Introduction
Ovarian cancer is the most lethal gynecological
malignancy, and one of the leading causes of morbidity and
mortality worldwide (1). In
ovarian carcinoma, cancer cells frequently metastasize by
implanting onto the peritoneal mesothelial surface of the abdominal
cavity. The resulting peritoneal implants are characterized by the
adhesion, migration, and invasion of the tumor cells into the
peritoneum and underlying organs (2).
Integrins are a large family of cell surface
adhesion proteins that are involved in epithelial cell-matrix
interactions. Upregulation of integrins has been associated with
malignancy (3,4), particularly during invasion,
metastasis, and angiogenesis (5,6).
Increasing evidence suggests that aberrantly expressed integrins
have a role in ovarian cancer pathophysiology (7–9).
Integrin β1 (ITGB1), regulates broader functional activities such
as cellular proliferation, adhesion, and invasion, and previous
studies have suggested its implication in therapeutic resistance in
numerous solid cancer models (10–12)
and hematopoietic malignancies (13,14).
Focal adhesion kinase (FAK) is upregulated in various malignancies,
such as breast, colon, prostate, neck, and ovarian cancer (15,16).
FAK is an important regulator of survival, proliferation,
migration, and invasion, processes that are all involved in
tumorigenesis and metastasis (17,18).
Therefore, FAK signaling may be regarded as a potential target in
the development of anticancer drugs. A previous study demonstrated
that FAK, identified as a downstream kinase of ITGB1, has an
important role in integrin-stimulated signaling events. Integrin
clustering induces autophosphorylation of FAK and activates
signaling pathway networks (19).
However, the mechanisms underlying ITGB1 inhibition-induced
suppression of ovarian cancer progression, and the contribution of
ITGB1 inhibition in anticancer therapy remains to be elucidated.
The present study investigated the effects of ITGB1 inhibition on
tumor apoptosis, invasion and migration, and bevacizumab anticancer
therapy in ovarian cancer cells. Furthermore, the present study
investigated the signaling pathways that may mediate the
anti-invasive and anti-migratory effects that resulted from ITGB1
inhibition.
Materials and methods
Ovarian cancer cell culture
Human ovarian surface epithelial cells (HOSEpic),
HO-8910 and HO-8910PM ovarian cancer cells were used in the present
study. HO-8910PM cells are highly metastatic ovarian cancer cells
derived from HO-8910 (20). The
cell lines were grown in RPMI 1640 (GE Healthcare Life Sciences,
Hudson, NH, USA) supplemented with 10% fetal bovine serum
(Invitrogen Life Technologies, Carlsbad, CA, USA), 1% penicillin
and streptomycin (Sigma-Aldrich, St. Louis, MO, USA), in an
atmosphere containing 5% CO2, at 37°C. The cell lines
were purchased from the Cell Bank of Shanghai Institutes for
Biological Sciences, the Chinese Academy of Science (Shanghai,
China).
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the cells with TRIzol
(Invitrogen Life Technologies) and reverse transcribed into cDNA
with reverse transcriptase from the Primescript RT Reagent kit
(Takara Biotechnology Co., Ltd., Dalian, China), as described
previously (21). The RT-qPCR
reactions were performed using the Thermal Cycler Dice Real-Time
PCR system (Takara Biotechnology Co., Ltd.) and the ABI 7300
Thermal Cycler (Applied Biosystems Life Technologies, Carlsbad, CA,
USA) with the following cycling conditions: Initial denaturation at
95°C for 10 min, and 40 cycles of 95°C for 30 sec, 55°C for 30 sec,
and 72°C for 2 min. In order to ensure the reproducibility of the
results, all the genes were tested in triplicate. The following
primers from BGI Tech (Shenzhen, China) were used for the present
study: ITGB1, sense 5′-GTCTCAGACTGGCTTCAGTG-3′, antisense
5′-ATGATTGTACCGAGGCTGTC-3′; FAK, sense 5′-GCGGCCCAGGTTTACTGAA-3′,
antisense 5′-GGCCTGTCTTCTGGACTCCAT-3′; and β-actin (human), sense
5′-AGGGGCCGGACTCGTCATACT-3′, and antisense
5′-GGCGGCACCACCATGTACCCT-3′. Reaction of each sample was performed
in triplicate. Relative expression levels were calculated using the
2-∆∆Ct method.
Lentivirus packaging and infection
Three precursor small hairpin RNA sequences
targeting ITGB1 (GenBank accession no. NM_133376; GTG TAC AGA TCC
GAA GTT TCA) were designed using an Internet application system
(http://rnaide-signer.lifetechnologies.com/rnaiexpress/plate.jsp;
Invitrogen Life Technologies). The anti-ITGB1 sequence was inserted
into a lentiviral vector gene (Shanghai GenePharma Co., Ltd.,
Shanghai, China) in order to construct a recombinant lentiviral
vector GV248, as previously described (22). HO-8910 and HO-8910PM cells in
6-well plates (2×105 cells/well) were transfected with
42 µl ITGB1-shRNA lentiviral vector (LV; 2.40×108
transducing units) or 12 µl negative control (NC) lentiviral
vector (LV-NC; 1.8×109 transducing units) at 37°C. A
total of 5 µg/ml polybrene (Sigma-Aldrich) was added to
enhance the transfection efficiency. A selection medium
supplemented with 2.5 µg/ml puromycin (Sigma-Aldrich) was
used to obtain a stable transfected cell line over the course of 2
weeks.
Treatment of cells with bevacizumab and
fludarabine
The ovarian cancer cells transfected with the
LV-ITGB1-RNAi vector were treated with 10 µg/ml bevacizumab
(Avastin®; Genentech, Inc., South San Francisco, CA,
USA) for 24 h, and then cell apoptosis, adhesion and invasion were
detected. The HO-8910 and HO-8910PM cells were treated with 1.54
µg/ml fludarabine (Selleck Chemicals, Houston, TX, USA) for
24 h, and then protein expression, adhesion and invasion were
detected.
