Introduction
Intermedin (IMD), also known as adrenomedullin 2
(ADM2), is a newly discovered member of the calcitonin family
peptides, which was independently identified by two groups in 2004
(1,2). IMD is expressed in a variety of organ
systems, including the gastrointestinal tract, lungs,
cardiovascular system, hypothalamus, pituitary gland, adrenal gland
and placenta (2–6). Similar to other CT family members,
including calcitonin gene-related peptide (CGRP) and AMD, IMD
exerts its actions through receptor complexes comprising calcitonin
receptor-like receptor (CRLR) and one of three receptor activity
modifying proteins, (RAMP1, RAMP2 and RAMP3) (1,7), and
stimulate the formation of cyclic AMP.
A series of physiological and pharmacological
functions of IMD have been reported. IMD protects the mammalian
vasculature, myocardium and kidney from acute ischemia-reperfusion
injury, chronic oxidative stress and pressure-loading (5,8). IMD
also inhibits apoptosis, attenuates maladaptive tissue remodelling
and preserves cardiac and renal functions (9,10).
Previously, it was reported that bone appears to be a common target
for all the peptides of the calcitonin family, although the
specific bone effects of the peptides vary (11). The administration of calcitonin
produces rapid lowering of serum calcium levels, predominantly
through the inhibition of bone resorption by osteoclasts (12). In vitro investigations and a
number of animal experimental models have demonstrated that amylin
and CGRP are also effective in inhibiting osteoclast activity and
bone resorption (12). Susanne
et al identified that IMD exhibits significant inhibitory
effects on mouse calvarial bone matrix degradation stimulated by
PTH and osteoclastogenesis (13),
and IMD receptors, RAMPd and CRLP are expressed in osteoblasts
(14). However, whether IMD can
affect osteoblasts and is involved in bone resorption remains to be
elucidated.
In the present study, the effects of IMD on the
proliferation and apoptosis of osteoblasts were investigated.
Subsequently, whether IMD is involved in osteoblastic and
osteoclastic activity was determined by assessing molecules, which
are closely associated with the processes, including receptor
activator of NF-κB ligand (RANKL), osteoprotegerin (OPG) and
macrophage colony-stimulating factor (M-CSF). The MC3T3-E1
osteoblast cell line was used as a cell model in vitro. In
addition, the present study investigated the possible mechanism
underlying the inhibitory effects of IMD on bone resorption.
Materials and methods
Reagents
Intermedin/Adrenomedullin-2 (IMD) was purchased from
Phoenix Biotech Company. MTT was obtained from Sigma-Aldrich (St.
Louis, MO, USA) and Annexin V-Fluorescein isothiocyanate
(FITC)/propidium iodide (PI) kit from BD Biosciences (San Jose, CA,
USA). The caspase-3 activity assay kit was purchased from Promega
Corporation (Madison, WI, USA). Alkaline phosphatase (ALP)
detection buffer was purchase from Sigma-Aldrich and the Micro BCA
protein assay kit was obtained from Thermo Fisher Scientific
(Waltham, MA, USA). All RNA primers and the PrimeScript RT-PCR kit
were obtained from Takara, Bio, Inc. (Otsu, Japan). TRIzol reagent
was obtained from Gibco Life Technologies (Carlsbad, CA, USA).
Anti-OPG, RANKL, M-CSF and caspase-3 antibodies were purchased from
Santa Cruz Biotechnology, Inc. (Dallas, TX, USA) The MC3T3-E1 mouse
osteoblastic cell line was obtained from American Type Culture
Collection (Manassas, VA, USA).
Mouse MC3T3-E1 cell culture
The MC3T3-E1 cells were cultured in minimum
essential medium (MEM) containing 10% fetal bovine serum (FBS;
Gibco Life Technologies), 100 U/ml penicillin, 100 mg/ml
streptomycin (Beyotime Institute of Biotechnology, Haimen, China)
and 50 mg/ml of ascorbic acid (Sigma-Aldrich). The cells were
maintained in a humidified, 95% air, 5% CO2 atmosphere
at 37°C. The medium was replaced every 3 days, and the cells were
subcultured using 0.05% trypsin with 0.01% EDTA (Beyotime Institute
of Biotechnology).
Cell proliferation assay
The MC3T3-E1 cells, at a density of 3×103
cells/well, were seeded into 96-well plates, cultured for 24 h at
37°C and then treated with different concentrations of IMD (1, 10
or 100 nM) for 48 h. Cell viability was detected by measuring the
absorbance of each well at 490 nm using a Bio-Rad 680 microplate
reader (Bio-Rad Laboratories, Inc., Hercules, CA, USA) and the
experiment was performed five times. The proliferation rate was
calculated from the optical density (OD) value and was used to
determine the effect of IMD on MC3T3-E1 proliferation.
Osteoblast differentiation
The MC3T3-E1 cells (3×105) were cultured
in MEM containing 10% FBS, 50 mg/ml ascorbic acid and 10 mM
β-glycerophosphate (Sigma-Aldrich) treated with IMD (1, 10 or 100
nM) at 37°C. At differentiation days 7 and 14, the activity of ALP
was measured. Briefly, the cells were washed twice with PBS and
then lysed. The lysates were incubated in ALP detection buffer for
30 min at 37°C and were monitored at 405 nm. Total protein was
detected using a Micro BCA protein assay kit and read at 562 nm.
