Introduction
Photo-aging of skin primarily refers to ultraviolet
(UV) irradiation-induced skin aging, which generally manifests as
exposed skin laxity, sagging, wrinkles, pigmentation, dryness,
roughness, telangiectasia and the appearance of large pores
(1). Primary histological features
of photo-aged skin include reduction of collagen and accumulation
of abnormal elastic fibers (2). UV
irradiation may induce abundant secretion of matrix
metalloproteinases (MMPs) from fibroblasts and keratinocytes,
particularly matrix metalloproteinase-1 (MMP-1), which results in
abnormal degradation of the extracellular matrix (1). MMP-1 expression during photo-aging is
primarily controlled by the mitogen-activated protein kinase (MAPK)
signaling pathway (3). A previous
study confirmed that UV-induced reactive oxygen species (ROS)
promote MMP-1 expression and induce the occurrence of photo-aging
by upregulating phosphorylation levels of the MAPK signaling
pathway elements c-Jun N-terminal kinase (JNK), extracellular
signal-regulated kinase (ERK) and p38 (4).
Long non-coding RNA (lncRNA) transcripts that are
>200 bp in length, which do not appear to exhibit coding
capacity, regulate gene expression at transcriptional,
post-transcriptional and translational levels (5,6).
Metastasis-associated lung adenocarcinoma transcript 1 (MALAT1), an
evolutionarily conserved lncRNA expressed in a wide range of
species and tissues, exhibits biological functions including
nuclear organization and architecture, gene splicing, and
epigenetic modulation of gene expression (7,8).
MALAT1 is important in the occurrence and development of cancer,
cardiovascular and diabetes-associated microvascular disease
(9–11). It has previously been demonstrated
that MALAT1 interacts with the MAPK signaling pathway. MALAT1 may
activate ERK/MAPK signaling to promote proliferation and metastasis
of gallbladder cancer cells (12).
Following silencing MALAT1 expression, the p38/MAPK signaling
pathway has been demonstrated to be downregulated in endothelial
cells (10). Activation of MAPK
signaling exerts a key effect on photo-aging, and the present study
aimed to investigate the role of MALAT1 in UV-induced photo-aging
and any potential interaction with the MAPK signaling pathway
elements.
The present study is the first, to the best of the
authors' knowledge, to demonstrate that 60 mJ/cm2 UVB
irradiation induces high expression of MALAT1 in fibroblasts.
MALAT1 siRNA potentially suppressed UVB-induced MMP-1 secretion and
fibroblast senescence by inhibiting activation of the ERK/MAPK
signaling pathway.
Materials and methods
Primary culture of fibroblasts
Foreskin tissue was donated (with signed informed
consent) by a healthy young male following circumcision. The
donor's health status was established through physical examinations
and laboratory investigations which revealed no obvious
abnormalities. The sample was immersed in an iodine complex for 15
min and washed three times with phosphate buffered saline (PBS).
Following removal of subcutaneous tissue, the sample was cut into
pieces and digested with trypsin to isolate fibroblasts.
Fibroblasts were collected, washed, and then incubated with
high-glucose Dulbecco's Modified Eagle's medium (DMEM; Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with
10% fetal bovine serum (HyClone; GE Healthcare Life Sciences,
Logan, UT, USA). Cells in logarithmic phase from the third to
eighth passage were used for experiments.
Ultraviolet B (UVB) irradiation
Cells that had reached ~70% confluence were
irradiated with UVB using a SS-07 light therapy device (36 W power;
time of irradiation automatically adjusted to dose of irradiation;
cat. no. 1,047,469; Shanghai Sigma Hi-Tech Co., Ltd., Shanghai,
China). Prior to irradiation, culture medium was removed from
cells, which were then washed twice with PBS. Fibroblasts, coated
with a thin layer of PBS, were vertically irradiated with UVB
immediately following opening the culture dish cover. The distance
of irradiation was 20 cm. Complete medium was added to the culture
dish immediately following irradiation and the culture dish was
then incubated in the original culture environment.
Chemical treatments
MALAT1 siRNA (5′-CACAGGGAAAGCGAGTGGTTGGTAA-3′) and
the negative control non-targeting siRNA were designed and
synthesized by GeneChem Co., Ltd. (Shanghai, China). When cells
reached 30–50% confluence, siRNAs (50 nM) were transfected into
cells using Lipofectamine® 3000 (Thermo Fisher
Scientific, Inc.,) in serum-free DMEM according to the
manufacturer's protocol. Following 6 h of incubation, cells were
returned to normal medium in the incubator. When cells reached 70%
confluence, they were exposed to irradiation. Cells were pretreated
with 10 mM N-acetyl-L-cysteine (NAC; Beyotime Institute of
Biotechnology, Haimen, China) 1 h prior to irradiation. Following
this, cells were supplemented with 10 mM NAC for 24 h.
