Introduction
The epithelial-to-mesenchymal transition (EMT) is
considered to be an essential biological process, which comprises a
series of epithelial plasticity alterations between the epithelial
and mesenchymal states, and has an important role in physiological
and pathological processes (1,2).
During the progression of EMT, epithelial cells, which form
monolayers that exist in body tissues and organs, lose their
cell-cell adhesiveness, attain an elongated morphology and exhibit
an increased motility and invasiveness (1). In addition to these phenotypic
changes, the cells may also exhibit alterations in their gene
expression, including an upregulation of a variety of transcription
factors, including Snail, a downregulation of epithelial markers,
including E-cadherin, and an increased expression of mesenchymal
markers, such as α-smooth muscle actin (α-SMA) (1). Concomitantly with the changes
observed in the EMT markers, the further progression through the
stages of EMT is marked by the stimulation of epithelial cells,
which are transformed into myofibroblasts. EMT is considered to
contribute to fibrogenesis by providing a source of
myofibro-blasts, and also by activating a paracrine signaling
pathway between the epithelial and stromal cells (3–5).
Furthermore, a number of reviews reported on the role of EMT in
epithelial-mesenchymal interactions in the development of fibrotic
diseases (6,7). These findings provide substantial
evidence for an important role for the epithelium in fibrogenesis,
as a source of myofibroblasts, and as a mediator influencing the
development of myofibroblasts from epithelia cells.
MicroRNAS (miRNAs) form a class of small,
single-stranded, non-coding RNAs, which are 19–22 nucleotides in
length and negatively regulate gene expression by base-pairing with
the 3′ untranslated region (UTR) of cognate mRNAs, which have the
conserved seed sequence (8). These
non-coding RNAs are considered to exert a critical role in
physiological and pathological processes, ranging from embryonic
develop ment to tumorigenesis, depending on their specific gene
targets (9,10). An accumulating body of evidence
supports that miRNAs exert a crucial role in EMT through the
modulation of EMT-associated genes. In the cell model of EMT
induced by transforming growth factor-β (TGFβ1), previous studies
have demonstrated that all five members of the miR-200 family and
miR-205 are capable of repressing EMT by targeting the
transcription factors, zinc finger E-box binding homeobox 1 and 2
(ZEB1/2) (11–13). A further study revealed that
miR-155 exerts an essential role in TGFβ1-induced EMT through
targeting the protein, RhoA (14).
Taken together, these results suggested that miRNAs may be
crucially important in EMT, however, the biological functions of
the majority of the miRNAs in EMT remain to be fully
elucidated.
In the present study, ample evidence is provided to
support a role for miR-590 in EMT, based on studies performed in
TGFβ1-treated human kidney 2 (HK2) cells and a unilateral ureteral
obstruction (UUO) kidney model. The study aimed to investigate
whether the overexpression of miR-590 exerted any influence on EMT
in HK2 cells and in human renal tubular epithelial cells.
Furthermore, the results of the present study are discussed with
respect to potential effects on epithelium homeostasis, and the
consequences of these effects in kidney fibrosis.
Materials and methods
Prediction of miRNAs which target TGFβ
receptor 2 (R2)
The miRNAs targeting TGFβR2 were predicted using the
prediction programs: miRDB (http://www.microrna.org/), PicTar(http://pictar.mdc-berlin.de/) and TargetScan
(http://www.targetscan.org/). Only those
miRNAs that were predicted by all three algorithms were selected as
putative regulators of TGFβR2, and were suggested for further
experimental identification.
Cell culture and transfection
Human embryonic kidney 293 (HEK293T) and HK2 cells
were cultivated in Dulbecco's modified Eagle's medium and RPMI-1640
media (Invitrogen Life Technologies, Carlsbad, CA, USA),
respectively, supple mented with 10% fetal bovine serum (Invitrogen
Life Technologies), 100 U/ml penicillin and 100 µg/ml
streptomycin at 37°C in an atmosphere of 5% CO2. The
HEK293T and the HK2 cells were purchased from the American Tissue
Culture Collection (Manassas, VA, USA). For the transfection
experiments, when the cells had reached 60% confluence, the HK2
cells were transfected with pCDNA3.1 (800 ng), pCDNA3.1-mir-590
(800 ng), small-interfering (si) RNA-TGFβR2 (20 nM; sense and
antisense sequences, 5′-GUGCCUGUAACAUGGAAGATT-3′ and
5′-UCUUCCAUGUUACAGGCACTT-3′, respectively) or siRNA-control (20 nM;
sense and antisense sequences, 5′-UUCUCCGAACGUGUCACGUTT-3′ and
5′-ACGUGACACGUUCGGAGAATT-3′, respectively) using Lipofectamine™
2000 transfection reagent (Invitrogen Life Technologies), according
to the manufacturer's instructions.
Plasmid construction
To overexpress the miR-590, the 81 base pairs (bp)
genomic sequence, which encodes mature miR-590, was cloned into the
PUC57 vector (manufactured by Genewiz, Inc., South Plainfield, NJ,
USA). The sequence used was:
5′-AGTCAGAAATGAGCTTATTCATAAAAGTGCAGTATGGTGAAGTCAATCTGTAATTTTATGTATAAGCTAGTCTCTGATTGA-3′.
Subsequently, the premature sequence was transferred into pcDNA3.1
(Invitrogen Life Technologies) downstream of the cytomegalovirus
(CMV) promoter between the restriction sites HindIII and
EcoRI. For the luciferase reporter plasmids, the 3′ UTR of
the mouse TGFβR2, or its mutant variations, was amplified by
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) from the mouse genome, and inserted downstream of the
luciferase coding sequence of the pcDNA3.1-luciferase reporter
plasmid between the restriction sites BamHI and
EcoRI. For the overexpression of TGFβR2, the coding sequence
was cloned to generate the plasmid pCMV-tag-2B-TGFβR2 using the
restriction sites of BamHI and Xhol, and (forward:
5′-CGCGGATCCCTGTCCACTTGCGACAACCAG-3′ and reverse:
5′-GGCCTCGAGTTTGGTAGTGTTCAGCGAGC-3′). All the plasmids used in the
present study were verified by sequencing.
Generation of the EMT model in vitro
EMT was induced in the HK2 cells in the presence of
TGFβ1. Cells that had reached 50% confluence were treated with
recombinant human TGFβ1 (10 ng/ml; R&D Systems, Minneapolis,
MN, USA) in RPMI-1640 medium for 48 h.
Generation of the UUO kidney model
The UUO kidney model was generated in male C57BL/6
mice (12–16 weeks old, weighing 24–28 g), which were purchased from
the Animal Center of Chongqing Medical University. Animals were
treated according to the recommendations in the Guide for the Care
and Use of Laboratory Animals of the National Institutes of Health.
