Introduction
Chronic peripheral nerve injury induces
morphological and phenotypic alterations in nerve fibers, leading
to hyperalgesia and allodynia (1,2). It
has been suggested that chronic nerve compression affects millions
of individuals (3). Pathological
changes have been found at the site of nerve compression after 1
month and, at 8 months, the damage has been observed to have
progressed throughout the nerve (4). However, the precise mechanism
underlying the pathogenesis of nerve compression injury remains to
be fully elucidated. Studies have shown that chronic sciatic nerve
compression triggers a series of pathological changes in the areas
close by, including breakdown of the blood nerve barrier,
sub-perineurial edema, fibrosis, and localized and diffuse
demyelination (3,5–8).
Details of the fibrotic process following nerve compression are
unclear, and phenotypic changes in the neurons of the dorsal root
ganglia (DRG) at the damaged site are accompanied by upregulated or
downregulated gene and protein expression levels (9,10).
In addition, neuronal inflammation of the DRG as a response to
chronic nerve compression has been noted (11).
Collagen fibers are the most important component of
the extracellular matrix, and the excessive production of collagen
type I may contribute to the pathogenesis of fibrosis (12). Transforming growth factor-β (TGF-β)
is reported to be prominent in sensory neurons, promote fibrosis
associated with pathological pain (13) and induce the expression of
connective tissue growth factor (CTGF) (14). It has been suggested that increased
CTGF in the painful disc is associated with disc fibrosis and
degeneration (15).
The mitogen-activated protein kinase (MAPK)
signaling pathway has been found to be activated in DRG neurons
during neuropathic pain in cell and rat models (16–18).
However, the involvement of the collagen type 1, TGF-β, CTGF and
MAPK signaling proteins in neuropathic pain remains to be fully
elucidated. The present study hypothesized that these factors and
mediators may support and contribute to the pathological changes
that are associated with chronic sciatic nerve compression,
particularly fibrosis, in DRG neurons and in the surrounding
tissues.
In the present study, a rat model of chronic sciatic
nerve compression was established to investigate the subsequent
signs of fibrosis, and the dynamics of the collagen type 1, TGF-β1,
CTGF and MAPK family members in the DRG neurons and surrounding
tissues. The results from the current study will provide further
understanding of the pathology of chronic sciatic nerve
compression-induced fibrosis in dorsal root ganglia.
Materials and methods
Reagents
Fluoro Ruby (FR) and Fluoro Gold (FG) were purchased
from Molecular Probes (Thermo Fisher Scientific, Inc., Waltham, MA,
USA) and Fluorochrome LLC (Denver, CO, USA), respectively. Silastic
tubes were purchased from Shanghai Daoguan Rubber Products
(Shanghai, China). Polymerase chain reaction (PCR) primers were
obtained from Invitrogen (Thermo Fisher Scientific, Inc). Rabbit
polyclonal anti-TGF-β1 (cat. no. sc-146) and rabbit polyclonal
anti-CTGF antibodies (cat. no. sc-25440) were obtained from Santa
Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). Rabbit monoclonal
anti-glyceraldehyde 3-phosphate dehydrogenase (GAPDH; cat. no.
2251-1) and rabbit polyclonal anti-collagen I (cat. no. ab34710)
antibodies were purchased from Epitomics, Inc. (Burlingham, CA,
USA) and Abcam (Cambridge, UK), respectively. Rabbit monoclonal
anti-phosphorylated (p-)ERK (cat. no. 4370) and rabbit monoclonal
anti-p-p38 (cat. no. 4511) antibodies were purchased from Cell
Signaling Technology, Inc. (Danvers, MA, USA). Rabbit polyclonal
anti-p-c-Jun N-terminal kinase (JNK; ab4821) antibody was purchased
from Abcam. Horseradish peroxidase (HRP)-conjugated goat
anti-rabbit secondary antibody (cat. no. 074-1506) was purchased
from KPL, Inc. (Gaithersburg, MD, USA). Secondary antibodies,
including AlexaFluor 488-conjugated goat anti-rabbit IgG (cat. no.
ANT030) and AlexaFluor 594-conjugated goat anti-rabbit IgG (cat.
no. ANT051) antibodies, were purchased from Antgene Biotech (Wuhan,
China).
Animals
A total of 100 male Sprague-Dawley rats (age, 7–8
weeks; weight, 250–300 g) were purchased from the Laboratory Animal
Center, Tongji Medical College, Huazhong University of Science and
Technology (Wuhan, China) and housed at 22°C with a 12 h light/dark
cycle. Each animal had ad libitum access to food and water.
All experimental procedures and protocols were approved by the
Board of Ethics of Tongji Medical College, Huazhong University of
Science and Technology.
Establishment of the chronic sciatic
nerve compression model
The chronic sciatic nerve compression model was
established in accordance with the method described by O'Brien
et al (19). The animals
were anesthetized using pentobarbital (40 mg/kg intraperitoneal
injection) and the right sciatic nerve of the rat was exposed
through a gluteal muscle splitting approach. In the model rat group
(n=8), a 1 cm-long Silastic tube (internal diameter 1 mm; outer
diameter 2 mm; Shanghai Daoguan Rubber Products Factory, Shanghai,
China) was cut longitudinally, and placed atraumatically around the
sciatic nerve. The longitudinal split in the tube was then closed
using 8–0 sutures (Shanghai Pudong Jinhuan Medical Product Co.,
Ltd., Shanghai, China). The left sciatic nerve was exposed, as
above, but remained unbanded and was returned to its original
position to serve as a comparative control. In the sham-operated
group (n=8), the right sciatic nerves were exposed, using the same
procedure as described above for the left sciatic nerve of the
model rats, but without Silastic tube placement.
Subsequently, 3 weeks following the above
procedures, the ipsilateral and contralateral L4–L6 dorsal root
ganglia (DRG) were harvested from the above experimental groups
(n=8 for each group). At 3 or 8 weeks after the establishment of
the animal model, rats were sacrificed using an overdose of
phenobarbital (Wuhan Boster Biological Technology, Ltd., Wuhan,
China), and the ipsilateral and contralateral gastrocnemius muscles
were collected from the above experimental groups (n=8 for each
group).
