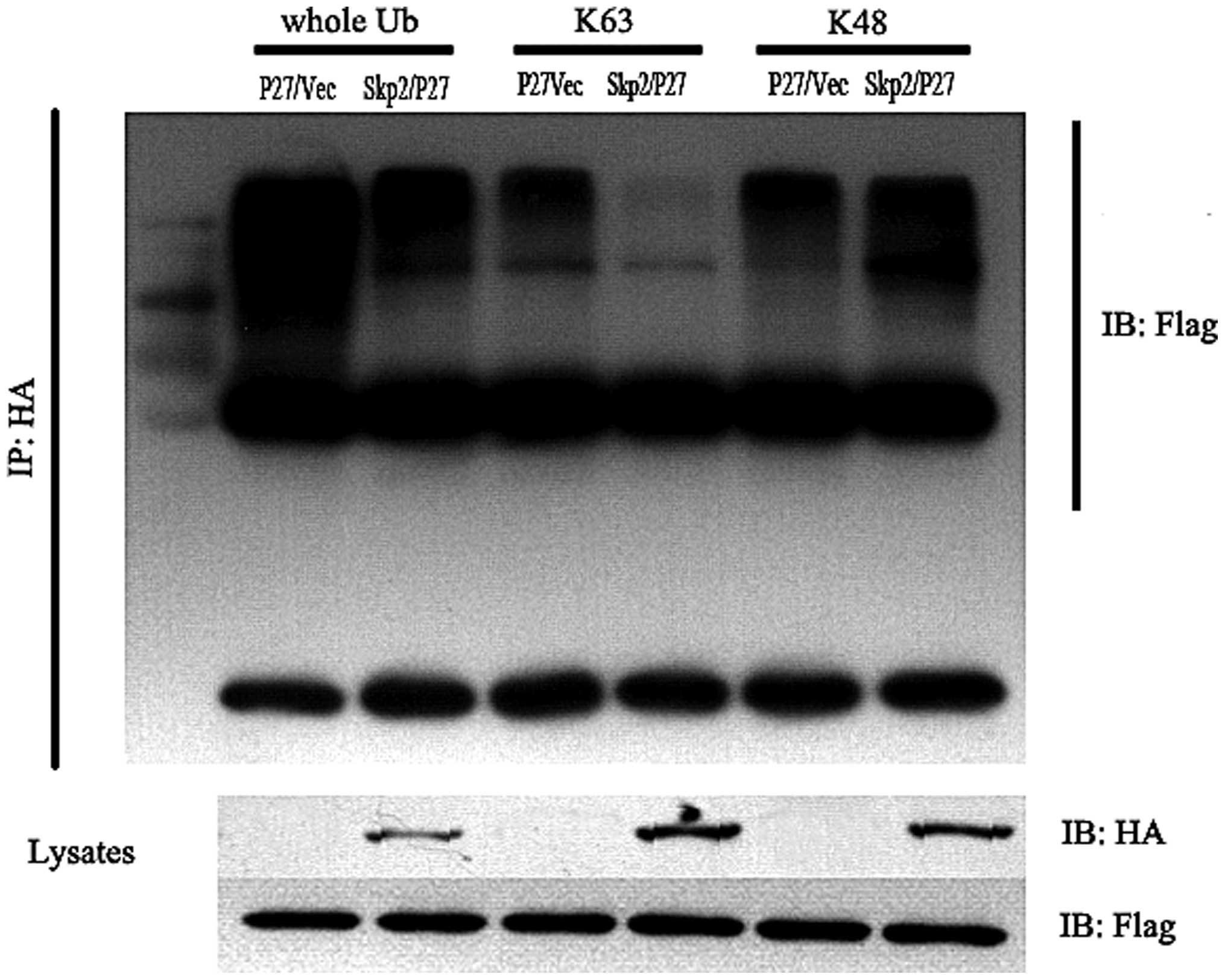
Skp2 is degraded by the
ubiquitin-proteasome pathway regulated by Skp2
In order to examine whether the degradation of
p27kip1 is regulated by Skp2, 293T cells, which
expressed HA-Skp2, Flag-p27 and ubiquitin together were used. After
48 h, a co-immunoprecipitation assay was performed and the data
demonstrated that the K48-linked p27kip1 increased and
the K63-linked p27kip1 decreased. The total quantity of
ubiquitinated p27kip1 was also downregulated (Fig. 4). Taken together, these data
demonstrated that p27kip1 was degraded by the
ubiquitin-proteasome pathway, which was mediated by the Skp2
complex.
