Introduction
Pulmonary hypertension (PH) is a complex and
multifactorial disorder characterized by increased pulmonary
arterial pressure and adverse remodeling of pulmonary arteries
(PAs), which result in right ventricular failure and premature
mortality (1). Excessive
proliferation and resistance to apoptosis in pulmonary artery
smooth muscle cells (PASMCs) is widely accepted as one of the
predominant causes of PA remodeling during hypoxia (2,3).
Hypoxia suppresses mitochondria-dependent apoptosis in human PASMCs
(HPASMCs) and increases proliferation in these cells (4,5).
However, the potential mechanism underlying the proliferation and
resistance to apoptosis in hypoxic PASMCs remains to be elucidated,
and there remains no curative therapeutic strategy. The overall
prognosis of patients with PH remains poor, and further
understanding of the mechanisms underlying the development of PH is
important to improve patient management and outcomes (6).
Several predisposing and disease-modifying
abnormalities, including the de novo expression of survivin
and the downregulated expression of the voltage-dependent
K+ (Kv)1.5 channel, have been reported to
contribute to the cancer-like, proliferative, apoptosis-resistant
phenotype of PASMCs (7).
Kv channels in PASMCs are inhibited by acute and chronic
exposure to hypoxia (8). Survivin
is a member of the inhibitor of apoptosis (IAP) protein gene
family, which negatively regulates programmed cell death and is
well documented to be overexpressed in almost all types of human
cancer (9). Additional data has
indicated a more selective role of survivin, also a chromosomal
passenger protein required for cell division (10), in antagonizing
mitochondria-dependent apop-tosis (11). Survivin expression is cell
cycle-dependent but it is also regulated by exposure to hypoxia
(12). It is almost undetectable
in the majority of normal adult tissues, and increased expression
of survivin correlates with a poor outcome (13). A previous study by McMurtry et
al (14) indicated that
survivin was expressed in the PAs of patients with PH, and that the
overexpression of survivin coincided with pulmonary vascular
remodeling in monocrotaline-induced rat PAH models. In addition,
the therapeutic effect of inhibition of survivin was achieved by
the induction of mitochondria-dependent apop-tosis and the
activation of Kv channels in PASMCs (14). These findings suggested that
inducing the expression of survivin may contribute to the abnormal
PASMC phenotype observed in PH; therefore, survivin may be an
attractive target for PH therapy.
As a novel small-molecule survivin inhibitor,
sepantronium bromide (YM155) suppresses the transactivation of
survivin via direct binding to its promoter (15) and, therefore, has little effect on
the expression levels of other IAP family members or B-cell
lymphoma 2-associated proteins (16). It has been demonstrated that YM155
induces tumor cell apoptosis and survivin suppression in various
human cancer models (16,17). A previous study by Liu et al
(18) demonstrated that survivin
was expressed in the PAs of rats with chronic hypoxic pulmonary
hypertension, but not in the PAs of normal rats. YM155 treatment
downregulated the expression levels of survivin in the distal PAs
and lung tissues of the rats exposed to chronic hypoxia, and
reduced mean pulmonary arterial pressure and right ventricular
hypertrophy, subsequently reversing hypoxia-induced PH. These
results suggested that YM155 may be a potential therapeutic agent
for hypoxic PH. However, no previous studies, to the best of our
knowledge, have evaluated the effects of YM155 on the expression of
survivin and apoptosis of HPASMCs exposed to hypoxia, or the
potential underlying mechanisms. The present study hypothesized
that YM155 may have anti-proliferative effects on hypoxia-induced
HP. Therefore, the protective effect of YM155 on hypoxic HPASMCs
was investigated, with a focus on the mechanisms of cell
proliferation and apoptosis, as well as the activation of
Kv1.5 and Kv2.1 channel in the PASMCs during
hypoxia.
Materials and methods
Cell culture
Human pulmonary artery smooth muscle cells (HPASMCs)
were purchased from ScienCell Research Laboratories (Carlsbad, CA,
USA) and cultured in smooth muscle cell medium (ScienCell Research
Laboratories) supplemented with 2% fetal bovine serum (Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA), 100 U/ml
penicillin, 100 µg/ml streptomycin and 1% smooth muscle cell
growth supplement (all ScienCell Research Laboratories) under an
atmosphere of 5% CO2 at 37°C. The HPASMCs at passages
4–10 were used in the following experimental assays.
Experimental protocol
The cultured HPASMCs were randomly divided into six
groups: i) Normoxia (N) group, in which the cells were cultured in
serum-free Dulbecco's modified Eagle's medium (DMEM; Beijing
Solarbio Science and Technology Co., Ltd., Beijing, China) under
normoxic conditions for 24 h at 37°C; ii) normoxia + 100 nmol/l
YM155 (NY100) group, in which the cells were cultured in serum-free
DMEM with 100 nmol/l YM155 under normoxic conditions for 24 h at
37°C; iii) hypoxia (H) group, in which the cells were cultured in
serum-free DMEM under hypoxic conditions for 24 h at 37°C; iv)
hypoxia + 1 nmol/l YM155 (HY1) group, in which the cells were
cultured in serum-free DMEM with 1 nmol/l YM155 under hypoxic
conditions for 24 h at 37°C; v) hypoxia + 10 nM YM155 (HY10) group,
in the which cells were cultured in serum-free DMEM with 10 nmol/l
YM155 under hypoxic conditions for 24 h; and vi) hypoxia + 100
nmol/l YM155 (HY100) group, in which the cells were cultured in
serum-free DMEM with 100 nmol/l YM155 under hypoxic conditions for
24 h at 37°C. YM155 was obtained from Selleck Chemicals (Houston,
TX, USA) and dissolved in dimethyl sulfoxide (Beijing Solarbio
Science and Technology, Co., Ltd.). The final concentration of DMEM
was <0.1% in medium.
The HPASMCs were exposed to the different
conditions, according to the group. For all experiments, the cells
were rendered quiescent by incubation in serum-free DMEM for 24 h
at 37°C prior to incubation in either hypoxic (2.5% O2,
5% CO2) or normoxic (21% O2, 5%
CO2) conditions. Hypoxic culture conditions, defined as
2.5% O2, were established using a Heraeus®
oxygen-regulated cell culture incubator (Thermo Fisher Scientific,
Inc.).