Western blot analysis
Whole cell extracts were prepared using
radioimmunoprecipitation assay lysis reagent (Sigma-Aldrich),
according to the manufacturer's instructions. The proteins were
then quantified using a bicinchoninic acid assay (Pierce
Biotechnology, Inc., Rockford, IL, USA), prior to being separated
by 10% SDS-PAGE, and detected by western blotting on a
polyvinylidene difluoride membrane (EMD Millipore, Billerica, MA,
USA). The membranes were blocked with 3 ml Tris-buffered saline
with Tween 20 (TBST; Auragene Bioscience Co., Changsha, China) with
5% bovine serum albumin (Auragene Bioscience Co.) for 2 h at room
temperature. The membranes were incubated overnight at 4°C with the
following primary antibodies: Rabbit polyclonal anti-ITGB1
(sc-13590; 1:200), anti-STAT1 (sc-345; 1:500), anti-phosphorylated
STAT1 (sc-135648; 1:500), anti-FAK (sc-558; 1:500), anti-matrix
metalloproteinase (MMP)-2 (sc-13594; 1:500) and MMP-9 (sc-21733;
1:200) primary antibodies (Santa Cruz Biotechnology, Inc., Dallas,
TX, USA). The membranes were incubated for 1 h at 37°C with the
anti-β-actin antibody (sc-8979; 1:6,000; Santa Cruz Biotechnology,
Inc.). The membranes were washed 3 times with TBST, then incubated
with the horseradish peroxidase-conjugated goat anti-rabbit
immunoglobulin G secondary antibody (Pierce Biotechnology, Inc.),
followed by 3 additional washes with TBST. A SuperSignal West Femto
Enhanced Chemiluminescence Detection system (Pierce Biotechnology,
Inc.) was used for detection. The blots were analyzed with the
CanoScan LiDE 110 scanner (Canon, Inc., Tokyo, Japan) and Image
Pro-Plus software, version 6.0 (Media Cybernetics, Inc., Rockville,
MD, USA).
In vitro adhesion assay
The cells were pretreated with or without
bevacizumab (10 µg/ml) for 24 h. The cells were then
suspended in serum-free Dulbecco's modified Eagle's medium (GE
Healthcare Life Sciences) to form a single-cell suspension, prior
to being seeded at a density of 105 cells/well into
96-well plates precoated with Matrigel™ (BD Biosciences, Franklin
Lakes, NJ, USA). The wells were incubated at 37°C for 50 min, and
washed three times with phosphate-buffered saline (PBS) in order to
remove non-adherent cells. Cell viability was determined using an
MTT assay.
Cell survival assay
An MTT assay was used to estimate cell viability
(23). Briefly, the cells were
plated at a density of 1×104 cells/well in 96-well
plates. Following exposure to the original and the stably
lentivirus infected cell lines of ovarian cancer, with or without
bevacizumab (10 µg/ml) treatment for 24 h, the cells were
incubated with MTT (Beijing Solarbio Science & Technology Co.,
Ltd., Beijing, China) at a final concentration of 0.5 mg/ml for 4 h
at 37°C. The medium was carefully discarded, and 150 mM dimethyl
sulfoxide (Beijing Solarbio Science & Technology Co., Ltd.) was
added to dissolve the formazan crystals. The absorbance was
measured at 570 nm using a multi-well scanning spectrophotometer
reader (756MC; Jinghua Instruments, Shanghai, China). The cells in
the control group were considered 100% viable.
Flow cytometry
An Annexin V and propidium iodide (PI) Fluorescein
Staining kit (Bender MedSystems GmbH, Wien, Austria) were used to
measure cell apoptosis, according to the manufacturer's
