Introduction
Lung cancer is one of the leading causes of
cancer-related mortality (1).
Non-small cell lung cancer (NSCLC) accounts for up to 85% of lung
cancer cases and is usually resistant to chemotherapy (2). Cancer cells in the lung experience
mechanical forces associated with normal breathing as well as
altered mechanical environments due to pre-existing lung disease or
tumor-associated changes. Recent studies have suggested that the
mechanical environment of cancerous tumors is involved in tumor
formation and metastasis (3,4).
Extracellular matrix (ECM) rigidity, which provides
biophysical cues from the microenvironment, has been shown to have
an effect on a number of aspects of tumor cell behavior, including
adhesion, migration, proliferation and invasion. When cells undergo
transformation and initiate the formation of a solid tumor mass,
they cause profound changes in the phenotypes of the cells that
surround them (5). It has been
demonstrated that the majority of solid tumors have increased ECM
stiffness, which may lead to increased activation of protumor
signaling pathways, such as focal adhesion kinase (FAK) and Ras
homolog gene family, member A (RhoA) pathways (6,7). The
transcription coactivator Yes-associated protein (YAP) is an
important downstream effector of the Hippo pathway (8). Investigation into the role of YAP is
a burgeoning field of research, with the focus on transcriptional
regulation of a number of aspects of cell behavior. It has been
shown that YAP and transcriptional coactivator with PDZ-binding
motif (TAZ) are regulators of the nuclear transduction of physical
cues in mesenchymal stem cells (9,10)
and that increased matrix stiffness may also result in increased
activity of the oncogenic YAP/TAZ complex (11). It is therefore evident that NSCLC
may develop in an environment with mechanical properties distinct
from those encountered in the normal lung.
Despite the growing evidence suggesting that YAP is
a crucial regulator of human cancer (11), its involvement in the development
of NSCLC in response to mechanical cues remains unclear. In the
present study, substrates with different rigidity were generated in
order to evaluate the role of YAP, and its upstream regulators in
the Hippo pathway, in the regulation of NSCLC cell growth. It was
hypothesized that YAP may be a bridging molecule between NSCLC
growth and the mechanical microenvironment. The findings
demonstrate that YAP may regulate the growth of NSCLC in response
to matrix stiffness, thereby suggesting a role of the Hippo-YAP
pathway in NSCLC growth in response to its physical
microenvironment.
Materials and methods
Generation of substrates with different
rigidity
Substrates with modifiable mechanical properties
were generated on variably compliant polyacrylamide (PA) gels,
according to a previously established method (12). Briefly, glass slides were treated
with 3-aminopropyltrimethoxysilane and 0.5% glutaraldehyde. PA gel
solutions with the desired concentrations of acrylamide and
bis-acrylamide were allowed to polymerize. The elastic modulus of
the PA gel substrates was controlled by adjusting the quantity of
acrylamide and bis-acrylamide crosslinker, as previously described
(13). Sulfo-SANPAH (Proteochem,
Denver, CO, USA) was used to link fibronectin (100 μg/ml;
Sigma, St. Louis, MO, USA) to the PA gel surface. The PA gel
modulus of elasticity was quantified using atomic force microscopy
