Introduction
Ewing's sarcoma is a primary malignant bone cancer
type that mainly affects children and adolescents, while it rarely
occurs in adults over the age of 30 years (1–3). It
is the second most common type of bone cancer in children with a
male/female ratio of 1.6:1 (1,2). The
currently used treatment of Ewing's sarcoma consists of a
combination of systemic anti-cancer chemotherapy and complete
surgical resection (3–5). However, the majority of patients with
Ewing's sarcoma develop resistance to chemotherapy (5). The underlying mechanisms of the
chemoresistance of Ewing's sarcoma have been identified to include
enhanced drug-efflux pump activity, upregulation of DNA repair
mechanisms, changes in the intracellular metabolic pathway
(6), the aberrant expression of
microRNA (7) and impairment of
apoptosis (6). Therefore, novel
treatment strategies with reduced chemoresistance as well as
methods to inhibit the abovementioned mechanisms of chemoresistance
are required, while a relatively low toxicity to surrounding normal
tissues is also essential.
Epidermal growth factor receptor (EGFR), an abundant
transmembrane glycoprotein of the tyrosine kinase growth factor
receptor family (8,9), is expressed in numerous types of
normal human tissues; the activation of this proto-oncogene by
ligand binding to its extra-cellular domain results in the
activation of downstream cellular signals that regulate cell
growth, resistance to anti-cancer drugs, promotion of tumor
invasion and angiogenesis (9,10).
Cetuximab (CTX) is an anti-cancer drug with a binding affinity to
EGFR greater than that of its natural ligand EGF, thereby blocking
the ligand-induced activation of EGFR. In addition, cetuximab
stimulates the internalization of EGFR thereby removing it from the
cell surface and any possible ligand interaction (11).
Lactate dehydrogenase-A (LDHA) is one of the
metabolic key enzymes controlling the conversion of pyruvate to
lactate as part of the cellular glycolytic process (12). Studies have reported that LDHA has
a key role in tumor growth, chemoresistance and tumor metastasis of
numerous types of cancer (13).
LDHA has been identified as a direct target of the c-myc and EGFR
oncogenes (12,14). In addition, LDHA is regulated by
hypoxia-inducible factor (HIF-1α) (14). The present study explored the
correlation between LDHA expression and CTX resistance in human
Ewing's sarcoma, revealing that LDHA is highly expressed in
CTX-resistant Ewing's sarcoma tissues and a cell line. Inhibition
of LDHA enhanced the sensitivity of Ewing's sarcoma cells to CTX.
The present study revealed a novel mechanism of cetuximab
resistance from the perspective of cancer cell metabolism.
Materials and methods
Cell culture and Ewing's sarcoma patient
samples
The A673 human Ewing's sarcoma cell line was
purchased from the American Type Culture Collection (Manassas, VA,
USA). Cells were cultured in Dulbecco's modified Eagle's medium
(Mediatech Inc., Manassas, VA, USA) with 10% fetal bovine serum in
an incubator containing a humidified atmosphere with 5%
CO2 at 37°C. Eight cetuximab-resistant and two
cetuximab-sensitive primary Ewing's sarcoma tissue samples were
used in the present study, which were in this project, which were
obtained from male and female patients (mean age, 24.2; age range,
15–43 years; male/female gender ratio, 1.4/1) undergoing surgery
for Ewing's sarcoma between January 2013 and October 2014 at the
Department of Orthopedics, Tianjin Third Central Hospital (Tianjin,
China). The CTX-resistant samples were obtained from patients who
had a negative response to CTX treatment and the CTX-sensitive
samples from patients who had a positive a response to CTX
treatment. All samples were stored in liquid nitrogen until
analysis. Tumors were obtained following a protocol approved by the
Ethics Committee of the Department of Orthopedics, Tianjin Third
Central Hospital (Tianjin, China). All patients provided written
informed consent.
Antibodies and reagents
The following antibodies were used in the present
study: LDHA (cat no. sc-130327; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA) and GAPDH (Scat no. sc-365062; Santa Cruz
Biotechnology, Inc.). Oxamate was purchased from Sigma-Aldrich (St.
Louis, MO, USA) and CTX was purchased from ImClone Systems
(Bridgewater, NJ, USA).
Generation of a CTX resistant cell
line
A673 human Ewing's sarcoma cells were treated with
CTX with stepwise dose increases of up to 20 µg/ml under
standard cell culture conditions for selection of resistant cells.
After successive treatments in continuous culture for four months,
several resistant cell clones were developed from the parental cell
line. CTX resistant pooled clones were used for all subsequent
experiments in the present study. The resistant cells were
re-selected by CTX treatments each month.
Western blot analysis
Cells were lysed in lysis buffer containing 50
mmol/l Tris-HCl (pH 7.4), 150 mmol/l NaCl, 0.5% NP-40, 50 mmol/l
NaF, 1 mmol/l Na3VO4, 1 mmol/l
phenylmethylsulfonyl fluoride, 25 mg/ml aprotinin and 25 mg/ml
leupeptin (Bio-Rad Laboratories, Inc., Hercules, CA, USA) and kept
on ice for 15 min. Following centrifugation at 12,000 × g for 15
min at 4°C, the supernatants were collected and the protein
concentration was determined using the Pierce Coomassie Plus
colorimetric protein assay (Thermo Fisher Scientific, Inc.).