The experimental procedures were approved by the Animal
Experimental Ethics Committee of Chongqing Medical University. The
animals were operated on by left ureteral ligation, as described
previously (15,16). Briefly, following anesthetization
with sodium pentobarbital (50 mg/kg), the middle portion of the
left ureter was ligated completely by surgical suture, whereas the
control group underwent an identical process without ligation. The
mice were sacrificed on day 14 following the ligation surgery, and
the kidneys were harvested for subsequent studies. These
experiments were approved by the Medicine Animal Care Committee of
the Chongqing Medical University.
Histochemical analysis
The kidneys were dissected, fixed with 10% neutral
formalin for 48 h, and embedded in paraffin. Briefly, tissues were
immersed in acetone at 4°C, transferred to a freezer at −20°C, and
fixed overnight. Then the tissues were dehydrated in acetone at 4°C
for 15 min and in acetone for another 15 min at room temperature.
Thereafter the tissues were cleared twice in methyl benzoate for 15
min, and twice in xylene for 15 min at room temperature. Finally
they were penetrated with paraffin (melting point, 58–60°C; Junsei
Chemical Co., Ltd., Tokyo, Japan) at 60°C for 2–3 h in a
vacuum-evaporating embedder and embedded in paraffin. Subsequently,
6-mm thick sections were selected and stained with Masson's stain
for the determination of the tubulointerstitial index, as
previously described (17–19).
TGFβR2 3′ UTR-luciferase reporter
assay
The luciferase reporter analyses were performed on
HEK293T cells. For the transfection assay, the HEK293T cells were
seeded (1×105 cells/well) in 24-well plates. The
plasmids pcDNA3.1-Luc-TTGFβR2WT, pcDNA3.1-Luc-TGFβR2Mut and
pcDNA3.1-Luc were transfected into HEK293T cells, which stably
expressed pcDNA3.1-miR-590 or pcDNA3.1, using Lipofectamine™ 2000
in OptiMEM medium (Thermo Fisher, Waltham, MA, USA). At 48 h later,
the cells were harvested with PBS, and lysed with
radioimmunoprecipitation assay (RIPA) buffer. The dual-luciferase
reporter assay system (Promega Corp., Madison, WI, USA) was used
for the analysis, and the data are shown as the ratio of luciferase
activity normalized against that of Renilla.
RT-qPCR
RT-qPCR was used to determine the mRNA levels. The
total RNA was isolated from cells or kidneys using
Trizol® (Invitrogen Life Technologies), and 5 µg
total RNA was selected to be reverse-transcribed using the
First-Strand cDNA Synthesis kit (Thermo Fisher Scientific)
according to the manufacturer's instructions. Aliquots of 0.5
µl transcription product served as the template for the qPCR
reactions with UltraSYBR mixture (CWBio, Inc., Beijing, China) or
UltraSYBR mixture (Takara Bio, Inc., Otsu, Japan). The mRNA
expression levels of E-cadherin, α-SMA and collagen were normalized
to 18S RNA, whereas the expression level of miR-590 was normalized
to U6 RNA. Primers for the detection of the miRNAs were as follows.
The primers for miR-590 were provided by Ruibo (Guangzhou, China);
the U6 primers were 5′-CTCGCTTCGGCAGCACA-3′ (forward) and
5′-AACGCTTCACGAATTTGCGT-3′ (reverse). The primers for the detection
of the mRNAs were as follows: 18S primers were
5′-GTAACCCGTTGAACCCCATT-3′ (forward) and 5′-CCATCCAATCGGTAGTAGCG-3′
(reverse); the mouse E-cadherin primers, 5′-GTCAACACCTACAACGCTGC-3′
(forward) and 5′-ACGTGCTTGGGTTGAAGA-3′ CA (reverse); the mouse
α-SMA primers, 5′-GCTATGCCCTGCCTCATGC-3′ (forward) and
5′-TCACGGACAATCTCACGCTC-3′ (reverse); the mouse collagen primers,
5′-AACTTTGCTTCCCAGATGTCCTATG-3′ (forward) and
5′-GCTTCCCCATCATCTCCATTCTTGC-3′ (reverse); the human E-cadherin
primers, 5′-GGTGCTCTTCCAGGAACCTC-3′ (forward) and
5′-GGAAACTCTCTCGGTCCAGC-3′ (reverse); the human α-SMA primers,
5′-GGGGTGATGGTGGGAATG-3′ (forward) and 5′-GCAGGGTGGGATGCTCTT-3′
(reverse); and the human collagen primers were
5′-AAGGTGTTGTGCGATGACG-3′ (forward) and 5′-TGGTCGGTGGGTGACTCTG-3′
(reverse). All the experiments were performed in triplicate. The
expression levels were quantified using the 2−ΔCt
method. ΔCt was calculated either as Ct (E-cadherin/
α-SMA/collagen) - Ct (i8s) or as Ct (miR-590) - Ct (U6).
Protein isolation and western
blotting
The HK2 cells were washed three times with 100
µl ice-cold PBS buffer and lysed by adding 200 µl
RIPA buffer/well immediately, with a 30-min incubation period. For
the kidney tissue, the kidneys were homogenized in lysis buffer
(RIPA) containing 1 mg/l protease cocktail (BioTool, Jupiter, FL,
USA; containing 104 mM AEBSF, 80 µM aprotinin, 5 mM
bestatin, 1.5 mM E-64, 2 mM leupeptin, 1.5 mM pepstatin A), prior
to sonication Xo-1200D (Heng Long Instrument Co., Ltd., Changzhou,
China) at 4°C for 60 sec. The total protein concentration of each
sample was measured using the bicinchoninic acid protein assay with
a microplate procedure (Beyotime Institute of Biotechnology,
Haimen, China), prior to western blotting. Samples of 30 µg
total protein were subjected to 12% SDS-PAGE, and the protein bands
were subsequently transferred onto polyvinylidene fluoride
membranes (Millipore Corp., Bedford, MA, USA). The membranes were
blocked with 5% non-fat milk in Tris-buffered saline (TBS)/Tween 20
(0.1%) at room temperature for 1 h. Subsequently, the membranes
were immersed in primary antibody solution [rabbit polyclonal
anti-E-cadherin, 1:3,000 (20648-1-AP; Proteintech Group Inc.,
Chicago, IL, USA); rabbit polyclonal anti-laminin, 1:2,500
(sc-20145; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA);
rabbit polyclonal anti-TGFβR2, 1:2,000 (our laboratory); or mouse
monoclonal anti-β-tubulin, 1:5,000 (HC101-01 TransGen, Beijing,
China)] at 4°C for an overnight incubation. After washing three
times with TBS/Tween 20 (0.1%; 5 min each wash), incubation with
horseradish peroxidase-conjugated goat anti-rabbit and mouse
secondary antibody (1:5,000, Beijing Zhongshan Jinqiao
Biotechnology Co., Ltd., Beijing, China) secondary antibodies was
performed at room temperature for 1 h (1:5,000 dilution). The
immunoreactivity was developed using chemiluminescent horseradish
peroxidase substrate reagent (Millipore Corp.), and the signal was
detected using the Bio-Rad ChemiDoc XRS+imaging system (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). β-tubulin was used as a
protein loading control, and western blot analyses were performed
in triplicate.
