Introduction
Malignant gliomas are the most common primary brain
tumors, and glioblastomas are among the most aggressive form of
human cancers (1). Astrocytic
tumors are the most common glial neoplasms, with an annual
incidence of 3–4/100,000 populations, and roughly 80% are
glioblastomas. Genetic defects are shared in GBM (2). GBM is a malignant brain tumor
typically resulting in death of the patient within one year
following diagnosis (3).
Angiogenesis is a hallmark of cancer. Tumor cells can initiate
angiogenesis, by firstly progressing an angiogenic phenotype
through angiogenic switch (4).
Their characteristics are a solid linkage of extremely circuitous
and large-diameter vessels (5).
The tumor vasculature is functionally unusual and this increased
vessel permeability can lead to vasogenic edema and hemorrhages
(6). As the vasculature in GBMs is
common with a seriously disordered vessel type, this may also limit
the ability of radio- and chemotherapy by blocking blood flow
(7). Accumulated preclinical and
clinical reports have provided innovative molecular targets, which
may improve the therapeutic advantage from anti-angiogenic
strategies (8). At this time
point, the promising drug for targeting vascular endothelial growth
factor (VEGF) is bevacizumab (BEV), a recombinant antibody to block
human VEGF-A (9). Nevertheless,
the reported positive effect of BEV treatment may be due to imaging
restrictions causing from reduced neoangiogenesis and reduced
vascular permeability at least in part, thus resulting to an
obvious but debatable reduction in the contrast-enhancing tumor
volume (10). The phase III trials
with avastin and radiation therapy has been performed to evaluate
the effectiveness of temozolomide (TMZ)-based radio-chemotherapy
with the BEV treatment for GBM patients (11). The case study of avastin treatment
(AVAglio) for GBM patients indicated a significant improvement of
progression-free survival (PFS) by 4.4 months with BEV co-treatment
whereas radiation therapy alone group (RTOG) with BEV co-treatment
failed to show significant benefits in terms of PFS and overall
survival. However, this PFS benefit did not improve the overall
survival (12). Hence it is
critical to find the new target for GBM patient in clinics with the
exception of the TMZ and VEGF targeting drug. Mitochondrial
transcription factor A (TFAM) is a main regulator of mitochondrial
transcription in mammals. Its expression level correlate to mtDNA
content and gene expression (13).
Also TFAM has a function in mitochondrial DNA replication, since
transcription generates an RNA primer required for initiation of
mtDNA replication (14). Abnormal
function of mitochondria has been found to be connected to many
metabolic disorders, including cancer (15). Therefore the putative role of TFAM
in glioblastoma was investigated since the novel therapeutic
strategy is urgently needed in brain cancer. In this study, we have
provided the clear evidence to support the upregulation of TFAM in
glioblastoma cell lines and the tissue from glioma patients. This
information will provide critical knowledge for better
understanding the regulatory mechanism of cancer as well as a
promising therapeutic strategy for brain cancer.
Materials and methods
Antibodies and reagents
Anti-TFAM antibody and Anti-actin antibodies were
purchased from Sigma-Aldrich (St. Louis, MO, USA). Horseradish
peroxidase-conjugated anti-mouse IgG or anti-rabbit IgG secondary
antibodies were purchased from Komabiotech (Seoul, Korea).
Cell culture
The glioblastoma cells (U87-MG, U251-MG, U343-MG and
U373-MG) were maintained in medium (RPMI) supplemented with 10%
FBS, 25 mM HEPES (Thermo Fisher Scientific, Waltham, MA, USA), 1%
antibiotics-antimycotics (Life Technologies, Carlsbad, CA,
USA).
Immunoblot analysis
The western blot analysis was performed as
previously described (16–18). Briefly, cells were placed on ice
and extracted with lysis buffer containing 50 mM Tris-HCl, pH 7.5,
1% v/v Nonidet P-40, 120 mM NaCl, 25 mM sodium fluoride, 40 mM
β-glycerol phosphate, 0.1 mM sodium orthovanadate, 1 mM
phenylmethylsulfonyl fluoride, 1 mM benzamidine, and 2 µM
microcystin-LR. Lysates were centrifuged for 15 min at 12,000 × g.
The cell extracts were resolved by 10–15% SDS-PAGE, and transferred
to Immobilon-P membranes (Millipore). The filters were blocked for
1 h in 1X Tris-buffered saline buffer (TBS; 140 mM NaCl, 2.7 mM
KCl, 250 mM Tris-HCl, pH 7.4), containing 5% skimmed milk and 0.2%
Tween-20, followed by an overnight incubation with the anti-TFAM
and anti-actin antibodies diluted 1,000-fold at 4°C. The secondary
antibody was horseradish peroxidase-conjugated anti-mouse IgG or
anti-rabbit IgG (Komabiotech), diluted 5,000-fold in the blocking
buffer. The detection of protein expression was visualized by
enhanced chemiluminescence, according to the manufacturer's
instructions (Thermo Fisher Scientific).
