Introduction
Silicosis is a form of lung disease, which is caused
by the inhalation and deposition of occupational dust containing
silicon dioxide (SiO2) (1). Silicosis is considered to be one of
the most severe occupational diseases, with no effective treatment
(2–7). Therefore, elucidation of the
mechanisms underlying the development and progression of silicosis
is critical in order to develop effective therapeutic methods. At
present, the mechanisms of silicosis remain to be fully elucidated.
Previous studies have indicated that pulmonary alveolar macrophages
are pivotal in the pathological process of silicosis (8,9).
Resident and recruited pulmonary macrophages have been observed to
be in intimate contact with silica at the beginning of deposition
and throughout the process, with the particles remaining in the
lung (9). These macrophages appear
to be stimulated to secrete mediator substances, which alter the
function and behavior of other cells. Tumor necrosis factor α
(TNF-α) and interleukin-1β (IL-1β) are two mediators, of which the
levels are commonly increased in airway inflammation, and are
involved in the initiation and progression of silicosis (2,8,10,11).
MicroRNAs (miRNAs) are small regulatory RNAs, which
control gene expression by translational suppression and
destabilization of target mRNAs (12). Numerous miRNAs have been implicated
in various biological processes, including miR-125b and let-7,
which are associated with cell proliferation control, miR-15a and
miR-16-1 that contribute to heart disease, lung disease and
oncogenesis, and miR-155 and miR17 that function as tumor
suppressors or as oncogenes (13,14).
There is increasing evidence that miRNAs are key regulators of
genes in inflammatory responses and fibrosis of the lung (15–18).
Aberrant expression of miR-146a and miR-181b have been reported to
be involved in the formation of lung cancer (19,20).
However, studies investigating their roles in silicosis have been
limited (21,22).
In our previous study, a large-scale screen for
miRNAs potentially involved in experimental silicosis in rats was
performed, and it was revealed that miR-146a is upregulated and
miR-181b is downregulated in response to stimulation with
SiO2 (21). However,
the specific roles of these two miRNAs and the underlying mechanism
of action in silicosis remain to be elucidated. Therefore, the aim
of the present study was to determine the roles of miR-146a and
miR-181b in inflammatory responses, and determine their effect on
the expression of the TNF-α and IL-1β inflammatory chemokines in
SiO2-induced NR8383 rat macrophages.
Materials and methods
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) for miRNA
All experimental procedures in the present study
were approved by the Experimental Animal Ethics Committee of
Sichuan University (Chengdu, China). RT-qPCR for miRNA was
performed using an EpiScript™ Reverse Transcriptase kit (EpiCentre,
Palmerston North, New Zealand), as reported previously (21). A total of 20 male Sprague Dawley
rats (age, 5–7 weeks; weight, 160–200 g; Dashuo Center of
Experimental Animals, Chengdu, China), were used for the
experiments of the present study. The animals were housed in
standard conditions at 22±1°C with relative humidity of 59% and a
12 h light/dark cycle, and were provided with ad libitum
access to food and water. The rats were randomly divided into two
experimental groups: A control group (n=10) and a silicosis group
(n=10). The rats were then sacrificed through loss of blood under
10% chloral hydrate anesthesia. Following sacrifice on day 40, the
lungs were harvested for total RNA isolation and histological
analysis. Briefly, 5 µm sections of lung tissue were
obtained from the inferior lobe of the left lungs and fixed with
10% formalin (Kelon, Beijing, China), embedded in paraffin (Beijing
Solarbio Science & Technology Co., Ltd., Beijing, China) and
stained with hematoxylin and eosin (Beijing Solarbio Science &
Technology Co., Ltd.) and Masson (Beijing Solarbio Science &
Technology Co., Ltd.) dyes for histological examination of the
collagen fibers. The tissue samples were cryopulverized using
Biopulverizer™ (Biospec Products, Inc., Bartlesville, OK, USA) and
homogenized using a Mini-Bead-Beater-16 (BioSpec, Shanghai Yuan
Sheng Co., Ltd., Shanghai, China). Total RNA was isolated from the
tissue samples using TRIzol® (Invitrogen Life
Technologies, Carlsbad, CA, USA) and further purified using an
RNeasy mini kit (Qiagen, Hilden, Germany) according to the
manufacturer's instructions. Total RNA (600 ng), 10 µM
stem-loop RT primer (Table I), 10X
RT buffer, 2.5 mM each of dATP, dGTP, dCTP and dTTP (HyTest Ltd.,
Turku, Finland), 10 U/µl reverse transcriptase and 40
U/µl RNase inhibitor (EpiCentre) were subjected to RT
reactions. PCR Master Mix (Qiagen) and 0.4 µM of each primer
(Table I) were used for RT-qPCR.
The reactions were conducted with the following thermocycling
conditions: 95°C for 5 min, followed by 40 cycles of 95°C for 10
sec, and 60°C for 60 sec. Each sample was normalized to endogenous
U6 RNA content. RT-qPCR was performed using a CFX96 Touch™
Real-Time PCR Detection system (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). Each experiment was performed in triplicate.