Immunofluorescent analysis
For the detection of immunofluorescence, HK2 cells
were cultured with sterile glass coverslips in six-well plates. The
glass slides were treated with 95% alcohol for 15 min at room
temperature prior to cell culture. Subsequently, the cell
coverslips were washed three times with 500 µl PBS and
permeabilized with 0.1% Triton X-100 for 30 min. Following blocking
with 10% normal goat serum for 1 h at room temperature, the cells
were incubated with rabbit anti-laminin primary antibody (1:100;
Santa Cruz Biotechnology, Inc.) for 3 h at room temperature. The
cover-slips were subsequently washed three times with PBS, prior to
incubation with fluorescein isothiocyanate-conjugated goat
anti-rabbit immunoglubulin G at room temperature for 1 h. The
slides were examined under a Nikon (Melville, NY, USA) Eclipse
TE300 fluorescence microscope, and images were captured with a SPOT
Diagnostic (Sterling Heights, MI, USA) charged-couple device
camera.
Statistical analysis
GraphPad Prism 5 (San Diego, CA, USA) software was
used to analyze the experimental data. The statistical differences
between groups, including the bar graph of the luciferase assay and
the results from the RT-qPCR experiments, were analyzed by one-way
analysis of variance. All the experiments were performed at least
in triplicate, and the statistical errors of averaged data are
presented as the mean ± standard error of the mean. P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-590 is decreased during EMT in vitro
and in vivo
To investigate whether miR-590 has a direct function
on EMT, TGFβ1-induced epithelial cells were used as a cell model,
and UUO mice were an animal model, for modulating EMT in vitro and
in vivo, respectively. TGFβ1-induced EMT experiments were performed
in HK2 cells according to the method described previously (20,21),
whereas the UUO model was generated in mice by left ureter ligation
(22). Following treatment with
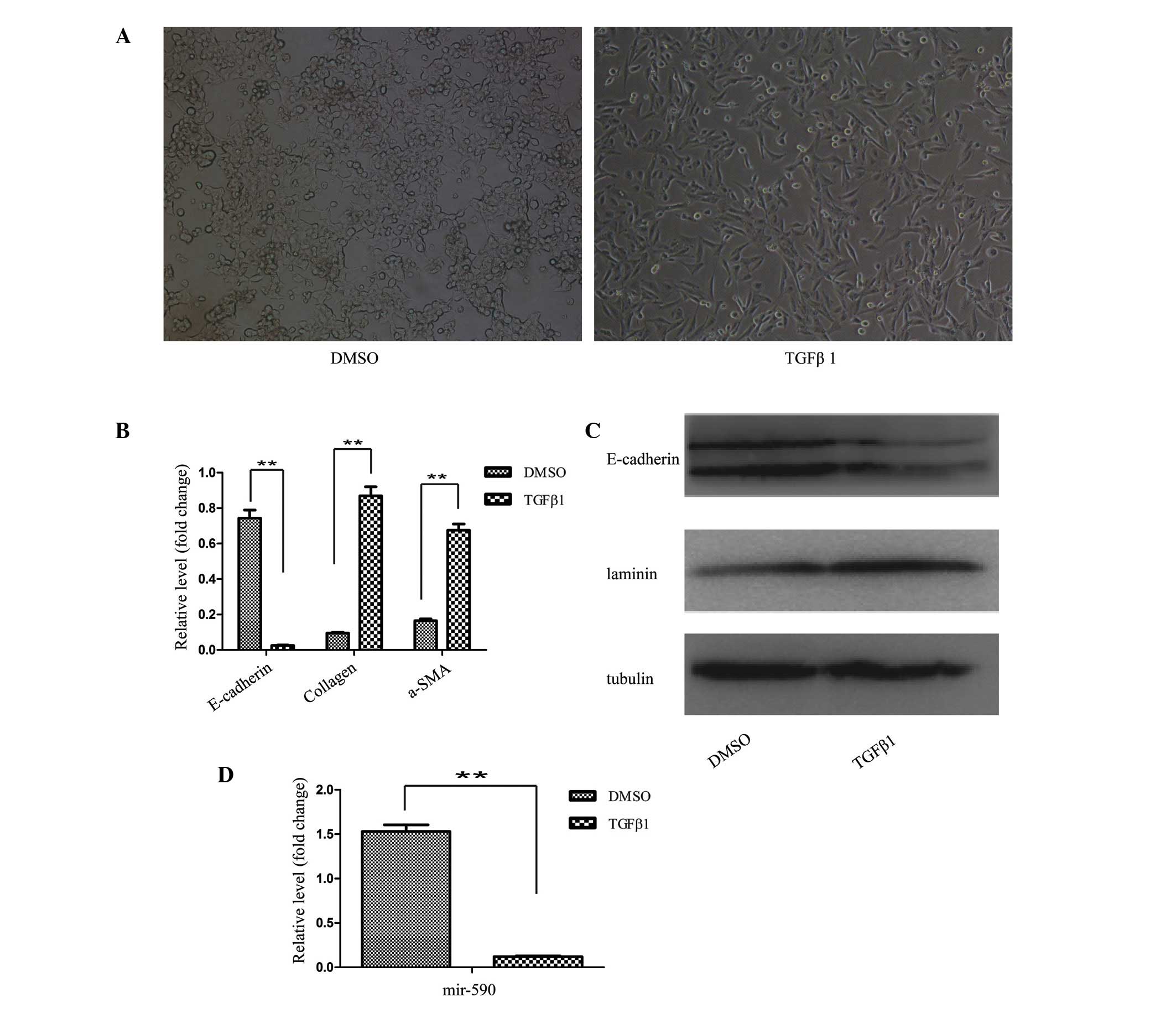
TGFβ1 for 48 h, the HK2 cells were transformed from the epithelial
state, characterized by polygonal morphology and tight connections,
into a spindle-like mesenchymal morphology, which appeared as the
HK2 cells underwent the process of EMT (Fig. 1A). Subsequently, the EMT process at
the molecular level was identified by determining the levels of the
marker genes. The results from the RT-qPCR analysis revealed
decreased mRNA expression levels of the epithelial marker
(E-cadherin), and increased expression levels of the mesenchymal
markers, collagen and α-SMA, following TGFβ1 treatment of the HK2
cells (Fig. 1B). Western blot
analysis also confirmed the EMT process.
As shown in Fig.
