Introduction
Breast cancer is a common malignant tumor in women,
with ~1,700,000 cases and 521,900 cases of mortality in 2012
worldwide (1). The incidence of
breast cancer is progressively increasing, particularly in the
urban regions of China. Official data predicted a continuing
increase in mortality rates in the ensuing 5 years (2). According to current understanding,
tumor metastasis remains the dominant cause for cancer-associated
mortality (3). Therefore, it is
necessary to identify or develop drugs with antimetastatic ability
for breast cancer therapy.
Tumor metastasis is a multi-step process, in which
the key step is the degradation of extracellular matrix (ECM) by
certain enzymes, including matrix metalloproteinases (MMPs) and
heparanase (4,5). Heparanase is a mammalian
endo-D-glucuronidase, which cleaves heparan sulfate (HS) involved
in the formation of ECM. The expression level of this enzyme
correlates with the metastatic potential of tumor cells (6,7). It
has been demonstrated that the overexpression of heparanase results
in intensive angiogenesis, lymph node metastasis, advanced clinical
stage and short overall survival rates in lung, breast, colon and
ovarian cancer (8–11). These poor outcomes can be partly
ascribed to the degradation of ECM by heparanase. The overexpressed
heparanase impairs the structural integrity of ECM. Subsequently,
the degradation of HS chains promotes the release of growth
factors, including fibroblast growth factor (FGF), vascular
endothelial growth factor (VEGF) and platelet-derived growth
factor, from ECM, which activate the downstream signaling pathways,
facilitating the proliferation and metastasis of cancer cells
(12,13). The nonenzymatic function of
heparanase also directly stimulates Akt-dependent endothelial cell
invasion and migration activities (14). Therefore, heparanase is a target of
interest for the prevention of cancer metastasis.
Elemene (ELE) is a natural plant drug extracted from
Curcuma wenyujin. A previous study demonstrated the
extensive spectrum of antitumor effects of ELE, involving lung
cancer, breast cancer, gastric cancer and brain tumors (15). The effects of ELE are not only on
the inhibition of cancer cells, but also on the regulation of the
tumor microenvironment, including inhibition of
epithelial-mesenchymal transition (EMT) (16), decreased angiogenesis (17) and inhibition of ECM degradation by
MMPs (18). As a key enzyme
involved in degrading the ECM in the tumor microenvironment,
whether the expression of heparanase can be inhibited by ELE
remains to be elucidated and requires further investigation.
Low-molecular weight heparin (LMWH), an analog of the natural
substrate of heparanase, is considered a potent inhibitor of
heparanase (7,19,20),
thus, serving as a positive control. In the present study, the
antiproliferative and antimetastatic effects of ELE were confirmed.
In addition, it was found that ELE downregulated the expression of
heparanase and potentially decreased the phosphorylation of
extracellular signal-regulated kinase (ERK) and AKT in 4T1 murine
breast cancer cells.
Materials and methods
Chemicals and reagents
β-elemene (purity, 98%; molecular formula, C15H24;
molecular weight, 204.35) was obtained from Dalian Jingang
Pharmaceuticals, Ltd. (Liaoning, China). The LMWH was purchased
from Aventis Intercontinental (Paris, France).
Primary antibodies against heparanase (cat no.
ab85543) and VEGF (cat no. ab46154) were purchased from Abcam
(Cambridge, UK). The primary antibodies against fibroblast growth
factor (FGF)-2 (cat. no. sc-79) and β-actin (cat no. sc-47778) were
purchased from Santa Cruz Biotechnology, Inc. (Houston, TX, USA).
The primary antibodies against ERK (cat no. #9102), phosphorylated
(p)-ERK (cat no. #4377), AKT (cat no. #9272) and p-AKT (cat no.
#4058) were purchased from Cell Signaling Technology, Inc.
(Danvers, MA, USA). The secondary antibodies, including Dylight
800-conjugated goat anti-mouse (cat no. 072-07-18-06) and Dylight
680-conjugated goat anti-rabbit IgG (cat no. 072-06-15-06) were
purchased from KPL, Inc. (Gaithersburg, MD, USA).
Cell culture
The 4T1 murine breast cancer cells (Cell Bank of the
Chinese Academy of Sciences, Shanghai, China) were cultured in
RPMI-1640 (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA). The MCF-7, MDA-MB-231 and MDA-MB-435S human breast cancer
cell lines were purchased from the Cell Center of Medical Research
Institute, Chinese Academy of Medical Sciences. The MCF-7 cells
were cultured in DMEM (Gibco; Thermo Fisher Scientific, Inc.),
whereas the MA-MB-231 and MDA-MB-435S cells were cultured in
Leibovitz's L15 (Hyclone; GE Healthcare Life Sciences, Logan, UT,
USA). The media was supplemented with 10% fetal bovine serum (FBS;
Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin-streptomycin (Hyclone; GE Healthcare Life Sciences). The
cells were trypsinized with 0.125% trypsin (Gibco) and were seeded
onto microplates for the subsequent experiments. All cell lines
were cultured at 37°C in a humidified incubator (Sanyo, Osaka,
Japan) supplied with 5% carbon dioxide (CO2).