The relative activity of ALP was normalized to the total protein
content of the sample (405/562 nm). In addition, the mRNA levels of
osteoblast differentiation markers, OCN and BSP were measured, and
Alizarin red staining (Sigma-Aldrich) was performed on
differentiation days 14 and 21. For Alizarin red staining, the
medium was removed, and the cell layer was rinsed with PBS and
fixed in 70% ethanol for 1 h at −20°C. The cell layer was washed
with deionized water and then the fixed cells were stained with 40
mM Alizarin red S (pH 4.2) for 10 min at room temperature.
Cell apoptosis analysis
The apoptosis of the MCT3-E1 cells was quantified
using flow cytometry. The cells (3×105) were cultured at
37°C in serum-free medium or 10−7 M dexamethasone (DEX;
Sigma-Aldrich) for 24 h in the absence or presence of 1–100 nM IMD.
Treatment with 1% FBS was used to determine the basal levels of
apoptosis. Briefly, following treatment with IMD for 48 h at 37°C,
the cells were freshly harvested and washed with PBS. Subsequently
150 µl chilled Annexin V binding buffer was added, followed
by 5 µl Annexin V-FITC and incubation for 15 min. During the
final 5 min, 5 µl PI was added and further incubated for 10
min. Following staining, 350 µl Annexin V binding buffer was
added and the cells were analyzed using a BD FACSCalibur (BD
Biosciences).
Caspase-3 activity was determined by measuring the
cleavage of the Ac-DEVD-pNA chromogenic caspase substrate, which is
a caspase-3 substrate. The OD value at 405 nm was determined to
reflect the quantity of caspase-3. The specific caspase-3 activity,
normalized to that of the total proteins of the cell, was then
expressed as the fold change from the baseline caspase-3 activity
of the control cells.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
The MC3T3-E1 cells were cultured for 48 h in the
presence of 1–100 nM IMD. Following treatment, the mRNA of cells
was extracted using TRizol reagent, and cDNA was synthesized using
the PrimeScript RT-PCR kit. The quantity of samples used in the
RT-qPCR were as follows: SYBR Green Mastermix 10 µl, cDNA 2
µl, forward primer (10 µM) 1 µl, reverse
primer (10 µM) 1 µl and H2O 5 µl.
The sequences of the primers of RANKL, OPG, M-CSF, osteocalcin
(OCN), bone sialoprotein (BSP) and the housekeeping β-actin gene
are presented in Table I. qPCR was
performed on a RotorGene real-time DNA amplification system using
the following cycling protocol: 95°C denaturation for 10 min,
followed by 40 cycles of 95°C (10 sec), 60°C annealing (20 sec) and
72°C extension (30 sec). The 2−ΔΔCT method was used to
determine the gene expression levels (15).
 | Table IPrimer sequences used in reverse
transcription-quantitative polymerase chain reaction analysis in
the present study. |
Table I
Primer sequences used in reverse
transcription-quantitative polymerase chain reaction analysis in
the present study.
| Gene | Primer | Primer sequence
(5′-3′) |
|---|
| OPG | Forward |
TGAGAGAACGAGAAAGACCTGC |
| Reverse |
CGGATTGAACCTGATTCCCTAT |
| RANKL | Forward |
TCCTGAGACTCCATGAAAACGCAG |
| Reverse |
GCCACATCCAACCATGAGCCTTC |
| M-CSF | Forward |
TTTTCCTGGGCATTGTGGTCT |
| Reverse |
AGGAGGTTCAGGGCTTCTTTG |
| OCN | Forward |
GAGGGCAATAAGGTAGTGAA |
| Reverse |
CATAGATGCGTTTGTAGGC |
| BSP | Forward |
CAGGGAGGCAGTGACTCTTC |
| Reverse |
AGTGTGGAAAGTGTGGCGTT |
| β-actin | Forward |
CTGTGCCCATCTACGAGGGCTAT |
| Reverse |
TTTGATGTCACGCACGATTTCC |
Western blot analysis
Following treatment with either serum-free medium or
10−7 M DEX in the presence or absence of 1–100 nM IMD
for 24 h, the MC3T3-E1 cells were lysed in lysis buffer, containing
150 mM TrisHCl (pH 7.4), 150 mM NaCl, 0.1% SDS, 1% Triton X-100, 1%
Nonidet P-40, protease inhibitor, 1 M NaF, 1 M b-glycerophosphate,
0.5 M Na3VO4, 1 M DTT, 1% sodium deoxycholate
and 5 mM EDTA. Following 30 min incubation at 4°C, the lysates were
sonicated and centrifuged at 12,000×g for 10 min at 4°C, and heated
for 5 min at 95°C. The samples were resolved on a SDS-PAGE gel and
transferred onto a nitrocellulose membrane (EMD Millipore,
Billerica, MA, USA). Immunoblotting was then performed using
antibodies against rabbit polyclonal RANKL (cat. no. SC-9073),
rabbit polyclonal OPG (cat. no. SC-11383), rabbit polyclonal M-CSF
(cat. no. SC-13103), rabbit polyclonal caspase-3 (cat. no. SC-7148)
and mouse monoclonal β-actin (cat. no. SC-8342). Following 1 h
incubation at 25°C with 1:2,000 goat anti-rabbit secondary
antibodies, the blots were visualized using enhanced
chemiluminescence (7Sea Biotech, Shanghai, China) and quantified
using a Gel-Pro-Analyzer 4.0 (Media Cybernetics, Inc., Rockville,
MD, USA).
