1. Introduction
Endometriosis affects ~10% of women of reproductive
age and is associated with dysmenorrhea, pelvic pain, infertility
and in rare cases with malignant transformation. It is an
estrogen-dependent, progesterone-resistant disorder, which
contributes to endometriotic cell proliferation and survival
(1). Estrogen has been shown to
have an important role in the growth of endometriotic lesions
(1). Epidemiological studies have
suggested that gene polymorphism of estrogen and progesterone
receptors is associated with a risk of endometriosis (2,3).
Furthermore, several studies have been designed to identify
candidate genes and their pathways of endometriosis susceptibility
(4,5). In line with those of earlier studies,
the results showed that the candidate pathways, which included
genes with functions as hormonal regulators, cell cycle regulators,
signal transducers, transcription factors, hormones, cytokines,
chemokines and (pro)inflammatory molecules, proteases, as well as
molecules with functions in cell adhesion, motility, stress
response, detoxification, immune response and metabolism, may be
associated with susceptibility to endometriosis (4,5).
In addition, aberrations of components at the
genetic, epigenetic, transcriptional, post-transcriptional,
translational and post-translational level may drive candidate
pathways that may affect the susceptibility to this disorder. It is
now widely accepted that epigenetic mechanisms critically repress,
maintain or induce encoded genetic information (6). Silencing of
endometriosis-susceptibility genes by epigenetic aberrations has
been reported (4). Among the
candidate genes, the differentially methylated genes are often
associated with steroidogenesis, implantation and placental
development (6). These results
indicated that numerous genes which are down-regulated in the
eutopic endometrium of endometriosis are decidualization-associated
genes.
Alterations in the gene expression profile influence
susceptibility to endometriosis; however, the potential mechanism
that drives these alterations has remained elusive. The aim of the
present review was to understand how aberrant hormonal signaling is
integrated to generate a transcriptional output during the process
of endometriosis development.
2. Aberrant expression of candidate and
susceptibility genes
Steroid receptor-mediated tissue-specific complex
functions, including DNA methylation and transcriptional repression
signaling are the most affected pathways in the eutopic and ectopic
endometrium in women with endometriosis (7). Certain studies proposed a molecular
model stating that hypomethylation of the ER-β promoter results in
overexpression of ER-β and reduced expression of ER-α and
progesterone receptor (PR) in endometriotic stromal cells (8). In conjunction with estrogen-priming,
PR signaling is also involved in a variety of reproductive
functions, including embryo implantation, trophoblast invasion,
decidualization and subsequent formation of a functional placenta
that all are crucial for establishment and maintenance of
pregnancy. Two major PR isoforms, PR-A and PR-B, have been
identified (9). PR-A acts as a
repressor of PR-B, a stronger activator of progesterone target
genes. A reduced PR-B/PR-A ratio has been demonstrated in ectopic
tissue, as the promoter region of PR-B is specifically
hypermethylated (10). Therefore,
a number of progesterone target genes that are essential for
implantation and pregnancy were dysregulated in eutopic endometrium
of women affected with endometriosis (11). Abnormalities of the epigenome,
chromatin remodeling in response to steroid hormones and the
decidualization process may contribute to progesterone resistance
commonly observed in women with this disease.