Subsequently, equal amounts of protein (50 µg for each lane)
were subjected to 4–20% SDS-PAGE gradient gel (Bio-Rad
Laboratories, Inc.) and transferred onto a nitrocellulose membrane
(Bio-Rad Laboratories, Inc.). The membrane was blocked by 4%
non-fat dry milk for 1 h at room temperature prior to the
incubation. The membrane was incubated with primary LDHA mouse
monoclonal anti-human IgG1 (cat. no., sc-130327; dilution, 1:1,000)
and/or GAPDH mouse monoclonal IgG1 antibodies (cat. no., sc-365062;
dilution, 1:1,000) overnight at 4°C in phosphate-buffered saline
(PBS) containing 5% non-fat dry milk. Both primary antibodies were
obtained from Santa Cruz Biotechnology, Inc., Dallas, TX, USA.
Following extensive washing with PBS, membranes were incubated with
a horse anti-mouse IgG (heavy and light chain) horseradish
peroxidase-conjugated secondary antibody (cat. no., 7074; dilution,
1:2,000; Cell Signaling Technology, Inc., Danvers, MA, USA). The
antigen-antibody complexes were then visualized using an Enhanced
Chemiluminescence Detection kit (GE Healthcare, Little Chalfont,
UK). The images were developed using ChemiDoc™ Touch Gel Imaging
System (1708370; Bio-Rad Laboratories, Inc., Hercules, CA, USA).
The quantification of the blots was performed using ImageJ
software.
cDNA preparation and
reverse-transcription quantitative polymerase chain reaction
(RT-qPCR) analysis
Cells and tissues were homogenized and total RNA was
extracted using an RNeasy mini kit (Qiagen, Hilden Germany), which
was simultaneously subjected to DNase digestion performed using the
High Capacity cDNA Reverse Transcription kit (Applied Biosystems;
Thermo Fisher Scientific, Inc., Waltham, MA, USA). Real-time qPCR
was then performed to amplify the obtained cDNA; the PCR mixture
contained TaqMan Gene Expression Assay primers and probes specific
for LDHA were supplied by Applied Biosystems and were as follows:
5′-ATCTTGACCTACGTGGCTTGGA-3′ forward and
5′-CCATACAGGCACACTGGAATCTC-3′ reverse. Ribosomal 18S RNA, was used
as an internal control and the primers used were as follows:
5′-CTACCACATCCAAGGAAGCA-3′ forward and 5′-TTTTTCGTCACTACCTCCCCG-3′
reverse (Applied Biosystems). PCR amplifications were performed in
a final reaction volume of 10 µl containing 0.5 µl of
the primers and probes mix, 5.5 µl TaqMan Universal PCR
Master Mix (Applied Biosystems) and 4.5 µg cDNA solution
diluted at 1:10. The thermocycling conditions were as follows: One
cycle of 50°C for 2 min and initial denaturation for 10 min at
95°C, followed by 40 cycles of denaturation (15 sec at 95°C) and
annealing extension (1 min at 60°C). The Step 1 Plus Real-Time PCR
Systems Thermocycler (Applied Biosystems) was used for all
reactions. RTqPCR was performed in triplicate with at least two
repetitions. The quantification cycle (Cq) of 18S ribosomal RNA was
used to determine the ΔCq value for mRNA expression. The formula
2(−ΔΔCq) was used to calculate the relative mRNA
expression (15).
Plasmid DNA and small interfering (si)RNA
transfections
Transfections were performed using the
Oligofectamine Transfection reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions.
Overexpression vector for wild type LDAH (cat no. RC209378) was
purchased from Origene (Cambridge, UK). siRNA oligonucleotides for
LDHA were purchased from Sigma-Aldrich, with a scrambled siRNA
(Sigma-Aldrich) used as a control. Following 48 h hours of
transfection, whole-cell lysates were prepared for subsequent
analysis.
Glucose consumption assay
Cells were seeded in six-well plates at
5×105 cells per well in 3 ml phenol red-free,
low-glucose, low-serum cell culture medium (Thermo Fisher
Scientific, Inc.). Following treatment with the cell culture
medium, a 50-µl aliquot of the conditioned medium was
collected from each well and diluted with 950 µl distilled
water (1:20). The glucose concentration in the diluted medium was
measured using the Glucose Assay kit (Sigma-Aldrich) according to
the manufacturer's instructions and the protocol of a previous
study (14). Glucose consumption
was calculated by subtracting the concentration of glucose
remaining in the medium at the indicated times from the
concentration of glucose present in fresh cell culture medium and
results were normalized to the amount of total protein compared
with that in the control group.
Lactate production assay
Cells were seeded in six-well plates at
5×105 cells per well in 3 ml phenol red-free,
low-glucose, low-serum cell culture medium. After the designated
treatments, a 50 µl aliquot of the conditioned medium was
collected from each well and diluted with 950 µl distilled
water (1:20). The lactate concentration in the diluted medium was
measured using the Lactate Assay kit from BioVision Inc.
(Cambridge, UK) according to the protocol of a previous study
(14). Results were normalized to
the amount of total protein compared with that in the control
group.
Cell survival assay
Cell survival was measured using a colorimetric
ELISA kit (Cell Death Detection ELISA; Roche Diagnostics Corp.,
Basel, Switzerland), which quantitatively measures cytoplasmic
histone-associated DNA fragments (mononucleosomes and
oligonucleosomes). The procedure was performed according to the
manufacturer's instructions. The results were verified by trypan
blue staining (Bio-Rad Laboratories, Inc. and direct cell counting
using a hemotocytometer (Thermo Fisher Scientific, Inc.).