Statistical analysis
Data are presented as the mean ± standard deviation,
and all experiments were performed at least three times with
similar results. Comparisons were made using a one-way analysis of
variance and an unpaired t-test. Statistical analysis was performed
using SPSS version 17.0 (SPSS, Inc., Chicago, IL, USA). P<0.05
was considered to indicate a statistically significant
difference.
Results
Effects of IMD on the proliferation and
differentiation of MC3T3-E1 cells
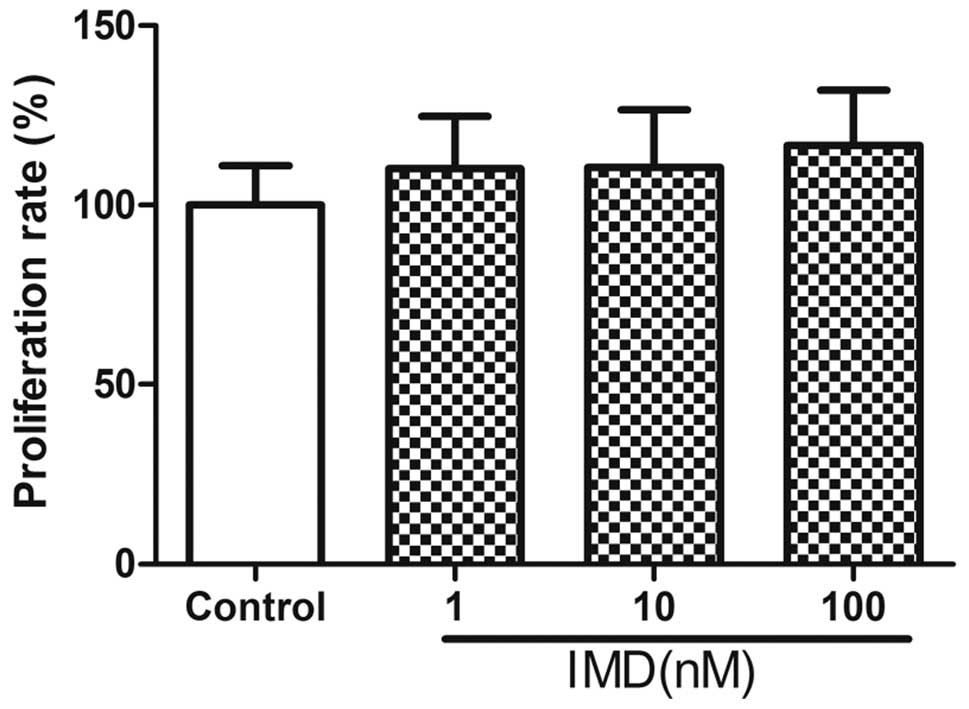
Following treatment with different doses of IMD (0,
10 or 100 nM) for 48 h, the effects of IMD on the growth of
MC3T3-E1 cells was determined using MTT assays (Fig. 1). The data revealed no significant
enhancement in the proliferation of MC3T3-E1 cells exposed to IMD,
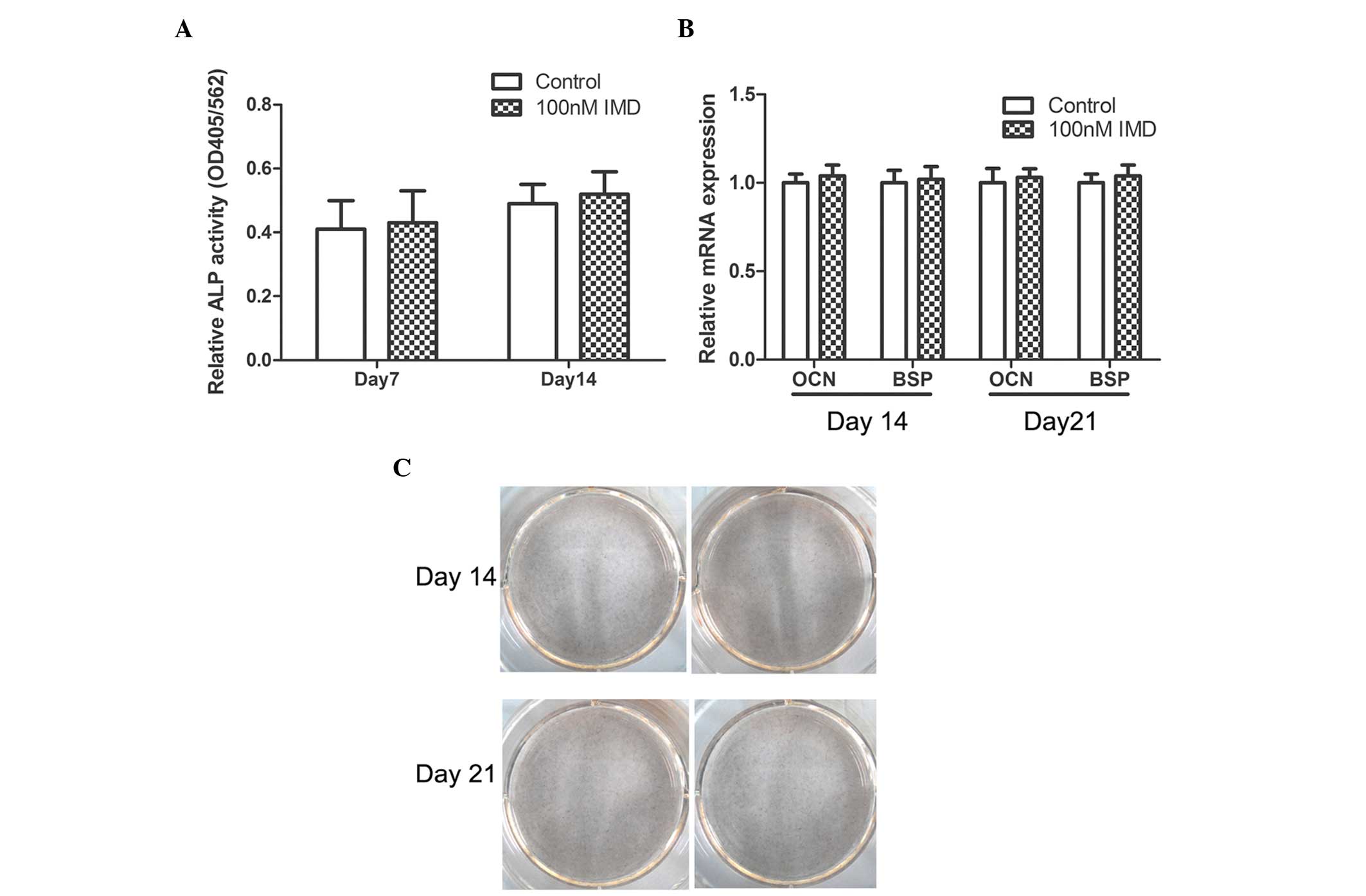
compared with the control group (P>0.05). In addition, no
significant effects on ALP activity, Alizarin red staining or the
mRNA expression levels of OCN or BSP were observed (P>0.05;
Fig. 2). Therefore, IMD had no
effect on the proliferation or differentiation of MC3T3-E1 cells,
and was unable to promote bone formation (data not shown).