Cell proliferation
Cells were seeded into a 96-well plate
(104 cells/well). Confluent cells were treated with
serum-free medium for 24 h and irradiated with UVB. Following
irradiation, cells were incubated with normal medium for 24 h and
then treated with 10 µl Cell Counting kit (CCK)-8 (Dojindo
Molecular Technologies, Inc., Kumamoto, Japan) for 4 h. Absorbance
values were measured at a wavelength of 450 nm using a microplate
reader (GEN5 CHS v.2.00, BioTek Instruments, Inc., Winooski, VT,
USA).
RT-qPCR
Total RNA was extracted from cells using
TRIzol® reagent (Thermo Fisher Scientific, Inc.).
Extracted total RNA, following purification and quality
assessments, was reverse-transcribed to cDNA. RT-qPCR was performed
to detect MALAT1 expression using the SYBR-Green PCR Master Mix
(Applied Biosystems; Thermo Fisher Scientific, Inc.) with a ABI
PRISM 7700 system (Applied Biosystems; Thermo Fisher Scientific,
Inc.). The thermocycling conditions were as follows: Initial
denaturation at 95°C for 2 min, then denaturation at 95°C for 15
sec, annealing at 56°C for 15 sec and extension at 68°C for 20 sec,
for a total of 40 cycles. β-actin served as an internal reference.
The expression level of MALAT1 was calculated using the
2−∆∆Cq method (13)
method. The primer sequences used were as follows: Forward,
5′-AAAGCAAGGTCTCCCCACAAG-3′ and reverse,
5′-GGTCTGTGCTAGATCAAAAGGCA-3′ for MALAT1. Experiments were
performed in triplicate.
Enzyme-linked immunosorbent assay
(ELISA) for MMP-1 expression in supernatants
Following irradiation, cells were incubated with
serum-free medium for 24 h. Cell culture medium was collected and
centrifuged at 1,000 × g/min for 10 min at room temperature,
and then supernatants were harvested. MMP-1 concentration was
detected in supernatants using the MMP-1 kit (cat. no. EK0458;
Boster Systems, Inc., Pleasanton, CA, USA) according to the
manufacturer's protocol.
Detection of senescent cells using a
β-galactosidase (SA-β-Gal) kit
Following 24 h of UVB irradiation, cells were
stained using an SA-β-Gal staining kit (Beyotime Institute of
Biotechnology) according to the manufacturer's protocol. Senescent
cells presented with a blue color around the nuclei and the
percentage of senescent cells was equal to number of cells stained
blue/total number of cells ×100%.
Detection of ROS content using flow
cytometry
Following 24 h of UVB irradiation, cells were
treated with serum-free medium containing 10 µM
dichloro-dihydro-fluorescein diacetate (DCFH-DA; cat. no. KGT010-1;
Nanjing Keygen Biotech Co., Ltd., Nanjing, China) for 30 min at
37°C, according to the manufacturer's protocol, and then washed
three times with DMEM. Images were captured using a fluorescence
microscope. Cells were collected and the level of green
fluorescence was detected to evaluate the levels of intracellular
ROS using flow cytometry (FACSCalibur, BD Biosciences, Franklin
Lakes, NJ, USA) with BD CellQuest Pro software version 6.0 (BD
Biosciences) according to the manufacturer's protocol.
Western blot analysis
Following fibroblast irradiation with UVB for 48 h,
total protein was collected. Expression levels of ERK,
phosphorylated ERK (p-ERK), p38, phosphorylated p38 (p-p38), JNK
and phosphorylated JNK (p-JNK) were measured using western blot
analysis. The process was conducted as previously described
(14). Briefly, the cells were
rinsed with PBS and lysed in radioimmunoprecipitation lysis buffer
(Beyotime Institute of Biotechnology) with a protease and
phosphatase inhibitor cocktail (Thermo Fisher Scientific, Inc.) on
ice for 30 min. Cell lysates were centrifuged at 13,000 × g
at 4°C for 20 min. The supernatants were collected and the proteins
were quantified using a bicinchoninic acid protein assay kit
(Beyotime Institute of Biotechnology). Protein samples (20 µg) were
separated by 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis. Gels were transferred to nitrocellulose membranes,
blocked with 5% non-fat-milk and then probed with anti-ERK
(dilution, 1:1,000; Cell Signaling Technology, Inc., Danvers, MA,
USA; cat. no. 9926), anti-p-ERK (dilution, 1:1,000; Cell Signaling
Technology, Inc.; cat. no. 9910), anti-p38 (dilution, 1:1,000; Cell
Signaling Technology, Inc.; cat. no. 9926), anti-p-p38 (dilution,
1:1000; Cell Signaling Technology, Inc.; cat. no. 9910), anti-JNK
(dilution, 1:1,000; Cell Signaling Technology, Inc.; cat. no.