Microarray database analysis
Data on the expression of LDHA in Ewing's sarcoma
were retrieved from the Oncomine cancer microarray database
(oncomine.org) in analogy with a previous study
(16,17). Oncomine contains 65 gene expression
datasets comprising ~48 million gene expression measurements form
>4700 microarray experiments.
Statistical analysis
The statistical analysis was performed using
GraphPad Prism 5.0 (GraphPad Software, Inc., La Jolla, CA, USA).
Student's t-test was used for all statistical analyses.
Values are expressed as the mean ± standard error. P<0.05 was
considered to indicate a statistically significant difference. All
experiments were repeated at least once with reproducible
results.
Results
LDHA is associated with CTX resistance in
Ewing's sarcoma
Since dysregulated glycolysis of cancer cells has
been studied as a target for overcoming chemoresistance and LDHA is
an important key enzyme of glycolysis, the present study explored
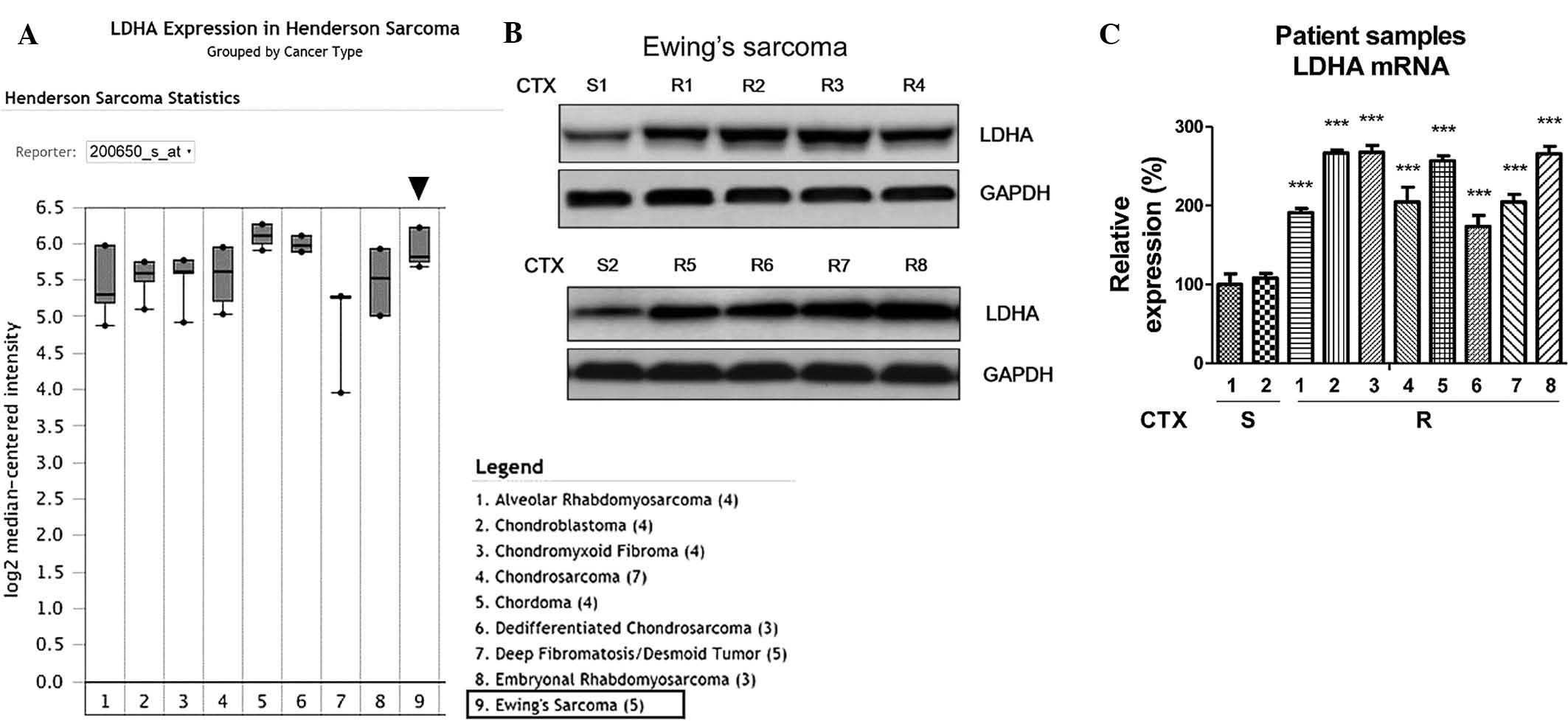
the link between LDHA and Ewing's sarcoma. Bioinformatics analysis
using the oncomine online microarray database revealed that LDHA
was upregulated in Ewing's sarcoma (Fig. 1A), indicating that LDHA may act as
an oncogene and contribute to chemoresistance. The present study
assessed the LDHA expression at the protein levels (Fig. 1B) as well as the mRNA levels
(Fig. 1C) in CTX-sensitive and
resistant Ewing's sarcoma tissues. As expected, the expression of
LDHA the in eight CTX-resistant Ewing's sarcoma tissues was
significantly upregulated compared with that in the nonresistant
tissues, suggesting that LDHA may be an important biomarker for the
clinical diagnosis and treatment of Ewing's sarcoma.