1C, downregulation of the protein expression level of
E-cadherin, and an upregulation of the protein expression level of
laminin, were observed in TGFβ1-treated HK2 cells. Furthermore, the
expression level of miR-590 in HK2 cells was significantly
decreased following TGFβ1 treatment, as revealed by the RT-qPCR
analysis (Fig. 1D). These results
indicated an inverse correlation between the expression of
EMT-associated genes and miR-590. To investigate the association
between miR-590 and the EMT process in renal fibrosis, the
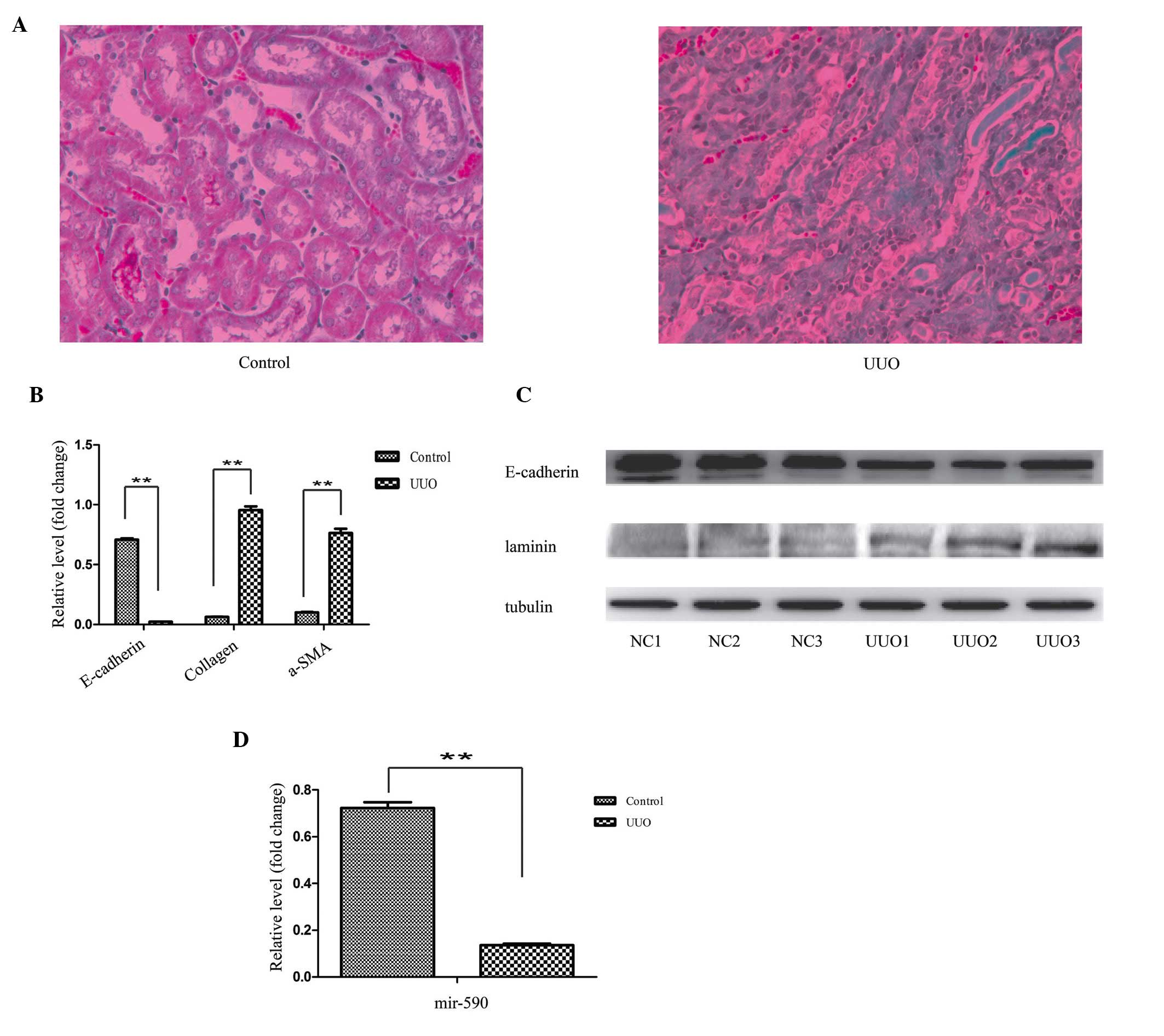
identical experiments were performed in the UUO kidney. Prior to
the experiments, the efficiency of the UUO model was confirmed by
Masson staining.
The histopathological results revealed substantial
structural alterations, induced by left ureter ligation. In the
control specimens, the kidney tissues were well aligned and
surrounded by only a small quantity of interstitial tissue, whereas
the section of the kidney from the UUO-induced mice featured a
large proportion of fibres (Fig.
2A). For the EMT analysis, consistently with the cell model, a
decreased mRNA expression level of E-cadherin, and an increased
mRNA expression level of collagen and α-SMA, were observed in the
RT-qPCR experiment (Fig. 2B). The
western blot analysis revealed a decreased protein expression level
of E-cadherin, and an increased protein expression level of
laminin, in the UUO kidney (Fig.
2C). Similarly, the expression level of miR-590 was
significantly decreased in the UUO animals compared with the
control (Fig. 2D).
mir-590 downregulates TGFβR2 by targeting
its 3′ UTR
Since a pronounced downregulation of miR-590 was
revealed in the EMT model, to investigate the role exerted by
miR-590 in EMT regulation, a search of the prediction software
programs, TargetScan, miRanda and PicTar, was performed. It was
observed that the 3′ UTR of the TGFβR2 mRNA contained a potential
site for miR-590 (23).
Subsequently, conservation of the 3′ UTR of TGFβR2 was analyzed by
blasting the sequences between different species, according to the
University of California Santa Cruz genome database (Santa Cruz).
As shown in Fig. 3A, the entire 3′
UTR of TGFβR2 was 2,698 bp long, and was highly conserved among
different species, including humans, rats, rabbits, shrews, tree
shrews and dogs. Subsequently, the validation of the association
between miR-590 and TGFβR2 was assessed. Given that it was
considered to be one of the most effective miRNAs in inhibiting the
luciferase activity, the plasmid pCDNA-3.1-miR-590 was used in the
present study. To examine whether miR-590 regulates TGFβR2
expression by directly binding to the predicted 3′ UTR sequence,
luciferase reporter plasmids containing the wild-type or the
mutated binding site for miR-590 were constructed (Fig. 3B). These results revealed that the
overexpression of miR-590 significantly suppressed the luciferase
activity of the wild-type TGFβR2 3′ UTR reporter, although not that
of the mutant reporter containing a mutation in the miR-590-binding
site (Fig. 3C). To examine the
physiological importance of miR-590 in regulating TGFβR2
production, the effect of miR-590 on TGFβR2 production in HK2 cells
was assessed. The western blot analysis revealed that the
transfection of HK2 cells with pCDNA-3.1-miR-590 markedly reduced
the protein expression level of TGFβR2 (Fig. 3D). These results demonstrated that
miR-590 is able to negatively regulate the expression of TGFβR2 by
directly targeting its 3′ UTR.