Effects of IMD on the apoptosis of
MC3T3-E1 cells
To examine the role of IMD on the apoptosis of
MC3T3-E1 cells, the cells were cultured in serum-free medium or
10−7 M DEX with IMD (1–100 nM) for 24 h, and apoptotic
progression was detected using flow cytometric analysis.
FITC-conjugated annexin V was used to stain the early apoptotic
biomarker, phosphoserine, at the cell surface, whereas PI was used
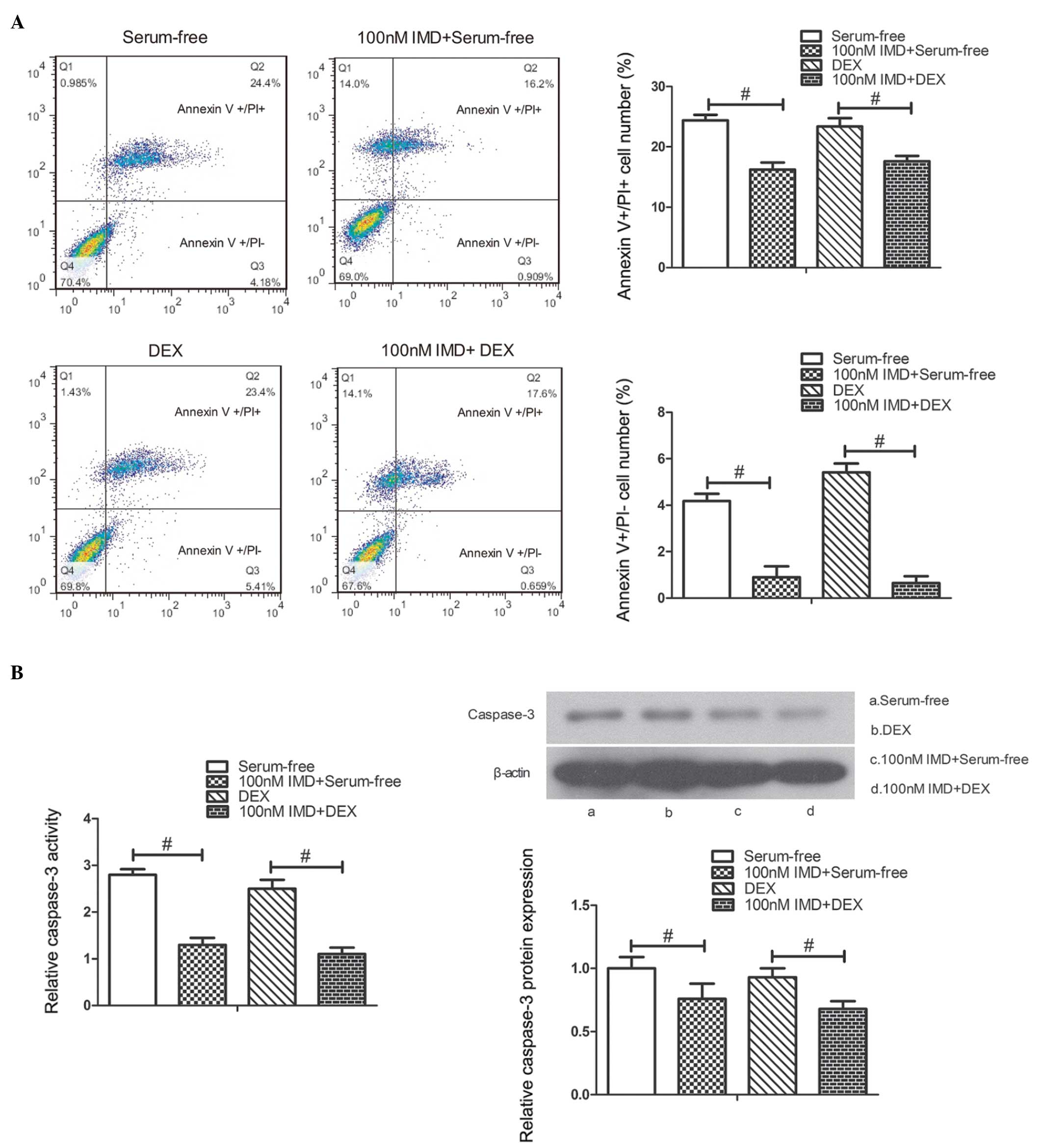
to stain later apoptotic and necrotic cells. As shown in Fig. 3A, the number of PI-positive and
annexin V-positive cells significantly decreased in the cells
treated with 100 nM IMD, compared with those in serum-free medium
or 10−7 M DEX alone (P<0.01).