(AFM, Bruker, Billerica, MA, USA). All substrates were sterilized
using UV light.
Cell culture
The SPCA-1 human non-small cell lung cancer cell
line was purchased from the American Type Culture Collection
(Manassas, VA, USA) and maintained in RPMI-1640 medium
(Sigma-Aldrich, St. Louis, MO, USA), supplemented with 10% fetal
bovine serum, 100 U/ml penicillin, and 100 U/ml streptomycin
(Sigma-Aldrich) at 37°C in 5% CO2 and 95%
relative humidity. The cells were then cultured on PA substrates
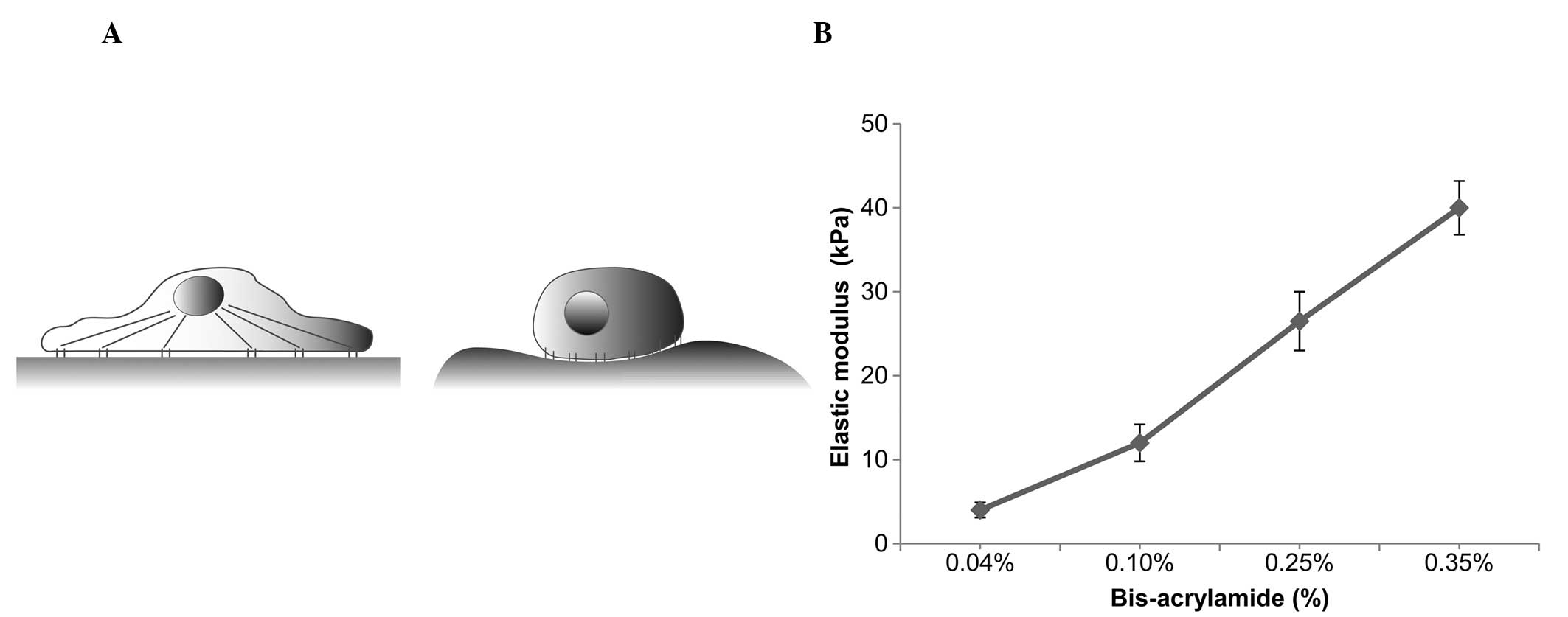
for up to 48 h (Fig. 1A). A
density of 1×105 cells/cm2 was used to seed
the substrates. The proliferation rates were determined by
quantifying the cells using a hemocytometer subsequent to
trypsinization in each condition. Growth curves for the cells were
then determined.
Small-interfering RNA (siRNA)
transfection of YAP
siRNAs (siControl and siYAP) specific for the target
genes were purchased from Santa Cruz Biotechnology, Inc. (Santa
Cruz, CA, USA). For the knockdown of YAP, the corresponding siRNAs
were delivered into SPCA-1 cells using Lipofectamine®
RNAiMAX (Invitrogen Life Technologies, Carlsbad, CA, USA) according
to the manufacturer’s instructions.
Immunofluorescence staining
Samples were fixed with 4% paraformaldehyde for 15
min and then permeabilized with 0.1% Triton X-100 for 10 min. After
washing with phosphate-buffered saline (PBS) three times, the
samples were blocked with normal goat serum (Invitrogen Life
Technologies) for 30 min to prevent nonspecific binding, incubated
with polyclonal rabbit primary antibodies against YAP (sc-15407;
Santa Cruz Biotechnology, Inc.) at 4°C overnight and then
incubated with goat anti-rabbit IgG fluorescein isothiocyanate
(FITC)-conjugated secondary antibodies (sc-2012; Santa Cruz
Biotechnology, Inc.) for 1 h. After staining, the samples were
washed with PBS two to three times and imaged using fluorescence
microscopy (Olympus IX71; Olympus, Tokyo, Japan). The percentage of
cells with YAP expression localized to the nucleus (nuclear YAP) in
10 different visual fields for three replicates. As in the method
described previously (14),
distribution patterns of Yap expression in the cells were
classified as nuclear (nuclear expression>cytoplasmic
expression), diffuse (nuclear expression=cytoplasmic expression)
and cytoplasmic (nuclear expression<cytoplasmic expression). In
the current study, only YAP distribution localized to the nucleus
(nuclear expression>cytoplasmic expression) was deemed as
‘nuclear YAP localization’; the ‘diffuse’ (nuclear and cytoplasmic
expression) and ‘cytoplasmic’ (nuclear<cytoplasmic expression)
cells were not included in the criteria.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RT-qPCR was used to analyze the transcriptional
levels of Survivin, connective tissue growth factor (CTGF),
amphiregulin (AREG) and Ki67. RT-qPCR was performed using an
M×3000P qPCR system (Agilent Technologies, Santa Clara, CA, USA)
with SYBR® Premix Ex Taq™ II (Takara Bio Inc., Shiga,
Japan). β-actin was used as the internal control gene to normalize
the levels of target gene expression. Thermocycling conditions were
applied as follows: 95°C for 30 sec, 40 cycles of
denaturation (95°C for 5 sec), annealing (60°C for 30 sec)
and extension (72°C for 30 sec). The primers used for the
RT-qPCR assays are listed in Table
1. The data were analyzed using the 2−∆∆Ct
method.
 | Table IPrimer sequences used for RT-qPCR. |
Table I
Primer sequences used for RT-qPCR.
