Introduction
Osteosarcoma is a rare type of sarcoma, however, it
is the most common histological form of bone cancer, which is
associated with a poor prognosis due to early pulmonary metastasis
(1,2). Although osteosarcoma occurs in
patients of all ages, osteosarcoma is more common in children and
comprises 2.4% of all malignancies in pediatric patients worldwide
(3). The cause of osteosarcoma is
unclear and risk factors, such as irradiation and genetic
influences contribute to the development of osteosarcoma (4).
Adriamycin (ADM) belongs to a group of
chemotherapeutic agents, termed anthracycline antibiotics that are
able to slow or stop the growth of cancer cells (5). It has been clinically applied to
treat patients exhibiting a variety of types of cancer, including
osteosarcoma. Similar to other chemotherapeutic agents, although it
has a killing effect on cancer cells, ADM eventually becomes
ineffective due to the cancer cells developing ADM resistance
(6). In addition, the toxic
side-effects reduce the success of the treatment in patients.
Therefore, a strategy is necessary to sensitize ADM in order to
decrease the dose required for administration in clinical
practice.
Phenethyl isothiocyanate (PEITC) is a compound of
naturally occurring isothiocyanates found in a variety of
cruciferous vegetables and is produced by enzymatic conversion of
glucosinolate. PEITC has been evaluated as a potential antitumor
agent in various types of cancer. For example, PEITC regulates
epigenetic process and inhibits histone deacetylases in prostate
cancer, leukemia and myeloma cells (7–9).
Increasing numbers of studies have demonstrated that PEITC inhibits
cell proliferation and induces apoptosis in various types of tumor
cell (10–13). Recently, PEITC has been shown to
sensitize tumor cells to chemotherapeutic agents and
synergistically enhance chemotherapeutic agent-induced apoptosis of
cancer cells (14–18). However, it remains unknown as to
whether PEITC influences ADM-induced apoptosis in osteosarcoma.
In the present study, U2-OS cells were used to
determine the synergistic effects of PEITC and ADM treatment on
osteosarcoma cells. The half maximal inhibitory concentration
(IC50) values of PEITC and ADM, either alone or in
combination, against U2-OS cells were measured. In addition, the
effects of PEITC and ADM treatment, either alone or in combination,
on proliferation and apoptosis of U2-OS cells were assessed.
Finally, the potential signaling pathways involved in PEITC and
ADM-induced apoptosis of U2-OS cells were investigated.
Materials and methods
Cells and reagents
U2-OS cells were purchased from the Cell Bank of
Chinese Academy of Sciences (Shanghai, China). An MTT cell counting
kit, Annexin V-fluorescein isothiocyanate (FITC) and propidium
iodide (PI) were purchased from Vazyme Biotech (Nanjing, China).
2-Phenylethyl Isothiocyanate was purchased from TSI Instrument
(Beijing) Co., Ltd. (Shanghai, China). ADM was purchased from
Sigma-Aldrich (St. Louis, MO, USA). 4′,6-diamidino-2-phenylindole
dihydrochloride (DAPI) and a caspase-3 activity assay kit were
purchased from Beyotime Institute of Biotechnology (Haimen, China).
A bicinchoninic acid assay (BCA) protein assay kit was purchased
from Geneseed Biotech Co., Ltd. (Guanzhou, China). Rabbit
anti-caspase-3 (1:1,000; cat. no. BS1518), rabbit anti-Fas (1:500;
cat. no. BS1745), and rabbit anti-FasL (1:500; cat. no. BS1122;
Bioworld Technology, Inc.) were purchased from Bioworld Technology
Inc. (St. Louis Park, MN, USA). Mouse anti-β-actin antibodies
(1:10,000; cat. no. DKM9001) were purchased from Sungene Biotech
Co., Ltd. (Tianjin, China). Horseradish peroxidase-conjugated goat
anti-rabbit (cat. no. LK2001) and anti-mouse (cat. no. KM9001) IgG
(H+L)-HRP secondary antibodies (1:10,000) were purchased from
Sungene Biotech Co., Ltd.
MTT assay
The proliferation of U2-OS cells was measured using
an MTT cell counting kit according to the manufacturer's
instruction. U2-OS cells (100 μl of the suspension) were
harvested at the log phase and seeded into a 96-well plate at
6×103 cells/well and cultured overnight at 37°C in a 5%
CO2 atmosphere and saturated humidity. Then, 100
μl PEITC and 100 μl ADM alone or in combination (100
μl total volume; 1:1) were added to the wells. Triplicate
wells were set for each treatment. After 24 h, 20 μl MTT
solution (5 mg/ml 0.5% MTT) was added into each well and incubated
at 37°C for 4 h. Following washing with phosphate-buffered saline
(PBS), dimethyl sulfoxide and glycine buffering solution (Takara
Bio, Inc., Otsu, Japan) were added to stop the reaction. The plates
were maintained at room temperature in the dark for 2 h and the
optical density (OD) value at 490 nm was measured using a
microtiter plate reader (CytoFluor® Series 4000
Fluorescence Multi-Well Plate Reader; Applied Biosystems Life
Technologies, Foster City, CA, USA).