instructions. Briefly, 5×105 cells were suspended in 500
µl 1X binding buffer containing 10 mM HEPES (pH 7.4), 140 mM
NaCl, and 2.5 mM CaCl2. The cells were then incubated
with annexin V (1:20) for 5 min, prior to being incubated with PI
(1 mg/ml) for 15 min. The rate of apoptosis was evaluated by flow
cytometry (MoFlo™ XDP; Beckman Coulter, Inc., Brea, CA, USA).
Invasion assay
The cells were cultivated to 80% confluence in
12-well plates. Cellular growth was subsequently observed for 24 h.
All the experiments were repeated in triplicate. Cell invasion was
determined by the Transwell assay. Cells suspended in serum-free
medium were added into the upper chamber of the insert with
Matrigel (BD Biosciences). Following incubation for 24 h at 37°C,
cells remaining on the upper side of the membrane were carefully
removed, while cells that had migrated through the membrane were
fixed with 75% alcohol (Auragene Bioscience Co.) and stained with
crystal violet (Sigma-Aldrich) for 25 min, washed with water and
air-dried. The cells that invaded the membrane were counted using a
light microscope (AE31; Motic, Xiamen, China).
Migration assay
For the wound healing assay, cells were seeded in
12-well plates and grown to 90% confluence. Monolayers in the
center of the wells were scraped with pipette tips and washed with
PBS. Cell movement into the wound area was monitored and the images
were captured at 0 and 24 h using the AE31 light microscope. The
migration distance between the leading edge of the migrating cells
and the edge of the wound was compared, as previously described
(24).
Statistical analysis
Each experiment was repeated in triplicate. Data are
presented as the mean ± standard deviation, and analyzed using SPSS
18.0 (SPSS, Inc., Chicago, IL, USA). Statistical comparisons
between the groups were analyzed using two-tailed Student's
t-tests. P<0.05 was considered to indicate a statistical
significant difference.
Results
Expression levels of ITGB1 and FAK in
ovarian cancer cells
To assess whether ITGB1 and FAK were dysregulated in
ovarian cancer cells, the expression levels of ITGB1 and FAK in
normal ovarian cells (HOSEpic) and ovarian cancer cells (HO-8910
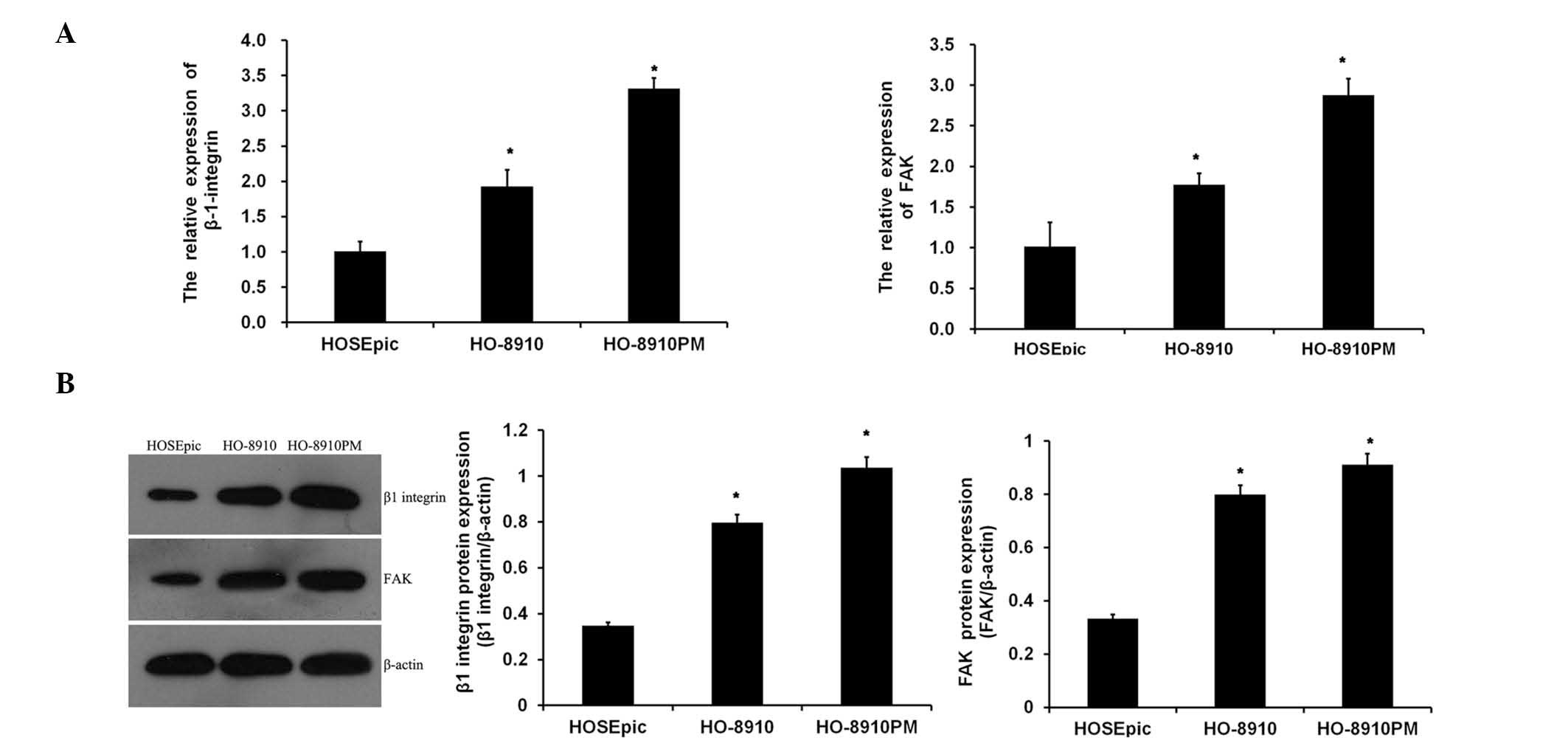
and HO-8910PM) were analyzed. As shown in Fig. 1A, the mRNA expression levels of
ITGB1 and FAK were significantly increased in metastatic ovarian
cancer cell lines, as compared with the normal ovarian cells, with
the expression levels of ITGB1 and FAK being the highest in the
highly metastatic HO-8910PM cell line. Similar results were
obtained from the western blot analysis (Fig. 1B). These data suggest that
suppression of ITGB1 may be associated with carcinogenesis of
ovarian cancer.
ITGB1 inhibition suppresses cell invasion
and MMP expression in ovarian cancer cells
Previous studies have demonstrated that ITGB1 may
mediate tumor growth, apoptosis, adhesion, and signaling in various
cancer cells (7,22). Due to the fact that invasion is an
important factor for cancer expansion and metastasis, the present
study examined whether ITGBI inhibition altered the rate of
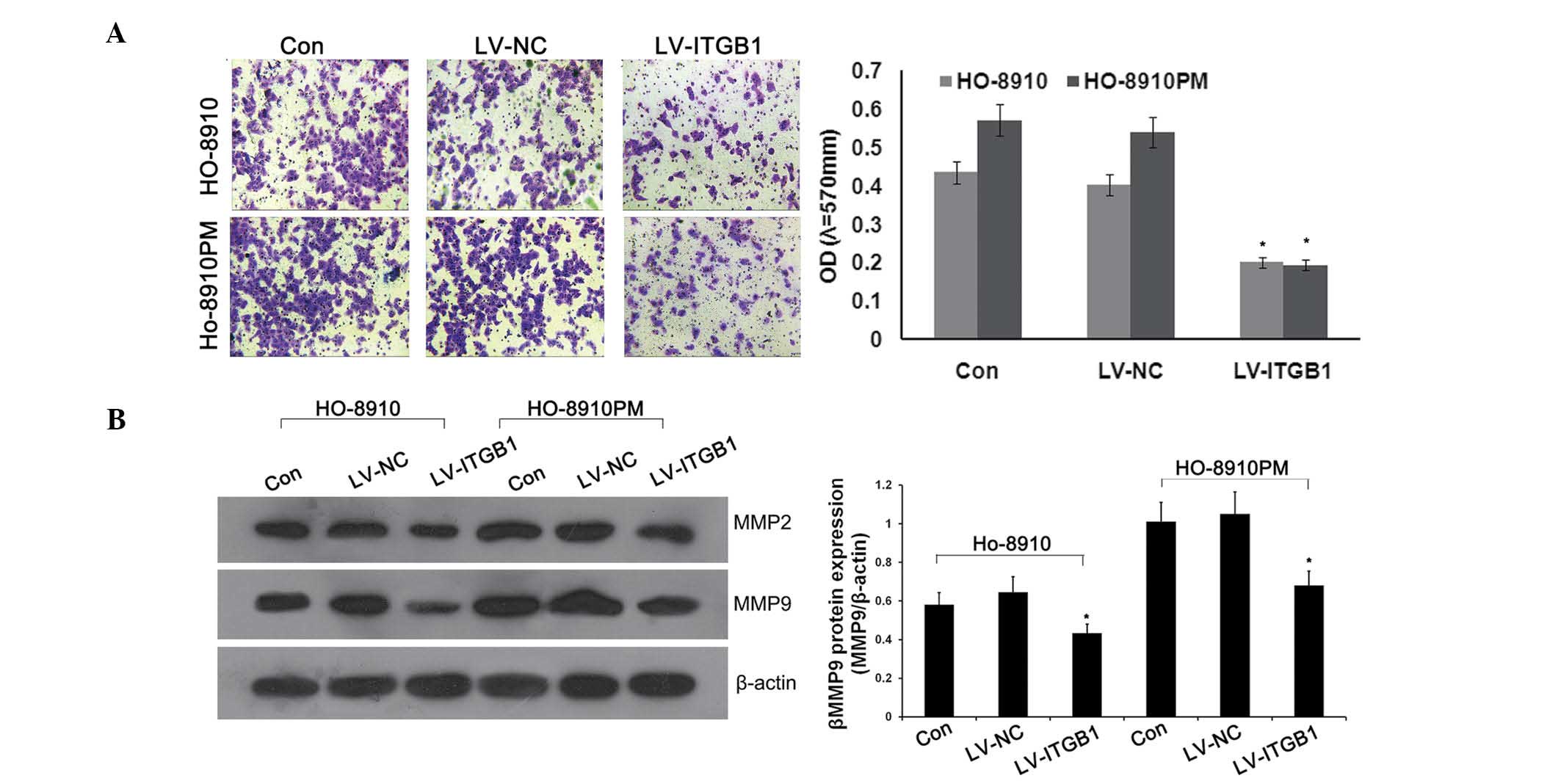
invasion of cancer cells through transwell chambers coated with
Matrigel™. Anti-ITGB1 lentiviral vectors were transfected into
HO-8910 and HO-8910PM cells to inhibit the expression of ITGB1.
