Introduction
Bone healing is characterized by different processes
consisting of the recruitment, migration, proliferation and
differentiation of repair cells, as well as autocrine and paracrine
mechanisms. Platelet-derived growth factor (PDGF) is a cell growth
factor that is released from platelet particles and presents near
to damaged tissues during the early stage of fracture healing.
PDGF, which a 30-kDa dimer of A and B polypeptide chains linked by
disulphide bonds, is critical in regulating osteoblast replication
and bone collagen degradation, controlling the proliferation of
repair cells, and inducing cartilage and bone formation (1). As a multiple mitogen, PDGF is
released by platelets and facilitates blood clotting via the
adhesion between platelets and blood vessels. Previous studies
demonstrated that PDGF was a stimulator for bone fracture healing
and was responsible for bone metabolism processes, including cell
proliferation, migration and apoptosis (2,3).
Furthermore, PDGF has three isoforms comprising of PDGF-AA, -BB and
-AB for bone metabolism and bone remodeling (3). Nash et al (4) demonstrated that the bone marrow
cavity volume and bone mineral density were markedly increased
after injecting recombinant human PDGF-BB into rabbits with tibial
fractures, indicating that PDGF participates in the stimulation of
fracture healing. In addition, cartilage cells cultured with
PDGF-BB and transforming growth factor β1 (TGF-β1) exhibited a
significantly higher proliferation rate in the study by Brandl
et al (3). Furthermore, the
findings of Lee et al (5)
and Schmidt et al (6)
support the hypothesis that PDGF promotes the proliferation of
cultured cartilage cells and repairs cartilage tissue.
G-protein-coupled receptor kinase interacting
protein-1 (GIT1) has been identified in various mammals and birds
(7). A yeast two-hybrid screen was
originally used to identify GIT1, which binds to G protein-coupled
receptor kinase (GRK2) for activation and membrane recruitment.
GIT1 was identified to be essential in focal adhesions, and cell
growth and migration, as it was located in the cytoplasmic membrane
and composite structures, such as inclusion bodies (8–11).
GIT1, a scaffold protein, has been demonstrated to act as a
GTPase-activating protein for the ADP-ribosylation factor family of
proteins in previous studies (8–11).
The results of recent studies have shown that GIT1 is important in
receptor endocytosis and cell motility (11,12).
Additionally, GIT1 is responsible for phosphorylation of
phospholipase Cγ (PLCγ1) and activation of extracellular
signal-regulated kinase (ERK)1/2 signaling induced by angiotensin
II and epidermal growth factor (12). Previously, it was found that GIT1
regulated cytoskeletal dynamics for cell growth and migration. For
example, cytoskeletal proteins, such as Paxillin, focal adhesion
kinase (FAK) and Src all participate in the promotion of cell
migration (13). Src,
characterized by protein tyrosine kinase activity, is one of many
signal transduction pathways and upstream core regulators, that is
important in cell proliferation, migration and differentiation in
addition to other mechanisms (13). It was identified that Src, acting
as a regulating kinase, is located at ERK1/2 upstream in the signal
transduction pathway of myocardial cells and nerve cells (14).
It is hypothesized that PDGF stimulates DNA
replication and protein synthesis of osteoblasts and chondrocytes,
and promotes osteoblast arrested in the G0/G1
phase to enter the S phase, inducing the cell division and
proliferation of osteoblasts (1).
Ren et al (15) revealed
that phosphorylation of GIT1 tyrosine 321 is critical in order to
associate with FAK and activate FAK in focal adhesions to promote
PDGF-induced osteoblastic cell migration. In addition, Rui et
al (9) indicated that the
phosphorylation of GIT1Y321 is critical for ERK1/2- and
PDGF-dependent vascular endothelial growth factor (VEGF) secretion
from osteoblasts for the purpose of angiogenesis and bone healing
stimulation. Thus, PDGF is important in the activation of GIT1
during bone repair. PDGF may promote osteoblast migration through
regulating cell division control protein 42 homolog/Rac and Src
activity in the phosphatidylinositol kinase signaling pathway
(16). The activation of Src and
its associated family members requires phosphorylation of tyrosine
residues located in the membrane proximal region of the PDGF
receptor (PDGFR). Src may undergo conformational reorganization of
the Src kinase domain through the connection between its SH2 domain
and the phosphorylated PDGFR in an attempt to activate Src
(17). Furthermore, Src-induced
formation of the calpain/FAK/ERK1/2 complex and ERK1/2 activation
are required for calpain-mediated proteolysis of FAK (17). Therefore, PDGF may mediate GIT1 via
the regulation of Src.
With regard to GIT1 and PDGF in bone cells, there
has been less research into the effects of GIT1 and PDGF on cell
proliferation and cell apoptosis in chondrocytes compared with cell
migration in osteoblasts. PDGF may activate specific tyrosine
kinases containing PDGF-α and -β when the corresponding receptors
are present on the cell membrane, contributing to increased
mitogenic signals in the signal transduction pathway and leading to
DNA synthesis, cell division and proliferation induction (18). Furthermore, PDGF has been
demonstrated to be critical in the ERK1/2 signaling pathway. Thus,
PDGF may activate the cytokine-mediated ERK1/2 signaling pathway to
regulate the typical early signaling pathways associated with cell
proliferation, differentiation and apoptosis.
Materials and methods
Cell culture
Animal care and use followed the ethical guidelines
of the Chinese Council on Animal Care and were reviewed and
approved by the Institutional Animal Care and Use Committee of Qilu
Hospital of Shandong University (Jinan, China). Three one-week-old
Sprague-Dawley neonatal rats were purchased from the Institute of
Laboratory Animal Sciences, Chinese Academy of Medical Sciences and
Peking Union Medical College (Beijing, China). The weight of the
rats was 300–350 g. The rats were sacrificed by an intraperitoneal
injection of euthatal solution (150 mg/kg; Sigma-Aldrich, St.