Real-time quantitative reverse
transcription-polymerase chain reaction (qRT-PCR)
Total RNA was extracted from frozen tissue samples
or from cells using the PureHelix RNA Extraction Solution
(Nanohelix, Korea). The cDNA was synthesized from total RNA with
the SuperScript III First-Strand Synthesis system for qRT-PCR
(Invitrogen, Grand Island, NY, USA). The qRT-PCR measurement of
individual cDNAs was performed using SYBR-Green dye to measure
duplex DNA formation with the StepOne Plus real-time PCR system
(Invitrogen) and normalized to the expression of glyceraldehyde
3-phosphate dehydrogenase (GAPDH) RNA. The following primers were
used in the qRT-PCR (F, forward; R, reverse); human TFAM,
F-5′-GGCAAGTTGTCCAAAGAAACC/R-5′-GCATCTGGGTTCTGAGCTTTA; human GAPDH,
F-5′-TCGACAGTCAGCCGCATCTTCTTT/R-5′-TACGACCAAATCCGTTGACTCCGA.
RNA sequencing and RNA-Seq data
analysis
Total RNA of U87-MG, U251-MG and normal brain was
extracted using TRIzol reagent (Invitrogen) following the
manufacturer's procedures. The total RNA quantity and purity were
analysis of Bioanalyzer 2100 and RNA 6000 Nano LabChip kit
(Agilent, Santa Clara, CA, USA). Roughly 10 µg of total RNA was
subjected to isolate Poly(A) mRNA with poly-T oligo attached
magnetic beads (Invitrogen). Following purification, the mRNA is
fragmented into small pieces using divalent cations under raised
temperature. Then the cleaved RNA fragments were
reverse-transcribed to create the final cDNA library in accordance
with the protocol for the mRNA-Seq sample preparation kit
(Illumina). The average insert size for the paired-end libraries
was 300 bp (± 50 bp). Next we performed the paired-end sequencing
on an Illumina HiSeq 2000 system at Macrogen (Seoul, Korea)
following the vendor's recommended protocol. For each sample,
sequenced reads were aligned to the UCSC human reference genome
(19) using the Tophat package
(20), which initially removes a
portion of the reads based on quality information accompanying each
read and then maps the reads to the reference genome. FPKM
(fragments per kilobase of exon per million fragments mapped) were
calculated to compare the expression level of TFAM mRNA variants in
each sample.
Confocal imaging analysis and indirect
immunofluorescence
U251-MG cells were grown on glass coverslips until
they were 50–70% confluent. After 24 h, the cells were fixed in 4%
paraformaldehyde at room temperature for 10 min and permeabilized
in 0.2% Triton X-100 for 5 min at room temperature. Then cells were
incubated in blocking buffer containing 5% bovine serum albumin
(Sigma) in 1X TBS for 1 h at 37°C. The rabbit polyclonal anti-TFAM
was diluted 200-fold for primary antibody and incubated for
overnight. The secondary antibody, FITC-conjugated anti-rabbit
antibody (BD Biosciences) was used. After appropriate rinsing,
cover slips were mounted with Vectashield (Vector Laboratories) and
visualized using a Zeiss confocal microscope.
Immunohistochemistry
The analysis of immunohistochemistry was performed
as the described previously (21).
A human cancer tissue array slide with paraffin sections was
purchased from Bio Max (US Biomax Inc). Histostain-Plus kits (Zymed
Laboratories Inc.) were used in accordance with the manufacturer's
instructions for the immunohistochemistry of tissue array. Briefly,
paraffin sections were deparaffinized with xylene and rehydrated in
a graded series of ethanol. The slide was submerged in peroxidase
quenching solution for 10 min. After it was washed twice with PBS
for 5 min, it was added with 2 drops of reagent A for blocking and
incubated for 30 min. Following two washes with PBS, the primary
antibody, anti-TFAM antibody, was applied at 4°C for overnight.
Then biotinylated secondary antibody, reagent B, was added after
rinsing with PBS. It was incubated at room temperature for 1 h. It
was rinsed with PBS and dropped with enzyme conjugated reagent C.
After it was washed with PBS, DAB chromogen, and a mixture of
reagent D1, D2, and D3, it was dropped, and signals were observed
with a florescence microscope. Then the reaction was stopped with
distilled water, and pictures were taken with a microscope.