Concordant with the previous results, significant attenuation of
ovarian cancer cell invasion was observed in the lentivirus
(LV)-ITGB1-RNA interference (RNAi) group, as compared with the
control and LV-NC groups (Fig.
2A).
MMPs, specifically MMP-2 and MMP-9, are the most
important proteases for the degradation of type IV collagen, which
is the predominant constituent of extracellular matrix (ECM)
(25,26), and has an important role in cancer
cell metastasis. In order to determine the effects of ITGB1
inhibition on MMP-2 and MMP-9 protein expression, western blotting
was performed. As shown in Fig.
2B, ITGB1 knockdown significantly downregulated the protein
expression levels of MMP-2 and MMP-9 in the ITGB1-RNAi group, as
compared with the control and LV-NC groups. These results suggest
that ITGB1 inhibition suppresses MMP-2 and MMP-9 expression.
ITGB1 inhibition enhances the effect of
bevacizumab treatment
Bevacizumab offers promise as an antiangiogenic in
the treatment of ovarian cancer (27); however, acquired resistance limits
the use of bevacizumab as a therapeutic agent. The present study
demonstrated the effects of the combination of ITGB1 inhibition and
bevacizumab treatment on apoptosis, adhesion and migration of
ovarian cancer cells. Ovarian cancer cells with knocked down ITGB1
were treated with 10 µg/ml bevacizumab for 24 h, and cell
apoptosis, adhesion, and invasion were detected.
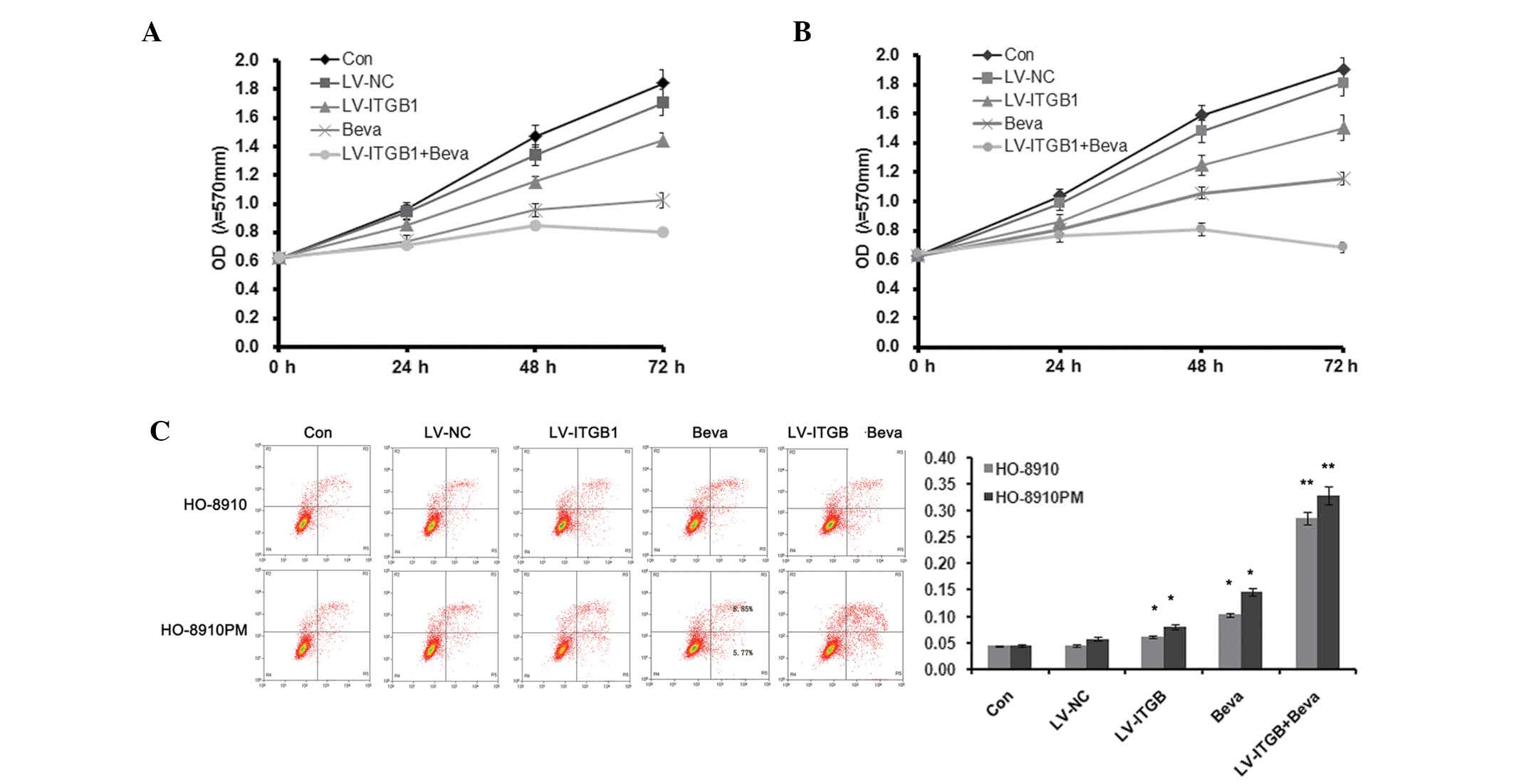
As shown in Fig. 3,
MTT assay and flow cytometry were used to detect HO-8910 and
HO-8910PM cell apoptosis. Cellular proliferation markedly decreased
following ITGB1 inhibition and treatment with 10 µg/ml
bevacizumab (Fig. 3A and B).
Furthermore, ITGB1 inhibition further reduced cell proliferation in
bevacizumab-treated ovarian cancer cells. Concordant with the
results of the MTT assay, Annexin V/PI staining demonstrated that
the percentage of apoptotic cells was significantly increased in
both the LV-ITGB1 and bevacizumab (Beva) groups, as compared with
the control group. In addition, ITGB1 inhibition further increased
cell apoptosis in bevacizumab-treated ovarian cancer cells. These
results suggest that ITGB1 inhibition regulates bevacizumab therapy
by decreasing cell proliferation and increasing cell apoptosis.