The Data were analyzed using Microsoft Excel. The relative
expression levels of miR-146a were evaluated by the
2−ΔΔCT method.
 | Table IOligonucleotide primers used in the
present study. |
Table I
Oligonucleotide primers used in the
present study.
| Primer | Primer sequence
|
|---|
| Reverse
transcription | Quantitative
polymerase chain reaction |
|---|
| U6 |
5′-CGCTTCACGAATTTGCGTGTCAT-3′ | F:
5′-GCTTCGGCAGCACATATACTAAAAT-3′
R: 5′-CGCTTCACGAATTTGCGTGTCAT-3′ |
| Rno-miR-146a |
5′-GTCTTATCCAGTGCGTGTCGTGGAGTCG
GCAATTGCACTGGATACGACAACCA-3′ | GSP:
5′-CAGTGCGTGTCGTGGAGT-3′
R: 5′-GGGTGAGAACTGAATTCCA-3′ |
| Rno-miR-181b |
5′-GTCGTATCCAGTGCGTGTCGTGGAGTCG
GCAATTGCACTGGATACGACAGCCTAT-3′ | GSP:
5′-GGGTTCAAGTAATCCAGG-3′
R: 5′-TGCGTGTCGTGGAGTC′-3′ |
Cell culture, transfection and
SiO2 treatment
NR8383 cells were purchased from Shanghai Institute
of Biochemistry and Cell Biology and were cultured in Ham's F12K
medium (Sigma-Aldrich, St. Louis, MO, USA) supplemented with 15%
fetal bovine serum (Gibco Life Technologies, Grand Island, NY,
USA). The NR8383 cells were seeded in triplicate into 6-well plates
at a density of 1×105 cells/well, and were grown to
30-50% confluence at 37°C in an atmosphere containing 5%
CO2. Turbofect transfection reagent from Thermo Fisher
Scientific (Pittsburgh, PA, USA) was used for transfection,
according to the manufacturer's instructions. The miRNA mimic
control, miR-146a mimic, miR-181b mimic, miRNA inhibitor control,
miR-146a inhibitor and miR-181b inhibitor (Guangzhou RiboBio Co.
Ltd., Guangzhou, China) were transfected at a final concentration
of 25 nM. Following 24 h of transfection, >90% of the cells
exhibited green fluorescence under an inverted fluorescence
microscope (Nikon ECLIPSE Ti-U; Nikon, Tokyo, Japan).. The cells
were then treated with 20 µg/cm2 SiO2
(cat. no. S5631; Sigma-Aldrich). The silica content of the quartz
dust was >99%, particle size of the dust was 0.5–10 µm,
and 80% of the dust particles were 1–5 µm. The cells and
culture supernatants were collected following SiO2
treatment for 12 h at 37°C in an atmosphere containing 5%
CO2. RT-qPCR, western blotting and ELISA were then
performed to detect the expression levels of TNF-α and IL-1β.
RT-qPCR for mRNA
The NR8383 cells were suspended at ~1×107
cells/well in TRIzol® reagent (Invitrogen Life
Technologies, Carlsbad, CA, USA), and the total RNA was isolated,
according to the manufacturer's instructions. Total RNA (1
µg) was subjected to reverse transcription using the
PrimeScript™ RT reagent kit with gDNA Eraser (Takara Bio, Inc.,
Tokyo, Japan). SYBR® Premix Ex Taq™ II (Takara Bio,
Inc.) and the following specific primers were used for RT-qPCR:
TNF-α, forward 5′-CAT GGA TCT CAA AGA CAA CCAA-3′ and reverse
5′-CTC CTG GTA TGA AAT GGC AAA T-3′; IL-1β, forward 5′-CTT CAA ATC
TCA CAG CAG AAT C-3′ and reverse 5′-GCT GTC TAA TGG GAA CAT CAC
A-3′; β-actin, forward 5′-GGA GAT TAC TGC CCT GGC TCC TA-3′ and
reverse 5′-GAC TCA TCG TAC TCC TGC TTG CTG-3′. The PCR reactions
were run using the following thermal cycling parameters: 95°C for
30 sec, followed by 40 cycles of 95°C for 5 sec, 60°C for 15 sec
and 72°C for 45 sec. Each experiment was performed in
triplicate.
Western blot analysis
Total protein (30 µg) was extracted using a
total protein extraction kit (Nanjing KeyGEN Biotech Co., Ltd.,
Nanjing, China) according to the manufacturer's instructions, and
loaded onto 15% SDS-PAGE and transferred onto polyvinylidene
difluoride membranes (EMD Millipore, Billerica, MA, USA) using a
semi-dry blotting system (Invitrogen Life Technologies). The
membranes were blocked with 5% skim milk (w/v) containing 0.2%
Tween-20 (Sigma-Aldrich) for 2 h at room temperature, and then were
incubated overnight with the following primary antibodies in
blocking buffer at 4°C: Polyclonal goat anti-rat TNF-α (1:300; cat.
no. sc-1349; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA)
and polyclonal goat anti-rat IL-1β (1:500; cat. no. sc-1251; Santa
Cruz Biotechnology, Inc.). These were used according to the
manufacturer's instructions. The membranes were washed three times
for 6 min with 1X Tris-buffered saline with Tween-20 were then
incubated with polyclonal rabbit anti-goat secondary antibody
(1:2,000; cat no. ZB-2306; ZSGB-BIO, Beijing, China) for 1 h at
37°C in an atmosphere containing 5% CO2. Following final
washes, the signals were detected using enhanced chemiluminescence
reagents (Beyotime Institute of Biotechnology, Shanghai, China).
The intensity of each signal spot was transformed into digital
data, with auto-background subtraction during spot density
analysis, using the Image-Pro Plus software, version 6.0 (Media
Cybernetics, Inc., Bethesda, MD, USA).
ELISA
The levels of TNF-α and IL-1β in the supernatants
were determined using TNF-α and IL-1β ELISA kits (NeoBioscience
Technology Co., Ltd., Beijing, China), according to the
manufacturer's instructions. Briefly, the samples were diluted and
added to the wells (100 µl) prior to being covered with a
closure plate membrane, and incubated for 90 min at 37°C. The
closure plate membrane was removed and the liquid was carefully
discarded, prior to five washes in phosphate-buffered saline
containing 1% bovine serum albumin and 0.05% Tween 20, each for 30
sec. Horseradish peroxidase-conjugated anti-rat TNF-α or IL-1β
polyclonal antibody (from the TNF-α and IL-1β ELISA kits;
NeoBioscience Technology Co., Ltd.; 100 µl) was added to
each well, except the blank well, and incubated for 60 min at 37°C.
