Introduction
Cisplatin (cis-diamminedichloroplatinum II)
is one of the most widely used chemotherapeutic drugs; however, the
associated side effects and drug resistance limit its usage
(1–3). In order to overcome the problems
associated with cisplatin, previous studies have investigated the
mechanism by which cisplatin kills tumor cells and the reasons for
its side effects and resistance. The mechanism underlying cell
apoptosis induction by cisplatin is complicated and remains to be
elucidated. It has been demonstrated that cisplatin may have
effects on multiple cellular targets in tumor cells, not only on
DNA in the nucleus (4–6). Previous studies revealed that
treatment with cisplatin induces endoplasmic reticulum stress (ER
stress) in vitro and in vivo (7–9).
Cisplatin-induced ER stress-associated apoptosis is hypothesized to
be one of the cisplatin-induced pathways, which contributes to its
cytotoxicity and is also involved in drug resistance (10). Therefore, targeting ER stress may
be a potential strategy to improve the chemotherapeutic effect of
cisplatin.
ER stress triggers the unfolded protein response
(UPR), which involves the ER molecular chaperone, glucose-regulated
protein 78/binding immunoglobulin protein (GRP78/BIP), ER stress
sensor protein, protein kinase R-like ER kinase, inositol-requiring
enzyme 1 and activating transcription factor 6, and also their
downstream signaling pathway. ER stress induces cell autophagy,
cell apoptosis and the complicated regulatory network between them,
through the UPR system (11). In
UPR, autophagy performed a protective role by transporting
misfolded proteins for degradation to avoid ER stress-mediated
apoptosis (12–14).
The present study analyzed the effect of the
autophagy inhibitor, 3-methyladenine (3-MA), on cisplatin
cytotoxicity in U251 human glioma cells. The aim of the present
study was to clarify the role of autophagy in cisplatin-induced
U251 human glioma cell death in vitro, and to determine the
relationship between ER stress-associated apoptosis and
cisplatin-induced autophagy, in order to identify a novel treatment
strategy for glioma.
Materials and methods
Cell culture
U251 human glioma cells were purchased from the
American Type Culture Collection (Rockville, MD, USA) and cultured
in Dulbecco’s modified Eagle’s media (Gibco Life Technologies,
Gaithersburg, MD, USA), supplemented with 10% (v/v) fetal bovine
serum (Gibco Life Technologies) at 37°C with 5% CO2.
MTT assay
Cell viability was determined using an MTT assay.
Briefly, the cells (1×104 cells/well) were plated for 24
h in 96-well plates in 200 µl complete medium and exposed to
different concentrations of inhibitors for various durations. Each
treatment was repeated in six separate wells. The cells were
incubated at 37°C with 5% CO2, and MTT reagent (20
µl, 5 mg/ml; Sigma-Aldrich, St. Louis, MO, USA) in
phosphate-buffered saline (PBS) was added to each well and
incubated for 4 h. The formazan crystals were dissolved in 150
µl dimethyl sulfoxide (Beijing Dingguo Changsheng
Biotechnology Co., Ltd., Beijing, China) and the absorbance was
recorded at a wavelength of 490 nm using a Microplate Reader
(Bio-Tek Instruments, Inc., Winooski, VT, USA). Cell viability was
calculated as follows: Cell viability (%) =
absorbanceexperiment/absorbancecontrol ×
100.
Western blotting
For protein analysis, the cells were harvested
following 12 h treatment, washed with cold PBS and incubated in
ice-cold radioimmunoprecipitation buffer, containing 50 mM Tris-HCl
(pH 6.8), 0.1% SDS, 150 mM NaCl, 1 mM EDTA, 0.1 mM
Na3VO4, 1 mM NaF, 1% Triton X-100, 1% NP40, 1
mM dithiothreitol, 1 mM PMSF, 1 µg/ml aprotinin, 1
µg/ml leupeptin and 1 µg/ml pepstatin A. The cells
were sonicated (Ningbo Scientz Biotechnology Co., Ltd., Ningbo,
China) for 30 sec on ice and subsequently lysed at 4°C for 60 min.
The cell lysates were centrifuged for 30 min at 12,000 × g and the
protein concentration in the supernatants was determined using
bicinchoninic acid reagent (Pierce, Rockford, IL, USA). For western
blot analysis, lysate proteins (30–60 µg) were resolved on
12–15% SDS-polyacrylamide gel electrophoresis gels and transferred
onto nitrocellulose transfer membranes (Whatman, London, UK). The
membranes were blocked with 5% non-fat dry milk in buffer,
containing 10 mM Tris-HCl (pH 7.6), 100 mM NaCl and 0.1% Tween-20,
for 1 h at room temperature and subsequently incubated with the
following primary antibodies: Mouse monoclonal anti-PDI antibody
(cat. no. sc-166474), mouse monoclonal anti-Grp78 antibody (cat.
no. sc-376768), mouse monoclonal anti-CCAAT-enhancer binding
protein homologous protein (CHOP) antibody (cat. no. sc-7351),
rabbit polyclonal anti-caspase-4 antibody (cat. no. sc-28229),
rabbit polyclonal anti-caspase-3 antibody (cat. no. sc-7148),
rabbit polyclonal anti-LC3 antibody (cat. no. sc-292354) and mouse
monoclonal anti-ubiquitin (Ub) antibody (cat. no. sc-8017) (1:200
dilution; Santa Cruz Biotechnology, Inc., Dallas, TX, USA)
overnight at 4°C. The membranes were then incubated with horse
radish peroxidase-conjugated goat anti-mouse (cat. no. 31431) or
goat anti-rabbit (cat. no. 31466) secondary antibodies (Thermo
Fisher Scientific, Waltham, MA, USA) at 1:2,000 dilution for 1 h at
room temperature. The immunoreactive bands were visualized by the
diaminobenzidine (Sigma-Aldrich) coloration method. The
representative bands of proteins were measured with Quantity one
v4.62 software (Bio-Rad Laboratories, Inc., Hercules, CA, USA) and
analyzed as described previously (15). The protein expression levels were
normalized against β-actin and the ratios against β-actin were
expressed as the mean ± standard deviation from three independent
experiments.