| Gene | PCR primer sequences
(forward and reverse) | GenBank accession
no. |
|---|
| Survivin | Forward:
5′-AGGACCACCGCATCTCTACAT-3′
Reverse: 5′-AAGTCTGGCTCGTTCTCAGTG-3′ | NM_001012270 |
| CTGF | Forward:
5′-ACCGACTGGAAGACACGTTTG-3′
Reverse: 5′-CCAGGTCAGCTTCGCAAGG-3′ | NM_001901 |
| AREG | Forward:
5′-CTGGGAAGCGTGAACCATTTT-3′
Reverse: 5′-TCTGAGTAGTCATAGTCGGCTC-3′ | NM_001657 |
| Ki67 | Forward:
5′-ACGAGACGCCTGGTTACTATC-3′
Reverse: 5′-GCTCATCAATAACAGACCCATTTAC-3′ | NM_002417 |
| β-actin | Forward:
5′-CATGTACGTTGCTATCCAGGC-3′
Reverse: 5′-CTCCTTAATGTCACGCACGAT-3′ | NM_001101 |
Western blot analysis
Cells were lysed in lysis buffer containing protease
and phosphatase inhibitors (Sigma-Aldrich). Proteins were separated
by sodium dodecyl sulfate-polyacrylamide gel electrophoresis,
electroblotted onto polyvinylidene fluoride membranes
(Sigma-Alrdich), blocked with 5% dry milk in Tris-buffered saline
with Tween-20, immunoblotted with specific primary antibodies and
horseradish peroxidase-conjugated secondary antibodies and detected
by enhanced chemiluminescence (Pierce Chemical Co., Rockford, IL,
USA). The following rabbit polyclonal IgG antibodies were used (all
from Santa Cruz Biotechnology, Inc. unless otherwise stated):
Anti-YAP (sc-15407), anti-p-YAP (#13008; Cell Signaling Technology,
Inc., Danvers, MA, USA), anti-LATS (sc-28223), anti-CTGF
(sc-25440), anti-AREG (sc-25436) anti-Survivin (sc-10811), and
anti-Ki67 (sc-15402), and anti-β-actin (ab1801; Abcam, Cambridge,
MA, USA). The secondary antibody used was the goat anti-rabbit
horseradish peroxidase-conjugated IgG H&L (ab97051; Abcam).
Statistical analysis
At least three replicates were performed in each
experiment. Comparison between pairs of groups was performed using
the two-samples t-test. SPSS version 13.0 (SPSS, Inc., Chicago, IL,
USA) software was used for statistical analysis. P<0.05 was
considered to indicate a statistically significant difference.
Results
Matrix elasticity increases with an
increase in bis-acrylamide
To control the substrate stiffness,
fibronectin-coated PA gels with various degrees of stiffness were
generated by adjusting the relative quantities of acrylamide and
bis-acrylamide crosslinker. When maintaining acrylamide at a
constant concentration of 10%, the PA gels with 0.04, 0.10, 0.25
and 0.35% bis-acrylamide ratios yielded elasticities of 4±0.9,
12±2.2, 26.5±3.5 and 40±3.2 kPa, respectively (mean±standard
deviation; Fig. 1B).
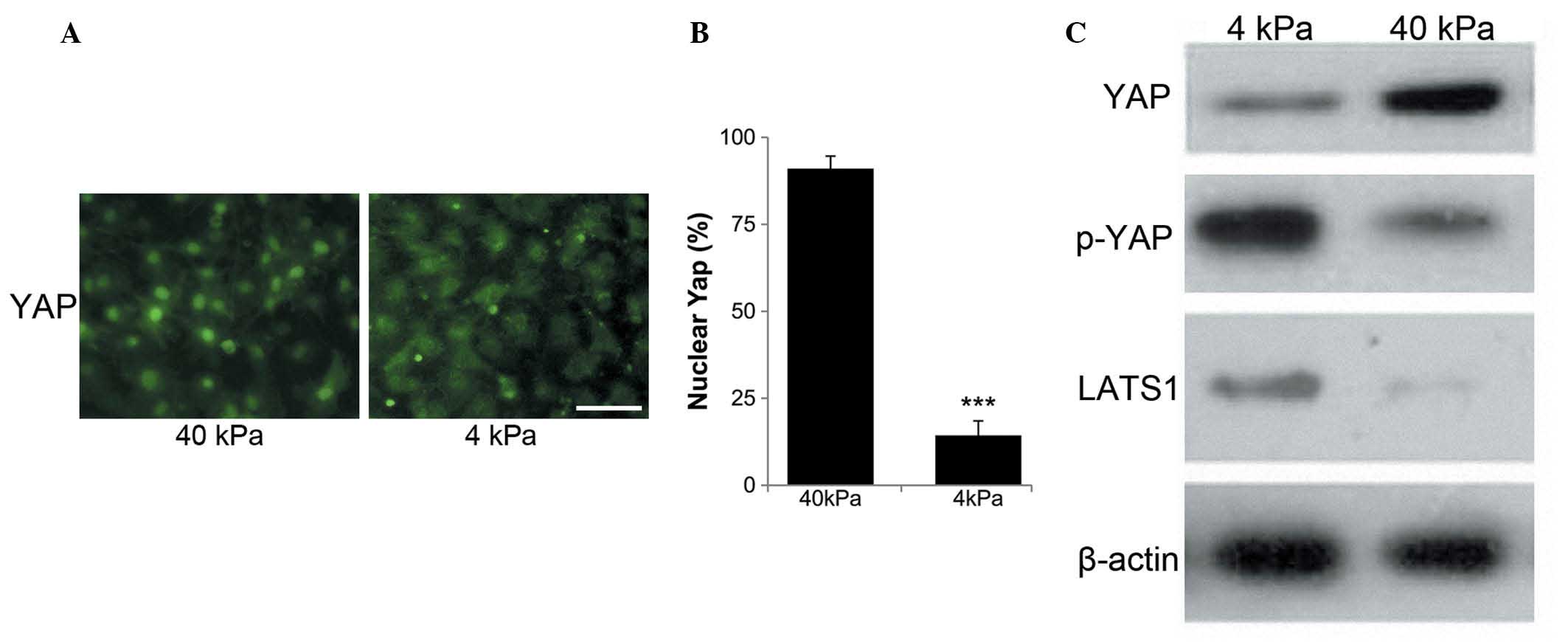
Substrate stiffness correlates with YAP
and LATS1 kinase expression in NSCLC cells
The SPCA-1 human NSCLC cells were seeded on soft (4
kPa) or stiff (40 kPa) substrates for up to two days.