Cell Counting kit-8 (CCK-8)
cytotoxicity assay
The cytotoxicity of ELE was measured using a CCK-8
assay. The cells were inoculated in a 96-well plate (Corning,
Steuben County, NY, USA) at 3×103 cells/well, incubated
overnight at 37°C with 5% CO2, and were exposed to
different concentrations of ELE (10–160 µg/ml) or LMWH (50–800
IU/ml as a positive control) for 24 or 48 h. Following ELE and LMWH
exposure, 15 µl CCK-8 (KeyGEN Biotech Co., Ltd., Nanjing, China)
was added to each of the wells. The cells were incubated for
another 4 h at 37°C with 5% CO2. The absorbance values
were measured at 450 nm using a microplate reader (Thermo Fisher
Scientific, Inc.). The optical density (OD) was determined to
calculate the rate of inhibition, which is expressed as follows:
Inhibition rate=[1-(OD of experimental sample-OD of blank
group)/(OD of control group-OD of blank group)]x100%.
Electric cell-substrate impedance
sensing (ECIS) wound healing assay
The ECIS assay was used to measure the healing
ability of the 4T1 cells (21). A
400 µl suspension of 4T1 with 8×104 cells were seeded in
8W10E+ ECIS arrays (Applied Biophysics, Inc., Troy, NY, USA) in
each well. Following inoculation of the cells, the procedure of
attachment and spreading was exhibited by impedance measurements.
Lethal electroporation (current, 6,500 µA; frequency, 100,000 Hz;
time, 60 sec) was performed when the cells were fully confluent
(~16 h). The dead cells were washed away and fresh medium
containing ELE (25 µg/ml) and LMWH (200 IU/ml) was added. The wound
healing was assessed by continuous impedance measurements for 20 h.
The experiment was performed in a humidified 5% CO2
incubator at 37°C and was repeated three times.
Real-time cell analysis (RTCA)
migration assay
RTCA was used for determining cell migration on an
xCELLigence DP device (Roche Diagnostics, Mannheim, Germany) as
described in the manufacturer's protocol and previous study
(22). The migration assay was
performed on a CIM-plate16 (Roche Diagnostics), comprising a
two-chamber device separated by an aporous membrane. Either cell
attachment or cell migration directly through pores to the lower
surface of the membrane, where electrodes exist, can be recorded. A
10% FBS solution was added to the bottom chamber, and the top
chamber was assembled using the CIM-plate assembly tool. The 4T1
cells treated with ELE (25 µg/ml) and LMWH (200 IU/ml) were
collected and counted, following which 6×104 cells in
100 µl serum-free medium were seeded into the top chamber of the
CIM-plate16. The xCELLigence device recorded the migratory
information from 4T1 cells for 24 h. The cell indices represented
the migration capacity of the 4T1 cells treated with drugs.
Transwell migration and invasion
assay
The 4T1 cells were exposed to ELE at 25 µg/ml and
LMWH at 200 IU/ml for 36 h. The cells were collected in a
serum-free medium at 5×105 cells/ml. A 100 µl suspension
was added to the top chambers, coated with Matrigel (BD
Biosciences, San Diego, CA, USA) with or without growth factors. In
the lower chamber, 600 µl of RPMI-1640 medium containing 10% FBS
was added to the 24-well plate. The cells were cultured at 37°C
with 5% CO2 for 24 h. After 24 h, the cells were fixed
with methanol for 10 min at −20°C. Cells on the apical side of the
top chamber were removed using a cotton swab. Those cells, which
migrated or invaded to the lower side of the membrane were stained
with Hoechst 33342 (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany)
for 15 min at 25°C. The cells were counted under a fluorescence
microscope (magnification, ×200; Leica Microsystems GmbH,
Heidelberg, Germany). Data were collected from three independent
experiments.
Western blot analysis
4T1 cells were cultured in 100 mm dishes and exposed
to ELE at 25 µg/ml and LMWH at 200 IU/ml for 36 h. Subsequently,
the cells were harvested and lysed with radioimmunoprecipitation
assay lysis buffer (Beyotime Institute of Biotechnology, Jiangsu,
China) containing protease inhibitor cocktail set III (Merck KGaA).
The protein content was measured using a bicinchoninic acid protein
assay kit (Pierce; Thermo Fisher Scientific, Inc.). Following
denaturation by boiling, equal amounts of extracted protein samples
(30 µg) were separated by 4–10% SDS-PAGE and transferred onto a
polyvinylidene difluoride membrane. The membrane was blocked with
5% skimmed milk for 2 h at room temperature and probed with the
following primary antibodies at 4°C overnight: Anti-heparanase
rabbit-polyclonal antibody (1:500), anti-FGF-2 rabbit-polyclonal
antibody (1:500), anti-VEGF rabbit-polyclonal antibody (1:1,000),
anti-ERK/p-ERK rabbit-polyclonal antibodies (1:1,000),
anti-AKT/p-AKT rabbit-polyclonal antibodies (1:1,000) and
anti-β-actin (1:1,000), which was followed by incubation with
secondary antibodies (1:10,000) for 1 h at room temperature. The
protein of interest was detected using Odyssey infrared imaging
system (LI-COR Biosciences, Lincoln, NE, USA). Blots were
semi-quantified by densitometric analysis using the Odyssey
software version 3.0 (LI-COR Biosciences) and protein expression
was normalized to β-actin. The experiments were repeated three
times.