Immunofluorescence staining and confocal
laser microscopy
U251 cells were cultured on coverslips at a density
of 5×104 cells/well in 500 µl complete medium and
exposed to different concentrations of inhibitors for various
durations. Following treatment, U251 cells were washed with cold
PBS three times and fixed in 4%(w/v) paraformaldehyde/PBS for 20
min. The cells were washed with cold PBS three times and were
digested with protein enzyme K (Beijing Dingguo Changsheng
Biotechnology Co., Ltd.) for 1 min and washed with PBS twice. The
cells were incubated with 0.1% (v/v) Triton X-100 for 6–10 min,
washed once with PBS and subsequently blocked for 30 min in 5%
(v/v) non-immune animal serum/PBS. The cells were incubated
overnight with the following primary antibodies at 1:200 dilution:
Mouse monoclonal anti-protein disulfide isomerase (PDI) antibody
(cat. no. sc-166474), rabbit polyclonal LC3 antibody (cat. no.
sc-292354) (Santa Cruz Biotechnology, Inc.); and rabbit monoclonal
anti-active caspase-3 antibody (cat. no. ab32042; Epitomics,
Burlingame, CA, USA), prior to three washes with PBS. The cells
were then incubated with the following secondary antibodies at
1:400 dilution: Tetramethylrhodamine (TRITC)-conjugated goat
anti-mouse (cat. no. 31660) and fluorescein
isothiocyanate/TRITC-conjugated goat anti-rabbit (cat. nos. 31635
and 31670) (Thermo Fisher Scientific) for 30 min in the dark. The
cells were washed three times with PBS, treated with Hoechst 33342
(Sigma-Aldrich)/H2O (1 µg/ml) for 2 min and
washed three times with PBS. The cells were examined on an Olympus
FV1000 confocal laser microscope (Olympus, Tokyo, Japan).
Statistical analysis
All statistical analyses were performed using SPSS
17.0 for Windows (SPSS, Inc., Chicago, IL, USA). Data were analyzed
by an one-way ANOVA, and Tukey’s post-hoc test was used to
determine the significance for all pairwise comparisons of
interest. P<0.05 was considered to indicate a statistically
significant difference. The data are representative of three
independent experiments performed in triplicate.
Results
Cisplatin induces ER stress-associated
apoptosis in U251 cells
The U251 cells were treated with different
concentrations of cisplatin for 24 h or with 10 µg/ml
cisplatin for different durations, prior to determining the
survival rate using an MTT assay. The results demonstrated that the
viability of U251 cells was decreased by treatment with cisplatin
in a dose- and time-dependent manner (Fig. 1A and B). The levels of cellular
apoptosis in U251 cells treated with cisplatin was assessed by
Hoechst 33342 staining. Cisplatin-induced apoptotic chromatin
condensation was more evident in the U251 cells following treatment
with cisplatin for 12 h compared with the control cells (Fig. 1C). The ER stress associated
apoptotic pathway was assessed and treatment with cisplatin
upregulated the expression levels of thioredoxin-like PDI, Grp78,
CHOP/growth arrest and DNA-damage-inducible protein 153, cleaved
caspase-4 and cleaved caspase-3 (Fig.
1D–F).