Retrograde tracing using FR or FG
FR and FG are fluorescent tracers used for the
retrograde labeling of afferent neurons in rodents (20). At 2 weeks post-surgery, the rats
were anesthetized. The right sciatic nerves were exposed again, and
the FR and FG fluorescent tracers were injected intraneurally. In
the rats in the chronic sciatic nerve compression model, 5
µl FR (5% dissolved in normal saline) was injected into the
proximal end of the compressed nerve, and 5 µl FG (5%
dissolved in normal saline) was injected into the distal nerve. In
the sham control rats, 5 µl 5% FR and 5 µl 5% FG were
injected into the same respective sites as in the model rats.
Following injection, the muscle and incision were closed.
Retrograde tracing analysis
At 1 week post-fluorescent tracer injection, the
ipsilateral DRG of L4, L5 and L6 were removed and fixed in 4%
paraformaldehyde (Wuhan Boster Biological Technology, Ltd.). The
DRGs were then dehydrated, embedded in optimal cutting temperature
compound (Applygen Technologies Inc., Beijing, China) and sectioned
coronally using a microtome (Leica CM1510S; Leica Microsystems,
Wetzlar, Germany). The fluorescence was examined under a
fluorescent microscope (Olympus IX71; Olympus Corporation, Tokyo
Japan), and the numbers of FR- and FG-positive neurons were
analyzed using ImagePro Plus version 6.0 (Media Cybernetics, Inc.,
Rockville, MD, USA).
Masson's trichrome staining of
gastrocnemius muscle
The ipsilateral and contralateral gastrocnemius
muscles were fixed and embedded in paraffin (Wuhan Boster
Biological Technology, Ltd.). The gastrocnemius muscles were
stained with Masson's trichrome (Harris hematoxylin, 1%
hydrochloric-alcohol solution, acid fuchsin, 1% phosphomolybdic
acid and 2% aniline blue). The morphological details of Masson's
trichrome staining were examined using a light microscope (CKX41;
Olympus Corporation). A total of six images, each from the
ipsilateral and contralateral gastrocnemius muscles, were randomly
selected to measure the average muscle fiber cross-sectional area
(CSA) and collagen volume fraction (CVF). The CVF was calculated as
follows: CVF (%) = (collagen area / total area) x 100 in muscle
fiber CSA.
Reverse transcription-quantitative PCR
(RT-qPCR)
At 3 weeks following the establishment of the
chronic sciatic nerve compression model, the contralateral and
ipsilateral L4–L6 DRG tissues were removed. Total RNA was extracted
from the homogenized tissue samples using TRIzol reagent,
chloroform and isopropyl alcohol (Wuhan Boster Biological
Technology, Ltd.). After centrifugation at 7,500 × g at 4°C for 5
min, samples were collected and dried at room temperature. The
total RNA was resolved in RNA-free water. The samples were
reverse-transcribed using a First Strand cDNA Synthesis kit (Toboyo
Co., Ltd., Osaka, Japan). The RT-qPCR was performed using a
sequence detection system (StepOne Real-Time PCR System; Thermo
Fisher Scientific, Inc.). PCR amplification was performed using
Thunderbird SYBR qPCR mix (12.5 µl; Toboyo Co., Ltd.) with 2
µl specific primers (2.5 µM; Table I), 2 µl cDNA and 8.5
µl double distilled water. The PCR reactions were as
follows: Pre-denaturation at 95°C for 1 min, denaturation at 95°C
for 30 sec, annealing at 58°C for 30 sec and extension at 72°C for
20 sec (for a total of 40 cycles). β-actin was used as the internal
control. The relative expression levels from the amplified RNA
samples were calculated using the 2−ΔΔCq method
(21).
 | Table ISequences of forward and reverse
primers used for reverse transcription-quantitative polymerase
chain reaction analysis. |
Table I
Sequences of forward and reverse
primers used for reverse transcription-quantitative polymerase
chain reaction analysis.
| Target gene | Primer
sequence |
|---|
| β-actin | F:
5′-CGTTGACATCCGTAAAGACCTC-3′
R: 5′-TAGGAGCCAGGGCAGTAATCT-3′ |
| TGF-β1 | F:
5′-TACTACGCCAAAGAAGTCACCC-3′
R: 5′-TCCCGAATGTCTGACGTATTGA-3′ |
| CTGF | F:
5′-CGGGAAATGCTGTGAGGAGT-3′
R: 5′-CAGTTGGCTCGCATCATAGTT-3′ |
| Col1agen I | F:
5′-CCTACAGCACGCTTGTGGATG-3′
R: 5′-AGATTGGGATGGAGGGAGTTTAC-3 |
Immunostaining
At 3 weeks following the establishment of the
chronic sciatic nerve compression model, the contra-lateral and
ipsilateral L4–L6 DRG tissues were removed, embedded in paraffin
and sectioned. The samples were blocked with normal goat serum
(Wuhan Boster Biological Technology, Ltd.) and probed with
anti-TGF-β1 (1:50 dilution), anti-CTGF (1:50 dilution) or
anti-collagen I (1:200 dilution) at 4°C overnight. The following
day, the slides were washed three times with phosphate-buffered
saline and incubated with secondary antibody at 4°C overnight, then
washed with Tris-buffered saline plus Tween-20 (TBS-T; 0.1%
Tween-20). Following washing, the nuclei were counterstained with
DAPI (100 µg/ml; Wuhan Boster Biological Technology, Ltd.),
and samples were mounted in antifade reagents (Beyotime Institute
of Biotechnology, Haimen, China). The fluorescence was examined
under a fluorescent microscope (Olympus IX71; Olympus
Corporation).