The closure plate membrane was removed, the liquid discarded, and
the plates were dried prior to further washing with washing buffer,
repeated five times for 30 sec. A total of 100 µl Chromogen
Solution B (NeoBioscience Technology Co., Ltd.) was added to each
well, and incubated in the dark for 15 min at 37°C. A total of 100
µl stopping solution (NeoBioscience Technology Co., Ltd.)
was then added to each well to stop the reaction. Absorbance was
measured at 450 nm using a Thermo Scientific Multiskan GO (Thermo
Fisher Scientific, Pittsburgh, PA, USA) as compared with the blank
well 15 min following the end of the reaction.
Statistical analysis
R software from the The Comprehensive R Archive
Network (http://cran.r-project.org/) was used
to perform all statistical analyses. A one-way analysis of variance
and two-tailed Student's t-test were used to analyze the data.
P<0.05 (two-tailed) was considered to indicate a statistically
significant difference.
Results
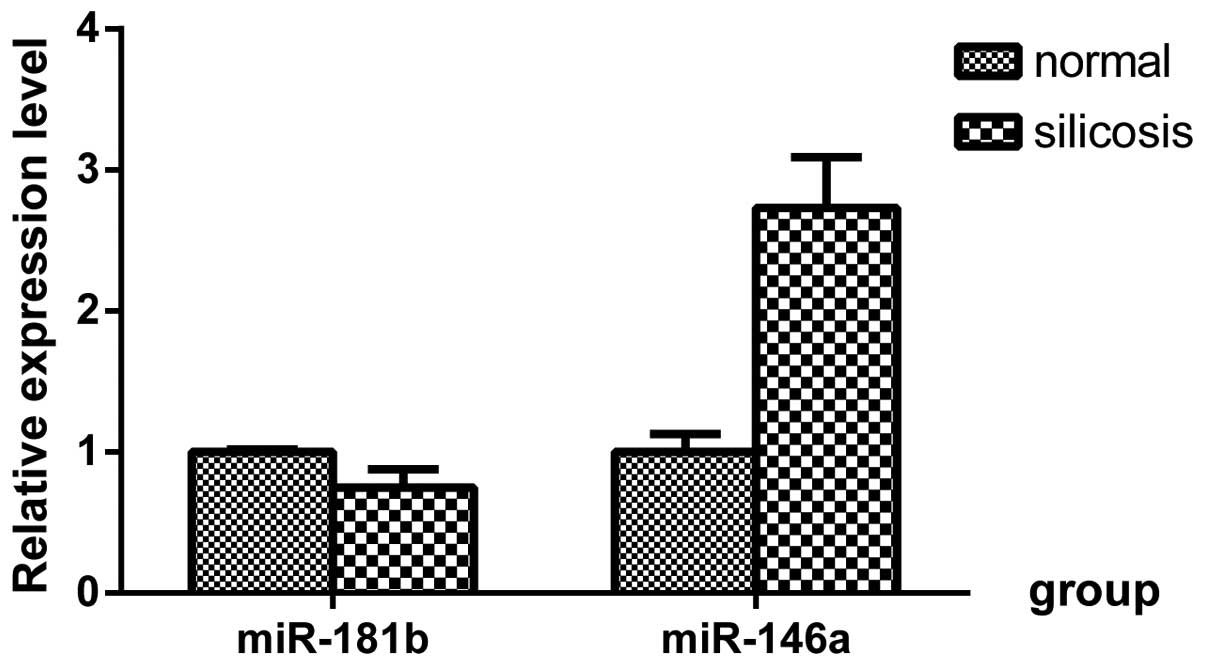
Expression levels of miR-146a increase
and miR-181b decrease in SiO2-treated lungs
Typical fibrosis was observed in the
SiO2-treated lungs, which involved infiltration of
macrophages in alveolar spaces and increased collagen deposition
(21). miRNA array analysis of the
total RNA samples was performed on the day
40-SiO2-treated lungs, and these were compared with the
respective controls, as described in our previous study (21). Of 1,890 miRNAs examined, 39 miRNAs
were either up- or downregulated (P<0.05; fold changes >2) in
the SiO2-treated lungs (21). In addition, the results of the
RT-qPCR revealed that the expression of miR-146a was significantly
increased and the expression of miR-181b was significantly
decreased in the SiO2-treated lungs (Fig. 1).