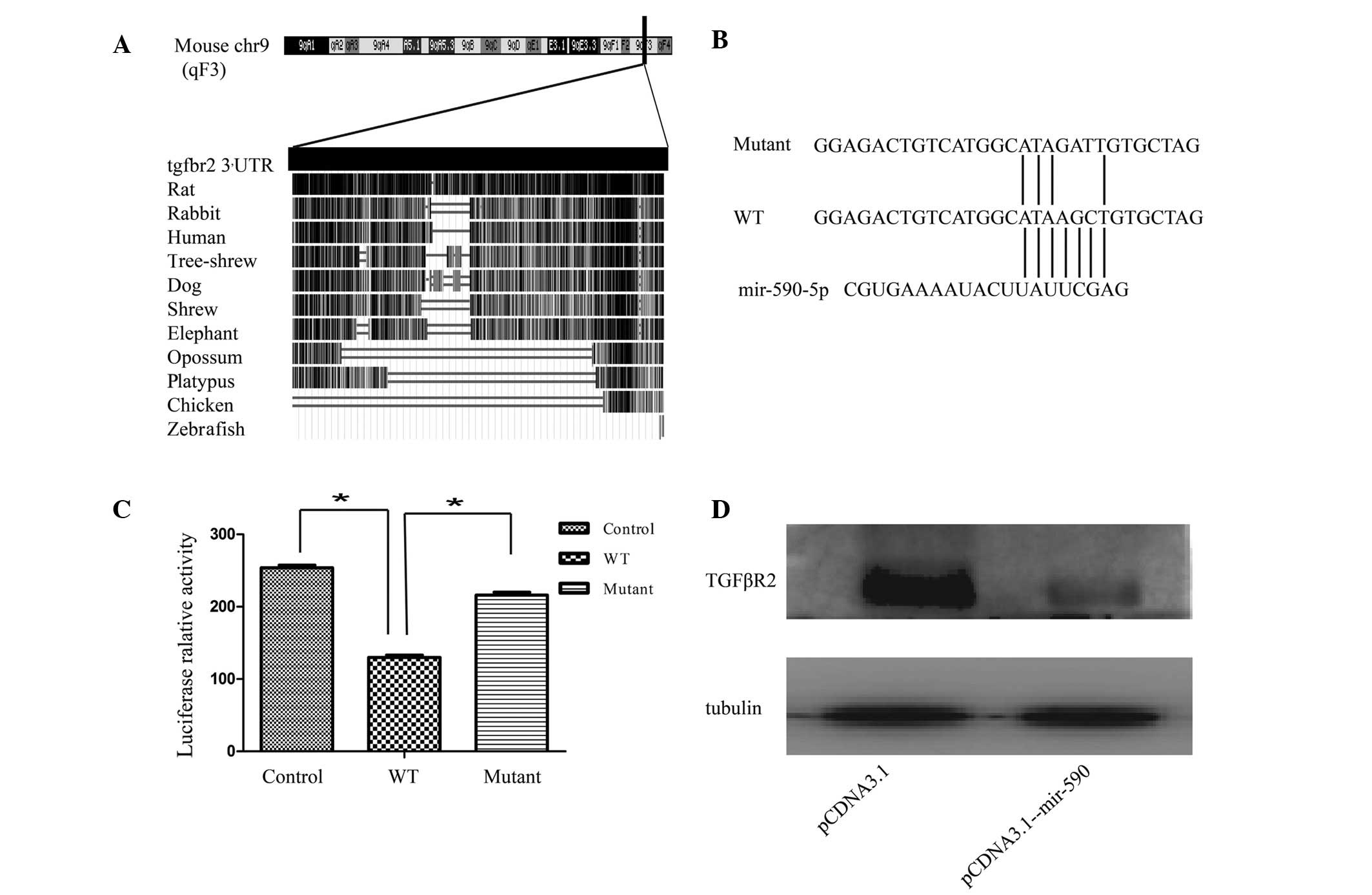 | Figure 3mir-590 downregulates TGFβR2 by
targeting its 3′ UTR. (A) Bioinformatics analysis of the 3′ UTR of
TGFβR2. The 3′ UTR of TGFβR2 is 2,698 bp in length, and is highly
conserved among mammals, as revealed by the University of
California, Santa Cruz genome database. (B and C) Analysis of the
luciferase activity in HEK293T cells. HEK293T cells were
transiently co-transfected with the luciferase reporter plasmid,
containing the wild-type TGFβR2 3′ UTR, or mutant variations, in
the presence of pCDNA3.1-mir-590 or pCDNA3.1. (D) Effects of
miR-590 on the endogenous TGFβR2 level in HK2 cells. The expression
level was analyzed by western blotting. The data are presented as
fold changes compared with the control cells. Data are presented as
the mean ± standard deviation, and are representative of at least
three independent experiments. *P<0.05, compared with
the control. chr9, chromosome 9; WT, wild-type; UTR, untranslated
region; TGFβR2, transforming growth factor receptor β2. |
Effects of miR-590 and TGFβR2 on EMT in
HK2 cells
A consistent correlation between the expression
levels of miR-590 and E-cadherin was observed, and a marked
downregulation of miR-590 was identified in the EMT model.
Furthermore, the expression of endogenous miR-590 was higher in HK2
cells compared with L929 cells (mice embryonic myofibroblasts; data
not shown) indicating that the downregulation of miR-590 may
explain phenotypic stabilization of mesenchymal features in the
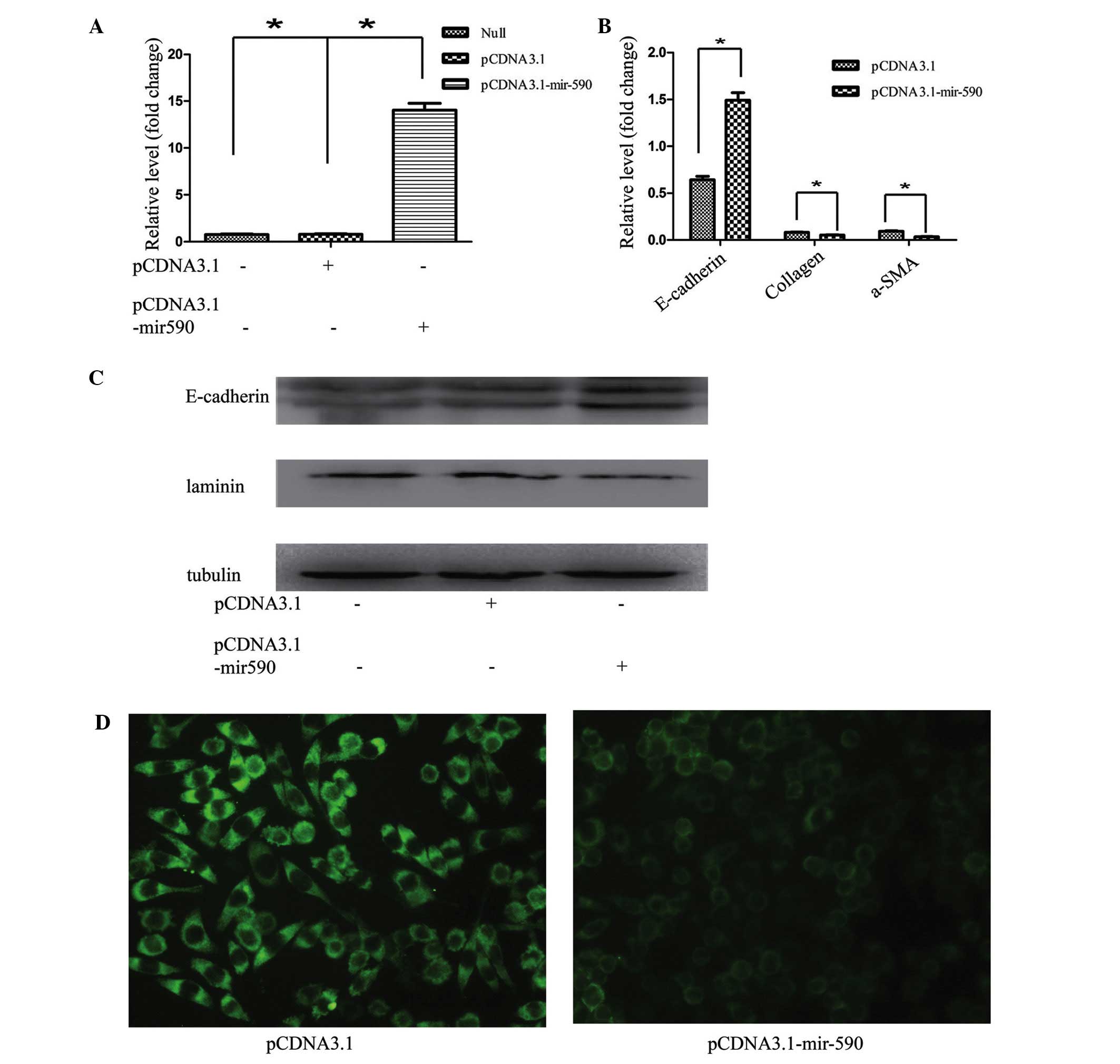
L929 cell line. To investigate the role of miR-590 in EMT, HK2
cells were transfected with pCDNA3.1-mir-590, and the expression
level of mir-590 was significantly increased (Fig. 4A). The induction of miR-590
markedly inhibited EMT in HK2 cells, as revealed by epithelial
marker changes. The RT-qPCR results demonstrated that the
expression of the miR-590 plasmid in HK2 cells also resulted in a