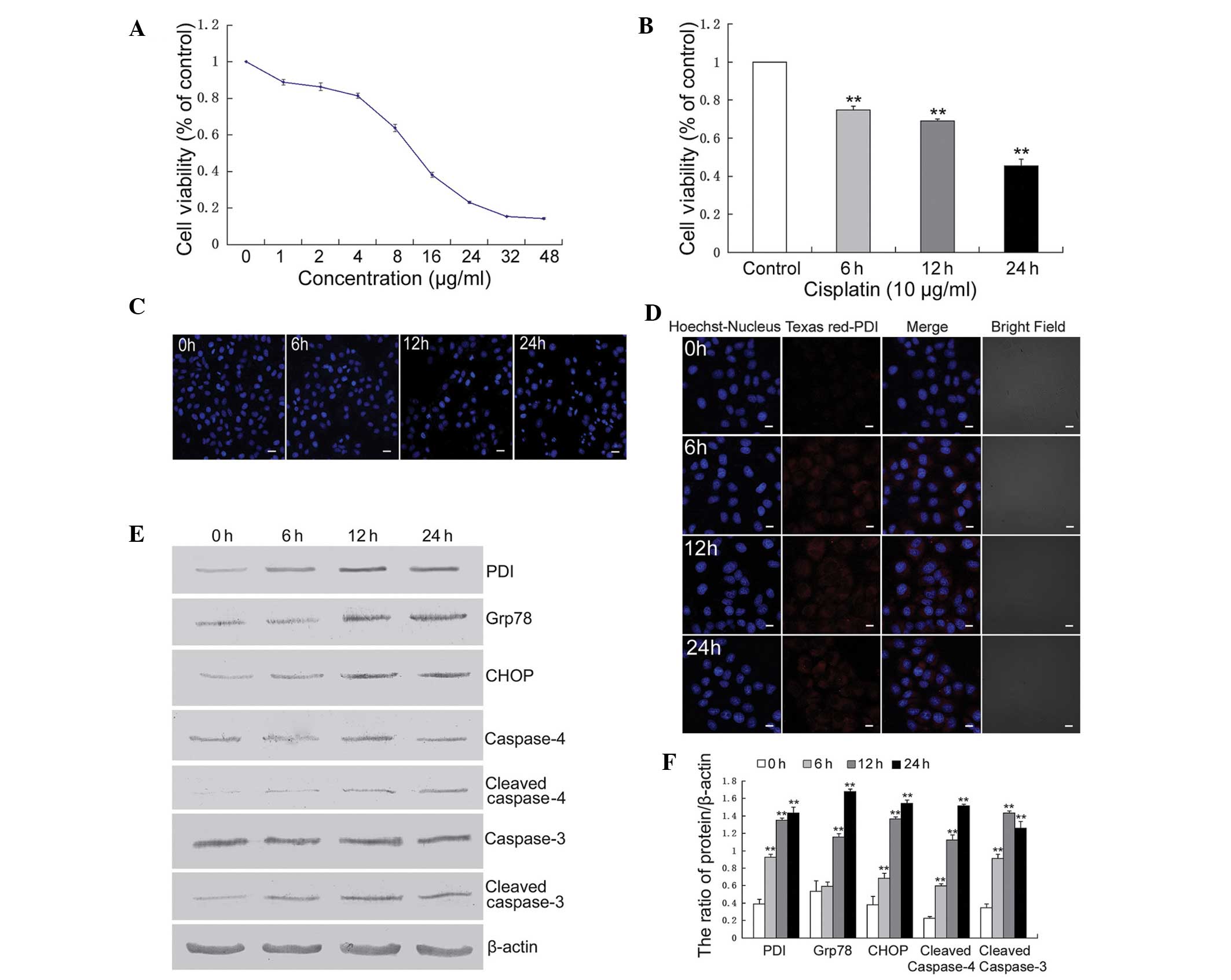 | Figure 1Cisplatin induced endoplasmic
reticulum stress-associated apoptosis in U251 cells. (A) U251 cells
were treated with varying concentrations of cisplatin for 24 h and
cell viability was determined by an MTT assay. (B) The cells were
treated with cisplatin (10 µg/ml) for 6, 12 and 24 h, and
cell viability was determined by an MTT assay
(**P<0.01, vs. control). (C) The cells were treated
with cisplatin (10 µg/ml) for 6, 12 and 24 h, stained with
Hoechst 33342 and cell morphology was observed by confocal
microscopy (scale bar, 20 µm). (D) The cells were treated
with cisplatin (10 µg/ml) for 6, 12 and 24 h, and the
expression of PDI was detected by confocal microscopy (scale bar,
10 µm, Texas red-conjugated secondary antibody). (E) The
cells were treated with cisplatin (10 µg/ml) for 6, 12 and
24 h, and western blot analysis was performed to detect the
expression levels of PDI, Grp78, CHOP, caspase-4, cleaved
caspase-4, caspase-3 and cleaved caspase-3. (F) Quantitation of the
protein expression levels. The data are expressed as the mean ±
standard deviation (n=3; **P<0.01, vs. Control). PDI,
protein disulfide isomerase; Grp, glucose regulated protein; CHOP,
CCAAT-enhancer-binding protein homologous protein. |
These results indicated that the ER
stress-associated apoptosis pathway was involved in
cisplatin-induced U251 cell death.
3-MA efficiently inhibits
cisplatin-induced autophagy in U251 cells
Previous studies have suggested that autophagy can
be induced by treatment with cisplatin; therefore, the activation
of autophagy was detected. LC3 puncta were observed in U251 cells
by confocal microscopy and it was demonstrated that the number of
puncta increased following treatment with cisplatin (Fig. 2A). LC3 is a molecular marker of
autophagy and is associated with the autophagosome membranes
following processing. LC3 transformation was assessed by western
blotting and revealed that following treatment with cisplatin, the
ratio of LC3II/I was increased at 6, 12 and 24 h (Fig. 2B and C).