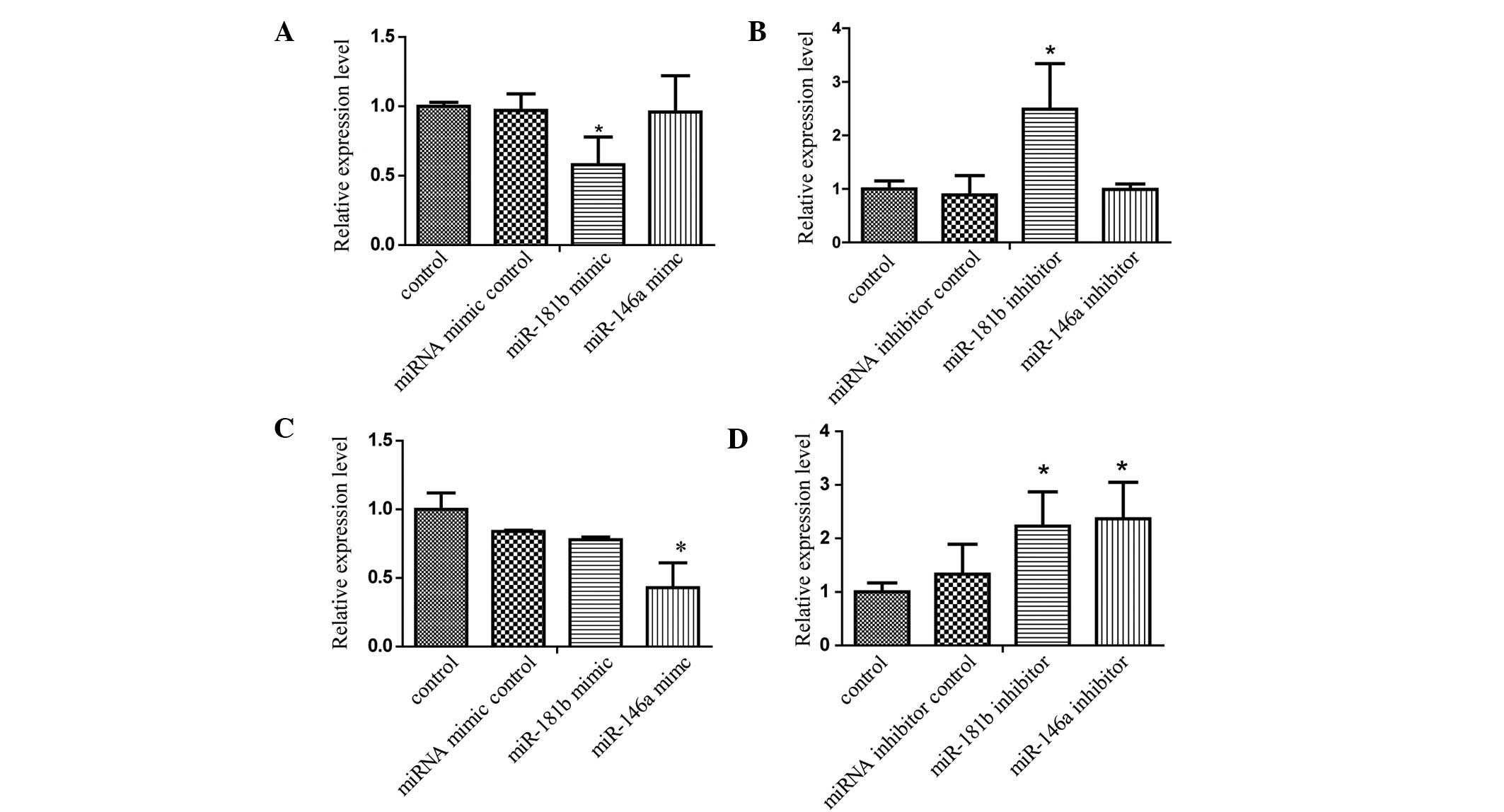
mRNA expression levels of TNF-α and
IL-1β, detected using RT-qPCR, are altered by miRNA
transfection
As shown in Fig.
2A, when the NR8383 cells were transfected with the miR-181b
mimic for 24 h following 12 h of SiO2 treatment, the
expression of TNF-α was significantly decreased, compared with the
miRNA mimic control group (P<0.05). Simultaneously, miR-181b
inhibitors increased the mRNA expression of TNF-α significantly
(P<0.05; Fig. 2B), whereas the
miR-146a mimic and inhibitors had no significant effect on the mRNA
expression levels of TNF-α (Fig. 2A
and B). As shown in Fig. 2C,
12 h following SiO2 treatment of the NR8383 cells
transfected with either the miR-146a mimics, miR-181b mimics or
miRNA mimic control, the miR-146a mimics significantly attenuated
the expression of IL-1β (P<0.05). The expression levels of IL-1β
in response to 12 h of SiO2 treatment, subsequent to
transfection with the miR-146a or miR-181b inhibitors were
significantly increased, compared with those in the inhibitor
control group (P<0.05; Fig.
2D).