Louis, MO, USA) and then dissected aseptically to obtain all the
limb joints. The fibrous tissue was removed from the articular
cartilage surface of the limb joints and the transparent central
portion of cartilage was sliced into small sections (size, ~1
mm3). The small tissue sections were digested using
0.25% trypsin (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) at 37°C for 30 min. The mixture was centrifuged at 1,000 × g
for 5 min to obtain cell pellets. The pellets were combined with
0.2% II collagenase (Sigma-Aldrich) and incubated at 37°C for 4 h
for digestion. A 200-mesh filter (Anping County Sheng Silk Factory,
Tainan, China) was used and a further centrifugation at 1,000 × g
for 5 min was conducted to collect the cells. Invitrogen Dulbecco's
modified Eagle's medium (DMEM; Thermo Fisher Scientific, Inc.)
containing Gibco 10% fetal bovine serum (FBS; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin (Gibco; Thermo Fisher
Scientific, Inc.) and 50 U/ml streptomycin (Gibco; Thermo Fisher
Scientific, Inc.) was used for maintaining the cells, which were
incubated at 37°C in a humidified incubator with a 5%
CO2 atmosphere. After repeated isolation and culture,
the morphology of the purified cells was observed under an inverted
phase contrast microscope (CKX41; Olympus Corporation, Tokyo,
Japan).
Transfection of siRNA
The siRNA transfection was conducted using
Invitrogen Lipofectamine 2000 (Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. Briefly, the cells
was seeded into 6-well plates (1×105 cells/ml) and
incubated for 24 h at 37°C. The primers, designed as previously
described (9) and synthesized by
Ambion (Thermo Fisher Scientific, Inc.) were as follows: Sense
5′-AAGCTGCCAAGAAGAAGCTAC-3′ for GIT1 siRNA and sense
5′-AATTCTCCGACACGTGTCACT-3′ for the negative control siRNA. The
GIT1 siRNA (100 nM) was prepared and transfected for 24 h as
previously described (10). The
Src inhibitor, PP2 (10 μM; CST Biological Reagents Co.,
Ltd., Shanghai, China) dissolved in 100% dimethyl sulfoxide (DMSO;
Enzo Life Sciences, Inc., Farmingdale, NY, USA) was used to
pretreat the cells for 1 h. The same volume of 0.1% DMSO served as
a control. PDGF (10 ng/ml; Sigma-Aldrich) was added for 24 h to
stimulate the cells. After 72 h, the transfected cells were
analyzed by quantitative polymerase chain reaction (qPCR) and
western blot analysis.
Fluorescence qPCR
The cells were collected and total RNA was extracted
using 1 ml Trizol (Invitrogen; Thermo Fisher Scientific, Inc.).
Total RNA (1 μg) was used for first-strand cDNA synthesis
with RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher
Scientific, Inc., Pittsburgh, PA, USA). First-strand cDNA was
synthesized using 1 μl of random hexamers and adjusted to a
volume of 12 μl using DEPC-treated water. The mix was
incubated at 65°C for 5 min. The following components were added in
to the reaction: 4 μl 5X Reaction buffer; 1 μl
Ribolock RNase inhibitor (20 U/μl), 2 μl dNTP (10
mM); and 1 μl RevertAid M-MuLV RT (200 U/μl). The
reactions were incubated at 25, 42 and 70°C for 5, 60 and 5 min,
respectively. Then, the cDNA was used to measure of mRNA levels by
qPCR using the SYBR-Green dye (Toyobo Co., Ltd., Osaka, Japan)
method with 100 ng cDNA in a 20-μl system. The primer
sequences were as follows: Forward, 5′-ATGGATGTGTATGACGAAGTG-3′ and
reverse, 5′-GGTTGACTGGCAGGAAGG-3′ for GIT1; forward,
5′-GTCCACCTTCCAGCAGATG-3′ and reverse, 5′-CTCAGTAACAGTCCGCCTAG-3′
for β-actin, which was used as a reference gene. The reaction
conditions were as follows: 95°C for 5 min; followed by 40 cycles
of 95°C for 30 sec, 55°C for 30 sec and 72°C for 30 sec. qPCR was
performed using the Applied Biosystems 7500 Real-Time PCR system
(Thermo Fisher Scientific, Inc.) All reactions were run in
triplicate and performed 3 times. Quantitative analysis was
performed by comparison of the 2−ΔΔCq values (19).
Western blot analysis
Total cellular proteins were extracted from the
cells using lysis buffer (Beyotime Institute of Biotechnology,
Haimen, China). The solution containing cells and lysis buffer was
agitated at 4°C for 20 min for mixing. The solution was centrifuged
at 10,000 × g at 4°C for 10 min to obtain the supernatant. The
protein concentration in the cell lysates was determined using a
Bradford protein assay (Pierce Biotechnology, Inc., Rockford, IL,
USA). SDS-PAGE containing 10% tricine gels and 20 μg of
protein was loaded in each lane. The proteins were separated by
electrophoresis (40 V; 5 h) and transferred onto nitrocellulose
membranes (Pierce Biotechnology, Inc.). The membranes were
incubated in a blocking buffer consisting of 5% non-fat milk
(Pierce Biotechnology, Inc.) and Tris-buffered saline with Tween-20
[TBST; Sunshine Biotechnology (Nanjing) Co., Ltd., Nanjing, China]
for 1 h. The membranes were incubated with polyclonal rabbit
anti-GIT1 (1:600; cat. no. 2919; Cell Signaling Technology Inc.,
Danvers, MA, USA), polyclonal rabbit anti-Src antibody (1:800; cat.
no. 2108; CST), polyclonal rabbit anti-p-Src (1:800; cat. no. 2105;
Cell Signaling Technology Inc.) and monoclonal rabbit anti-β-actin
(1:1,000; cat. no. 4970; Cell Signaling Technology Inc.) antibodies
in 5% non-fat milk overnight at 4°C. The membranes were washed with
TBST every 10 min, for 30 min. Subsequently, goat anti-rabbit
horseradish peroxidase-conjugated secondary antibody (cat. no.
