Introduction
Squamous cell carcinoma (SCC) of the oral cavity is
the sixth most frequent solid cancer worldwide (1). Tongue squamous cell carcinoma (TSCC)
is the most common type of oral cancer and is well known for its
high rate of proliferation and lymph node metastasis. The majority
of TSCC patients are associated with smoking, heavy alcohol use and
HPV infection (2–4). According to the American Cancer
Society (5), while overall new
cancer cases increased ~8%, new cases of TSCC increased by >37%
in the same period. This indicates a major health problem
associated with TSCC and suggests the immediate requirement for an
improved understanding of this disease. To prevent and improve the
outcomes of TSCC, it is necessary to further understand the
molecular mechanism underlying the development and progression of
TSCC and to develop new target therapies.
Four and a half LIM protein 1 (FHL1) is a member of
the FHL protein family, which contains four complete LIM domains
and an N-terminal half LIM domain (6). It has been reported that LIM domains
function in protein-protein interactions with transcription
factors, cell-signaling molecules and cytoskeleton-associated
proteins (6,7). Previously, FHL1 has been demonstrated
to be important in carcinogenesis. FHL1 expression is downregulated
in various types of malignancy, including breast cancer, liver
cancer, kidney cancer, prostate cancer, gastric cancer, lung cancer
and oral squamous cell carcinoma (OSCC) (8–13).
FHL1 exerts its tumor suppressive role via multiple mechanisms.
FHL1 activates the tumor suppressor gene p21 (WAF1/CIP1) and
represses the oncogene c-myc through interaction with Smad2, Smad3
and Smad4 in liver cancer cells (10). In breast cancer cells, FHL1
interacted with estrogen receptors and thus decreases the
expression of pS2 and cathepsin D, two estrogen-responsive genes
(9). In addition, FHL1 induces G1
and G2/M cell cycle arrest in lung cancer cells by inhibiting the
expression of cyclin A, cyclin B and cyclin D as well as the
induction of the cyclin-dependent kinase (CDK) inhibitors p21 and
p27 (Kip1) (12). Although it has
been reported that FHL1 expression is downregulated in OSCC,
including TSCC (13), the
biological function and the underlying molecular mechanisms of FHL1
in TSCC remain to be elucidated.
The present study aimed to investigate the function
and mechanism of FHL1 in TSCC. Cell proliferation and soft agar
assays were performed to detect whether FHL1 regulated
anchorage-dependent and -independent growth of TSCC cells. The
effects of FHL1 on cell migration and invasion were also examined
by wound healing and Transwell assays. In addition, cell cycle
assay, western blotting and reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis were performed to
detect the regulatory effects of FHL1 on the cell cycle, and the
expression of cell cycle-associated regulators. Finally, a tumor
formation assay was performed to detect the regulation of FHL1 on
TSCC cell proliferation in vivo.
Materials and methods
Plasmids and small-interfering RNAs
(siRNAs)
The expression vector for FLAG-tagged FHL1 has been
described previously (10). The
cDNA target sequences of siRNA for FHL1 were siRNA-1: 5-AAG GAG GTG
CAC TAT AAG AAC-3 and siRNA-2: 5-AAT CTG GCC AAC AAG CGC TTT-3 and
were cloned into the vector pSilencer2.1-U6 (Ambion, Austin, TX,
USA), respectively. All plasmids were verified by DNA
sequencing.
Cell culture and transfection
The human TSCC Tca8113 cell line (Cell Institute,
Chinese Academy of Sciences, Shanghai, China) and SCC6 cell line
(provided by Dr Jing Sun, Department of Oral and Maxillofacial
Surgery, Hospital of Stomatology, Tongji University, Shanghai,
China) at passage 20 were cultured in Dulbecco's modified Eagle's
medium (DMEM) supplemented with 10% fetal bovine serum at 37°C in
an humidified atmosphere of 5% CO2 in air. For the
transfection assay, cells were seeded in 24-well or 6-well plates
and transfected with the indicated plasmids using Lipofectamine
2000 according to the manufacturer's instructions (Invitrogen Life
Technologies, Carlsbad, CA, USA).
Anchorage-dependent and -independent
assays
Anchorage-dependent cell proliferation was analyzed
by crystal violet assay as described previously (14). For the anchorage-independent growth
assay, cells (1×103) were seeded in a 12-well plate,
with a bottom layer of 0.7% low-melting temperature agar in DMEM
and a top layer of 0.35% agar in DMEM. Colonies were scored after 5
weeks of growth.
