Introduction
Atopic dermatitis (AD) is an inflammatory skin
disease, which is associated with allergic diseases and has a
genetic background (1). Atopic
dermatitis occurs mainly in response to an infiltration of mast
cells (2) and dysregulation in T
helper (Th) cell reactivities (3).
The production of interleukin-4 (IL-4) from Th2 cells and mast
cells and interferon-γ (IFN-γ), which is produced from Th1 or mixed
Th1/Th2 cells, has been observed to trigger isotype-switching to
immunoglobulins in AD (4).
Excessive immunoglobulin E (IgE) production and histamine release
from activated mast cells are indicative of AD in humans and have
been observed in mice with AD-like skin lesions (5,6).
Skin mast cells contribute to AD inflammation. The increased levels
of IL-4 and IFN-γ, IgE production and histamine release during AD
are all closely associated with the infiltration of mast cells into
the site of inflammation (2). Mast
cells are involved in AD through releasing Th2 polarizing
cytokines, IL-4 and generating pruritus symptoms through release of
histamine and tryptase (7). In
addition, mast cells may increase histamine release and enhance
IgE-dependent skin inflammation in mice (8).
Previous studies on complement anaphylatoxin
receptors have been undertaken in multiple fields, including
cardiovascular disease and inflammation (9–11).
C5a is a risk factor for skin inflammation and specifically binds
to its receptor C5aR on immune cells, including mast cells and
leads to proinflammatory activation (12). It has been demonstrated that C5a
can induce chemoattraction and induce secretion in mast cells
during inflammation through a C5aR-mediated pathway (13); therefore, the aim of the present
study was to determine whether a C5aR antagonist (C5aRA) inhibits
AD in mice through suppressing the C5aR-mediated cascade action of
mast cells.
In the present study, the role of C5aR-mediated
activity during AD was assessed for the first time, to the best of
our knowledge. Dorsal skin of BALB/c mice was treated with
2,4-dinitrochlorobenzene (DNCB) and C5aR expression in skin tissue
was assessed. The effect of DNCB or DCNB combined with C5aRA on
skin-fold thickness the number of total infiltrating leukocytes
(mast cells, neutrophils, lymphocytes, monocytes and a few
eosinophils and basophils) as well as the levels of IL-4, IFN-γ,
histamine and IgE was also in determined. The results indicated
that C5aRA inhibited AD in mice, possibly through suppressing the
C5aR-mediated cascade action of mast cells, which may represent a
novel therapeutic strategy for AD.
Materials and methods
Animals
BALB/c mice (specific pathogen-free males, 7 weeks
old) were purchased from Shanghai Laboratory Animal Centre
(Shanghai, China). The animal care and protocol of the present
study was in accordance with the Animal Experiment Guidelines of
Harbin Medical University (Harbin, China). The mice were kept under
a 12-h light/dark cycle at 22°C and fed an unlimited quantity of
water and feed throughout the duration of the experiment. The mice
were divided into three groups: Control group, AD-like symptom
group and C5aRA-treated group.
Induction of an atopic dermatitis-like
skin disorder with corresponding immunology
DNCB was applied onto the hairless dorsal skin of
the mice. Following complete removal of dorsal hair in an area of
~8 cm2, 100 μl 1% DNCB dissolved in olive oil was
applied onto the dorsal skin once a day to enable sensitization in
the first week. The following week, 100 μl 0.2% DNCB was
applied to their dorsal skin twice every three days. DNCB was
dissolved in a 4:1 mixture of acetone and olive oil. C5aRA JPE-1375
(Jerini AG, Berlin, Germany) 1 μg in 100 μl
phosphate-buffered saline (PBS; 137 mM NaCl, 2.7 mM KCl, 4.3 mM
Na2HPO4, 1.4 mM KH2PO4;
pH adjusted to 7.4 with HCl, Sigma-Aldrich, Shanghai, China) was
injected intradermally. After a 2-week DNCB application period with
or without C5aRA intracutaneous injection, blood samples from the
posterior vena cava of the mice were used for immunological or
hematological analysis. Furthermore, skin tissue from each mouse
examined was sampled, a section was treated by sonication and
immediately centrifuged at 1,000 × g for 10 min for ELISA, and the
remainder was fixed with 10% neutral buffered formalin solution for
histopathological examination.
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR)
Following DNCB application for 2 weeks, total RNA
was extracted from skin tissue and relative levels of messenger
ribonucleic acid (mRNA) expression of C5aR were normalized to 18s.
The following primers were used: for C5aR forward,
5′-GACCCCATAGATAACAGCA-3′ and reverse, 5′-CAGAGGCAACACAAAACCCA-3′;
18s forward, 5′-GTAACCCGTTGAACCCCATT-3′ and reverse,
5′-CCATCCAATCGGTAGTAGCG-3′; and GAPDH forward,
5′-TTGCCATCAATGACCCCTTCA-3′ and reverse, 5′-CGCCCCACTTGATTTTGGA-3′.
RT-qPCR was performed using a PrimeScript™ RT reagent kit (Takara
Bio, Inc., Tokyo, Japan) with a Veriti™ Thermal Cycler (Applied
Biosystem, Foster City, CA, USA) at 37°C for 15 min, 85°C for 5 sec
and maintained at 4°C; and an SYBR Premix Ex Taq kit (Takara Bio,
Inc.) using a 7300 Fast Real-Time PCR system (Applied Biosystems)
under cycling conditions of 95°C for 30 sec for one cycle and 40
cycles of 95°C for 5 sec and 60°C for 31 sec.