Cell viability
Cell viability was determined using a Cell Counting
Kit-8 (CCK-8) assay (Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan) in accordance with the manufacturer's protocol.
The HPASMCs were seeded in 96-well plates at 1×104
cells/well (four wells for each group) and cultured for 24 h in
complete medium in an atmosphere of 5% CO2 at 37°C. The
cells were treated, as described above, and 10 µl/well CCK-8
was added to each well at the end of the experiment and incubated
at 37°C for a further 2 h. The absorbance of each well was measured
at 450 nm on a microplate reader (HR801; Rayto Life and Analytical
Sciences Co., Ltd., Shenzhen, China).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the HPASMCs using
TRIzol (Tiangen Biotech Co., Ltd., Beijing, China), according to
the manufacturer's protocol, and the RNA was treated with
RNase-Free DNase I (Tiangen Biotech Co., Ltd.) to eliminate
contaminating DNA. The RNA concentration (552 ng/µl) was
determined using an ultra-violet spectrophotometer (NanoDrop 2000,
Thermo Fisher Scientific, Inc.), and the quality of RNA was checked
by 1.5% agarose gel electrophoresis (Sangon Biotech Co., Ltd.,
Shanghai, China). cDNA was synthesized from 10 µl total RNA
using the QuantScript RT kit (KR106; Tiangen Biotech Co., Ltd.).
qPCR was then performed on the ABI 7500 Real-Time PCR System
(Applied Biosystems; Thermo Fisher Scientific, Inc.). The reaction
mixture (25 µl) consisted of 6 µl cDNA, 12.5
µl 2X FastFire qPCR PreMix (SYBR Green; Tiangen Biotech Co.,
Ltd.) and 0.75 µl each of forward and reverse primers. The
PCR cycling conditions were as follows: 94°C for 5 min, followed by
45 cycles at 94°C for 40 sec, 58°C for 40 sec and 72°C for 40 sec.
The primers were obtained from Sunny Biotech Co., Ltd. (Shanghai,
China) and their sequences are presented in Table I. Melting curve analysis was
performed to examine the specificity of the amplification
reactions. GAPDH served as an internal control. Experiments were
performed in triplicate. The relative gene expression levels were
calculated for each sample using the 2−ΔΔCq method
(19).
 | Table ISequences of primers used for reverse
transcription-polymerase chain reaction analysis. |
Table I
Sequences of primers used for reverse
transcription-polymerase chain reaction analysis.
| Gene | Primer (5′→3′) |
|---|
| Survivin | Forward:
ACTTGGCCCAGTGGGTTTTT |
| Reverse:
CAGAAAGGAAAGCGCAACCG |
|
Kv1.5 | Forward:
CTACTTCGACCCCCTGAGGA |
| Reverse:
CAGGGTCTCCAAGCAGAAGG |
|
Kv2.1 | Forward:
TTTGCCCGGAGCATTGAGAT |
| Reverse:
GAACGTTCAGGTGCTGAGGA |
| GAPDH | Forward:
CACCATCTTCCAGGAGCGAG |
| Reverse:
AAATGAGCCCCAGCCTTCTC |
Immunocytochemical analysis
The HPASMCs were seeded onto 96-well plates at a
density ~1×105 cells/ml for immunocytochemical analysis.
Following washing with phosphate-buffered saline (PBS; Pujingkangli
Science & Technology, Beijing, China), the HPASMCs were fixed
with 4% paraformaldehyde solution (Beijing Huayueyang Biotechnology
Co., Ltd., Beijing, China) and permeabilized with 0.2% Triton X-100
(Sigma-Aldrich, St. Louis, MO, USA) in PBS. The cells were then
incubated with goat anti-human survivin polyclonal antibody
(ab27468; Abcam, Cambridge, UK) overnight at 4°C. After washing
three times with PBS for 5 min each, the cells were incubated with
tetraethyl rhodamine isothiocyanate-conjugated rabbit anti-goat IgG
(heavy and light chains) secondary antibody (31650; Thermo Fisher
Scientific, Inc.) at room temperature for 30 min. The cells were
observed under a fluorescent microscope (Axio Scope.A1; Carl Zeiss
AG, Oberkochen, Germany), and the images were acquired and analyzed
using Image-Pro Plus software, version 6.0 (Media Cybernetics,
Inc., Rockville, MD, USA).
Western blot analysis
The HPASMCs were placed in 200 µl
radioimmunoprecipitation assay lysis buffer (Tiangen Biotech Co.,
Ltd.) supplemented with 1% protease inhibitor cocktail
(Pujingkangli Science & Technology). Following centrifugation
at 12,000 × g at 4°C for 10 min, the supernatants were collected
for Western blot analysis. The amount of protein was quantified
using a Bio-Rad Protein Assay kit (500-0116; Bio-Rad Laboratories,
Inc., Hercules, CA, USA), according to the manufacturer's protocol.
Protein samples (40 µg) were run on a 10% acrylamide gel
(Huaxing Biotechnology Co., Ltd., Beijing, China) and transferred
onto a nitrocellulose membrane (Merck Millipore, Darmstadt,
Germany). The membranes were blocked with 5% skimmed milk powder in
Tris-buffered saline and 0.1% Tween 20 (TBST; Huaxing Biotechnology
Co., Ltd.) for 1 h at room temperature, followed by washing three
times with TBST for 10 min. Subsequently, the membranes were
immunoblotted overnight at 4°C with rabbit anti-human survivin
polyclonal antibody (ab469; 1:1,000; Abcam, Cambridge, UK), rabbit
anti-human Kv1.5 polyclonal antibody (ab181798; 1:1,000;
Abcam), mouse anti-human Kv2.1 monoclonal antibody
(ab105586; 1:1,000; Abcam) or rabbit anti-human GAPDH monoclonal
antibody (14C10; 1:2,000; Cell Signaling Technology, Inc., Danvers,
MA, USA). The membranes were washed three times with PBS and then
incubated with horseradish peroxidase-conjugated goat anti-rabbit
or goat anti-mouse secondary antibodies (ZB-2305 and ZB-2301;
1:5,000; Beijing Zhongshan Golden Bridge Biotechnology Co., Ltd.).