Immunofluorescent staining showed that YAP expression in SPCA-1
cells was largely localized to the nucleus when seeded on the
40-kPa substrate, but was predominantly cytoplasmic on the 4-kPa
substrate (Fig. 2A). The
percentage of cells with nuclear YAP expression was significantly
lower on the 4-kPa substrate than the 40-kPa substrate (Fig. 2B). In addition, expression of the
YAP upstream regulators in the Hippo pathway in response to
changing substrate stiffness was investigated. The protein
expression levels of YAP, p-YAP and LATS1 kinase in SPCA-1 human
NSCLC cells in response to substrate stiffness, were analyzed by
western blotting following two days of culturing (Fig. 2C). The results showed that levels
of LATS1 kinase and p-YAP were extremely low in cells on the stiff
substrate (40 kPa) but were markedly increased in cells on the soft
substrate (4 kPa). Thus, expression of the YAP protein
significantly increased after culturing on the stiff substrate,
whereas expression of p-YAP and LATS1 protein markedly decreased
after culturing on the stiff substrate (Fig. 2C).
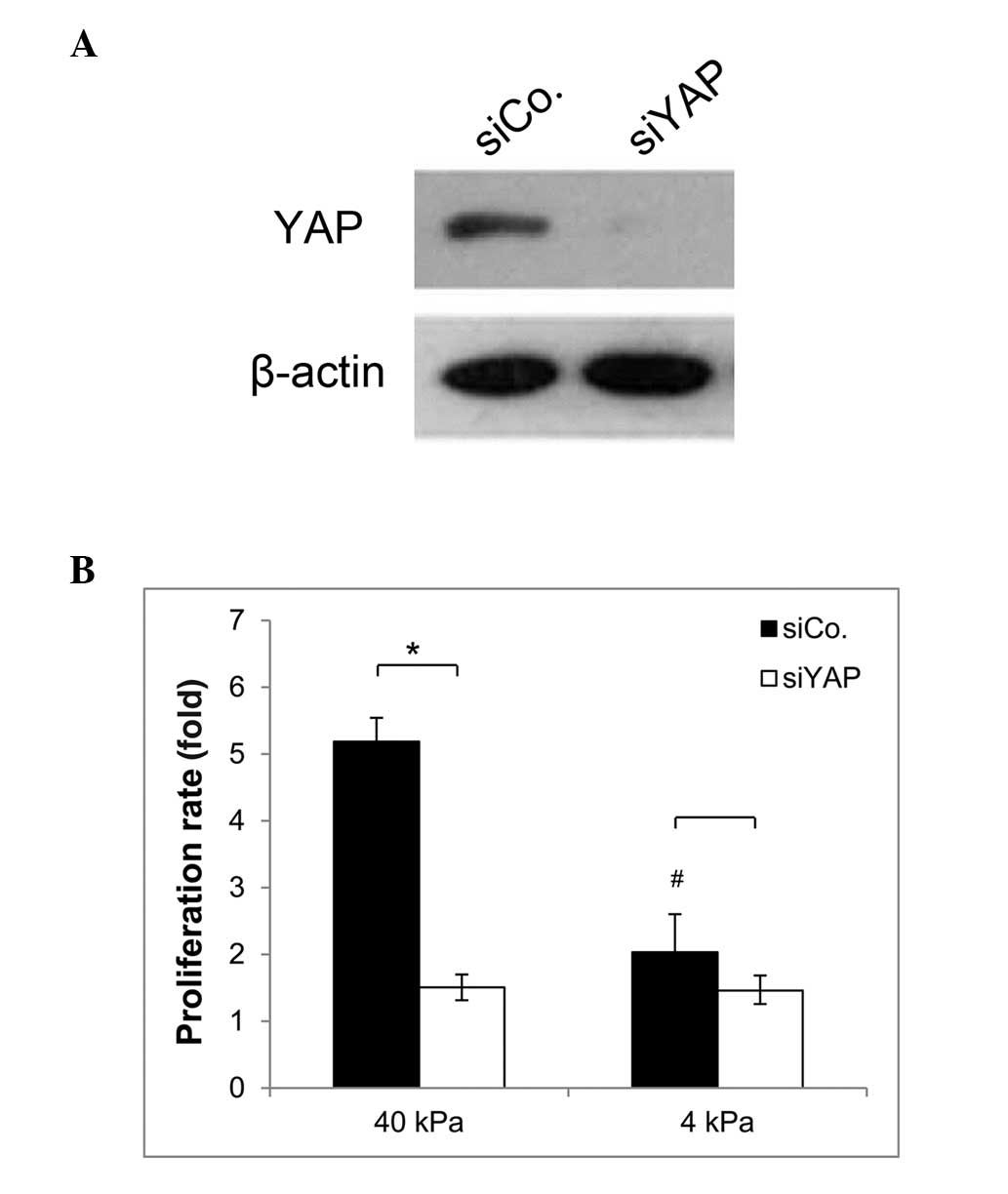
Higher levels of YAP expression and
culturing on a stiffer substrate lead to increased proliferation of
SPCA-1 NSCLC cells
To evaluate the function of YAP in SPCA-1 NSCLC
cells, a knockdown of YAP in these cells was performed. Cells were
then cultured on substrates of different stiffness. As shown in
Fig. 3A, efficient knockdown of
YAP endogenous protein was achieved. The proliferation rates of YAP
knockdown cells and control cells were calculated after two days of
culture by counting. As shown in Fig.
3B, the proliferation rate of the YAP knockdown cells decreased
significantly compared with that of the control cells on the stiff
substrate. However, there was no statistical difference in the
proliferation rate between the YAP knockdown cells and the YAP