Western blotting
Cells were lysed in radioimmunoprecipitation assay
buffer (50 mM Tris-HCl pH 7.4, 150 mM NaCl, 1% NP40, 0.1% SDS, 0.5%
DOC, 1 mM PMSF, and supplemented with a protease inhibitor
cocktail) for 30 min on ice. The protein concentration was
determined using a bicinchoninic acid assay (Pierce Biotechnology,
Inc., Rockford, IL, USA). Equivalent quantities of protein were
separated by 10% SDS-PAGE and blotted onto a nitrocellulose
membrane (GE Healthcare, Amersham, UK). The membranes were blocked
in Tris-buffered saline containing Tween (TBST) supplemented with
10% nonfat milk for 1 h at room temperature. The membranes were
then incubated with primary antibodies, diluted in TBST containing
10% nonfat milk. Following extensive washing with TBST, the
membranes were incubated with a horseradish peroxidase-conjugated
secondary antibody, followed by chemiluminescent detection with ECL
detection reagent, according to the manufacturer's instructions
(Pierce Biotechnology, Inc.). Images were captured and analysed
using the Tanon-5200 Chemiluminescent imaging system (Tanon,
Shanghai, China). The antibodies used in the present study were as
follows: Rabbit polyclonal anti-human FHL1 (1:200; cat. no.
sc-28691; Santa Cruz Biotechnology, Inc. Dallas, TX, USA), rabbit
polyclonal anti-human GAPDH (1:5,000; cat. no. sc-25778; Santa Cruz
Biotechnology, Inc.), rabbit monoclonal anti-human cyclin D
(1:1,000; cat. no. ab134175; Abcam, Cambridge, UK), rabbit
polyclonal anti-human cyclin E (1:500; cat. no. ab101324; Abcam),
and horseradish peroxidase-conjugated goat anti-rabbit
immunoglobulin G secondary antibody (1:10,000; cat. no. sc-2004;
Santa Cruz Biotechnology, Inc.).
Cell migration and invasion assays
Wound healing assays were used to determine cell
migration. Briefly, cells grown in 6-well plates as confluent
monolayers were mechanically scratched using a 1-ml pipette tip to
create the wound. Cells were washed with phosphate-buffered saline
(PBS) and the debris was removed. Cells were cultured for 24 h in
DMEM without serum to allow wound healing. A cell invasion assay
was performed using a Transwell chamber according to the
manufacturer's instructions (Corning Inc., Corning, NY, USA). Cells
invaded through the Matrigel membrane were fixed with 4%
paraformaldehyde and stained with crystal violet 24 h after
seeding.
RT-qPCR
Total RNA was isolated using TRIzol reagent
(Invitrogen Life Technologies) and reverse transcribed using
SuperScript II Reverse Transcriptase (Invitrogen Life
Technologies). qPCR was performed using the following primers:
Cyclin D, sense GCT TCC TCT CCA GAG TGATC and antisense GTC CAT GTT
CTG CTG GGCCT; cyclin E, sense GAA GAT TCC TAT GGA AGA CAGAC and
antisense GCA CAC TGG TGA CAA CTGTC; FHL1, sense GAA GTG TGC TGG
ATG CAAGA and antisense GGG GGC TTC CTA GCT TTAGA; β-actin, sense
ATC ACC ATT GGC AAT GAGCG and antisense TTG AAG GTA GTT TCG
TGGAT.
Animal experiments
A total of 10 BALB/c nude mice were purchased from
Vital River Laboratories (Beijing, China). The mice were housed
under specific pathogen-free conditions and fed a normal diet.
Tca8113 (1×107) cells were subcutaneously inoculated
into the right flank of the 5-week old nude mice. Tumor size was
monitored every week by measuring length and width with a caliper.
Tumor volume was calculated using the following formula:
Width2 × length / 2. The mice were sacrificed by
cervical dislocation 4 weeks later, the tumors were dissected out
and immunohistochemistry was performed to detect the expression of
FHL1. The present study was approved by the ethics committee of
Anqing Municipal Hospital of Anhui Medical University (Anqing,
China).