Detection was performed using an enhanced chemiluminescence kit
(ZLI-9036; Beijing Zhongshan Golden Bridge Biotechnology Co., Ltd.,
Beijing, China). Proteins bands were visualized following exposure
of the membrane to X-ray film (Kodak, Rochester, NY, USA). The
protein expression levels of survivin, Kv1.5 and
Kv2.1 were normalized to the protein expression of
GAPDH.
Terminal deoxynucleotidyl
transferase-mediated deoxyuridine triphosphate nick end labeling
(TUNEL) assay
Following hypoxia and/or pretreatment, the HPASMCs
were fixed with 4% paraformaldehyde in PBS for 1 h at room
temperature. The TUNEL assay was performed for apoptotic cell
determination using an In situ Cell Death Detection kit
(Roche Diagnostics GmbH, Mannheim, Germany), according to the
manufacturer's protocol. Counterstaining of nuclei with DAPI
(Thermo Fisher Scientific, Inc.) was performed for 10 min at 20°C,
and sealed with nail varnish. All TUNEL-positive cells (indicated
by green fluorescence in the nuclei) were counted in each field of
view, and subsequently expressed as a percentage of the total
number of nuclei in that same field.
Statistical analysis
Data are presented as the mean ± standard deviation
and were analyzed using the SPSS 17.0 statistical software package
(SPSS, Inc, Chicago, IL. USA). One-way analysis of variance was
used for comparison of variance among groups. Comparison between
groups was analyzed via Student-Newman-Keuls q test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
YM155 inhibits the expression of survivin
in hypoxic HPASMCs
RT-qPCR, immunocytochemistry and Western blot
analyses were performed to determine the expression levels of
survivin in hypoxic or normoxic HPASMCs with or without YM155
treatment. As shown in Fig. 1A,
survivin was expressed in the HPASMCs cultured under hypoxic
