Introduction
Postoperative cognitive dysfunction (POCD) is one of
the most common postoperative complications in elderly patients
(1). A previous study demonstrated
that, in patients >60 years of age that had undergone major
non-cardiac surgery, 25.8% developed cognitive dysfunction after 1
week and 9.9% developed cognitive dysfunction after 3 months
(2). In addition, a significant
association between early and late-onset POCD and advancing age was
observed. POCD is associated with impairment of daily functional
activities, an increased risk of dismissal from employment, as well
as increased expenses, and risk of morbidity and mortality
(2). However, the etiology and
pathogenesis of POCD remain elusive.
Neuroinflammation, which involves the activation of
microglia, astrocytes and neurons (3), as well as an elevation in the
production of proinflammatory cytokines (4), has been reported to serve a role in
the pathogenesis of neurodegenerative diseases. Although
neuroinflammation is known to be involved in the pathogenesis of
POCD (5), multiple perioperative
factors, including surgical trauma (6) and inhaled anesthetics (7), may facilitate this process. Nuclear
factor-κB (NF-κB) is activated as a result of canonical and
non-canonical signaling pathways, and has been reported to mediate
neuroinflammation in multiple pathological processes, such as
Alzheimer's disease (AD) (8) and
cerebral ischemia reperfusion injury (9). In a previous study, the canonical
NF-κB pathway was observed to be involved in isoflurane-induced
neuroinflammation (5); however,
the upstream regulatory factors involved in this signaling pathway
remain elusive and impede further studies.
Deficiencies in mitochondrial respiratory chain
components, the generation of reactive oxygen species (ROS) and
mitochondrial DNA mutations result in mitochondrial membrane
potential (ΔΨm) loss, and may induce mitochondrial retrograde
signaling (10). This signaling
pathway is an adaptive mechanism that transmits signals from
dysfunctional mitochondria to activate nuclear gene expression.
Zhang et al (11) reported
that inhaled anesthetics induce ROS generation and mitochondrial
permeability transition pore (mPTP) opening, and reduce ΔΨm and ATP
production in cells in vitro. Therefore, inhaled anesthetics
may initiate mitochondrial retrograde signaling. ΔΨm initiates
mitochondrial Ca2+ uptake (12), and its loss leads to cytosolic
Ca2+ elevation and the activation of calcineurin (CaN).
Ca2+/CaN signaling serves an important role in
mitochondrial retrograde signaling (13), and NF-κB is also involved in the
process (14). Therefore, inhaled
anesthetics may activate CaN signaling, which may subsequently
facilitate multiple pathological processes, such as
neuroinflammation.
In the present study, an anesthesia model in aged
rats was established using the inhaled anesthetic, isoflurane. This
study aimed to elucidate the role of mitochondrial retrograde
signaling in isoflurane-induced hippocampal neuroinflammation and
cognitive impairment in aged rats, and to explore the effects of
CaN and NF-κB on neuroinflammation and cognitive impairment, in
order to provide potential therapeutic targets for the prevention
and treatment of POCD.
Materials and methods
Animals
A total of 102 male Sprague-Dawley rats (age, 18
months; weight, 550–600 g; obtained from Dongchuang Laboratory
Animal Center, Changsha, China) were used in the present study.
Rats were maintained on a standard pellet food and water ad
libitum, a with a 12:12 h light-dark cycle, and a temperature
of 24±2°C for 2 weeks prior to isoflurane exposure.
Study protocol
The study protocol was approved by the Peking
University Biomedical Ethics Committee, Experimental Animal Ethics
Branch (approval no. LA201412). Rats were randomly divided into
isoflurane (n=12) or control groups (n=6), and received isoflurane
(Baxter International, Inc., Deerfield, IL, USA) or vehicle gas
(50% O2 and 50% air), respectively. CaN and
interleukin-1β (IL-1β) expression levels, and NF-κB activation,
including inhibitor of NF-κB (IκBα) phosphorylation and NF-κB RelA
(also known as p65) nuclear translocation in the hippocampus were
assessed at 6 or 12 h after isoflurane exposure.
Rats were treated with the CaN inhibitor cyclosporin
A (CsA) (15) and the NF-κB
inhibitor pyrrolidine dithiocarbamate (PDTC) (16) to investigate the role and
interaction of CaN and NF-κB in isoflurane-induced
neuroinflammation. To achieve this, rats were randomly divided into
control, CsA, PDTC, isoflurane, isoflurane + CsA or isoflurane +
PDTC groups (n=6 for each group). Rats in CsA and isoflurane + CsA
groups received intraperitoneal CsA (7 mg/kg; Abcam, Cambridge,
UK), and rats in the PDTC and isoflurane + PDTC groups received 100
mg/kg intraperitoneal PDTC (Sigma-Aldrich, St. Louis, MO) in 0.9%
saline (total volume 0.5 ml) at 30 min prior to isoflurane
exposure. Rats in the control and isoflurane groups received an
identical volume of 0.9% saline delivered by intraperitoneal
injection. Hippocampal CaN and IL-1β expression levels, and IκBα
phosphorylation were then assessed, and the spatial learning and
memory function of the rats were evaluated.
Isoflurane exposure
Isoflurane exposure was performed according to the
methods described previously (17). Briefly, rats received 2% isoflurane
for 4 h in an anesthetic chamber. During this time, the arterial
oxygen saturation, heart rate, blood pressure and rectal
temperature were monitored to ensure that they were maintained
within the physiological range. The rats then received 100% oxygen
until they regained consciousness. Isoflurane was well tolerated,
with all monitored variables remaining within the physiological
range. Rats in the control, CsA and PDTC groups only received
vehicle gas.
ROS assay
An ROS assay kit (Cell Biolabs, Inc., San Diego, CA,
USA) was used to determine ROS levels in the rat hippocampus.
Briefly, rats were sacrificed by decapitation at the end of the
experiments. The brain tissues were removed rapidly, and
hippocampus was dissected out on ice for subsequent experiments.
Subsequently, the hippocampus was homogenized and centrifuged at
7,155 × g for 5 min at 4°C, and the total protein
concentration of the supernatant was determined using a
bicinchoninic acid assay (Applygen Technologies Inc., Beijing,
China). Protein samples were subsequently incubated with catalyst
and dichlorodihydrofluorescein solutions included in the ROS assay
kit. The fluorescence was read at 480/530 nm using a microplate
reader (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
ΔΨm assay
A tetramethylrhodamine ethyl ester perchlorate
(TMRE) assay (Sigma-Aldrich, EMD Millipore, Billerica, MA, USA) was
used to determine ΔΨm levels in the rat hippocampus. Briefly, the
hippocampus was homogenized and centrifuged at 7,155 × g for
5 min at 4°C, and the total protein concentration of the sample
supernatant was determined using a bicinchoninic acid assay. The
samples were subsequently incubated with 50 nM TMRE solutions for
15 min at room temperature, and the fluorescence was read at
549/574 nm using a microplate reader (Thermo Fisher Scientific,
Inc.).
Western blot analysis
Western blot analysis was performed to determine the
protein expression levels of CaN and IκBα in the rat hippocampus.