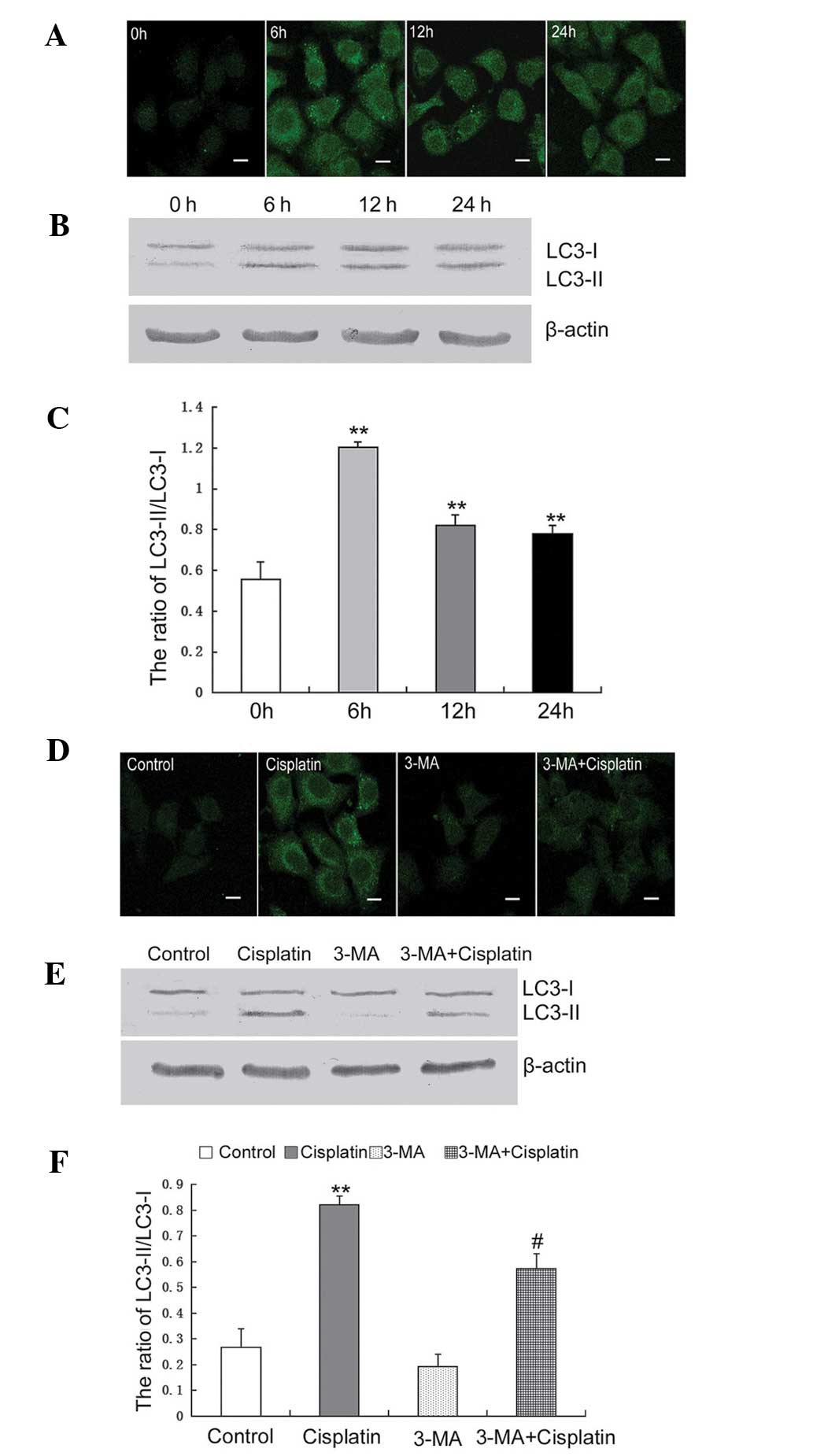 | Figure 23-MA efficiently inhibits
cisplatin-induced autophagy in U251 cells. (A) U251 cells were
treated with cisplatin (10 µg/ml) for 6, 12 and 24 h. The
expression of LC3 was detected by confocal microscopy (scale bar,
10 µm; FITC-conjugated secondary antibody). (B) The cells
were treated with cisplatin (10 µg/ml) for 6, 12 and 24 h,
and the expression of LC3 was detected by western blot analysis.
(C) The protein levels were quantified for LC3-II/I. The data are
expressed as the mean ± standard deviation (n=3;
**P<0.01, vs. control). (D) The cells were treated
with cisplatin (10 µg/ml) and/or 3-MA (10 mM) for 12 h, and
the expression of LC3 was detected by confocal microscopy (Bar, 10
µm; FITC-conjugated secondary antibody). (E) The cells were
treated with cisplatin (10 µg/ml) and/or 3-MA (10 mM) for 12
h, and the expression of LC3 was detected by western blot analysis.
(F) Quantitation of the protein expression of LC3-II/I. The data
are expressed as the mean ± standard deviation (n=3;
**P<0.01, vs. control; #P<0.05, vs.
cisplatin). 3-MA, 3-methyladenine; LC3, microtubule-associated
protein 1 light chain 3; FITC, fluorescein isothiocyanate. |
Based on these results, the autophagy-specific
inhibitor, 3-MA, was used to assess the onset of autophagy in U251
cells treated with cisplatin. Using confocal microscopy, LC3 puncta
were observed in U251 cells treated with cisplatin alone and in
combination with 3-MA. Following treatment for 12 h, LC3 puncta
were clearly observed in cells treated with cisplatin alone and
less LC3 puncta were observed in cells treated with cisplatin
combined with 3-MA (Fig. 2D). The
LC3 transformation in cells treated with cisplatin combined with
3-MA was significantly decreased, compared with the cells treated
with cisplatin alone (Fig. 2E and
F).
These results indicated that treatment with
cisplatin induced autophagy in the U251 cells and this was
efficiently inhibited by treatment with 3-MA.
3-MA increases cisplatin-induced ER
stress
Previous studies demonstrated that cisplatin
treatment induces the ER stress response, which regulates autophagy
and apoptosis (16,17). Activation of autophagy can
attenuate ER stress, therefore, the present study assessed the
effect of 3-MA on cisplatin-induced ER stress in U251 cells.