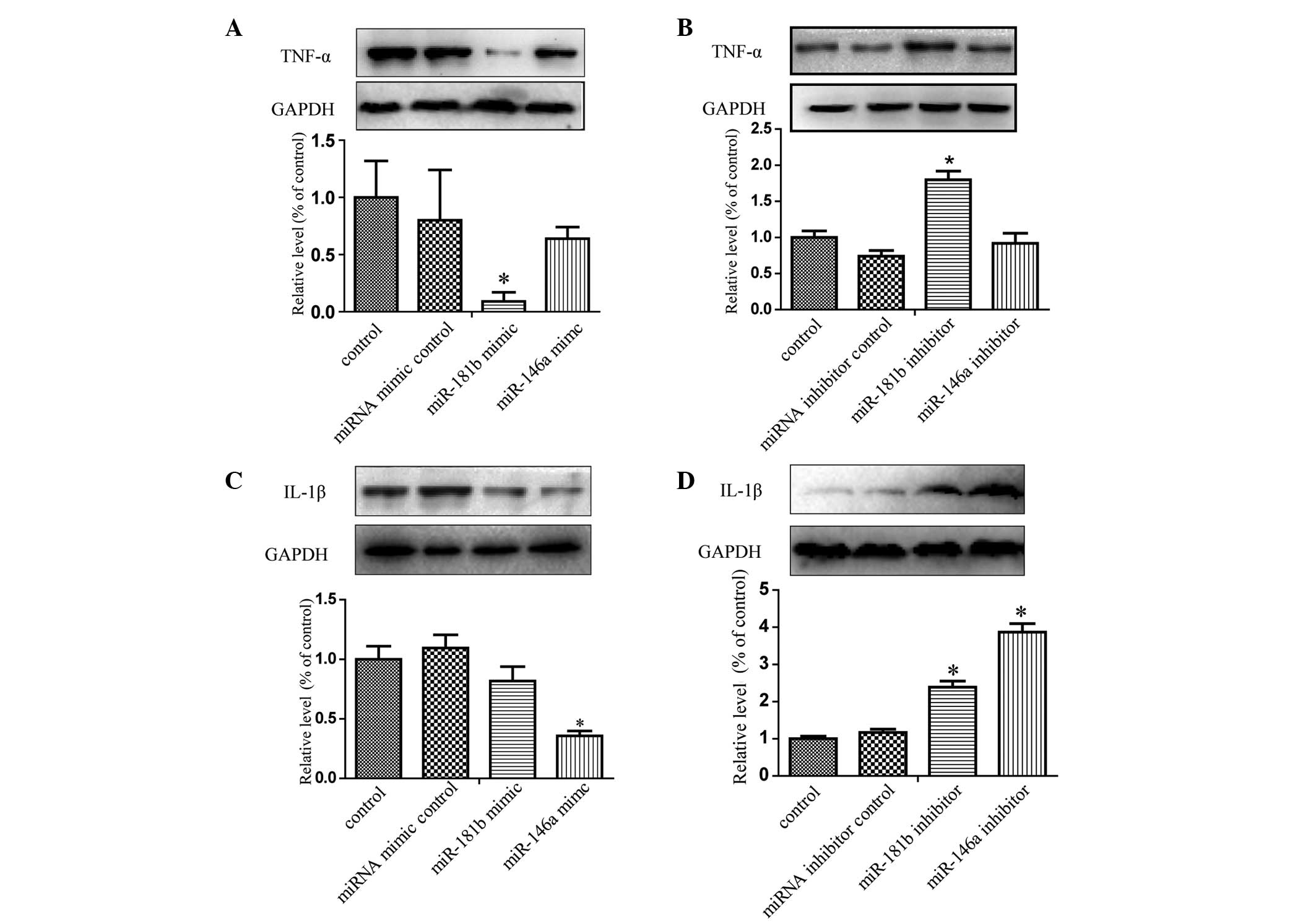
Protein expression levels of TNF-α and
IL-1β, detected using western blot analysis, are significantly
altered by miRNA transfection
The results of the western blot analysis
demonstrated that overexpression of miR-181b following
SiO2 treatment for 12 h significantly reduced the
protein levels of TNF-α, compared with the miRNA mimic control
group (P<0.05; Fig. 3A).
Subsequent to transfection with miR-181b inhibitors, followed by
SiO2 treatment for 12 h, the protein levels of TNF-α
were significantly upregulated (P<0.05; Fig. 3B). However, the miR-146a mimics or
inhibitors had no significant effect on the protein expression
levels of TNF-α. The NR8383 cells transfected with the miR-146a
mimic exhibited lower expression levels of IL-1β in response to the
subsequent SiO2 stimulation (P<0.05; Fig. 3C) and IL-1β was significantly
upregulated in the NR8383 cells, which were transfected with the
miR-146a or miR-181b inhibitors (P<0.05; Fig. 3D).
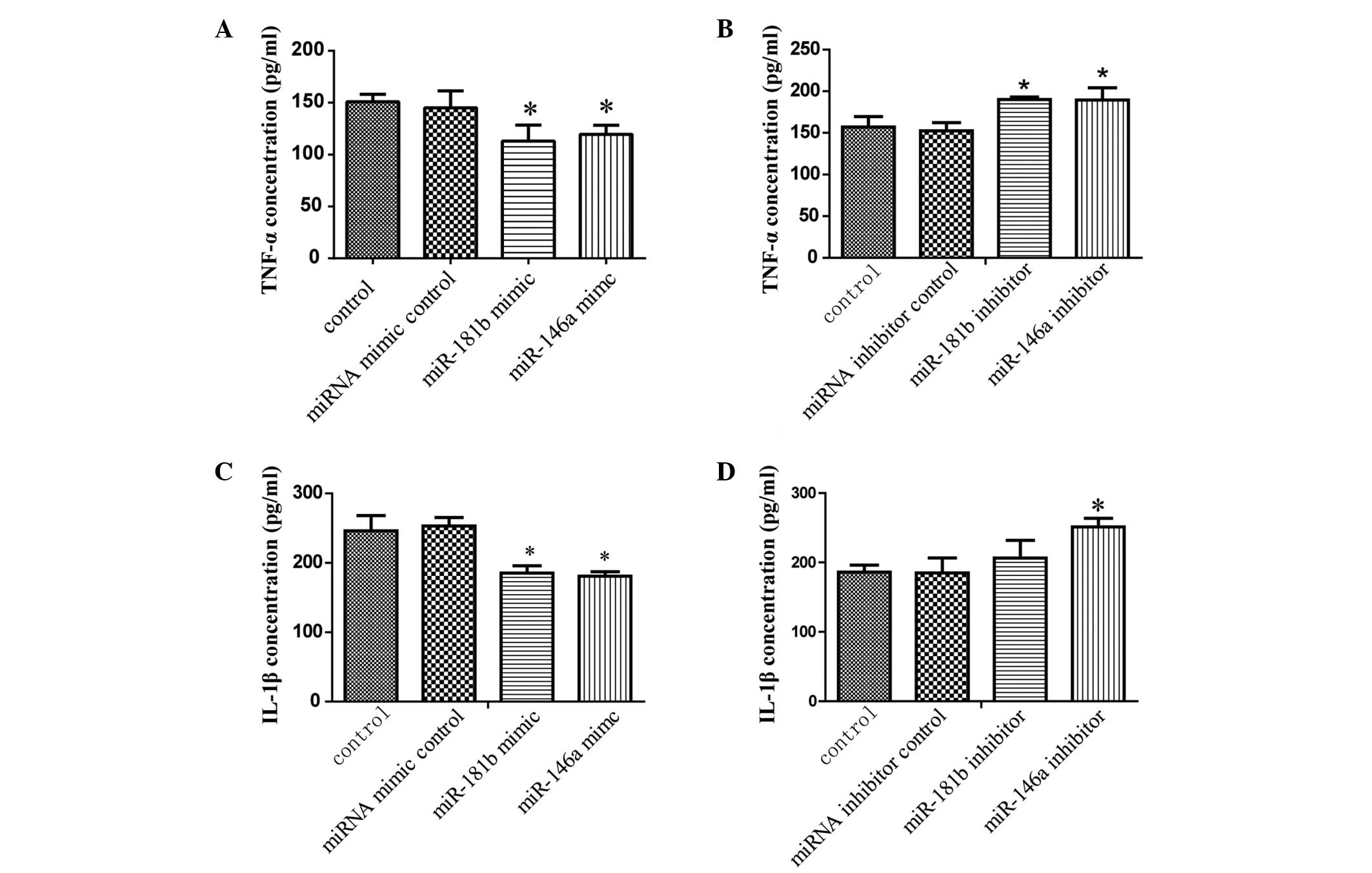
Expression levels of TNF-α and IL-1β,
detected using ELISA, are significantly altered by miRNA
transfection
When the NR8383 cells were transfected with the
miR-181b mimic, the expression of TNF-α was significantly decreased
in the culture supernatants, compared with the miR-181b mimic
control group (P<0.05; Fig.
4A). Simultaneously, miR-181b inhibitors increased the protein
levels of TNF-α significantly (P<0.05), while the miR-146a mimic
and inhibitors had no effect on the protein expression of TNF-α
(Fig. 4A and B). As shown in
Fig. 4C, when the NR8383 cells
were transfected with miR-146a or miR-181b mimics for 24 h,
followed by SiO2 treatment for 12 h, the expression
levels of IL-1β were significantly decreased, compared with the
mimic control group (P<0.05). The expression of IL-1β in
response to SiO2 treatment, following transfection with
miR-146a or miR-181b inhibitors, was significantly increased,
compared with the inhibitor control group (P<0.05; Fig. 4D).
Discussion
It has been increasingly accepted that aberrant
expression levels of certain miRNAs are significantly involved in
the underlying pathophysiology of lung diseases (15,17,18).
However, investigations focusing on the association between
alterations in miRNA expression and the formation of silicosis are
limited. In order to identify miRNAs potentially involved in
SiO2-induced lung fibrosis, our previous study
characterized the miRNA profile of SiO2-treated lungs
using an miRCURY™ LNA array (version 16.0; Exiqon A/S, Vedbaek,
Denmark) (21). In the present