Protein extraction and western blot
analysis
At 3 weeks following establishment of the chronic
sciatic nerve compression model, the contralateral and ipsilateral
DRG tissues were removed. Total proteins were extracted from
tissues using radioimmunoprecipitation assay lysis buffer
containing 1% phenylmethanesulfonyl fluoride and phosphatase
inhibitor (Wuhan Boster Biological Technology, Ltd.), and lysed on
ice for 30 min. After centrifugation at 12,000 × g for 10 min at
4°C, the supernatant was collected and stored at −80°C until used.
The protein concentration was measured using a bicinchoninic acid
protein assay kit (Beyotime Institute of Biotechnology) in
accordance with the manufacturer's protocol. The samples were
resolved using either 10% SDS-PAGE (for CTGF, collagen I and GAPDH;
Wuhan Boster Biological Technology, Ltd.) or 15% (for TGF-β1)
SDS-PAGE, followed by transfer onto a polyvinylidene fluoride
membrane (eBioscience, Inc., San Diego, CA, USA), and blocking with
5% w/v non-fat dry milk. The membranes were then incubated with
primary antibodies at 4°C overnight as follows: Anti-TGF-β1 (1:500
dilution), anti-CTGF (1:1,000 dilution), anti-collagen I (1:5,000
dilution), anti-GAPDH (1:10,000 dilution), anti-p-ERK (1:1,000
dilution), anti-p-p38 (1:1,000 dilution) and anti-p-JNK (1:1,000
dilution). Following washing, the membranes were incubated with
HRP-labeled secondary antibodies (1:10,000 dilution) at 4°C
overnight, then washed with TBS-T (0.1% Tween-20).
The resulting immunobands were visualized using an
Enhanced Chemiluminescence kit (EMD Millipore, Billerica, MA, USA),
and the densitometric values were used for statistical analysis.
The housekeeping protein, GAPDH, was used as an internal
control.
Statistical analyses
The data were analyzed using SPSS 17.0 software
(SPSS, Inc., Chicago, IL, USA). Data are presented as the mean ±
standard deviation. P<0.05 was considered to indicate a
statistically significant difference. Statistical significance
between two groups was determined using Student's
t-test.
Results
Chronic sciatic nerve compression-induced
pathology
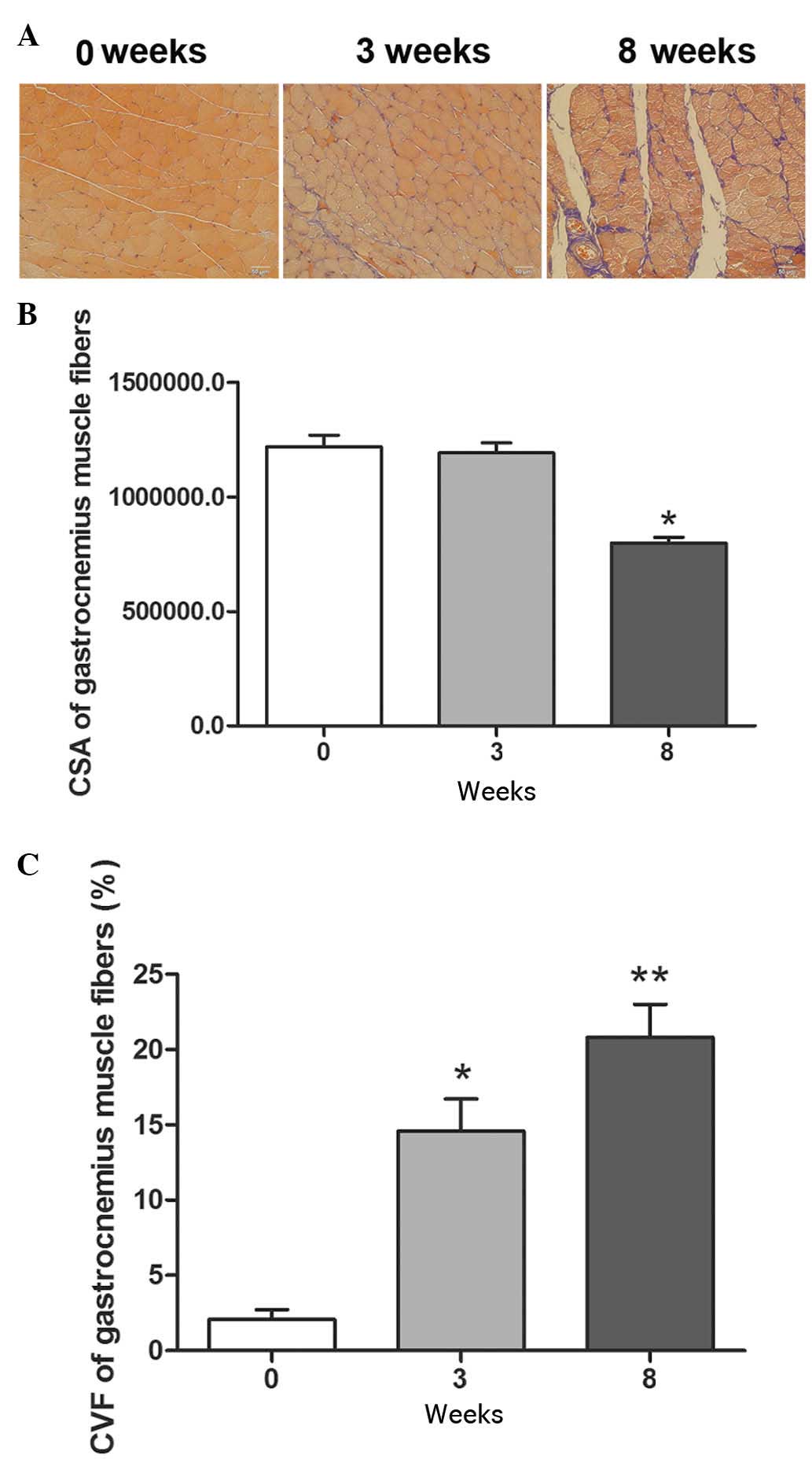
The present study examined the pathological changes
in the rat chronic sciatic nerve compression model. At 3 weeks and
8 weeks post-surgery, Masson's trichrome staining showed normal
gastrocnemius muscle fiber structure and collagen fiber morphology
on the contralateral side. However, at 3 weeks, collagen fiber
accumulation was detected on the ipsilateral side and, at 8 weeks,
excessive collagen fiber formation and muscle atrophy were observed
(Fig. 1A).