7074; Cell Signaling Technology, Inc.) at 1:7,000-8,000 dilution
was incubated with the membranes for 1 h at room temperature. After
washing with TBST every 10 min for 30 min, the protein bands were
detected using ECL Western Blotting Substrate (Pierce
Biotechnology, Inc.). The western blots were scanned using a gel
imaging system (UVP, LLC, Upland, CA, USA) and the gray values of
the bands were measured using Gel-Pro Analyzer software (version
6.3; Media Cybernetics, Inc., Rockville, MD, USA).
5-bromo-2′-deoxyuridine (BrdU) assay of
cell proliferation
To examine the roles of PDGF, Src, GIT1 and the
apoptotic pathway in cell proliferation, a BrdU assay was conducted
to investigate the effects of different inhibitors on cell
proliferation. Briefly, the cells were seeded into 6-well plates
(1×105 cells/ml), incubated for 24 h at 37°C and siRNA
transfection was conducted using Lipofectamine 2000. After
pretreating the cells with 30 μM PP2, or 30 μM 0.1%
DMSO for 1 h, 10 ng/ml PDGF was added to stimulate phosphorylation
for 24 h. Cell proliferation was quantified using a BrdU
colorimetric immunoassay kit (Cell Proliferation ELISA kit; Roche
Diagnostics GmbH, Mannheim, Germany) according to the
manufacturer's protocol. Cell proliferation was expressed as the
mean percentage of the control values (set at 100%).
Terminal deoxynucleotidyl transferase
dUTP nick end labeling (TUNEL)-DAPI co-staining assay
A TUNEL-DAPI co-staining assay was performed
to analyze the apoptotic cells using an In Situ cell Death
Detection kit (Roche Diagnostics GmbH) according to the
manufacturer's instructions. Briefly, 50 ml 4% formaldehyde
(Sigma-Aldrich) was used for fixation at 4°C for 25 min, then the
cells were washed twice with phosphate-buffered saline (PBS).
Subsequently, 50 ml 0.2% Triton X-100 (Sigma-Aldrich) was then
added to cells for a 5-min incubation at room temperature. The
cells were subsequently mixed with 100 μl equilibration
buffer (Roche Diagnostics GmbH) at room temperature for 10 min. The
cells were washed with 2 ml saline sodium citrate (Beijing
Kangpuhuiwei Technology Co., Ltd., Beijing, China) for 15 min
following incubation with 50 μl TUNEL reaction mixture
(Roche Diagnostics GmbH) containing 5 μl nucleotide mixture
(Roche Diagnostics GmbH) and 1 μl terminal deoxynucleotidyl
transferase (Roche Diagnostics GmbH) for 60 min at 37°C.
Furthermore, the cells were incubated with 50ml 0.3%
H2O2 (Sigma-Aldrich) for 10 min and 100
μl streptavidin working solution (Roche Diagnostics GmbH)
for 30 min at room temperature. The cells were incubated with 0.5
μg/ml DAPI in a humidified chamber in the dark for 5 min at
room temperature. A fluorescence microscope (Nikon Eclipse 80i;
Nikon Corporation, Tokyo, Japan) was used to examine and obtain
images of the stained cells.
Examination of migration ability by
Transwell migration assay
The Transwell migration assay is used to examine the
migratory response of cells to angiogenic inducers or inhibitors.
Determination of cell migration rates was conducted in a Transwell
chamber (BD Biosciences, San Jose, CA, USA). DMEM (0.6 ml) with
0.5% FBS was added to the lower chamber. The cells were diluted to
2×106 cells/ml in FBS-free DMEM medium following
trypsinization. Cells (100 μl) were added to the upper
chamber of filters precoated with Matrigel and incubated in a 5%
CO2 atmosphere at 37°C for 6 h. After removing the
medium, 4% glutaraldehyde (Sigma-Aldrich) was used to fix the cells
on the lower side of the insert filter for 15 min. Subsequently, a
cotton swab was used to remove the remaining cells on the upper
side of the filter membrane. The cells on the lower side of the
insert filter were stained with 0.1% crystal violet (Sigma-Aldrich)
for 10 min. Finally, the number of cells on the lower side of the
filter was counted under a microscope after washing with PBS. The
cells that had migrated through the membrane were stained and
counted, therefore, the migration of chondrocytes was expressed by
the relative number of migrated cells.
Statistical analysis
The experiments were performed in triplicate, at
minimum, and results were expressed as means ± standard deviation.
SPSS statistical package 13.0 (SPSS, Inc., Chicago, IL, USA) was
used for statistical analysis. The difference between two groups
was analyzed by two-tailed Student's t-test and differences
between three or more groups were analyzed by one-way analysis of
variance and multiple comparisons. P<0.05 was considered to
indicate a statistically significant difference.