9926), or anti-p-JNK (dilution, 1:1,000; Cell Signaling Technology,
Inc.; cat. no. 9910) antibody at 4°C overnight. Following
incubation with a rabbit horseradish peroxidase-conjugated
secondary antibody (dilution, 1:2,000; Abcam, Cambridge, UK; cat.
no. ab6721) at room temperature for 2 h, immunoreactive bands were
visualized using a electrochemiluminescence detection reagent
(Yeasen; Shanghai Yi Sheng Biological Technology Co., Ltd.,
Shanghai, China; cat. no. 36208ES60) according to the
manufacturer's instructions. Integrated optical density (IOD) was
calculated using ImageJ version 1.46 software (http://rsb.info.nih.gov/ij). The relative intensity of
the level of the protein of interest was calculated using the
following formula: IODphosphorylated protein band of
interest/IODcorresponding total protein band.
Representative blots of at least three independent experiments are
presented.
Statistical analysis
Data were analyzed using SPSS software, version 22.0
(IBM SPSS, Armonk, NY, USA) and expressed as the mean ± standard
deviation. All experiments were performed at least in triplicate.
Significance tests were conducted on the data groups using analysis
of variance followed by a comparison between the specific groups
using a Student-Newman-Keuls test and analysis of differences
between only two groups was performed using an unpaired Student's
t-test. P<0.05 was considered to indicate a statistically
significant difference.
Results
MALAT1 expression increases following
UVB irradiation
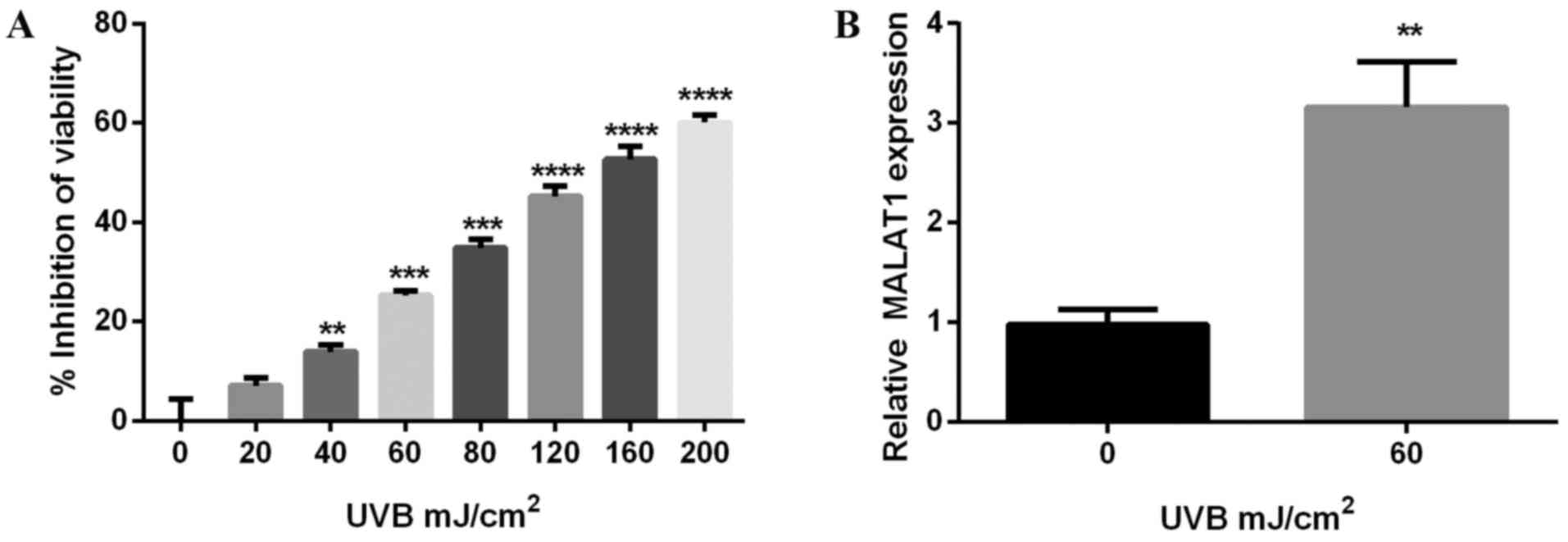
To investigate the phototoxicity of UVB on
fibroblasts, cell viability was detected using the CCK-8 method, on
cells irradiated with various doses of UVB for 24 h. The results
demonstrated UVB suppressed melanocyte viability, an inhibitory
effect enhanced by increasing doses of UVB. The inhibitory rate of
60 mJ/cm2 UVB was ~25% (P<0.001; Fig. 1A) and results further demonstrated
that 60 mJ/cm2 UVB increased MALAT1 expression
(P<0.01; Fig. 1B).