Bioinformatics data set
Glioma data sets and corresponding clinical data
were downloaded from the publicly available databases (446 cases
from the Repository of Molecular Brain Neoplasia Data (REMBRANDT;
http://www.betastasis.com/glioma/rembrandt/). Normal,
n=21; GBM, n=214; oligodendroglioma, n=66; astrocytomas, n=145.
Statistical analysis
Data are expressed as the mean ± SD from at least
three separate experiments performed triplicate. The differences
between groups were analyzed using a Student's t-test and
P<0.05 was considered significant, and P<0.01 was highly
significant compared with corresponding control values. Statistical
analyses were carried out using SPSS software ver. 13.0 (SPSS Inc.,
Chicago, IL, USA).
Results
Enhancement of TFAM expression in
glioblastoma cell lines
In order to investigate the putative roles of TFAM
in brain cancer, western blot analysis with anti-TFAM antibody were
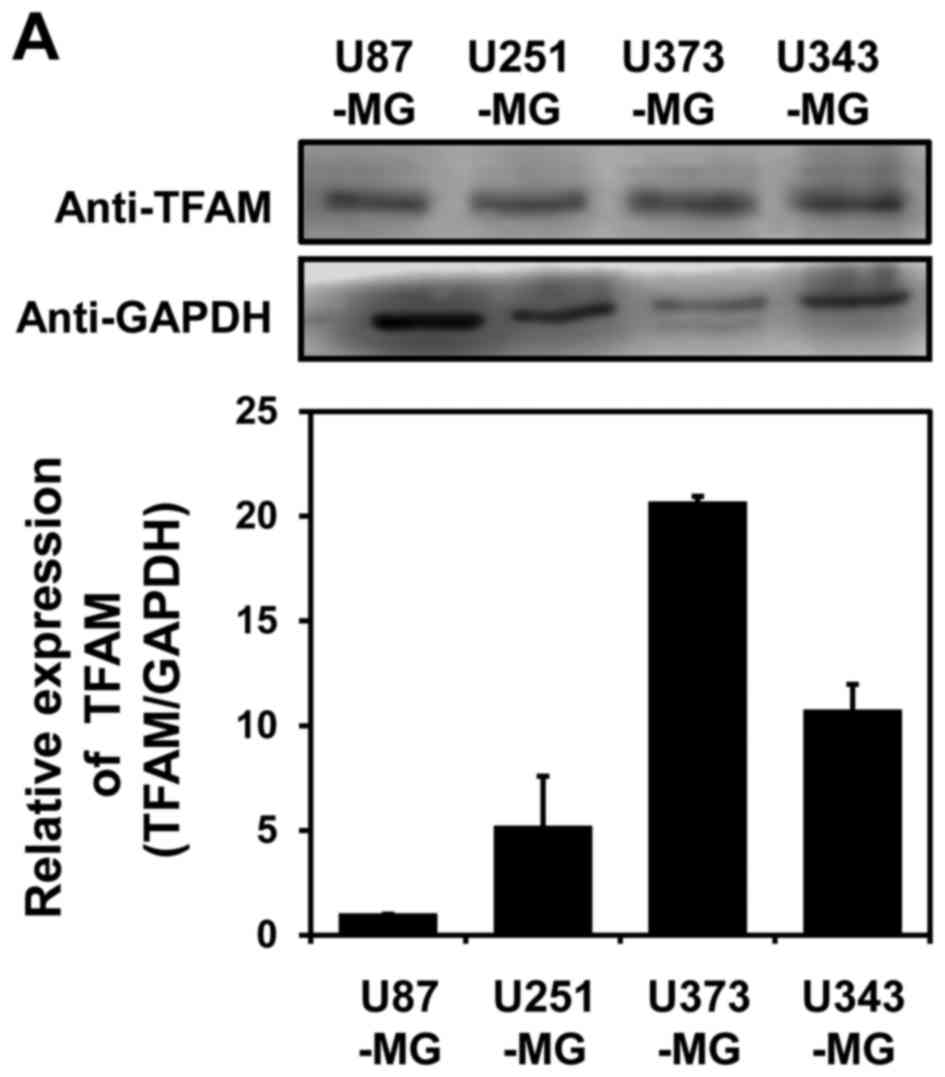
performed. As shown in Fig. 1A,
the protein expression of TFAM is markedly increased especially in
U343-MG cells and U373-MG cells compared to other cells. Analysis
of quantitative real-time PCR (qRT-PCR) with glioblastoma cell
lines also showed that mRNA level of TFAM is elevated in U251-MG
cells, U343-MG cells and U373-MG cells (Fig. 1B).
Transcriptional induction of TFAM mRNA
in U87-MG cells and U251-MG cells
Based on the observation of Fig. 1A and B, the mRNA levels of TFAM
were also monitored by using RNA sequencing of glioblastoma cell
lines. Total RNA were isolated from two cell lines (U87-MG and
U251-MG) which showed the low expression of TFAM by western blot
analysis and normal brain for the comparison of TFAM expression.