As caspase-3 is important in the process of
apoptosis, caspase-3 activity and protein expression were measured
to assess apoptosis in the present study. To evaluate the effect of
IMD on caspase-3 activity and protein expression, the MC3T3-E1
cells cultured in serum-free medium or 10−7 M DEX were
treated with different doses of IMD between 1 and 100 nM for 24 h.
The results indicated that IMD significantly decreased caspase-3
activity and protein expression at a concentration of 100 nM,
compared with the untreated cells at 24 h (P<0.01; Fig. 3B). These findings suggested that
IMD inhibited the apoptosis of MC3T3-E1 cells (data not shown).
Effects of IMD on ratio of RANKL/OPG
The expression ratio of RANKL/OPG, which are
synthesized by osteoblasts, represent bone resorption. To verify
the expression of RANKL and OPG in the MC3T3-E1 cells treated with
IMD, the present study performed RT-qPCR and western blot analysis.
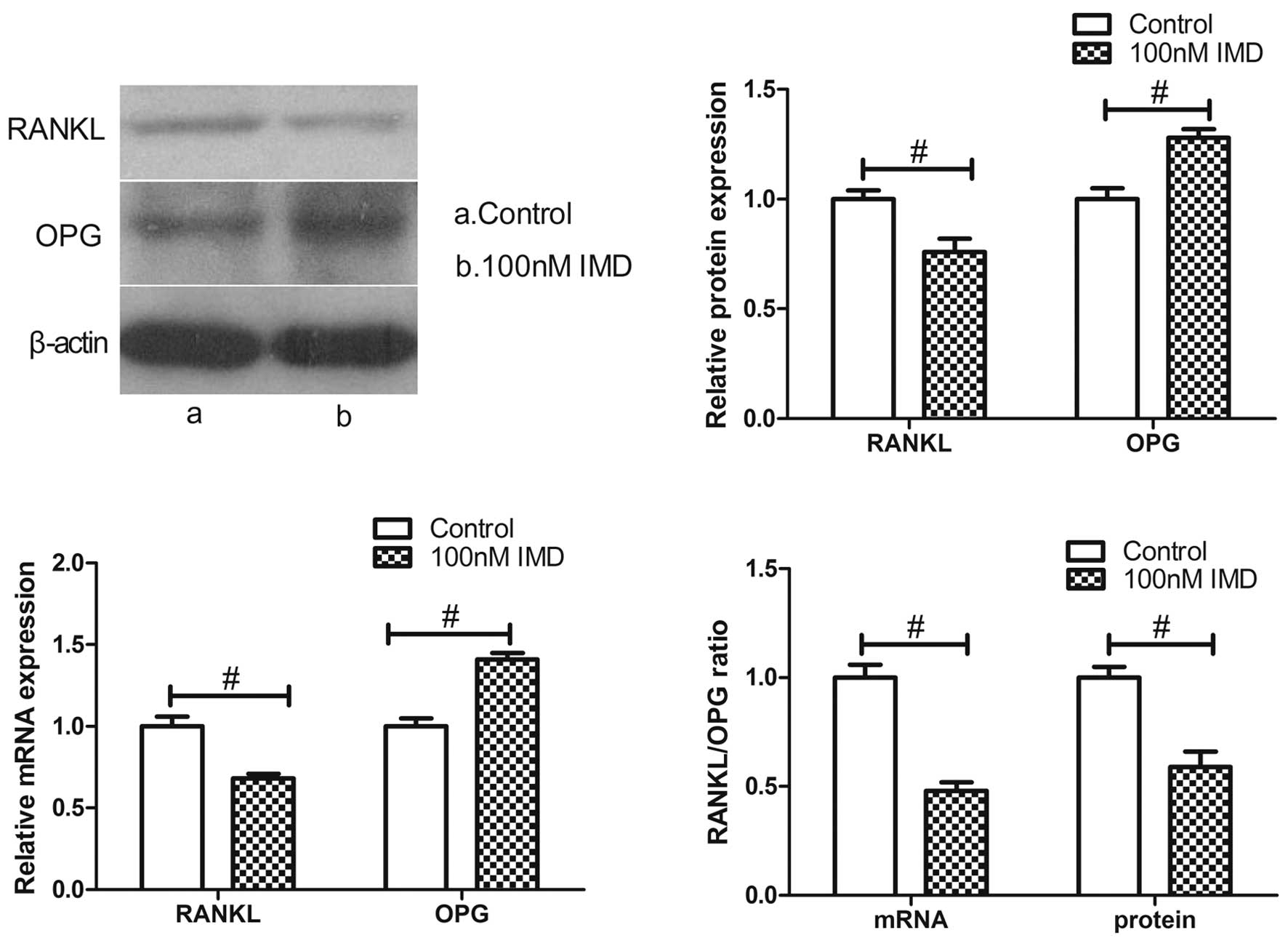
The results revealed that the expression of RANKL at the mRNA and
protein levels were markedly downregulated by various doses of IMD,
whereas the levels of OPG were significantly upregulated (Fig. 4). IMD at 100 nM significantly
decreased the mRNA and protein expression levels of RANKL
(P<0.01), and upregulated the mRNA and protein expression levels
of OPG (P<0.01). Therefore, IMD significantly improved the
imbalance in the RANKL/OPG ratio (P<0.01), which is likely to
induce an inhibitory effect on bone resorption.