Measurement of skin-fold thickness and
histological analysis
Skin-fold thickness was measured using a digital
thickness gauge (Mitutoyo, Kawasaki, Japan) by pulling up the skin
from shoulder to hip. The skin biopsies were fixed with 10% neutral
buffered formalin solution (~4% formaldehyde) and then embedded in
paraffin. Paraffin sections (3 μm each; Sigma-Aldrich) were
stained with hematoxylin and eosin (H&E) solution for detecting
skin histological features and various inflammatory cells (mast
cells, neutrophils, lymphocytes, monocytes and a few eosinophils
and basophils) and toluidine blue solution was used for detecting
mast cells. In brief, the skin sections were washed with distilled
water, stained with the alum hematoxylin (Sigma-Aldrich), rinsed in
running tap water and then treated with 0.3% hydrochloric acid
alcohol (Baowanchem, Nantong, China). The sections were then rinsed
under running tap water again and then washed in Scott’s tap water
substitute (RY Bio-tech Co., Shanghai, China) prior to rinsing
again in tap water. Sections were stained with eosin
(Sigma-Aldrich) for 2 min and then dehydrated in 95% and absolute
alcohols, two changes of 2 min each until excess eosin was cleared,
and mounted. The microscopy images were prepared using an automatic
microscope, Provis AX (Olympus, Tokyo, Japan), and a digital CCD
camera, Penguin 600CL (Pixera, Los Gatos, CA, USA). For toluidine
blue solution for mast cells, the skin sections were hydrated with
distilled water, stained in toluidine blue for 2–3 min, washed in
distilled water 3 times, dehydrated quickly through 95% and 2
changes of 100% alcohol, and then cleared in xylene or xylene
substitute with 2 changes of 3 min each. For morphometric analysis,
digital images were captured of three different areas at least at a
microscopic high-power field and the numbers of infiltrating
leukocytes (objective lens, ×100) and mast cells (objective lens,
×400) were counted as described previously (14).
Immunological observation
Blood from the posterior vena cava of mice and skin
tissue from each mouse examined were sampled and used for
immunological analysis. Production of IL-4 and IFN-γ in skin tissue
and IL-4, IFN-γ, histamine and IgE levels in serum were determined
using ELISA kits (Mouse IL-4, Mouse IFN-γ, Porcine Histamine HIS
and Mouse IgE ELISA kits; Sigma-Aldrich) according to the
manufacturer’s instructions. The absorbance at 450 nm was measured
using an ELISA reader (MTP-800; Corona Electric Co., Ltd, Tokyo,
Japan).
Statistical analysis
All statistical analyses were performed using SPSS
19.0 software (International Business Machines, Armonk, NY, USA).
Values are expressed as the mean ± standard error of the mean or
the mean ± standard deviation. Each experiment was repeated at
least three times. Student’s t-test was used and P<0.05 was
considered to indicate a statistically significant difference.
Results
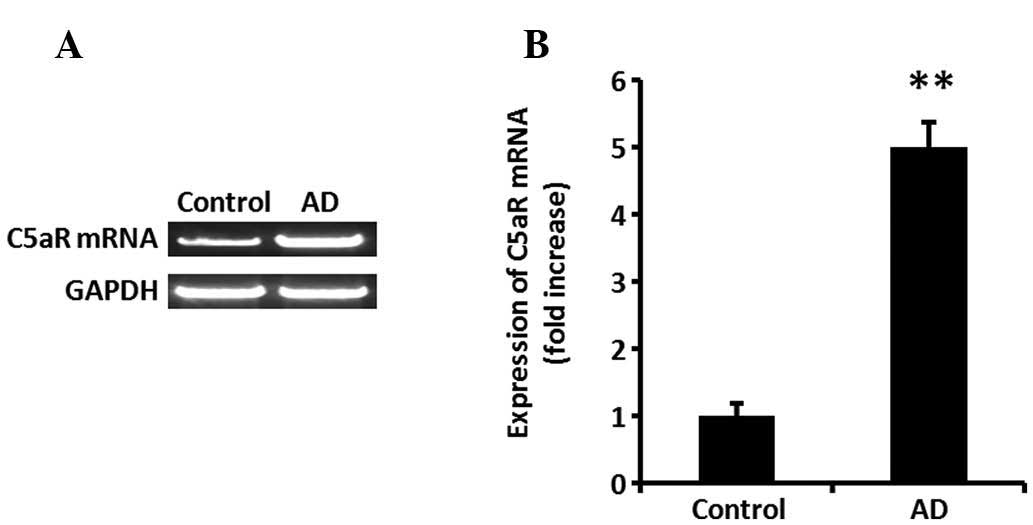
C5aR expression significantly increases
in skin tissue of mice with AD
To investigate the expression of C5aR, BALB/c mice
were treated with DNCB. Following DNCB application onto dorsal skin
for 2 weeks, the mRNA expression of C5aR in the tissue was assessed
using RT-qPCR. C5aR expression was significantly increased in AD
mouse skin tissue (Fig. 1A and B;
P<0.01).
C5aRA reduces the increased skin-fold
thickness in mice with AD
DNCB was applied onto BALB/c mouse dorsal skin for 2
weeks with or without C5aRA intracutaneous injection. Skin-fold
thickness was measured using a digital thickness gauge. The
skin-fold thickness was significantly increased in mice with AD
(Fig. 2; P<0.01) which was
significantly attenuated in the mice that had received
intracutaneous C5aRA injections (P<0.01). C5aRA intracutaneous
injection alone did not affect the skin-fold thickness in normal
mice (data not shown).