A previous review analyzed 20 published datasets
that are specifically regulated during the process of
decidualization (12). Genetic and
epigenetic studies identified a plethora of
candidate/susceptibility genes and their downstream targets with
potential functional roles in decidualization. During the process
of decidualization, estrogen and progesterone bind to their
endometrial receptors and activate downstream targets, including
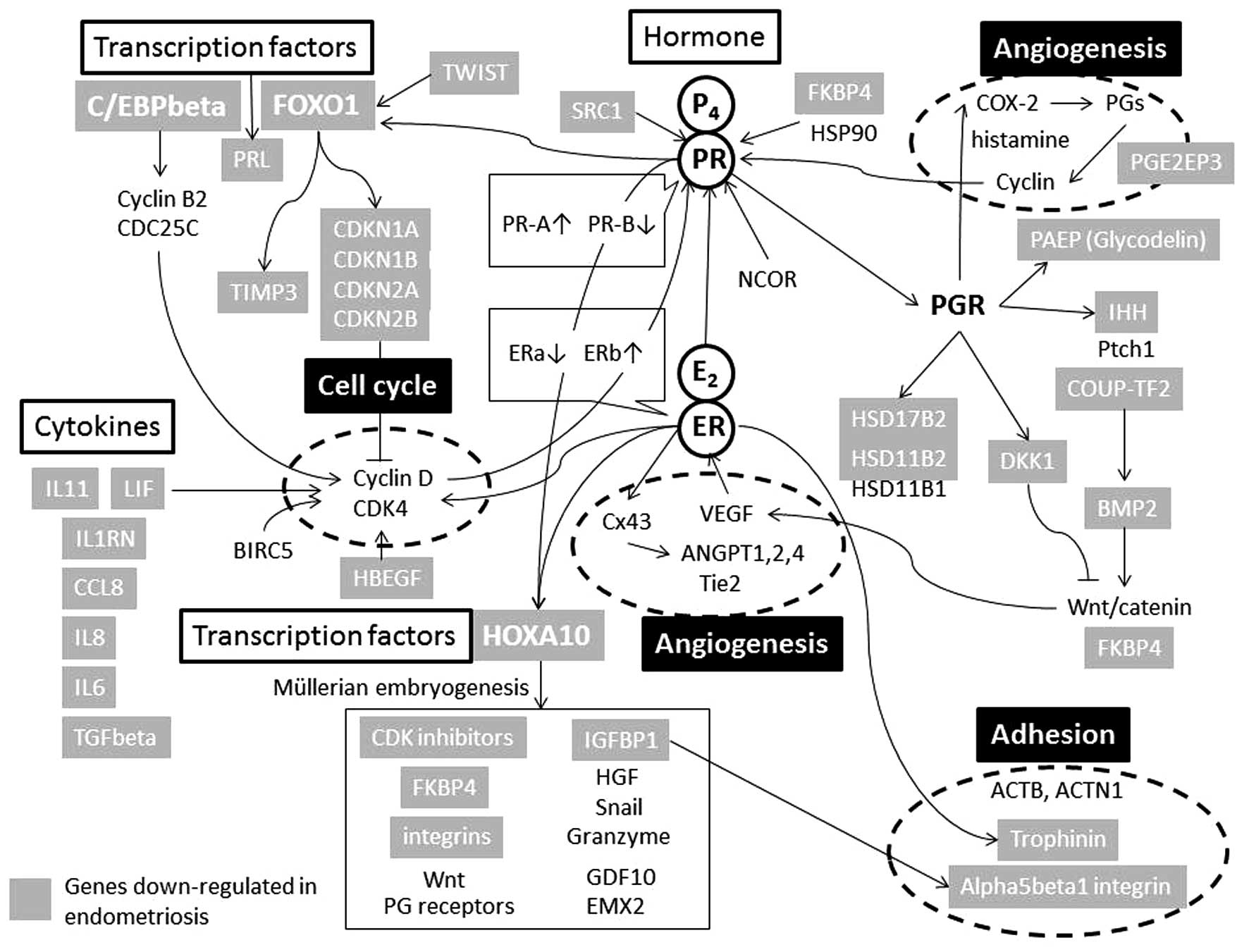
HOXA10, FOXO1, C/EBP-β and IHH, which in turn regulate cell
differentiation, resulting in an endometrium receptive to embryo
implantation (Fig. 1). Abrogation
of these genes causes stromal cell defects due to deregulation of
implantation and decidualization (13).
Recent studies based on (epi)genome-wide technology
have noted specific expression of candidate genes and
susceptibility genes for endometriosis (12). Reduced expression of the
decidualization susceptibility genes whose expression is regulated
by transcription factors (HOXA10, FOXO1 and C/EBP-β), growth
factors (the IGF and IGFBP families), cell cycle regulators (the
CDKN family) and cytokines/chemokines (LIF, IL11 and TGF-β) can
influence the expression of specific target genes and proteins with
central roles in the development of endometriosis (12). Bioinformatics analysis revealed a
substantial degree of overlapping candidate gene transcriptomes
between decidualization process and endometriosis from an
(epi)genetic point of view (5).
Downregulated genes identified in endometriosis among genes
overexpressed in the decidualization process encode proteins
directly involved in hormonal regulation as well as transcription
factors. Interestingly, in endometriosis, 28% of differentially
overexpressed genes are markedly downregulated during the
decidualization process (12).
This overlap is significantly greater than expected by chance. The
expression pattern of the endometriosis susceptibility genes
resembles that of the defective decidualization process. These
findings support the hypothesis that retrograde menstruation of
regurgitated endometrial cells with insufficient decidualization
possibly caused by epigenetic aberrations is implicated in the
pathogenesis of endometriosis.