decrease in the mRNA expression of other known fibroblast markers,
including α-SMA and collagen I, and an increased mRNA expression of
E-cadherin (Fig. 4B). Furthermore,
the western blotting and immunofluorescence results revealed that
transfection of the miR-590 plasmid markedly reduced the expression
of laminin, and upregulated E-cadherin expression in HK2 cells
(Fig. 4C and D). A previous study
in our laboratory revealed that TGFβ1 induced EMT in vitro,
accompanied by the upregulation of laminin, and the downregulation
of E-cadherin, at the mRNA and protein levels in HK2 cells, whereas
the transfection of exogenous miR-590 markedly attenuated the EMT
process in HK2 cells.
The TGFβR2 3′ UTR has a functional site for miR-590,
and it is well known that TGFβR2 exerts an important role in the
process of TGFβ-mediated signal transduction by linking TGFβ1 to
its receptor (TGFβR2) and causing the activation of TGFβ receptor
type I (TGFβR1) kinase, leading to phosphorylation of the Smad2 and
-3 complexes (24). To examine the
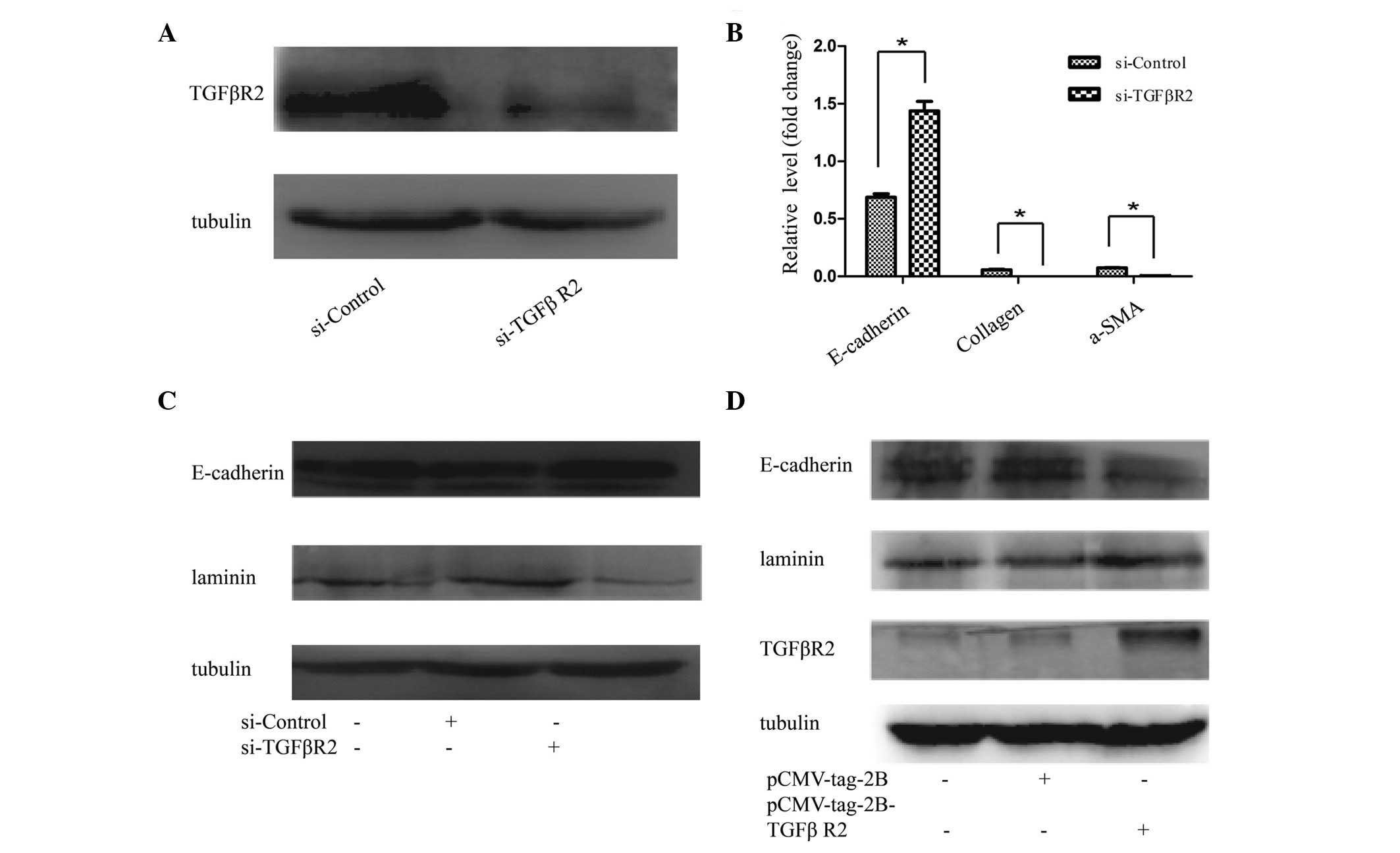
effect of TGFβR2 on EMT, TGFβR2 siRNA or scrambled siRNA was
introduced into HK2 cells. As shown in Fig. 5A, TGFβR2 siRNA effectively
suppressed the protein levels of TGFβR2, and prevent EMT in HK2
cells. The results revealed that E-cadherin expression was induced,
whereas the mRNA expression levels of collagen and α-SMA were
reduced (Fig. 5B). HK2 cells
transfected with TGFβR2-specific siRNA exhibited increased protein
levels of E-cadherin and a reduced protein expression of laminin,
compared with the negative controls (Fig. 5C). By contrast, the upregulation of
TGFβR2 with pcmv-tag2b-TGFβR2 in the HK2 cells was able to reverse
the EMT-associated gene expression, as demonstrated by the
increased protein expression levels of laminin, and reduced protein
expression levels of E-cadherin, revealed in the western blot
analysis of cells transfected with pcmv-tag2b-TGFβR2 compared with
transfected pcmv-tag2b vector (Fig.