Results
PDGF induced Src phosphorylation via
upregulation of GIT1 expression
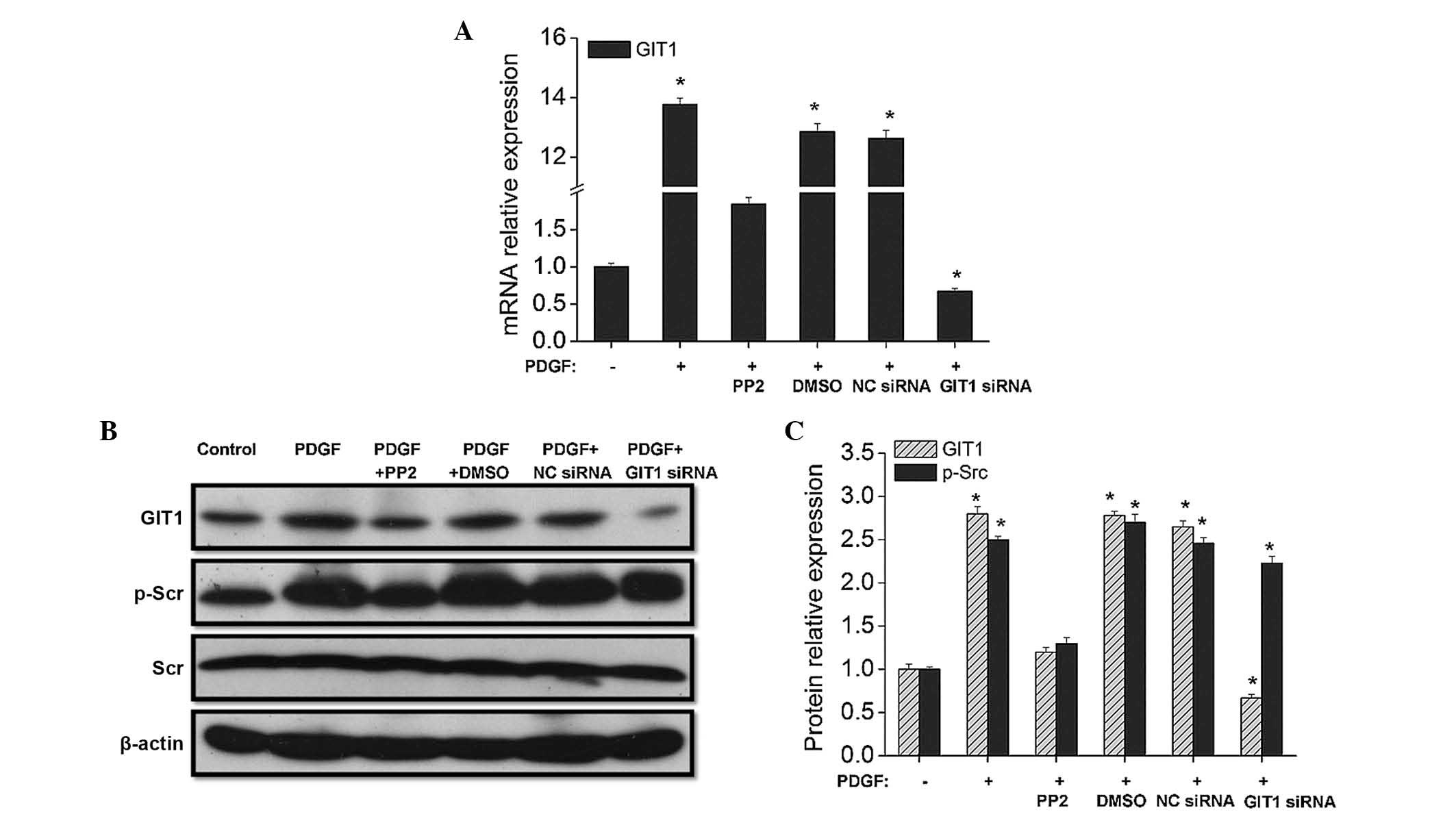
In the present study, the roles of PDGF, GIT1 and
Src were analyzed by siRNA silencing techniques and western blot
analysis (Fig. 1). The western
blot analysis indicated that PDGF promotes Src phosphorylation,
while upregulating GIT1 expression. The results revealed that PDGF
increased the expression level of GIT1 2.8-fold, thereby promoting
a 2.5-fold increase in Src phosphorylation (Fig. 1B). Although PDGF was combined with
cartilage cells for stimulation, the GIT1 expression level reduced
to 60% of the control when GIT1 was knocked down by siRNA. However,
the effects of PDGF on GIT1 overexpression were suppressed after
adding the Src inhibitor, PP2. As shown in Fig. 1, the effect of PDGF on promotion of
Src phosphorylation was not statistically significant, however, the
expression of GIT1 was inhibited by knock down of GIT1 by siRNA.
Therefore, the results demonstrate that PDGF initially activates
Src phosphorylation, which in turn increases the level of GIT1
expression, suggesting that Src is upstream of GIT1.