Statistical analysis
Statistical significance in the cell growth assays
between the control group and FHL1 overexpression or the knockdown
group was examined by two-tailed Student's t-test. Statistical
calculations were performed using SPSS 13.0 (SPSS, Inc., Chicago,
IL, USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
FHL1 inhibits TSCC cell proliferation in
vitro
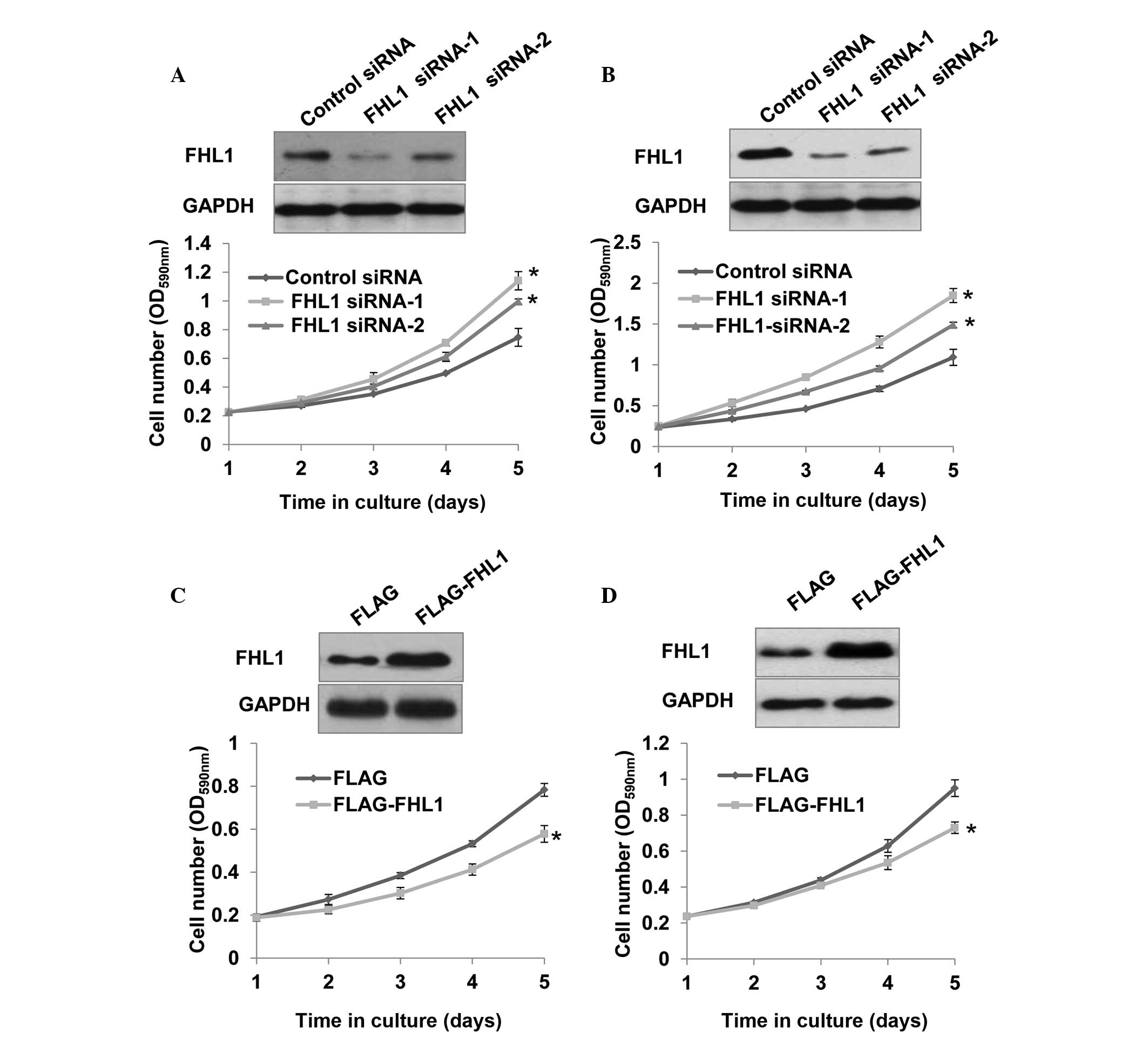
To investigate the effect of FHL1 on the
proliferation of the TSCC cell lines Tca8113 and SCC6, stable cell
lines expressing control siRNA, FHL1 siRNA-1 and FHL1 siRNA-2 were
constructed. FHL1 protein expression was downregulated in cells
transfected with FHL1 siRNA-1 and FHL1 siRNA-2, particularly with
FHL1 siRNA-1, compared with cells transfected with control siRNA
(Fig. 1A and B). Notably, FHL1
knockdown with the two siRNAs, particularly FHL1 siRNA-1, markedly
promoted the proliferation of Tca8113 and SCC6 cells (Fig. 1A and B). By contrast,
overexpression of FHL1 inhibited Tca8113 and SCC6 cell
proliferation (Fig. 1C and D).
FHL1 siRNA-1 was used in the following experiments due to its more
efficient knockdown of endogenous FHL1 and more notable effect on
TSCC cell proliferation.