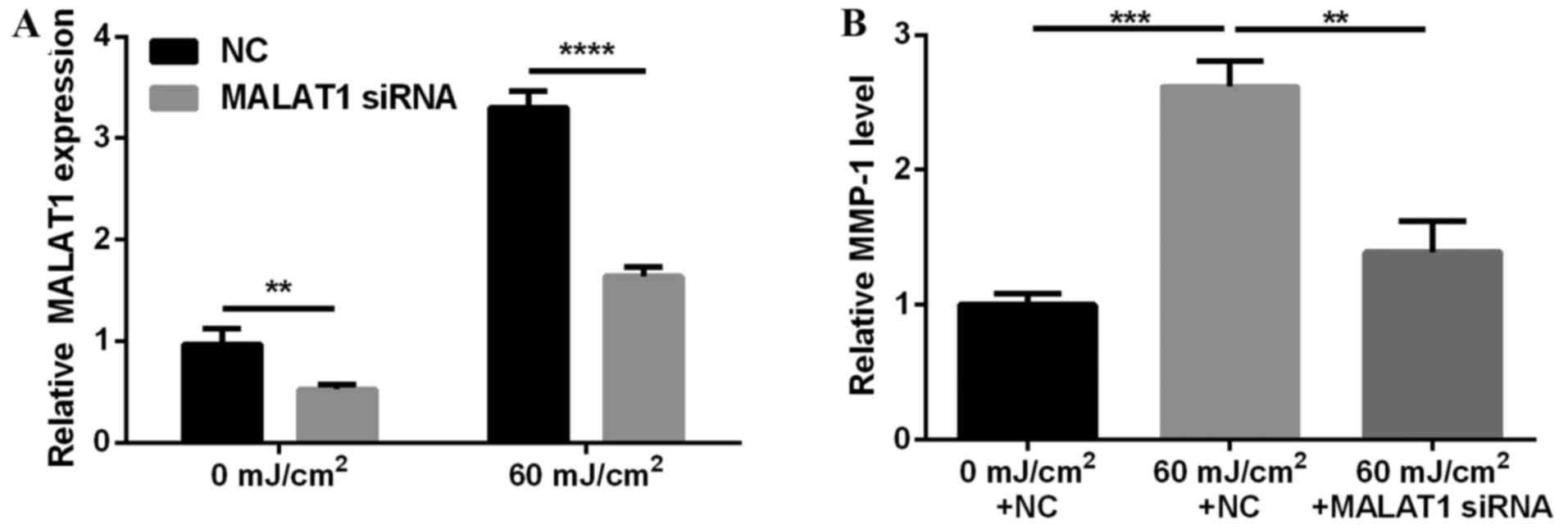
MALAT1 siRNA inhibits UVB-induced
MMP-1 secretion
To investigate the effects of MALAT1 on photo-aging,
MALAT1 expression in fibroblasts was silenced and effects on MMP-1
secretion observed. The results demonstrated that MALAT1 siRNA
suppressed MALAT1 expression (P<0.01 and P<0.0001; Fig. 2A) and inhibited 60
mJ/cm2 UVB-induced MMP-1 secretion (P<0.01 and
P<0.001; Fig. 2B).
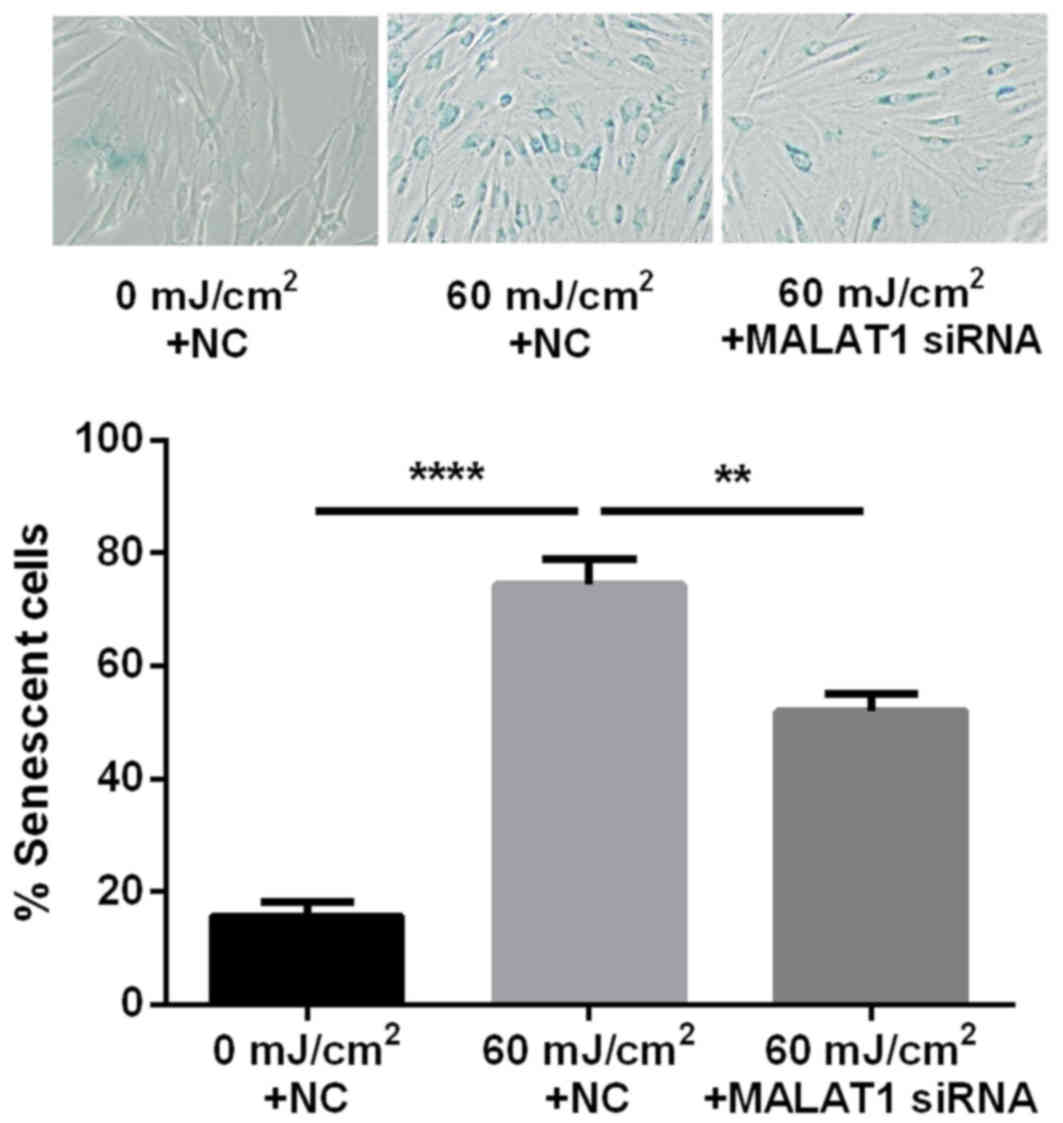
MALAT1 siRNA inhibits UVB-induced
fibroblast senescence
The ratio of senescent cells was markedly greater
following UVB irradiation (74.4%) compared with cells that had not
been exposed to irradiation (15.7%). In addition, the ratio of