Briefly, the hippocampus was homogenized and centrifuged at 7,155 ×
g for 5 min at 4°C, and the total protein concentration of
the supernatant was determined using a bicinchoninic acid assay.
Proteins were separated by 10% SDS-PAGE, prior to transfer to the
polyvinylidene fluoride membranes. Membranes were then incubated
with the anti-CaN (dilution, 1:1,000; cat. no. ab3673; Abcam),
anti-IκBα and anti-phosphorylated (p)-IκBα (dilution, 1:1,000; cat.
nos. 4812 and 9246; Cell Signaling Technology, Inc., Danvers, MA,
USA) and anti-β-actin (dilution, 1:10,000; cat. no. 4970; Santa
Cruz Biotechnology, Inc., Dallas, TX, USA) primary antibodies
overnight at 4°C, followed by incubation with the IRDye®
800CW-conjugated goat anti-rabbit secondary antibody (dilution,
1:10,000; cat. no. 926-32211; LI-COR Biosciences, Inc., Lincoln,
NE, USA) for 2 h at room temperature. Immunoreactivity was
visualized by scanning membranes with an Odyssey 9120 infrared
imaging system (LI-COR Biosciences, Inc.). The densitometric
analysis was conducted using β-actin as a control for loading
differences, the protein levels in the experimental groups were
calculated as percentages of the control group.
CaN activity
A colorimetric assay was performed using a CaN
activity kit (cat. no. ab139461; Abcam) to determine CaN activity
in the rat hippocampus. Briefly, the hippocampus was homogenized
and centrifuged at 7,155 × g for 5 min at 4°C, and the total
protein concentration of the supernatant was determined using a
bicinchoninic acid assay. Protein lysates and calcineurin assay
buffer were mixed, calcineurin substrate was added and reaction
proceeded for 10 min. Green assay reagent was added and optical
density (OD)620nm was read on a microplate reader
(Thermo Fisher Scientific, Inc.). OD620nm data were
converted into released phosphate amount, and calcineurin activity
was calculated as a ratio of phosphate amount/reaction time.
Enzyme-linked immunosorbent assay
(ELISA)
ELISA was performed using an sandwich ELISA kit
(cat. no. ab100768; Abcam) to determine the IL-1β protein
expression level in the rat hippocampus. Briefly, the hippocampus
was homogenized and centrifuged at 7,155 × g for 5 min at
4°C, and the total protein concentration of the supernatant was
determined using a bicinchoninic acid assay. Protein lysates,
biotinylated IL-1β antibody, horseradish peroxidase-streptavidin
and TMB One-Step Substrate reagent were incubated in turn in
96-well plates for 2.5, 1 h, 45 min and 30 min at room temperature.
Stop solution was then added and OD450nm was read on a
microplate reader (Thermo Fisher Scientific, Inc.).
Immunofluorescence
Immunofluorescence was performed in order to
determine the nuclear translocation of RelA. Briefly, the rat
hippocampus was fixed with 4% paraformaldehyde for 24 h,
cryoprotected with 30% sucrose for 48 h, and sectioned using a
cryostat (Leica Microsystems, Inc., Buffalo Grove, IL, USA).
Coronal sections (of 10-µm thickness) were incubated with a RelA
primary antibody (dilution, 1:50; cat. no. 8242; Cell Signaling
Technology, Inc.) overnight at 4°C, followed by incubation with a
goat anti-rabbit IgG-fluorescein isothiocyanate-conjugated
secondary antibody (dilution, 1:200; cat. no. sc-2012; Santa Cruz
Biotechnology, Inc.) for 1 h at room temperature. Nuclei were
subsequently counterstained with DAPI (1:5,000; Roche Diagnostics
GmbH, Mannheim, Germany). Images were captured using a confocal
fluorescence microscope (Zeiss GmbH, Jena, Germany). Due to the
results of a previous study demonstrating that the CA1 region of
hippocampus serves an important role in memory formation (18), this region was analyzed for RelA
expression.
Morris water maze
The spatial learning ability and memory of rats was
evaluated using a Morris water maze test as described previously
(5). The place navigation test was
performed 24 h following isoflurane exposure, at which point rats
received four training trials daily for 5 days. In each trial, rats
were randomly placed in water at one of four equally spaced
starting positions. The time to locate a submerged platform (the
escape latency, defined by a cut-off time of 120 sec) and the swim
speed were recorded. A probe trial (duration, 120 sec) was
performed 24 h after the last trial, during which time the platform
was removed and the target quadrant dwell time was recorded.
Statistical analysis
Statistical analyses were performed using GraphPad
Prism software (version 5.0; GraphPad Software, Inc., La Jolla, CA,
USA). Data are expressed as the mean ± standard deviation or the
mean ± standard error. One-way or two-way analysis of variance
(ANOVA) was used to compare ROS, MMP, CaN, IκBα, p-IκBα, IL-1β, and
probe test results between groups. Two-way repeated-measures ANOVA
followed by a post-hoc Bonferroni test was used to compare place
navigation test results. P<0.05 was considered to indicate a
statistically significant difference.