Flow cytometry
U2-OS cells were digested with 0.05% trypsin/0.5 mM
EDTA (pH 8.0) for 3 min, harvested and washed with PBS. Cells were
resuspended in 95 μl Annexin V staining buffer [10 μM
HEPES (pH 7.4), 140 mM NaCl, 2.5 mM CaCl2] (GE
Healthcare Life Sciences, Piscataway, NJ, USA) and incubated with 5
μl Annexin V-FITC for 15 min at 4°C. Following washing, the
cells were resuspended in Annexin V staining buffer and
10-μg/ml PI (10 μl) was added to each sample. Flow
cytometry was immediately performed to measure U2-OS cell apoptosis
using a FACSCalibur (BD Biosciences, Franklin Lakes, NJ, USA).
Terminal deoxynucleotidyl transferase
dUTP nick end labeling (TUNEL) assay
A TUNEL assay was performed with a
Fluorescein-FragEL™ DNA Fragmentation Detection kit (EMD Millipore,
Gibbstown, NJ, USA) according to the manufacturer's instructions.
Briefly, U2-OS cells were grown on glass slides and then fixed with
4% formaldehyde at 4°C for 25 min. This was followed by incubation
with 0.2% Triton X-100 (Life Technologies, Grand Island, NY, USA)
at room temperature for 5 min. The slides were immersed in 1xTdT
Equilibration buffer for 30 min, and incubated with Fluorescein-TdT
labeling mixture and TdT enzyme (Sigma-Aldrich) for 1 h at 37°C.
Staining was completed by a 1-min incubation with DAPI (Molecular
Probes Life Technologies, Carlsbad, CA, USA) and the coverslips
were mounted on slides. Measurements of TUNEL-positive nuclei were
performed on 10–15 images/slide (captured by an independent
observer blinded to the experiment) using a microscope (BX43;
Olympus Corporation, Tokyo, Japan) connected to a digital camera,
which was attached to an image processor (Quantity One 4.6.2;
Bio-Rad Laboratories, Inc., Hercules, CA, USA). The images were
recorded and saved using Adobe Photoshop 6 (Adobe Systems, San
Jose, CA, USA).
Measurement of caspase-3 activity
Caspase-3 activity was assayed using a caspase-3
activity assay kit according to the manufacture's instructions. The
assay is based on spectrophotometric detection of the chromophore
p-nitroaniline (pNA) following cleavage from the labeled substrate,
Ac-DEVD-pNA. The free pNA is quantified using a spectrophotometer
or a microtiter plate reader at 405 nm. Comparison of the
absorbance of pNA from an apoptotic sample with an uninduced
control allows determination of the fold increase in caspase-3
activity. Briefly, U2-OS cells were digested, harvested and lysed
with lysis buffer. Total proteins were measured using the BCA
protein assay kit and the cell lysates were incubated with 0.2 mM
Ac-DEVD-pNA for 60–120 min at 37°C. The OD values at A405 were
measured by spectrophotometry detection (MK3; Tocan Trading Co.,
Ltd., Shanghai, China).
Western blotting
Total proteins from U2-OS cells were extracted using
a lysis buffer containing 60 μg/ml phenyl-methylsulfonyl
fluoride (Tokyo Chemical Industry Co., Ltd., Tokyo, Japan). Protein
concentrations were assessed using the BCA protein assay kit. The
protein samples were denatured at 100°C for 5 min and separated by
10% SDS-PAGE (Life Technologies). After transferring and blocking
with blocking buffer (1X Tris-buffered saline-Tween with 5% w/v
nonfat dry milk) for 1–2 h, the membranes were hybridized with
primary antibodies on a shaker (cat. no. 4637-1CECN; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) overnight at 4°C. After
washing, the membranes were incubated with a secondary antibodies
on a shaker for 1 h at 37°C. Enhanced chemiluminescence regents
were used for detection. The films were developed using autographed
films and scanned with a laser densiDetect (Applied Biosystems Life
Technologies).
Statistical analysis
Statistical analysis was performed using Student's
t-test. P<0.05 was considered to indicate a statistically
significant difference and P-values were two-tailed.