senescent cells was significantly lower following intervention with
MALAT1 siRNA (52.1%) compared with the UVB irradiation group
(P<0.01 and P<0.0001; Fig.
3).
MALAT1 siRNA inhibits UVB-induced ERK
phosphorylation in fibroblasts
Activation of MAPK signaling is important in
photo-aging. The present study investigated the effect of the
silencing of MALAT1 on activation of MAPK signaling pathway
elements, induced by UVB. The results demonstrated that 60
mJ/cm2 UVB upregulated ERK, JNK and p38 phosphorylation
levels, however MALAT1 siRNA inhibited UVB-induced ERK
phosphorylation (P<0.01), with no significant influence on JNK
and p38 phosphorylation levels (P>0.05; Fig. 4).
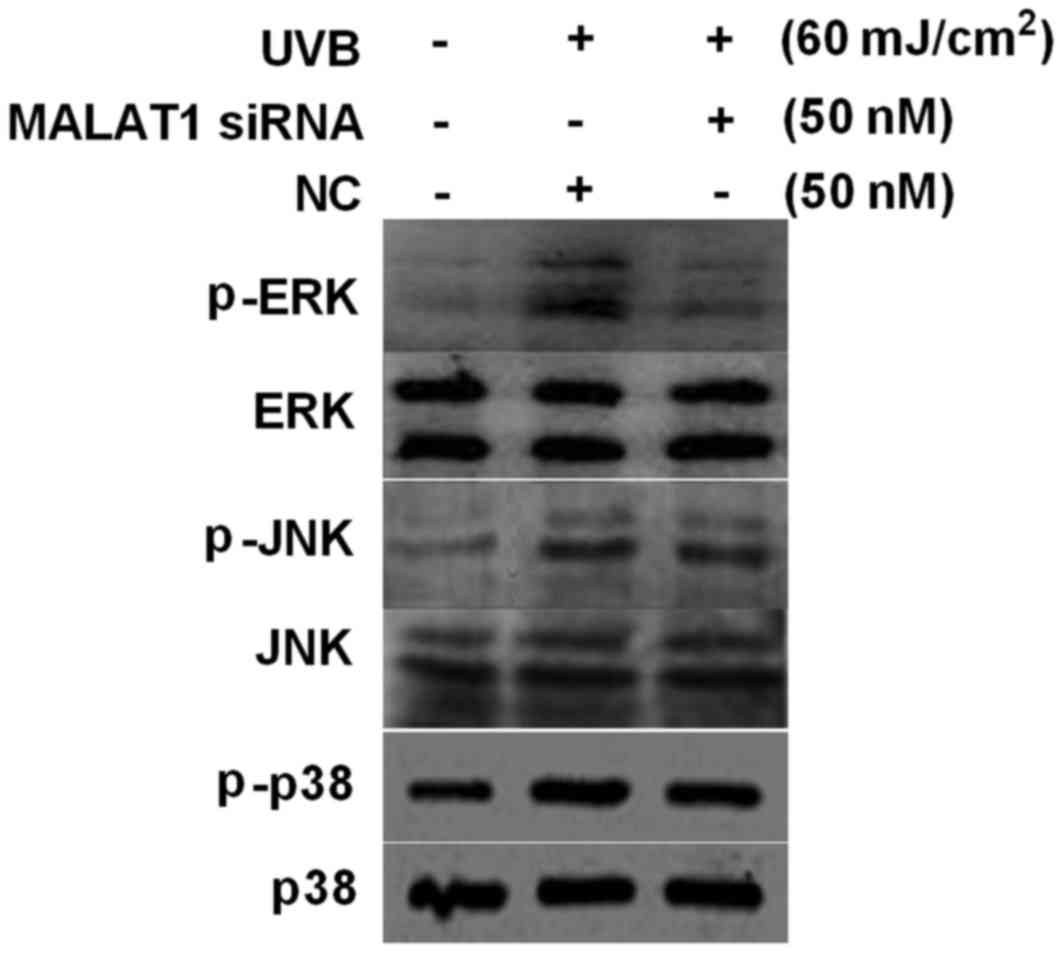 | Figure 4.MALAT1 siRNA suppresses UVB-induced