control cells when cultured on the soft substrate. Moreover, the
proliferation rate of SPCA-1 cells expressing YAP was significantly
decreased when plated on the soft substrate, compared with cells
plated on the stiff substrate. These findings indicate that
increasing matrix stiffness promotes SPCA-1 NSCLC cell
proliferation, and that the higher levels of YAP expression in
cells grown on a stiff substrate, may in part be the cause of this
effect.
CTGF, AREG, Survivin and Ki67 expression
alters in response to substrate stiffness and the presence of
YAP
To further investigate whether YAP expression in
NSCLC, in response to substrate stiffness, is correlated with cell
proliferation, the mRNA and protein expression levels of CTGF,
AREG, Survivin and Ki-67 (cellular proliferation markers) were
examined under the different conditions. As shown in Fig. 4, it was found that when culturing
on the stiff substrate, the YAP knockdown cells displayed
relatively low mRNA expression levels of CTGF, AREG, Survivin and
Ki67, compared with the control cells. However, when cells were
cultured on the soft substrate, mRNA expression levels of CTGF and
Survivin between the knockdown cells and the control cells were not
significantly different. For the YAP control cells, the mRNA
expression levels of CTGF, AREG, Survivin and Ki67 on the soft
substrate were significantly decreased compared with the levels
following culture on the stiff substrate.
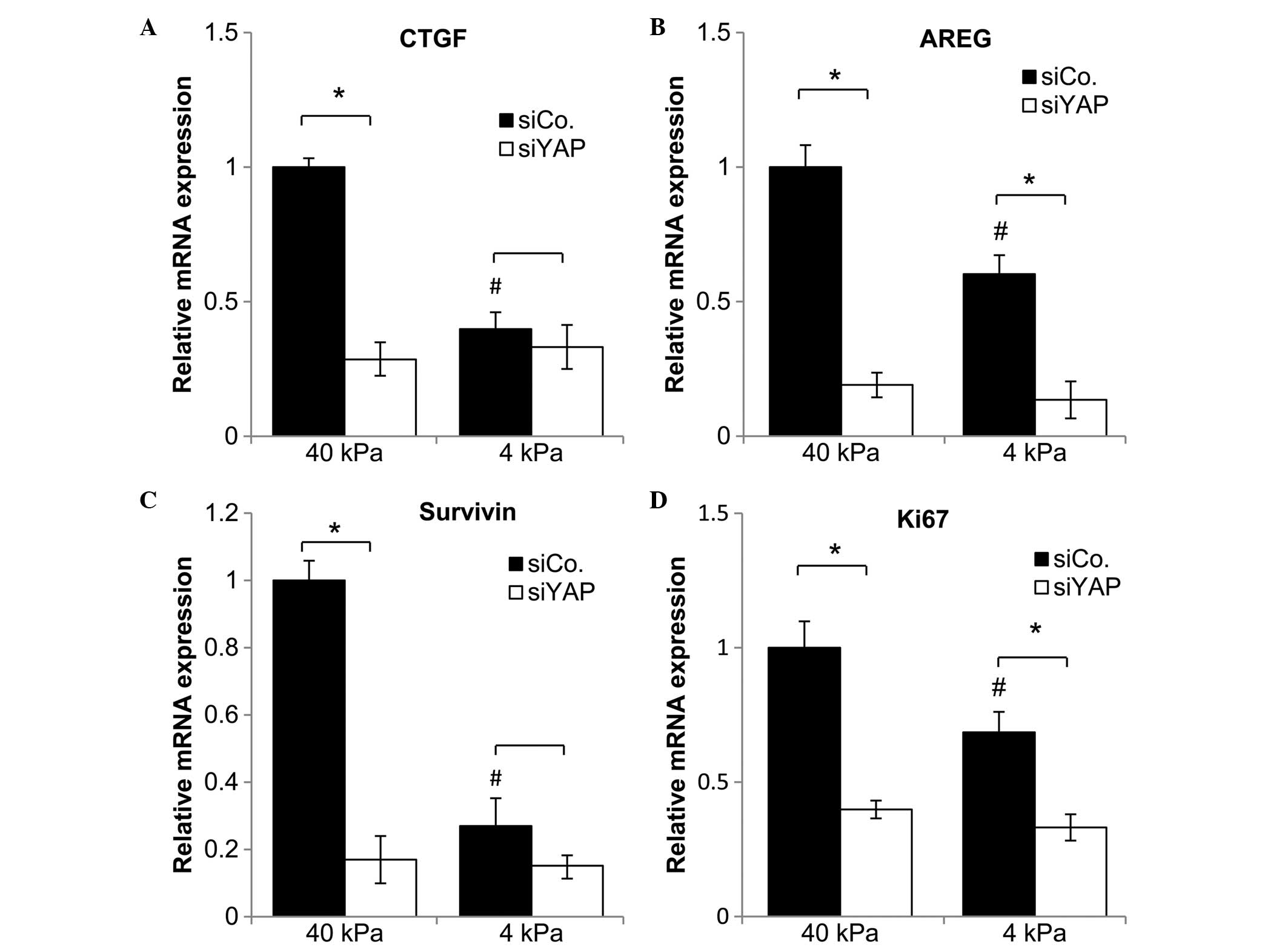 | Figure 4Gene expression levels of (A) CTGF,
(B) AREG, (C) Survivin and (D) Ki67 in NSCLC cells, with or without
YAP knockdown, after two days of culture on a soft substrate (4
kPa) or a stiff substrate (40 kPa). Data represent the mean ±
standard error of the mean. *P<0.05 compared with
siControl. #P<0.05, cells plated on the 4 kPa substrate compared
with cells plated on the 40 kPa substrate. CTGF, connective tissue