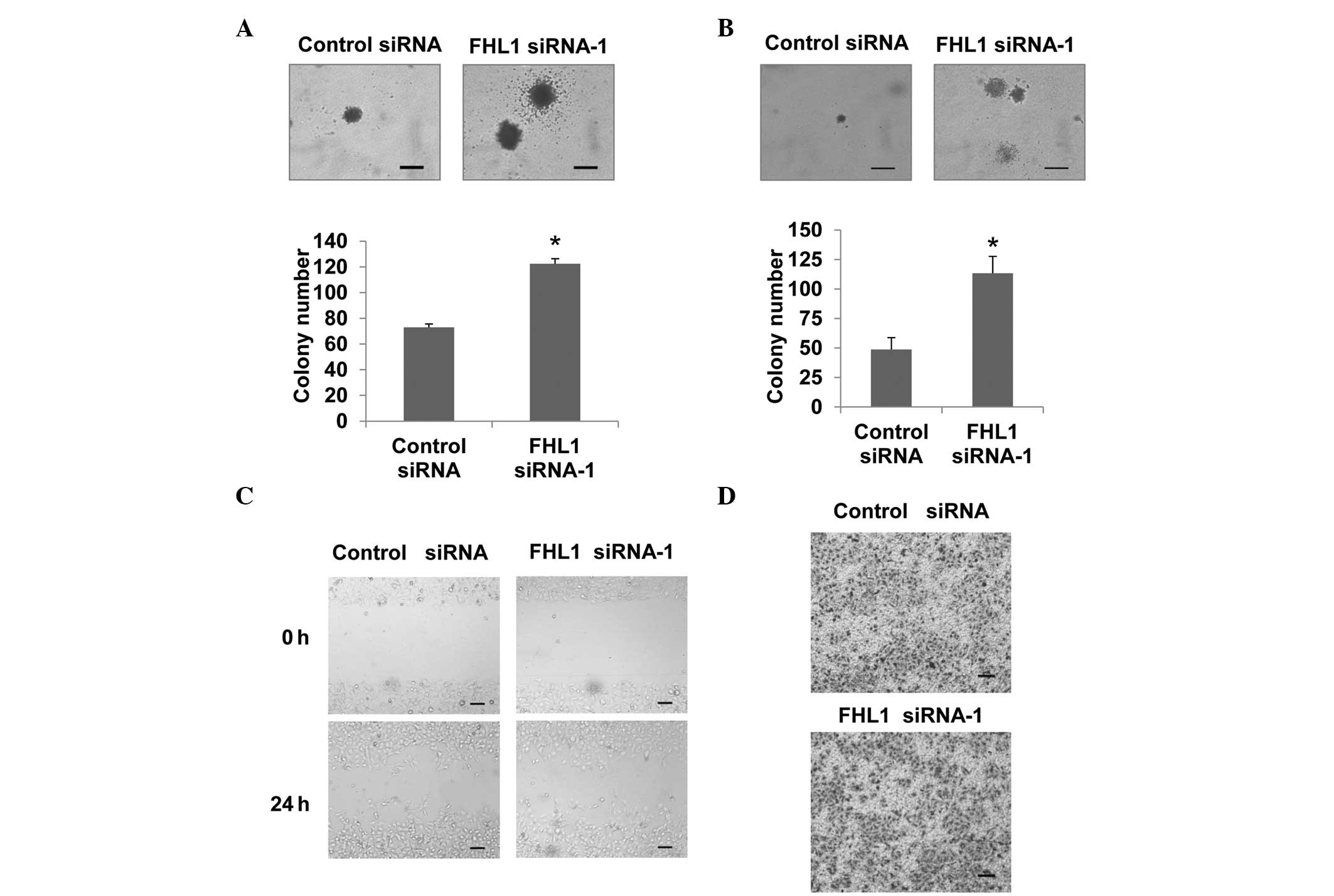
Since anchorage-independent growth is one of the
hallmarks of cancer cells, the effect of FHL1 on this phenotype was
investigated using a soft agar assay. Knockdown of FHL1 increased
anchorage-independent Tca8113 cell growth based on the size and the
number of colonies (Fig. 2A).
Similar results were observed in SCC6 cells (Fig. 2B). Taken together, these results
demonstrated that FHL1 inhibits anchorage-dependent and
-independent growth of TSCC cells.
FHL1 does not affect Tca8113 cell
migration and invasion
Since TSCC is characterized by a high metastatic
rate (15,16), the present study aimed to determine
whether FHL1 modulates TSCC migration and invasion in Tca8113
cells. Wound healing assays were performed to evaluate cell
migration ability. Knockdown of FHL1 did not affect Tca8113 cell
migration (Fig. 2C). In addition,
FHL1 did not affect cell invasion according to the transwell assay
(Fig. 2D).