PDGF promotes cell proliferation via
regulation of GIT1 and Src
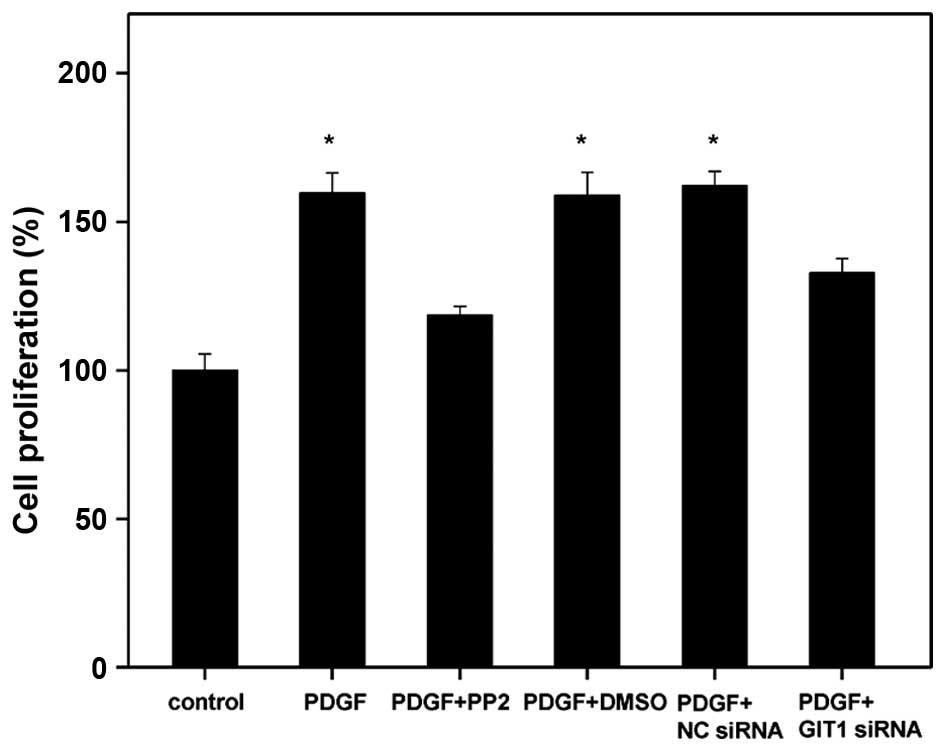
In order to investigate the biological functional
roles of Src, GIT1 and PDGF in cartilage cells, the effects of
their siRNA and specific inhibitors on the PDGF-activated cell
proliferation were further investigated in the present study. A
BrdU assay was used to analyze the cell proliferation of
chondrocytes. The results demonstrated that PDGF stimulated
chondrocyte proliferation by regulating GIT1 and Src (Fig. 2). According to Fig. 2, the cell proliferation of
chondrocytes was significantly suppressed by the Src inhibitor,
PP2. Indicating that Src is key in chondrocyte proliferation. Since
Src is an upstream core regulator in various signal transduction
pathways, the addition of Src inhibitor, PP2, decreased the cell
proliferation ability to a greater extent than cells treated with
GIT1 siRNA. However, although GIT1 expression was downregulated,
cell proliferation was still greater than that of the control
group. Therefore, there may be certain additional signaling
pathways that involve the induction of chondrocyte proliferation by
PDGF. Furthermore, the chondrocyte protein levels were greater than
those of the negative control in the experiments (Fig. 1B and C), which may indicate that
PDGF affects chondrocyte proliferation by mediating other proteins
whilst inhibiting Src or GIT1.
PDGF affects cell apoptosis via GIT1 and
Src regulation
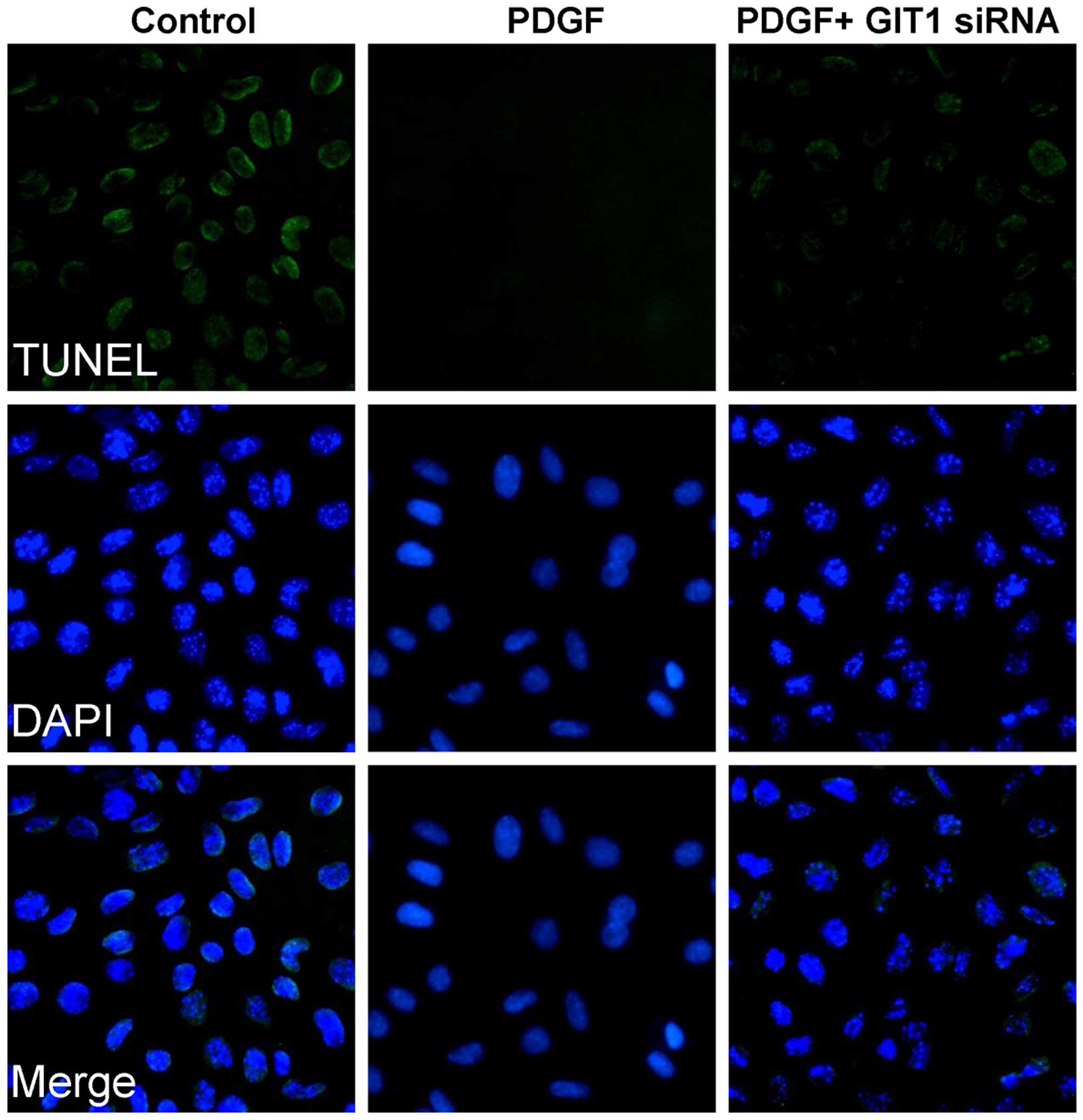
Apoptosis is an important process in programmed cell
death that occurs in multicellular organisms. In an attempt to
examine DNA fragmentation resulting from apoptotic signaling
cascades, the TUNEL-DAPI assay was used to conduct further
analyses. The TUNEL assay was used to investigate the role of GIT1
in chondrocyte apoptosis depending on the presence of identifiable
nicks in the DNA. As shown in Fig.