Roughly 10 µg of total RNA was further isolated to mRNA. Then mRNA
were fragmented into small pieces, followed by the
reverse-transcription of cleaved mRNA into the final cDNA library.
Fragments per kilobase of exon per million fragments mapped (FPKM)
were calculated to compare the expression level of TFAM mRNA
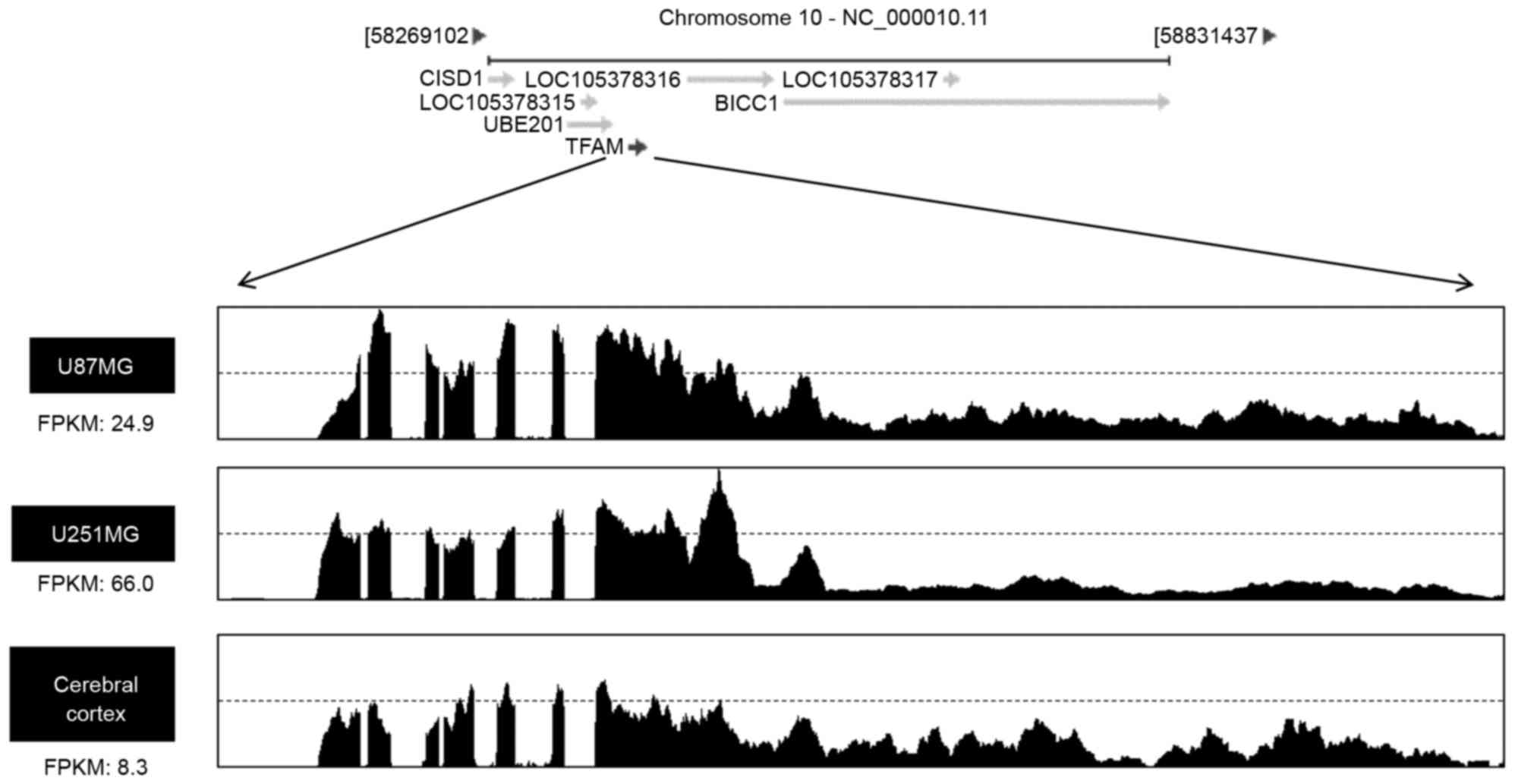
variants in each sample. As shown in Fig. 2, FPKM values were markedly higher
in U87-MG cells (24.9) and U251-MG cells (66.0) than its of
cerebral cortex (8.3), indicating that TFAM is transcriptionally
upregulated in glioblastoma cells.