p-ERK activation in fibroblasts. Effect of 60 mJ/cm2 UVB
irradiation and MALAT1 siRNA on fibroblast p-ERK, p-p38 and p-JNK
expression levels, detected by western blot analysis. MALAT1,
metastasis-associated lung adenocarcinoma transcript 1; siRNA,
small interfering; UVB, ultraviolet B; NC, negative control; p,
phosphorylated; ERK, extracellular signal-regulated kinase, JNK,
c-Jun N-terminal kinase. |
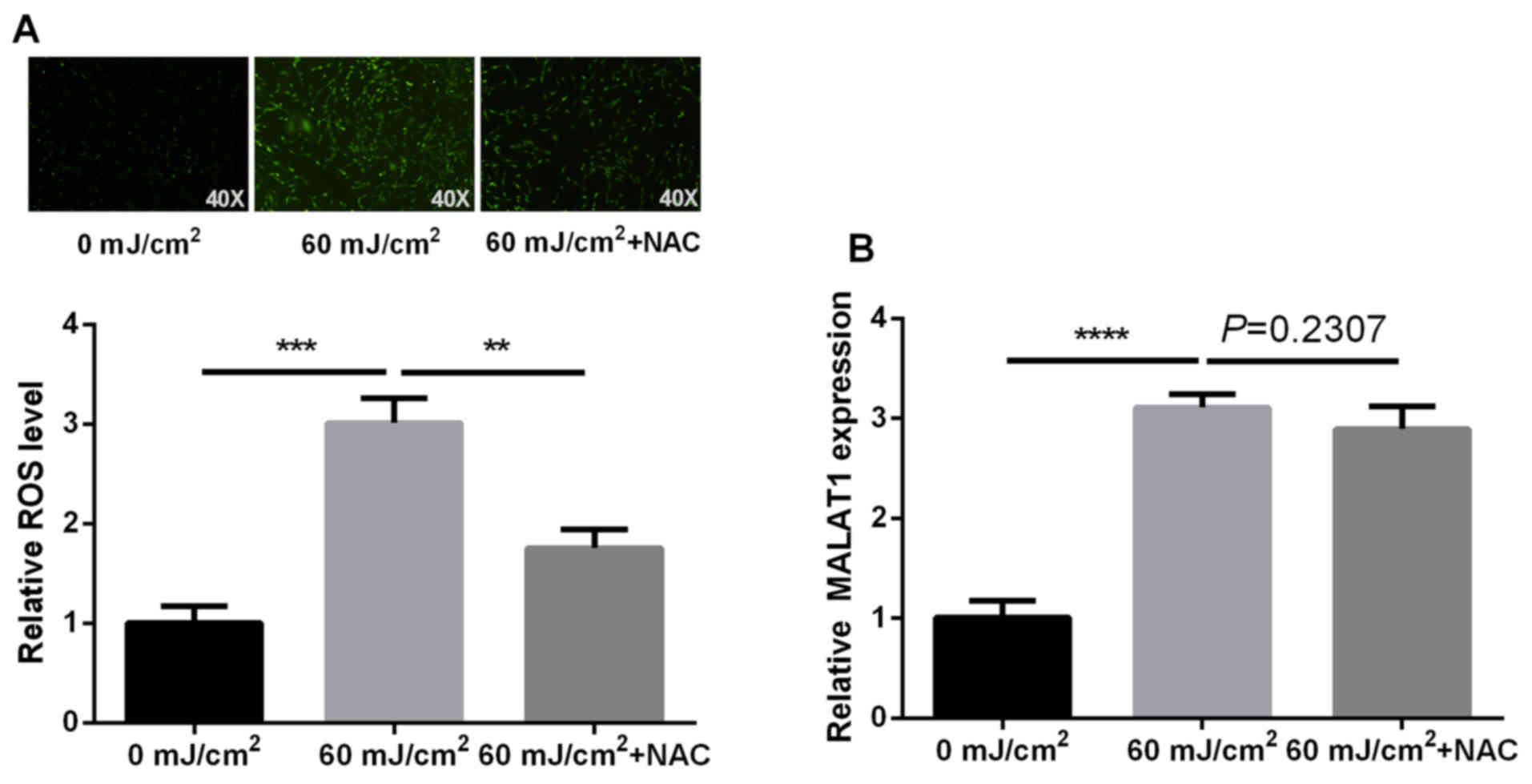
UVB-induced MALAT1 expression is
independent of ROS generation
ROS alters the expression levels of numerous
molecules. The present study examined whether ROS underlies
UVB-induced MALAT1 alteration using NAC, which is a ROS scavenger.