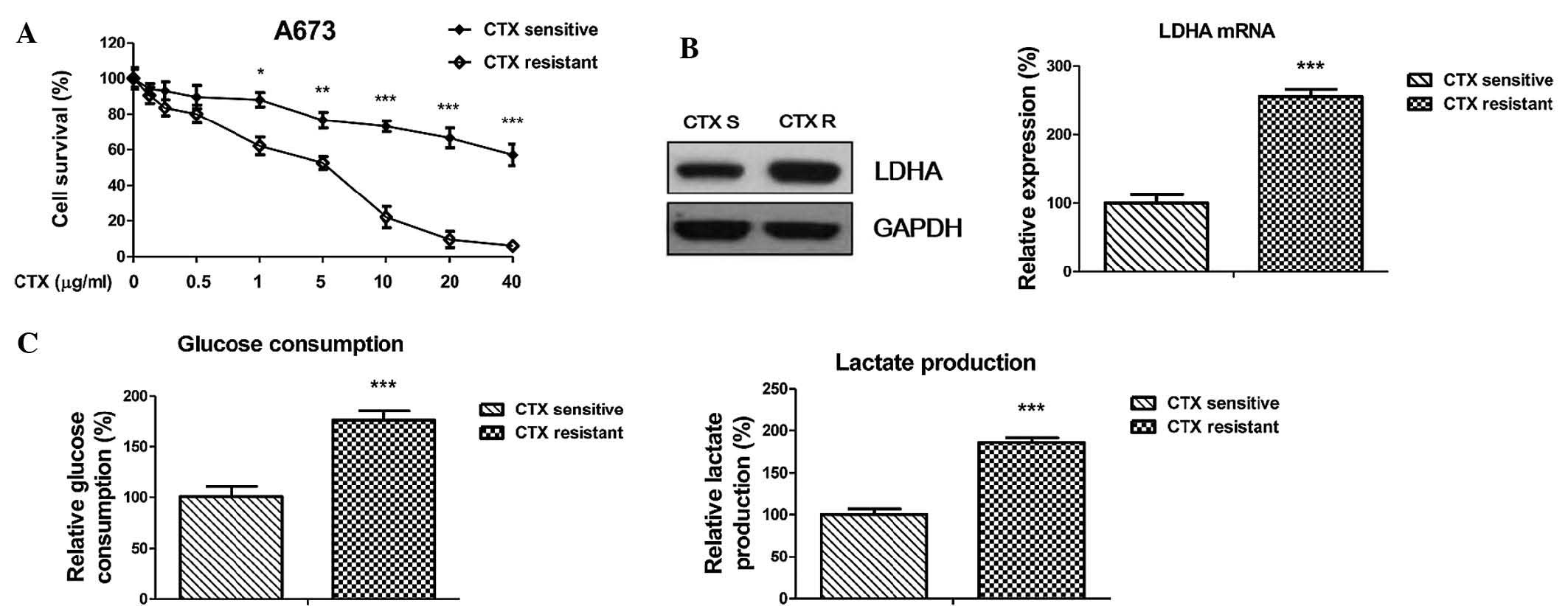
CTX-resistant Ewing's sarcoma cells
display an upregulated LDHA expression and glycolysis rate
To assess the underlying mechanisms of the
chemoresistance to CTX and to determine any possible links with the
dysregulated glycolysis of cancer cells, an isogenic CTX-resistant
human Ewing's sarcoma cell line, A673 CTX-R, was generated by
incubating parental cells to cetuximab with stepwise dose increases
of up to 20 µg/ml in continuous culture for four months. To
verify the resistance, A673 CTX-R cells and native A673 cells were
treated with CTX under identical experimental conditions (Fig. 2A). As expected, A673 CTX-R cells
tolerated significantly higher concentrations of cetuximab compared
with CTX-sensitive cells, whose cell survival was significantly
impaired by CTX at 5–40 µg/ml. Since the abovementioned
results revealed that LDHA was linked with CTX resistance in
Ewing's sarcoma patient samples, the LDHA levels in CTX-sensitive
and resistant cells were determined. In consistency with the
results obtained from the tissue samples, LDHA expression in CTX-R
A673 cells was upregulated at the protein and mRNA level compared
with that in A673 cells (Fig. 2B).
To assess the effects of CTX on aerobic glycolysis, glucose
consumption and lactate production were directly measured in the
supernatants of CTX-sensitive and resistant cells. The results
revealed that glucose consumption and lactate production were
significantly increased in CTX-resistant cells compared with those
in native A673 cells (Fig. 2C),
which was an indicator of the dysregulated glycolysis by LDHA in
CTX-resistant cells; this aberration may be utilized as a
therapeutic target for Ewing's sarcoma.