Subcellular distribution of TFAM in
U251-MG cells
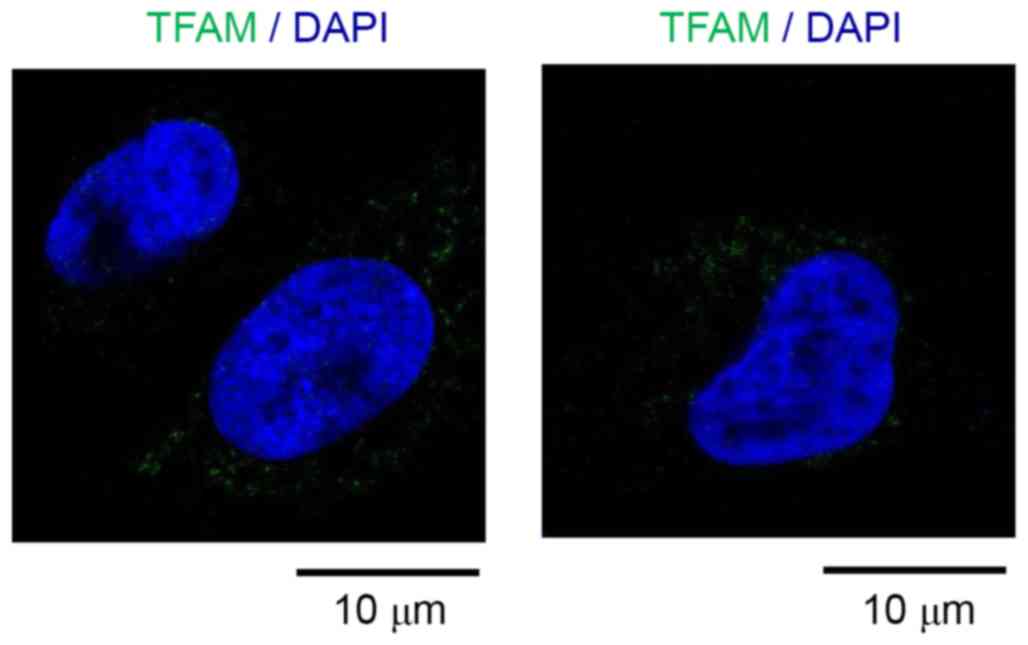
In order to investigate the function of TFAM, the
subcellular localization of TFAM were monitored in U251-MG cells by
using immunocytochemistry. Interestingly TFAM is located to the
dot-like structure close to nucleus, probably mitochondria
(Fig. 3), suggesting that the TFAM
is mainly located to mitochondria in glioblastoma cells.
Positive relationship between high
TFAM expression and glioblastoma tissue specimens
In order to further elucidate the previous results
(Figs. 1 and 2) in clinical setting,
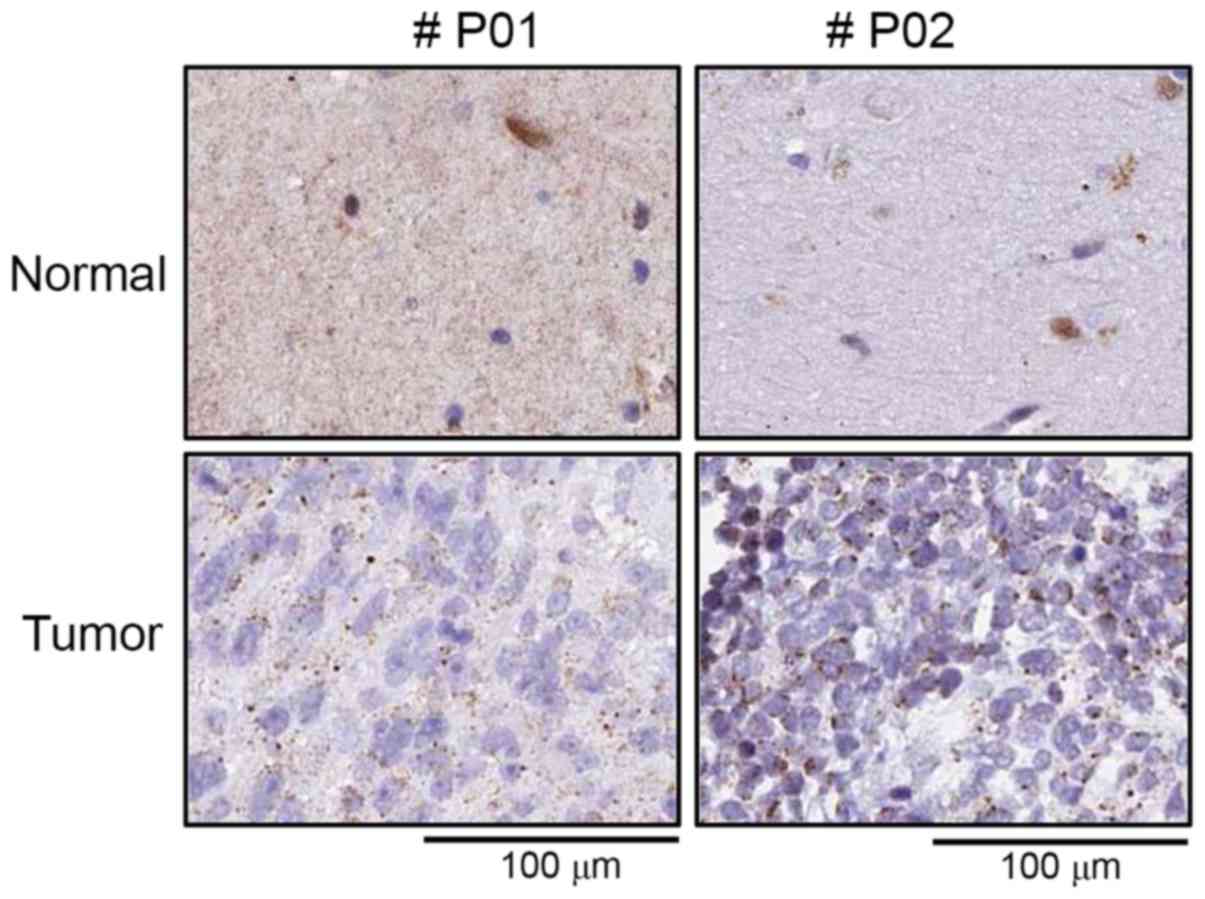
immunohistochemical analysis with human cancer tissue array were
employed. As shown in Fig. 4,
tumor tissue were well stained with anti-TFAM antibody compared to