Results
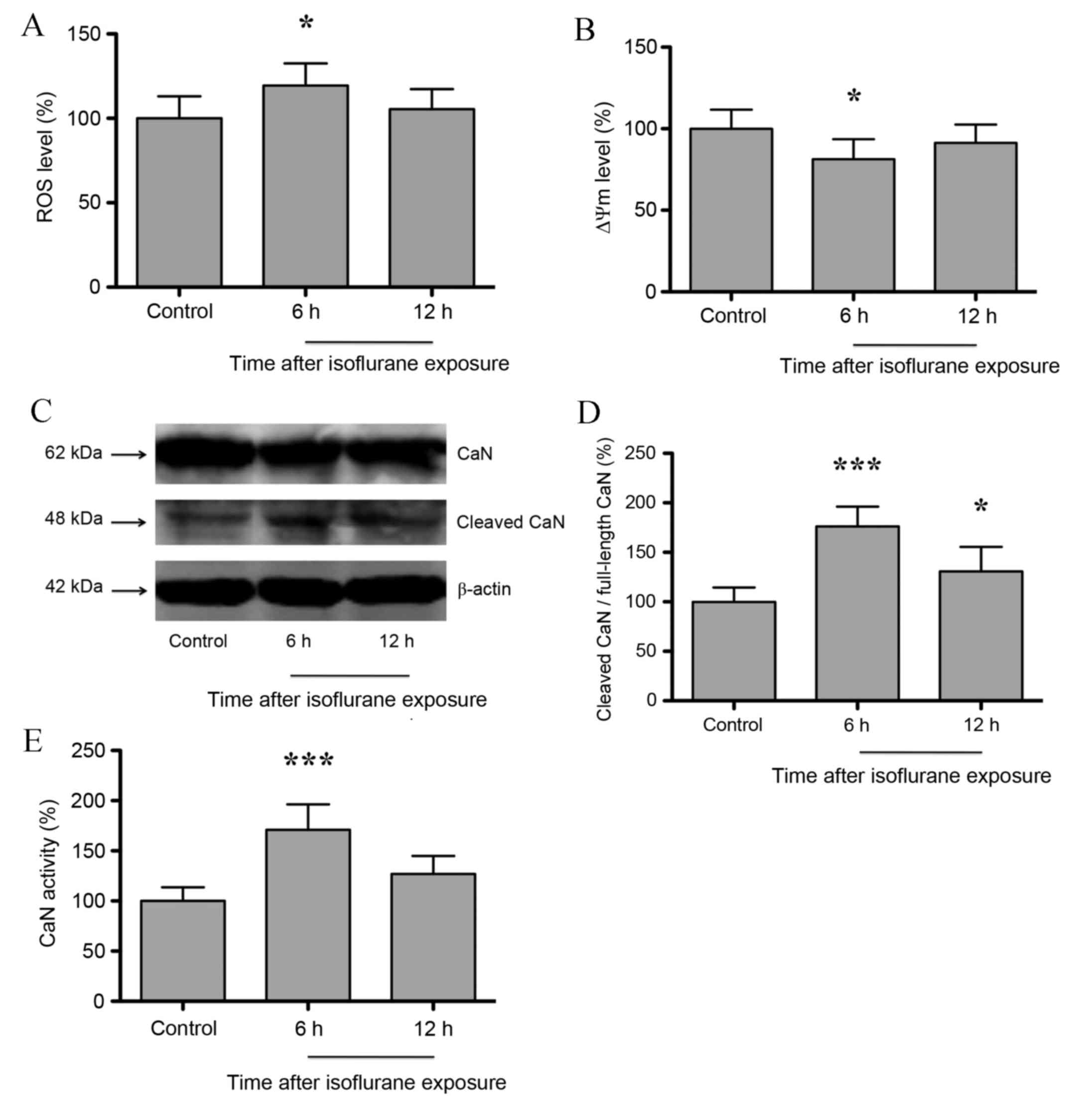
Isoflurane induces mitochondrial
dysfunction in the hippocampus of aged rats and is associated with
increased CaN activity
Zhang et al (11) demonstrated that isoflurane induced
mitochondrial dysfunction in neuroglioma/neurons. As shown in
Fig 1A, hippocampal ROS and ΔΨm
levels increased significantly at 6 h following isoflurane exposure
compared with untreated controls (P<0.05), but returned to
baseline levels at 12 h post-isoflurane exposure (P>0.05). In
addition, ΔΨm levels decreased significantly at 6 h (P<0.05),
and returned to baseline levels at 12 h following isoflurane
exposure (P>0.05; n=6; Fig.
1B). Consistent with Zhang et al (11), the results of the present study
indicate that isoflurane induces mitochondrial dysfunction in the
hippocampus of aged rats. Hypoxia, genetic or pharmacological
intervention-induced mitochondrial dysfunction may activate
mitochondrial retrograde signaling, and CaN serves an important
role in the initiation of mitochondrial retrograde signaling
(10). As shown in Fig. 1C and D, the expression of cleaved
CaN in the rat hippocampus increased significantly at 6 h
(P<0.001) following isoflurane exposure, and was maintained at a
relatively high level at 12 h after exposure when compared with
untreated controls (P<0.05; n=6;). In addition, CaN activity was
observed to increase significantly at 6 h (P<0.001) following
isoflurane exposure, but returned to baseline levels following 12 h
(P>0.05; n=6; Fig. 1E). These
results indicate that isoflurane induces mitochondrial dysfunction
and subsequent CaN activation in the hippocampus.
Isoflurane exposure leads to increased
IκBα phosphorylation and RelA nuclear translocation in the rat
hippocampus
NF-κB has been demonstrated to be involved in CaN
signaling (19), and consists of
RelA, RelB, c-Rel, p50 and p52 protein dimers. RelA/p50 is the most
abundant NF-κB heterodimer, and its nuclear translocation and
transcriptional activity is inhibited by IκBα (20). Thus, hippocampal IκBα and p-IκBα
protein expression following isoflurane exposure was determined. As
shown in Fig. 2A-C, the protein
expression levels of p-IκBα increased significantly, whereas IκBα
expression decreased significantly at 6 h (P<0.01 and
P<0.001, respectively) post-isoflurane exposure. IκBα expression
returned to baseline levels (P>0.05) at 12 h following
isoflurane exposure, whereas p-IκBα expression was maintained at a
significantly higher level (P<0.05) compared with untreated
controls (n=6; Fig. 2A-C). With
IκBα phosphorylation and subsequent degradation, NF-κB dimers may
enter the nucleus to regulate target gene expression. Therefore,
the nuclear translocation of RelA was investigated at 6 h following
isoflurane exposure. In untreated controls, RelA was primarily
distributed in the cytosol of the pyramidal cell layer in the CA1
region of hippocampus. However, following isoflurane exposure, RelA
exhibited a nuclear distribution (Fig.
2D).
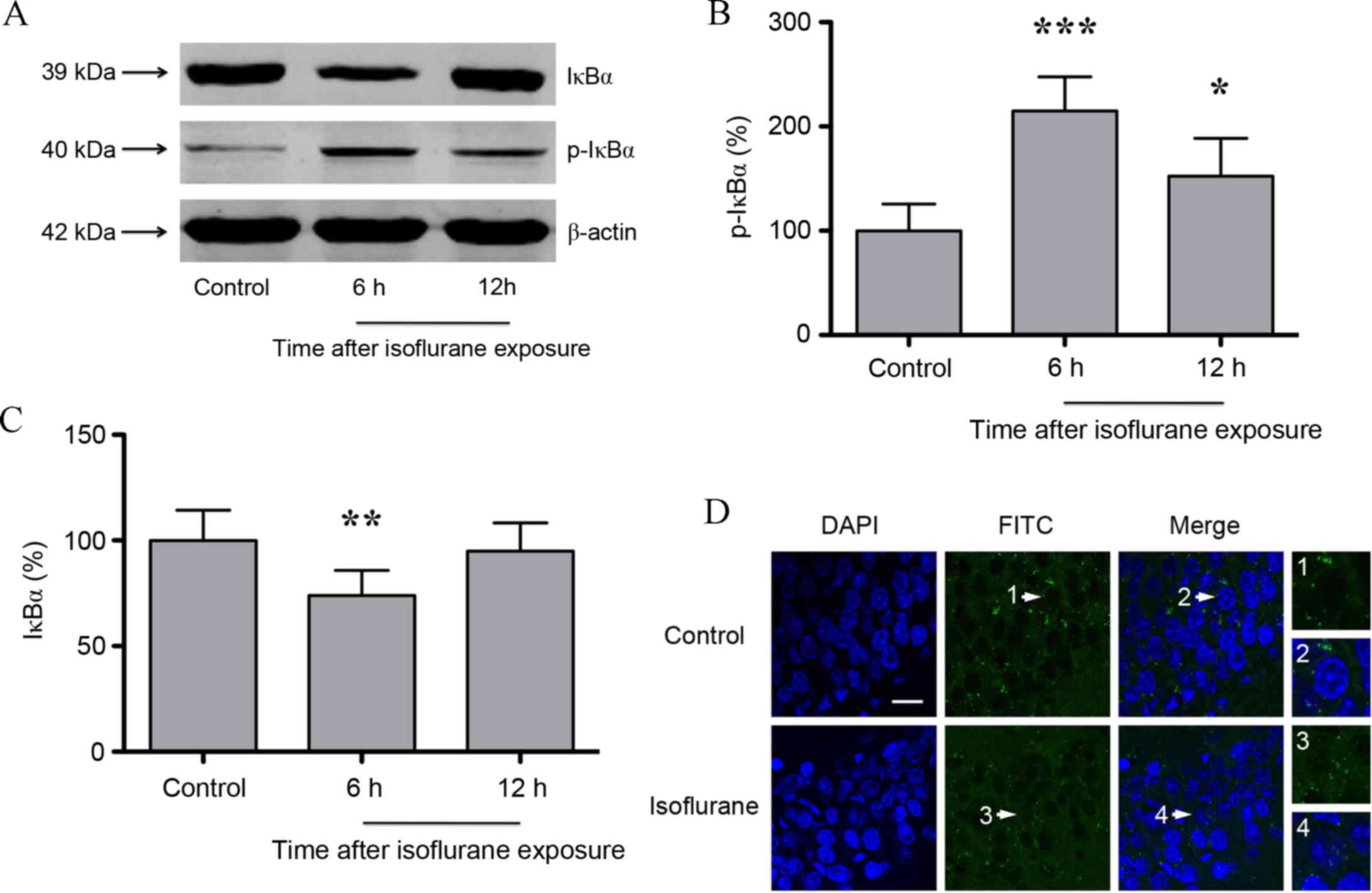 | Figure 2.(A) Western blot analysis and
quantification of (B) p-IκBα and (C) IκBα protein expression
levels. IκBα levels were significantly decreased, and p-IκBα levels
were significantly increased, at 6 h following isoflurane exposure.