The chronic sciatic nerve compression in the model
group was associated with lower levels of gastrocnemius muscle
fiber CSA ag 3 weeks post-surgery, which was significantly lower at
8 weeks, compared with the contralateral side (Fig. 1B). In the model rats, the
percentage of gastrocnemius muscle fiber CVF was significantly
higher on the injured side, compared with the contralateral side at
3 weeks, and this difference was more marked at 8 weeks (Fig. 1C). In the subsequent experiments,
the molecular and cellular changes in the DRG neurons 3 weeks
following nerve compression.
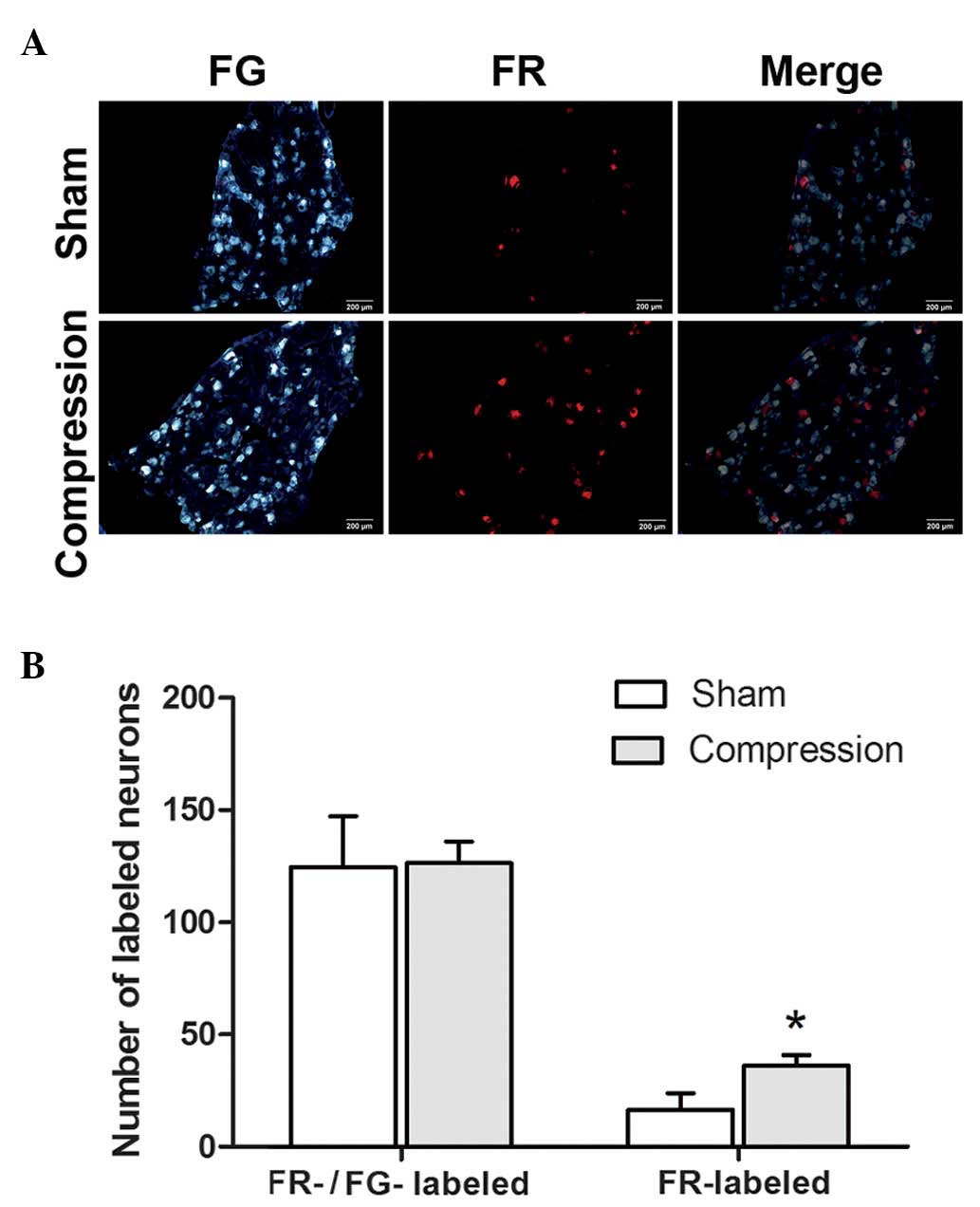
Retrograde fluorescent tracing of DRG
neurons
In the present study, the FR-labeled neurons
exhibited red fluorescence, predominantly in the cytoplasm, and the
FG-labeled neurons appeared bright blue under the fluorescent
microscope (Fig. 2). The majority
of the DRG neurons were stained with FG, and no significant
difference was detected in the number of FG-labeled neurons between
the sham control group and the group of rats with compression on
the injured side (P>0.05).
By contrast, the number of FR-labeled DRG neurons
was significantly higher in the rats with chronic sciatic nerve
compression, compared with the sham control group (P<0.05). Few
neurons were stained with both FR and FG. No significant
differences were observed in the total number of neurons labeled
with FR, FG or the two together, between the two groups
(P>0.05).