Results
PEITC and ADM treatment inhibits
proliferation of U2-OS cells in a dose-dependent manner
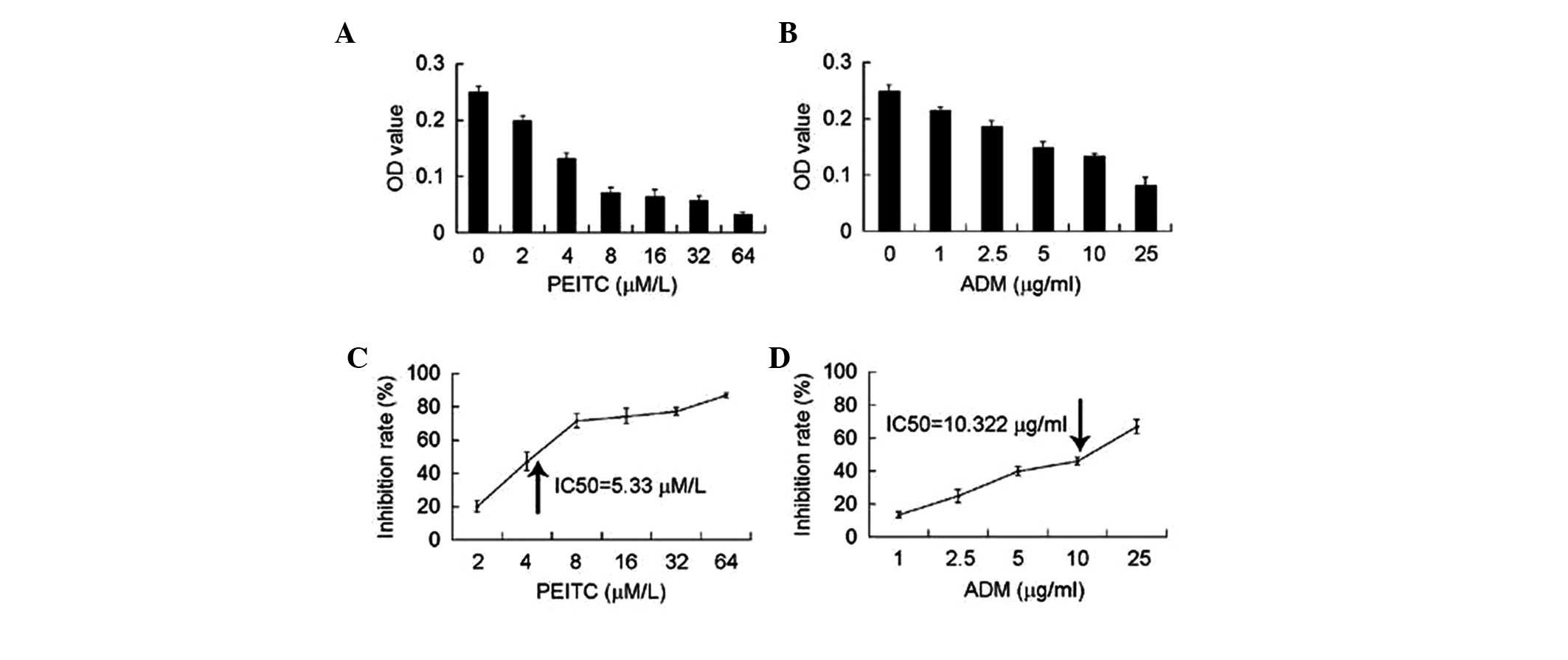
To investigate the synergistic treatment effect of
PEITC and ADM on osteosarcoma cells, the IC50 value of
PEITC or ADM against U2-OS cells was assessed. U2-OS cells were
treated with a series of doses of PEITC or ADM for 24 h and the OD
values were measured using an MTT assay. As shown in Fig. 1A, PEITC and ADM had a
dose-dependent effect on U2-OS cells. The OD values decreased as
the dose of PEITC or ADM increased. When the OD values were
converted into inhibition rates [(OD value - OD baseline)/OD value
× 100], PEITC and ADM were found to dose-dependently inhibit the
proliferation of U2-OS cells. SPSS Statistics 17.0 software (SPSS,
Inc., Chicago, IL, USA) was used to determine that the
concentration of PEITC and ADM, which caused 50% inhibition of
U2-OS cells (the IC50 value) was 5.33 μM and
10.322 μg/ml, respectively.
Following this, whether PEITC and ADM treatment
exerts synergistic, additive or antagonistic effects on U2-OS cells
was determined. The cells were treated with PEITC and ADM alone or
with a combination of the two at varying concentrations for 24 h,
and the OD values were measured by MTT assay. The untreated cells
served as a control when calculating the inhibition rates of PEITC
or ADM. As shown in Table I, while
treatment with each therapeutic agent alone dose-dependently
produced a substantial inhibition rate of the U2-OS cells, a
combination of the two therapeutic agents led to an increased
inhibition rate when compared with either PEITC or ADM treatment
alone. Q-values were calculated using the following formula: Q =
Ea+b / (Ea+Eb - Ea × Eb). Ea and Eb represent the inhibition rate
of ADM and PEITC alone, respectively; Ea+b represents the
inhibition rate of ADM and PEITC in combination. A Q-value >1.15
signifies a synergistic effect; a Q-value >0.85 and <1.15
signifies an additive effect and a Q-value <0.85 signifies an
antagonistic effect. Based on this, the combination of low doses of
PEITC and ADM was observed to produce a synergistic effect, while
the combination of high doses of the two therapeutic agents
resulted in an additive effect (Table
I). No antagonistic effect was observed for PEITC + ADM
treatment.
 | Table IEffects of ADM and PEITC on
osteosarcoma cells. |
Table I
Effects of ADM and PEITC on
osteosarcoma cells.