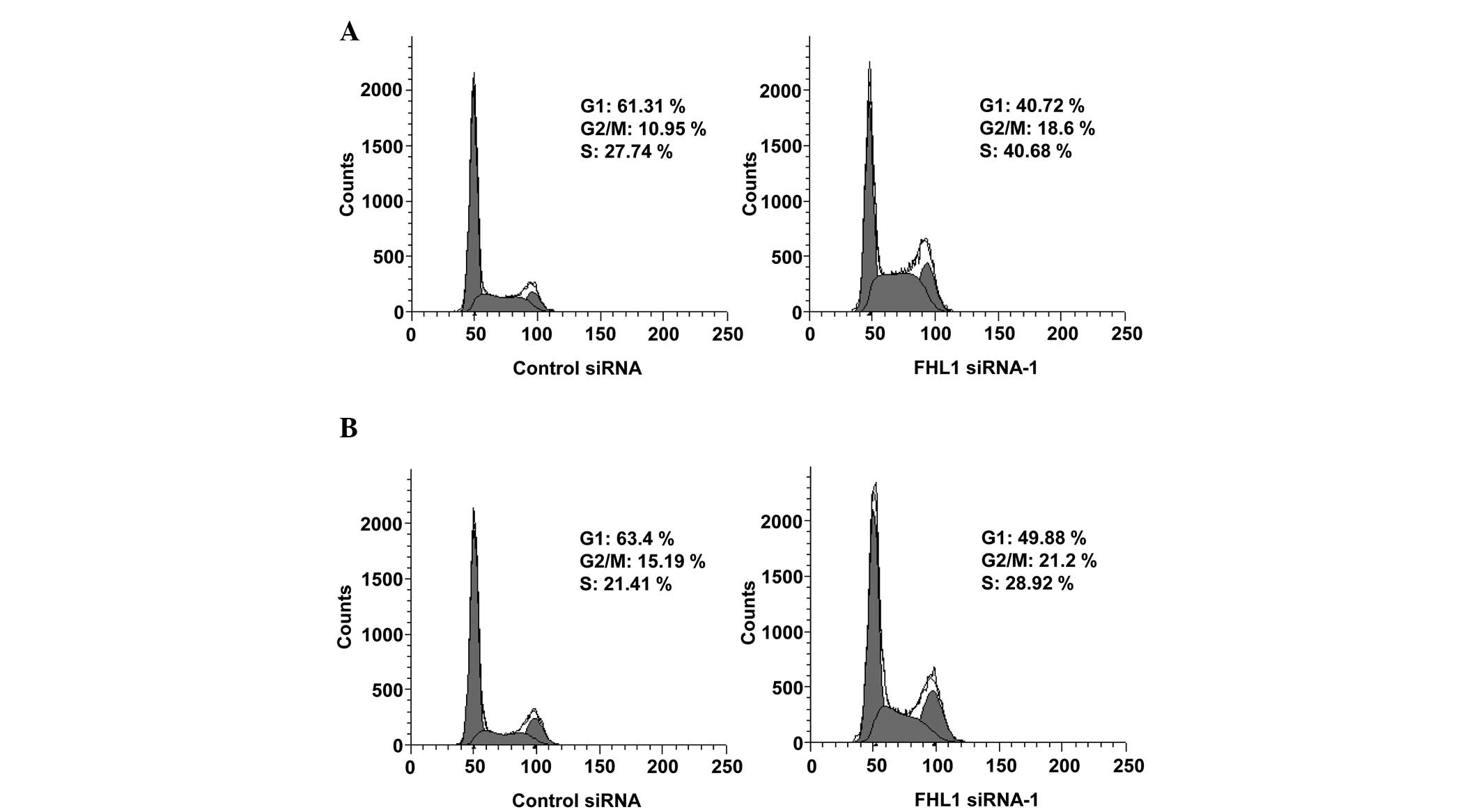
FHL1 induces G1/S cell cycle arrest in
TSCC cells
To elucidate the mechanism underlying the growth
inhibition of TSCC cells by FHL1, the effect of FHL1 on the cell
cycle of Tca8113 and SCC6 cells was examined. In comparison with
control siRNA, the reduction of endogenous FHL1 cells using FHL1
siRNA caused a clear decrease in the proportion of cells in the G1
phase (from 61.31 to 40.72% for Tca8113 cells; from 63.4 to 49.88%
for SCC6 cells) and an increase in the proportion of cells in S
phase (from 27.74 to 40.68% for Tca8113 cells; from 21.41 to 28.92%
for SCC6 cells; Fig. 3A and B).
Taken together, these data suggest that FHL1 induces G1/S cell
cycle arrest in TSCC cells.
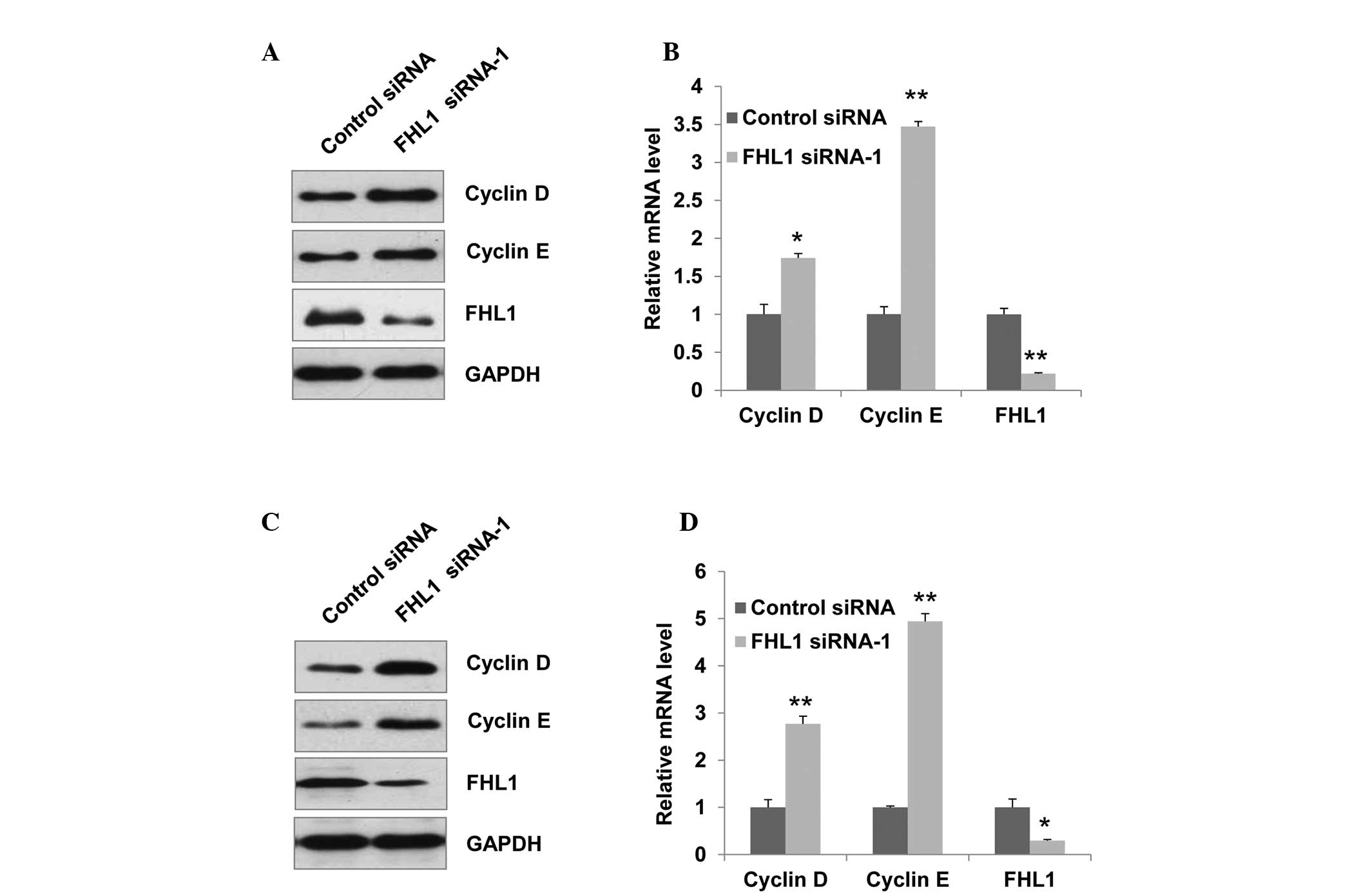
FHL1 regulates the expression of cyclin D
and cyclin E in TSCC cells
To further elucidate the molecular mechanism by
which FHL1 expression induces G1/S cell cycle arrest, the
expression of cyclin D and cyclin E, which promote G1/S transition,
was determined by western blot analysis. Knockdown of endogenous
FHL1 in Tca8113 cells increased the expression of cyclin D and
cyclin E (Fig. 4A). Consistent
with the results of FHL1 modulation of protein expression, FHL1
knockdown increased the expression of cyclin D and cyclin E at the
mRNA level (Fig. 4B). Similar
results were observed in SCC6 cells (Fig. 4C and D).
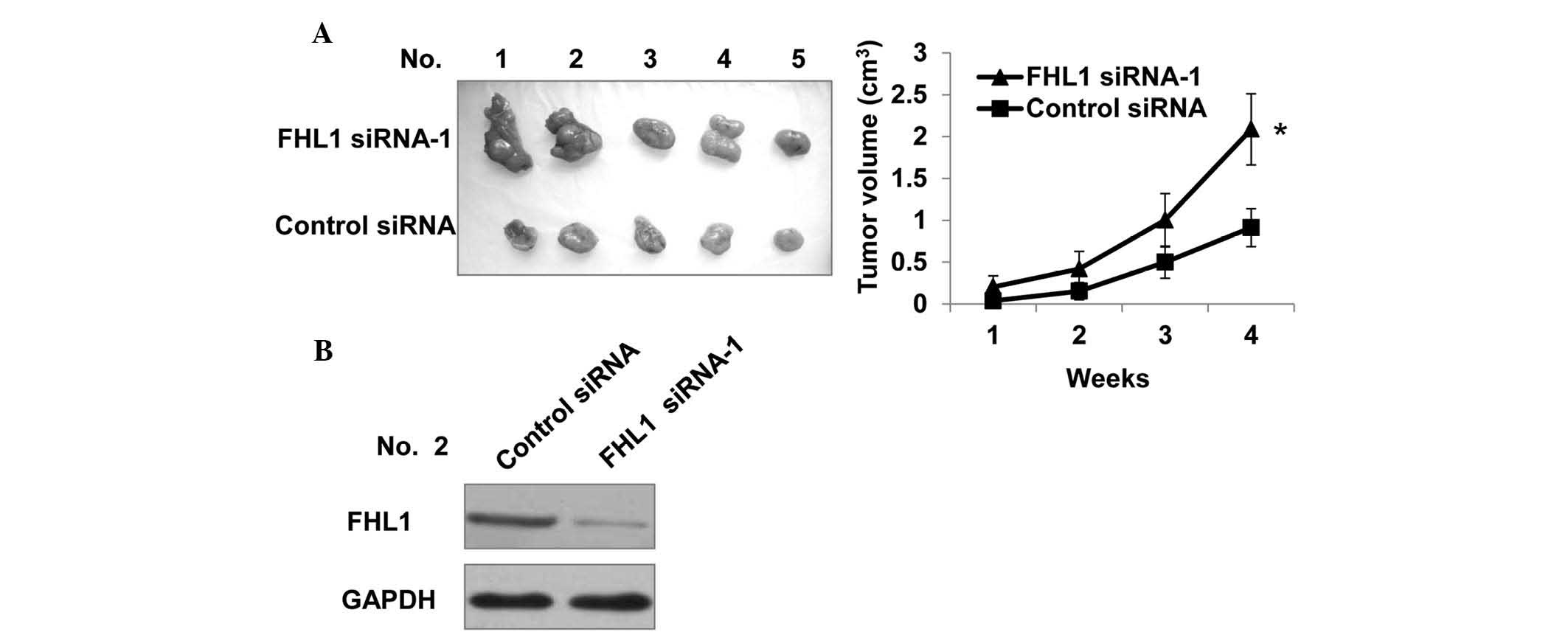
Knockdown of FHL1 promotes TSCC cell
growth in nude mice
Subsequently, the effect of FHL1 on TSCC cell growth
in nude mice was determined. A total of 10 mice were injected with
Tca8113 cells stably transfected with control siRNA or FHL1
siRNA-1. As shown in Fig. 5A, all
mice inoculated with Tca8113 cells developed tumors after 4 weeks.