Statistical analysis
Data are expressed as the mean ± standard deviation.
The statistical significance of the differences between the control
and drug treatment groups was assessed using Student's t-test for
pair-wise comparisons, or a one-way analysis of variance followed
by a post hoc Dunnett's test for multiple comparisons. Statistical
analysis was performed using the GraphPad Prism software version
5.0 (GraphPad Software Inc., San Diego, CA, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
Inhibitory effect of ELE on cell
proliferation in breast cancer cells
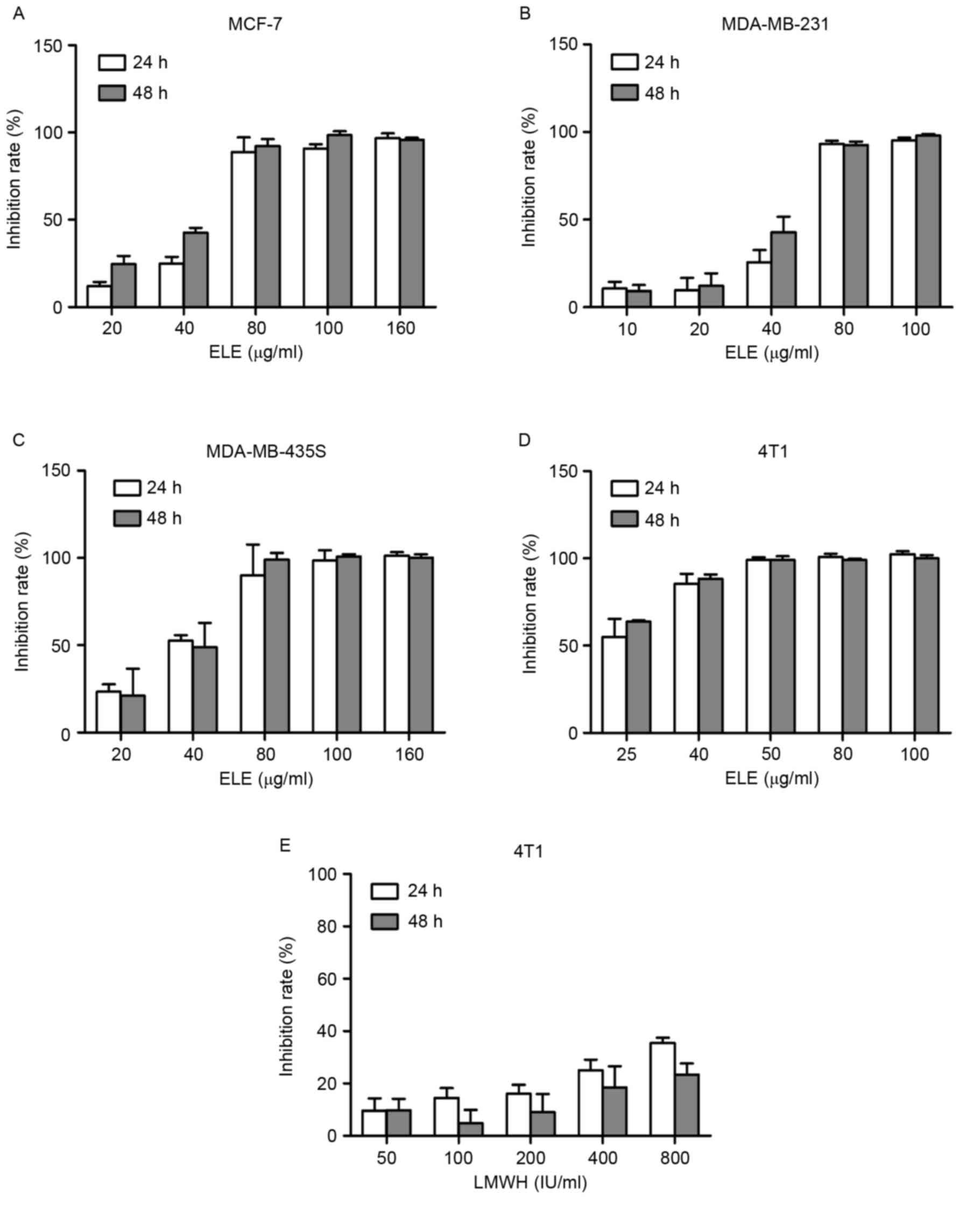
To examine the effect of ELE on cell viability in
different breast cancer cells, the cells were treated with ELE
(10–160 µg/ml) for 24 or 48 h, and the inhibitory rate of the cells
was determined using a CCK-8 assay. The half-inhibitory
concentrations of ELE were 25.31 and 21.31 µg/ml in the 4T1 cells
at 24 and 48 h respectively. The half-inhibitory concentrations
values of ELE for MCF-7 (51.26 and 38.88 µg/ml for 24 and 48 h),
MDA-MB-231 (49.9 and 41.92 µg/ml for 24 and 48 h) and MDA-MB-435s
cells (35.5 and 37.07 µg/ml for 24 and 48 h) were higher, compared
with those of the 4T1 cells. The results showed that ELE (10–80
µg/ml) resulted in a dose-dependent inhibition of 4T1, MCF-7,
MDA-MB-231 and MDA-MB-435s cells (Fig.