5D).
Discussion
Regardless of the type of kidney disease, fibrosis
is unavoidably the outcome, resulting in the marked destruction of
the kidney structure, with consequential functional deterioration
(24). Myofibroblasts contribute
to the accumulation of the extracellular matrix observed in
fibrotic disease, and it was demonstrated that myofibroblasts may
originate from renal residential fibroblasts (25,26).
In addition to the residential fibroblasts, other sources of
fibroblasts have been studied, including fibrocytes and pericytes.
The EMT of mature tubular epithelial cells is another source of
fibroblasts, which are fundamentally linked to the process of renal
fibrosis (27,28). As revealed in the present study,
the mRNA expression level of E-cadherin in the epithelial cells was
markedly decreased, whereas the mesenchymal markers, collagen I and
α-SMA, were increased in the EMT model in vitro and in vivo.
Consistent with the results of the RT-qPCR analysis, the western
blotting data further confirmed that the EMT process existed in
TGFβ1-treated HK2 cells and the UUO model, as demonstrated by the
lower expression levels of E-cadherin and higher expression levels
of laminin. Furthermore, previous studies revealed that a
proportion of the fibroblasts, which are predominantly the
functional cells in this process, originate from the tubular
epithelial cells in the diseased kidney by EMT (28–30),
which has an important role in the process of kidney disease.
Therefore, understanding the regulatory mechanisms of EMT may help
to elucidate the events involved in the process of fibrosis,
leading to the development of novel therapeutics for treating
kidney disease.
miRNAs are endogenously encoded, non-coding RNAs
which predominantly post-transcriptionally regulate gene expression
through either translational suppressors or mRNA degradation
(8). miRNAs, a class of newly
identified regulators, were demonstrated to be involved in the
process of EMT. In vitro and animal studies revealed that these
miRNAs (miR-200a, miR-200b and miR-205) suppress EMT by targeting
ZEB1 and ZEB2 (11–13). Other studies demonstrated that
miR-655 and miR-34a are associated with EMT-suppressive miRNAs
(31,32). The present study has revealed that
miR-590 causes EMT, as demonstrated by the evidence that, to the
best of our knowledge, for the first time, miR-590 affects EMT in
HK2 cells. Transfection of the HK2 cells with pCDNA3.1-mir-590
resulted in an inhibition of a number of the fibrotic changes, and
the promotion of various epithelial characteristics. Therefore,
miR-590 appears to exert a key role in EMT, and may promote
fibrosis.
Previous studies revealed that miR-590 has multiple,
experimentally validated target genes, including TGFβR2, protein
polybromo-1, S100A10 and the cell adhesion molecule L1-like gene
(33–36). Among these targets, TGFβR2 was
revealed to be associated with EMT in epithelial cells. To
investigate the mechanism underlying the involvement of miR-590 in
EMT, the present study has revealed that miR590 directly targets
TGFβR2, as demonstrated by the luciferase reporter activity assays
and western blot analyses. Protein levels of TGFβR2 were markedly
decreased following miR-590 overexpression. Additionally,
E-cadherin expression was upregulated and the expression of laminin
was decreased. TGFβR2 was identified as a functional target of
miR-590, and this target is well established as a component of the
TGFβ signaling pathway. The TGFβ signaling pathway consists of
TGFβ, TGFβR, the Smad protein family and several other important
transcriptional regulatory factors. The TGFβR family comprises
three subtypes, TGFβR1, TGFβR2 and TGFβR3, although only TGFβR2 may
accept TGFβ1 as a protein partner independently of the other two
receptors, which are unable to function in the absence of TGFβR2
(37). Furthermore, a previous
study demonstrated that mir-655 is an EMT-suppressive miRNA, which
targets TGFβR2 (32). In the
present study, it was also revealed that the knockdown of TGFβR2
with siRNA elicited a similar pattern of alteration of the
EMT-associated genes, whereas the transfection of pcmv-tag2B-TGFβR2
reversed the protein expression, results which are consistent with
the previous hypothesis. Therefore, mir-590 was identified as an
EMT-suppressive miRNA in the present study.
In conclusion, the results reported in the present
study provide the first evidence, to the best of our knowledge,
that miR-590 is involved in the process of EMT in tubular
epithelial cells. The overexpression of miR-590 markedly increased
E-cadherin expression and suppressed laminin expression in HK2
cells, clearly indicating that this miRNA is an inhibitor of EMT.
Furthermore, TGFβR2, which is a core component of the TGFβ
signaling pathway, was characterized as a direct target of miR-590.
The present results suggest that the EMT-suppressor miR-590,
targeting TGFβR2, has the potential to serve as a biomarker of, and
a potential therapeutic target for, fibrosis.
Acknowledgments
This study was supported by the National Natural
Science Foundation of China (no. 31271563), the National Basic
Research Program of China (no. 2011CB944002) and the Chongqing
Application Development Project (no. cstc2014yykfB10003).
References
|
1
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu Y: Epithelial to mesenchymal
transition in renal fibrogenesis: Pathologic significance,
molecular mechanism and therapeutic intervention. J Am Soc Nephrol.
15:1–12. 2004. View Article : Google Scholar
|
|
3
|
Schrimpf C and Duffield JS: Mechanisms of
fibrosis: the role of the pericyte. Curr Opin Nephrol Hypertens.
20:297–305. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lin SL, Kisseleva T, Brenner DA and
Duffield JS: Pericytes and perivascular fibroblasts are the primary
source of collagen-producing cell sin obstructive fibrosis of the
kidney. Am J Pathol. 173:1617–1627. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Humphreys BD, Lin SL, Kobayashi A, Hudson
TE and Nowlin BT: Fate tracing reveals the pericyte and not
epithelial origin of myofibroblasts in kidney fibrosis. Am J
Pathol. 176:85–97. 2010. View Article : Google Scholar :
|
|
6
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Thiery JP: Epithelial-mesenchymal
transitions in development and pathologies. Curr Opin Cell Biol.