| Group | Conc.a | Blank | OD1 | OD2 | OD3 | Mean | Adj. OD value | Inhibition rate
(%) | Q-value |
|---|
| Untreated | 0 | 0.097 | 0.376 | 0.399 | 0.407 | 0.394 | 0.297 | – | – |
| ADM | 1 | 0.091 | 0.344 | 0.329 | 0.324 | 0.332 | 0.241 | 18.743 | – |
| PEITC | 2 | 0.119 | 0.346 | 0.356 | 0.327 | 0.343 | 0.224 | 24.579 | – |
| ADM + PEITC | 1 + 2 | 0.121 | 0.255 | 0.281 | 0.254 | 0.263 | 0.142 | 52.076 | 1.345 |
| ADM | 3 | 0.102 | 0.333 | 0.323 | 0.324 | 0.327 | 0.225 | 24.242 | – |
| PEITC | 2.5 | 0.105 | 0.330 | 0.327 | 0.312 | 0.323 | 0.218 | 26.599 | – |
| ADM + PEITC | 3 + 2.5 | 0.118 | 0.204 | 0.245 | 0.214 | 0.221 | 0.103 | 65.320 | 1.471 |
| ADM | 10 | 0.134 | 0.308 | 0.297 | 0.303 | 0.303 | 0.169 | 43.210 | – |
| PEITC | 4 | 0.099 | 0.277 | 0.253 | 0.255 | 0.262 | 0.163 | 45.230 | – |
| ADM + PEITC | 10 + 4 | 0.098 | 0.179 | 0.193 | 0.181 | 0.184 | 0.086 | 70.887 | 1.029 |
Treatment with PEITC and ADM alone and in
combination increases U2-OS cell apoptosis
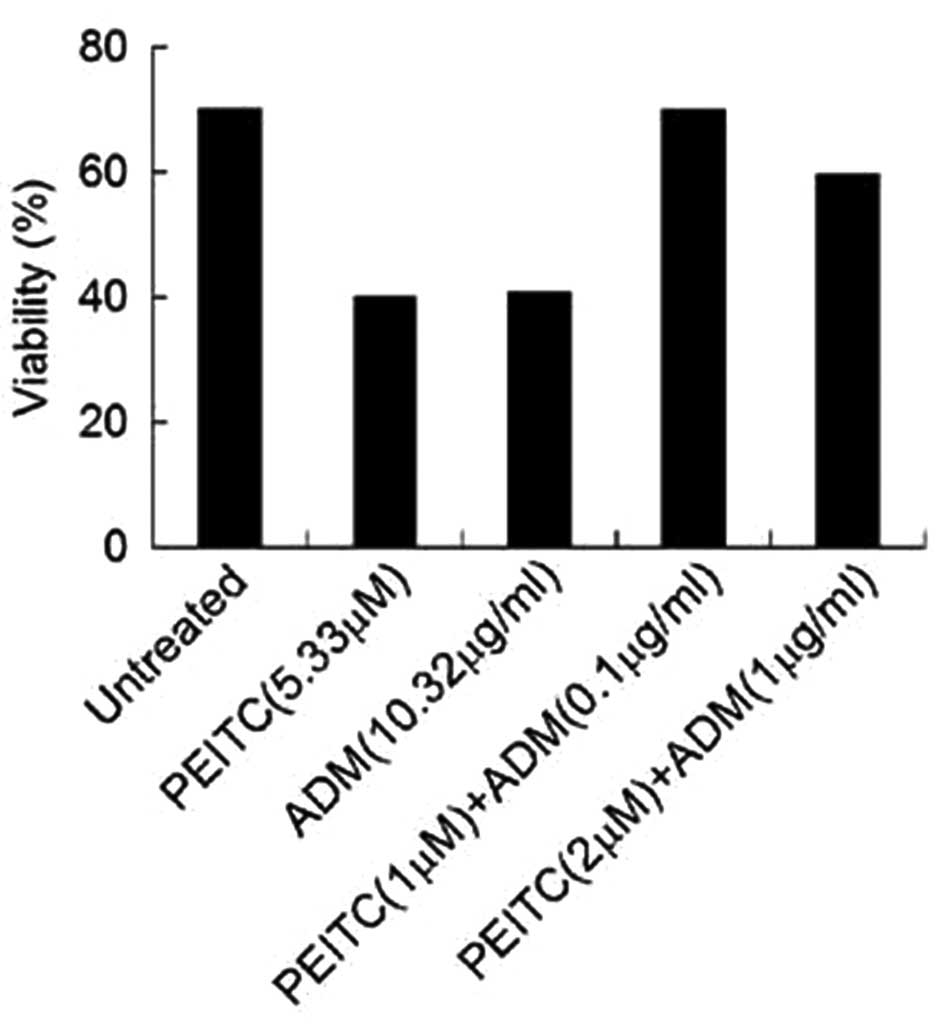
Given the inhibitory effects of PEITC or ADM
observed in the osteosarcoma cells, whether PEITC and ADM alone or
in combination impacted survival of osteosarcoma cells was
subsequently investigated. To do this, U2-OS cells were stained
with Annexin V and PI and the U2-OS cell apoptosis was measured.
The viable cells were defined as Annexin V− and
PI− cells. As shown in Fig.
2, U2-OS cells treated with the IC50 concentration
of PEITC or ADM displayed a decreased number of viable cells
[viability from 70% (observed in the untreated cells) to 40%]. The
findings demonstrate that a combination treatment of PEITC and ADM
dose-dependently marginally decreased the viability of U2-OS
cells.