1A-D). In the 4T1 cells, LMWH did not exhibit an inhibitory
effect, compared with ELE (Fig.
1E).
ELE reduces the wound healing ability
of 4T1 cells
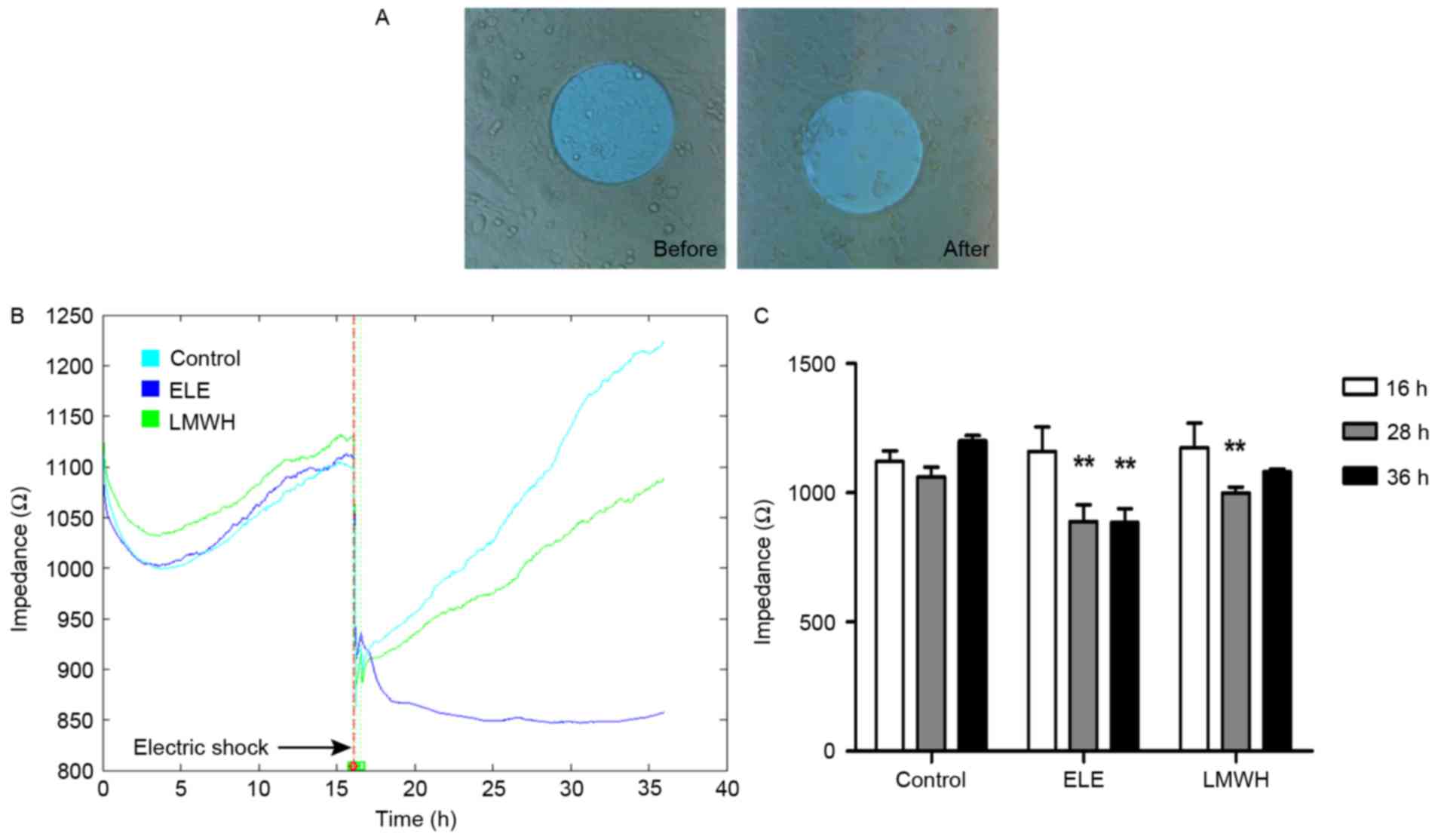
To assess the wound healing ability of the
ELE-treated 4T1 cells, impedance was determined using ECIS
following electric shock wounding. Following wounding of the cells
electrically, the impedance recorded using ECIS returned to the
basal level, as the cells attached on the electrode were
compromised (Fig. 2A).
Subsequently, the healthy neighboring cells migrated inwards and
replaced the dead cells. With drug treatment, the speed and ability
of cell migration were altered to a certain extent. The impedance
of the control group healed to its primary level (16 h, fully
confluent) at 28 h, whereas the LMWH group required a longer
duration to heal (~36 h). Additionally, the ELE group failed to
repair over the 36 h (Fig. 2B),
and the differences in impedance between pre- and post-wounding
were significant (Fig. 2C). These
results suggested that the wound healing ability of the ELE-treated
cells was limited.