15:740–746. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
He L and Hannon GJ: MicroRNAs: Small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang J and Ma L: MicroRNA control of
epithelial-mesenchymal transition and metastasis. Cancer Metastasis
Rev. 31:653–662. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Akkina S and Becker BN: MicroRNAs in
kidney function and disease. Transl Res. 157:236–240. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gregory PA, Bert AG, Paterson EL, Barry
SC, Tsykin A, Farshid G, Vadas MA, Khew-Goodall Y and Goodall GJ:
The miR-200 family and miR-205 regulate epithelial to mesenchymal
transition by targeting ZEB1 and SIP1. Nat Cell Biol. 10:593–601.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Paterson EL, Kolesnikoff N, Gregory PA,
Bert AG, Khew-Goodall Y and Goodall GJ: The microRNA-200 family
regulates epithelial to mesenchymal transition. Scientific World
Journal. 8:901–904. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Korpal M, Lee ES, Hu G and Kang Y: The
miR-200 family inhibits epithelial-mesenchymal transition and
cancer cell migration by direct targeting of E-cadherin
transcriptional repressors ZEB1 and ZEB2. J Biol Chem.
283:14910–14914. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kong W, Yang H, He L, Zhao JJ, Coppola D,
Dalton WS and Cheng JQ: MicroRNA-155 is regulated by the
transforming growth factor beta/Smad pathway and contributes to
epithelial cell plasticity by targeting RhoA. Mol Cell Biol.
28:6773–6784. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen YT, Chang FC, Wu CF, Chou YH, Hsu HL,
Chiang WC, Shen J, Chen YM, Wu KD, Tsai TJ, Duffield JS and Lin SL:
Platelet-derived growth factor receptor signaling activates
pericyte myofibroblast transition in obstructive and post-ischemic
kidney fibrosis. Kidney Int. 80:1170–1181. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yang J, Shultz RW, Mars WM, Wegner RE, Li
Y, Dai C, Nejak K and Liu Y: Disruption of tissue type plasminogen
activator gene in mice reduces renal interstitial fibrosis in
obstructive nephropathy. J Clin Invest. 110:1525–1538. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Morales C, González GE, Rodríguez M,
Bertolasi CA and Gelpi RJ: Histopathologic time course of
myocardial infarct in rabbit hearts. Cardiovasc Pathol. 11:339–345.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Junqueira LC, Bignolas G and Brentani RR:
Picrosirius staining plus polarization microscopy, a specific
method for collagen detection in tissue sections. Histochem J.
11:447–455. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen L, Faulhaber-Walter R, Wen Y, Huang
Y, Mizel D, Chen M, Sequeira Lopez ML, Weinstein LS, Gomez RA,
Briggs JP and Schnermann J: Renal failure in mice with Gsalpha
deletion in juxtaglomerular cells. Am J Nephrol. 32:83–94. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kaimori A, Potter J, Kaimori JY, Wang C,
Mezey E and Koteish A: Transforming growth factor-beta 1 induces an
epithelial-to-mesenchymal transition state in mouse hepatocytes in
vitro. J Biol Chem. 282:22089–22101. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Aresu L, Rastaldi MP, Scanziani E, Baily
J, Radaelli E, Pregel P and Valenza F: Epithelial-mesenchymal
transition (EMT)of renal tubular cells in canine
glomerulonephritis. Virchows Arch. 451:937–942. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Huang XR, Chung AC, Wang XJ, Lai KN and
Lan HY: Mice overexpressing latent TGF-beta 1 are protected against
renal fibrosis in obstructive kidney disease. Am J Physiol Renal
Physiol. 295:F118–F127. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang J, Zhang H, Liu J, Tu X, Zang Y, Zhu
J, Chen J, Dong L and Zhang J: MiR-30 inhibits TGF-b1-induced
epithelial-to-mesenchymal transition in hepatocyte by targeting
Snail1. Biochem Biophys Res Commun. 417:1100–1105. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu Y: Cellular and molecular mechanisms
of renal fibrosis. Nat Rev Nephrol. 7:684–696. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hills CE and Squires PE: The role of TGF-β
and epithelial-to mesenchymal transition in diabetic nephropathy.
Cytokine Growth Factor Rev. 22:131–139. 2011.PubMed/NCBI
|
|
26
|
Barnes JL and Gorin Y: Myofibroblast
differentiation during fibrosis: Role of NAD(P)H oxidases. Kidney
Int. 79:944–956. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lan HY: Tubular epithelial-myofibroblast
transdifferentiation mechanisms in proximal tubule cells. Curr Opin
Nephrol Hypertens. 12:25–29. 2003. View Article : Google Scholar
|
|
28
|
Zeisberg M and Duffield JS: Resolved: EMT
produces fibroblasts in the kidney. J Am Soc Nephrol. 21:1247–1253.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang J and Liu Y: Blockage of tubular
epithelial to myofibroblast transition by hepatocyte growth factor
prevents renal interstitial fibrosis. J Am Soc Nephrol. 13:96–107.
2002.
|
|
30
|
Iwano M: EMT and TGF-beta in renal
fibrosis. Front Biosci (Schol Ed). 2:229–238. 2010. View Article : Google Scholar
|
|
31
|
Du R, Sun W, Xia L, Zhao A, Yu Y, Zhao L,
Wang H, Huang C and Sun S: Hypoxia-induced downregulation of
microRNA-34a promotes EMT by targeting the Notch signaling pathway
in tubular epithelial cells. PLoS One. 7:e307712012. View Article : Google Scholar
|
|
32
|
Harazono Y, Muramatsu T, Endo H, Uzawa N,
Kawano T, Harada K, Inazawa J and Kozaki K: MiR-655 is an EMT-
suppressive microRNA targeting ZEB1 and TGFBR2. PLoS One.
8:e627572013. View Article : Google Scholar
|
|
33
|
Chu Y, Ouyang Y, Wang F, Zheng A, Bai L,
Han L, Chen Y and Wang H: MicroRNA-590 promotes cervical cancer
cell growth and invasion by targeting CHL1. J Cell Biochem.
115:847–853. 2014. View Article : Google Scholar
|
|
34
|
Xiao X, Tang C, Xiao S, Fu C and Yu P:
Enhancement of proliferation and invasion by MicroRNA-590-5p via
targeting PBRM1 in clear cell renal carcinoma cells. Oncol Res.
20:537–544. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shan X, Miao Y, Fan R, Qian H, Chen P, Liu
H, Yan X, Li J and Zhou F: MiR-590-5P inhibits growth of HepG2
cells via decrease of S100A10 expression and inhibition of the Wnt
pathway. Int J Mol Sci. 14:8556–8569. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jiang X, Xiang G, Wang Y, Zhang L, Yang X,
Cao L, Peng H, Xue P and Chen D: MicroRNA-590-5p regulates
proliferation and invasion in human hepatocellular carcinoma cells
by targeting TGF-β RII. Mol Cells. 33:545–551. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Shi Y and Massagué J: Mechanisms of
TGF-beta signaling from cell membrane to the nucleus. Cell.
113:685–700. 2003. View Article : Google Scholar : PubMed/NCBI
|