Interference of Skp2 induces the
apoptosis and inhibits the proliferation of SSMC-7721 cells
Skp2 is an oncogene, which is highly expressed in
tumor cells. The present study investigated whether interference of
Skp2 with specific siRNA induced the apoptosis of liver cancer
cells or promoted liver cancer cell death. As shown in Fig. 5, the apoptotic rates of the
SSMC-7721 cells transfected with Skp2-siRNA were significantly
higher compared with the control cells (P<0.01).
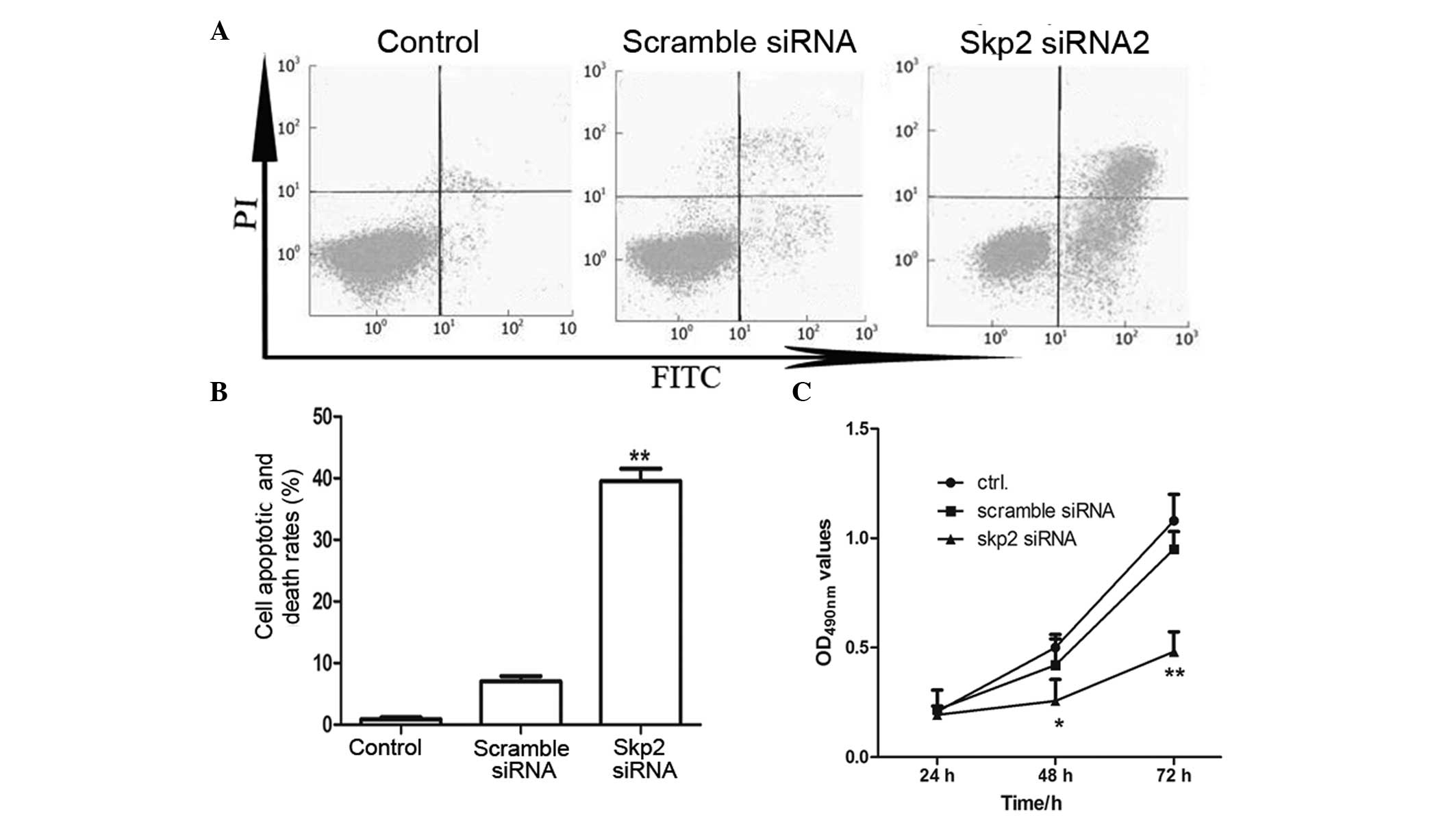 | Figure 5Interference of Skp2 promotes the
apoptosis and death of SSMC-7721 cells. (A) Fluorescence-activated
cell sorting assay. The SSMC-7721 cells were transfected with Skp2
siRNA. After 48 h, the apoptotic rate was detected by Annexin V-PI
staining. The untreated cells and the cells transfected with
scramble siRNA were used as negative controls. (B) Histograms of
the apoptotic rates by Annexin V-PI staining.