3, chondrocytes demonstrated DNA fragmentation, which was
evidenced by apoptotic cell death. The results showed that the
presence of PDGF markedly decreased the number of apoptotic cells.
Furthermore, siRNA silencing of GIT1 resulted in the reversal of
the protective effects of PDGF. Therefore, these results
demonstrated that GIT1 is key in PDGF-induced chondrocyte
survival.
PDGF promotes chondrocyte migration
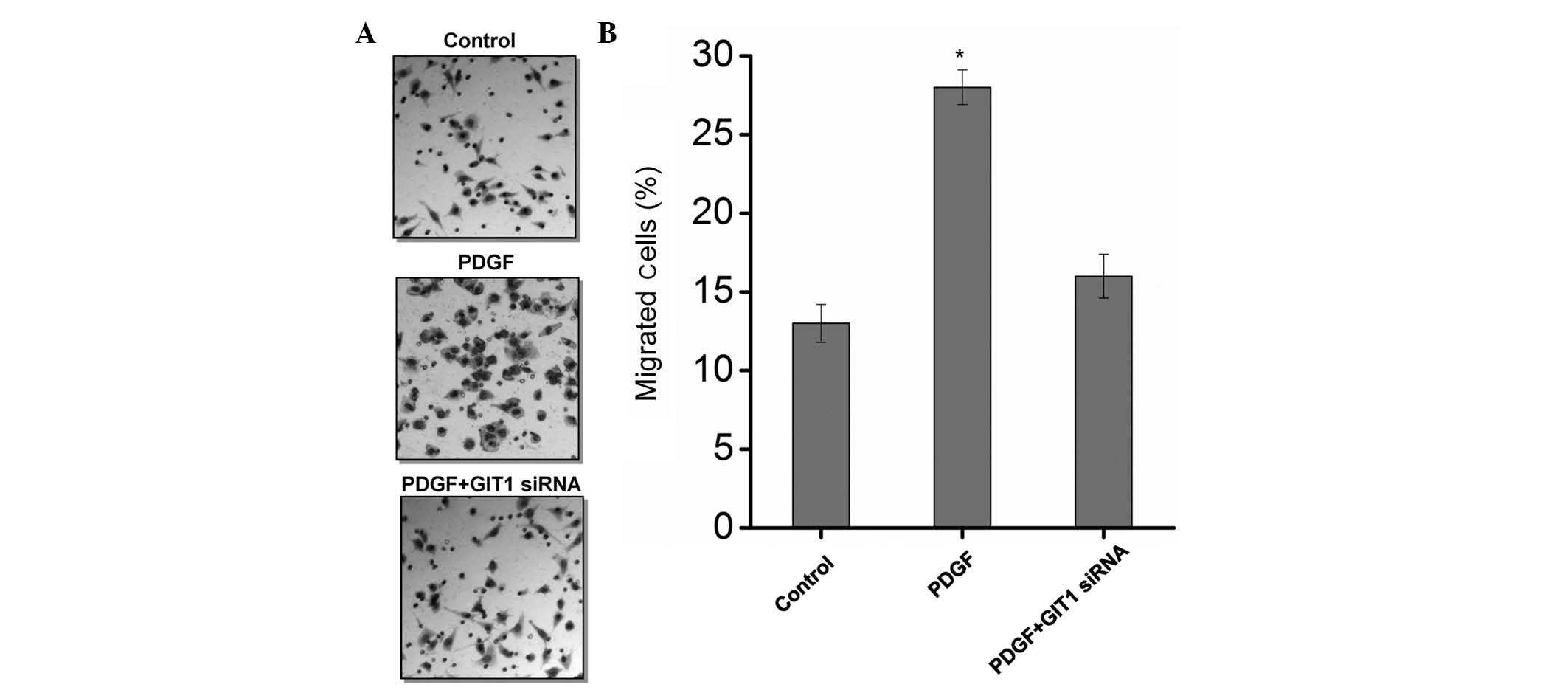
The cell migration ability of cartilage cells
induced by PDGF and GIT1 was detected by Transwell assay. The
results revealed that PDGF induced chondrocyte migration ability
(Fig. 4). When GIT1 was knocked
down using siRNA, the effect of PDGF on the migration ability of
cartilage cells was significantly suppressed (Fig. 4). This finding indicated that GIT1
is critical in chondrocyte migration.
Discussion
Local growth factors in bones and cytokines are
critical in the processes of fibrocartilage, cartilaginous and bony
callus formation for fracture healing following bone tissue damage
(1–4). Previous research has demonstrated
that various growth factors promote bone healing with synergistic
effects (5,6). PDGF, a growth factor (associated with
healing damaged tissue) is released following bone fracture and is
considered to be essential growth factor for processes, including
osteogenesis and bone remodeling (1–4).
Previous studies demonstrated that PDGF promotes bone cell
differentiation, cell proliferation and regulation of bone cells,
and participates in bone formation (20). PDGF is essential in promotion of
bone formation and bone remodeling by interacting with a variety of
cells during fracture healing. Furthermore, it has been
demonstrated that PDGF collaborates with other factors, such as
NGF, to promote fracture healing (21). The aim of the present study was to
examine the effects of PDGF on chondrocyte proliferation and
apoptosis via regulation of GIT1 and Src.