surrounding normal tissues, indicating that TFAM expression is
upregulated in glioblastoma patients.
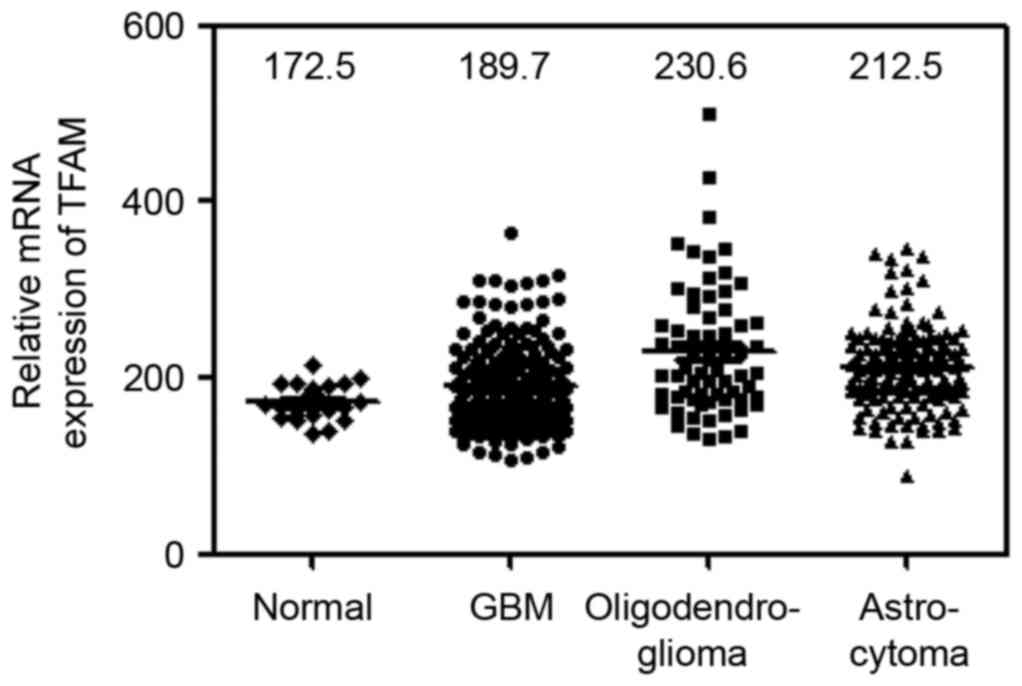
The differential mRNA expression and
prognostic value of TFAM was further validated in the REMBRANDT
cohort
To further evaluate the previous observation
(Figs. 1–3), the differential expression and
prognostic significance of TFAM in glioma of REMBRANDT cohort
(http://www.betastasis.com/glioma/rembrandt/) were
analyzed. Consistent with above mentioned analysis including tissue
microarray, western blotting and RNA-sequencing, TFAM gene
expression was markedly increased in various glioma (GBM, n=214;
oligodendroglioma, n=66; astrocytomas, n=145) than in normal
controls (n=21; one way ANOVA P<0.0001; Fig. 5).