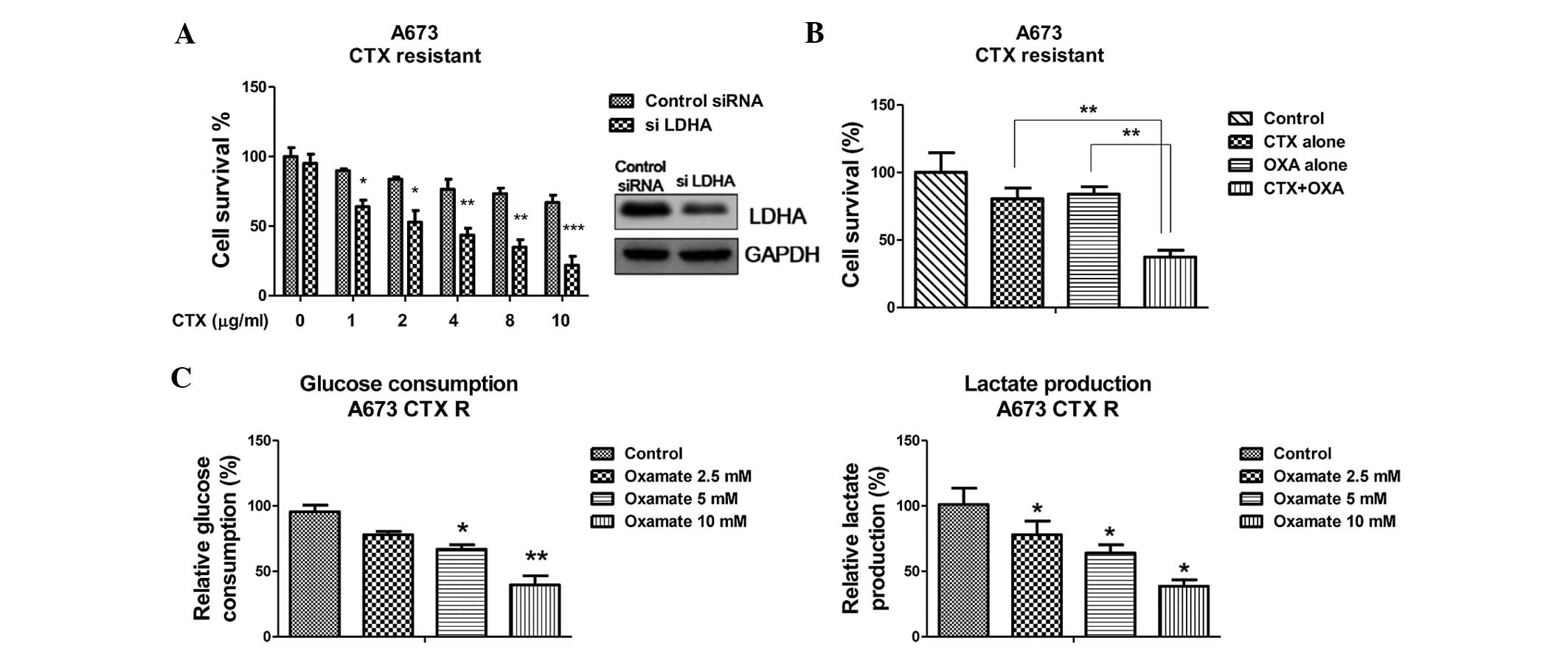
Inhibition of LDHA enhances sensitivity
to CTX
To further elucidate the link between the
upregulation of LDHA and the resistance of CTX as well as its
targetability for enhancing the efficiency of chemotherapy, a cell
survival analysis after subjecting the cells to treatments with CTX
and/or knockdown of LDHA by siRNA. LDHA knockdown significantly
reduced the survival of A673 cells in the presence of CTX (Fig. 3A). The reversed phenotypes verified
that LDHA was upregulated in CTX-resistant Ewing's sarcoma cells.
The IC50 of CTX on control-transfected A673 cells was ~4
µg/ml, while it was decreased to ~0.5 µg/ml following
knockdown of LDHA. Next, the present study investigated the
inhibitory effects of a combination of CTX and oxamate (18), which is an inhibitor of LDHA, on
the A673 cells. Treatment of parental A673 cells with CTX alone at
0.5 µg/ml or oxamate alone at 2 mM slightly reduced the cell
viability (to 90 and 85% live cells, respectively), while the
combination of CTX and oxamate synergistically reduced the cell
viability to a significantly greater extent (38% live cells)
(Fig. 3B), indicating that the
inhibition of LDHA either by siRNA or the small-molecule inhibitor
oxamate enhanced the chemotherapeutic efficiency of CTX in Ewing's
sarcoma. As expected, the glucose consumption and lactate
production were significantly decreased following inhibition of
LDHA (Fig. 3C and D).