ELE decreases cell migration and
invasion
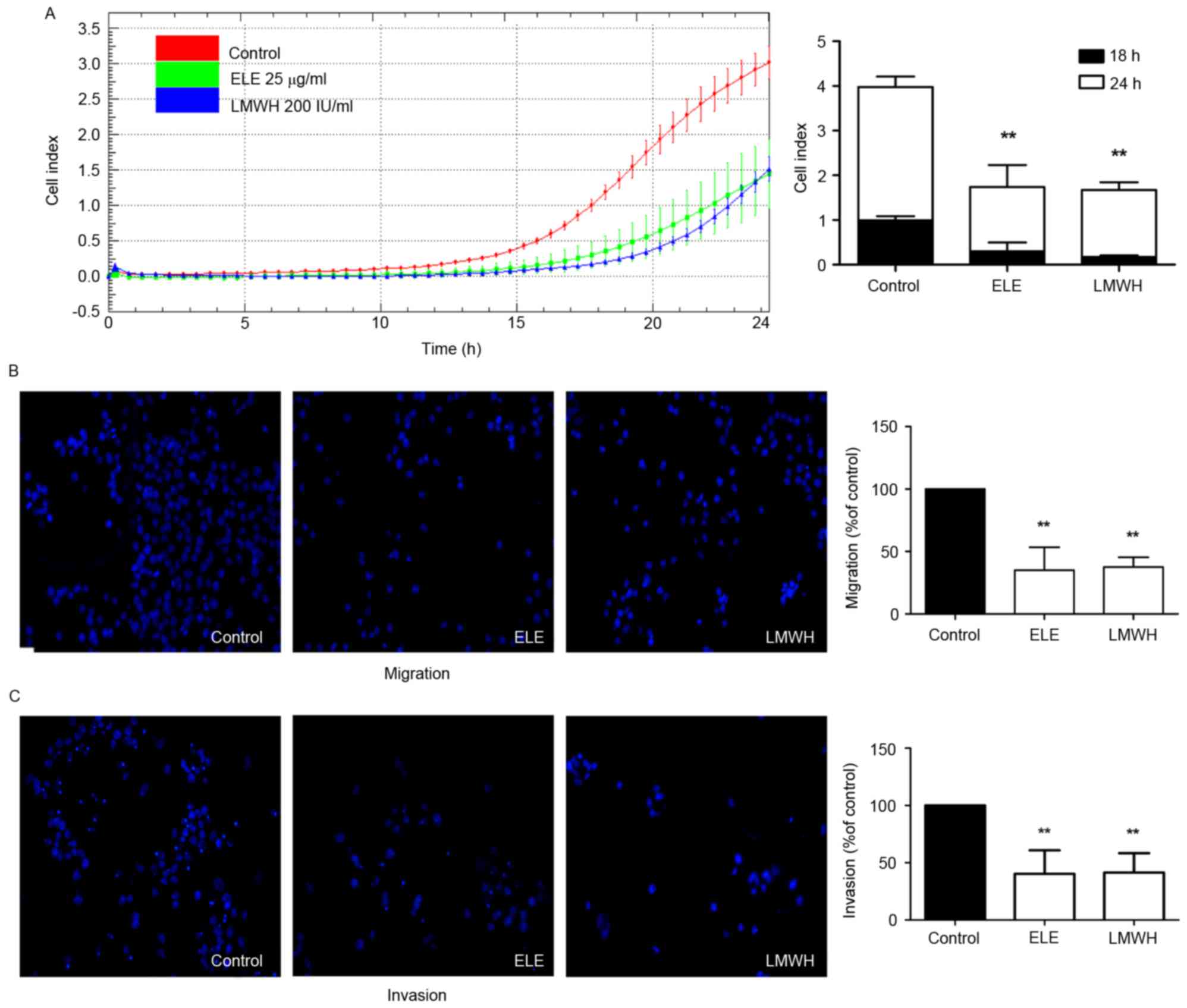
The present study further investigated whether ELE
can inhibit the migratory and invasive ability of 4T1 cells using
impedance-based detection of migration and a Transwell assay. The
cells in the control group began to migrate earlier, compared with
those in the ELE and LMWH groups. The migration ability of the
control group also remained higher until the end of the experiment.
The migration abilities of the cells in the ELE and LMWH groups
were lower. The differences in cell indices between the
drug-treated and control groups at 18 and 24 h were significant
(P<0.01; Fig. 3A). The results
of the Transwell assay also demonstrated that the 4T1 cells
exhibited reduced migration and invasion in response to ELE and
LMWH treatment (P<0.01; Fig. 3B and
C). The above data indicated that ELE inhibited the migratory
and invasive capacity of the 4T1 cells.
ELE downregulates the expression of
heparanase, and suppresses the phosphorylation of ERK and AKT
In order to analyze the mechanisms underlying the
effect of ELE on 4T1 cell migration and invasion, the present study
examined the expression of heparanase and associated proteins in
ELE-treated cells. ELE treatment resulted in significant decreases
in the expression of heparanase, FGF-2 and VEGF (P<0.05;
Fig. 4B and C). LMWH treatment had
a similar effect on the expression of heparanase (P<0.01),
however, only marginal decreases were observed in the expression
levels of FGF-2 and VEGF (Fig. 4B and
C). As is already known, the overexpression of heparanase or
released growth factors from ECM enhance the activation of ERK and
AKT. In the present study, the phosphorylation levels of ERK and
AKT were suppressed following drug treatment (P<0.01; Fig. 4D).