Notably, knockdown of FHL1 increased Tca8113 tumor growth, which is
consistent with the stimulatory role of FHL1 knockdown in cell
growth in vitro. As expected, the tumors in mice inoculated
with Tca8113 cells expressing FHL1 siRNA had reduced protein levels
of FHL1 compared with control siRNA (Fig. 5B).
Discussion
The present study provided for the first time, to
the best of our knowledge, several lines of evidence demonstrating
a novel role for FHL1 in TSCC. Firstly, FHL1 inhibits
anchorage-dependent and -independent growth of TSCC cells.
Secondly, knockdown of FHL1 inhibits G1/S cell cycle arrest,
accompanied by upregulation of important cell cycle regulators,
including cyclin D and cyclin E. Finally, knockdown of endogenous
FHL1 promotes tumor growth in nude mice. These results suggest that
FHL1 functions as a tumor suppressor in TSCC and that FHL1 may be a
useful target for TSCC gene therapy.
FHL1 does not affect TSCC cell migration and
invasion, which is consistent with a previous study demonstrating
that expression of FHL1 does not correlate with node metastasis
(13). These findings indicate
that FHL1 may be important in the development of TSCC, but not in
the progression of TSCC.
Knockdown of endogenous FHL1 upregulates the protein
and mRNA levels of cyclin D and cyclin E. In normal human cells,
cellular division is an ordered, tightly regulated process,
involving multiple cell cycle checkpoints that ensure genomic
integrity. Cyclins and their associated CDKs are the central
machinery that govern cell cycle progression (17,18).
During the G1 phase, cyclin D binds and activates CDK4 and CDK6,
which leads to partial inactivation of RB, RBL1 and RBL2 proteins.