growth factor; AREG, amphiregulin; NSCLC, non-small cell lung
cancer; YAP, Yes-associated protein; siCo, small interfering RNA
control; siYAP, small interfering YAP RNA. |
As shown in Fig. 5,
the protein expression levels of CTGF, AREG, Survivin and Ki67 in
the YAP knockdown cells were decreased compared with those of
control cells on the stiff and soft substrates. Furthermore,
protein expression of CTGF, Survivin and Ki67 in SPCA-1 NSCLC cells
on the soft substrate also decreased markedly compared with those
cultured on the stiff substrate.
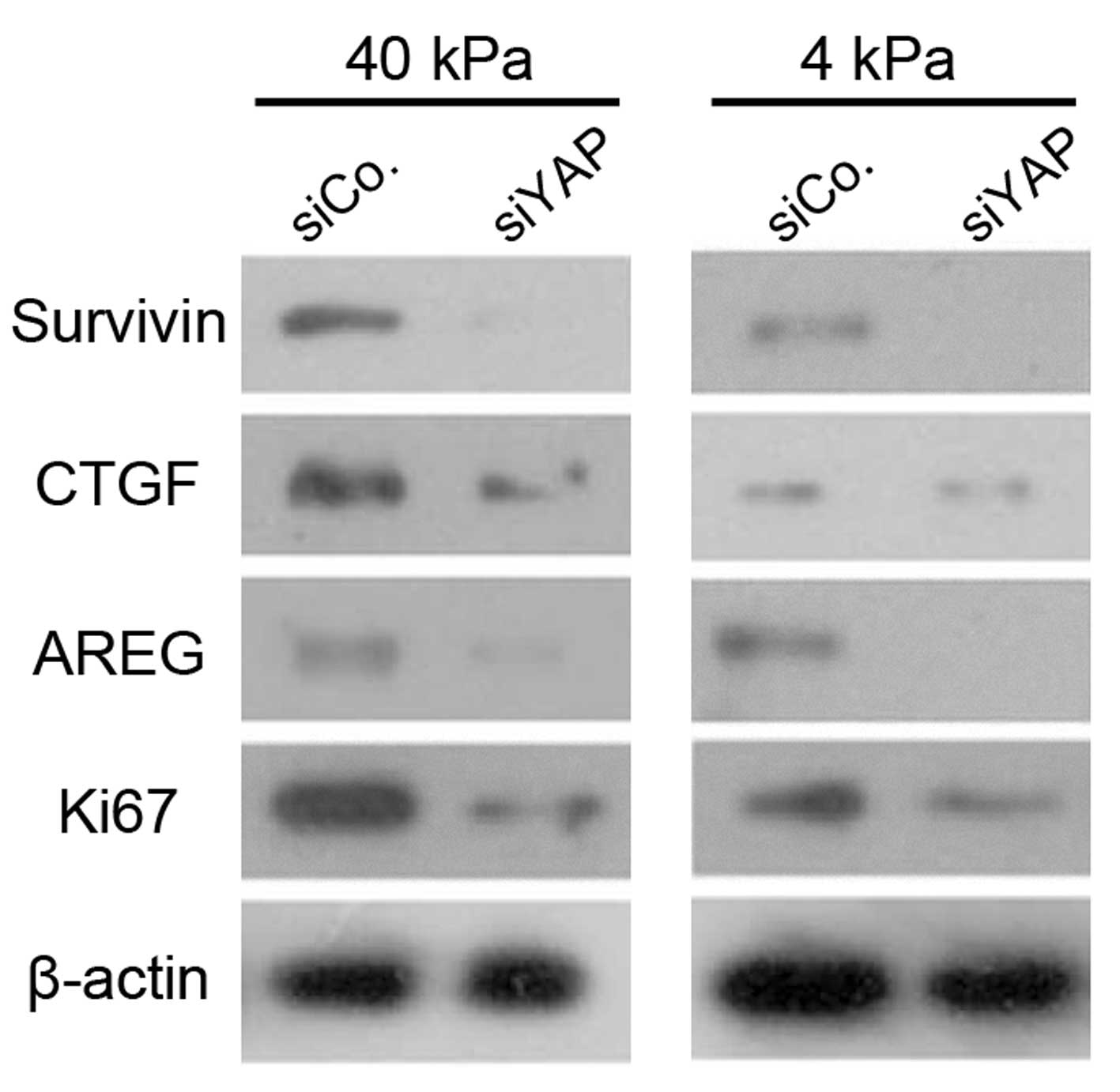 | Figure 5Western blot showing protein
expression levels of CTGF, AREG, Survivin and Ki67 in NSCLC cells,
with or without YAP knockdown, after two days of culture on a soft
substrate (4 kPa) or a stiff substrate (40 kPa). Data represent the
mean ± standard error of the mean. *P<0.05, compared
with siControl. CTGF, connective tissue growth factor; AREG,
amphiregulin; NSCLC, non-small cell lung cancer, YAP,
Yes-accosiated protein; SiCon, small interfering RNA control;
siYAP, small interfering YAP RNA. |
Discussion
The Hippo pathway is an evolutionarily conserved
regulator of tissue growth and cell fate (11). The transcription coactivator, YAP,
is inhibited by the LATS 1/2 kinases and is a major downstream
effector of the Hippo pathway (15). In this study, the role of YAP, and
its upstream regulators in the Hippo pathway, in the regulation of
NSCLC cell growth on substrates with different stiffnesses was
evaluated. The results support the hypothesis that YAP is a
bridging molecule between the growth of SPCA-1 NSCLC cells and the
mechanical microenvironment. The findings demonstrate that the
stiffness of the subcellular matrix alters the behavior of NSCLC
cells in vitro, and that YAP can regulate the growth of
NSCLC cells in response to matrix stiffness.
It has previously been demonstrated that mechanical