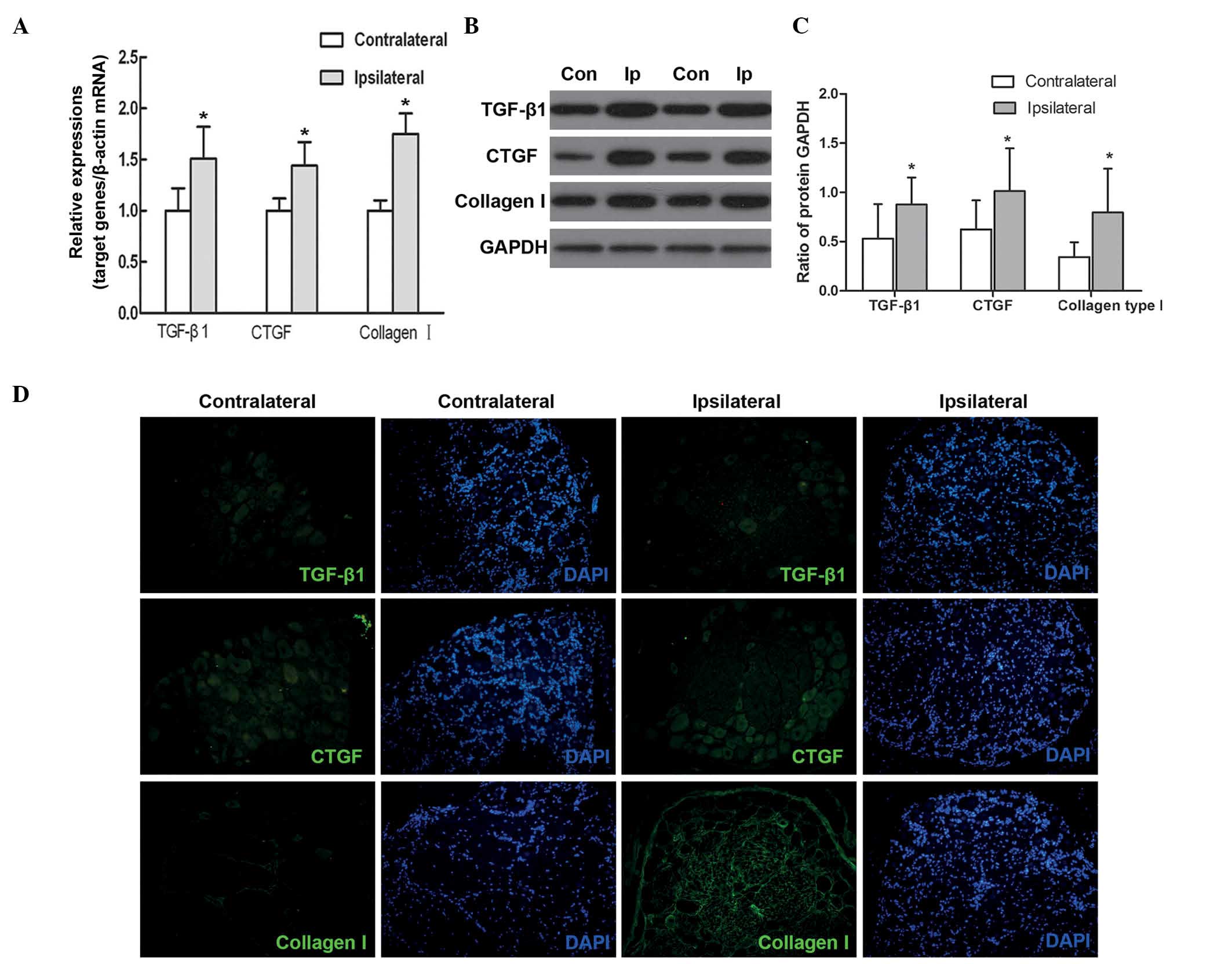
Expression of TGF-β1, CTGF and collagen
type I in injured DRG neurons and surrounding tissues
In the model rats, the present study found that the
mRNA expression levels of TGF-β1, CTGF and collagen type I were
significantly higher in the ipsilateral DRG neurons, compared with
the contralateral DRG neurons (P<0.05; Fig. 3A). Consistent with this, the
protein levels of TGF-β1, CTGF and collagen type I were also higher
in the ipsilateral DRG neurons and surrounding tissues, compared
with the those on the contralateral side (P<0.05; Fig. 3B and C).
Immunohistochemical analysis showed low levels of
expression of TGF-β1, CTGF and collagen type I in the
contra-lateral DRG neurons and surrounding tissues, whereas their
levels of expression were higher in the ipsilateral DRG neurons and
surrounding tissues (Fig. 3D).
Additionally, TGF-β1 and CTGF were predominantly distributed in the
cytoplasm and axons of the DRG neurons, and collagen type I was
expressed in the tissues surrounding the neurons.
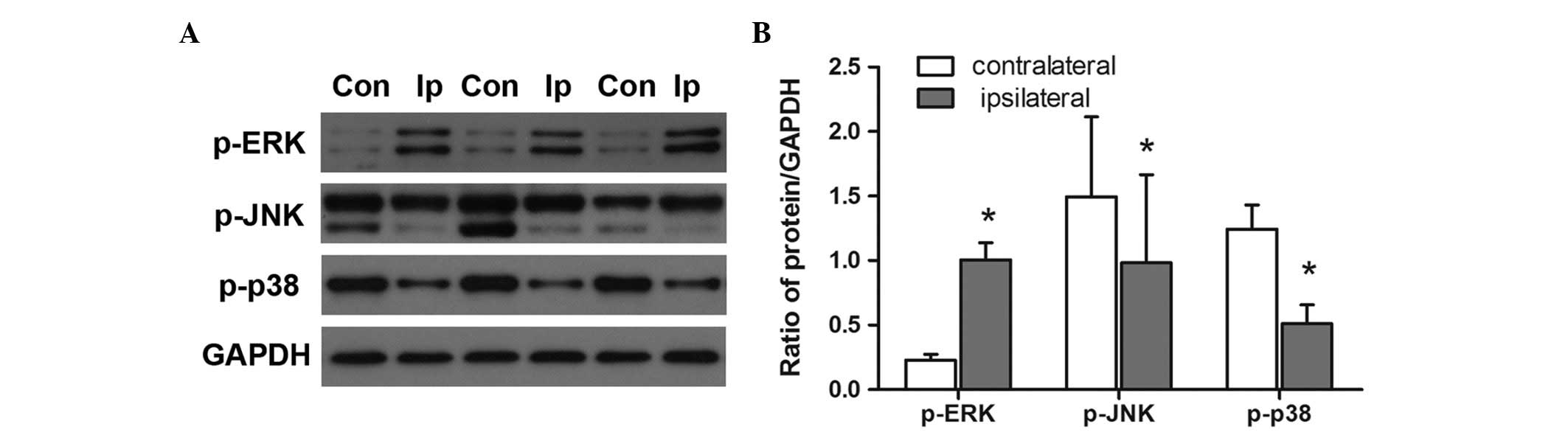
Involvement of the MAPK signaling pathway
in chronic sciatic nerve compression-induced DRG injury
Activation of the three major MAPK members, ERK1/2,
JNK and p38 MAPK is known to modulate the pathogenesis of fibrosis
in multiple organs (22–24). In the present study, immunoblotting
revealed that, in the model rats, the protein levels of p-ERK1/2
were higher in the ipsilateral DRGs following chronic nerve
compression, compared with the levels in the contralateral DRGs
(P<0.05; Fig. 4). By contrast,
chronic nerve compression was associated with significantly lower
levels of p-JNK and p-p38 in the DRGs (P<0.05).