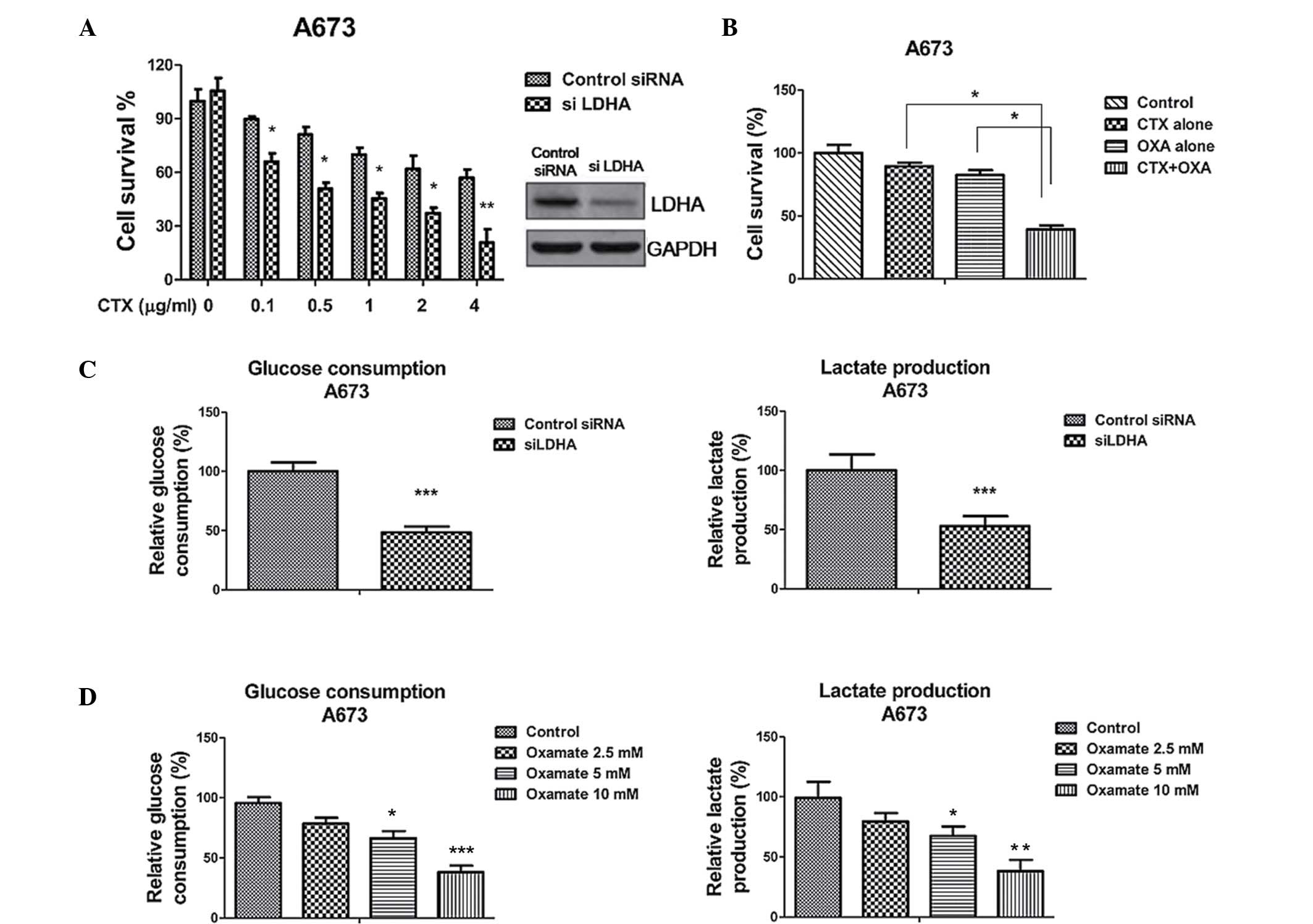 | Figure 3Inhibition of LDHA sensitizes Ewing's
sarcoma cells to CTX. (A) LDHA knockdown enhanced the cells'
sensitivity to cetuximab. A673 cells were transfected with control
siRNA or siLDHA for 48 hours, followed by treatment with cetuximab
at the indicated concentrations for 24 h. The cell survival ratio
was determined by ELISA of DNA fragments following lysis. (B) A673
cells were transfected with control siRNA or siLDHA for 48 h, and
levels of glucose (left) and lactate (right) in the supernatant
were determined following incubation in low-glucose (1 g/l),
low-serum (0.5%) medium for 24 h. (C) A673 cells were treated with
CTX alone at 0.5 µg/ml, oxamate alone at 2 mM or with their
combination for 48 h, followed by the measurement of cell survival
using ELISA. (D) A673 cells were treated with oxamate (2.5, 5 or 10
mM) for 48 h, and levels of glucose (left) and lactate (right) in
the supernatant were determined following incubation in low-glucose
(1 g/l), low-serum (0.5%) medium for 24 h. Values are expressed as
the mean ± standard error of three independent experiments.
*P<0.05; **P<0.01;
***P<0.001 vs. control. CTX, cetuximab; LDHA, lactate
dehydrogenase-A; S, sensitive; R, resistant; siRNA, small
interfering RNA. |
Synergistic effects of LDHA inhibitor and
CTX against CTX-resistant Ewing's sarcoma cells
Since the results of the present study demonstrated
that LDHA was upregulated in CTX-resistant Ewing's sarcoma cells
and that downregulation of LDHA by siRNA or oxamate significantly
inhibited the viability of the parental A673 cells, it was
hypothesized that inhibition of LDHA in CTX-resistant cells has
synergistic effects with CTX. Fig.
4A displays the synergistic inhibitory effects of the
combination of LDHA knockdown with CTX in A673-resistant cells.
Similarly, CTX combined with oxamate was significantly more potent
at inhibiting cell survival compared with either agent alone
(Fig. 4B). In consistency with
these results, glycolysis was inhibited by either LDHA knockdown as
well as oxamate treatment (Fig. 3C and
D). In conclusion, CTX and oxamate exhibited a synergistic
effect to reduce the viability of CTX-resistant cells, indicating
that LDHA inhibition may be an efficient adjuvant in the treatment
of Ewing's sarcoma with CTX.