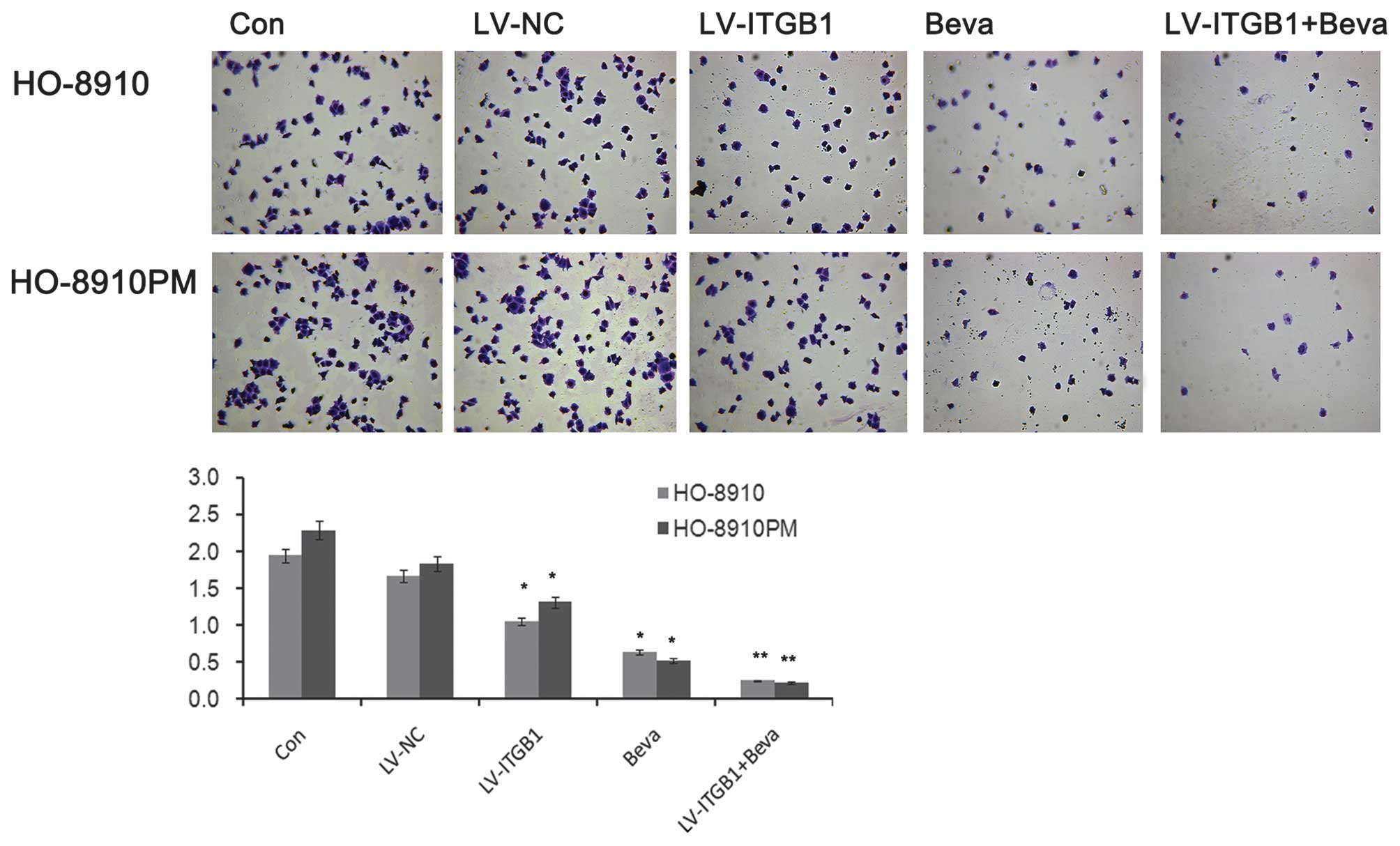
The adhesion of cancer cells to endothelial cells or
the ECM has an important role in establishing metastasis (20). Integrin regulates adhesion and
signaling in ovarian cancer (7).
The present study evaluated the effects of ITGB1 silencing on
ovarian cancer cell adhesion to ECM proteins. The number of
adherent cells markedly decreased following ITGB1 inhibition
(Fig. 4). The inhibitory effects
of cell adhesion induced by ITGB1 silencing may contribute to the
inhibition of cell metastasis. Concordant with the results of the
apoptosis analysis, ITGB1 inhibition enhanced the effects of
bevacizumab treatment against cell adhesion in ovarian cancer. As
shown in Fig. 4, significant
attenuation of ovarian cancer cell adhesion was observed in the
LV-ITGB1 + Beva group, as compared with the Beva group.
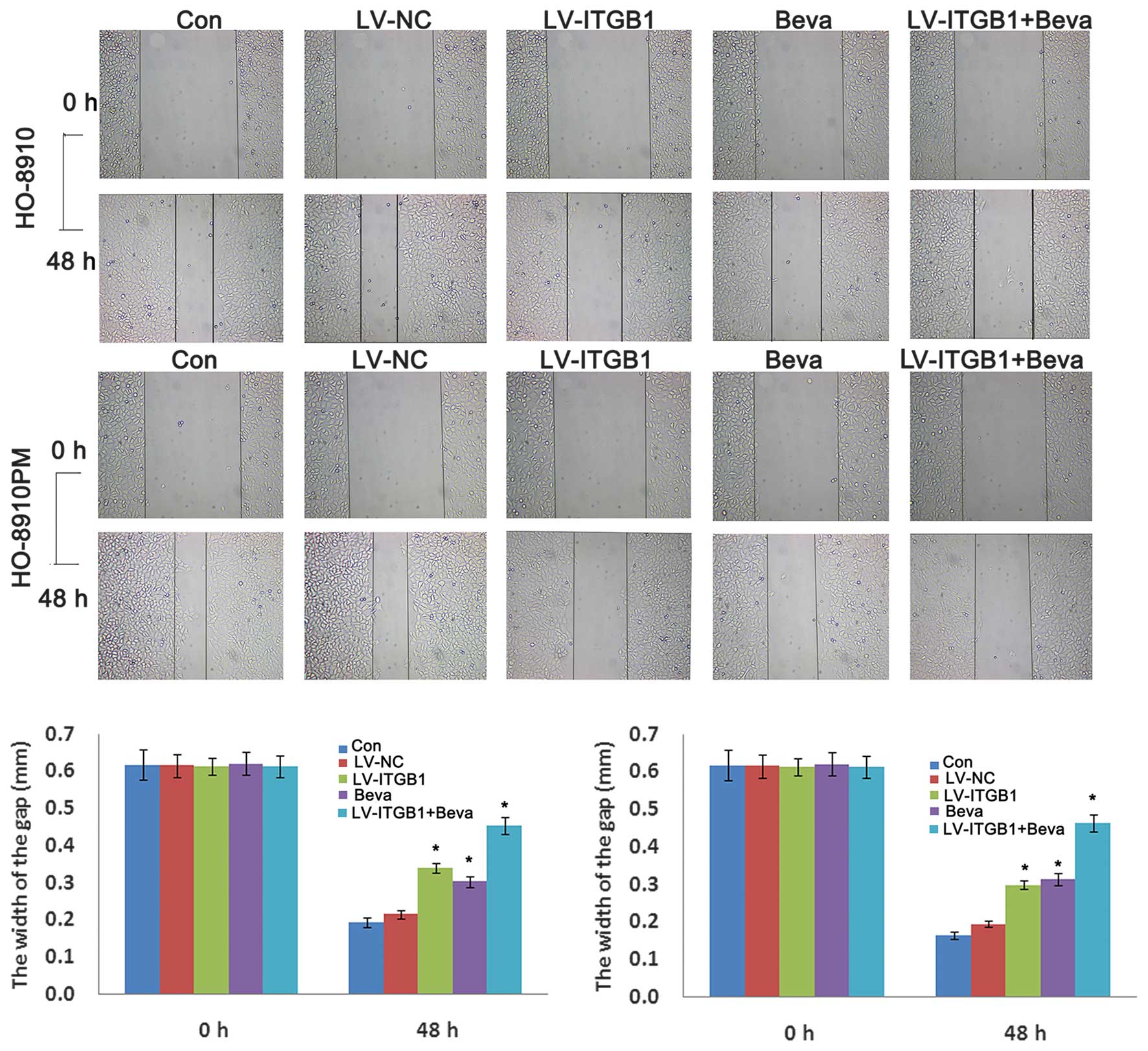
The inhibitory effects of ITGB1 suppression on
ovarian cancer cell migration were determined by wound healing
assay. HO-8910 and HO-8910PM cells were transfected with ITGB1-RNAi
and treated with bevacizumab, and wound closure was monitored and
the images of the wound closure were captured. The results of the
wound healing assay indicated that ITGB1 inhibition significantly
attenuated HO-8910 and HO-8910PM cell migration after 48 h
(Fig. 5). These data indicate that
ITGB1 silencing may effectively inhibit HO-8910 and HO-8910PM cell
motility. Furthermore, cell migration was significantly decreased
in the LV-ITGB1 + Beva group, as compared with the Beva group.