study, RT-qPCR revealed that the expression of miR-146a was
significantly increased, while the expression of miR-181b was
significantly reduced in fibrotic lungs. In order to elucidate the
underlying mechanisms, the NR8383 cells were transfected with
miRNA-146a and miR-181b mimics or inhibitors, and the cells and
culture supernatants were collected following SiO2
treatment for 12 h. The results demonstrated that the mRNA and
protein levels of TNF-α were significantly reduced by miR-181b
mimic, and the opposite effect was observed following knockdown of
miR-181b. Simultaneously, the expression mRNA and protein
expression levels of IL-1β were significantly decreased by the
miR-146a mimic and increased by the miR-146a inhibitor. Taken
together, the results of the present study provided support for the
roles of miR-146a and miR-181b in the pathogenesis of silicosis,
and suggested that they may be candidate therapeutic targets in
this disease.
Alterations in the expression of miR-146a are
important events in the pathogenesis of several human diseases,
including autoimmune disorders, viral infections, cancer, muscle
disorders and myelodysplastic syndrome (23-28).
In the present study, transfection of NR8383 cells with the
miR-146a mimic resulted in reduced IL-1β release, wheraes
transfection with the miR-146a inhibitor increased IL-1β release in
response to SiO2 stimulation. This suggested that
miR-146a may directly or indirectly suppress the expression of
IL-1β. Liu et al (29)
reported that miRNA-146a was upregulated in human bronchial
epithelial cells in response to stimulation by transforming growth
factor β1 and cytomix, a mixture of IL-1β, interferon γ and TNF-α.
Zhong et al (30) provided
evidence that miR-146a exerts negative feedback in neutrophil
elastase-stimulated MUC5AC production from human bronchial
epithelial cells. Sato et al (31) demonstrated that reduced levels of
miR-146a increase the release of the inflammatory mediator
prostaglandin E2, indicating a crucial role for this miRNA in the
abnormal inflammatory response in chronic obstructive pulmonary
disease. A number of previous reports have suggested that miR-146a
transcription is regulated by nuclear factor κB (30,32,33).
The results of the present study suggested that miR-146a may act
through post-translational inhibition of target genes and regulate
innate responses of macrophages to SiO2 stimulation.
miR-181b belongs to the miR-181 family and has an
important regulatory role in cell cycle and differentiation
(34-36). miR-181b is also involved in
different cancer cell lines (37,38).