**P<0.01, compared with the negative control cells.
(C) MTT assay, in which the cells (5×105) were seeded
into 48-well plates and transfected with Skp2 siRNA or scramble
siRNA for 24, 48 and 72 h, respectively. Data are expressed as the
mean ± standard deviation of at least three independent experiments
on different individuals. *P<0.05 and
**P<0.01, compared with the control group. Skp2,
S-phase kinase-associated protein 2; FITC, fluorescein
isothiocyanate; PI, propidium iodide; ctrl, control; OD, optical
density. |
In the MTT assay (Fig.
5C), the optical density at 490 nm in the cells transfected
with Skp2-siRNA were significantly lower compared with that of the
control cells (P<0.05 and P<0.01). This was consistent with
the results observed by fluorescence-activated cell sorting (FACS),
suggesting that the proliferation of cancer cells was inhibited
following transfection with Skp2-siRNA.
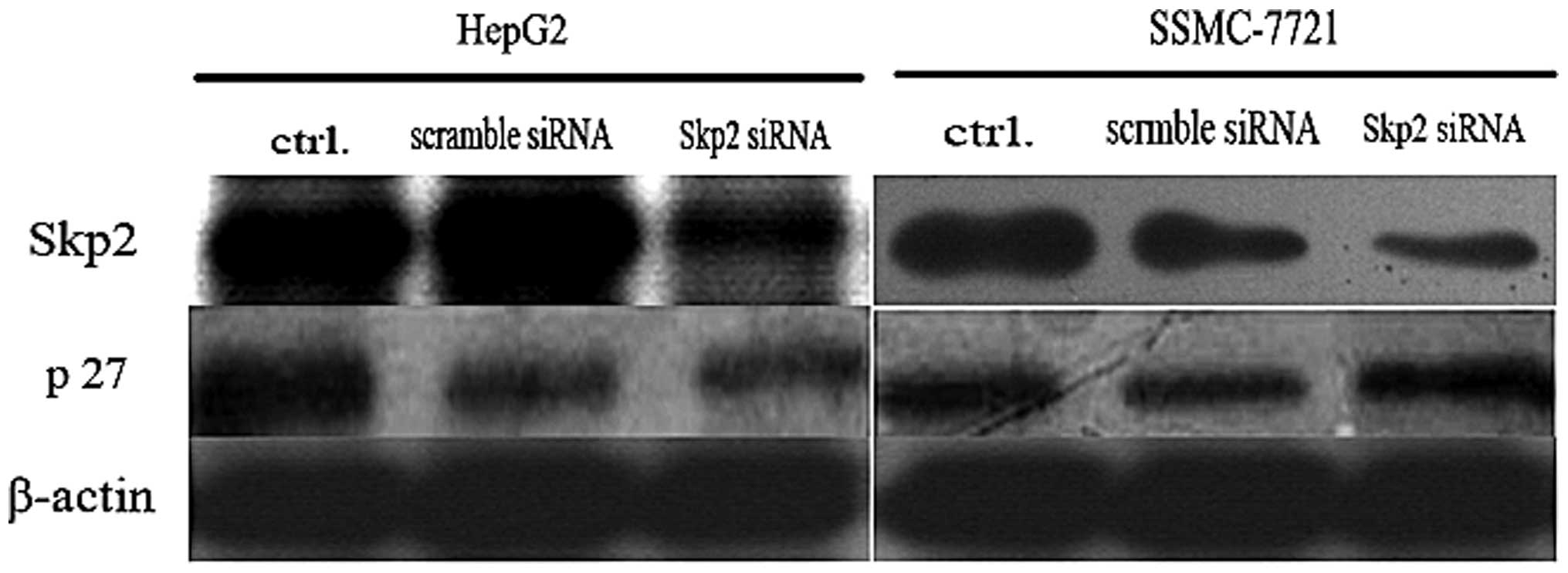
Interference of the expression of Skp2
increases the expression of p27kip1
The present study demonstrated that interference of
Skp2 induced apoptosis and inhibited proliferation of the SSMC-7721
cells. In order to further examine the mechanism involved and
clarify the association with the expression of p27kip1,
the expression of p27kip1 was examined by western blot
analysis. As shown in Fig. 6,
following depletion of the expression of endogenous Skp2 by RNA
interference, the levels of endogenous p27kip1 increased
in the HepG2 and SSMC-7721 cells suggesting that Skp2-mediated
degradation of p27kip1 was important for proliferation
of the tumor cells.