These data suggest that the the inhibitory effects of combination
treatment of bevacizumab and ITGB1 have an improved effect compared
with bevacizumab simple treatment alone on HO-8910 and HO-8910PM
cell migration.
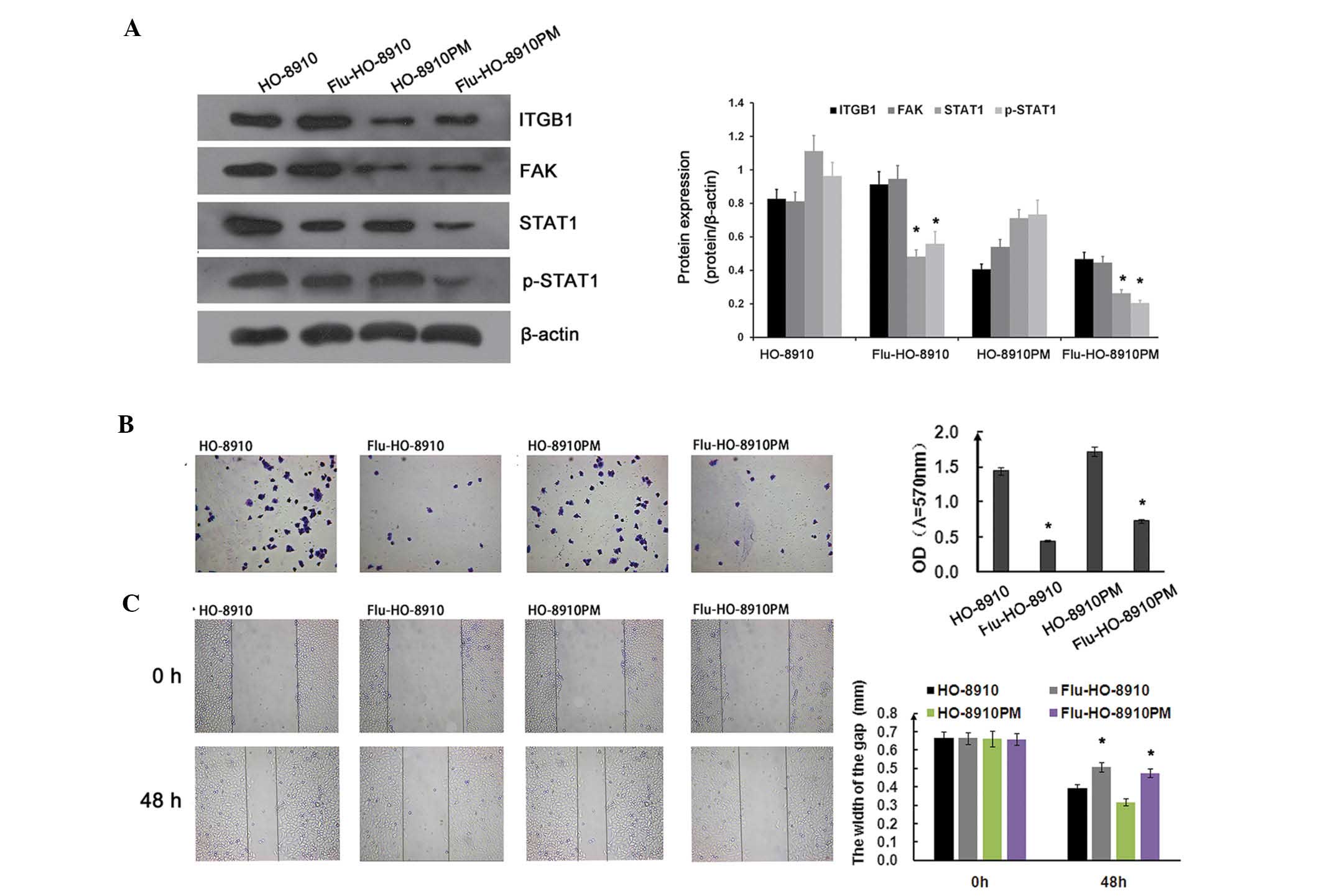
ITGB1/FAK/STAT1 pathway in ovarian cancer
cells
The subsequent experiments of the present study
focused on the signaling pathways involved in ITGB1-mediated
suppression of metastatic potential in ovarian cancer cells. To
determine whether the STAT1 signaling pathway was involved in cell
adhesion and migration in ovarian cancer, HO-8910 and HO-8910PM
cells were treated with 1.54 µg/ml fludarabine for 24 h.
ITGB1 and FAK expression were not significantly affected by
fludarabine. However, STAT1 and p-STAT1 expression were markedly
downregulated by fludarabine (Fig.
6A). As shown in Fig. 6B, the
adhesion ratio was significantly decreased in the fludarabine
group, as compared with the control group. Concordant with the
adhesion assay results, fludarabine significantly inhibited ovarian
cancer cell migration, as determined by the wound healing assay
(Fig. 6C). FAK and c-Src form a
dual kinase complex, which functions to promote cell motility, cell
cycle progression, and cell survival. Previous studies have
demonstrated that ITGB1-mediated FAK signaling is closely
associated with tumor growth and metastasis (19,28).