loading can trigger proliferation of tumor cells (16). This implies that signaling pathways
that are sensitive to the mechanical properties of their
environment may modulate tumor cell behavior. More recently, a
study demonstrated that ECM rigidity, cell tension and changes in
cell geometry mediate cell contractility, which in turn activates a
YAP-dependent mechanoresponse independently of the Hippo cascade
(9). It was shown that upon
YAP/TAZ depletion, the proliferation rate and the expression levels
of cellular proliferation markers in cells plated on a stiff
substrate behaved as if they had been plated on a soft substrate.
In addition, previous data indicated that the interaction between
YAP and TEA domain protein (TEAD) is an integral part of the
mechano-tuning mechanism that permits fibroblasts to increase their
actomyosin contractility in response to elevated ECM stiffness
(17). The current study extended
these findings to NSCLC cells. It was shown that a hard substrate
promoted the growth of NSCLC cells in vitro, as well as an
increase in the transcription levels of CTGF, AREG, Survivin and
Ki67. This was associated with decreased cytoplasmic expression of
YAP and increased nuclear expression, as well as downregulation of
p-YAP expression. The results indicated that YAP regulates the
proliferation of NSCLC cells in response to the stiffness of the
surrounding environment.
Furthermore, CTGF, AREG and Survivin have been
identified as potential transcriptional targets of YAP, required
for cell growth and differentiation (18,19).
Thus, ECM rigidity may influence the expression of CTGF, AREG,
Survivin and Ki67 by mediating the expression patterns of YAP,
thereby impacting the growth of NSCLC cells. Furthermore, TEAD is
also required for YAP-induced cell growth (18), and therefore may also be involved
in the stiffness-mediated regulation of NSCLC cell
proliferation.