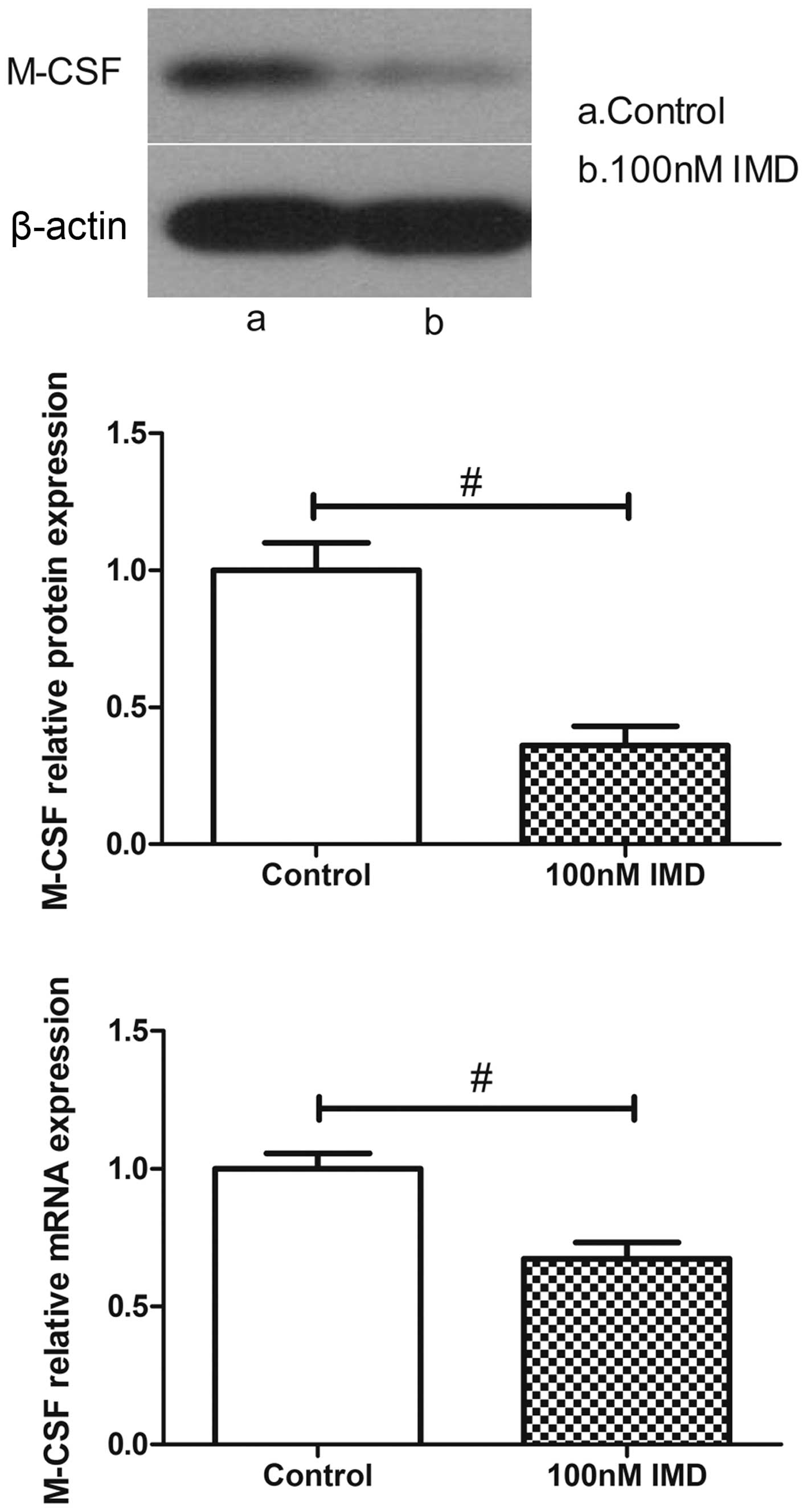
Effects of IMD on the expression of
M-CSF
M-CSF is another osteoblast-mediated activation
marker of bone resorption. As shown in Fig. 5, the expression levels of M-CSF
were measured using RT-qPCR and western blot analysis. In the
untreated group, the MC3T3-E1 cells expressed a basal level of
M-CSF. By contrast, the level of M-CSF in the MC3T3-E1 cells was
significantly altered following IMD treatment. The data
demonstrated that IMD significantly downregulated the mRNA and
protein expression levels of M-CSF (P<0.01), which suggested
suppression of the osteoclast-mediated bone resorption by IMD.
Discussion
Osteoporosis is a common bone disease characterized
by a reduction in bone mineral density and an increase in fracture
risk, which affects quality of life (16). Bone is a dynamic tissue that
constantly undergoes a process of renewal and repair (17) termed 'bone remodeling', in which a
balance of bone formation by osteoblasts and resorption by
osteoclasts continues throughout life (18,19).