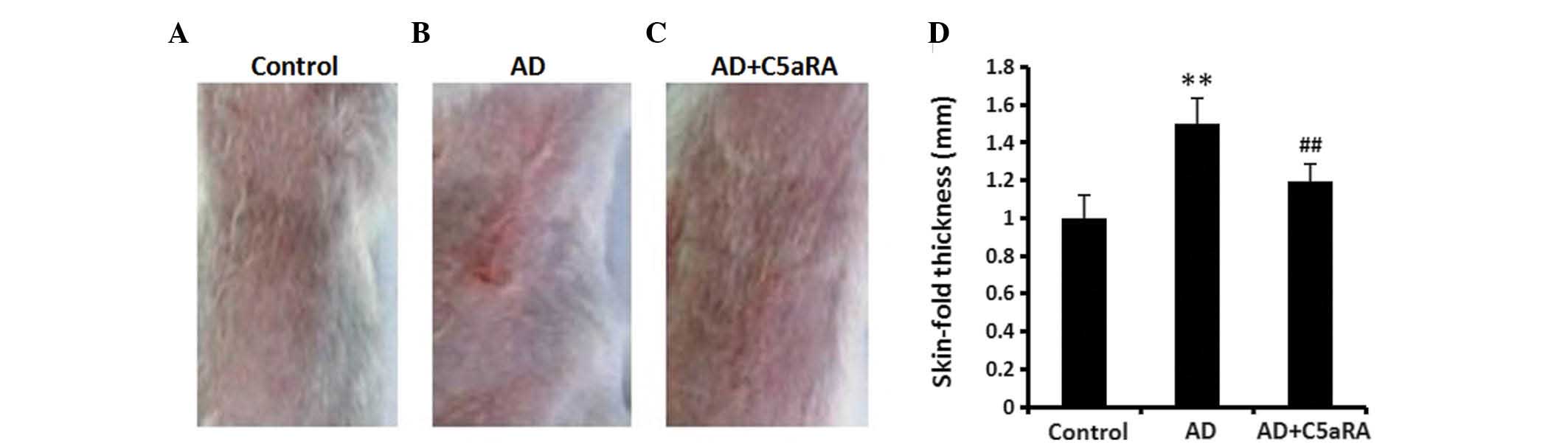 | Figure 2C5aRA reduces the increased skin-fold
thickness in mice with AD. DNCB was applied onto the dorsal skin of
BALB/c mice for 2 weeks with or without C5aRA (1 μg)
intracutaneous injection. Skin-fold thickness was measured using a
digital thickness gauge. The skin-fold thickness was significantly
increased in mice with AD, which was significantly attenuated by
intracutaneous C5aRA injection. Images of the dorsal skin of mice
in (A) the control, (B) AD and (C) AD+C5aRA groups (Magnification,
×10). (D) Quantification of A, B and C. Values are expressed as the
mean ± standard error of the mean (n=5). **P<0.01, AD
vs. control; ##P<0.01, AD+C5aRA vs. AD. AD, atopic
dermatitis; DNCB, 2,4-dinitrochlorobenzene; C5aRA, C5a receptor
antagonist. |
C5aRA attenuates inflammatory cell
infiltration in skin tissue of mice with AD
DNCB was applied onto BALB/c mouse dorsal skin for 2
weeks with or without C5aRA intracutaneous injection. Following
DNCB application, skin tissue from each mouse examined was sampled
and fixed with 10% neutral buffered formalin solution. Skin was
stained with H&E solution for detecting histological features
and various inflammatory cells and toluidine blue solution for
detecting mast cells. The numbers of total infiltrating leukocytes
(mast cells, neutrophils, lymphocytes, monocytes and a few
eosinophils and basophils) and mast cells in skin tissue were
counted in a microscopic high-power field. The numbers of
infiltrating leukocytes and mast cells in skin tissue were
significantly increased in mice with AD (Figs. 3 and 4; P<0.01), while the increased
skin-fold thickness as well as numbers of infiltrating leukocytes
(Fig. 3; P<0.01) and mast cells
(Fig. 4; P<0.05) were
significantly decreased in the mice that had received
intracutaneous C5aRA injections.
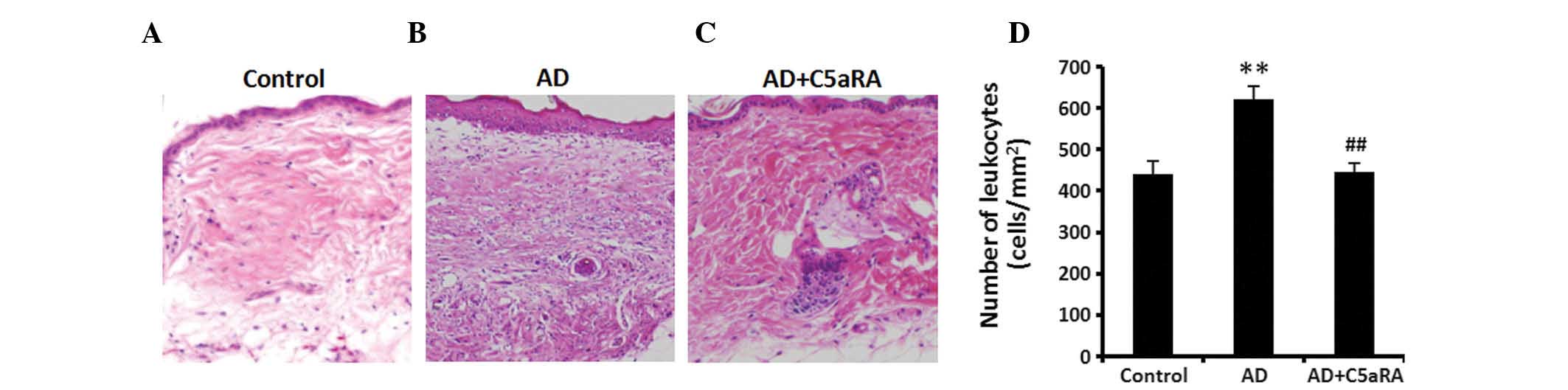 | Figure 3C5aRA attenuates inflammatory cell
infiltration in the skin tissue of mice with AD. DNCB was applied
onto the dorsal skin of BALB/c mice for 2 weeks with or without
C5aRA (1 μg) intracutaneous injection. Following DNCB
application, skin tissue from each mouse examined was sampled and
fixed with 10% neutral buffered formalin solution. Skin was stained
with hematoxylin and eosin solution to detect histological features
and various inflammatory cells. The number of infiltrating cells
(mast cells, neutrophils, lymphocytes, monocyte and a few
eosinophils and basophils) in skin tissue was counted at a
microscopic high-power field (objective lens, ×100). The number of
infiltrating cells in skin tissue was significantly increased in AD
mice, which was significantly attenuated following intracutaneous
C5aRA injection. Images of inflammatory cell infiltration in skin
tissue of mice in (A) control, (B) AD and (C) AD+C5aRA groups
(magnification, ×20). (D) Quantification of A, B and C. Values are
expressed as the mean ± standard deviation (n=5).