conditions, but was not expressed in the normoxia-cultured cells.
Treatment with YM155 significantly inhibited the hypoxia-induced
upregulated gene and protein expression levels of survivin, and
this effect was enhanced by increasing the concentration of YM155
(P<0.05; Fig. 1B and C).
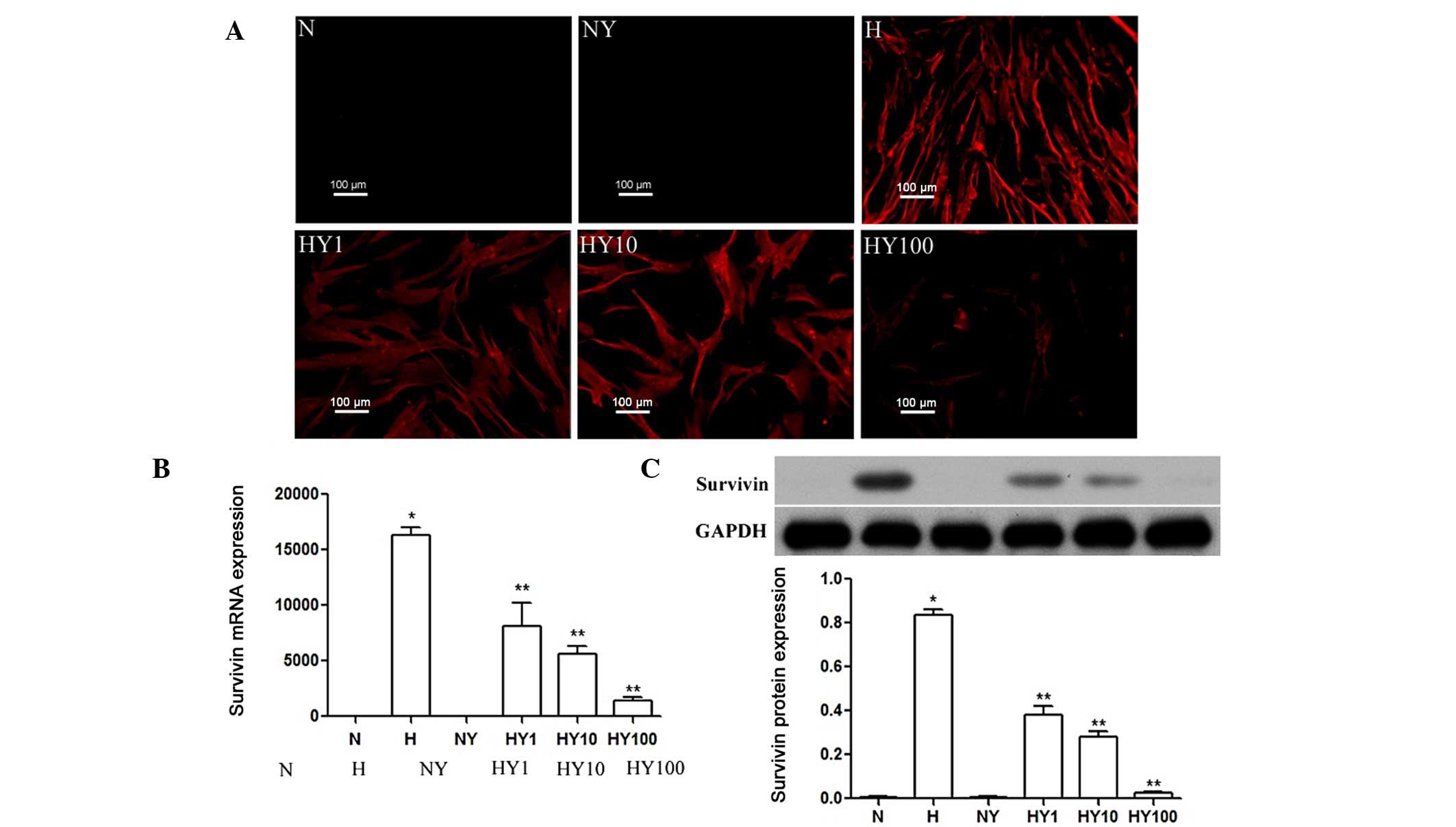 | Figure 1YM155 treatment inhibits the
expression of survivin in hypoxic HPASMCs. Expression levels of
survivin in hypoxic or normoxic HPASMCs, with or without YM155
treatment, were determined using (A) immunocytochemical (scale
bar=100 µm), (B) quantitative reverse
transcription-polymerase chain reaction and (C) Western blot
analyses. Data are presented as the mean ± standard deviation
(n=3). *P<0.05 vs. N group; **P<0.05
vs. H group. HPAMSCs, human pulmonary artery smooth muscle cells;
YM155, sepantronium bromide; N, normoxia; H, hypoxia; NY, normoxia
+ 100 nmol/l YM155; HY1, hypoxia + 1 nmol/l YM155; HY10, hypoxia +
10 nM YM155; HY100, hypoxia + 100 nmol/l YM155. |
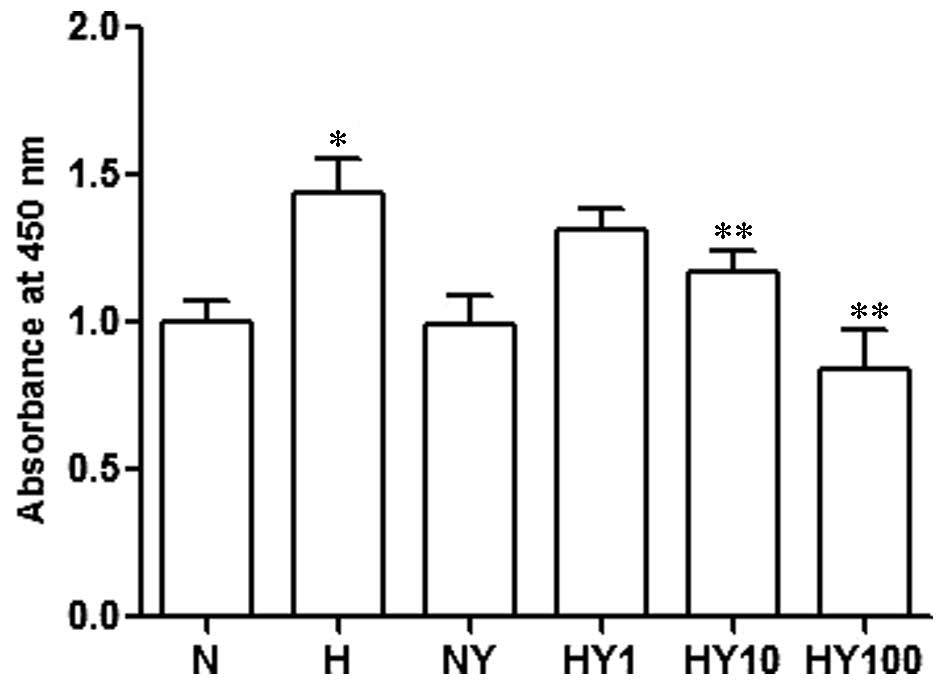
YM155 suppresses the cell viability of
hypoxic HPASMCs
To investigate the cell viability of the hypoxic or
normoxic HPASMCs with or without YM155 treatment, a CCK-8 cell
proliferation assay was performed. The results demonstrated that
cell proliferation was significantly increased by hypoxia, compared
with the normoxia-cultured HPASMCs (P<0.05). By contrast, YM155
treatment ameliorated this hypoxia-induced increase in cell
proliferation, in a concentration-dependent manner (P<0.05;
Fig. 2). YM155 treatment in the
normoxic HPASMCs had no significant effect on the proliferation of
the HPASMCs.
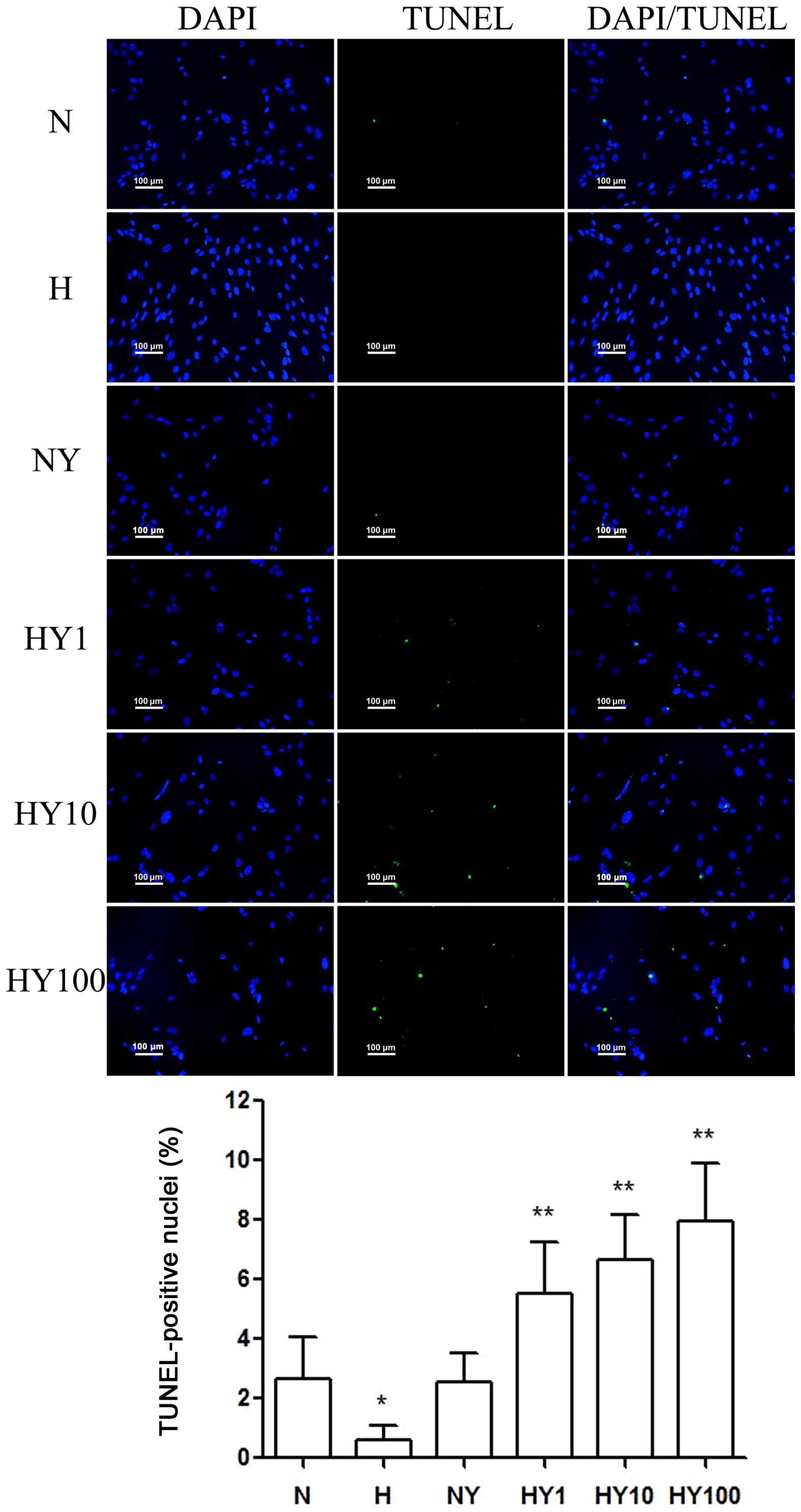
YM155 induced apoptosis of hypoxic
HPASMCs
To investigate whether YM155 suppressed cell
viability by increasing apoptosis of the HPASMCs during hypoxia, a
TUNEL assay was performed (Fig.