The expression of PDI was determined by confocal
microscopy. The expression of PDI in cells treated with cisplatin
combined with 3-MA was markedly increased, compared with the cells
treated with cisplatin alone (Fig.
3A). Western blotting was performed to determine the expression
levels of ubiquitinated proteins, PDI and Grp78. The expression
levels of ubiquitinated PDI and Grp78 were increased following the
inhibition of autophagy by treatment with 3-MA (Fig. 3B and C).
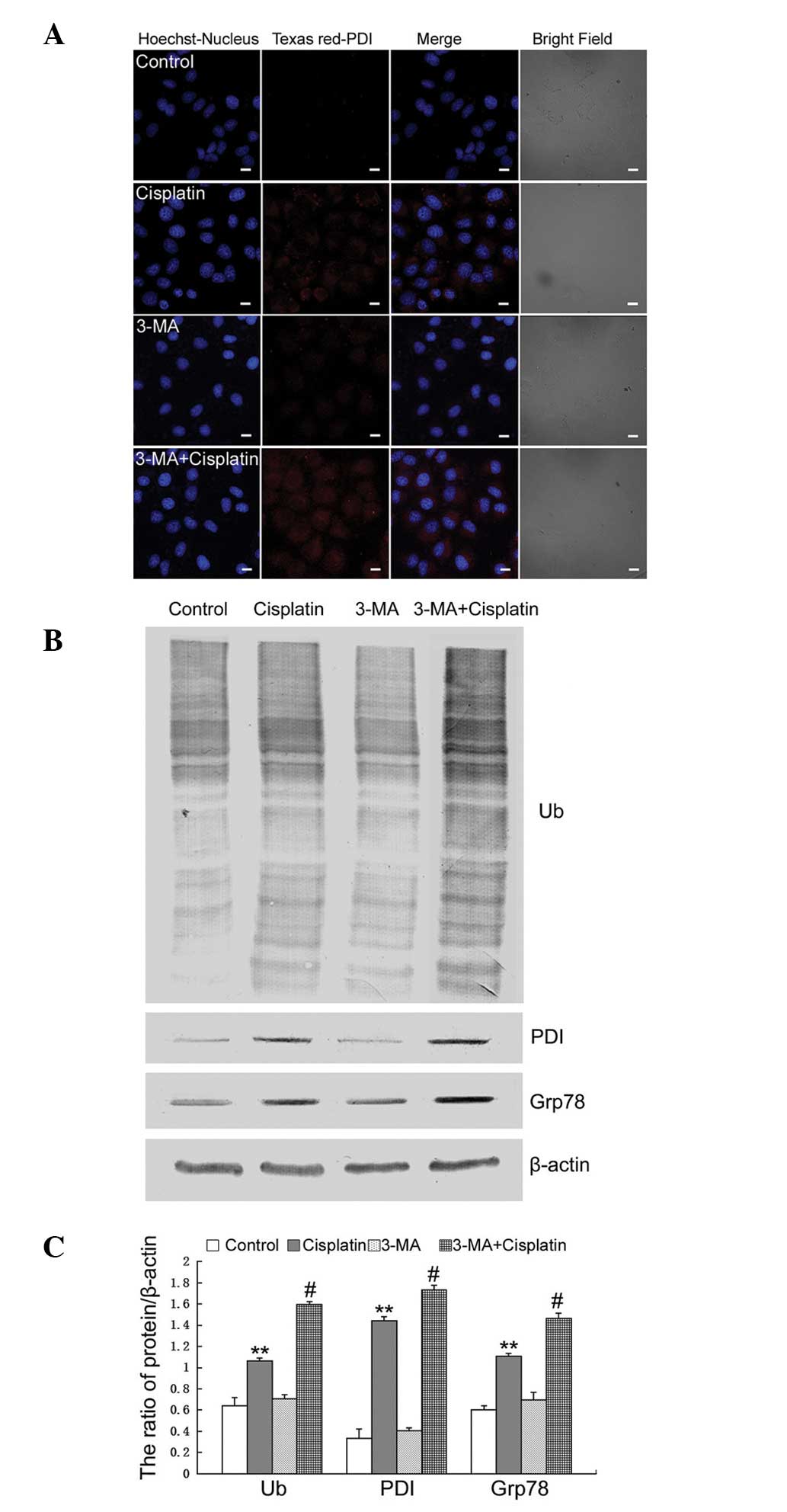 | Figure 33-MA increases cisplatin-induced
endoplasmic reticulum stress. (A) U251 cells were treated with
cisplatin (10 µg/ml) and/or 3-MA (10 mM) for 12 h, and the
expression of PDI was detected by confocal microscopy (scale bar,
10 µm; Texas red-conjugated secondary antibody). (B) The
cells were treated with cisplatin (10 µg/ml) and/or 3-MA (10
mM) for 12 h, and western blot analysis was performed to determine
the expression levels of ubquitinated proteins, PDI, and Grp78. (C)
Quantitation of the proteins level. The data are expressed as the
mean ± standard deviation (n=3; **P<0.01, vs.
control; #P<0.05, vs. cisplatin). 3-MA,
3-methyladenine; Ub, ubiqutin; PDI, protein disulfide isomerase;
Grp, glucose regulated protein. |
These results indicated that treatment with 3-MA
increased cisplatin-induced ER stress by inhibiting autophagy in
U251 cells.