(D) Immunofluorescence images showing RelA (FITC, green) and DAPI
(blue) counter-staining. In controls, RelA was primarily
distributed in the cytosol of the pyramidal cell layer in the CA1
region. At 6 h following isoflurane exposure, RelA was distributed
in the nucleus and cytosol. The white arrows indicate the typical
RelA distribution, which is shown in the high-magnification images
(magnification, ×400; scale bar, 20 µm). Data are presented as the
mean ± standard deviation (n=6). *P<0.05, **P<0.01 and
***P<0.001 vs. the control group. p-IκBα, phosphorylated
inhibitor of κB; FITC, fluorescein isothiocyanate. |
Inhibition of CaN attenuates
isoflurane-induced hippocampal IκBα phosphorylation and the nuclear
translocation of RelA, whereas NF-κB inhibition has no effect on
CaN activity
In order to investigate the effect of increased CaN
expression and activity on NF-κB following isoflurane exposure,
untreated and isoflurane-treated rats were treated with CsA. In
isoflurane + CsA-treated rats at 6 h following treatment, the
protein expression levels of IκBα increased significantly, whereas
the expression levels of p-IκBα decreased significantly when
compared with rats treated with isoflurane alone (P<0.01;
Fig. 3A-C). CsA treatment alone
did not affect IκBα or p-IκBα expression (P>0.05; n=6; Fig. 3A-C). It is therefore possible that
CsA attenuated isoflurane-induced hippocampal NF-κB activation. As
shown in Fig. 3D, CsA attenuated
isoflurane-induced RelA nuclear translocation. However, CsA
treatment alone did not affect RelA distribution, which was
primarily localized to the cytosol of the pyramidal cell layer in
the CA1 region of hippocampus (Fig.
3D). Subsequently, rats were treated with the NF-κB inhibitor,
PDTC, in order to investigate the effect of NF-κB on CaN. As shown
in Fig. 3E-G, PDTC did not affect
the expression level of cleaved CaN or its activity following
isoflurane exposure or in isoflurane-untreated controls (P>0.05;
n=6). The results suggest that CaN is an upstream mediator of NF-κB
activation in the hippocampus following isoflurane exposure.
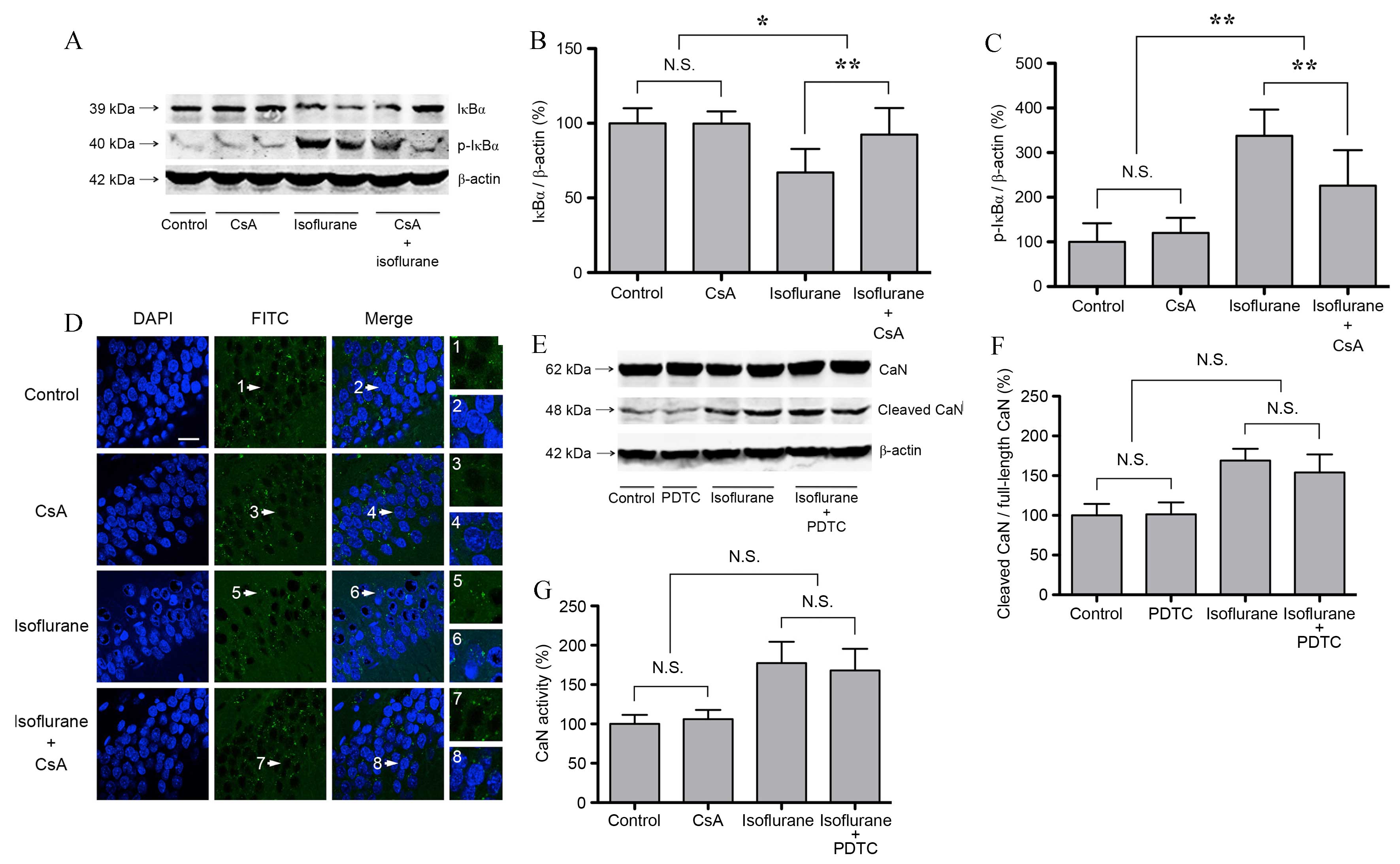 | Figure 3.CaN activation mediates
isoflurane-induced NF-κB activation in the hippocampus of aged
rats. CsA (7 mg/kg) or PDTC (100 mg/kg) was administered 30 min
prior to isoflurane exposure through intraperitoneal injection. (A)
Western blot analysis and quantification of (B) IκBα and (C) p-IκBα