3). Hypoxia induced HPASMC proliferation and suppressed
apoptosis. Following treatment with YM155, a significant decrease
in cell viability was detected in the hypoxic HPASMCs, and the
percentage of TUNEL-positive cells decreased in a dose-dependent
manner. However, no significant effects were observed in the
normoxic HPASMCs treated with 10 nmol/l YM155 for 24 h.
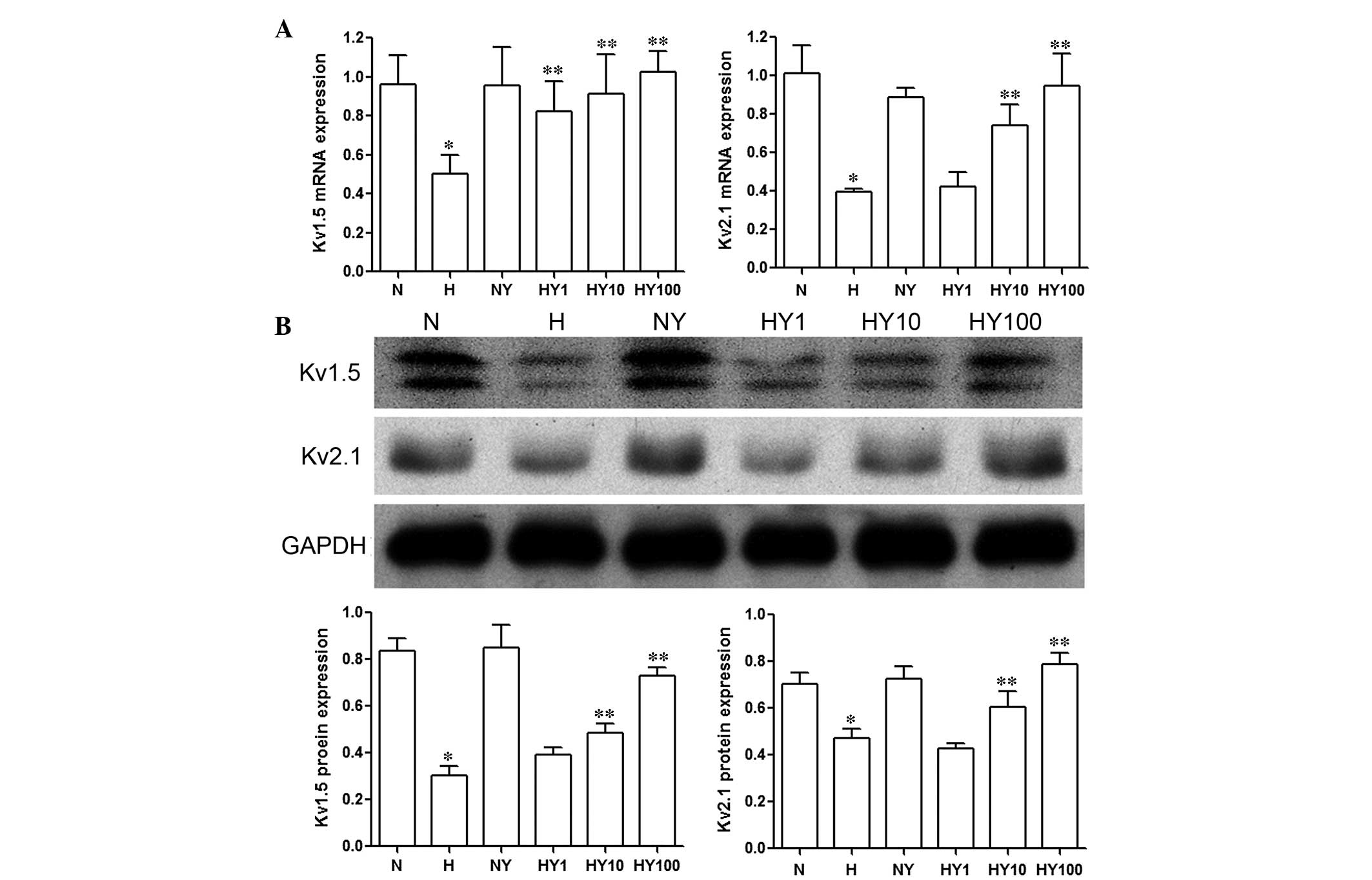
YM155 upregulates the expression levels
of Kv1.5 and Kv2.1 in hypoxic HPASMCs
The results of the RT-qPCR and Western blot analyses
demonstrated that the expression levels of Kv1.5 and
Kv2.1 were markedly inhibited by hypoxia, compared with
the HPASMCs cultured under normoxic conditions (P<0.05; Fig. 4). Treatment with YM155 resulted in
a concentration-dependent increase in the mRNA (Fig. 4A) and protein (Fig. 4B) expression levels of
Kv1.5 and Kv2.1 in the HPASMCs cultured in
hypoxia (P<0.05). No significant changes were observed in the
expression levels of Kv1.5 and Kv2.1 in the
normoxic HPASMCs following YM155 treatment (Fig. 4).