Differentiation, proliferation, secretory function
and apoptosis of chondrocytes have been regarded as critical steps
for the development, growth and remodeling of bones (22). PDGF, as well as PDGF mRNA, are
apparent at the early stage of callus formation in cartilage,
whereas the expression of PDGF becomes weak or disappears in the
latter hypertrophy of mature chondrocytes (23). Therefore, it is essential to
investigate the role of PDGF and its underlying mechanisms in
cartilage cells. In the present study, the results indicated that
PDGF promotes chondrocyte proliferation and migration, and
suppresses apoptosis. Brandl et al (3) demonstrated that there was a markedly
higher proliferation rate of chondrocytes cultured with PDGF-BB and
TGF-β1 than that of control cells. Schmidt et al (6) found that PDGF promoted chondrocyte
proliferation and proteoglycan synthesis. Furthermore, the role of
ectopic cartilage formation was promoted, endochondral bone
formation was inhibited and chondrocyte phenotype stability was
maintained without mechanical stimulation by PDGF in the
experiments (6). In addition, it
has been reported that PDGF is able to stimulate cultured
chondrocytes and restore cartilage tissue proliferation (3–6).
A potential mechanism underlying the effects of PDGF
on the proliferation, migration and apoptosis of cartilage cells is
that PDGF mediates the induction of Src phosphorylation by
upregulating the expression GIT1. Western blotting demonstrated
that PDGF promoted Src phosphorylation, while upregulating GIT1
expression. Furthermore, the effect of PDGF on GIT1 overexpression
was significantly inhibited by the presence of Src inhibitor, PP2.
However, knock down of GIT1 expression using siRNA did not
influence the effect of PDGF on Src phosphorylation induction.
These results demonstrated that PDGF activated the phosphorylation
of Src first, and subsequently increased GIT1 expression,
indicating that Src is upstream of GIT1. Additionally, after using
the Src inhibitor, PP2, the effect of PDGF on induction of
chondrocyte proliferation was markedly decreased. Additionally,
knocking down GIT1 using siRNA showed a similar result that was
comparable to that of PP2. Src, one of the most important members
of the Src family, is characterized by tyrosine kinase activity.
The phosphorylation of Src-Tyr418 in the Src protein
kinase activation loop triggers kinase activation; activated Src
results in the transfer of a phosphate group to the target
molecules. As a result, the target molecule phosphorylates and
transmits the signal to the nucleus, which eventually promotes
mechanisms, including cell proliferation, migration and
differentiation (23,24). The findings of Ren et al
(15) indicated that periodic
mechanical stress activated the Src-PLCγ1-mitogen-activated protein
kinase kinase (MEK)1/2-ERK1/2 signaling pathway to induce
chondrocyte proliferation and matrix synthesis. In addition, the
study of Ren et al (25)
demonstrated that the Src-PLCγ1-MEK1/2 signaling pathway involved
chondrocyte area expansion and migration using periodic mechanical
stress.
GIT1, present in mammals and birds, is a multidomain
scaffold protein that participates in the internalization and
membrane transportation of G protein-coupled receptors (7–11).
GIT1 is essential in cell proliferation and migration, and is
located in the cytoplasmic membrane of focal adhesions and
composite structures, such as inclusion bodies of cells in
vivo (8–11). Yin et al (26) indicated that the phosphorylation of
GIT1 was enhanced in the presence of Src by angiotensin II and EGF.
In the present study, GIT1 was observed to be critical for
chondrocyte proliferation, migration and apoptosis via Src
regulation induced by PDGF. Furthermore, Menon et al
(27) found that GIT1 is a key
regulator of receptor activator of nuclear factor-κB signaling,
comparable with Src-mediated tyrosine phosphorylation and
phosphorylation of PLCγ1, and regulates osteoclast function and
bone mass. However, recent studies have focused on the effects of
GIT1 on cell migration in osteoblasts rather than chondrocytes. For
example, Ren et al (15)
demonstrated that GIT1 tyrosine 321 phosphorylation is required for
association with FAK, FAK activation in focal adhesions and for
PDGF-induced osteoblastic cell migration. In the present study,
downregulation of GIT1 was identified to inhibit the effects of
PDGF on promotion of chondrocyte proliferation and migration, and
inhibition of apoptosis in cartilage cells, indicating that GIT1
plays a similar role to PDGF in the promotion of chondrocyte
proliferation and migration, and inhibition of apoptosis.
In conclusion, PDGF may promote chondrocyte
proliferation and migration, and inhibit apoptotic cell death by
upregulation of GIT1 expression and Src phosphorylation. However,
the underlying mechanisms of bone healing are complex and
intertwined and the results of the present study indicated that
multiple signaling molecules are involved. Thus, further
investigation is required into the role of PDGF in other signaling
pathways.
References
|
1
|
Yang D, Chen J, Jing Z and Jin D:
Platelet-derived growth factor (PDGF)-AA: A self-imposed cytokine
in the proliferation of human fetal osteoblasts. Cytokine.
12:1271–1274. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Filardo G, Kon E, Di Martino A, Iacono F
and Marcacci M: Arthroscopic second-generation autologous
chondrocyte implantation: A prospective 7-year follow-up study. Am
J Sports Med. 39:2153–2160. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Brandl A, Angele P, Roll C, Prantl L,
Kujat R and Kinner B: Influence of the growth factors PDGF-BB,
TGF-beta1 and bFGF on the replicative aging of human articular
chondrocytes during in vitro expansion. J Orthop Res. 28:354–360.