protein expression levels relative to β-actin. IκBα expression
decreased and p-IκBα expression increased significantly at 6 h
following isoflurane exposure. CsA attenuated isoflurane-induced
alterations in IκBα expression, and CsA treatment alone did not
affect IκBα expression. (D) Immunofluorescence images,
demonstrating that the nuclear distribution of RelA increased
following isoflurane exposure in the pyramidal cell layer of the
CA1 hippocampal region. CsA attenuated isoflurane-induced RelA
nuclear translocation, whereas CsA treatment alone did not affect
RelA distribution. The white arrows indicate the typical RelA
distribution, which are provided as high magnification images
(magnification, ×400; scale bar, 20 µm). (E) Western blot analysis
and (F) quantification of cleaved CaN expression. The expression of
cleaved CaN increased significantly at 6 h following isoflurane
exposure. PDTC did not affect the expression of cleaved CaN in
untreated controls or isoflurane treated rats. (G) CaN activity
increased significantly at 6 h following isoflurane exposure, and
PDTC did not affect CaN activity after isoflurane exposure or in
the control group. Data are expressed as mean ± standard deviation
(n=6). *P<0.05 and **P<0.01 vs. the control group. CaN,
calcineurin; NF-κB, nuclear factor-κB; CsA, cyclosporin A; PDTA,
pyrrolidine dithiocarbamate; p-IκBα, phosphorylated inhibitor of
κB; N.S., not significant. |
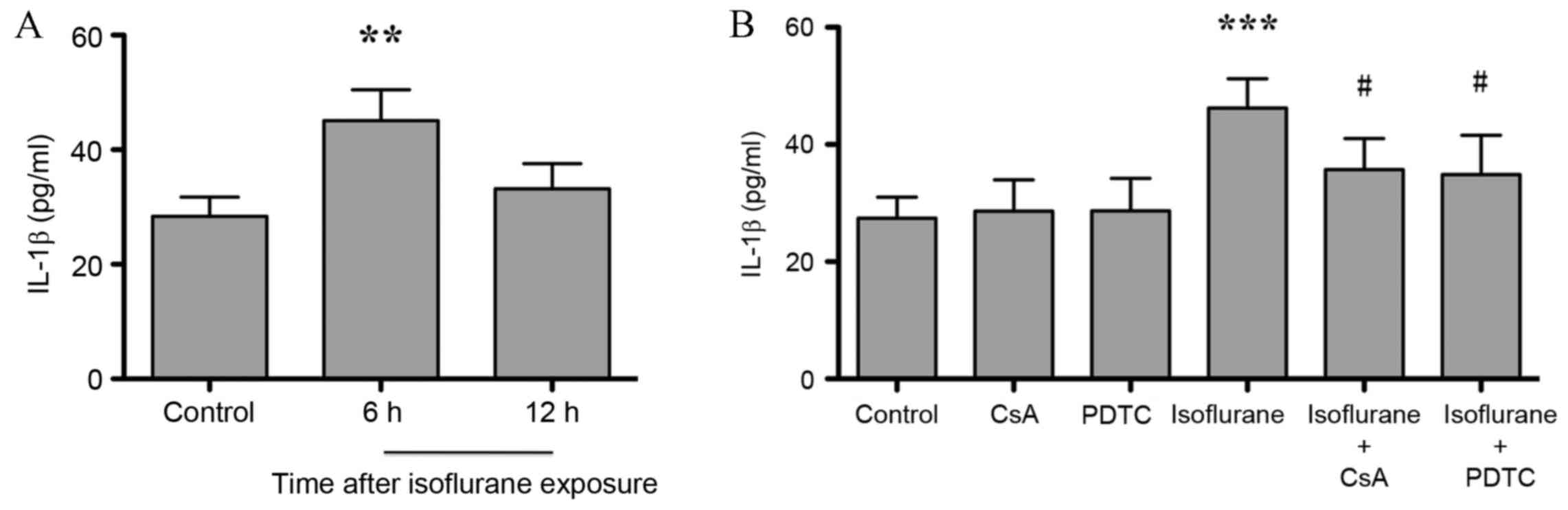
IL-1β is a downstream target of the
CaN/NF-κB signaling pathway in the hippocampus, and its expression
is increased in response to isoflurane exposure
NF-κB regulates the expression of genes involved in
inflammation and the innate immune response (21). As shown in Fig. 4A, the expression level of
hippocampal inflammatory factor IL-1β increased significantly at 6
h (P<0.001) compared with untreated controls, and returned to
baseline levels at 12 h after isoflurane exposure (P>0.05; n=6).
Treatment of isoflurane-exposed rats with CsA and PDTC
significantly attenuated isoflurane-induced IL-1β elevation
(P<0.05) and no difference between isoflurane + CsA and
isoflurane + PDTC groups was observed (P>0.05, Fig. 4B). CsA or PDTC treatment alone did
not affect IL-1β expression levels, when compared with untreated
controls (P>0.05, Fig. 4B).
These results indicate that CaN/NF-κB signaling may mediate
neuroinflammation in the hippocampus following isoflurane
exposure.