Discussion
Hepatocellular carcinoma is the fifth most common
type of malignant tumor worldwide and the third most common cause
of cancer-associated death (34,35).
Investigating the variation in oncogenes and suppressor genes
during hepatocarcinogenesis assists in identifying the prognostic
markers of the disease. Regulators controlling the cell cycle,
including the G1, S and M phases, regulate the progression and
development of several types of cancer, including liver cancer
(36,37). The present study investigated the
clinical predictive value of altered expression levels of
p27kip1 and Skp2 in hepatocellular carcinoma tissues
compared with normal liver tissues. The results revealed higher
expression levels of Skp2, often accompanied by lower expression
levels of p27kip1 in hepatocellular carcinoma, which
suggested that they may be important in the pathogenesis and
progression of liver cancer.
Skp2 is an F-box substrate-recognition subunit in
the SCF ubiquitin-protein ligase complex, which is important in
degrading the tumor suppressor gene p27kip1 by the
ubiquitin-proteasome system (38–42).
The present study demonstrated that the expression of
p27kip1 was negatively associated with that of Skp2,
which was detected by immunohistochemical and western blot
analysis. Additionally, the results also demonstrated that
interference of Skp2 by Skp2-siRNA significantly inhibited the
proliferation of the HepG2 and SSMC-7721 cells, which was detected
using an MTT assay and FACS analysis. Decreased levels of Skp2 are
usually accompanied by increased levels of p27kip1,
which are considered to be associated with the inhibition of cancer
cells (43,44). The results of the present study
demonstrated that inhibition of hepatocellular carcinoma by
interference of Skp2 was mainly dependent on upregulating the
protein expression of p27kip1.
P27kip1 degradation is mediated mainly by
the ubiquitin-proteasome system and Skp2 is an E3 ligase, one of
the major components of the SCF complex. This system specifically
recognizes the phosphorylated Thr187 of p27kip1 and
promotes the degradation of p27kip1 by ubiquitination
(30,45). When the protein levels of Skp2
decrease, less p27kip1 is recognized and degraded. Thus,
accumulation of the tumor inhibitor protein p27kip1 is
accompanied by downregulation of Skp2, which was consistent with
the results of the MTT and FACS assays in the present study.
In conclusion, the present study confirmed increased
expression of Skp2 accompanied by decreased expression of
p27kip1 in hepatocellular carcinoma, with the two
proteins having a negative correlation. Additionally, the RNA
interference technique may provide a novel approach for the therapy
and treatment of liver carcinoma.
References
|
1
|
Singh S, Fujii LL, Murad MH, et al: Liver
stiffness is associated with risk of decompensation, liver cancer,
and death in patients with chronic liver diseases: a systematic
review and meta-analysis. Clin Gastroenterol Hepatol. 11:1573–1584.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tanaka K, Ichikawa Y and Endo I: Liver
resection for advanced or aggressive colorectal cancer metastases
in the era of effective chemotherapy: a review. Int J Clin Oncol.
16:452–463. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Leenders MW, Nijkamp MW and Borel Rinkes
IH: Mouse models in liver cancer research: a review of current
literature. World J Gastroenterol. 14:6915–6923. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Coviello E, Caputi G, Martinelli D,
Germinario CA and Prato R: Mortality trends for primary liver
cancer in Puglia, Italy. Eur J Cancer Prev. 19:417–423. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shiraha H, Yamamoto K and Namba M: Human
hepatocyte carcinogenesis (review). Int J Oncol. 42:1133–1138.
2013.PubMed/NCBI
|
|
6
|
Baillargeon J, Snyder N, Soloway RD, et
al: Hepatocellular carcinoma prevalence and mortality in a male
state prison population. Public Health Rep. 124:120–126.
2009.PubMed/NCBI
|
|
7
|
Chen JG and Zhang SW: Liver cancer
epidemic in China: past, present and future. Semin Cancer Biol.
21:59–69. 2011. View Article : Google Scholar
|
|
8
|
Okuda K, Ohtsuki T, Obata H, et al:
Natural history of hepatocellular carcinoma and prognosis in
relation to treatment. Study of 850 patients. Cancer. 56:918–928.