In addition, CDK2-cyclin E complexes further phos-phorylate these
proteins and drive the G1/S transition. Altered regulation of the
cell cycle is a hallmark of several types of human cancer (19). Overexpression of cyclin D is common
in human cancers of epithelial cell origin (20). In tongue tumors, cyclin D gene
amplification was detected in 88% of the tumors (21). Patients with head and neck squamous
cell carcinoma (HNSCC) that were strongly positive for cyclin D had
reduced overall and disease-free survival. Cyclin D may be used as
a predictor of long-term outcomes for patients with HNSCC (22). Various types of cancer, including
breast cancer, lung cancer, cervical cancer, endometrial cancer and
gastrointestinal cancer, overexpress cyclin E protein or mRNA
(23). In addition, cyclin E
overexpression has been proposed as a marker of poor clinical
outcome in breast cancer (24).
The fact that FHL1 can regulate cyclin D and cyclin E suggests a
critical role for FHL1 in cell cycle regulation and cellular
division.
It has been reported that FHL1 is frequently
downregulated in primary OSCC tissues compared with the
corresponding normal oral tissues (13). One of the mechanisms of FHL1
downregulation in OSCC is hypermethylation of CpG islands within
FHL1 gene promoter regions, which is also found in bladder cancer
(13,25). Due to the importance of FHL1 in the
regulation of TSCC cell growth, it would be interesting to
investigate whether the FHL1 promoter is methylated in TSCC and to
determine other mechanisms underlying FHL1 downregulation in
TSCC.
Acknowledgments
This study was supported by the National Natural
Science Foundation of China (grant nos. 31200565, 31071174 and
81330053), the Beijing Natural Science Foundation (grant no.