The effects on MALAT1 expression in NAC-pretreated cells were
investigated. The results indicated an increase in ROS content
following fibroblast irradiation with UVB, which was then inhibited
by NAC. However, NAC did not have a significant effect on MALAT1
expression (P=0.2307). These findings indicated that UVB-induced
MALAT1 expression is not dependent on ROS generation (P<0.01,
P<0.001 and P<0.0001 respectively; Fig. 5).
Discussion
It has recently been demonstrated that microRNA
expression profiles within non-coding RNA alter in fibroblasts
following administration of UV irradiation (15). An additional study verified that
two microRNAs, miR-146a and miR-155, potentially regulate
photo-aging via targeting Smad4 and c-Jun regulation
(16,17). The role of lncRNAs in UV-induced
photo-aging of skin remains to be elucidated.
The potential of UV radiation to alter lncRNA
expression profiles within skin cells has implications on numerous
important biological functions associated with lncRNAs. Hall et
al (18) demonstrated that
UVB-induced lincRNA-p21 is important in keratinocyte apoptosis. The
authors previously demonstrated that >two-fold alterations in
the expression of 807 lncRNAs occurred, following melanocyte
irradiation with UVB (14),
including a markedly increased expression of MALAT1 (data not
shown). MALAT1, a well-known cancer-promoting gene within lncRNAs,
regulates numerous biological activities including tumor
proliferation, metastasis and epithelial-mesenchymal transitioning
(19). The current study revealed
increases in MALAT1 expression following UVB irradiation of
fibroblasts, indicating a potentially important role of MALAT1 in
the photo-aging of fibroblasts.
MMP-1 is an important indicator of photo-aging
within fibroblasts. In the present study, MMP-1 content increased
within fibroblasts following 60 mJ/cm2 UVB irradiation,
suggesting aging of fibroblasts. MMP-1 secretory volume was
markedly reduced in photo-aged fibroblasts following silencing of
MALAT1 expression, indicating MALAT1 l promoted photo-aging of
fibroblasts. Cell senescence staining experiments confirmed the
ability of MALAT1 siRNA to suppress UVB-induced photo-aging of
fibroblasts.
The MAPK signaling pathway is activated during
photo-aging of fibroblasts (4).
Furthermore, recent studies have demonstrated an interaction
between MALAT1 and the MAPK signaling pathway (10,12).
Therefore, the present study explored the effects of MALAT1 on MAPK
signaling in fibroblasts. The results demonstrated MALAT1 siRNA
inhibited UVB-induced p-ERK activation, indicating MALAT1 may
participate in the photo-aging of fibroblasts via activation of the
ERK/MAPK signaling pathway elements.
The authors previously demonstrated that UVB-induced
lncRNAs (including lnc-CD1D-2:1) depend on ROS generation (14), and therefore, further proceeded to
investigate whether UVB-induced MALAT1 upregulation is
ROS-dependent. The results demonstrated no inhibition of MALAT1
expression by NAC (a ROS scavenger), indicating UVB-induced MALAT1
upregulation is independent of ROS. In conclusion, MALAT1 may
participate in UVB-induced photo-aging via regulation of the
ERK/MAPK signaling pathway elements and UVB-induced MALAT1
upregulation does not depend on ROS generation.
Acknowledgements
The present study was supported by the Science and
Technology Plan of Hunan province (grant no. 2013SK3056) and the
New Xiangya Talent Projects of the Third Xiangya Hospital of
Central South University (grant no. JY201623).
Glossary
Abbreviations
Abbreviations:
|
DCFH-DA
|
dichloro-dihydro-fluorescein
diacetate
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
|
ERK
|
extracellular signal-regulated
kinase
|
|
JNK
|
c-Jun N-terminal kinase
|
|
lncRNA
|
long non-coding RNA
|
|
MALAT1
|
metastasis-associated lung
adenocarcinoma transcript 1
|
|
MMP-1
|
metalloproteinase-1
|
|
MAPK
|
mitogen-activated protein kinase
|
|
NAC
|
N-acetyl-L-cysteine
|
|
PBS
|
phosphate-buffered saline
|
|
P-ERK
|
phosphorylated ERK
|
|
P-JNK
|
phosphorylated JNK
|
|
P-p38
|
phosphorylated p38
|
|
ROS
|
reactive oxygen species
|
|
siRNA
|
small interference RNA
|
|
UVB
|
ultraviolet B
|
References
|
1
|
Ham SA, Kang ES, Lee H, Hwang JS, Yoo T,
Paek KS, Park C, Kim JH, Lim DS and Seo HG: PPARδ inhibits
UVB-induced secretion of MMP-1 through MKP-7-mediated suppression
of JNK signaling. J Invest Dermatol. 133:2593–2600. 2013.