1985. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lobry C, Oh P, Mansour MR, Look AT and
Aifantis I: Notch signaling: switching an oncogene to a tumor
suppressor. Blood. 123:2451–2459. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Elliman SJ, Howley BV, Mehta DS, Fearnhead
HO, Kemp DM and Barkley LR: Selective repression of the oncogene
cyclin D1 by the tumor suppressor miR-206 in cancers. Oncogenesis.
3:e1132014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu Z, Fu Q, Lv J, Wang F and Ding K:
Prognostic implication of p27Kip1, Skp2 and Cks1 expression in
renal cell carcinoma: a tissue microarray study. J Exp Clin Cancer
Res. 27:512008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Newbold A, Salmon JM, Martin BP, Stanley K
and Johnstone RW: The role of p21 and p27 in HDACi-mediated tumor
cell death and cell cycle arrest in the Eμ-myc model of B-cell
lymphoma. Oncogene. Dec 2–2013.(Epub ahead of print). View Article : Google Scholar
|
|
13
|
Gao J, Zhao Y, Lv Y, et al: Mirk/Dyrk1B
mediates G0/G1 to S phase cell cycle progression and cell survival
involving MAPK/ERK signaling in human cancer cells. Cancer Cell
Int. 13:22013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wei W, Ayad NG, Wan Y, et al: Degradation
of the SCF component Skp2 in cell-cycle phase G1 by the
anaphase-promoting complex. Nature. 428:194–198. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bashir T, Dorrello NV, Amador V,
Guardavaccaro D and Pagano M: Control of the SCF (Skp2-Cks1)
ubiquitin ligase by the APC/C(Cdh1) ubiquitin ligase. Nature.
428:190–193. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen G, Wang Y, Garate M, Zhou J and Li G:
The tumor suppressor ING3 is degraded by SCF(Skp2)-mediated
ubiquitin-proteasome system. Oncogene. 29:1498–1508. 2010.
View Article : Google Scholar
|
|
17
|
Cheng H, Meng J, Wang G, et al: Skp2
regulates subcellular localization of PPARgamma by MEK signaling
pathways in human breast cancer. Int J Mol Sci. 14:16554–16569.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang Z, Gao D, Fukushima H, et al: Skp2: a
novel potential therapeutic target for prostate cancer. Biochim
Biophys Acta. 1825:11–17. 2012.
|
|
19
|
Wei Z, Jiang X, Qiao H, et al: STAT3
interacts with Skp2/p27/p21 pathway to regulate the motility and
invasion of gastric cancer cells. Cell Signal. 25:931–938. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pateras IS, Apostolopoulou K, Koutsami M,
et al: Downregulation of the KIP family members p27 (KIP1) and p57
(KIP2) by SKP2 and the role of methylation in p57 (KIP2)
inactivation in nonsmall cell lung cancer. Int J Cancer.
119:2546–2556. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Saleem M, Maddodi N, Abu Zaid M, et al:
Lupeol inhibits growth of highly aggressive human metastatic
melanoma cells in vitro and in vivo by inducing apoptosis. Clin
Cancer Res. 14:2119–2127. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Adhami VM, Siddiqui IA, Ahmad N, Gupta S
and Mukhtar H: Oral consumption of green tea polyphenols inhibits
insulin-like growth factor-I-induced signaling in an autochthonous
mouse model of prostate cancer. Cancer Res. 64:8715–8722. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nishitani H, Sugimoto N, Roukos V, et al:
Two E3 ubiquitin ligases, SCF-Skp2 and DDB1-Cul4, target human Cdt1
for proteolysis. EMBO J. 25:1126–1136. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Peng L, Xu Z, Zhou Y, et al: Effect of
rosiglitazone on cells cycle, apoptosis and expression of Skp2 and
p27Kip1 in hepatocellular carcinoma cell line. Zhonghua Gan Zang
Bing Za Zhi. 18:148–149. 2010.(In Chinese). PubMed/NCBI
|
|
25
|
Schulman BA, Carrano AC, Jeffrey PD, et
al: Insights into SCF ubiquitin ligases from the structure of the
Skp1–Skp2 complex. Nature. 408:381–386. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li Y, Huang W, Huang S, Du J and Huang C:
Screening of anti-cancer agent using zebrafish: comparison with the
MTT assay. Biochem Biophys Res Commun. 422:85–90. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sarzaeem A, Zare Mirakabadi A, Moradhaseli
S, Morovvati H and Lotfi M: Cytotoxic effect of ICD-85
(venom-derived peptides) on HeLa cancer cell line and normal LK
cells using MTT assay. Arch Iran Med. 15:696–701. 2012.PubMed/NCBI
|
|
28
|
Sylvester PW: Optimization of the
tetrazolium dye (MTT) colorimetric assay for cellular growth and
viability. Methods Mol Biol. 716:157–168. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu Y, Wang Y, Cheng C, et al: A
relationship between p27 (kip1) and Skp2 after adult brain injury:
implications for glial proliferation. J Neurotrauma. 27:361–371.