Discussion
PH is an obstructive vasculopathy characterized by
the increased proliferation and suppressed apoptosis of PASMCs.
This phenotype is associated with activation of the expression of
survivin, which is important in PH pathogenesis (20). In the present study, the potential
role for survivin, an inhibitor of apoptosis, was investigated in
normoxic and hypoxic HPASMCs. The results demonstrated that
survivin was expressed in HPASMCs during hypoxia but not in
normoxic conditions. Treatment of the PASMCs under hypoxic
condition with YM155, a novel small-molecule inhibitor of survivin,
reversed the hypoxia-induced overexpression of survivin at the
transcriptional and translational levels, as demonstrated by
RT-qPCR, immunocytochemical and Western blot analyses. In addition,
the results of the present study also indicated that the
proliferation of the PASMCs was significantly induced in the
hypoxia-cultured cells, compared with the normoxia-cultured cells.
In addition, the apoptosis of the PASMCs was signifi-cantly
inhibited, and the activation of Kv1.5 and
Kv2.1 were reduced in the hypoxic HPASMCs. The
therapeutic benefit of YM155 on HPASMCs during hypoxia was
associated with the inhibited growth and increased apoptosis of
PASMCs, as well as the reactivation of the expression of
Kv1.5 and Kv2.1. These results suggested that
survivin may be key in the pathophysi-ology of hypoxia-induced PH,
and that the YM155 survivin inhibitor may offer therapeutic
potential for use in the treatment of PH, although future
investigations in animal models are required.
Gene microarray analysis has demonstrated that
dysregulation of apoptotic mediators in the PA wall of patients
with PH favored the suppression of apoptosis (21). Survivin is the smallest member of
the IAP family, which is structurally unique and involved in
essential cellular functions, and is prominently expressed in human
cancer (22). High expression
levels of survivin in cancer cells are associated with poor patient
prognosis and survival rates, as well as resistance to therapy and
an increased rate of cancer recurrence (13,23).
However, the expression of survivin is not cancer specific, and it
is involved in essential cellular functions. It has been observed
to be upregulated in the media of small and medium-sized PAs from
monocrotaline-induced pulmonary arterial hypertension rats, as well
as idiopathic PH patients (14,21).
Increasing evidence has indicated that the expression of survivin
is increased in PAs of PH (24). A
previous study on chronic hypoxia-induced PH demonstrated the
overexpression of survivin in the media of the distal PAs of rats
following chronic hypoxia (18).
Hyperplasia of PASMCs is a characteristic pathological feature of
pulmonary hypertension (25).
Survivin may facilitate the transition of PASMCs to a proliferative
state and inhibit apoptosis by hyperpolarizing mitochondria
(14). This would result in the
impaired activation of Kv channels and, subsequently,
mitochondria-dependent apoptosis (9,26).
Increased proliferation of PASMC has been reported in animals with
hypoxia-induced pulmonary hypertension, which is often found to be
associated with vascular remodeling (27) Consistent with the results of
previous studies, the results of the present study indicated that
exposure of PASMCs to hypoxia enhanced the proliferation and
decreased the apoptosis of PASMCs, significantly upregulated the
expression of prosurvival factor, survivin, and downregulated the
expression of Kv1.5 and Kv2.1 channel
proteins. This is consistent with the results of a previous study
by McMurtry et al (14),
which indicated that survivin allowed the PASMCs to enter a
proliferative phase, parallel with a decrease in Kv1.5
expression. This previous study also provided direct evidence that
survivin inhibition led to selective apoptosis of the proliferating
PASMCs. This suggested that survivin may be central in the
hyperplasia of hypoxia-induced PASMCs, which may also elucidate the
therapeutic potential of survivin targeting in PH.
Multiple strategies to modulate the expression and
activation of survivin have been developed. As a selective survivin
inhibitor, YM155, a small molecule of sepantronium bromide, exerts
a marked antiproliferative activity in a large panel of tumor cell
models. Preclinical studies have indicated that continuous infusion
of YM155 resulted in a reduction of intratumor survivin in
tumor-bearing immunodeprived mice leading to marked tumor
regression due to an enhanced apoptotic response (28). Survivin was demonstrated to be
expressed at high levels in a previous study of chronic
hypoxia-induced PH rats, and indicated that YM155 treatment
downregulated the expression of survivin in the distal PAs and lung
tissues during chronic hypoxia, reversing hypoxia-induced PH
(18). This suggests YM155 as an
ideal candidate in the treatment of PH. In the present study, the
therapeutic effects of YM155 on the proliferation and apop-tosis of
HPASMCs during hypoxia were investigated. The cells were treated
with different concentrations of the YM155 survivin inhibitor. The
results of demonstrated that, in addition to reversing the
hypoxia-induced upregulation of survivin in the PASMCs, YM155
treatment significantly promoted apoptosis and suppressed PASMC
proliferation, consistent with the results of the
survivin-targeting strategy developed by McMurtry et al
(14). The disturbance in the
balance of apoptosis and proliferation in pulmonary vascular wall
cells, favoring proliferation, induces the remodeling of PAs
(29,30). Effective therapies require a shift
in this balance towards apoptosis (31). The data of the present study
indicated that the hypoxia-induced increase in survivin may have
been associated with the imbalance of apoptosis and proliferation
in the PASMCs, and the downregulation of survivin as a result of
YM155 administration in the hypoxic PASMCs resulted in an enhanced
apoptotic response.
Kv channels are an important factor in
hypoxic PH progression (32,33).
The inhibition or lack of Kv channels contributes to
resistance to apoptosis (16),
whereas Kv upregulation contributes to the promotion of
apoptosis via K+ activity of caspases (8). Furthermore, PH is associated with
selective inhibition of Kv channels. Prolonged hypoxia
inhibits the mRNA and protein expression of Kv channels,
and decreases the number of functional Kv channels in
PASMCs. The resulting membrane depolarization raises cytoplasmic
free calcium concentrations, stimulating PASMC proliferation and,
ultimately, increasing pulmonary vascular resistance and PH
(34). The upregulated expression
of survivin in patients with PH has been suggested to be associated
with the impaired activation of Kv channels and
consequent mitochondria-dependent apoptosis (9,26).