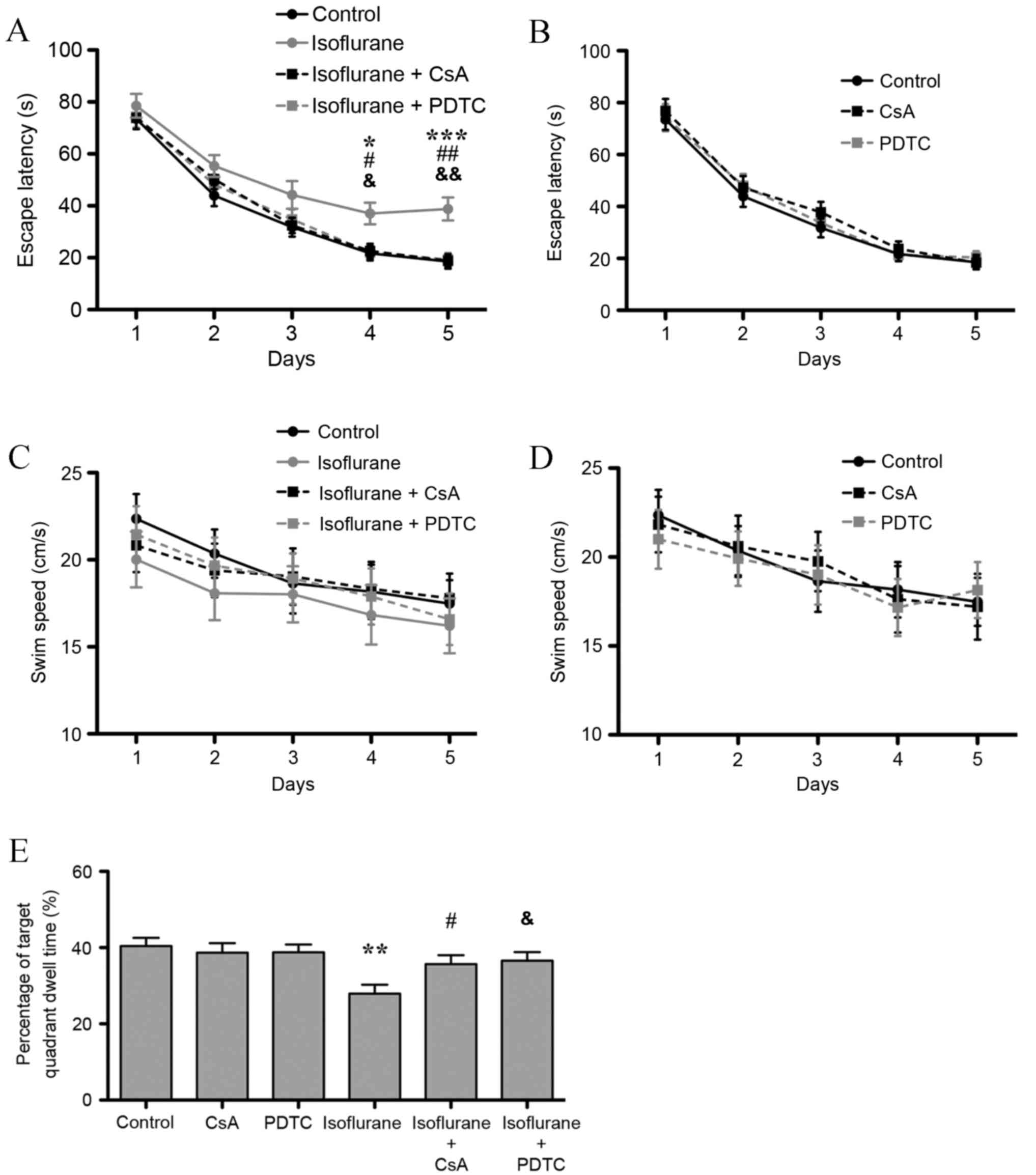
Isoflurane exposure decreased the
memory and spatial learning abilities of rats
Neuroinflammation is involved in AD, the stress
response (22) and in
hypoxic-ischemic encephalopathy (23), and may lead to cognitive
impairment. Due to the observed increase in the expression of IL-1β
(a marker of neuroinflammation) in the hippocampus of aged rats
following isoflurane exposure, the cognitive function of
isoflurane-treated rats was further investigated. The results of
the Morris water maze's place navigation test demonstrated that the
group (isoflurane exposure, CsA and PDTC treatment) and repeated
(Morris water maze training time) factors significantly affected
escape latency (P=0.014 and P<0.001), and no interaction was
found (P>0.05). The Bonferroni test demonstrated that exposure
to isoflurane prolonged escape latency on the 4th and 5th days
following isoflurane exposure (P<0.05 and P<0.001,
respectively; Fig. 5A).
Furthermore, CsA and PDTC treatment attenuated isoflurane-induced
escape latency prolongation also on the 4th and 5th days (P<0.05
and P<0.01, respectively), and no difference between isoflurane
+ CsA and isoflurane + PDTC groups was observed (P>0.05;
Fig. 5A). CsA or PDTC treatment
alone did not affect escape latency (Fig. 5B). As shown in Fig. 5C and D, no significant difference
in swim speed was observed among all the groups (P>0.05). In the
probe test, isoflurane treatment significantly decreased target
quadrant dwell time compared with untreated controls (P<0.01;
Fig. 5E). CsA or PDTC treatment
attenuated the isoflurane-induced decrease in target quadrant dwell
time (P<0.05), and no difference between isoflurane + CsA and
isoflurane + PDTC groups was observed (P>0.05, Fig. 5E). However, treatment with CsA or
PDTC alone did not affect target quadrant dwell time (P>0.05,
Fig. 5E). The results indicate
that isoflurane induced hippocampal mitochondrial dysfunction and
activated the CaN/NF-κB signaling pathway, which led to
neuroinflammation and subsequent cognitive impairment in aged
rats.
Discussion
Ca2+ is involved in the regulation of
multiple cellular functions (24),
and mitochondrial Ca2+ uptake and release serve
fundamental roles in a number of physiological processes, such as
ATP generation and cellular metabolism (25). In addition, disruption of
mitochondrial Ca2+ may induce apoptotic processes
through mPTP opening (26). The
results of the present study indicate that isoflurane impairs
mitochondrial function, and mitochondrial impairment may result in
cytosolic Ca2+ elevation. Furthermore, isoflurane may
induce overactivation of inositol 1,4,5-trisphosphate or ryanodine
receptors located on the endoplasmic reticulum (ER) membrane
(27), leading to Ca2+
escaping from the ER. Based on the results presented in the current
study and a previous study (28),
CaN, a Ca2+-dependent serine/threonine protein kinase,
is activated following cytosolic Ca2+ elevation, which
may initiate mitochondrial retrograde signaling.
NF-κB signaling participates in mitochondrial
biogenesis and metabolism (29).
In the central nervous system, NF-κB functions as a pleiotropic
regulator of target gene expression, hippocampal neurogenesis
(30) and neurodegeneration
(31). The results of the present
study suggest that isoflurane activates NF-κB following
mitochondrial dysfunction and CaN activation, and CaN is an
upstream regulator of NF-κB activation. A complex interaction
between CaN and NF-κB has been reported previously (32,33).
CaN may dephosphorylate Bcl-10, which leads to IKKβ
phosphorylation, IκBα degradation, RelA nuclear translocation and
DNA binding in T helper cells (19). These results are consistent with
those of the current study demonstrating that an increase in CaN
expression and activity in the hippocampus, is associated with a
reduction in IκBα expression. However, it has also been reported
that CaN inhibition may promote NF-κB activation in kidney tubular
cells (34). In addition, NF-κB
has been reported to activate regulator of calcineurin 1 gene
expression (35), which may
subsequently interact with calcineurin A and activate downstream
signaling pathways thereby affecting central nervous system
development. In contrast, the results of the present study indicate
that NF-κB inhibition does not affect CaN activity following
isoflurane exposure.