**P<0.01, AD vs. control; ##P<0.01,
AD+C5aRA vs. AD. AD, atopic dermatitis; DNCB,
2,4-dinitrochlorobenzene; C5aRA, C5a receptor antagonist. |
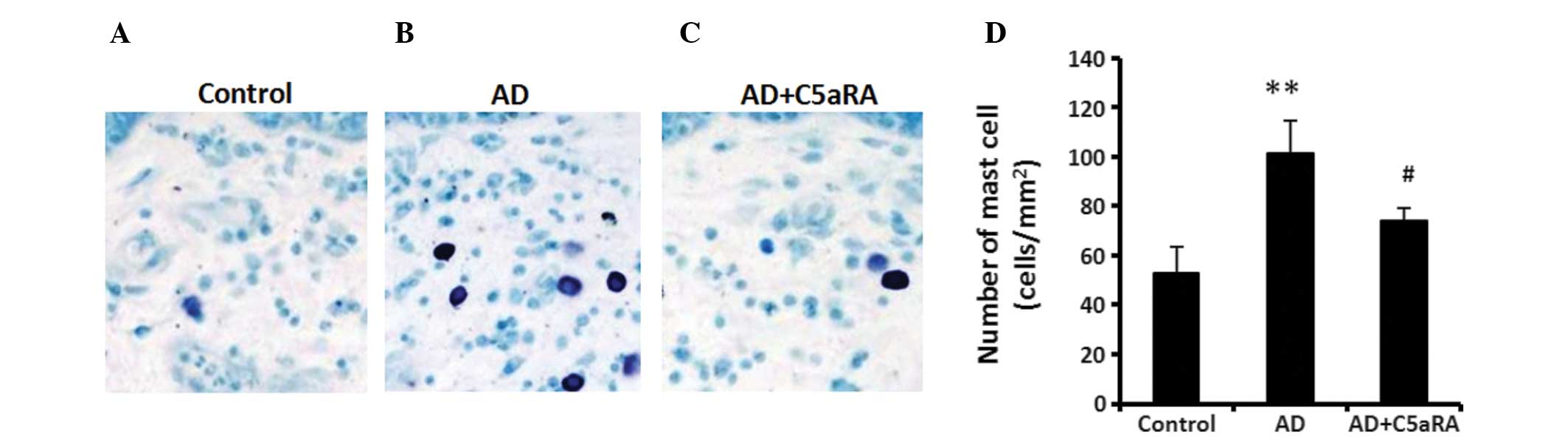 | Figure 4C5aRA attenuates mast cell
infiltration in AD mouse skin tissue. Following application of DNCB
onto the dorsal skin of BALB/c mice for 2 weeks with or without
C5aRA (1 μg) intracutaneous injection, skin tissue from each
mouse examined was sampled and fixed with 10% neutral buffered
formalin solution. Skin was stained with toluidine blue solution
for detecting mast cells, which were counted in a microscopic
high-power field (objective lens, ×400). The number of mast cells
in skin tissue was significantly increased in AD mice, which was
significantly attenuated in the mice receiving C5aRA intracutaneous
injection. Images of mast cells in the skin tissue of mice in (A)
control, (B) AD and (C) AD+C5aRA groups (magnification, ×200). (D)
Quantification of A, B and C. Values are expressed as the mean ±
standard deviation (n=5). **P<0.01, AD vs. control;
#P<0.05, AD+C5aRA vs. AD. AD, atopic dermatitis;
DNCB, 2,4-dinitrochlorobenzene; C5aRA, C5a receptor antagonist. |
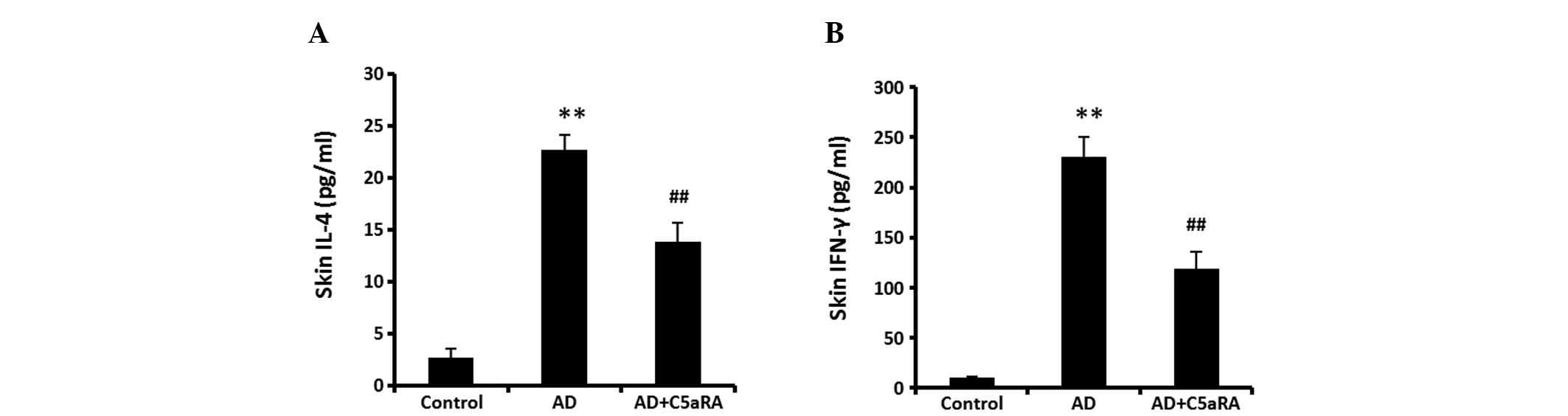
C5aRA attenuates DNCB-mediated increases
in IL-4 and IFN-γ levels in skin tissue
BALB/c mice were administered DNCB onto the dorsal
skin for 2 weeks with or without C5aRA intracutaneous injection.