A previous study found that mechanical loading
promoted Lewis lung cancer cell growth through induction of
periostin expression in the mouse (20). However, whether periostin function
correlates with YAP-mediated cell growth in the mechanical
microenvironment requires further investigation. In addition,
although this study found a positive association between YAP
expression and Ki-67 expression (a marker for cell proliferation)
in NSCLC cells, the precise mechanisms that are involved in the
oncogenic processes of NSCLC development in response to changes in
the mechanical microenvironment require further investigation. It
is possible that YAP function changes depending on the upstream
input, and on variations in the repertoire of binding partners
between cells.
This ability of the Hippo pathway to respond to
mechanical cues may partially underlie the hyperactivation of YAP
and TAZ that has been observed in human tumors, since tumor tissue
has altered mechanical properties due to extracellular matrix
modifications and the infiltration of stromal and immune cells,
which render it more rigid (21).
This may also explain why NSCLC tumor tissue is stiffer than the
surrounding tissue.
In conclusion, the findings suggest that YAP may be
a bridging molecule between NSCLC growth and its mechanical
microenvironment, and that it may regulate the growth of NSCLC in
response to matrix stiffness. This study provides novel insight
into how the growth of NSCLC cells is mediated by physical cues.
Modulation of proliferation and apoptosis, controlled by the
Hippo-Yap signaling pathway, in lung cancer cells presents an
opportunity to develop novel drugs, which may be successfully
combined with conventional chemotherapy and radiotherapy treatment
strategies.
References
|
1
|
Jemal A, Siegel R, Ward E, et al: Cancer
statistics, 2007. CA Cancer J Clin. 57:43–66. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Schiller JH, Harrington D, Belani CP, et
al: Comparison of four chemotherapy regimens for advanced
non-small-cell lung cancer. New Engl J Med. 346:92–98. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Menon S and Beningo KA: Cancer cell
invasion is enhanced by applied mechanical stimulation. PLoS One.
6:e172772011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tse JM, Cheng G, Tyrrell JA, et al:
Mechanical compression drives cancer cells toward invasive
phenotype. Proc Natl Acad Sci USA. 109:911–916. 2012. View Article : Google Scholar :
|
|
5
|
Boudreau A, van’t Veer LJ and Bissell MJ:
An ̔elite hacker̓: breast tumors exploit the normal
microenvironment program to instruct their progression and
biological diversity. Cell Adh Migr. 6:236–248. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Guilluy C, Swaminathan V, Garcia-Mata R,
et al: The Rho GEFs LARG and GEF-H1 regulate the mechanical
response to force on integrins. Nat Cell Biol. 13:722–727. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Levental KR, Yu H, Kass L, et al: Matrix
crosslinking forces tumor progression by enhancing integrin
signaling. Cell. 139:891–906. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dong J, Feldmann G, Huang J, et al:
Elucidation of a universal size-control mechanism in Drosophila and
mammals. Cell. 130:1120–1133. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dupont S, Morsut L, Aragona M, et al: Role
of YAP/TAZ in mechanotransduction. Nature. 474:179–183. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Halder G, Dupont S and Piccolo S:
Transduction of mechanical and cytoskeletal cues by YAP and TAZ.
Nat Rev Mol Cell Biol. 13:591–600. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Harvey KF, Zhang X and Thomas DM: The
Hippo pathway and human cancer. Nat Rev Cancer. 13:246–257. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Park JS, Chu JS, Tsou AD, et al: The
effect of matrix stiffness on the differentiation of mesenchymal
stem cells in response to TGF-β. Biomaterials. 32:3921–3930. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tse JR and Engler AJ: Preparation of
hydrogel substrates with tunable mechanical properties. Current
Protoc Cell Biol. Chapter 10: Unit 10. 162010.
|
|
14
|
Wada K, Itoga K, Okano T, et al: Pathway
regulation by cell morphology and stress fibers. Development.
138:3907–3914. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yu FX, Zhao B, Panupinthu N, et al:
Regulation of the Hippo-YAP pathway by G-protein-coupled receptor
signaling. Cell. 150:780–791. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hofmann M, Guschel M, Bernd A, et al:
Lowering of tumor interstitial fluid pressure reduces tumor cell
proliferation in a xenograft tumor model. Neoplasia. 8:89–95. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Calvo F, Ege N, Grande-Garcia A, et al:
Mechanotransduction and YAP-dependent matrix remodelling is
required for the generation and maintenance of cancer-associated
fibroblasts. Nat Cell Biol. 15:637–646. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao B, Ye X, Yu J, et al: TEAD mediates
YAP-dependent gene induction and growth control. Genes Dev.
22:1962–1971. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huang JM, Nagatomo I, Suzuki E, et al: YAP
modifies cancer cell sensitivity to EGFR and survivin inhibitors
and is negatively regulated by the non-receptor type protein
tyrosine phosphatase 14. Oncogene. 32:2220–2229. 2013. View Article : Google Scholar
|
|
20
|
Ma D, Lu H, Xu L, et al: Mechanical
loading promotes Lewis lung cancer cell growth through periostin.
In Vitro Cell Dev Biol Anim. 45:467–472. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Butcher DT, Alliston T and Weaver VM: A
tense situation: forcing tumour progression. Nat Rev Cancer.
9:108–122. 2009. View
Article : Google Scholar : PubMed/NCBI
|