View Article : Google Scholar
|
|
2
|
Stoebner PE and Meunier L: Photoaging of
face. Ann Dermatol Venereol. 135:1S21–1S26. 2008.(In French).
|
|
3
|
Park G, Baek S, Kim JE, Lim TG, Lee CC,
Yang H, Kang YG, Park JS, Augustin M, Mrosek M, et al: Flt3 is a
target of coumestrol in protecting against UVB-induced skin
photoaging. Biochem Pharmacol. 98:473–483. 2015. View Article : Google Scholar
|
|
4
|
Kim MJ, Woo SW, Kim MS, Park JE and Hwang
JK: Anti-photoaging effect of aaptamine in UVB-irradiated human
dermal fibroblasts and epidermal keratinocytes. J Asian Nat Prod
Res. 16:1139–1147. 2014. View Article : Google Scholar
|
|
5
|
He X, Bao W, Li X, Chen Z, Che Q, Wang H
and Wan XP: The long non-coding RNA HOTAIR is upregulated in
endometrial carcinoma and correlates with poor prognosis. Int J Mol
Med. 33:325–332. 2014.
|
|
6
|
Li Y and Wang X: Role of long noncoding
RNAs in malignant disease (Review). Mol Med Rep. 13:1463–1469.
2016.
|
|
7
|
Xu S, Sui S, Zhang J, Bai N, Shi Q, Zhang
G, Gao S, You Z, Zhan C, Liu F and Pang D: Downregulation of long
noncoding RNA MALAT1 induces epithelial-to-mesenchymal transition
via the PI3K-AKT pathway in breast cancer. Int J Clin Exp Pathol.
8:4881–4891. 2015.
|
|
8
|
Dey BK, Mueller AC and Dutta A: Long
non-coding RNAs as emerging regulators of differentiation,
development, and disease. Transcription. 5:e9440142014. View Article : Google Scholar :
|
|
9
|
Uchida S and Dimmeler S: Long noncoding
RNAs in cardiovascular diseases. Circ Res. 116:737–750. 2015.
View Article : Google Scholar
|
|
10
|
Liu JY, Yao J, Li XM, Song YC, Wang XQ, Li
YJ, Yan B and Jiang Q: Pathogenic role of lncRNA-MALAT1 in
endothelial cell dysfunction in diabetes mellitus. Cell Death Dis.
5:e15062014. View Article : Google Scholar :
|
|
11
|
Wei Y and Niu B: Role of MALAT1 as a
prognostic factor for survival in various cancers: A systematic
review of the literature with meta-analysis. Dis Markers.
2015:1646352015. View Article : Google Scholar :
|
|
12
|
Wu XS, Wang XA, Wu WG, Hu YP, Li ML, Ding
Q, Weng H, Shu YJ, Liu TY, Jiang L, et al: MALAT1 promotes the
proliferation and metastasis of gallbladder cancer cells by
activating the ERK/MAPK pathway. Cancer Biol Ther. 15:806–814.
2014. View Article : Google Scholar :
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
14
|
Zeng Q, Wang Q, Chen X, Xia K, Tang J,
Zhou X, Cheng Y, Chen Y, Huang L, Xiang H, et al: Analysis of
lncRNAs expression in UVB-induced stress responses of melanocytes.
J Dermatol Sci. 81:53–60. 2016. View Article : Google Scholar
|
|
15
|
Syed DN, Khan MI, Shabbir M and Mukhtar H:
MicroRNAs in skin response to UV radiation. Curr Drug Targets.
14:1128–1134. 2013. View Article : Google Scholar :
|
|
16
|
Li W, Zhou BR, Hua LJ, Guo Z and Luo D:
Differential miRNA profile on photoaged primary human fibroblasts
irradiated with ultraviolet A. Tumour Biol. 34:3491–3500. 2013.
View Article : Google Scholar
|
|
17
|
Song J, Liu P, Yang Z, Li L, Su H, Lu N
and Peng Z: MiR-155 negatively regulates c-Jun expression at the
post-transcriptional level in human dermal fibroblasts in vitro:
Implications in UVA irradiation-induced photoaging. Cell Physiol
Biochem. 29:331–340. 2012. View Article : Google Scholar
|
|
18
|
Hall JR, Messenger ZJ, Tam HW, Phillips
SL, Recio L and Smart RC: Long noncoding RNA lincRNA-p21 is the
major mediator of UVB-induced and p53-dependent apoptosis in
keratinocytes. Cell Death Dis. 6:e17002015. View Article : Google Scholar :
|
|
19
|
Gutschner T, Hammerle M and Diederichs S:
MALAT1-a paradigm for long noncoding RNA function in cancer. J Mol
Med (Berl). 91:791–801. 2013. View Article : Google Scholar
|