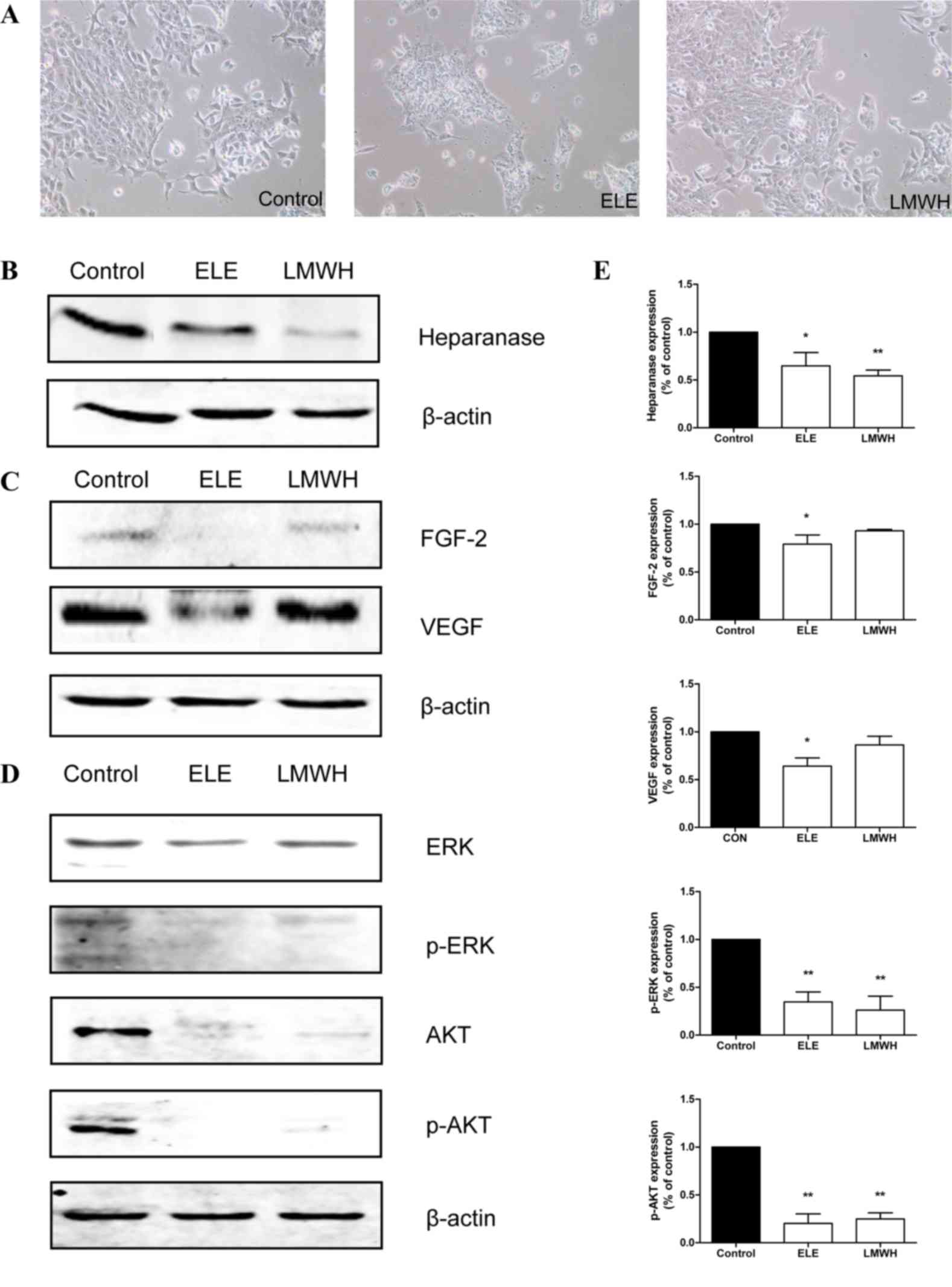 | Figure 4.ELE reduces the expression of
heparanase, and suppresses the phosphorylation of ERK and AKT. (A)
4T1 cells were cultured in medium containing ELE and LMWH.
Photomicrographs were captured under ×100 magnification. The
expression levels of (B) heparanase, (C) FGF-2, VEGF, (D) ERK/p-ERK
and AKT/p-AKT were analyzed using western blot analysis. The
relative abundance of each band to β-actin was quantified and the
control level was set to 1. (E) Data representation of the
expression levels. **P<0.01 and *P<0.05, compared with the
control. ELE, elemene; LMWH, low-molecular weight heparin; FGF-2,
fibroblast growth factor-2; VEGF, vascular endothelial growth
factor; ERK, extracellular signal-regulated kinase; p-,
phosphorylated. |
Discussion
In the present study, ELE was demonstrated to induce
significant cell toxicity in human and murine breast cancer cells
in a dose-dependent manner. The 4T1 cells were selected for further
investigation due to their higher sensitivity to ELE. These results
are consistent with previous findings that ELE has cytotoxic
effects on several types of solid tumor cell (15,23,24).