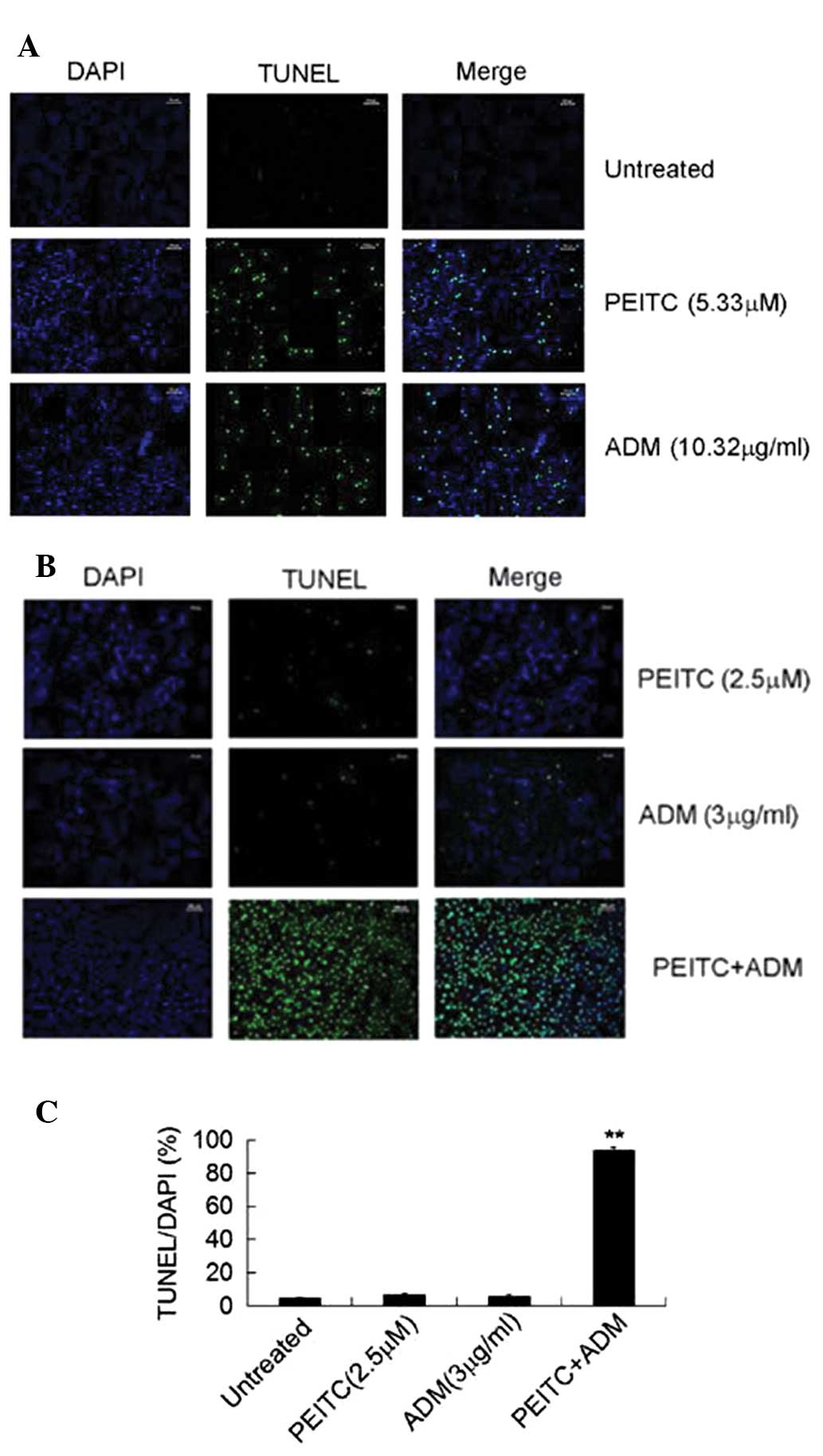
In addition to Annexin V/PI staining, a TUNEL assay
was conducted to determine the apoptotic DNA fragmentation. A
fluorescence-labeled TdT was incubated with fixed and permeabilized
U2-OS cells and the apoptotic cells were visualized using a
fluorescent microscope (cells exhibiting fluorescence were
apoptotic). Analysis of the TUNEL assay of the U2-OS cells treated
with or without the IC50 concentration of ADM or PEITC
was conducted first. As shown in Fig.
3A, while the viability of the untreated U2-OS cells was high,
treatment with 5.33 μM PEITC or 10.322 μg/ml ADM
significantly induced U2-OS cell apoptosis, as indicated by the
increased number of fluorescent cells. Subsequently, the effect of
the combination treatment (ADM + PEITC) at lower doses on TUNEL in
the U2-OS cells was assessed. Treatment with 2.5 μM PEITC or
3 μg/ml ADM marginally increased TUNEL in the U2-OS cells;
however, a combination of the two therapeutic agents markedly
enhanced the number of TUNEL cells (Fig. 3B). The percentage of TUNEL cells
from DAPI-stained cells was then quantified and calculated. The
data from three experiments is summarized in Fig. 3C. As expected, treatment with a
combination of PEITC and ADM significantly increased the percentage
of TUNEL cells when compared with the untreated group or with the
cells treated with PEITC or ADM alone (Fig. 3C).