Following DNCB application, the levels of IL-4 and IFN-γ were
determined using ELISA kits. The levels of IL-4 and IFN-γ in skin
tissue were significantly increased (Fig. 5, P<0.01). However, these
increases in levels of IL-4 and IFN-γ were significantly attenuated
in the mice receiving C5aRA intracutaneous injection
(P<0.01).
C5aRA reduces the levels of IL-4, IFN-γ,
histamine and IgE in serum
DNCB was applied onto the dorsal skin of BALB/c mice
for 2 weeks with or without C5aRA intracutaneous injection.
Following DNCB application, blood was sampled from the posterior
vena cava of the mice. The levels of IL-4, IFN-γ, histamine and IgE
in serum were significantly increased (Fig. 6; P<0.01). The increased levels
of IL-4 (P<0.05), IFN-γ (P<0.01), histamine (P<0.01) and
IgE (P<0.05) in serum were significantly decreased in the mice
receiving intracutaneous injection of C5aRA.
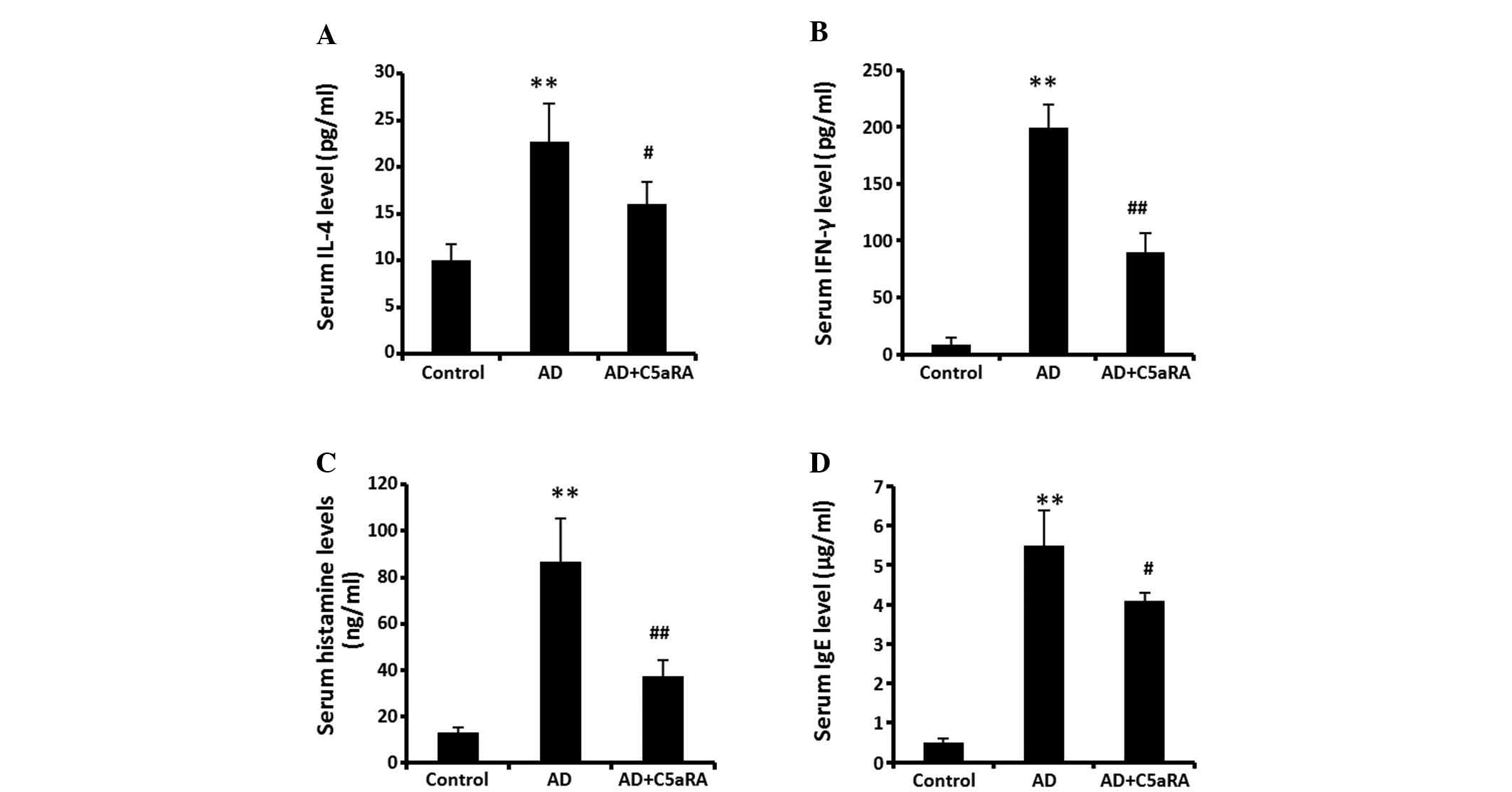 | Figure 6C5aRA reduces the levels of IL-4,
IFN-γ, histamine and IgE in mouse serum. Following DNCB application
onto the dorsal skin of BALB/c mice for 2 weeks with or without
C5aRA (1 μg) intracutaneous injection, blood was sampled
from the posterior vena cava of mice. Levels of IL-4, IFN-γ,
histamine and IgE in serum were determined using ELISA kits. Levels
of (A) IL-4, (B) IFN-γ, (C) histamine and (D) IgE were
significantly increased. The increases in levels of IL-4, IFN-γ,
histamine and IgE in serum were significantly attenuated in the
mice receiving C5aRA intracutaneous injection. Values are expressed
as the mean ± standard deviation (n=5). **P<0.01, AD
vs. control; ##P<0.01, #P<0.05,
AD+C5aRA vs. AD. AD, atopic dermatitis; DNCB,
2,4-dinitrochlorobenzene; Th, T helper; IL, interleukin; IFN,
interferon; IgE, immunoglobulin E; C5aRA, C5a receptor
antagonist. |
Discussion
In the present study, it was demonstrated for the
first time, to the best of our knowledge, that C5aRA inhibits AD in
mice by suppressing the C5aR-mediated cascade action of mast cells.