Furthermore, the present study demonstrated the inhibitory effect
of ELE on cell invasion and migration using three cellular models
of metastasis. ELE inhibited the migratory and invasive ability of
4T1 cells, and the inhibitory rates were comparable to those of
LMWH, which is a potential inhibitor of cancer metastasis (25,26).
During cancer metastasis, ECM degradation is an
important process. The overexpression of heparanase in cancer
cells, involved in the degradation of ECM, has been considered a
promising target for anticancer metastatic therapy. The inhibition
of the expression of hepranase can effectively reduce the potential
of cancer metastasis (11,27). Therefore, it is necessary to
identify drugs with antiheparanase effects. Previous studies have
shown that ELE decreases breast cancer cell invasion and migration
through upregulating the expression of E-cadherin, downregulating
the expression of MMPs or inhibiting EMT by suppressing nuclear
transcription factors (16,18,28).
By contrast, the results of the present study demonstrated that ELE
downregulated the expression of heparanase, reduced levels of
growth factors, and inactivated the phosphorylation of ERK and AKT,
which is associated with cancer growth and metastasis. Various
antiheparanase agents, including heparin, PI-88, PG545, M402 and
SST0001, have been developed and shown to inhibit cancer metastasis
(7,29–33).
Consistent with the results of the present study, these previous
studies revealed that the downregulation of heparanase and growth
factors decreased the phosphorylation of ERK and AKT. Therefore,
the findings of the present study suggested that the inhibition of
heparanase may contribute to the response of ELE in the regulation
of cell invasion and migration.
In conclusion, the results of the present study
confirmed the antiproliferative and antimetastatic effects of ELE
in vitro. Furthermore, the inhibitory effects of ELE were,
at least partly, associated with downregulation of the expression
of heparanase. Considering the aforementioned observations, ELE may
be a promising agent for the antitumor and antimetastatic treatment
of breast cancer.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81373815 and
81202840), the Specialized Research Fund for the Doctoral Program
of Higher Education of China (grant no. 20131107110014), the
Beijing Natural Science Foundation of China (grant no. 7162084) and
the Beijing Municipal Science and Technology Project (grant no.
D161100005116005).
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shi XJ, Au WW, Wu KS, Chen LX and Lin K:
Mortality characteristics and prediction of female breast cancer in
China from 1991 to 2011. Asian Pac J Cancer Prev. 15:2785–2791.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Guan X: Cancer metastases: Challenges and
opportunities. Acta Pharm Sin B. 5:402–418. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Brown GT and Murray GI: Current
mechanistic insights into the roles of matrix metalloproteinases in
tumour invasion and metastasis. J Pathol. 237:273–281. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Vlodavsky I, Friedmann Y, Elkin M, Aingorn
H, Atzmon R, Ishai-Michaeli R, Bitan M, Pappo O, Peretz T, Michal
I, et al: Mammalian heparanase: Gene cloning, expression and
function in tumor progression and metastasis. Nat Med. 5:793–802.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Maxhimer JB, Quiros RM, Stewart R,
Dowlatshahi K, Gattuso P, Fan M, Prinz RA and Xu X: Heparanase-1
expression is associated with the metastatic potential of breast
cancer. Surgery. 132:326–333. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li JP: Heparin, heparan sulfate and
heparanase in cancer: Remedy for metastasis? Anticancer Agents Med
Chem. 8:64–76. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wu BW, Li DF, Ke ZF, Ma D, Li YJ, Gang D,
Zheng ZG, Zhang KJ and Zhang YH: Expression characteristics of
heparanase in colon carcinoma and its close relationship with
cyclooxygenase-2 and angiogenesis. Hepatogastroenterology.
57:1510–1514. 2010.PubMed/NCBI
|
|
9
|
dos Santos Fernandes TC, Gomes AM,
Paschoal ME, Stelling MP, Rumjanek VM, Ado Junior R, Valiante PM,
Madi K, de Souza Pereira HS, Pavão MS and Castelo-Branco MT:
Heparanase expression and localization in different types of human
lung cancer. Biochim Biophys Acta. 1840:2599–2608. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Davidson B, Shafat I, Risberg B, Ilan N,
Trope' CG, Vlodavsky I and Reich R: Heparanase expression
correlates with poor survival in metastatic ovarian carcinoma.
Gynecol Oncol. 104:311–319. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang L, Sullivan PS, Goodman JC,
Gunaratne PH and Marchetti D: MicroRNA-1258 suppresses breast
cancer brain metastasis by targeting heparanase. Cancer Res.
71:645–654. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Vlodavsky I, Fuks Z, Ishai-Michaeli R,
Bashkin P, Levi E, Korner G, Bar-Shavit R and Klagsbrun M:
Extracellular matrix-resident basic fibroblast growth factor:
Implication for the control of angiogenesis. J Cell Biochem.
45:167–176. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Vlodavsky I, Goldshmidt O, Zcharia E,
Atzmon R, Rangini-Guatta Z, Elkin M, Peretz T and Friedmann Y:
Mammalian heparanase: Involvement in cancer metastasis,
angiogenesis and normal development. Semin Cancer Biol. 12:121–129.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gingis-Velitski S, Zetser A, Flugelman MY,
Vlodavsky I and Ilan N: Heparanase induces endothelial cell
migration via protein kinase B/Akt activation. J Biol Chem.