5132027) and the Beijing Nova Program (grant no. Z131102000413034).
Anqing Municipal Hospital of Anhui Medical University and Beijing
Institute of Biotechnology contributed equally to this work.
References
|
1
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jemal A, Siegel R, Ward E, Murray T, Xu J
and Thun MJ: Cancer statistics, 2007. CA Cancer J Clin. 57:43–66.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Heaton CM, Durr ML, Tetsu O, van Zante A
and Wang SJ: TP53 and CDKN2a mutations in never-smoker oral tongue
squamous cell carcinoma. Laryngoscope. 124:E267–E273. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Thavaraj S, Stokes A, Mazuno K, et al:
Patients with HPV-related tonsil squamous cell carcinoma rarely
harbour oncogenic HPV infection at other pharyngeal sites. Oral
Oncol. 50:241–246. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J and
Thun MJ: Cancer statistics, 2009. CA Cancer J Clin. 59:225–249.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Matthews JM, Lester K, Joseph S and Curtis
DJ: LIM-domain-only proteins in cancer. Nat Rev Cancer. 13:111–122.
2013. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cowling BS, Mcgrath MJ, Nguyen MA, et al:
Identification of FHL1 as a regulator of skeletal muscle mass:
Implications for human myopathy. J Cell Biol. 183:1033–1048. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li X, Jia Z, Shen Y, Ichikawa H, Jarvik J,
Nagele RG and Goldberg GS: Coordinate suppression of Sdpr and Fhl1
expression in tumors of the breast, kidney and prostate. Cancer
Sci. 99:1326–1333. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ding L, Niu C, Zheng Y, et al: FHL1
interacts with oestrogen receptors and regulates breast cancer cell
growth. J Cell Mol Med. 15:72–85. 2011. View Article : Google Scholar
|
|
10
|
Ding L, Wang Z, Yan J, et al: Human
four-and-a-half LIM family members suppress tumor cell growth
through a TGF-beta-like signaling pathway. J Clin Invest.
119:349–361. 2009.PubMed/NCBI
|
|
11
|
Sakashita K, Mimori K, Tanaka F, et al:
Clinical significance of loss of Fhl1 expression in human gastric
cancer. Ann Surg Oncol. 15:2293–2300. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Niu C, Liang C, Guo J, et al:
Downregulation and growth inhibitory role of FHL1 in lung cancer.
Int J Cancer. 130:2549–2556. 2012. View Article : Google Scholar
|
|
13
|
Koike K, Kasamatsu A, Iyoda M, et al: High
prevalence of epigenetic inactivation of the human four and a half
LIM domains 1 gene in human oral cancer. Int J Oncol. 42:141–150.
2013.
|
|
14
|
Cheng L, Li J, Han Y, et al: PES1 promotes
breast cancer by differentially regulating ERα and ERβ. J Clin
Invest. 122:2857–2870. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cao Z, Xiang J and Li C: Expression of
extracellular matrix metalloproteinase inducer and enhancement of
the production of matrix metalloproteinase-1 in tongue squamous
cell carcinoma. Int J Oral Maxillofac Surg. 38:880–885. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li S, Jiao J, Lu Z and Zhang M: An
essential role for N-cadherin and beta-catenin for progression in
tongue squamous cell carcinoma and their effect on invasion and
metastasis of Tca8113 tongue cancer cells. Oncol Rep. 21:1223–1233.
2009.PubMed/NCBI
|
|
17
|
Malumbres M and Barbacid M: Cell cycle,
CDKs and cancer: A changing paradigm. Nat Rev Cancer. 9:153–166.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hochegger H, Takeda S and Hunt T:
Cyclin-dependent kinases and cell-cycle transitions: Does one fit
all? Nat Rev Mol Cell Biol. 9:910–916. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Musgrove EA, Caldon CE, Barraclough J,
Stone A and Sutherland RL: Cyclin D as a therapeutic target in
cancer. Nat Rev Cancer. 11:558–572. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mahdey HM, Ramanathan A, Ismail SM,
Abraham MT, Jamaluddin M and Zain RB: Cyclin D1 amplification in
tongue and cheek squamous cell carcinoma. Asian Pac J Cancer Prev.
12:2199–2204. 2011.
|
|
22
|
Rasamny JJ, Allak A, Krook KA, et al:
Cyclin D1 and FADD as biomarkers in head and neck squamous cell
carcinoma. Otolaryngol Head Neck Surg. 146:923–931. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hwang HC and Clurman BE: Cyclin E in
normal and neoplastic cell cycles. Oncogene. 24:2776–2786. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Keyomarsi K, Tucker SL, Buchholz TA, et
al: Cyclin E and survival in patients with breast cancer. N Engl J
Med. 347:1566–1575. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Matsumoto M, Kawakami K, Enokida H, et al:
CpG hypermeth-ylation of human four-and-a-half LIM domains 1
contributes to migration and invasion activity of human bladder
cancer. Int J Mol Med. 26:241–247. 2010.PubMed/NCBI
|