Discussion
In the present study, a rat model was used to
investigate the changes associated with chronic sciatic nerve
compression in neurons of the DRG. The evidence suggested that
chronic sciatic nerve compression led to higher levels of TGF-β1,
CTGF and collagen type I, which may have been responsible for
inducing fibrosis in the tissues close to the lesions. Furthermore,
differences associated with chronic sciatic nerve compression in
the levels of the major MAPK members, ERK1/2, JNK and p38 MAPK,
suggested the involvement of the MAPK signaling pathway in the
pathogenesis of nerve compression injury.
Chronic sciatic nerve compression is characterized
by early damage to the blood-nerve barrier, which leads to swelling
and fibrosis, Schwann cell proliferation, local demyelination, and
Wallerian degeneration of the nerve (5). To understand the precise pathological
changes, as well as the molecular and cellular changes of DRG
neurons following chronic nerve compression, the present study
established a rat model of chronic sciatic nerve compression, as
previously described (19).
It has been demonstrated that demyelinated axons can
be differentially labeled with FR, and retrograde axonal transport
of this fluorescent tracer has also been shown, in which FR evident
in axonal fibers at the site of the compressed nerve can be used to
distinguish damaged neurons from healthy neurons (9). In accordance with this observation,
the present study detected a significantly higher number of
FR-labeled neurons in the model of nerve compression, compared with
the contralateral control. By contrast, the uptake of FG was
demonstrated in all neurons, whether injured or normal, which is
consistent with a previous report (25). In addition, moderate pathological
changes were observed 3 weeks following sciatic nerve compression,
and nerve injury and muscle atrophy were evident at 8 weeks.
A previous study examined injured nerves and
surrounding muscles using magnetic resonance imaging, and found
structural abnormalities (26). In
addition, enhanced FDG uptake was detected in these area using
(18F)-2-fluoro-2-deoxy-d-glucose positron emission tomography
(26). Therefore, the present
study investigated the molecular mechanism involved in chronic
sciatic nerve compression-induced DRG damage and muscle fibrosis 3
weeks following surgery.
Collagen type I is an important component of
mammalian extracellular matrix (27). During injury, fibroblasts migrate
into the damaged area, proliferate and synthesize extracellular
matrix components, particularly collagen type I (28). Collagen deposition is essential in
the regulation of Schwann cell function following nerve injury
(29), and the overproduction of
collagen type I may contribute to the pathogenesis of fibrosis
(12). In the present study,
higher levels of collagen type I were observed surrounding the
ipsilateral DRG, compared with the contralateral DRG. This
suggested that proliferated fibroblasts may generate collagen type
I, and that extracellular matrix accumulation and tissue fibrosis
may occur in damaged nerve tissues.
In pathological conditions, TGF-β1 and CTGF, which
are crucial mediators of tissue fibrosis, lead to the accumulation
of extracellular matrix (30).
TGF-β1 mediates tissue fibrosis by inducing the activation of CTGF
(30,31), which upregulates the expression of
CTGF by activating small mothers against decapentaplegic. The
activated CTGF subsequently mediates downstream effectors, which
collectively contribute to fibrosis (31,32).
CTGF is key in the development of tissue fibrosis by inducing
collagen type I in the surrounding tissues (33,34).
Reduced liver fibrosis has been observed in rats lacking CTGF
expression (35). Therefore,
TGF-β1 and CTGF are recognized as multifaceted regulatory proteins
in fibrosis (36). In the present
study, the levels of TGF-β1 and CTGF were higher in ipsilateral DRG
neurons, compared with contralateral DRG neurons. It is possible
that chronic sciatic nerve compression triggered the generation of
TGF-β1, and the consequent activation and release of CTGF in the
DRG neurons; and this increased expression of TGF-β1 and CTGF led
to the production of excessive collagen type I in the surrounding
tissues, which finally contributed to fibrosis. It is also possible
that the TGF-β1 released from DRG neurons (37) had a direct connection with the
paracrine CTGF signal. The cysteine-rich domain of CTGF directly
binds to and induces the activation of TGF-β1 (38), and activated TGF-β1 binds to its
receptor on fibroblasts, glia or Schwann cells to induce the
generation of collagen type I (39,40).
However, the precise mechanism requires further clarification.
The ERK1/2, JNK and p38 MAPK pathway transduction
signals are known to be involved in various intracellular signaling
pathways, which control a wide spectrum of cellular processes,
including fibrosis (41,42). The activation of MAPK signals,
mediated by TGF-β1, contribute to the deposition of extracellular
matrix and fibrosis (30). In
addition, activation of the ERK signaling molecule has been shown
to accompany fibroblast activation and collagen synthesis (43). In the present study, ERK1/2 was
activated in the ipsilateral DRG neurons, indicated by higher
levels of p-ERK1/2. Consistent with the observations in the present
study of increased activation of ERK1/2 in compressed DRGs, it has
been reported that p-ERK1/2 induces neuropathic pain in DRG
neurons, and gabapentin has an analgesic effect by inhibiting the
activation of ERK1/2 activation (44). However, the effects of the
activation of ERK1/2 and the downstream effectors of ERK1/2 remain
to be elucidated. Although the activation of JNK and p38 has been
detected in the development of neuropathic pain in cultured DRG
neurons (45), chronic nerve
compression in the present study was associated with significantly
lower levels of p-JNK and p-p38 in the DRGs. This discrepancy may
be due to differences between in vitro and in vivo
conditions. The evidence suggested that MAPK members may be
involved in chronic sciatic nerve compression-induced DRG damage,
and future investigations aim to investigate the underlying
mechanism involved.