NF-κB is a nuclear transcription factor that
regulates the expression of genes that are critical for the
regulation of apoptosis, inflammation and the immune system. In the
nervous system, NF-κB was observed to inhibit neurite growth
through the phosphorylation of RelA on Ser-536 (36). RelA-containing dimers induce
proapoptotic gene expression in ischemic neurons, and its
inhibition exerts protective effects in neuronal cells (37). Alcohol induces IL-6, IL-1β and
tumor necrosis factor-α elevation in the mouse brain through NF-κB
signaling (38). In addition,
NF-κB inhibition reduces proinflammatory inducible nitric oxide
synthase and cytochrome c oxidase subunit 2 expression
levels, and ameliorates inflammation and locomotor recovery during
spinal cord injury (39). The
results of the present study indicate that isoflurane elevates
IL-1β expression (a typical marker of neuroinflammation) in the
hippocampus of aged rats, and inhibition of CaN and NF-κB may
attenuate this process. This indicates that the CaN/NF-κB-mediated
mitochondrial retrograde signaling pathway may be involved in the
upstream mechanism for isoflurane-induced neuroinflammation in the
hippocampus.
The results of the present study indicate that
isoflurane induces cognitive impairment in aged rats. The
pathological consequences of anesthetic-induced cognitive
impairment includes amyloid-β accumulation and tau phosphorylation
(40). In addition, the immune
system exacerbates POCD pathology (7), and inflammatory cytokines and
microglia may be potential therapeutic targets. In the present
study, inhibition of CaN and NF-κB attenuated hippocampal
neuroinflammation and cognitive dysfunction, which confirms the
role of neuroinflammation in inhaled anesthetic-induced cognitive
impairment. This indicates that the CaN/NF-κB signaling pathway may
present a potential therapeutic target for the treatment and/or
prevention of hippocampal neuroinflammation and memory impairment
in aged rats.
CaN dephosphorylates the Ca2+
sensor/translocation domain of nuclear factor of activated T-cells
(NFAT), which leads to NFAT nuclear translocation and activation of
target gene expression (41). A
previous study indicated that consistent with intermediate to
severe AD, the nuclear translocation of NFATc4 in the hippocampus
may also involve in CaN-mediated memory impairment following
isoflurane exposure (28).
Moreover, the cAMP response element-binding protein, CCAAT enhancer
binding protein δ, and the heterogeneous ribonucleoprotein A2 are
reportedly involved in CaN-mediated mitochondrial retrograde
signaling and contribute to the regulation of downstream target
gene expression (42). Thus, the
role of these factors in isoflurane-induced memory impairment, and
the mechanisms underlying these processes, may require further
investigation.
In conclusion, the results of the present study
indicate that isoflurane induces hippocampal mitochondrial
dysfunction and CaN activation, and CaN functions as an upstream
mediator of NF-κB activation. In addition, the protein expression
levels of IL-1β were elevated following isoflurane exposure, which
is a classical marker of neuroinflammation, and may result in
cognitive impairment in aged rats. Furthermore, both CaN and NF-κB
inhibition attenuated isoflurane-induced neuroinflammation and
subsequent cognitive impairment. Collectively, these results reveal
the role of mitochondrial retrograde signaling and the factors
involved in inhaled anesthetic induced-neuroinflammation and
cognitive impairment. These factors may therefore present promising
therapeutic targets for the treatment and/or prevention of
POCD.
Acknowledgements
The present study was supported by the National
Natural Science Foundation Of China (grant no. 81400869) and the
Scientific Research Foundation for Returned Scholars (awarded to Dr
Cheng Ni, Peking University Third Hospital, Beijing, China).
References
|
1
|
Newman MF, Kirchner JL, Phillips-Bute B,
Gaver V, Grocott H, Jones RH, Mark DB, Reves JG and Blumenthal JA:
Neurological Outcome Research Group and the Cardiothoracic
Anesthesiology Research Endeavors Investigators: Longitudinal
assessment of neurocognitive function after coronary-artery bypass
surgery. N Engl J Med. 344:395–402. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Steinmetz J, Christensen KB, Lund T, Lohse
N and Rasmussen LS: ISPOCD Group: Long-term consequences of
postoperative cognitive dysfunction. Anesthesiology. 110:548–555.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Heneka MT and O'Banion MK: Inflammatory
processes in Alzheimer's disease. J Neuroimmunol. 184:69–91. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tan ZS, Beiser AS, Vasan RS, Roubenoff R,
Dinarello CA, Harris TB, Benjamin EJ, Au R, Kiel DP, Wolf PA and
Seshadri S: Inflammatory markers and the risk of Alzheimer disease:
The framingham study. Neurology. 68:1902–1908. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li ZQ, Rong XY, Liu YJ, Ni C, Tian XS, Mo
N, Chui DH and Guo XY: Activation of the canonical nuclear
factor-κB pathway is involved in isoflurane-induced hippocampal
interleukin-1β elevation and the resultant cognitive deficits in
aged rats. Biochem Biophys Res Commun. 438:628–634. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xu Z, Dong Y, Wang H, Culley DJ,
Marcantonio ER, Crosby G, Tanzi RE, Zhang Y and Xie Z:
Age-dependent postoperative cognitive impairment and
Alzheimer-related neuropathology in mice. Sci Rep.
4:37662014.PubMed/NCBI
|
|
7
|
Wu X, Lu Y, Dong Y, Zhang G, Zhang Y, Xu
Z, Culley DJ, Crosby G, Marcantonio ER, Tanzi RE and Xie Z: The
inhalation anesthetic isoflurane increases levels of
proinflammatory TNF-α, IL-6, and IL-1β. Neurobiol Aging.
33:1364–1378. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Solberg NO, Chamberlin R, Vigil JR, Deck
LM, Heidrich JE, Brown DC, Brady CI, Jagt TA Vander, Garwood M,
Bisoffi M, et al: Optical and SPION-enhanced MR imaging shows that
trans-stilbene inhibitors of NF-κB concomitantly lower Alzheimer's
disease plaque formation and microglial activation in AbetaPP/PS-1
transgenic mouse brain. J Alzheimers Dis. 40:191–212.
2014.PubMed/NCBI
|
|
9
|
Liu T, Zhang T, Yu H, Shen H and Xia W:
Adjudin protects against cerebral ischemia reperfusion injury by
inhibition of neuroinflammation and blood-brain barrier disruption.
J Neuroinflammation. 11:1072014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Guha M and Avadhani NG: Mitochondrial
retrograde signaling at the crossroads of tumor bioenergetics,
genetics and epigenetics. Mitochondrion. 13:577–591. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang Y, Xu Z, Wang H, Dong Y, Shi HN,
Culley DJ, Crosby G, Marcantonio ER, Tanzi RE and Xie Z:
Anesthetics isoflurane and desflurane differently affect
mitochondrial function, learning and memory. Ann Neurol.