2010.
|
|
4
|
Nash TJ, Howlett CR, Martin C, Steele J,
Johnson KA and Hicklin DJ: Effect of platelet-derived growth factor
on tibial orteotomies in rabbits. Bone. 15:203–208. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lee JY, Nam SH, Im SY, Park YJ, Lee YM,
Seol YJ, Chung CP and Lee SJ: Enhanced bone formation by controlled
growth factor delivery from chitosan-based biomaterials. J Control
Release. 78:187–197. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schmidt MB, Chen EH and Lynch SE: A review
of the effects of insulin-like growth factor and platelet derived
growth factor on in vivo cartilage healing and repair.
Osteoarthritis Cartilage. 14:403–412. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Manabe R, Kovalenko M, Webb DJ and Horwitz
AR: GIT1 function sin a motile, multi-molecular signaling complex
that regulates protrusive activity and cell migration. J Cell Sci.
115:1497–1510. 2002.PubMed/NCBI
|
|
8
|
Premont RT, Claing A, Vitale N, Freeman
JL, Pitcher JA, Patton WA, Moss J, Vaughan M and Lefkowitz RJ:
Beta2-Adrenergic receptor regulation by GIT1, a GIT1, a G protein
coupled receptor kinase-associated ADP ribosylation factor and
GTPase-activating protein. Proc Natl Acad Sci USA. 95:14082–14087.
1998. View Article : Google Scholar
|
|
9
|
Rui Z, Li X, Fan J, Ren Y, Yuan Y, Hua Z,
Zhang N and Yin G: GIT1Y321 phosphorylation is required for ERK1/2-
and PDGF-dependent VEGF secretion from osteoblasts to promote
angiogenesis and bone healing. Int J Mol Med. 30:819–825.
2012.PubMed/NCBI
|
|
10
|
Schmalzigaug R, Phee H, Davidson CE, Weiss
A and Premont RT: Differential expression of the ARF GAP genes GIT1
and GIT2 in mouse tissues. J Histochem Cytochem. 55:1039–1048.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Claing A, Perry SJ, Achiriloaie M, Walker
JK, Albanesi JP, Lefkowitz RJ and Premont RT: Multiple endocytic
pathways of G protein-coupled receptors delineated by GIT1
sensitivity. Proc Natl Acad Sci USA. 97:1119–1124. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang H, Webb DJ, Asmussen H and Horwitz
AF: Synapse formation is regulated by the signaling adaptor GITI. J
Cell Biol. 161:131–142. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chaturvedi LS, Marsh HM and Basson MD: Src
and focal adhesion kinase mediate mechanical strain-induced
proliferation and ERK1/2 phosphorylation in human H441 pulmonary
epithelial cells. Am J Physiol Cell Physiol. 292:C1701–C1713. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shen E, Fan J, Chen R, Yee SP and Peng T:
Phospholipase Cgamma1 signalling regulates
lipopolysaccharide-induced cyclooxygenase-2 expression in
cardiomyocytes. J Mol Cell Cardiol. 43:308–318. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ren Y, Yu L, Fan J, Rui Z, Hua Z, Zhang Z,
Zhang N and Yin G: Phosphorylation of GIT1 tyrosine 321 is required
for association with FAK at focal adhesions and for PDGF-activated
migration of osteoblasts. Mol Cell Biochem. 365:109–118. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Golden LH and Insogna KL: The expanding
role of PI3-kinase in bone. Bone. 34:3–12. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fredriksson L, Li H and Eriksson U: The
PDGF family: Four gene products form five dimeric isoforms.
Cytokine Growth Factor Rev. 15:197–204. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Claesson-Welesh L: Platelet-derived growth
factor receptor signals. J Biol Chem. 269:32023–32026. 1994.
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCt method. Methods. 25:402–408. 2001. View Article : Google Scholar
|
|
20
|
Marcopoulou CE, Vavouraki HN, Dereka XE
and Vrotsos IA: Proliferative effect of growth factors TGF-betal,
PDGF-BB and rhBMP-2 on human gingival fibroblasts and periodontal
ligament cells. J Int Acad Periodontol. 5:63–70. 2003.PubMed/NCBI
|
|
21
|
Oreffo RO: Growth factors for skeletal
reconstruction and fracture repair. Curr Opin Investig Drugs.
5:419–423. 2004.PubMed/NCBI
|
|
22
|
Adams CS and Shapiro IM: The fate of the
terminally differentiated chondrocyte: Evidence for
microenvironmental regulation of chondrocyte apoptosis. Crit Rev
Oral Biol Med. 13:465–473. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chaturvedi LS, Gayer CP, Marsh HM and
Basson MD: Repetitive deformation activates Src-independent
FAK-dependent ERK motogenic signals in human Caco-2 intestinal
epithelial cells. Am J Physiol Cell Physiol. 294:C1350–C1361. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Morita N, Iizuka K, Murakami T and
Kawaguchi H: N-terminal kinase and c-Src are activated in human
aortic smooth muscle cells by pressure stress. Mol Cell Biochem.
262:71–78. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ren K, Ma Y, Huang Y, Liang W, Liu F, Wang
Q, Cui W, Liu Z, Yin G and Fan W: Periodic mechanical stress
activates MEK1/2-ERK1/2 mitogenic signals in rat chondrocytes
through Src and PLCγ1. Braz J Med Biol Res. 44:1231–1242. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yin G, Zheng Q, Yan C and Berk BC: GIT1 is
a scaffold for ERK1/2 activation in focal adhesions. J Biol Chem.
280:27705–27712. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Menon P, Yin G, Smolock EM, Zuscik MJ, Yan
C and Berk BC: GPCR kinase 2 interacting protein 1 (GIT1) regulates
osteoclast function and bone mass. J Cell Physiol. 225:777–785.
2010. View Article : Google Scholar : PubMed/NCBI
|