In the present study, the mRNA and protein levels of TNF-α were
significantly reduced by the mimic, and the opposite effect was
observed following knockdown of miR-181b with the inhibitor. This
indicated that the miR-181b mimic enhanced the phagocytic ability
of the NR8383 cells towards SiO2. Despite the prominent
biological importance of miR-181b, its response to SiO2
stimulation remains to be fully elucidated. The effects of miR-181b
in NR8383 cells predominantly depend on the target genes and their
co-regulatory functions. For example, miR-181b has been reported to
sensitize pancreatic ductal adenocarcinoma, gastric and lung cancer
cells to chemotherapy by targeting B cell lymphoma-2 (39–41).
Yang et al (20)
demonstrated that miR-181b is markedly downregulated in clinical
non-small cell lung cancer tissues, compared with non-tumorous lung
tissues. The precise molecular mechanisms for the altered
expression of miR-181b in silicosis, and how this affects TNF-α
require investigation.
The present study investigated the roles of miR-146a
and miR-181b in regulating TNF-α and IL-1β secretion in
SiO2-induced NR8383 rat macrophages. miR-146a and
miR-181b were characterized in SiO2-treated lungs, which
indicated the importance of miR-146a and miR-181b in silicosis, and
assisted in elucidating the mechanisms underlying the development
and progression of silicosis. A limitation of the present study was
that the investigation was in vitro, thus it remains to be
elucidated whether manipulation of the expression of miR-146a and
miR-181b in vivo has a significant effect on the development
of pulmonary fibrosis. Therefore, examination of the effect of
miR-146a and miR-181b in vivo is a priority for future
investigations. In addition, the basis for cell-type-specific
expression of individual miR-146 and miR-181 family members and, in
certain cases, their ability to target specific genes, requires
further investigation. The present study, to the best of our
knowledge, is the first to indicate that alterations in the
expression of miR-146a and miR-181b may be correlated with
silicosis, suggesting that miR-146a and miR-181b may be involved in
silicosis and act as potential therapeutic targets for silicosis.
Further investigation of the molecular mechanisms by which miR-146a
and miR-181b contribute to the initiation and progression of
silicosis are required.
In conclusion, the present study investigated the
association between alterations in the expression of miR-146a and
miR-181b, and silicosis. The results provide support for the role
for miR-146a and miR-181b in the pathogenesis of silicosis, and
suggested the possibility for their use as therapeutic targets in
this disease. The specific mechanism underlying this interaction
requires further investigation.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81102107 and
81202181).
References
|
1
|
Leung CC, Yu IT and Chen W: Silicosis.
Lancet. 379:2008–2018. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Greenberg MI, Waksman J and Curtis J:
Silicosis: A review. Dis Mon. 53:394–416. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mossman BT and Churg A: Mechanisms in the
pathogenesis of asbestosis and silicosis. Am J Respir Crit Care
Med. 157:1666–1680. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Castranova V and Vallyathan V: Silicosis
and coal workers' pneumoconiosis. Environ Health Perspect.
108:675–684. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Thomas CR and Kelley TR: A brief review of
silicosis in the United States. Environ Health Insights.
4:212010.PubMed/NCBI
|
|
6
|
Cohen RA, Patel A and Green FH: Lung
disease caused by exposure to coal mine and silica dust. Semin
Respir Crit Care Med. 29:651–661. 2008. View Article : Google Scholar
|
|
7
|
Sirajuddin A and Kanne JP: Occupational
lung disease. J Thorac Imaging. 24:310–320. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rimal B, Greenberg AK and Rom WN: Basic
pathogenetic mechanisms in silicosis: Current understanding. Curr
Opin Pulm Med. 11:169–173. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Davis GS: Pathogenesis of silicosis:
Current concepts and hypotheses. Lung. 164:139–154. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yucesoy B, Vallyathan V, Landsittel DP,
Simeonova P and Luster MI: Cytokine polymorphisms in silicosis and
other pneumoconioses. Mol Cell Biochem. 234:219–224. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Vanhée D, Gosset P, Boitelle A, Wallaert B
and Tonnel A: Cytokines and cytokine network in silicosis and coal
workers' pneumoconiosis. Eur Respir J. 8:834–842. 1995.PubMed/NCBI
|
|
12
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Van Rooij E and Olson EN: MicroRNAs:
Powerful new regulators of heart disease and provocative
therapeutic targets. J Clin Invest. 117:2369–2376. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Calin GA and Croce CM: MicroRNA-cancer
connection: The beginning of a new tale. Cancer Res. 66:7390–7394.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cushing L, Kuang PP, Qian J, Shao F, Wu J,
Little F, Thannickal VJ, Cardoso WV and Lü J: miR-29 is a major
regulator of genes associated with pulmonary fibrosis. Am J Respir
Cell Mol Biol. 45:287–294. 2011. View Article : Google Scholar :
|
|
16
|
Guo L, Ji X, Yang S, Hou Z, Luo C, Fan J,
Ni C and Chen F: Genome-wide analysis of aberrantly expressed
circulating miRNAs in patients with coal workers' pneumoconiosis.