3-MA increases cisplatin-induced
apoptosis by increasing ER stress
Previous studies have suggested that autophagy is
important for protecting against cisplatin treatment in tumor
cells. Therefore, the present study aimed to detect the effect of
inhibiting autophagy on cisplatin-induced apoptosis in U251
cells.
It was demonstrated by an MTT assay that treatment
with 3-MA increased the growth inhibitory rate in cells treated
with cisplatin (Fig. 4A). Using
Hoechst 33342 staining, it was demonstrated that 3-MA increased
cisplatin-induced apoptotic chromatin condensation (Fig. 4B). Confocal microscopy revealed the
expression of active caspase-3. The expression of active caspase-3
was increased in the cells treated with cisplatin combined with
3-MA, compared with the cells treated with cisplatin alone
(Fig. 4C). The expression levels
of CHOP, cleaved caspase-4 and cleaved caspase-3 were assessed by
western blotting and revealed that the expression levels of CHOP,
cleaved caspase-4 and cleaved caspase-3 were increased in cells
treated with cisplatin combined with 3-MA compared with the cells
treated with cisplatin alone (Fig. 4D
and E).
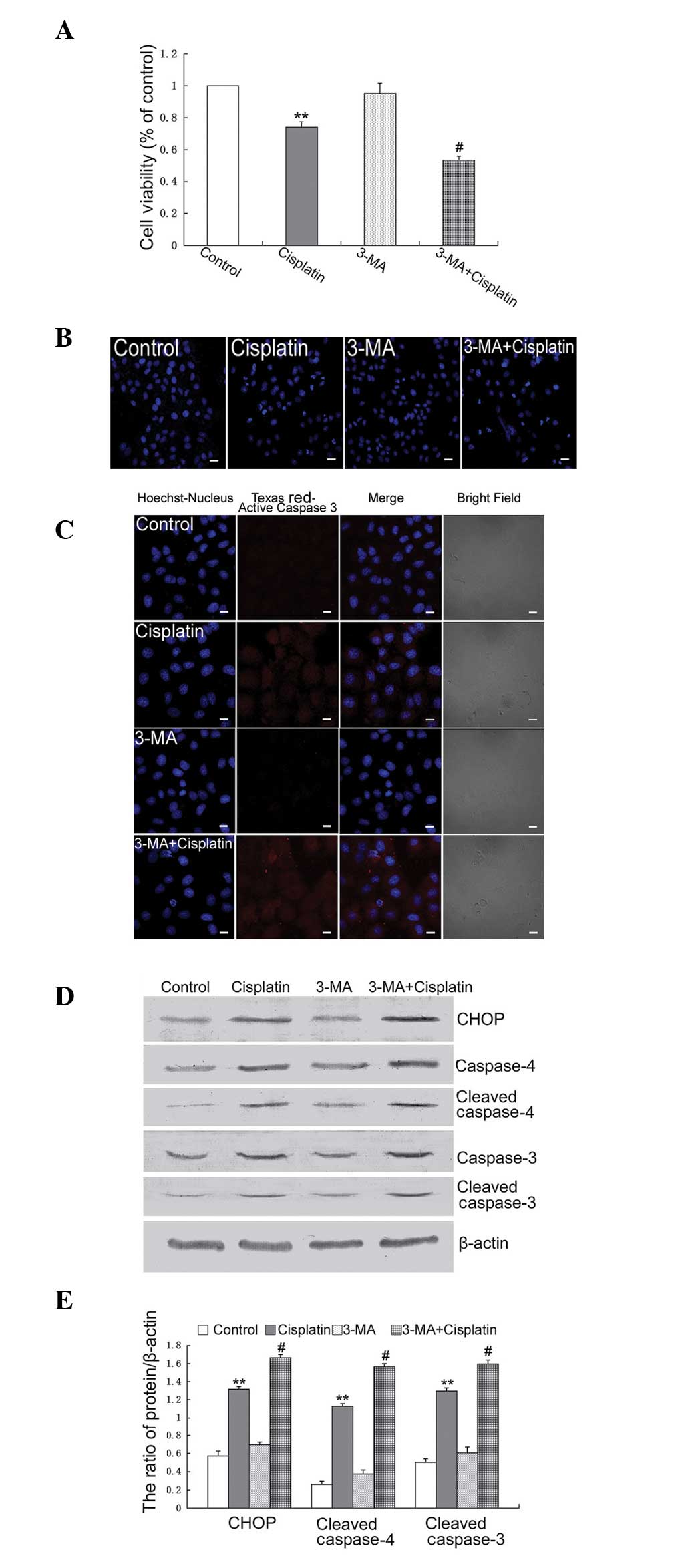 | Figure 43-MA increases cisplatin-induced
apoptosis by increasing endoplasmic reticulum stress. (A) U251
cells were treated with cisplatin (10 µg/ml) and/or 3-MA (10
mM) for 12 h, and cell viability was determined by MTT assay
(**P<0.01, vs. control; #P<0.05, vs.
Cisplatin). (B) The cells were treated with cisplatin (10
µg/ml) and/or 3-MA (10 mM) for 12 h, stained with Hoechst
33342 and cell morphology was observed by confocal microscopy
(Scale bar, 20 µm). (C) The cells were treated with
cisplatin (10 µg/ml) and/or 3-MA (10 mM) for 12 h, and the
expression of cleaved caspase-3 was detected by confocal microscopy
(Scale bar, 10 µm; Texas red-conjugated secondary antibody).