2010. View Article : Google Scholar
|
|
30
|
Rosner M and Hengstschläger M: Tuberin
binds p27 and negatively regulates its interaction with the SCF
component Skp2. J Biol Chem. 279:48707–48715. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hu D, Liu W, Wu G and Wan Y: Nuclear
translocation of Skp2 facilitates its destruction in response to
TGFbeta signaling. Cell Cycle. 10:285–292. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Foster JS, Fernando RI, Ishida N, Nakayama
KI and Wimalasena J: Estrogens down-regulate p27Kip1 in breast
cancer cells through Skp2 and through nuclear export mediated by
the ERK pathway. J Biol Chem. 278:41355–41366. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pavlides SC, Huang KT, Reid DA, et al:
Inhibitors of SCF-Skp2/Cks1 E3 ligase block estrogen-induced growth
stimulation and degradation of nuclear p27kip1: therapeutic
potential for endometrial cancer. Endocrinology. 154:4030–4045.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kudo M: Targeted therapy for liver cancer:
updated review in 2012. Curr Cancer Drug Targets. 12:1062–1072.
2012.PubMed/NCBI
|
|
35
|
Montalvo-Jave EE, Villegas-Alvarez F,
Montalvo-Arenas CE, et al: Liver transplantation: some advances in
liver cancer, live liver donation, and cell transplantation. A
literature review. Rev Gastroenterol Mex. 74:341–348. 2009.
|
|
36
|
Shanmugasundaram K, Block K, Nayak BK, et
al: PI3K regulation of the SKP-2/p27 axis through mTORC2. Oncogene.
32:2027–2036. 2013. View Article : Google Scholar :
|
|
37
|
Assoian RK and Yung Y: A reciprocal
relationship between Rb and Skp2: implications for restriction
point control, signal transduction to the cell cycle and cancer.
Cell Cycle. 7:24–27. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bashir T, Pagan JK, Busino L and Pagano M:
Phosphorylation of Ser72 is dispensable for Skp2 assembly into an
active SCF ubiquitin ligase and its subcellular localization. Cell
Cycle. 9:971–974. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Calvisi DF, Pinna F, Ladu S, et al: The
degradation of cell cycle regulators by SKP2/CKS1 ubiquitin ligase
is genetically controlled in rodent liver cancer and contributes to
determine the susceptibility to the disease. Int J Cancer.
126:1275–1281. 2010.
|
|
40
|
Wei Z, Jiang X, Liu F, et al:
Downregulation of Skp2 inhibits the growth and metastasis of
gastric cancer cells in vitro and in vivo. Tumour Biol. 34:181–192.
2013. View Article : Google Scholar
|
|
41
|
Wu J, Lee SW, Zhang X, et al: Foxo3a
transcription factor is a negative regulator of Skp2 and Skp2 SCF
complex. Oncogene. 32:78–85. 2013. View Article : Google Scholar :
|
|
42
|
Wu L, Grigoryan AV, Li Y, et al: Specific
small molecule inhibitors of Skp2-mediated p27 degradation. Chem
Biol. 19:1515–1524. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zheng XY, Ding W, Xie LP and Chen ZD:
Correlation of Skp2 and P27kip1 protein expression and
clinicopathological features of prostate cancer. Ai Zheng.
23:215–218. 2004.(In Chinese). PubMed/NCBI
|
|
44
|
Ben-Izhak O, Lahav-Baratz S, Meretyk S, et
al: Inverse relationship between Skp2 ubiquitin ligase and the
cyclin dependent kinase inhibitor p27Kip1 in prostate cancer. J
Urol. 170:241–245. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Radke S, Pirkmaier A and Germain D:
Differential expression of the F-box proteins Skp2 and Skp2B in
breast cancer. Oncogene. 24:3448–3458. 2005. View Article : Google Scholar : PubMed/NCBI
|