Treatment with PEITC and ADM alone and in
combination increases caspase-3 activity in U2-OS cells
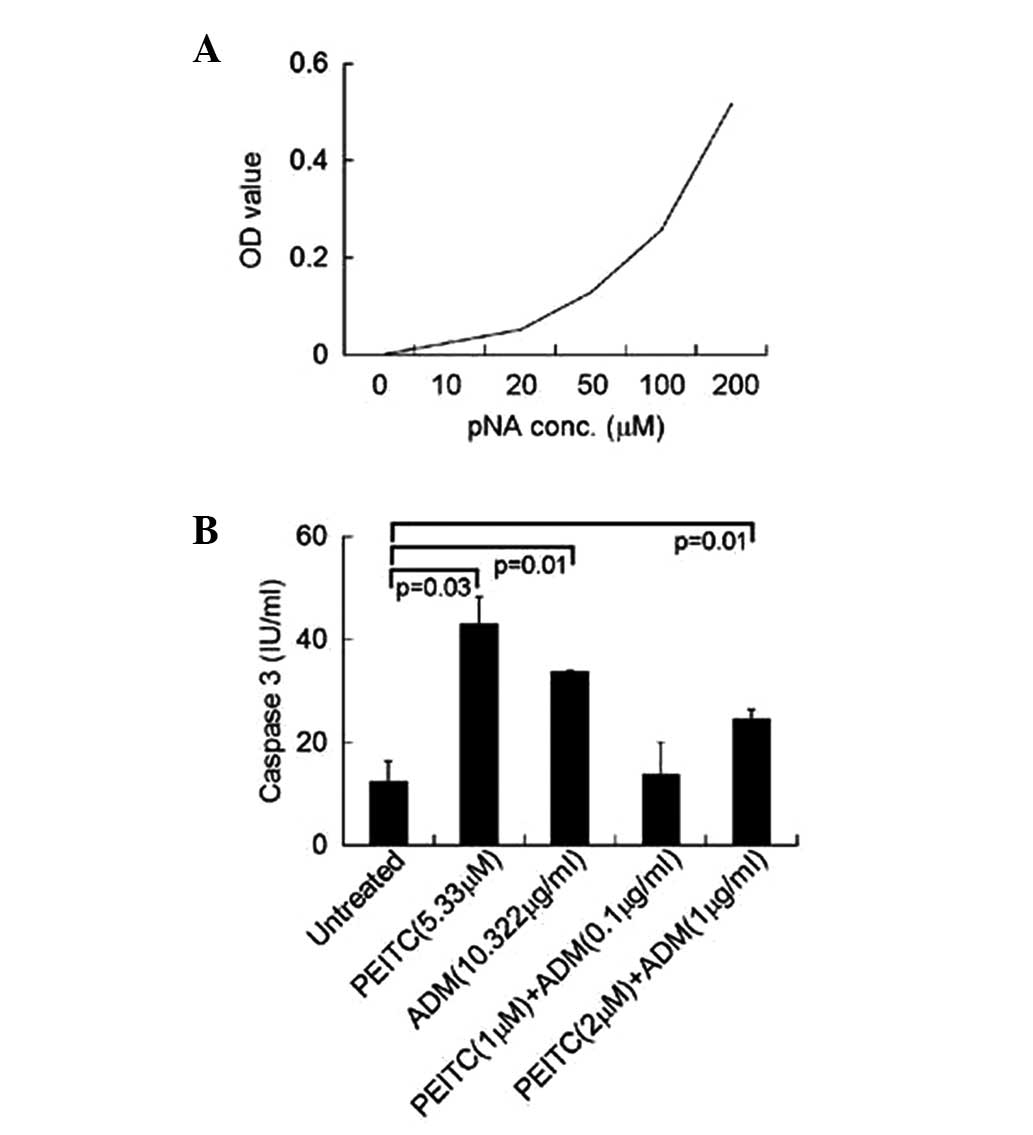
Caspase-3 is key in mediating cell apoptosis and the
increased activity of caspase-3 causes or is the result of
apoptosis. To further determine the apoptotic induction effect of
PEITC and ADM on U2-OS cells, U2-OS cells were treated with PEITC
and ADM to determine whether they activate caspase-3 and increase
its activity. A caspase-3 activity assay kit was used and a linear
standard curve was produced, which presents the increased OD values
against increasing concentrations of pNA (Fig. 4A). Subsequently, caspase-3 activity
of the U2-OS cells treated with the IC50 concentration
of PEITC or ADM was measured (untreated cells served as the
control). As shown in Fig. 4B,
treatment with PEITC and ADM alone at the IC50
concentration significantly increased capspase-3 activity in U2-OS
cells compared with the untreated cells (P<0.05). The effect of
the combination treatment (PEITC + ADM) at decreased doses on
caspase-3 activity in U2-OS cells was assessed. Although a
combination of PEITC (1 μM) and ADM (0.1 μg/ml)
marginally increased caspase-3 activity, 2 μM PEITC combined
with 1 μg/ml ADM significantly enhanced caspase-3 activity
(P<0.05).