(D) The cells were treated with cisplatin (10 µg/ml) and/or
3-MA (10 mM) for 12 h, and western blot analysis was performed to
determine the expression levels of CHOP, caspase-4, cleaved
caspase-4, caspase-3 and cleaved caspase-3. (E) Quantitation of the
proteins level. The data are expressed as the mean ± standard
deviation (n=3; **P<0.01, vs. control;
#P<0.05, vs. cisplatin). 3-MA, 3-methyladenine; CHOP,
CCAAT-enhancer-binding protein homologous protein. |
These results indicated that treatment with 3-MA
increased cisplatin-induced apoptosis by increasing ER stress in
U251 cells.
Discussion
Cisplatin is one of the most efficient
chemotherapeutic drugs and is widely used for the treatment of
solid tumors, including glioma (18,19).
However, side effects and acquired drug resistance limit its
application. Although it has a satisfactory effect on several types
of tumor, the mechanisms by which is kills tumor cells remain to be
elucidated. As a cytotoxic agent, the effect of cisplatin causing
tumor cell death is hypothesized to be by DNA damage and the
inhibition of DNA synthesis. The DNA damage induced by cisplatin
activates multiple signaling pathways, which increase cell death,
mainly via the apoptotic pathway (20). Previous studies demonstrated that
cisplatin induces ER stress and non-nucleus dependent apoptotic
signal activation (7–9).
ER is an important organelle inside the cell and is
the location of protein synthesis regulation, protein folding
following synthesis and accumulation, stress reaction and calcium
ion level modulation. Previous studies indicated that the changes
in the microenvironment of tumor cells (glucose deprivation,
hypoxia) or antitumor drugs, can induce ER stress, including
misfolded and unfolded protein accumulation in the ER and
intracellular calcium ion balance abnormality (21–23).
ER stress triggers the UPR, which in solid tumors inhibits the
majority of translational processes, reduces the protein processing
burden in the ER and upregulates molecular chaperones, including
Grp78 and Grp94, to increase the ER protein folding capability.
Eventually, proteins that fail to be folded correctly will be
degraded by the proteasomal and autophagy pathways. When ER stress
becomes severe, it induces cell apoptosis by activating the
downstream apoptotic signaling pathway (24,25).
The decisive factor in this change is CHOP. Increased expression of
the transcription factor CHOP, changing the transcription of
several proteins, induces the activation of the pro-apoptotic
process, activates caspases, integrates mitochondrial events and
amplifies the death signal (26,27).
The present study demonstrated that cisplatin induced apoptosis in
U251 cells in a dose- and time-dependent manner. Treatment with
cisplatin increased the expression levels of PDI, Grp78, CHOP,
cleaved caspase-4 and cleaved caspase-3, which indicated the
activation of ER stress-associated apoptosis.
Autophagy is a reaction of cells to environmental
changes. The physical function of autophagy is to degrade
macromolecules, including proteins, RNA, redundant glycogen, and
aged or damaged organelles in membrane enclosed vesicles, which
provides a recycle role to maintain cellular homeostasis (28–30).
The formation of the autophagosome is the key event during this
process. LC3 exists as two forms, termed LC3-I and -II. LC3-II is
associated with the autophagosomal membranes following processing
in various cells. The ratio of LC3-II/I is used to assess the
levels of autophagy (31). The
present study revealed that cisplatin caused the accumulation of
LC3 puncta and the transformation of LC3-I to LC3-II, which
indicated that cisplatin activated autophagy. Combining the
autophagy inhibitor, 3-MA, and treatment with cisplatin revealed
that the expression levels of PDI, ubiquitinated proteins and Grp78
were significantly increased. These results indicated that the
inhibition of autophagy leads to high level ER stress. Furthermore,
increased ER stress increased the expression levels of CHOP,
cleaved caspase-4 and cleaved caspase-3, which led to increased
cisplatin-induced apoptosis.
In conclusion, the present study demonstrated that
cisplatin induced ER stress-associated apoptosis and autophagy in
U251 cells. The inhibition of autophagy using 3-MA increased the
expression levels of PDI, ubiquitinated proteins, Grp78 and CHOP,
and induced the activation of caspase-4 and caspase-3. Treatment
with 3-MA combined with cisplatin increased cisplatin-induced
apoptosis by increasing ER stress. This indicated that the
inhibition of autophagy may be a therapeutic target for the
improvement of cisplatin chemotherapy in glioma.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (grant nos. 30973072 and
81372683).
References
|
1
|
Stathopoulos GP: Cisplatin: process and
future. J BUON. 18:564–569. 2013.PubMed/NCBI
|
|
2
|
Köberle B, Tomicic MT, Usanova S and Kaina
B: Cisplatin resistance: preclinical findings and clinical
implications. Biochim Biophys Acta. 1806:172–182. 2010.PubMed/NCBI
|
|
3
|
Sleijfer DT, Meijer S and Mulder NH:
Cisplatin: a review of clinical applications and renal toxicity.
Pharm Weekbl Sci. 7:237–244. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Macciò A and Madeddu C: Cisplatin: an old
drug with a newfound efficacy – from mechanisms of action to
cytotoxicity. Expert Opin Pharmacother. 14:1839–1857. 2013.
View Article : Google Scholar
|
|
5
|
Sancho-Martínez SM, Prieto-García L,
Prieto M, López-Novoa JM and López-Hernández FJ: Subcellular
targets of cisplatin cytotoxicity: an integrated view. Pharmacol
Ther. 136:35–55. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yu F, Megyesi J and Price PM: Cytoplasmic
initiation of cisplatin cytotoxicity. Am J Physiol Renal Physiol.