3. Estrogen receptor (ER) as an epigenetic
driver
Epigenetic modifications control cell development,
differentiation and programming. Alterations encompassing
epigenetic changes have a key role in regulating gene expression
through post-translational modifications of histones (histone
methyltransferases, histone acetyltransferases and histone
deacteylases), DNA methylation, microRNA silencing, non-coding RNA,
long intergenic non-coding RNA and DNA repair mechanisms (14). Recent analyses identified several
genes that were hypoacetylated in endometriosis. These include
ER-α, homeobox A10 (HOXA10), CCAAT/enhancer-binding protein alpha
(CEBPA), p16 (INK4a, CDKN2A), p21 (Waf1/Cip1, CDKN1A), p27 (Kip1,
CDKN1B), checkpoint kinase 2 (CHEK2), death receptor 6 (DR6), and
E-cadherin (CDH1) (15). An
inverse correlation between the histone acetylation status and DNA
methylation in regulating gene silencing is well established.
Analysis of cytosine-phosphate-guanine (CpG)
dinucleotides identified >40,000 CpGs that were differentially
methylated in endometriosis (16).
Promoter hypermethylation may contribute to the understanding of
epigenetic regulation in this disorder. DNA hypermethylation in
endometriosis affects the expression of several key genes,
including PR-B, HOXA10, insulin-like growth factor binding protein
1 (IGFBP1), steroidogenic factor 1 (NR5A1) and aromatase (CYP19A1)
(16). Highly focused deviations
in endometriotic cell DNA methylation may affect key genes involved
in progesterone responsiveness and the decidualization process. As
shown in Fig. 1, hypermethylation
of the PR-B-specific promoter is considered to be a cause for the
development of progesterone resistance. ER activation may be a main
driving force for the progression to endometriosis. Expression
levels of ER-α in endometriosis were shown to be lower than those
in the eutopic endometrium, while ER-β was overexpressed in
endometriotic stromal cells as compared with that in endometrial
cells (17). An increased
ER-β/ER-α ratio in endometriosis compared to that in endometrial
tissues is associated with suppressed PR levels, contributing to
the loss of progesterone signaling or progesterone resistance
(7). Total PR and PR-B were lower
in endometriotic stromal cells compared to those in endometrial
cells. Several lines of evidence suggested that endometriosis is a
chronic disease with substantial epigenetic dysregulation, leading
to the imbalance between estrogen and progesterone (2,3).
For the present review, nine published datasets of
endometriosis susceptibility genes were analyzed (4,10,18–24).
Table I summarizes 29 genes
silenced by epigenetic aberration. A majority of
endometriosis-specific downregulated genes overlap with those known
to be regulated by ER or its downstream targets. These data
revealed important ER-dependent signaling pathways, regulatory
association and molecular connectivity among the downregulated
genes identified in the eutopic endometrium of women affected with
endometriosis (12). ER may act as
a direct epigenetic driver for endometriosis. Therefore, the
ER-dependent (epi)genomic regulation provides novel insight into
the pathogenesis of endometriosis.
 | Table IGenes downregulated in endometriosis
(n=29). |
Table I
Genes downregulated in endometriosis
(n=29).
| Function | Genes |
|---|
|
Cytokines/chemokines | LIF, IL6, IL8,
IL11, TGFB1, IL1RN, CCL8, BMP2 |
| Cell cycle | CDKN1A, CDKN1B,
CDKN2A, CDKN2B |
| Transcription
factors | FOXO1, HOXA10,
CEBPB, TWIST1 |
| Hormones | FKBP4, PRL,
PGR |
| Growth factors | IGFBP1, HBEGF,
SRC |
| Signaling | IHH, DKK1,
PTGER3 |
| Adhesion | CDH1, TRO |
| Immune system | PAEP |
| Proteases | TIMP3 |
4. Defective chromatin organization and
remodeling
Recent studies may support defective chromatin
remodeling in endometriosis. DNA methylation and histone
modification are interrelated in regulating chromatin remodeling
and gene expression (25–28). Knowledge of the role of
three-dimensional (3D) architecture of chromatin on the
transcriptional regulation is evolving.
5. Somatic mutations of AT-rich interactive
domain 1A (SWI-like) (ARID1A)
Whole-exome sequencing identified that genes
involved in chromatin-remodeling complexes are frequently altered
in endometriosis, suggesting that genetic alterations in
chromatin-remodeling proteins may be involved in the pathogenesis
of this disorder (25). Somatic
mutations of a chromatin remodeler, such as ARID1A, were frequently
found in the endometriotic cyst epithelium in direct continuity
with the carcinoma (26). It has
been reported that defective or remodeled chromatin organization
has a key role in tumor development and growth (27). One may speculate that chromatin
architectures present in normal endometrial cells may be lost in
endometriotic cells.