In a previous study, co-immunoprecipitation and in vitro
binding assays demonstrated that STAT1 was transiently and directly
associated with FAK during cell adhesion (29), and its activity was induced by the
integrin signaling pathway. These results indicate that the
ITGB1/FAK/STAT1 pathway is involved in cell adhesion and migration
in ovarian cancer.
Discussion
Integrin-mediated cell adhesion and migration have
essential roles in cell growth and development. Previous studies
have demonstrated that ITGB1 is able to mediate ovarian carcinoma
cell adhesion, invasion, and migration (8,30).
In the present study, the anti-metastatic effects of ITGB1
inhibition on the HO-8910 and HO-8910PM ovarian cancer cell lines,
as well as its molecular mechanism of action, were investigated.
ITGB1 inhibition induced cell apoptosis, which was determined by
the inhibition of cell adhesion, migration, and invasion, as well
as by the suppression of MMP-2 and MMP-9 expression. The results of
the present study also demonstrated that ITGB1 inhibition enhanced
bevacizumab treatment in ovarian cancer. Furthermore, the
inhibition of STAT1 signaling by fludarabine revealed that the
ITGB1/FAK/STAT1 pathway may be associated with the molecular
mechanisms that underlie the anti-invasive effects of ITGB1
inhibition.
Metastasis is closely associated with cancer
therapeutic efficacy and patient prognosis. Metastasis is a
multistep process involving numerous factors. Cellular migration,
the attachment of cancer cells to the ECM components, and invasion
into surrounding tissues are critical to metastasis. Therefore,
decreased migration, cell-matrix adhesion, and invasive potential
may contribute to the prevention of metastasis. In the present
study, the effects of ITGB1 inhibition on apoptosis, migration,
invasion, and adhesion to ECM proteins were determined. The results
indicated that ITGB1 inhibition significantly increased cell
apoptosis, as determined by flow cytometry, and suppressed the
migration and invasion of ovarian cancer cells, as determined by
wound healing and transwell invasion assays. The cell adhesion
assay revealed that inhibition of ITGB1 attenuated the adhesion of
ovarian cancer cells to Matrigel™. These results indicated that
anti-migration, anti-invasion, and anti-adhesion functions may be
important contributors to the anti-metastatic activity of ITGB1
inhibition. MMPs are a well-known family of zinc-binding enzymes
that have been reported to be upregulated in cancer, and numerous
studies have demonstrated that overexpression of MMPs facilitates
cancer cell progression, suggesting that MMPs are also involved in
metastasis (31,32). In the present study, the inhibition
of ITGB1 suppressed MMP-2 and MMP-9 protein expression. These
results suggested that ITGB1 inhibition has the potential to
inhibit ovarian cancer metastasis by suppression of MMP-2 and MMP-9
expression. In conclusion, inhibition of ITGB1 resulted in tumor
cell apoptosis and disrupted tumor mass formation.
Previous studies demonstrated that ITGB1 may be
associated with therapeutic resistance to various agents and
ionizing radiation in the treatment of cancer (33–36).
Notably, ITGB1-mediated resistance is thought to occur at the level
of the tumor cells themselves. A recent study demonstrated that
ITGB1 inhibition combined with bevacizumab treatment reduced the
risk of resistance in glioblastoma (34). In the present study, ITGB1
inhibition enhanced the effects of bevacizumab on apoptosis,
adhesion, and migration of ovarian cancer cells. The results lead
to the hypothesis that ITGB1 inhibition combined with bevacizumab
treatment may reduce the required dose of the bevacizumab
anticancer agent, thus potentially reducing drug-related morbidity
in ovarian cancer.
It has been suggested that integrin/FAK has an
important role in regulating various cellular functions, including
adhesion, migration, invasion, survival, growth, and
differentiation (37). FAK
activates STAT1 in integrin-mediated cell migration and adhesion
(29). A previous study
demonstrated that FAK/STAT1 increased the malignant potential of
ovarian epithelium (28).
Therefore, ITGB1/FAK/STAT1 signaling is a promising therapeutic
target for ovarian cancer. In the present study the adherence and
migratory potentials of ovarian cancer cells were significantly
reduced following the inhibition of the ITGB1/FAK/STAT1 signaling
pathway by fludarabine. These results revealed that inhibiting
FAK/STAT1 signaling exerts anti-metastatic effects on ovarian
cancer cells.
These data suggested that ITGB1 inhibition
effectively reduced tumorigenesis and disease exacerbation, and
contributed to bevacizumab-induced anticancer therapy via the
FAK/STAT1 signaling pathway. This further elucidated the anti-tumor
molecular mechanisms underlying ITGB1 inhibition, suggesting that
targeting ITGB1 is a potential strategy for the prevention and
treatment of ovarian cancer metastasis.
References
|
1
|
Yang W, Han W, Ye S, Liu D, Wu J, Liu H,
Li C and Chen H: Fibroblast activation protein-α promotes ovarian
cancer cell proliferation and invasion via extracellular and
intracellular signaling mechanisms. Exp Mol Pathol. 95:105–110.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Heyman L, Kellouche S, Fernandes J, Dutoit
S, Poulain L and Carreiras F: Vitronectin and its receptors partly
mediate adhesion of ovarian cancer cells to peritoneal mesothelium
in vitro. Tumour Biol. 29:231–244. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gotzmann J, Mikula M, Eger A,
Schulte-Hermann R, Foisner R, Beug H and Mikulits W: Molecular
aspects of epithelial cell plasticity: Implications for local tumor
invasion and metastasis. Mutat Res. 566:9–20. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Foubert P and Varner JA: Integrins in
tumor angiogenesis and lymphangiogenesis. Methods Mol Biol.
757:471–486. 2012. View Article : Google Scholar
|
|
5
|
Garmy-Susini B and Varner JA: Roles of
integrins in tumor angiogenesis and lymphangiogenesis. Lymphat Res
Biol. 6:155–163. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tchaicha JH, Reyes SB, Shin J, Hossain MG,
Lang FF and McCarty JH: Glioblastoma angiogenesis and tumor cell
invasiveness are differentially regulated by β8 integrin. Cancer
Res. 71:6371–6381. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Buczek-Thomas JA, Chen N and Hasan T:
Integrin-mediated adhesion and signalling in ovarian cancer cells.
Cell Signal. 10:55–63. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lau MT, So WK and Leung PC: Integrin β1
mediates epithelial growth factor-induced invasion in human ovarian
cancer cells. Cancer Lett. 320:198–204. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tucker GC: Integrins: Molecular targets in
cancer therapy. Curr Oncol Rep. 8:96–103. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cordes N and Park CC: Beta1 integrin as a
molecular therapeutic target. Int J Radiat Biol. 83:753–760. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Desgrosellier JS and Cheresh DA: Integrins
in cancer: Biological implications and therapeutic opportunities.