This balance involves the coordinated regulation and interaction of
two cell types, osteoclasts and osteoblasts (17,20).
Either a decrease in osteoblastic activity or an increase in
osteoclastic activity in the bone can lead to imbalance by
accelerating bone resorption (21).
Osteoblasts, which are derived from mesenchymal stem
cells (MSCs), are responsible for the production of bone
extracellular matrix and are able to mineralize into bone matrix
(22,23). ALP, OCN and BSP have been reported
to be involved in the molecular events of osteoblast
differentiation (24,25). Using MTT analysis, the present
study demonstrated that IMD had no effect on the proliferation of
the MC3T3-E1 osteoblast lineage. In addition, IMD treatment had no
effect on ALP activity, Alizarin red staining or the mRNA
expression levels of OCN and BSP, suggesting that IMD had no effect
on the differentiation of osteoblasts. Notably, the results of the
Annexin V/PI flow cytometry revealed that IMD significantly
inhibited the apoptosis of the MC3T3-E1 cells. The present study
subsequently measured caspase-3 activity and protein expression
levels and, in accordance with the above findings, IMD markedly
attenuated caspase-3 activity and decreased protein expression
levels. These results indicated that IMD suppressed the apoptosis
induced by serum-free medium or DEX in the MC3T3-E1 cells.
Osteoclasts originate from the monocyte/macrophage
hematopoietic lineages and are responsible for bone resorption
(26,27). Previous studies have confirmed that
osteoblasts can modulate osteoclast function by secreting RANKL,
OPG and M-CSF (28–30). The OPG/RANKL/M-CSF system is
fundamental for regulation between osteoblasts and osteoclasts
(31). The expression of RANKL by
osteoblast cells, is essential for osteoclast differentiation and
maturity via its receptor, RANK, located on the osteoclast membrane
(32) and M-CSF, which binds to
its receptor, c-Fms, and appears to be necessary for osteoclast
development (33,34). OPG, produced by osteoblastic cells,
inhibits osteoclast differentiation through its binding to RANKL
(34). Thus, the RANKL/OPG ratio
and the expression of M-CSF determine osteoclast activity in
addition to bone resorption (35).
The present study is the first, to the best of our knowledge, to
provide insight into the effects of IMD on the imbalance of
RANKL/OPG ratio and the expression of M-CSF. The data obtained in
the present study demonstrated that IMD downregulated the ratio of
RANKL/OPG and the expression of M-CSF at the mRNA and protein
levels, suggesting that IMD is a novel negative regulator of OC
function and resists bone resorption.
In conclusion, the results of the presents study
demonstrated that IMD decreased apoptosis and also decreased the
RANKL/OPG ratio and the expression of M-CSF in MC3T3-E1 cells.
These results provide insight into the molecule mechanism
underlying the inhibitory effects of IMD on bone resorption.
Acknowledgments
This study was supported by the National Science
Foundation of China (grant no. 81370981).
References
|
1
|
Roh J, Chang CL, Bhalla A, Klein C and Hsu
SY: Intermedin is a calcitonin/calcitonin gene-related peptide
family peptide acting through the calcitonin receptor-like
receptor/receptor activity-modifying protein receptor complexes. J
Biol Chem. 279:7264–7274. 2004. View Article : Google Scholar
|
|
2
|
Takei Y, Inoue K, Ogoshi M, Kawahara T,
Bannai H and Miyano S: Identification of novel adrenomedullin in
mammals: A potent cardiovascular and renal regulator. FEBS Lett.
556:53–58. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chauhan M, Balakrishnan M, Yallampalli U,
Endsley J, Hankins GD, Theiler R and Yallampalli C: Adrenomedullin
2/intermedin regulates HLA-G in human trophoblasts. Biol Reprod.
85:1232–1239. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Takahashi K, Kikuchi K, Maruyama Y, Urabe
T, Nakajima K, Sasano H, Imai Y, Murakami O and Totsune K:
Immunocytochemical localization of adrenomedullin 2/intermedin-like
immunore-activity in human hypothalamus, heart and kidney.
Peptides. 27:1383–1389. 2006. View Article : Google Scholar
|
|
5
|
Takahashi K, Morimoto R, Hirose T, Satoh F
and Totsune K: Adrenomedullin 2/intermedin in the
hypothalamo-pituitary-adrenal axis. J Mol Neurosci. 43:182–192.
2011. View Article : Google Scholar
|
|
6
|
Chauhan M, Yallampalli U, Dong YL, Hankins
GD and Yallampalli C: Expression of adrenomedullin 2
(ADM2)/intermedin (IMD) in human placenta: Role in trophoblast
invasion and migration. Biol Reprod. 81:777–783. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
McLatchie LM, Fraser NJ, Main MJ, Wise A,
Brown J, Thompson N, Solari R, Lee MG and Foord SM: RAMPs regulate
the transport and ligand specificity of the
calcitonin-receptor-like receptor. Nature. 393:333–339. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pan CS, Yang JH, Cai DY, Zhao J, Gerns H,
Yang J, Chang JK, Tang CS and Qi YF: Cardiovascular effects of
newly discovered peptide intermedin/adrenomedullin 2. Peptides.