Discussion
Glioblastoma multiform (GBM) is the most aggressive
and common brain tumor, accounts for about 30% of all brain and
central nervous system tumors with poor prognosis. During the past
10 years, the 5-year survival rate of glioblastoma patients was
only 2%, and the average survival rate was 1 year (22). However, the diagnosis for
glioblastoma has not developed for a certain period of time, due to
the resistance against radiotherapy and chemotherapy (23). Therefore, the putative analytical
biomarkers and potential molecular targets for gliomas are
immediately required to deal with this disease.
In the present study, we have identified TFAM as a
new prognostic marker for GBM, which was upregulated in GBM cell
line and patients tissues. It has been well established that TFAM
is required for mtDNA replication and transcription (14). TFAM has been reported to
participate in the regulation of cell survival, proliferation and
migration. Vascular smooth muscle cell proliferation is dependent
on the upregulation of TFAM expression in damaged rat carotid
artery (24). In addition, TFAM
regulates p21 (WAF1/CIP1), a critical regulator of cell cycle
progression, since TFAM depletion induced p21-dependent G1 cell
cycle arrest (25). Moreover, TFAM
is associated to the development and progression of malignant
tumors. In this regard, TFAM mutation induced mtDNA depletion and
apoptotic resistance in colorectal cancer (26). However, TFAM function in glioma is
poorly understood. Therefore we have investigated if TFAM is
potentially involved in brain tumor including GBM. Interestingly,
TFAM was highly expressed in glioblastoma cell lines, including
U343-MG cell and U373-MG cell (Fig.
1A). Analysis of qRT-PCR with TFAM-specific primer have
indicated that U343-MG cell and U373-MG cells showed the higher
expression of TFAM mRNA (Fig. 1B)
compared to U87-MG cells. In addition, RNA deep-sequencing of
U87-MG cell and U251-MG cells provide the evidence that FPKM of
TFAM mRNA is higher in these cells compared to in normal brain
(Fig. 2).
In order to further evaluate the possible function
of TFAM in brain tumor, subcellular localization of TFAM were
examined in U251-MG cells. As expected, TFAM is located to the
dot-like structure close to nucleus, probably mitochondria
(Fig. 3), suggesting that the
mitochondria-located function of TFAM is essential for its role in
glioblastoma cells. To further investigate the central role of TFAM
in GBM development, tissue micro array were performed. As shown in
Fig. 4, TFAM was upregulated in
glioblastoma sample. By using clinical data from REMBRANDT cohort,
TFAM is greatly expressed in all glioma including GBM (Fig. 5), clearly indicating that TFAM
expression is somehow associated to the clinical prognosis of
glioma. Taken together TFAM is appeared to be associated to glioma
development in addition to be a new prognostic tool for
glioblastoma.
Acknowledgements
This study was financially supported by research
fund of Chungnam National University in 2014 (Jongsun Park) and by
the National Research Foundation of Korea (NRF) grant funded by the
Korea Government (MEST) (nos. NRF-2012 M3A9B6055302,
NRF-2014R1A1A3050752, NRF-2015 R1A2A2A01003597 and
NRF-2015R1D1A3A01015694).
References
|
1
|
Friedmann-Morvinski D, Bushong EA, Ke E,
Soda Y, Marumoto T, Singer O, Ellisman MH and Verma IM:
Dedifferentiation of neurons and astrocytes by oncogenes can induce
gliomas in mice. Science. 338:1080–1084. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Arshad A Arif: S.H: EGFR and PTEN gene
mutation status in glioblastoma patients and their prognostic
impact on patient's survival. J Carcinog Muta. 6:2015.
|
|
3
|
Virk SM, Gibson RM, Quinones-Mateu ME and
Barnholtz-Sloan JS: Identification of variants in primary and
recurrent glioblastoma using a cancer-specific gene panel and whole
exome sequencing. PLoS One. 10:e01241782015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bergers G and Benjamin LE: Tumorigenesis
and the angiogenic switch. Nat Rev Cancer. 3:401–410. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wesseling P, Ruiter DJ and Burger PC:
Angiogenesis in brain tumors; pathobiological and clinical aspects.
J Neurooncol. 32:253–265. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jain RK, di Tomaso E, Duda DG, Loeffler
JS, Sorensen AG and Batchelor TT: Angiogenesis in brain tumours.
Nat Rev Neurosci. 8:610–622. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Das S and Marsden PA: Angiogenesis in
glioblastoma. N Engl J Med. 369:1561–1563. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Carmeliet P and Jain RK: Molecular
mechanisms and clinical applications of angiogenesis. Nature.