6. Defective chromatin interactions of ER
with its inducible genes
ER binding to chromatin promotes correct assembly of
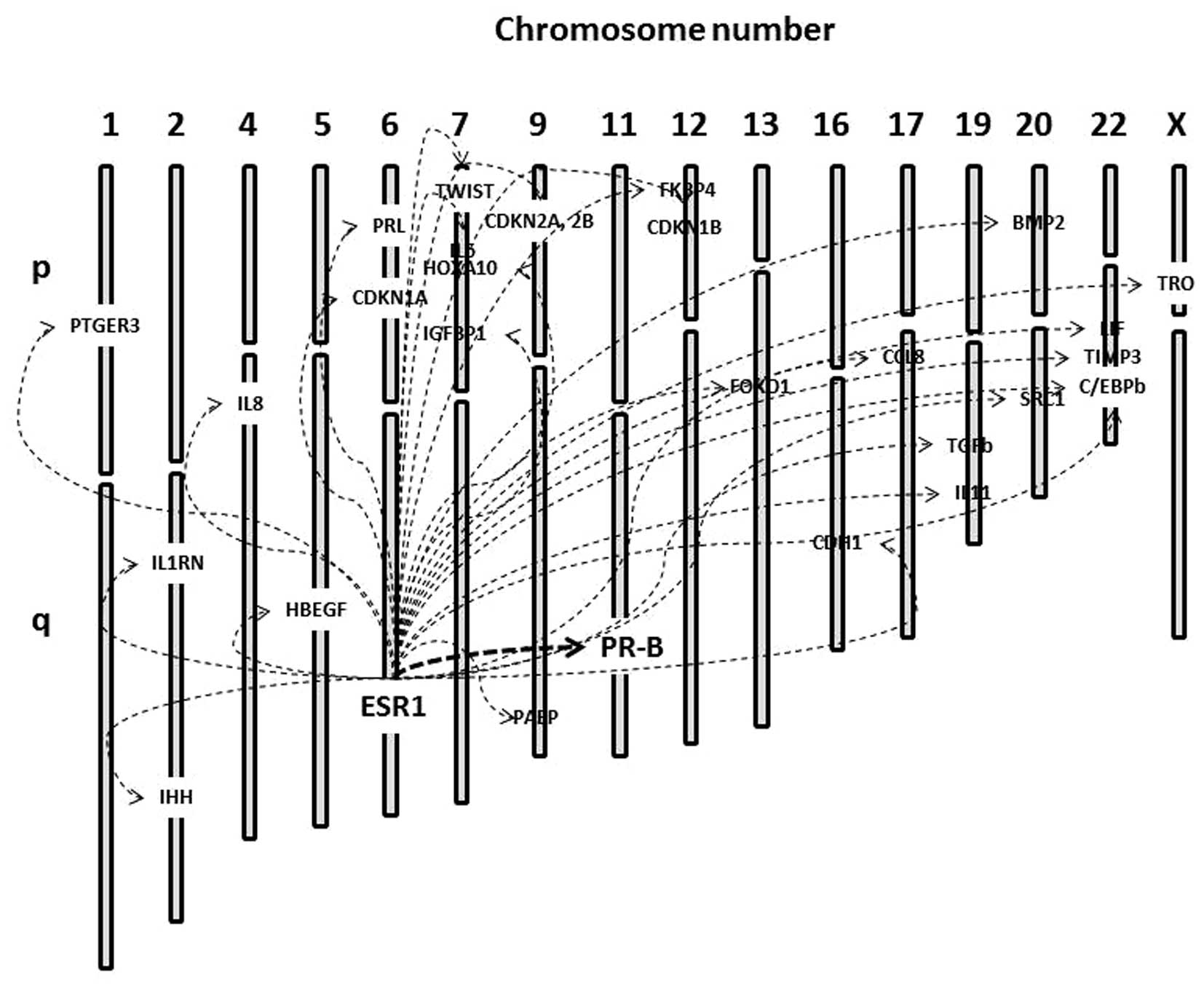
the transcriptional machinery of its target genes (28). As shown in Fig. 2 (29), interactions of ER with its
inducible target genes located in the same or different chromosomes
may be regulated by 3D long-distance chromatin interactions
(27,28). Future studies will provide a
catalog of aberrations pertinent to chromatin topology that
contribute to endometriosis development. The 3D organization of the
genome may achieve proper temporal and spatial patterns of gene
expression. ER is distributed within (near the promoters of
estrogen-regulated genes) and outside (distal regions) of gene
bodies (27). ER has numerous
binding sites along the genome and forms robust long-range
enhancer-promoter interactions as an intrachromosomal looping. In
addition, these remote binding sites are able to communicate with
their putative target genes via long-distance interchromosomal
interactions (30). We hypothesize
that a majority of endometriosis susceptibility genes may be
present in downstream targets of ER and associated with not only
various intragenic or intergenic chromatin epigenetic features, but
also long-range topological interactions among different
chromosomes. Modified chromatin interactions may be created in
temporal orchestration of the recruitment of transcription factors
specifically downregulated in endometriosis (Table I).