AD is an inflammatory skin disease that is associated with
allergies and genetics (1). Atopic
dermatitis occurs mainly from infiltration of various inflammatory
cells (mast cells, neutrophils, lymphocytes, monocytes and a few
eosinophils and basophils), particularly mast cells (2) and dysregulation in Th cell
reactivities (3,15). Previously, studies on the
complement anaphylatoxin receptors have been undertaken in multiple
fields, including cardiovascular disease and inflammation (9–11)
and it has been reported that anaphylatoxin receptor expression is
involved in skin inflammation (8).
C5a specifically binds to its receptor C5aR on immune cells,
including mast cells and leads to proinflammatory activation
(12). It has been demonstrated
that C5a can induce chemoattraction and secretion in mast cells in
inflammation through a C5aR-mediated pathway (13). As C5a is a risk factor for skin
inflammation and the expression of C5aR was significantly increased
in AD mouse skin tissue, the present study further investigated
whether C5aRA inhibited AD in mice by suppressing the C5aR-mediated
cascade action of mast cells.
AD is an inflammatory skin disease and is
distinguished by an increase in skin-fold thickness and
infiltration of numerous inflammatory cells into the site of
inflammation (1,2). In the present study, following DNCB
application onto the BALB/c mouse dorsal skin for 2 weeks, the
skin-fold thickness was significantly increased in AD mice and in
addition, the increase in skin-fold thickness was significantly
reduced in the mice that had received C5aRA intracutaneous
injection. Similarly, the quantity of total infiltrating leukocytes
(mast cells, neutrophils, lymphocytes, monocytes and a few
eosinophils and basophils) and mast cells in the skin tissue were
significantly increased in mice with AD, which was significantly
attenuated in the mice that had received C5aRA intracutaneous
injection.
IL-4, which is produced by Th2 cells and mast cells
(4,16) and IFN-γ, which is produced by Th1
or mixed Th1/Th2 cells, are known to trigger isotype-switching to
immunoglobulins in AD (6).
Excessive IgE production is a hallmark of atopic dermatitis in
humans and has been observed in mice with atopic dermatitis-like
skin lesions (16,18). The release of histamine from
activated mast cells is an indicator of the occurrence of allergic
diseases, including atopic dermatitis, which causes itching,
increased vascular permeability and the wheal-and-flare response of
immediate hypersensitivity (19).
As histamine is released immediately following exposure to
allergens, the increase in serum histamine levels is considered a
parameter for diagnosing the onset of allergic diseases (6,20).
In the present study, it was identified that in skin tissue, the
levels of IL-4 and IFN-γ were significantly increased in mice with
AD. The increases in levels of IL-4 and IFN-γ were significantly
attenuated in the mice receiving C5aRA intracutaneous injection
(Fig. 5). In the serum, the levels
of IL-4, IFN-γ, histamine and IgE were also significantly
increased, which was significantly attenuated in the mice receiving
C5aRA intracutaneous injection. Results of serum analysis confirmed
the findings in skin tissue.
The increased levels of IL-4 and IFN-γ, IgE
production and histamine release during the AD period are all
closely associated with the infiltration of mast cells to the site
of inflammation (2). Mast cells
are involved in AD by releasing Th2 and Th1/Th2, polarizing
cytokines to increase IL-4 and IFN-γ expression levels, while
generating pruritus symptoms through release of histamine and
tryptase (7). In addition, mast
cells may increase histamine release and enhance IgE-dependent skin
inflammation in mice (8). In
general, there is a balance between mast cells and other
pathogenesis factors of AD. There is also a balance between two
types of Th cells and any disruption of the balance may cause a
variety of disorders, including AD (21,22).
Corresponding with the mechanism shown in Fig. 7, following DNCB application onto
mouse dorsal skin for 2 weeks, C5aR expression in skin tissue was
significantly increased and the infiltration of mast cells was
significantly enhanced as well. C5aR is expressed on multiple types
of immune cells, including mast cells in skin tissue. C5a
specifically binds to the increased C5aR (23). Subsequently, the binding reaction
feeds back to mast cells and induces the chemoattraction of mast
cells (24). As shown in Fig. 7, the activated mast cells released
polarizing cytokines to increase IL-4 production from Th2 and IFN-γ
production from mixed Th1/Th2. In addition, the increased C5aR
expression on mast cells enhanced IgE in B cells. At the same time,
mast cells released histamine to generate pruritus symptoms. These
pathological processes constitute the pathogenesis of AD.
Intracutaneous injection of C5aRA directly prevented the binding of
C5a to C5aR on mast cells, then decreased the skin-fold thickness,
numbers of infiltrating leukocytes and mast cells levels of IL-4,
IFN-γ, histamine and IgE, thereby inhibiting the symptoms of AD.