Nat Rev Cancer. 10:9–22. 2010. View
Article : Google Scholar
|
|
12
|
Meads MB, Gatenby RA and Dalton WS:
Environment-mediated drug resistance: A major contributor to
minimal residual disease. Nat Rev Cancer. 9:665–674. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hazlehurst LA, Landowski TH and Dalton WS:
Role of the tumor microenvironment in mediating de novo resistance
to drugs and physiological mediators of cell death. Oncogene.
22:7396–7402. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Matsunaga T, Fukai F, Miura S, Nakane Y,
Owaki T, Kodama H, Tanaka M, Nagaya T, Takimoto R, Takayama T and
Niitsu Y: Combination therapy of an anticancer drug with the
FNIII14 peptide of fibronectin effectively overcomes cell
adhesion-mediated drug resistance of acute myelogenous leukemia.
Leukemia. 22:353–360. 2008. View Article : Google Scholar
|
|
15
|
Golubovskaya VM, Kweh FA and Cance WG:
Focal adhesion kinase and cancer. Histol Histopathol. 24:503–510.
2009.PubMed/NCBI
|
|
16
|
Lark AL, Livasy CA, Dressler L, Moore DT,
Millikan RC, Geradts J, Iacocca M, Cowan D, Little D, Craven RJ and
Cance W: High focal adhesion kinase expression in invasive breast
carcinomas is associated with an aggressive phenotype. Mod Pathol.
18:1289–1294. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ucar DA and Hochwald SN: FAK and
interacting proteins as therapeutic targets in pancreatic cancer.
Anticancer Agents Med Chem. 10:742–746. 2010. View Article : Google Scholar
|
|
18
|
Lechertier T and Hodivala-Dilke K: Focal
adhesion kinase and tumour angiogenesis. J Pathol. 226:404–412.
2012. View Article : Google Scholar
|
|
19
|
Mitra SK and Schlaepfer DD:
Integrin-regulated FAK-Src signaling in normal and cancer cells.
Curr Opin Cell Biol. 18:516–523. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jiang J, Grieb B, Thyagarajan A and Sliva
D: Ganoderic acids suppress growth and invasive behavior of breast
cancer cells by modulating AP-1 and NF-kappaB signaling. Int J Mol
Med. 21:577–584. 2008.PubMed/NCBI
|
|
21
|
Bird IM: Extraction of RNA from cells and
tissue. Methods Mol Med. 108:139–148. 2005.
|
|
22
|
Song J, Zhang J, Wang J, Cao Z, Wang J,
Guo X and Dong W: β1 integrin modulates tumor growth and apoptosis
of human colorectal cancer. Oncol Rep. 32:302–308. 2014.
|
|
23
|
Liu B, Che W, Xue J, Zheng C, Tang K,
Zhang J, Wen J and Xu Y: SIRT4 prevents hypoxia-induced apoptosis
in H9c2 cardiomyoblast cells. Cell Physiol Biochem. 32:655–662.
2013. View Article : Google Scholar
|
|
24
|
Liang CC, Park AY and Guan JL: In vitro
scratch assay: A convenient and inexpensive method for analysis of
cell migration in vitro. Nat Protoc. 2:329–333. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Monaco S, Gioia M, Rodriguez J,
Fasciglione GF, Di Pierro D, Lupidi G, Krippahl L, Marini S and
Coletta M: Modulation of the proteolytic activity of matrix
metalloproteinase-2 (gelatinase A) on fibrinogen. Biochem J.
402:503–513. 2007. View Article : Google Scholar :
|
|
26
|
Omanakuttan A, Nambiar J, Harris RM, Bose
C, Pandurangan N, Varghese RK, Kumar GB, Tainer JA, Banerji A,
Perry JJ, et al: Anacardic acid inhibits the catalytic activity of
matrix metalloproteinase-2 and matrix metalloproteinase-9. Mol
Pharmacol. 82:614–622. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tomao F, Papa A, Rossi L, Caruso D, Panici
PB, Venezia M and Tomao S: Current status of bevacizumab in
advanced ovarian cancer. Onco Targets Ther. 6:889–899.
2013.PubMed/NCBI
|
|
28
|
Zhang L, Wang D, Jiang W, Edwards D, Qiu
W, Barroilhet LM, Rho JH, Jin L, Seethappan V, Vitonis A, et al:
Activated networking of platelet activating factor receptor and
FAK/STAT1 induces malignant potential in BRCA1-mutant at-risk
ovarian epithelium. Reprod Biol Endocrinol. 8:742010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xie B, Zhao J, Kitagawa M, Durbin J, Madri
JA, Guan JL and Fu XY: Focal adhesion kinase activates Stat1 in
integrin-mediated cell migration and adhesion. J Biol Chem.
276:19512–19523. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Casey RC and Skubitz AP: CD44 and beta1
integrins mediate ovarian carcinoma cell migration toward
extracellular matrix proteins. Clin Exp Metastasis. 18:67–75. 2000.
View Article : Google Scholar
|
|
31
|
Mannello F, Tonti G and Papa S: Matrix
metalloproteinase inhibitors as anticancer therapeutics. Curr
Cancer Drug Targets. 5:285–298. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Radisky ES and Radisky DC: Matrix
metalloproteinase-induced epithelial-mesenchymal transition in
breast cancer. J Mammary Gland Biol Neoplasia. 15:201–212. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pontiggia O, Sampayo R, Raffo D, Motter A,
Xu R, Bissell MJ, Joffé EB and Simian M: The tumor microenvironment
modulates tamoxifen resistance in breast cancer: A role for soluble
stromal factors and fibronectin through β1 integrin. Breast Cancer
Res Treat. 133:459–471. 2012. View Article : Google Scholar
|
|
34
|
Carbonell WS, DeLay M, Jahangiri A, Park
CC and Aghi MK: β1 integrin targeting potentiates antiangiogenic
therapy and inhibits the growth of bevacizumab-resistant
glioblastoma. Cancer Res. 73:3145–3154. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kanda R, Kawahara A, Watari K, Murakami Y,
Sonoda K, Maeda M, Fujita H, Kage M, Uramoto H, Costa C, et al:
Erlotinib resistance in lung cancer cells mediated by integrin
β1/Src/Akt-driven bypass signaling. Cancer Res. 73:6243–6253. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ju L, Zhou C, Li W and Yan L: Integrin
beta1 over-expression associates with resistance to tyrosine kinase
inhibitor gefitinib in non-small cell lung cancer. J Cell Biochem.
111:1565–1574. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yamamoto N, Kinoshita T, Nohata N, Itesako
T, Yoshino H, Enokida H, Nakagawa M, Shozu M and Seki N: Tumor
suppressive microRNA-218 inhibits cancer cell migration and
invasion by targeting focal adhesion pathways in cervical squamous
cell carcinoma. Int J Oncol. 42:1523–1532. 2013.PubMed/NCBI
|