295:F44–F52. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Peyrou M, Hanna PE and Cribb AE:
Cisplatin, gentamicin and p-aminophenol induce markers of
endoplasmic reticulum stress in the rat kidneys. Toxicol Sci.
99:346–353. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mandic A, Hansson J, Linder S and Shoshan
MC: Cisplatin induces endoplasmic reticulum stress and
nucleus-independent apoptotic signaling. J Biol Chem.
278:9100–9106. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu H and Baliga R: Endoplasmic reticulum
stress-associated caspase 12 mediates cisplatin-induced LLC-PK1
cell apoptosis. J Am Soc Nephrol. 16:1985–1992. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu Y, Wang C and Li Z: A new strategy of
promoting cisplatin chemotherapeutic efficiency by targeting
endoplasmic reticulum stress. Mol Clin Oncol. 2:3–7.
2014.PubMed/NCBI
|
|
11
|
Gardner BM, Pincus D, Gotthardt K,
Gallagher CM and Walter P: Endoplasmic reticulum stress sensing in
the unfolded protein response. Cold Spring Harb Perspect Biol.
5:a0131692013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Høyer-Hansen M and Jäättelä M: Connecting
endoplasmic reticulum stress to autophagy by unfolded protein
response and calcium. Cell Death Differ. 14:1576–1582. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Benbrook DM and Long A: Integration of
autophagy, proteasomal degradation, unfolded protein response and
apoptosis. Exp Oncol. 34:286–297. 2012.PubMed/NCBI
|
|
14
|
Ogata M, Hino S, Saito A, Morikawa K,
Kondo S, Kanemoto S, Murakami T, Taniguchi M, Tanii I, Yoshinaga K,
Shiosaka S, Hammarback JA, Urano F and Imaizumi K: Autophagy is
activated for cell survival after endoplasmic reticulum stress. Mol
Cell Biol. 26:9220–9231. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tian Z, Yang Z, Gao J, Zhu L, Jiang R and
Jiang Y: Lower esophageal microbiota species are affected by the
eradication of Helicobacter pylori infection using antibiotics. Exp
Ther Med. 9:685–692. 2015.PubMed/NCBI
|
|
16
|
Xu Y, Yu H, Qin H, Kang J, Yu C, Zhong J,
Su J, Li H and Sun L: Inhibition of autophagy enhances cisplatin
cytotoxicity through endoplasmic reticulum stress in human cervical
cancer cells. Cancer Lett. 314:232–243. 2012. View Article : Google Scholar
|
|
17
|
Song L, Liu H, Ma L, Zhang X, Jiang Z and
Jiang C: Inhibition of autophagy by 3-MA enhances endoplasmic
reticulum stress-induced apoptosis in human nasopharyngeal
carcinoma cells. Oncol Lett. 6:1031–1038. 2013.PubMed/NCBI
|
|
18
|
Stewart DJ: The role of chemotherapy in
the treatment of gliomas in adults. Cancer Treat Rev. 16:129–160.
1989. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lesser GJ and Grossman SA: The
chemotherapy of adult primary brain tumors. Cancer Treat Rev.
19:261–281. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Basu A and Krishnamurthy S: Cellular
responses to Cisplatin-induced DNA damage. J Nucleic Acids.
2010:2013672010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Perlmutter DH: Misfolded proteins in the
endoplasmic reticulum. Lab Invest. 79:623–638. 1999.PubMed/NCBI
|
|
22
|
Kopito RR and Ron D: Conformational
disease. Nat Cell Biol. 2:E207–E209. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ruiz A, Matute C and Alberdi E:
Intracellular Ca2+ release through ryanodine receptors contributes
to AMPA receptor-mediated mitochondrial dysfunction and ER stress
in oligodendrocytes. Cell Death Dis. 1:e542010. View Article : Google Scholar
|
|
24
|
Brewer JW: Regulatory crosstalk within the
mammalian unfolded protein response. Cell Mol Life Sci.
71:1067–1079. 2014. View Article : Google Scholar
|
|
25
|
Xu C, Bailly-Maitre B and Reed JC:
Endoplasmic reticulum stress: cell life and death decisions. J Clin
Invest. 115:2656–2664. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Szegezdi E, Logue SE, Gorman AM and Samali
A: Mediators of endoplasmic reticulum stress-induced apoptosis.
EMBO Rep. 7:880–885. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang XZ and Ron D: Stress-induced
phosphorylation and activation of the transcription factor CHOP
(GADD153) by p38 MAP Kinase. Science. 272:1347–1349. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Klionsky DJ and Codogno P: The mechanism
and physiological function of macroautophagy. J Innate Immun.
5:427–433. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ryter SW, Cloonan SM and Choi AM:
Autophagy: a critical regulator of cellular metabolism and
homeostasis. Mol Cells. 36:7–16. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Parzych KR and Klionsky DJ: An overview of
autophagy: morphology, mechanism and regulation. Antioxid Redox
Signal. 20:460–473. 2014. View Article : Google Scholar :
|
|
31
|
Klionsky DJ, Cuervo AM and Seglen PO:
Methods for monitoring autophagy from yeast to human. Autophagy.
3:181–206. 2007. View Article : Google Scholar : PubMed/NCBI
|