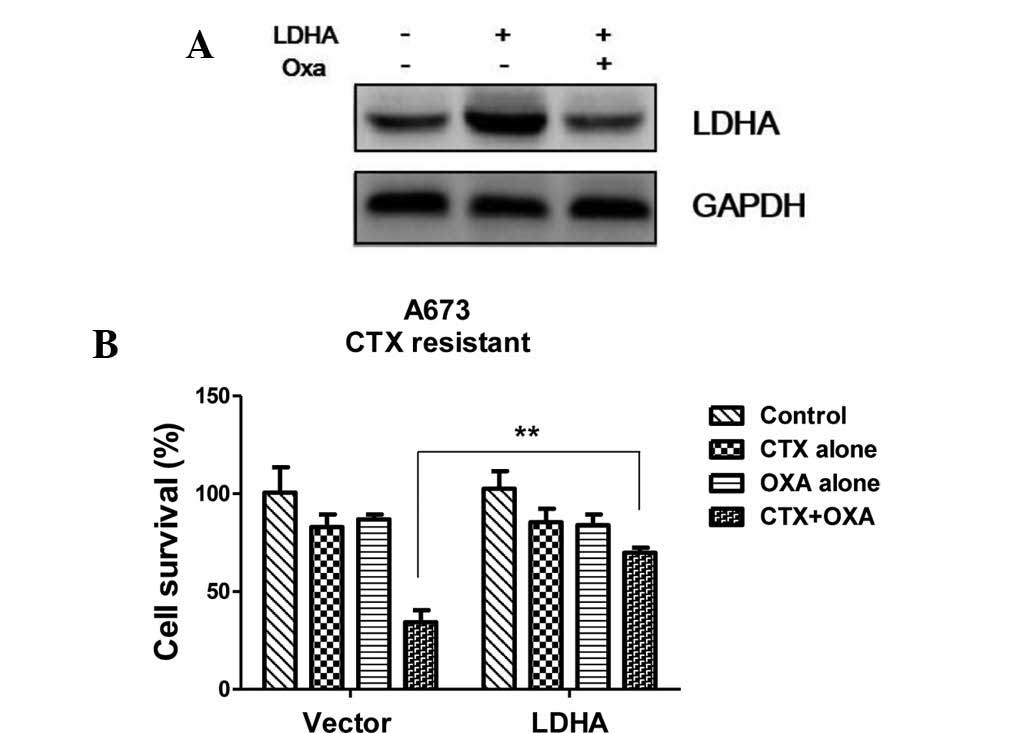
Restoration of LDHA reduces the
sensitivity of CTX-resistant A673 cells to CTX and oxamate
To further support the hypothesis that LDHA is
associated with resistance to chemotherapy in Ewing's sarcoma, LDHA
expression in CTX-resistant cells was restored by transient
transfection with LDHA overexpression vector. Western blot analysis
showed that the expression of LDHA was enhanced following
transfection, while treatment with oxamate reduced LDHA levels to
those of untransfected CTX-resistant cells (Fig. 5A). Furthermore, the survival rate
of LDHA-overexpressing, CTX-resistant cells was assessed following
treatment with CTX alone, oxamate alone or their combination. The
chemo-resistance of A673 CTX-R cells with restoration of LDHA was
shown to be enhanced in the presence of CTX and oxamate combined
(Fig. 5B).
Discussion
The treatment of Ewing's sarcoma requires surgery
along with intensive chemotherapy due to the aggressiveness of this
cancer type. As resistance to chemotherapy is common in Ewing's
sarcoma, the development of strategies to overcome this
chemoresistance is urgently required (1,5). The
present study compared cetuximab-sensitive and resistant Ewing's
sarcoma tissues, indicating that upregulation of LDHA is linked
with resistance and may therefore be a target for novel therapeutic
strategies. To the best of our knowledge, the present study was the
first to report the link between LDHA and CTX resistance in Ewing's
sarcoma.
Since cancer cells rely on an uninterrupted supply
of energy and nutrients to fuel their unlimited proliferation, they
utilize aerobic glycolysis to generate these; for this, they direct
the pyruvate pathway towards the production of lactate in the
cytosol under catalysis by LDHA, but not through oxidative
phosphorylation (19). This unique
phenomenon is known as the 'Warburg effect' and leads to an
elevated glycolytic flux in cancer cells. It has been reported that
CTX reverses the Warburg effect by inhibiting HIF-1-regulated LDHA
in cancer cells, indicating that elevated LDHA levels may be
associated with resistance to CTX. The present study established a
CTX-resistant A673 Ewing's sarcoma cell line and demonstrated
elevated levels of LDHA and are a higher glycolytic rate compared
with those of their parental cells. Inhibition of LDHA by siRNA or
oxamate decreased the glycolysis rate of CTX-sensitive and
resistant cells. Furthermore, knockdown or inhibition of LDHA
expression in Ewing's sarcoma cells sensitized them to CTX.
Consistent with the results of the present study, a previous study
reported that upregulated LDHA expression was correlated with Taxol
resistance in breast cancer (20).
Another recent study reported that inhibition of LDHA by microRNA
contributed to the re-sensitization of 5-fluorouracil-resistant
colon cancer cells (19). In
addition, the present study assessed the potency of the combination
of CTX with LDHA inhibitor oxamate against CTX-resistant cells. As
expected, the combination of CTX with oxamate showed a synergistic
effect to inhibit cell viability. In addition, forced
overexpression of LDHA to counteract the oxamate-induced depression
of LDHA levels rendered the cells resistant to the combination of
CTX and oxamate. The present study suggested that the inhibition of
glycolysis is a major mechanism of action of CTX on Ewing's
sarcoma. Further experiments are currently underway in our group to
explore the underlying mechanisms of CTX-mediated depression of
LDHA expression in Ewing's sarcoma cells in more detail.