Effect of PEITC and ADM treatment on the
apoptotic signaling pathway of U2-OS cells
As PEITC and ADM treatment significantly increased
caspase-3 activity, whether caspase-3 expression was induced in
U2-OS cells treated with PEITC and ADM was evaluated. Western blot
analysis revealed that while treatment with 2.5 μM PEITC or
3 μg/ml ADM alone increased the expression of caspase-3
protein, a combination treatment with the two therapeutic agents
further upregulated caspase-3 expression in the U2-OS cells. In
addition, the expression of Fas and Fas ligand (FasL) was observed
to decrease in U2-OS cells treated with either PEITC and ADM alone
or in combination, indicating that Fas/FasL may not be involved in
PEITC/ADM-induced apoptosis.
Discussion
The significant finding of the present study is that
ADM achieves a comparable effect in osteosarcoma cells at a lower
dose when incubated with PEITC. This finding is particularly
important as drug resistance and side-effects due to the
administration of high doses of ADM lead to failure of this cancer
therapy.
In the present study, PEITC synergistically enhanced
the ADM-mediated inhibition of U2-OS cell proliferation. The
IC50 values of PEITC and ADM against the U2-OS cells
were identified to be 5.33 μM and 10.322 μg/ml,
respectively. However, when incubated together, the IC50
values of PEITC and ADM against U2-OS cells significantly decreased
to a dose <2 μM and 1 μg/ml, respectively (data
not shown). Clinical administration of a reduced dose of ADM, which
achieves a comparable effect to its original dose, is considered to
be beneficial to patient outcome.
As a chemotherapeutic agent, ADM induces apoptosis
in various types of cancer cell. Similar to ADM, PEITC treatment
has been shown to induce apoptosis in tumor cells (19). Supporting these findings, the in
vitro results of the present study demonstrate that cell
viability was low in U2-OS cells that were treated with either ADM
or PEITC. When administered in combination, the rate of apoptosis
in U2-OS cells significantly increased when compared with cells
treated with ADM or PEITC alone. These results indicate that PEITC
sensitizes U2-OS cells to ADM-induced apoptosis, which is
consistent with previous studies (20,21).
Various signaling pathways have been proposed to be
involved in ADM-induced apoptosis of cancer cells. For example, it
has been shown that ADM induces Fas-mediated apoptosis in human
thyroid carcinoma cells (22);
however, a study found that ADM-induced apoptosis was primarily
dependent on tumor necrosis factor-related apoptosis-inducing
ligand (TRAIL)/TRAIL-receptor signaling, although not on FasL,
perforin, NKG2D or DNAX Accessory Molecule-1 (23). No increased expression of Fas/FasL
was noted in U2-OS cells that were treated with PEITC and ADM,
indicating that the Fas/FasL signaling pathway is not involved in
ADM-induced apoptosis in U2-OS cells.
Although Fas/FasL was not found to be involved in
U2-OS cells treated with PEITC and ADM, the activity and expression
of caspase-3 were found to be significantly enhanced in PEITC- and
ADM-treated U2-OS cells. While this result validates the finding
that PEITC and ADM treatment induces apoptosis of U2-OS cells due
to upregulation of caspases, including caspase-3, cause or is the
result of apoptosis (24), it
remains unknown which apoptotic signaling pathway is involved in
PEITC/ADM-induced apoptosis in U2-OS cells. Therefore, further
investigation is required.
In conclusion, PEITC and ADM treatment
synergistically inhibited the proliferation of U2-OS cells. In
addition to proliferation inhibition, PEITC and ADM treatment
synergistically induced the apoptosis of U2-OS cells. This
induction was associated with activity elevation and upregulated
expression of caspase-3. These findings determined the biological
effects of PEITC and ADM treatment on U2-OS cells, as well as
revealing the therapeutic potential of treatment with PEITC in
conjunction with ADM in patients exhibiting osteosarcoma.
References
|
1
|
Dotan A, Dadia S, Bickels J, Nirkin A,
Flusser G, Issakov J, Neumann Y, Cohen I, Ben-Arush M, Kollender Y,
et al: Expandable endoprosthesis for limb-sparing surgery in
children: Long-term results. J Child Orthop. 4:391–400. 2010.
View Article : Google Scholar :
|
|
2
|
Broadhead ML, Clark JC, Myers DE, Dass CR
and Choong PF: The molecular pathogenesis of osteosarcoma: A
review. Sarcoma. 2011:9592482011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ottaviani G and Jaffe N: The epidemiology
of osteosarcoma. Cancer Treat Res. 152:3–13. 2009. View Article : Google Scholar
|
|
4
|
Ottaviani G and Jaffe N: The etiology of
osteosarcoma. Cancer Treat Res. 152:15–32. 2009. View Article : Google Scholar
|
|
5
|
Weiss RB: The anthracylcines: Will we ever
find a better doxorubicin? Semin Oncol. 19:670–686. 1992.PubMed/NCBI
|
|
6
|
Thorna CF, Oshiroa C, Marshe S,
Hernandez-Boussard T, McLeod H, Kleina TE and Altman RB:
Doxorubicin pathways: Pharmacodynamics and adverse effects.