26:1640–1646. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fujisawa Y, Nagai Y, Miyatake A, Takei Y,
Miura K, Shoukouji T, Nishiyama A, Kimura S and Abe Y: Renal
effects of a new member of adrenomedullin family, adrenomedullin2,
in rats. Eur J Pharmacol. 497:75–80. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Holmes D, Campbell M, Harbinson M and Bell
D: Protective effects of intermedin on cardiovascular, pulmonary
and renal diseases: Comparison with adrenomedullin and CGRP. Curr
Protein Pept Sci. 14:294–329. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Granholm S, Lundberg P and Lerner UH:
Expression of the calcitonin receptor, calcitonin receptor-like
receptor, and receptor activity modifying proteins during
osteoclast differentiation. J Cell Biochem. 104:920–933. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Naot D and Cornish J: The role of peptides
and receptors of the calcitonin family in the regulation of bone
metabolism. Bone. 43:813–818. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Granholm S, Henning P and Lerner UH:
Comparisons between the effects of calcitonin receptor-stimulating
peptide and intermedin and other peptides in the calcitonin family
on bone resorption and osteoclastogenesis. J Cell Biochem.
112:3300–3312. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Uzan B, de Vernejoul MC and Cressent M:
RAMPs and CRLR expressions in osteoblastic cells after
dexamethasone treatment. Biochem Biophys Res Commun. 321:802–808.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
16
|
Hsu WL, Chen CY, Tsauo JY and Yang RS:
Balance control in elderly people with osteoporosis. J Formos Med
Assoc. 113:334–339. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Manolagas SC and Jilka RL: Bone marrow,
cytokines and bone remodeling. Emerging insights into the
pathophysiology of osteoporosis. N Engl J Med. 332:305–311. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Brunner M, Jurdic P, Tuckerman JP, Block
MR and Bouvard D: New insights into adhesion signaling in bone
formation. Int Rev Cell Mol Biol. 305:1–68. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Roodman GD: Advances in bone biology: The
osteoclast. Endocr Rev. 17:308–332. 1996.PubMed/NCBI
|
|
20
|
Roux S and Orcel P: Bone loss. Factors
that regulate osteoclast differentiation: An update. Arthritis Res.
2:451–456. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Reid IR: Anti-resorptive therapies for
osteoporosis. Semin Cell Dev Biol. 19:473–478. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Long F: Building strong bones: Molecular
regulation of the osteoblast lineage. Nat Rev Mol Cell Biol.
13:27–38. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Matsumura S, Hiranuma H, Deguchi A, Maeda
T, Jikko A and Fuchihata H: Changes in phenotypic expression of
osteoblasts after X irradiation. Radiat Res. 149:463–471. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Matsuguchi T, Chiba N, Bandow K, Kakimoto
K, Masuda A and Ohnishi T: JNK activity is essential for Atf4
expression and late-stage osteoblast differentiation. J Bone Miner
Res. 24:398–410. 2009. View Article : Google Scholar
|
|
25
|
Kim HK, Cho SG, Kim JH, Doan TK, Hu QS,
Ulhaq R, Song EK and Yoon TR: Mevinolin enhances osteogenic genes
(ALP, type I collagen and osteocalcin), CD44, CD47 and CD51
expression during osteogenic differentiation. Life Sci. 84:290–295.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Boyle WJ, Simonet WS and Lacey DL:
Osteoclast differentiation and activation. Nature. 423:337–342.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhao Q, Shao J, Chen W and Li YP:
Osteoclast differentiation and gene regulation. Front Biosci.
12:2519–2529. 2007. View
Article : Google Scholar
|
|
28
|
Simonet WS, Lacey DL, Dunstan CR, Kelley
M, Chang MS, Lüthy R, Nguyen HQ, Wooden S, Bennett L, Boone T, et
al: Osteoprotegerin: A novel secreted protein involved in the
regulation of bone density. Cell. 89:309–319. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Martin T, Gooi JH and Sims NA: Molecular
mechanisms in coupling of bone formation to resorption. Crit Rev
Eukaryot Gene Expr. 19:73–88. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Khosla S: Minireview: The OPG/RANKL/RANK
system. Endocrinology. 142:5050–5055. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kaji H, Kanatani M, Sugimoto T and Chihara
K: Statins modulate the levels of osteoprotegerin/receptor
activator of NFkappaB ligand mRNA in mouse bone-cell cultures. Horm
Metab Res. 37:589–592. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Giuliani N, Colla S, Sala R, Moroni M,
Lazzaretti M, La Monica S, Bonomini S, Hojden M, Sammarelli G,
Barillè S, et al: Human myeloma cells stimulate the receptor
activator of nuclear factor-kappa B ligand (RANKL) in T
lymphocytes: A potential role in multiple myeloma bone disease.
Blood. 100:4615–4621. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Udagawa N, Takahashi N, Akatsu T, Tanaka
H, Sasaki T, Nishihara T, Koga T, Martin TJ and Suda T: Origin of
osteoclasts: Mature monocytes and macrophages are capable of
differentiating into osteoclasts under a suitable microenvironment
prepared by bone marrow-derived stromal cells. Proc Natl Acad Sci
USA. 87:7260–7264. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jimi E, Akiyama S, Tsurukai T, Okahashi N,
Kobayashi K, Udagawa N, Nishihara T, Takahashi N and Suda T:
Osteoclast differentiation factor acts as a multifunctional
regulator in murine osteoclast differentiation and function. J
Immunol. 163:434–442. 1999.PubMed/NCBI
|
|
35
|
Abdallah BM, Stilgren LS, Nissen N, Kassem
M, Jorgensen HR and Abrahamsen B: Increased RANKL/OPG mRNA ratio in
iliac bone biopsies from women with hip fractures. Calcif Tissue
Int. 76:90–97. 2005. View Article : Google Scholar
|