7. CCCTC-binding factor (CTCF)-mediated
chromatin interactions
CTCF, a zinc finger DNA binding protein, is one of
the most important transcription factors that participate in
numerous processes associated with global chromatin organization
and remodeling (31). CTCF is the
vertebrate insulator protein that affects gene expression by
mediating intra- and inter-chromosomal interactions (32). This transcription factor functions
mainly as a transcriptional repressor, including hormone-responsive
gene silencing, long-range chromatin interactions, enhancer
blocking and/or barrier gene insulation, genomic imprinting, and
X-chromosome inactivation (33).
CTCF can regulate a wide range of target gene functions over long
distances in the genome by promoting the formation of chromatin
loops. DNA loops form between ER bound to enhancer regions and
promoter regions of target genes, suggesting the possible
importance of CTCF in the 3D chromatin reorganization of the
ER-specific target gene loci (34). It has been reported that ER
activates its downstream target HOXA10 (16,35).
CTCF functions as a controller of HOXA cluster silencing (36). CTCF also acts upstream of the FOX
gene and influences differential interactions between ER and
chromatin (37). CTCF binding to
the specific insulator sites of the target gene loci with a tight
loop formation contributes to its silencing. Loss of the CTCF
machinery may result in defective chromatin architecture at the ER
target locus. We hypothesized that the CTCF-mediated chromatin
conformational 3D architecture may be dysregulated in
endometriosis. It remains largely unexplored, however, whether
chromatin loops are lost during gene repression in
endometriosis.
8. Mechanistic link between genomic
imprinting and endometriosis-associated methylation profiles
Several studies have identified the entirely
different DNA methylation profiles in endometriosis, which include
a large number of genes associated with transcription factors (HOX
and GATA) and steroidogenesis (NR5A1, STAR, STRA6 and HSD17B2)
(6,16). However, it remains elusive whether
tissue-specific global alterations in the methylome profile pattern
occur in endometriosis. The extent to which histone/DNA methylation
contributes to its pathogenesis also remains elusive.
To date, two studies investigating this context have
been published (4,38). Firstly, the presence of
hypermethylated regions located at the ends of chromosomes among
endometriosis samples suggested that the distribution pattern of
the hypermethylation was not random (38). This indicated that altered
non-random methylation of CpG sites which do not carry the mutation
has a possible role in the development of endometriosis. Secondly,
among the 29 hypermethylated genes in endometriosis, 19 genes
(65.5%) were located near the known imprinted foci (4). Genes designated to genomic imprinting
are associated with allele-specific expression. Imprinted genes are
highly conserved and organized in clusters. According to an
imprinting gene database (http://www.geneimprint.com/site/home), >100
imprinted genes have been identified in humans thus far (39). It was demonstrated that a number of
hypermethylated genes were located in close proximity to the
imprinted foci, which was significantly higher than expected by
chance.
9. Conclusions
Several lines of evidence have indicated that
endometriosis is an epigenetic disease. Firstly, bioinformatics
analysis revealed a substantial degree of overlapping
transcriptomes between endometriosis candidate genes and the
decidualization process (5).
Interestingly, in endometriosis, 28% differentially overexpressed
genes are markedly downregulated during the decidualization process
in endometrial stromal cells (12). Downregulated genes identified in
endometriosis among genes overexpressed in the decidualization
process encode proteins directly involved in cytokine/chemokine
signaling, cell cycle regulation, adhesion, the immune system, as
well as transcription factors, hormones, growth factors and
proteases (12) (Table I). In addition to the genes
described in Fig. 2, several
important genes, including DNA methyltransferases (40), COX-2 (41), estrogen receptor 2 (ESR2) (42) and steroidogenic factor-1 (SF-1)
(43) are aberrantly methylated.
The mechanism responsible for insufficient decidualization may be
associated with the development of endometriosis. Consistent with
this hypothesis, enhanced estrogenic activity and reduced
progesterone responsiveness are considered to be involved in the
etiology of endometriosis (11).