71:687–698. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rizzuto R, De Stefani D, Raffaello A and
Mammucari C: Mitochondria as sensors and regulators of calcium
signalling. Nat Rev Mol Cell Biol. 13:566–578. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Guha M, Srinivasan S, Biswas G and
Avadhani NG: Activation of a novel calcineurin-mediated
insulin-like growth factor-1 receptor pathway, altered metabolism,
and tumor cell invasion in cells subjected to mitochondrial
respiratory stress. J Biol Chem. 282:14536–14546. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Biswas G, Tang W, Sondheimer N, Guha M,
Bansal S and Avadhani NG: A distinctive physiological role for
IkappaBbeta in the propagation of mitochondrial respiratory stress
signaling. J Biol Chem. 283:12586–12594. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wu X, Nguyen BC, Dziunycz P, Chang S,
Brooks Y, Lefort K, Hofbauer GF and Dotto GP: Opposing roles for
calcineurin and ATF3 in squamous skin cancer. Nature. 465:368–372.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Crack PJ, Taylor JM, Ali U, Mansell A and
Hertzog PJ: Potential contribution of NF-kappaB in neuronal cell
death in the glutathione peroxidase-1 knockout mouse in response to
ischemia-reperfusion injury. Stroke. 37:1533–1538. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ni C, Tan G, Luo A, Qian M, Tang Y, Zhou
Y, Wang J, Li M, Zhang Y, Jia D, et al: Melatonin premedication
attenuates isoflurane anesthesia-induced β-amyloid generation and
cholinergic dysfunction in the hippocampus of aged rats. Int J
Neurosci. 123:213–220. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Whitlock JR, Heynen AJ, Shuler MG and Bear
MF: Learning induces long-term potentiation in the hippocampus.
Science. 313:1093–1097. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Frischbutter S, Gabriel C, Bendfeldt H,
Radbruch A and Baumgrass R: Dephosphorylation of Bcl-10 by
calcineurin is essential for canonical NF-κB activation in Th
cells. Eur J Immunol. 41:2349–2357. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ferreiro DU and Komives EA: Molecular
mechanisms of system control of NF-kappaB signaling by
IkappaBalpha. Biochemistry. 49:1560–1567. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rius J, Guma M, Schachtrup C, Akassoglou
K, Zinkernagel AS, Nizet V, Johnson RS, Haddad GG and Karin M:
NF-kappaB links innate immunity to the hypoxic response through
transcriptional regulation of HIF-1alpha. Nature. 453:807–811.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee W, Moon M, Kim HG, Lee TH and Oh MS:
Heat stress-induced memory impairment is associated with
neuroinflammation in mice. J Neuroinflammation. 12:1022015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mirza MA, Ritzel R, Xu Y, McCullough LD
and Liu F: Sexually dimorphic outcomes and inflammatory responses
in hypoxic-ischemic encephalopathy. J Neuroinflammation. 12:322015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Williams GS, Boyman L, Chikando AC,
Khairallah RJ and Lederer WJ: Mitochondrial calcium uptake. Proc
Natl Acad Sci USA. 110:10479–10486. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mallilankaraman K, Cardenas C, Doonan PJ,
Chandramoorthy HC, Irrinki KM, Golenár T, Csordás G, Madireddi P,
Yang J, Müller M, et al: MCUR1 is an essential component of
mitochondrial Ca2+ uptake that regulates cellular
metabolism. Nat Cell Biol. 14:1336–1343. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Duchen MR: Mitochondria and Ca(2+) in cell
physiology and pathophysiology. Cell calcium. 28:339–348. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang H, Liang G, Hawkins BJ, Madesh M,
Pierwola A and Wei H: Inhalational anesthetics induce cell damage
by disruption of intracellular calcium homeostasis with different
potencies. Anesthesiology. 109:243–250. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ni C, Li Z, Qian M, Zhou Y, Wang J and Guo
X: Isoflurane induced cognitive impairment in aged rats through
hippocampal calcineurin/NFAT signaling. Biochem Biophys Res Commun.
460:889–895. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bakkar N, Ladner K, Canan BD,
Liyanarachchi S, Bal NC, Pant M, Periasamy M, Li Q, Janssen PM and
Guttridge DC: IKKα and alternative NF-αB regulate PGC-1β to promote
oxidative muscle metabolism. J Cell Biol. 196:497–511. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Crampton SJ and O'Keeffe GW: NF-κB:
Emerging roles in hippocampal development and function. Int J
Biochem Cell Biol. 45:1821–1824. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Camandola S and Mattson MP: NF-kappa B as
a therapeutic target in neurodegenerative diseases. Expert Opin
Ther Targets. 11:123–132. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
de la Fuente V, Federman N, Fustiñana MS,
Zalcman G and Romano A: Calcineurin phosphatase as a negative
regulator of fear memory in hippocampus: Control on nuclear
factor-κB signaling in consolidation and reconsolidation.
Hippocampus. 24:1549–1561. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lim D, Iyer A, Ronco V, Grolla AA,
Canonico PL, Aronica E and Genazzani AA: Amyloid beta deregulates
astroglial mGluR5-mediated calcium signaling via calcineurin and
Nf-kB. Glia. 61:1134–1145. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
González-Guerrero C, Ocaña-Salceda C,
Berzal S, Carrasco S, Fernández-Fernández B, Cannata-Ortiz P, Egido
J, Ortiz A and Ramos AM: Calcineurin inhibitors recruit protein
kinases JAK2 and JNK, TLR signaling and the UPR to activate
NF-κB-mediated inflammatory responses in kidney tubular cells.
Toxicol Appl Pharmacol. 272:825–841. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zheng L, Liu H, Wang P, Song W and Sun X:
Regulator of calcineurin 1 gene transcription is regulated by
nuclear factor-kappaB. Curr Alzheimer Res. 11:156–164. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gutierrez H, O'Keeffe GW, Gavaldà N,
Gallagher D and Davies AM: Nuclear factor kappa B signaling either
stimulates or inhibits neurite growth depending on the
phosphorylation status of p65/RelA. J Neurosci. 28:8246–8256. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Inta I, Paxian S, Maegele I, Zhang W,
Pizzi M, Spano P, Sarnico I, Muhammad S, Herrmann O, Inta D, et al:
Bim and Noxa are candidates to mediate the deleterious effect of
the NF-kappa B subunit RelA in cerebral ischemia. J Neurosci.
26:12896–12903. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang Y, Wei G, Di Z and Zhao Q:
miR-339-5p inhibits alcohol-induced brain inflammation through
regulating NF-κB pathway. Biochem Biophys Res Commun. 452:450–456.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rafati DS, Geissler K, Johnson K, Unabia
G, Hulsebosch C, Nesic-Taylor O and Perez-Polo JR: Nuclear
factor-kappaB decoy amelioration of spinal cord injury-induced
inflammation and behavior outcomes. J Neurosci Res. 86:566–580.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Dong Y, Wu X, Xu Z, Zhang Y and Xie Z:
Anesthetic isoflurane increases phosphorylated tau levels mediated
by caspase activation and Aβ generation. PLoS One. 7:e393862012.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Canellada A, Cano E, Sánchez-Ruiloba L,
Zafra F and Redondo JM: Calcium-dependent expression of TNF-alpha
in neural cells is mediated by the calcineurin/NFAT pathway. Mol
Cell Neurosci. 31:692–701. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Guha M, Tang W, Sondheimer N and Avadhani
NG: Role of calcineurin, hnRNPA2 and Akt in mitochondrial
respiratory stress-mediated transcription activation of nuclear
gene targets. Biochim Biophys Acta. 1797:1055–1065. 2010.
View Article : Google Scholar : PubMed/NCBI
|