The complement receptor cascade is a complex process, although it
was demonstrated that C5aRA inhibited AD in mice by suppressing
C5aR-mediated cascade action of mast cells. The complex inhibitory
process and the intracellular pathway require further
investigation.
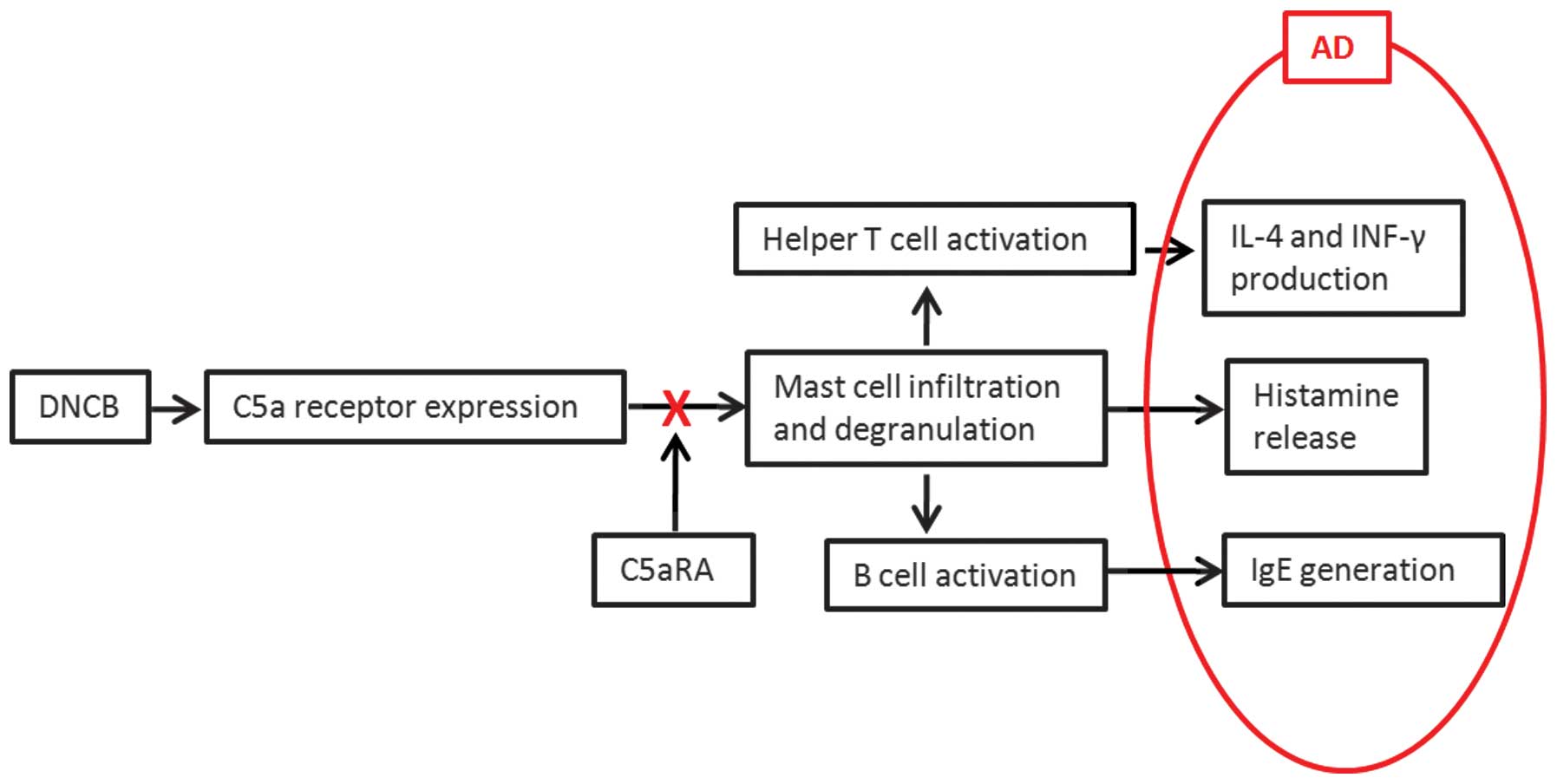 | Figure 7Inhibitory mechanism of C5aRA on AD.
Following DNCB application onto mouse dorsal skin for 2 weeks, C5aR
expression in skin tissue was significantly increased and the
infiltration of mast cells was significantly enhanced as well. The
activated mast cells released polarizing cytokines to increase IL-4
production from Th2 and IFN-γ production from mixed Th1/Th2. In
addition, the increased C5aR expression on mast cells enhanced IgE
in B cells. At the same time, mast cells released histamine to
generate pruritus symptoms. Intracutaneous injection of C5aRA
directly prevented the binding of C5a to C5aR on mast cells, then
decreased the skin-fold thickness, number of infiltrating
leukocytes and mast cells as well as levels of IL-4, IFN-γ,
histamine and IgE, and thereby inhibited the symptoms of AD. AD,
atopic dermatitis; DNCB, 2,4-dinitrochlorobenzene; Th, T helper;
IL, interleukin; IFN, interferon; IgE, immunoglobulin E; C5aRA, C5a
receptor antagonist. |
The present study was the first, to the best of our
knowledge, to demonstrate a crucial role of C5a-C5aR-C5aRA action
in the pathogenesis of AD, suggesting that C5aRA inhibited AD in
mice by suppressing the C5aR-mediated cascade action of mast cells
and providing a rationale for the potential use of C5aRA in
clinical practice for AD. These observations indicated that
C5a-C5aR-C5aRA may represent a novel therapeutic target for AD.