Therefore, endometriosis is a chronic disease with substantial
epigenetic dysregulation of decidualization susceptibility genes
(4).
Secondly, a large number of endometriosis
susceptibility genes are frequently associated with downstream
targets of ER. Approximately two thirds of the downregulated genes
associated with endometriosis susceptibility are not random, but
instead are located near the known paternally or maternally
expressed imprinting genes (4).
Hypermethylated regions showed a pattern of non-random
distribution, which serves to facilitate coordinated
transcriptional modulation in groups of genes (38). There appears to be a functional
association in the genomic location between genes specifically
downregulated in endometriosis and epigenetically imprinted
genes.
Finally, various chromatin architectures can
reportedly regulate ER-mediated gene expression, supporting a role
for long-range topological interactions in ER biology (28). CTCF was shown to have a role in
ER-mediated gene expression (28).
The transcription factor CTCF is involved in imprinting, long-range
chromatin interactions and transcription. Loss of CTCF machinery or
chromatin remodeling may lead to defective chromatin architecture
at the ER target locus. However, a global analysis of the role of
CTCF in chromatin looping, chromatin remodeling, various chromatin
architectures and the physical access of distal chromosomes in
endometriotic stromal cells has yet to be performed. Further
studies are required for molecular (epi)genomic approaches based on
the chromosome conformation capture method in the ER target gene
regions, and to then identify elements that can act over a
distance.
In conclusion, transcriptional repression at the ER
target locus may be an emerging key factor in endometriosis,
possibly through the non-random distribution of various DNA
epigenetic architectures, including histone acetylation,
methylation, CTCF-dependent insulation, genomic imprinting and 3D
chromatin features. Additional studies are required to further
elucidate the association of transcriptional profiles with
defective chromatin architectures in endometriosis.
Acknowledgments
The present review was supported by a grant-in-aid
for the Scientific Research from the Ministry of Education,
Science, and Culture of Japan to H.K (grant no. 26293361).
Abbreviations:
|
ER
|
estrogen receptor
|
|
CDKN
|
cyclin-dependent kinase inhibitor
|
|
CEBPA
|
CCAAT/enhancer-binding protein
alpha
|
|
CTCF
|
CCCTC-binding factor
|
|
HOXA10
|
homeobox A10
|
|
PR
|
progesterone receptor
|
References
|
1
|
Tamaresis JS, Irwin JC, Goldfien GA,
Rabban JT, Burney RO, Nezhat C, DePaolo LV and Giudice LC:
Molecular classification of endometriosis and disease stage using
high-dimensional genomic data. Endocrinology. 155:4986–4999. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hsieh YY, Wang YK, Chang CC and Lin CS:
Estrogen receptor alpha-351 XbaI*G and −397
PvuII*C-related genotypes and alleles are associated
with higher susceptibilities of endometriosis and leiomyoma. Mol
Hum Reprod. 13:117–122. 2007. View Article : Google Scholar
|
|
3
|
Wieser F, Schneeberger C, Tong D, Tempfer
C, Huber JC and Wenzl R: PROGINS receptor gene polymorphism is
associated with endometriosis. Fertil Steril. 77:309–312. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kobayashi H, Higashiura Y, Koike N,
Akasaka J, Uekuri C, Iwai K, Niiro E, Morioka S and Yamada Y: Genes
downregulated in endometriosis are located near the known
imprinting genes. Reprod Sci. 21:966–972. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kobayashi H, Uekuri C and Shigetomi H:
Towards an understanding of the molecular mechanism of
endometriosis: Unbalancing epithelial-stromal genetic conflict.
Gynecol Endocrinol. 30:7–15. 2014. View Article : Google Scholar
|
|
6
|
Yamagata Y, Nishino K, Takaki E, Sato S,
Maekawa R, Nakai A and Sugino N: Genome-wide DNA methylation
profiling in cultured eutopic and ectopic endometrial stromal
cells. PLoS One. 9:e836122014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bulun SE, Monsavais D, Pavone ME, Dyson M,
Xue Q, Attar E, Tokunaga H and Su EJ: Role of estrogen receptor-β
in endometriosis. Semin Reprod Med. 30:39–45. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bulun SE, Cheng YH, Pavone ME, Xue Q,
Attar E, Trukhacheva E, Tokunaga H, Utsunomiya H, Yin P, Luo X, et
al: Estrogen receptor-beta, estrogen receptor-alpha, and
progesterone resistance in endometriosis. Semin Reprod Med.