473:298–307. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Norden AD, Drappatz J and Wen PY: Novel
anti-angiogenic therapies for malignant gliomas. Lancet Neurol.
7:1152–1160. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chinot OL, Macdonald DR, Abrey LE,
Zahlmann G, Kerloëguen Y and Cloughesy TF: Response assessment
criteria for glioblastoma: Practical adaptation and implementation
in clinical trials of antiangiogenic therapy. Curr Neurol Neurosci
Rep. 13:3472013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chinot OL, Wick W, Mason W, Henriksson R,
Saran F, Nishikawa R, Carpentier AF, Hoang-Xuan K, Kavan P, Cernea
D, et al: Bevacizumab plus radiotherapy-temozolomide for newly
diagnosed glioblastoma. N Engl J Med. 370:709–722. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gilbert MR, Dignam JJ, Armstrong TS, Wefel
JS, Blumenthal DT, Vogelbaum MA, Colman H, Chakravarti A, Pugh S,
Won M, et al: A randomized trial of bevacizumab for newly diagnosed
glioblastoma. N Engl J Med. 370:699–708. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ekstrand MI, Falkenberg M, Rantanen A,
Park CB, Gaspari M, Hultenby K, Rustin P, Gustafsson CM and Larsson
NG: Mitochondrial transcription factor A regulates mtDNA copy
number in mammals. Hum Mol Genet. 13:935–944. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Picca A and Lezza AM: Regulation of
mitochondrial biogenesis through TFAM-mitochondrial DNA
interactions: Useful insights from aging and calorie restriction
studies. Mitochondrion. 25:67–75. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cheng Z and Ristow M: Mitochondria and
metabolic homeostasis. Antioxidants & redox signaling.
19:240–242. 2013. View Article : Google Scholar
|
|
16
|
Kim AY, Kwak JH, Je NK, Lee YH and Jung
YS: Epithelial-mesenchymal transition is associated with acquired
resistance to 5-fluorocuracil in HT-29 colon cancer cells. Toxicol
Res. 31:151–156. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li Y, Park J, Piao L, Kong G, Kim Y, Park
KA, Zhang T, Hong J, Hur GM, Seok JH, et al: PKB-mediated PHF20
phosphorylation on Ser291 is required for p53 function in DNA
damage. Cell Signal. 25:74–84. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim IS, Yang SY, Han JH, Jung SH, Park HS
and Myung CS: Differential gene expression in GPR40-overexpressing
pancreatic β-cells treated with linoleic acid. Korean J Physiol
Pharmacol. 19:141–149. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bioinformatics UG: 2015, Human genome
sequencing consortium. http://genome.ucsc.eduApril 17–2016
|
|
20
|
Kim D and Salzberg S: 2015, TopHat 2.1.0
release 6/29/2015. https://ccb.jhu.edu/software/tophat/index.shtml
|
|
21
|
Na CH, Hong JH, Kim WS, Shanta SR, Bang
JY, Park D, Kim HK and Kim KP: Identification of protein markers
specific for papillary renal cell carcinoma using imaging mass
spectrometry. Mol Cells. 38:624–629. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jiang J, Yang J, Wang Z, Wu G and Liu F:
TFAM is directly regulated by miR-23b in glioma. Oncol Rep.
30:2105–2110. 2013.PubMed/NCBI
|
|
23
|
Chargari C, Moncharmont C, Lévy A, Guy JB,
Bertrand G, Guilbert M, Rousseau C, Védrine L, Alphonse G, Toillon
RA, et al: Cancer stem cells, cornerstone of radioresistance and
perspectives for radiosensitization: Glioblastoma as an example.
Bull Cancer. 99:1153–1160. 2012.(In French). PubMed/NCBI
|
|
24
|
Yoshida T, Azuma H, Aihara K, Fujimura M,
Akaike M, Mitsui T and Matsumoto T: Vascular smooth muscle cell
proliferation is dependent upon upregulation of mitochondrial
transcription factor A (mtTFA) expression in injured rat carotid
artery. Atherosclerosis. 178:39–47. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim AJ, Jee HJ, Song N, Kim M, Jeong SY
and Yun J: p21 (WAF1/C1P1) deficiency induces mitochondrial
dysfunction in HCT116 colon cancer cells. Biochem Biophys Res
Commun. 430:653–658. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Guo J, Zheng L and Liu W, Wang X, Wang Z,
Wang Z, French AJ, Kang D, Chen L, Thibodeau SN and Liu W: Frequent
truncating mutation of TFAM induces mitochondrial DNA depletion and
apoptotic resistance in microsatellite-unstable colorectal cancer.
Cancer Res. 71:2978–2987. 2011. View Article : Google Scholar : PubMed/NCBI
|