In vitro and in vivo investigations have demonstrated
that survivin targeting induces mitochondria-dependent apoptosis of
PASMCs, and the activation of K+ channels by survivin
targeting may be important in this process (14,35).
The present study further investigated the potential therapeutic
mechanisms of YM155 on hypoxic HPASMCs, predominantly focussing on
the expression of the specific PASMC Kv channels,
Kv1.5 and Kv2.1. The results revealed that
hypoxia exposure significantly inhibited the expression levels of
Kv1.5 and Kv2.1 in the HPASMCs. YM155
treatment reversed the hypoxia-induced downregulation of
Kv channel proteins in a dose-dependent manner,
suppressed proliferation and enhanced apoptosis of the PASMCs. The
expression levels of Kv1.5 and Kv2.1
increased, parallel with the decrease in survivin. By contrast,
YM155 had little effect on the expression levels of Kv1.5 and Kv2.1
in the normoxic-cultured HPASMCs. These data suggested that
survivin may regulate the balance of cell proliferation and
apoptosis in hypoxic PASMCs through a Kv
channel-associated mechanism, and YM155 may offer therapeutic
potential for use in hypoxia-induced PH.
In conclusion, the present study demonstrated that
exposure to hypoxia of HPASMCs significantly increased cell
proliferation and suppressed apoptosis, upregulated the mRNA and
protein expression levels of survivin, and reduced the activation
of the Kv1.5 and Kv2.1 channels. YM155, a
selective survivin inhibitor, reversed the hypoxia-induced
apoptosis suppression and proliferation enhancement in the HPASMCs,
in a dose-dependent manner, which was associated with survivin
inhibition and reactivation of the Kv channels. These
results are the first, to the best of our knowledge, to demonstrate
that YM155 has a beneficial therapeutic effect on hypoxic HPASMCs,
and provided evidence that the pro-apoptotic effects induced by
YM155 involved downregulation of the apoptosis inhibitor, survivin,
possibly via a Kv channel-mediated mechanism.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (grant no. 81170045).
Abbreviations:
|
CCK-8
|
Cell Counting Kit-8
|
|
H
|
hypoxia
|
|
Kv
|
voltage-dependent K+
channels
|
|
IAP
|
inhibitor of apoptosis
|
|
N
|
normoxia
|
|
PAs
|
pulmonary arteries
|
|
PASMCs
|
pulmonary artery smooth muscle
cells
|
|
HPASMCs
|
human PASMCs
|
|
PBS
|
phosphate-buffered saline
|
|
PH
|
pulmonary hypertension
|
|
TUNEL
|
terminal deoxynucleotidyl
transferase-mediated deoxyuridine triphosphate nick end
labeling
|
|
YM155
|
sepantronium bromide
|
References
|
1
|
Guignabert C and Dorfmuller P: Pathology
and pathobiology of pulmonary hypertension. Semin Respir Crit Care
Med. 34:551–559. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
McMurtry MS, Bonnet S, Wu X, Dyck JR,
Haromy A, Hashimoto K and Michelakis ED: Dichloroacetate prevents
and reverses pulmonary hypertension by inducing pulmonary artery
smooth muscle cell apoptosis. Circ Res. 95:830–840. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang J, Weigand L, Lu W, Sylvester JT,
Semenza GL and Shimoda LA: Hypoxia inducible factor 1 mediates
hypoxia-induced TRPC expression and elevated intracellular
Ca2+ in pulmonary arterial smooth muscle cells. Circ
Res. 98:1528–1537. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hu HL, Zhang ZX, Chen CS, Cai C, Zhao JP
and Wang X: Effects of mitochondrial potassium channel and membrane
potential on hypoxic human pulmonary artery smooth muscle cells. Am
J Respir Cell Mol Biol. 42:661–666. 2010. View Article : Google Scholar
|
|
5
|
Chen C, Chen C, Wang Z, Wang L, Yang L,
Ding M, Ding C, Sun Y, Lin Q, Huang X, et al: Puerarin induces
mitochondria-dependent apoptosis in hypoxic human pulmonary
arterial smooth muscle cells. PLoS One. 7:e341812012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yung GL: Evaluation of patients with
pulmonary hypertension for lung transplantation. Textbook of
Pulmonary Vascular Disease. Yuan JXJ, Garcia JGN, Hales CA, Rich S,
Archer SL and West JB: Springer; New York, NY: pp. 1593–1598. 2011,
View Article : Google Scholar
|
|
7
|
Archer SL, Gomberg-Maitland M, Maitland
ML, Rich S, Garcia JG and Weir EK: Mitochondrial metabolism, redox
signaling and fusion: A mitochondria-ROS-HIF-1alpha-Kv1.5
O2-sensing pathway at the intersection of pulmonary hypertension
and cancer. AM J Physiol Heart Circ Physiol. 294:H570–H578. 2008.
View Article : Google Scholar
|
|
8
|
Park WS, Firth AL, Han J and Ko EA:
Patho-, physiological roles of voltage-dependent K+
channels in pulmonary arterial smooth muscle cells. J Smooth Muscle
Res. 46:89–105. 2010. View Article : Google Scholar
|
|
9
|
Altieri DC: Validating survivin as a
cancer therapeutic target. Nature Reviews Cancer. 3:46–54. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zyada MM: Relationship of survivin to
clinical drug resistance in Burkitt's lymphoma of the head and neck
region. Med Oncol. 28:1565–1569. 2011. View Article : Google Scholar
|
|
11
|
Dohi T, Beltrami E, Wall NR, Plescia J and
Altieri DC: Mitochondrial survivin inhibits apoptosis and promotes
tumorigenesis. J Clin Invest. 114:1117–1127. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Salvesen GS and Duckett CS: IAP proteins:
Blocking the road to death's door. Nat Rev Mol Cell Biol.