Mol Biol Rep. 40:3739–3747. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tomankova T, Petrek M and Kriegova E:
Involvement of microRNAs in physiological and pathological
processes in the lung. Respir Res. 11:1592010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Oglesby IK, McElvaney NG and Greene CM:
MicroRNAs in inflammatory lung disease-master regulators or target
practice? Respir Res. 11:1482010. View Article : Google Scholar
|
|
19
|
Chen G, Umelo IA, Lv S, Teugels E, Fostier
K, Kronenberger P, Dewaele A, Sadones J, Geers C and De Grève J:
miR-146a inhibits cell growth, cell migration and induces apoptosis
in non-small cell lung cancer cells. PloS one. 8:e603172013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang J, Liu H, Wang H and Sun Y:
Down-regulation of microRNA-181b is a potential prognostic marker
of non-small cell lung cancer. Pathol Res Pract. 209:490–494. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sato T, Liu X, Nelson A, Nakanishi M,
Kanaji N, Wang X, Kim M, Li Y, Sun J, Michalski J, et al: Reduced
miR-146a increases prostaglandin E2 in chronic
obstructive pulmonary disease fibroblasts. Am J Respir Crit Care
Med. 182:1020–1029. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Faxuan W, Qin Z, Dinglun Z, Tao Z, Xiaohui
R, Liqiang Z and Yajia L: Altered microRNAs expression profiling in
experimental silicosis rats. J Toxicol Sci. 37:1207–1215. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Urbich C, Kuehbacher A and Dimmeler S:
Role of microRNAs in vascular diseases, inflammation and
angiogenesis. Cardiovasc Res. 79:581–588. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nakasa T, Miyaki S, Okubo A, Hashimoto M,
Nishida K, Ochi M and Asahara H: Expression of microRNA-146 in
rheumatoid arthritis synovial tissue. Arthritis Rheum.
58:1284–1292. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li L, Chen XP and Li YJ: MicroRNA-146a and
Human Disease. Scand J Immunol. 71:227–231. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cameron JE, Yin Q, Fewell C, Lacey M,
McBride J, Wang X, Lin Z, Schaefer BC and Flemington EK:
Epstein-Barr virus latent membrane protein 1 induces cellular
MicroRNA miR-146a, a modulator of lymphocyte signaling pathways. J
Virol. 82:1946–1958. 2008. View Article : Google Scholar :
|
|
27
|
Stanczyk J, Pedrioli DM, Brentano F,
Sanchez-Pernaute O, Kolling C, Gay RE, Detmar M, Gay S and Kyburz
D: Altered expression of MicroRNA in synovial fibroblasts and
synovial tissue in rheumatoid arthritis. Arthritis Rheum.
58:1001–1009. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Perry MM, Williams AE, Tsitsiou E,
Larner-Svensson HM and Lindsay MA: Divergent intracellular pathways
regulate interleukin-1beta-induced miR-146a and miR-146b expression
and chemokine release in human alveolar epithelial cells. FEBS
Lett. 583:3349–3355. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu X, Nelson A, Wang X, Kanaji N, Kim M,
Sato T, Nakanishi M, Li Y, Sun J, Michalski J, et al: MicroRNA-146a
modulates human bronchial epithelial cell survival in response to
the cytokine-induced apoptosis. Biochem Biophys Res Commun.
380:177–182. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhong T, Perelman JM, Kolosov VP and Zhou
XD: MiR-146a negatively regulates neutrophil elastase-induced
MUC5AC secretion from 16HBE human bronchial epithelial cells. Mol
Cell Biochem. 358:249–255. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sato T, Liu X, Nelson A, Nakanishi M,
Kanaji N, Wang X, Kim M, Li Y, Sun J, Michalski J, et al: Reduced
miR-146a increases prostaglandin E2 in chronic
obstructive pulmonary disease fibroblasts. Am J Respir Crit Care
Med. 182:1020–1029. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ma X, Becker Buscaglia LE, Barker JR and
Li Y: MicroRNAs in NF-kapppaB signaling. J Mol Cell Biol.
3:159–166. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Taganov KD, Boldin MP, Chang KJ and
Baltimore D: NF-kappaB-dependent induction of microRNA miR-146, an
inhibitor targeted to signaling proteins of innate immune
responses. Proc Natl Acad Sci USA. 103:12481–12486. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Carroll AP, Tran N, Tooney PA and Cairns
MJ: Alternative mRNA fates identified in microRNA-associated
transcriptome analysis. BMC Genomics. 13:5612012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Visone R, Veronese A, Rassenti LZ, Balatti
V, Pearl DK, Acunzo M, Volinia S, Taccioli C, Kipps TJ and Croce
CM: miR-181b is a biomarker of disease progression in chronic
lymphocytic leukemia. Blood. 118:3072–3079. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sun X, Icli B, Wara AK, Belkin N, He S,
Kobzik L, Hunninghake GM, Vera MP; MICU Registry; Blackwell TS, et
al: MicroRNA-181b regulates NF-κB-mediated vascular inflammation. J
Clin Invest. 122:1973–1990. 2012.PubMed/NCBI
|
|
37
|
Visone R, Veronese A, Balatti V and Croce
CM: MiR-181b: New perspective to evaluate disease progression in
chronic lymphocytic leukemia. Oncotarget. 3:195–202. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen L, Yang Q, Kong WQ, Liu T, Liu M, Li
X and Tang H: MicroRNA-181b targets cAMP responsive element binding
protein 1 in gastric adenocarcinomas. IUBMB Life. 64:628–635. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cai B, An Y, Lv N, Chen J, Tu M, Sun J, Wu
P, Wei J, Jiang K and Miao Y: miRNA-181b increases the sensitivity
of pancreatic ductal adenocarcinoma cells to gemcitabine in vitro
and in nude mice by targeting BCL-2. Oncol Rep. 29:1769–1776.
2013.PubMed/NCBI
|
|
40
|
Zhu DX, Zhu W, Fang C, Fan L, Zou ZJ, Wang
YH, Liu P, Hong M, Miao KR, Liu P, et al: miR-181a/b significantly
enhances drug sensitivity in chronic lymphocytic leukemia cells via
targeting multiple anti-apoptosis genes. Carcinogenesis.
33:1294–1301. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhu W, Shan X, Wang T, Shu Y and Liu P:
miR-181b modulates multidrug resistance by targeting BCL2 in human
cancer cell lines. Int J Cancer. 127:2520–2529. 2010. View Article : Google Scholar : PubMed/NCBI
|