28:36–43. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Conneely OM, Maxwell BL, Toft DO, Schrader
WT and O’Malley BW: The A and B forms of the chicken progesterone
receptor arise by alternate initiation of translation of a unique
mRNA. Biochem Biophys Res Commun. 149:493–501. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wu Y, Strawn E, Basir Z, Halverson G and
Guo SW: Promoter hypermethylation of progesterone receptor isoform
B (PR-B) in endometriosis. Epigenetics. 1:106–111. 2006. View Article : Google Scholar
|
|
11
|
Burney RO, Talbi S, Hamilton AE, Vo KC,
Nyegaard M, Nezhat CR, Lessey BA and Giudice LC: Gene expression
analysis of endometrium reveals progesterone resistance and
candidate susceptibility genes in women with endometriosis.
Endocrinology. 148:3814–3826. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kobayashi H, Iwai K, Niiro E, Morioka S
and Yamada Y: Fetal programming theory: Implication for the
understanding of endometriosis. Hum Immunol. 75:208–217. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lim H, Ma L, Ma WG, Maas RL and Dey SK:
Hoxa-10 regulates uterine stromal cell responsiveness to
progesterone during implantation and decidualization in the mouse.
Mol Endocrinol. 13:1005–1017. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sharma S, Kelly TK and Jones PA:
Epigenetics in cancer. Carcinogenesis. 31:27–36. 2010. View Article : Google Scholar :
|
|
15
|
Nasu K, Kawano Y, Kai K, Aoyagi Y, Abe W,
Okamoto M and Narahara H: Aberrant histone modification in
endometriosis. Front Biosci (Landmark Ed). 19:1202–1214. 2014.
View Article : Google Scholar
|
|
16
|
Dyson MT, Roqueiro D, Monsivais D, Ercan
CM, Pavone ME, Brooks DC, Kakinuma T, Ono M, Jafari N, Dai Y, et
al: Genome-wide DNA methylation analysis predicts an epigenetic
switch for GATA factor expression in endometriosis. PLoS Genet.
10:e10041582014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shao R, Cao S, Wang X, Feng Y and Billig
H: The elusive and controversial roles of estrogen and progesterone
receptors in human endometriosis. Am J Transl Res. 6:104–113.
2014.PubMed/NCBI
|
|
18
|
Guo SW: Epigenetics of endometriosis. Mol
Hum Reprod. 15:587–607. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Monteiro JB, Colón-Díaz M, García M,
Gutierrez S, Colón M, Seto E, Laboy J and Flores I: Endometriosis
is characterized by a distinct pattern of histone 3 and histone 4
lysine modifications. Reprod Sci. 21:305–318. 2014. View Article : Google Scholar :
|
|
20
|
Nasu K, Kawano Y, Tsukamoto Y, Takano M,
Takai N, Li H, Furukawa Y, Abe W, Moriyama M and Narahara H:
Aberrant DNA methylation status of endometriosis: Epigenetics as
the pathogenesis, biomarker and therapeutic target. J Obstet
Gynaecol Res. 37:683–695. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Izawa M, Taniguchi F, Terakawa N and
Harada T: Epigenetic aberration of gene expression in
endometriosis. Front Biosci (Elite Ed). 5:900–910. 2013. View Article : Google Scholar
|
|
22
|
Colón-Díaz M, Báez-Vega P, García M, Ruiz
A, Monteiro JB, Fourquet J, Bayona M, Alvarez-Garriga C, Achille A,
Seto E, et al: HDAC1 and HDAC2 are differentially expressed in
endo-metriosis. Reprod Sci. 19:483–492. 2012. View Article : Google Scholar
|
|
23
|
Cakmak H and Taylor HS: Implantation
failure: Molecular mechanisms and clinical treatment. Hum Reprod
Update. 17:242–253. 2011. View Article : Google Scholar :
|
|
24
|
Kawano Y, Nasu K, Li H, Tsuno A, Abe W,
Takai N and Narahara H: Application of the histone deacetylase
inhibitors for the treatment of endometriosis: Histone
modifications as pathogenesis and novel therapeutic target. Hum
Reprod. 26:2486–2498. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li X, Zhang Y, Zhao L, Wang L, Wu Z, Mei
Q, Nie J, Li X, Li Y, Fu X, et al: Whole-exome sequencing of
endometriosis identifies frequent alterations in genes involved in
cell adhesion and chromatin-remodeling complexes. Hum Mol Genet.