3:401–410. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hingorani P, Dickman P, Garcia-Filion P,
White-Collins A, Kolb EA and Azorsa DO: BIRC5 expression is a poor
prognostic marker in ewing sarcoma. Pediatr Blood Cancer. 60:35–40.
2013. View Article : Google Scholar
|
|
14
|
McMurtry MS, Archer SL, Altieri DC, Bonnet
S, Haromy A, Harry G, Bonnet S, Puttagunta L and Michelakis ED:
Gene therapy targeting survivin selectively induces pulmonary
vascular apoptosis and reverses pulmonary arterial hypertension. J
Clin Invest. 115:1479–1491. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ryan BM, O'Donovan N and Duffy MJ:
Survivin: A new target for anti-cancer therapy. Cancer Treat Rev.
35:553–562. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yamanaka K, Nakata M, Kaneko N, Fushiki H,
Kita A, Nakahara T, Koutoku H and Sasamata M: YM155, a selective
survivin suppressant, inhibits tumor spread and prolongs survival
in a spontaneous metastatic model of human triple negative breast
cancer. Int J Oncol. 39:569–575. 2011.PubMed/NCBI
|
|
17
|
Tao YF, Lu J, Du XJ, Sun LC, Zhao X, Peng
L, Cao L, Xiao PF, Pang L, Wu D, et al: Survivin selective
inhibitor YM155 induce apoptosis in SK-NEP-1 Wilms tumor cells. BMC
Cancer. 12:6192012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu B, Fan Z, Li J, Liu Y, Wang N, Wang D,
Liu Y and Zhang B: Expression of survivin in pulmonary artery of
rats exposed to normoxia and hypoxia. International Journal of
Respiration. 33:994–998. 2013.
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2− ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar
|
|
20
|
Paulin R, Courboulin A, Meloche J, et al:
Signal transducers and activators of transcription-3/pim1 axis
plays a critical role in the pathogenesis of human pulmonary
arterial hypertension. Circulation. 123:1205–1215. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Geraci MW, Moore M, Gesell T, Yeager ME,
Alger L, Golpon H, Gao B, Loyd JE, Tuder RM and Voelkel NF: Gene
expression patterns in the lungs of patients with primary pulmonary
hypertension: A gene microarray analysis. Circ Res. 88:555–562.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Altieri DC: Targeting survivin in cancer.
Cancer Lett. 332:225–228. 2013. View Article : Google Scholar :
|
|
23
|
Stauber RH, Mann W and Knauer SK: Nuclear
and cytoplasmic survivin: Molecular mechanism, prognostic and
therapeutic potential. Cancer Res. 67:5999–6002. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Paulin R, Meloche J, Jacob MH, Bisserier
M, Courboulin A and Bonnet S: Dehydroepiandrosterone inhibits the
Src/STAT3 constitutive activation in pulmonary arterial
hypertension. Am J Physiol Heart Circ Physiol. 301:H1798–H1809.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Marcos E, Fadel E, Sanchez O, Humbert M,
Dartevelle P, Simonneau G, Hamon M, Adnot S and Eddahibi S:
Serotonin-induced smooth muscle hyperplasia in various forms of
human pulmonary hypertension. Circ Res. 94:1263–1270. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ekhterae D, Platoshyn O, Krick S, Yu Y,
McDaniel SS and Yuan JX: Bcl-2 decreases voltage-gated
K+ channel activity and enhances survival in vascular
smooth muscle cells. Am J Physiol Cell Physiol. 281:C157–C165.
2001.PubMed/NCBI
|
|
27
|
Reid L: Vascular remodeling. The Pulmonary
Circulation: Normal and Abnormal Mechansimc, Management and the
National Registry. Fishman A: University of Pennsylvania Press;
Philadelphia, PA: pp. p2641990
|
|
28
|
Nakahara T, Kita A, Yamanaka K, Mori M,
Amino N, Takeuchi M, Tominaga F, Kinoyama I, Matsuhisa A, Kudou M
and Sasamata M: Broad spectrum and potent antitumor activities of
YM155, a novel small-molecule survivin suppressant, in a wide
variety of human cancer cell lines and xenograft models. Cancer
Sci. 102:614–621. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mandegar M, Fung YC, Huang W, Remillard
CV, Rubin LJ and Yuan JX: Cellular and molecular mechanisms of
pulmonary vascular remodeling: Role in the development of pulmonary
hypertension. Microvasc Res. 68:75–103. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fernandez RA, Sundivakkam P, Smith KA,
Zeifman AS, Drennan AR and Yuan JX: Pathogenic role of
store-operated and receptor-operated ca(2+) channels in
pulmonary arterial hypertension. J Signal Transduct.
2012:9514972012. View Article : Google Scholar
|
|
31
|
Gurbanov E and Shiliang X: The key role of
apoptosis in the pathogenesis and treatment of pulmonary
hypertension. Eur J Cardiothorac Surg. 30:499–507. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Michelakis ED, McMurtry MS, Wu XC, Dyck
JR, Moudgil R, Hopkins TA, Lopaschuk GD, Puttagunta L, Waite R and
Archer SL: Dichloroacetate, a metabolic modulator, prevents and
reverses chronic hypoxic pulmonary hypertension in rats: Role of
increased expression and activity of voltage-gated potassium
channels. Circulation. 105:244–250. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Moudgil R, Michelakis ED and Archer SL:
The role of K+ channels in determining pulmonary
vascular tone, oxygen sensing, cell proliferation and apoptosis:
Implications in hypoxic pulmonary vasoconstriction and pulmonary
arterial hypertension. Microcirculation. 13:615–632. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sweeney M and Yuan JX: Hypoxic pulmonary
vasoconstriction: Role of voltage-gated potassium channels. Respir
Res. 1:40–48. 2000. View
Article : Google Scholar
|
|
35
|
Pozeg ZI, Michelakis ED, McMurtry MS,
Thébaud B, Wu XC, Dyck JR, Hashimoto K, Wang S, Moudgil R, Harry G,
et al: In vivo gene transfer of the O2-sensitive
potassium channel Kv1. 5 reduces pulmonary hypertension and
restores hypoxic pulmonary vasoconstriction in chronically hypoxic
rats. Circulation. 107:2037–2044. 2003. View Article : Google Scholar : PubMed/NCBI
|