In conclusion, the present study demonstrated that
chronic sciatic nerve compression induced DRG impairment. It is
possible that increased expression levels of TGF-β1 and CTGF in the
DRG neurons, and elevated expression of collagen type I in the
surrounding tissues, may contribute to the fibrosis induced by
chronic sciatic nerve compression.
Acknowledgments
This study was supported by the National Natural
Science Foundation of China (grant nos. 81271967, 81471270 and
81252899).
References
|
1
|
Pelletier J, Fromy B, Morel G, Roquelaure
Y, Saumet JL and Sigaudo-Roussel D: Chronic sciatic nerve injury
impairs the local cutaneous neurovascular interaction in rats.
Pain. 153:149–157. 2012. View Article : Google Scholar
|
|
2
|
Berger JV, Knaepen L, Janssen SP, Jaken
RJ, Marcus MA, Joosten EA and Deumens R: Cellular and molecular
insights into neuropathy-induced pain hypersensitivity for
mechanism-based treatment approaches. Brain Res Rev. 67:282–310.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pham K and Gupta R: Understanding the
mechanisms of entrapment neuropathies. Review article. Neurosurg
Focus. 26:E72009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gupta R, Rowshan K, Chao T, Mozaffar T and
Steward O: Chronic nerve compression induces local demyelination
and remyelination in a rat model of carpal tunnel syndrome. Exp
Neurol. 187:500–508. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mackinnon SE: Pathophysiology of nerve
compression. Hand Clin. 18:231–241. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mackinnon SE, Dellon AL, Hudson AR and
Hunder DA: Chronic human nerve compression - a histological
assessment. Neuropathol Appl Neurobiol. 12:547–565. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Prinz RA, Nakamura-Pereira M, De-Ary-Pires
B, Fernandes D, Fabião-Gomes BD, Martinez AM, de Ary-Pires R and
Pires-Neto MA: Axonal and extracellular matrix responses to
experimental chronic nerve entrapment. Brain Res. 1044:164–175.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pham Khoa, Nassiri Nima and Gupta Ranjan:
c-Jun, krox-20, and integrin β4 expression following chronic nerve
compression injury. Neurosci Lett. 465:194–198. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chao T, Pham K, Steward O and Gupta R:
Chronic nerve compression injury induces a phenotypic switch of
neurons within the dorsal root ganglia. J Comp Neurol. 506:180–193.
2008. View Article : Google Scholar
|
|
10
|
Zhang Y, Wang YH, Zhang XH, Ge HY,
Arendt-Nielsen L, Shao JM and Yue SW: Proteomic analysis of
differential proteins related to the neuropathic pain and
neuroprotection in the dorsal root ganglion following its chronic
compression in rats. Exp Brain Res. 189:199–209. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dubovy P, Brazda V, Klusakova I and
Hradilova-Svizenska I: Bilateral elevation of interleukin-6 protein
and mRNA in both lumbar and cervical dorsal root ganglia following
unilateral chronic compression injury of the sciatic nerve. J
Neuroinflammation. 10:552013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ponticos M, Papaioannou I, Xu S, et al:
The failure to degrade JunB contributes to Collagen type I
over-production and dermal fibrosis in Scleroderma. Arthritis
Rheumatol. 1:242–253. 2014.
|
|
13
|
Zhu Y, Colak T, Shenoy M, Liu L, Mehta K,
Pai R, Zou B, Xie XS and Pasricha PJ: Transforming growth factor
beta induces sensory neuronal hyperexcitability and contributes to
pancreatic pain and hyperalgesia in rats with chronic pancreatitis.
Mol Pain. 8:652012. View Article : Google Scholar
|
|
14
|
Fujii M, Nakanishi H, Toyoda T, Tanaka I,
Kondo Y, Osada H and Sekido Y: Convergent signaling in the
regulation of connective tissue growth factor in malignant
mesothelioma: TGF β signaling and defects in the Hippo signaling
cascade. Cell Cycle. 11:3373–3379. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Peng B, Chen J, Kuang Z, Li D, Pang X and
Zhang X: Expression and role of connective tissue growth factor in
painful disc fibrosis and degeneration. Spine (Phila Pa 1976).
34:E178–E182. 2009. View Article : Google Scholar
|
|
16
|
Yang Y, Wu H, Yan JQ, Song ZB and Guo QL:
Tumor necrosis factor-α inhibits angiotensin II receptor type 1
expression in dorsal root ganglion neurons via β-catenin signaling.
Neuroscience. 248:383–391. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Muralidharan A, Wyse BD and Smith MT:
Analgesic efficacy and mode of action of a selective small molecule
angiotensin II type 2 receptor antagonist in a rat model of
prostate cancer-induced bone pain. Pain Med. 15:93–110. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Obata K and Noguchi K: MAPK activation in
nociceptive neurons and pain hypersensitivity. Life Sci.
74:2643–2653. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
O'Brien JP, Mackinnon SE, MacLean AR,
Hudson AR, Dellon AL and Hunter DA: A model of chronic nerve
compression in the rat. Ann Plast Surg. 19:430–435. 1987.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zele T, Sketelj J and Bajrovic FF:
Efficacy of fluorescent tracers in retrograde labeling of cutaneous
afferent neurons in the rat. J Neurosci Methods. 191:208–214. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCt method. Methods. 25:402–408. 2001. View Article : Google Scholar
|
|
22
|
Madala SK, Schmidt S, Davidson C, Ikegami
M, Wert S and Hardie WD: MEK-ERK pathway modulation ameliorates
pulmonary fibrosis associated with epidermal growth factor receptor
activation. Am J Respir Cell Mol Biol. 46:380–388. 2012. View Article : Google Scholar :
|
|
23
|
Gao X, Wu G, Gu X, Fu L and Mei C:
Kruppel-like factor 15 modulates renal interstitial fibrosis by
ERK/MAPK and JNK/MAPK pathways regulation. Kidney Blood Press Res.