Bioinformatics and proteomics tools will be utilized for the
identification of novel targets for overcoming CTX resistance.
Acknowledgments
The authors would like to thank the following
members staff and faculty members of the Department of Orthopedics,
Tianjin Third Central Hospital (Tianjin, China): Dr Yi Jiang for
providing the patient samples, and Dr Lianping Xiao and Dr Yonggang
Tian for their editorial assistance.
References
|
1
|
Iwamoto Y: Diagnosis and treatment of
Ewing's sarcoma. Jpn J Clin Oncol. 37:79–89. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Grier HE, Krailo MD, Tarbell NJ, Link MP,
Fryer CJ, Pritchard DJ, Gebhardt MC, Dickman PS, Perlman EJ, Meyers
PA, et al: Addition of ifosfamide and etoposide to standard
chemotherapy for Ewing's sarcoma and primitive neuroectodermal
tumor of bone. N Engl J Med. 348:694–701. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Miser JS, Krailo MD, Tarbell NJ, Link MP,
Fryer CJ, Pritchard DJ, Gebhardt MC, Dickman PS, Perlman EJ, Meyers
PA, et al: Treatment of metastatic Ewing's sarcoma or primitive
neuroectodermal tumor of bone: Evaluation of combination ifosfamide
and etoposide - a Children's cancer group and pediatric oncology
group study. J Clin Oncol. 22:2873–2876. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Longtin R: Ewing sarcoma: A miracle drug
waiting to happen? J Natl Cancer Inst. 95:1574–1576. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
May WA, Grigoryan RS, Keshelava N, Cabral
DJ, Christensen LL, Jenabi J, Ji L, Triche TJ, Lawlor ER and
Reynolds CP: Characterization and drug resistance patterns of
Ewing's sarcoma family tumor cell lines. PLoS One. 8:e800602013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fodale V, Pierobon M, Liotta L and
Petricoin E: Mechanism of cell adaptation: When and how do cancer
cells develop chemoresistance? Cancer J. 17:89–95. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Iida K, Fukushi J, Matsumoto Y, et al:
miR-125b develops chemo-resistance in Ewing sarcomaprimitive
neuroectodermal tumor. Cancer Cell Int. 13:212013. View Article : Google Scholar
|
|
8
|
Bardelli A and Jänne PA: The road to
resistance: EGFR mutation and cetuximab. Nat Med. 18:199–200. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yewale C, Baradia D, Vhora I, Patil S and
Misra A: Epidermal growth factor receptor targeting in cancer: A
review of trends and strategies. Biomaterials. 34:8690–8707. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Red Brewer M, Yun CH, Lai D, Lemmon MA,
Eck MJ and Pao W: Mechanism for activation of mutated epidermal
growth factor receptors in lung cancer. Proc Natl Acad Sci USA.
110:E3595–E3604. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Graham J, Muhsin M and Kirkpatrick P:
Cetuximab. Nat Rev Drug Discov. 3:549–550. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fantin VR, St-Pierre J and Leder P:
Attenuation of LDH-A expression uncovers a link between glycolysis,
mitochondrial physiology and tumor maintenance. Cancer Cell.
6:425–434. 2006. View Article : Google Scholar
|
|
13
|
Maftouh M, Avan A, Sciarrillo R, Granchi
C, Leon LG, Rani R, Funel N, Smid K, Honeywell R, Boggi U, et al:
Synergistic interaction of novel lactate dehydrogenase inhibitors
with gemcitabine against pancreatic cancer cells in hypoxia. Br J
Cancer. 110:172–182. 2014. View Article : Google Scholar :
|
|
14
|
Lu H, Li X, Luo Z, Liu J and Fan Z:
Cetuximab reverses the Warburg effect by inhibiting HIF-1-regulated
LDH-A. Mol Cancer Ther. 12:2187–2199. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
16
|
Rhodes DR, Yu J, Shanker K, Deshpande N,
Varambally R, Ghosh D, Barrette T, Pandey A and Chinnaiyan AM:
ONCOMINE: A cancer microarray database and integrated data-mining
platform. Neoplasia. 6:1–6. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Henderson SR, Guiliano D, Presneau N,
McLean S, Frow R, Vujovic S, Anderson J, Sebire N, Whelan J,
Athanasou N, et al: A molecular map of mesenchymal tumors. Genome
Biol. 206:R762005. View Article : Google Scholar
|
|
18
|
Zhou M, Zhao Y, Ding Y, Liu H, Liu Z,
Fodstad O, Riker AI, Kamarajugadda S, Lu J, Owen LB, et al: Warburg
effect in chemosensitivity: Targeting lactate dehydrogenase-A
re-sensitizes taxol-resistant cancer cells to taxol. Mol Cancer.
9:332010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Vander Heiden MG, Cantley LC and Thompson
CB: Understanding the Warburg effect: The metabolic requirements of
cell proliferation. Science. 324:1029–1033. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li X, Zhao H, Zhou X and Song L:
Inhibition of lactate dehydrogenase A by microRNA-34a resensitizes
colon cancer cells to 5-fluorouracil. Mol Med Rep. 11:577–582.
2015.
|