Pharmacogenet Genomics. 21:440–446. 2011. View Article : Google Scholar
|
|
7
|
Beklemisheva AA, Fang Y, Feng J, Ma X, Dai
W and Chiao JW: Epigenetic mechanism of growth inhibition induced
by phenylhexyl isothiocyanate in prostate cancer cells. Anticancer
Res. 26:1225–1230. 2006.PubMed/NCBI
|
|
8
|
Ma X, Fang Y, Beklemisheva A, Dai W, Feng
J, Ahmed T, Liu D and Chiao JW: Phenylhexyl isothiocyanate inhibits
histone deacetylases and remodels chromatins to induce growth
arrest in human leukemia cells. Int J Oncol. 28:1287–1293.
2006.PubMed/NCBI
|
|
9
|
Lu Q, Lin X, Feng J, Zhao X, Gallagher R,
Lee MY, Chiao JW and Liu D: Phenylhexyl isothiocyanate has dual
function as histone deacetylase inhibitor and hypomethylating agent
and can inhibit myeloma cell growth by targeting critical pathways.
J Hematol Oncol. 9:1–6. 2008.
|
|
10
|
Stan SD, Singh SV, Whitcomb DC and Brand
RE: Phenethyl isothiocyanate inhibits proliferation and induces
apoptosis in pancreatic cancer cells in vitro and in a MIAPaca2
xenograft animal model. Nutr Cancer. 66:747–755. 2014.Nov 6 2013
[Epub ahead of print]. View Article : Google Scholar :
|
|
11
|
Huang SH, Hsu MH, Hsu SC, Yang JS, Huang
WW, Huang AC, Hsiao YP, Yu CC and Chung JG: Phenethyl
isothiocyanate triggers apoptosis in human malignant melanoma
A375.S2 cells through reactive oxygen species and the
mitochondria-dependent pathways. Hum Exp Toxicol. 33:270–283. 2014.
View Article : Google Scholar
|
|
12
|
Sarkars R, Mukherjee S and Roy M:
Targeting heat shock proteins by phenethyl isothiocyanate results
in cell-cycle arrest and apoptosis of human breast cancer cells.
Nutr Cancer. 65:480–493. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xiao D and Singh SV: p66Shc is
indispensable for phenethyl isothiocyanate-induced apoptosis in
human prostate cancer cells. Cancer Res. 70:3150–3158. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cang S, Ma Y, Chiao JW and Liu D:
Phenethyl isothiocyanate and paclitaxel synergistically enhanced
apoptosis and alpha-tubulin hyperacetylation in breast cancer
cells. Exp Hematol Oncol. 3:52014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lee DH, Kim DW, Lee HC, Lee JH and Lee TH:
Phenethyl isothiocyanate sensitizes glioma cells to TRAIL-induced
apoptosis. Biochem Biophys Res Commun. 446:815–521. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tang T, Song X, Liu YF and Wang WY: PEITC
reverse multi-drug resistance of human gastric cancer SGC7901/DDP
cell line. Cell Biol Int. 38:502–510. 2014. View Article : Google Scholar
|
|
17
|
Liu K, Cang S, Ma Y and Chiao JW:
Synergistic effect of paclitaxel and epigenetic agent phenethyl
isothiocyanate on growth inhibition, cell cycle arrest and
apoptosis in breast cancer cells. Cancer Cell Int. 13:102013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xiao D and Singh SV: Phenethyl
isothiocyanate sensitizes androgen-independent human prostate
cancer cells to docetaxel-induced apoptosis in vitro and in vivo.
Pharm Res. 27:722–731. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gupta P and Srivastava SK: Antitumor
activity of phenethyl isothiocyanate in HER2-positive breast cancer
models. BMC Med. 10:802012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yuang P, Chen BA, Cheng J, Ma XD, Liu DL
and Lu QY: Effect of phenylhexyl isothiocyanate on adriamycin
resistance of K562/A02 cell line. Zhongguo Shi Yan Xue Ye Xue Za
Zhi. 17:352–357. 2009.In Chinese. PubMed/NCBI
|
|
21
|
Mukherjee S, Bhattacharya RK and Roy M:
Targeting protein kinase C (PKC) and telomerase by phenethyl
isothiocyanate (PEITC) sensitizes PC-3 cells towards
chemotherapeutic drug-induced apoptosis. J Environ Pathol Toxicol
Oncol. 28:269–282. 2009. View Article : Google Scholar
|
|
22
|
Massart C, Barbet R, Genetet N and
Gibassier J: Doxorubicin induces Fas-mediated apoptosis in human
thyroid carcinoma cells. Thyroid. 14:263–270. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wennerberg E, Sarhan D, Carlsten M,
Kaminskyy VO, D'Arcy P, Zhivotovsky B, Childs R and Lundqvist A:
Doxorubicin sensitizes human tumor cells to NK cell- and
T-cell-mediated killing by augmented TRAIL receptor signaling. Int
J Cancer. 133:1643–1652. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Degterev A, Boyce M and Yuan J: A decade
of caspases. Oncogene. 22:8543–8567. 2003. View Article : Google Scholar : PubMed/NCBI
|