37:631–640. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ma FY, Tesch GH and Nikolic-Paterson DJ:
ASK1/p38 signaling in renal tubular epithelial cells promotes renal
fibrosis in the mouse obstructed kidney. Am J Physiol Renal
Physiol. 307:F1263–F1273. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kawarai Y, Suzuki M, Yoshino K, Inoue G,
Orita S, Yamauchi K, Aoki Y, Ishikawa T, Miyagi M, Kamoda H, et al:
Transient receptor potential vanilloid 1-immunoreactive innervation
increases in fractured rat femur. Yonsei Med J. 55:185–190. 2014.
View Article : Google Scholar :
|
|
26
|
Lasko L, Huang X, Voorbach MJ, Lewis LG,
Stavropoulos J, Carriker J, Seifert TR, Baker SJ and Upadhyay J:
Multimodal assessment of nervous and immune system responses
following sciatic nerve injury. Pain. 154:2782–2793. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jones CA, Liang L, Lin D, Jiao Y and Sun
B: The spatial-temporal characteristics of type I collagen-based
extracellular matrix. Soft Matter. 10:8855–8863. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cheret J, Lebonvallet N, Buhe V, Carre JL,
Misery L and Le Gall-Ianotto C: Influence of sensory neuropeptides
on human cutaneous wound healing process. J Dermatol Sci.
74:193–203. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Koopmans G, Hasse B and Sinis N: Chapter
19: The role of collagen in peripheral nerve repair. Int Rev
Neurobiol. 87:363–379. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ihn H: Pathogenesis of fibrosis: Role of
TGF-beta and CTGF. Curr Opin Rheumatol. 14:681–685. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Leask A, Holmes A, Black CM and Abraham
DJ: Connective tissue growth factor gene regulation. Requirements
for its induction by transforming growth factor-beta 2 in
fibroblasts. J Biol Chem. 278:13008–13015. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Igarashi A, Okochi H, Bradham DM and
Grotendorst GR: Regulation of connective tissue growth factor gene
expression in human skin fibroblasts and during wound repair. Mol
Biol Cell. 4:637–645. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Boerma M, Wang J, Sridharan V, Herbert JM
and Hauer-Jensen M: Pharmacological induction of transforming
growth factor-beta1 in rat models enhances radiation injury in the
intestine and the heart. PLoS One. 8:e704792013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sonnylal S, Xu S, Jones H, Tam A, Sreeram
VR, Ponticos M, Norman J, Agrawal P, Abraham D and De Crombrugghe
B: Connective tissue growth factor causes EMT-like cell fate
changes in vivo and in vitro. J Cell Sci. 126:2164–2175. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
George J and Tsutsumi M: siRNA-mediated
knockdown of connective tissue growth factor prevents
N-nitrosodimethylami ne-induced hepatic fibrosis in rats. Gene
Ther. 14:790–803. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gressner OA and Gressner AM: Connective
tissue growth factor: A fibrogenic master switch in fibrotic liver
diseases. Liver Int. 28:1065–1079. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Stark B, Carlstedt T and Risling M:
Distribution of TGF-beta, the TGF-beta type I receptor and the R-II
receptor in peripheral nerves and mechanoreceptors; observations on
changes after traumatic injury. Brain Res. 913:47–56. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Abreu JG, Ketpura NI, Reversade B and De
Robertis EM: Connective-tissue growth factor (CTGF) modulates cell
signalling by BMP and TGF-beta. Nat Cell Biol. 4:599–604.
2002.PubMed/NCBI
|
|
39
|
Liu Y, Liu Z, Liu X, Luo B, Xiong J, Gan
W, Jiang M and Zhang Z, Schluesener HJ and Zhang Z: Accumulation of
connective tissue growth factor+ cells during the early phase of
rat traumatic brain injury. Diagn Pathol. 9:1412014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Petito RB, Amadeu TP, Pascarelli BM,
Jardim MR, Vital RT, Antunes SL and Sarno EN: Transforming growth
factor-β1 may be a key mediator of the fibrogenic properties of
neural cells in leprosy. J Neuropathol Exp Neurol. 72:351–366.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Peti W and Page R: Molecular basis of MAP
kinase regulation. Protein Sci. 22:1698–1710. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Okada Y, Shirai K, Reinach PS,
Kitano-Izutani A, Miyajima M, Flanders KC, Jester JV, Tominaga M
and Saika S: TRPA1 is required for TGF-β signaling and its loss
blocks inflammatory fibrosis in mouse corneal stroma. Lab Invest.
94:1030–1041. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhang YP, Wang WL, Liu J, Li WB, Bai LL,
Yuan YD and Song SX: Plasminogen activator inhibitor-1 promotes the
proliferation and inhibits the apoptosis of pulmonary fibroblasts
by Ca (2+) signaling. Thromb Res. 131:64–71. 2013. View Article : Google Scholar
|
|
44
|
Zhang JL, Yang JP, Zhang JR, Li RQ, Wang
J, Jan JJ and Zhuang Q: Gabapentin reduces allodynia and
hyperalgesia in painful diabetic neuropathy rats by decreasing
expression level of Nav1.7 and p-ERK1/2 in DRG neurons. Brain Res.
1493:13–18. 2013. View Article : Google Scholar
|
|
45
|
Zang Y, Xin WJ, Pang RP, Li YY and Liu XG:
Upregulation of Nav1.3 Channel Induced by rrTNF in Cultured Adult
Rat DRG Neurons via p38 MAPK and JNK Pathways. Chin J Physiol.
54:241–246. 2011. View Article : Google Scholar : PubMed/NCBI
|