279:23536–23541. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lu JJ, Dang YY, Huang M, Xu WS, Chen XP
and Wang YT: Anti-cancer properties of terpenoids isolated from
Rhizoma Curcumae-a review. J Ethnopharmacol. 143:406–411.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang X, Li Y, Zhang Y, Song J, Wang Q,
Zheng L and Liu D: Beta-elemene blocks epithelial-mesenchymal
transition in human breast cancer cell line MCF-7 through
Smad3-mediated down-regulation of nuclear transcription factors.
PLoS One. 8:e587192013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yan B, Zhou Y, Feng S, Lv C, Xiu L, Zhang
Y, Shi J, Li Y, Wei P and Qin Z: β-Elemene-attenuated tumor
angiogenesis by targeting notch-1 in gastric cancer stem-like
cells. Evid Based Complement Alternat Med. 2013:2684682013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shi H, Liu L, Liu L, Geng J, Zhou Y and
Chen L: β-Elemene inhibits the metastasis of B16F10 melanoma cells
by downregulation of the expression of uPA, uPAR, MMP-2, and MMP-9.
Melanoma Res. 24:99–107. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Teoh ML, Fitzgerald MP, Oberley LW and
Domann FE: Overexpression of extracellular superoxide dismutase
attenuates heparanase expression and inhibits breast carcinoma cell
growth and invasion. Cancer Res. 69:6355–6363. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gong F, Jemth P, Galvis Escobar ML,
Vlodavsky I, Horner A, Lindahl U and Li JP: Processing of
macromolecular heparin by heparanase. J Biol Chem. 278:35152–35158.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Szulcek R, Bogaard HJ and van Nieuw
Amerongen GP: Electric cell-substrate impedance sensing for the
quantification of endothelial proliferation, barrier function, and
motility. J Vis Exp. 2014.doi: 10.3791/51300. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mandel K, Seidl D, Rades D, Lehnert H,
Gieseler F, Hass R and Ungefroren H: Characterization of
spontaneous and TGF-β-induced cell motility of primary human normal
and neoplastic mammary cells in vitro using novel real-time
technology. PLoS One. 8:e565912013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li QQ, Wang G, Huang F, Banda M and Reed
E: Antineoplastic effect of beta-elemene on prostate cancer cells
and other types of solid tumour cells. J Pharm Pharmacol.
62:1018–1027. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen W, Lu Y, Wu J, Gao M, Wang A and Xu
B: Beta-elemene inhibits melanoma growth and metastasis via
suppressing vascular endothelial growth factor-mediated
angiogenesis. Cancer Chemother Pharmacol. 67:799–808. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ettelaie C, Fountain D, Collier ME, Beeby
E, Xiao YP and Maraveyas A: Low molecular weight heparin suppresses
tissue factor-mediated cancer cell invasion and migration in
vitro. Exp Ther Med. 2:363–367. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhong GX, Gong Y, Yu CJ, Wu SF, Ma QP,
Wang Y, Ren J, Zhang XC, Yang WH and Zhu W: Significantly
inhibitory effects of low molecular weight heparin (Fraxiparine) on
the motility of lung cancer cells and its related mechanism. Tumour
Biol. 36:4689–4697. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu H, Chen X, Gao W and Jiang G: The
expression of heparanase and microRNA-1258 in human non-small cell
lung cancer. Tumour Biol. 33:1327–1334. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang X, Zhang Y and Li Y: β-elemene
decreases cell invasion by upregulating E-cadherin expression in
MCF-7 human breast cancer cells. Oncol Rep. 30:745–750.
2013.PubMed/NCBI
|
|
29
|
Joyce JA, Freeman C, Meyer-Morse N, Parish
CR and Hanahan D: A functional heparan sulfate mimetic implicates
both heparanase and heparan sulfate in tumor angiogenesis and
invasion in a mouse model of multistage cancer. Oncogene.
24:4037–4051. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhou H, Roy S, Cochran E, Zouaoui R, Chu
CL, Duffner J, Zhao G, Smith S, Galcheva-Gargova Z, Karlgren J, et
al: M402, a novel heparan sulfate mimetic, targets multiple
pathways implicated in tumor progression and metastasis. PLoS One.
6:e211062011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Winterhoff B, Freyer L, Hammond E, Giri S,
Mondal S, Roy D, Teoman A, Mullany SA, Hoffmann R, von Bismarck A,
et al: PG545 enhances anti-cancer activity of chemotherapy in
ovarian models and increases surrogate biomarkers such as VEGF in
preclinical and clinical plasma samples. Eur J Cancer. 51:879–892.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hammond E, Brandt R and Dredge K: PG545, a
heparan sulfate mimetic, reduces heparanase expression in vivo,
blocks spontaneous metastases and enhances overall survival in the
4T1 breast carcinoma model. PLoS One. 7:e521752012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ritchie JP, Ramani VC, Ren Y, Naggi A,
Torri G, Casu B, Penco S, Pisano C, Carminati P, Tortoreto M, et
al: SST0001, a chemically modified heparin, inhibits myeloma growth
and angiogenesis via disruption of the heparanase/syndecan-1 axis.
Clin Cancer Res. 17:1382–1393. 2011. View Article : Google Scholar : PubMed/NCBI
|