23:6008–6021. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wiegand KC, Shah SP, Al-Agha OM, Zhao Y,
Tse K, Zeng T, Senz J, McConechy MK, Anglesio MS, Kalloger SE, et
al: ARID1A mutations in endometriosis-associated ovarian
carcinomas. N Engl J Med. 363:1532–1543. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu MH and Cheung E: Estrogen
receptor-mediated long-range chromatin interactions and
transcription in breast cancer. Mol Cell Endocrinol. 382:624–632.
2014. View Article : Google Scholar
|
|
28
|
Ross-Innes CS, Brown GD and Carroll JS: A
co-ordinated interaction between CTCF and ER in breast cancer
cells. BMC Genomics. 12:5932011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liang XH, Deng WB, Li M, Zhao ZA, Wang TS,
Feng XH, Cao YJ, Duan EK and Yang ZM: Egr1 protein acts downstream
of estrogen-leukemia inhibitory factor (LIF)-STAT3 pathway and
plays a role during implantation through targeting Wnt4. J Biol
Chem. 289:23534–23545. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hu Q, Kwon YS, Nunez E, Cardamone MD, Hutt
KR, Ohgi KA, Garcia-Bassets I, Rose DW, Glass CK, Rosenfeld MG, et
al: Enhancing nuclear receptor-induced transcription requires
nuclear motor and LSD1-dependent gene networking in inter-chromatin
granules. Proc Natl Acad Sci USA. 105:19199–19204. 2008. View Article : Google Scholar
|
|
31
|
Franco MM, Prickett AR and Oakey RJ: The
role of CCCTC-binding factor (CTCF) in genomic imprinting,
development, and reproduction. Biol Reprod. 91:1252014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yang J and Corces VG: Chromatin
insulators: A role in nuclear organization and gene expression. Adv
Cancer Res. 110:43–76. 2011.PubMed/NCBI
|
|
33
|
Ong CT and Corces VG: Enhancer function:
new insights into the regulation of tissue-specific gene
expression. Nat Rev Genet. 12:283–293. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fullwood MJ, Liu MH, Pan YF, Liu J, Xu H,
Mohamed YB, Orlov YL, Velkov S, Ho A, Mei PH, et al: An
oestrogen-receptor-alpha-bound human chromatin interactome. Nature.
462:58–64. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fambrini M, Sorbi F, Bussani C, Cioni R,
Sisti G and Andersson KL: Hypermethylation of HOXA10 gene in
mid-luteal endometrium from women with ovarian endometriomas. Acta
Obstet Gynecol Scand. 92:1331–1334. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Heger P, Marin B, Bartkuhn M, Schierenberg
E and Wiehe T: The chromatin insulator CTCF and the emergence of
metazoan diversity. Proc Natl Acad Sci USA. 109:17507–17512. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang Y, Liang J, Li Y, Xuan C, Wang F,
Wang D, Shi L, Zhang D and Shang Y: CCCTC-binding factor acts
upstream of FOXA1 and demarcates the genomic response to estrogen.
J Biol Chem. 285:28604–28613. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Borghese B, Barbaux S, Mondon F, Santulli
P, Pierre G, Vinci G, Chapron C and Vaiman D: Research resource:
Genome-wide profiling of methylated promoters in endometriosis
reveals a subtelomeric location of hypermethylation. Mol
Endocrinol. 24:1872–1885. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Edwards CA and Ferguson-Smith AC:
Mechanisms regulating imprinted genes in clusters. Curr Opin Cell
Biol. 19:281–289. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wu Y, Strawn E, Basir Z, Halverson G and
Guo SW: Aberrant expression of deoxyribonucleic acid
methyltransferases DNMT1, DNMT3A, and DNMT3B in women with
endometriosis. Fertil Steril. 87:24–32. 2007. View Article : Google Scholar
|
|
41
|
Wang D, Chen Q, Zhang C, Ren F and Li T:
DNA hypomethylation of the COX-2 gene promoter is associated with
up-regulation of its mRNA expression in eutopic endometrium of
endometriosis. Eur J Med Res. 17:122012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Xue Q, Lin Z, Cheng YH, Huang CC, Marsh E,
Yin P, Milad MP, Confino E, Reierstad S, Innes J, et al: Promoter
methylation regulates estrogen receptor 2 in human endometrium and
endo-metriosis. Biol Reprod. 77:681–687. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Xue Q, Lin Z, Yin P, Milad MP, Cheng YH,
Confino E, Reierstad S and Bulun SE: Transcriptional activation of
steroidogenic factor-1 by hypomethylation of the 5′ CpG island in
endometriosis. J Clin Endocrinol Metab. 92:3261–3267. 2007.
View Article : Google Scholar : PubMed/NCBI
|