References
|
1
|
Matsumoto Y, Imai Y, Sugita Y, Tanaka T,
Tsujimoto G, Saito H and Oshida T: CCDC132 is highly expressed in
atopic dermatitis T cells. Mol Med Rep. 3:83–87. 2010.PubMed/NCBI
|
|
2
|
Abramovits W: Atopic dermatitis. J Am Acad
Dermatol. 53:S86–S93. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kurt-Jones EA, Hamberg S, Ohara J, Paul WE
and Abbas AK: Heterogeneity of helper/inducer T lymphocytes. I.
Lymphokine production and lymphokine responsiveness. J Exp Med.
166:1774–1787. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bonness S and Bieber T: Molecular basis of
atopic dermatitis. Curr Opin Allergy Clin Immunol. 7:382–386. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chan LS, Robinson N and Xu L: Expression
of interleukin-4 in the epidermis of transgenic mice results in a
pruritic inflammatory skin disease: an experimental animal model to
study atopic dermatitis. J Invest Dermatol. 117:977–983. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kapp A: The role of eosinophils in the
pathogenesis of atopic dermatitis - eosinophil granule proteins as
markers of disease activity. Allergy. 48:1–5. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kawakami T, Ando T, Kimura M, Wilson BS
and Kawakami Y: Mast cells in atopic dermatitis. Curr Opin Immunol.
21:666–678. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schäfer B, Piliponsky AM, Oka T, Song CH,
Gerard NP, Gerard C, Tsai M, Kalesnikoff J and Galli SJ: Mast cell
anaphylatoxin receptor expression can enhance IgE-dependent skin
inflammation in mice. J Allergy Clin Immunol. 131:541–548. 2013.
View Article : Google Scholar :
|
|
9
|
Tsuruta T, Yamamoto T, Matsubara S,
Nagasawa S, Tanase S, Tanaka J, Takagi K and Kambara T: Novel
function of C4a anaphylatoxin. Release from monocytes of protein
which inhibits monocyte chemotaxis. Am J Pathol. 142:1848–1857.
1993.PubMed/NCBI
|
|
10
|
Zhao Y, Xu H, Yu W and Xie BD: Complement
anaphylatoxin C4a inhibits C5a-induced neointima formation
following arterial injury. Mol Med Rep. 10:45–52. 2014.PubMed/NCBI
|
|
11
|
Jia N, Semba U, Nishiura H, Kuniyasu A,
Nsiama TK, Nishino N and Yamamoto T: Pivotal Advance:
Interconversion between pure chemotactic ligands and
chemoattractant/secretagogue ligands of neutrophil C5a receptor by
a single amino acid substitution. J Leukoc Biol. 87:965–975. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mayr M and Xu Q: Smooth muscle cell
apoptosis in arteriosclerosis. Exp Gerontol. 36:969–987. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Murakami Y, Yamamoto T, Imamichi T and
Nagasawa S: Cellular responses of guinea pig macrophages to C4a;
inhibition of C3a-induced O2-generation by C4a. Immunol Lett.
36:301–304. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nishiura H, Shibuya Y and Yamamoto T: S19
ribosomal protein cross-linked dimer causes monocyte-predominant
infiltration by means of molecular mimicry to complement C5a. Lab
Invest. 78:1615–1623. 1998.
|
|
15
|
Fiorentino DF, Bond MW and Mosmann TR: Two
types of mouse T helper cell. IV. Th2 clones secrete a factor that
inhibits cytokine production by Th1 clones. J Exp Med.
170:2081–2095. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kepron MR, Chen YW, Uhr JW and Vitetta ES:
IL-4 induces the specific rearrangement of gamma 1 genes on the
expressed and unexpressed chromosomes of lipopolysaccharide
activated normal murine B cells. J Immunol. 143:334–339.
1989.PubMed/NCBI
|
|
17
|
Matsumoto M, Ra C, Kawamoto K, Sato H,
Itakura A, Sawada J, Ushio H, Suto H, Mitsuishi K, Hikasa Y and
Matsuda H: IgE hyperproduction through enhanced tyrosine
phosphorylation of Janus kinase 3 in NC/Nga mice, a model for human
atopic dermatitis. J Immunol. 162:1056–1063. 1999.PubMed/NCBI
|
|
18
|
Takakura M, Takeshita F, Aihara M, Xin KQ,
Ichino M, Okuda K and Ikezawa Z: Hyperproduction of IFN-γ by CpG
oligodeoxynucleotide-induce exacerbation of atopic dermatitis-like
skin lesion in some NC/Nga mice. J Invest Dermatol. 125:1156–1162.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tamura T, Amano T, Ohmori K and Manabe H:
The effects of olopatadine hydrochloride on the number of
scratching induced by repeated application of oxazolone in mice.
Eur J pharmacol. 524:149–154. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Simon D, Braathen LR and Simon HU:
Eosinophils and atopic dermatitis. Allergy. 59:561–570. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sanderson CJ, Strath M, Warren DJ, O’Garra
A and Kirkwood TB: The production of lymphokines by primary
alloreactive T-cell clones: a co-ordinate analysis of 233 clones in
seven lymphokine assays. Immunology. 56:575–854. 1985.PubMed/NCBI
|
|
22
|
Bardana EJ Jr: Immunoglobulin E- (IgE) and
non-IgE-mediated reactions in the pathogenesis of atopic
eczema/dermatitis syndrome (AEDS). Allergy. 59(Suppl 78): 25–29.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Monk PN, Scola AM, Madala P and Fairlie
DP: Function, structure and therapeutic potential of complement C5a
receptors. Br J Pharmacol. 152:429–448. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xie P, Nishiura H, Semba U, Chen J, Zhao
R, Kuniyasu A and Yamamoto T: Inhibitory effects of C4a on
chemoattractant and secretagogue functions of the other
anaphylatoxins via Gi protein-adenylyl cyclase inhibition pathway
in mast cells. Int Immunopharmacol. 12:158–168. 2012. View Article : Google Scholar
|